Embed Size (px)
DESCRIPTION
Orban Et Al., 2011
Citation preview

7/18/2019 Orban Et Al., 2011
http://slidepdf.com/reader/full/orban-et-al-2011 1/10
FUNCTIONAL NEUROANATOMY ASSOCIATED WITH THEEXPRESSION OF DISTINCT MOVEMENT KINEMATICS IN MOTORSEQUENCE LEARNING
P. ORBAN,a,b* P. PEIGNEUX,b,c O. LUNGU,a K. DEBAS,a
M. BARAKAT,a P. BELLEC,a H. BENALI,d P. MAQUETb,e
AND J. DOYONa,d
aFunctional Neuroimaging Unit, Geriatric Institute Research Center,
Université de Montreal, Queen Mary 4565, Montreal, Canada H3W
1W5
bCyclotron Research Center, Université de Liège, Belgium
c Neuropsychology and Functional Neuroimaging Research Unit, Uni-
versité Libre de Bruxelles, Belgium
d Unité Mixte de Recherche-S 678, Institut National de la Santé et de laRecherche Médicale/University of Paris 6, Centre Hospitalier Univer-
sitaire Pitié-Salpêtrière, Paris, France
eDepartment of Neurology, CHU, Université de Liège, Belgium
Abstract—A broad range of motor skills, such as speech and
writing, evolves with the ability to articulate elementary motor
movements into novel sequences that come to be performed
smoothly through practice. Neuroimaging studies in humans
have demonstrated the involvement of the cerebello-cortical
and striato-cortical motor loops in the course of motor se-
quence learning. Nonetheless, the nature of the improvement
and brain mechanisms underlying different parameters of
movement kinematics are not yet fully ascertained. We aimed
at dissociating the cerebral substrates related to the increase
in performance on two kinematic indices: velocity, that is the
speed with which each single movement in the sequence is
produced, and transitions, that is the duration of the gap
between these individual movements. In this event-related
fMRI experiment, participants practiced an eight-element se-
quence of finger presses on a keypad which allowed to re-
cord those kinematic movement parameters. Velocity was
associated with activations in the ipsilateral spinocerebellum
(lobules 4-5, 8 and medial lobule 6) and in the contralateral
primary motor cortex. Transitions were associated with in-
creased activity in the neocerebellum (lobules 6 bilaterally
and lobule 4-5 ipsilaterally), as well as with activations within
the right and left putamen and a broader bilateral network of
motor cortical areas. These findings indicate that, rather than
being the product of a single mechanism, the general im-
provement in motor performance associated with early motor
sequence learning arises from at least two distinct kinematic
processes, whose behavioral expressions are supported by
partially overlapping and segregated brain networks. © 2011
IBRO. Published by Elsevier Ltd. All rights reserved.
Key words: fMRI, humans, motor control, sequence learning,
putamen, cerebellum.
From early childhood, the human brain builds and retains abroad repertoire of complex motor behaviors. Many of them may be conceived as single movements, for instancewhen reaching tools and throwing objects, or may consistof even simpler ballistic actions, for example when per-forming pinch finger presses. Numerous motor actionsadditionally require the capacity to combine and articulateelementary movements into a well-coordinated sequencelike when speaking or writing. Due to brain plasticity, our innate, limited motor repertoire expands over life in order toadapt to novel environmental demands and to carry outmore complex actions. Thus, adjustments with practicemay occur through motor adaptation at the level of singlemovements, and through learning of novel motor se-quences whereby complex chains of movements eventu-ally come to be performed as fast and automatic actionunits. Although most often treated as separate abilities inlaboratory settings, it should be stressed that motor adap-tation of single movements and motor sequence learning
may actually coexist in real-life behaviors, suggesting thatthe motor system can acquire them simultaneously (Over-duin et al., 2008; Moisello et al., 2009). In fact, in manycases, learning a novel motor sequence involves the con-comitant motor adaptation of the sequence’s elementarycomponents.
Brain imaging research has used a variety of experi-mental paradigms to identify the brain areas associatedwith the acquisition of motor sequences in humans (Hiko-saka et al., 1999, 2002; Doyon and Benali, 2005; Ashe etal., 2006; Orban et al., 2008; Doyon et al., 2009). In theserial reaction time task, subjects are asked to learn, inci-dentally or intentionally, a new motor sequence by press-
ing spatially compatible key presses corresponding to spe-cific series of stimuli (Peigneux et al., 2000; Doyon et al.,2002; Seidler et al., 2002; Destrebecqz et al., 2005). Other paradigms require to discover a repeating pattern of motor responses by trial-and-error (Jueptner et al., 1997a,b), toexecute an explicitly known motor sequence paced viavisual or auditory cues (Karni et al., 1995; Lehéricy et al.,2005), or to learn to draw simple Chinese word charactersforming a graphomotor sequence in order to study hand-writing in a more naturalistic context (Swett et al., 2010).Together, such studies have provided valuable insightsinto the brain structures mediating this form of learning,highlighting the role of the cortico-striatal and cortico-cer-
*Correspondence to: P. Orban, University of Montreal, Geriatric Insti-tute Research Center, Functional Neuroimaging Unit, 4565 QueenMary, Montreal, QC, Canada H3W1W5. Tel: 1-514-340-3540, ext:4114.E-mail address: [email protected] (P. Orban).
Abbreviations: FDR, false discovery rate; fMRI, functional magneticresonance imaging; MNI, Montreal Neurological Institute; PPM, pos-terior probability maps.
Neuroscience 179 (2011) 94–103
0306-4522/11 $ - see front matter © 2011 IBRO. Published by Elsevier Ltd. All rights reserved.doi:10.1016/j.neuroscience.2011.01.040
94

7/18/2019 Orban Et Al., 2011
http://slidepdf.com/reader/full/orban-et-al-2011 2/10
ebellar systems in particular. Nonetheless, most of theseexperimental paradigms have essentially explored thesubjects’ ability to acquire an “abstract representation” of the order of the sequence elements, rather than investi-gating the nature of the motoric improvement measuredduring skill sequential motor learning (Krakauer and Shad-mehr, 2006). Indeed, the behavioral expression of learninghas usually been assessed through global measures of performance such as the subjects’ overall improvement inaccuracy or reaction time, rather than through trial-by-trialchanges in different movement kinematics subtending theunconstrained execution of a motor sequence. A recentcombined fMRI and kinematic approach however demon-strates how studying the precise relationship betweenchanges in neural activity and the improvement on a re-fined performance measure such as normalized jerk caninform on motor sequence learning processes (Swett et al.,2010). Nonetheless, it is still unclear how different aspectsof the motor sequence learning process, in particular per-
formance changes in velocity and transitions between sin-gle movements, are individually mediated at the brainlevel.
Using skill finger tapping tasks that favor the uncon-fined execution of a series of digit movements (Korman etal., 2003; Walker et al., 2003; Hotermans et al., 2006),recent behavioral studies have revealed that some move-ment transitions embedded within the motor sequence candifferently benefit from practice (Kuriyama et al., 2004;Sheth et al., 2008). Such results indicate that the temporalstructure of the motor sequence comes to be reorganizedduring learning through the development of chunking andcoarticulation mechanisms, which enable grouping individ-
ual motor items into larger units (Gobet et al., 2001; Sakaiet al., 2003, 2004). However, the neural substrates specif-ically involved in the overall transition gains induced bytraining have not yet been identified. Furthermore, thebehavioral effect of training on velocity of finger pinchmovements and its associated brain mechanisms have notbeen specifically assessed in the context of a sequencelearning task involving multiple fingers. Indeed, velocitygains, which reflect dynamic motor adaptation processes,have only been tested while subjects produce repetitiveballistic contractions of a single finger, typically the thumb(Classen et al., 1998; Muellbacher et al., 2001, 2002).Thus, how changes in elementary parameters of singlemovements may affect the behavioral manifestation and
brain mechanisms engaged during the course of motor sequence learning still remains unknown.
We therefore sought to provide a refined behavioraland neurophysiological description of the effects of im-proved performance on distinct movement kinematics dur-ing the course of skill finger sequence learning. In partic-ular, this study aimed at better characterizing the functionalroles that motor regions like the cerebellum, the putamen,and other motor-related cortical areas bear on genuinemotoric properties (compared to more cognitive aspects)as subjects acquire a new motor sequence. To do so, weused an event-related functional magnetic resonance im-aging (fMRI) paradigm and a dedicated keypad that can
precisely measure the kinematics of finger displacementsduring a sequential tapping task. This approach allowed usto test the hypothesis that the performance improvementsrelated to velocity (V) and transitions (T) contribute differ-ently to motor sequence learning, and that these kinematicproperties are mediated by partially segregated networksof motor areas in the human brain.
EXPERIMENTAL PROCEDURES
Participants
Twelve volunteers (six females; group mean age 26.53.8 years)gave informed consent to take part in this study, which wasapproved by the local Ethics Committee at the Geriatric InstituteResearch Center, University of Montreal. All subjects were right-handed as assessed by a questionnaire (Oldfield, 1971) and hadno history of neurological or psychiatric disorder. Musicians andprofessional typists were excluded in order to control for pre-existing skills that require highly coordinated finger dexterities.
Experimental task
Participants were trained on a sequential finger tapping taskadapted for use in an event-related fMRI design. The task wasabout 20 min long and included 60 practice trials that were inter-spersed with rest epochs lasting 12 s on average (jittering rangingfrom 6 s to 18 s). Before scanning began (i.e., during the pre-training session) subjects were asked to memorize the sequenceusing a verbal code (3 4 1 3 2 1 4 2; where 1, 2, 3, and 4respectively refer to the index, middle, ring, and little fingers),without actual training on the keypad. This was done so as tostudy brain mechanisms related to implicit motor learning pro-cesses rather than to the explicit acquisition of the sequenceorder. During scanning, each trial required subjects to prepare,and then execute the explicitly known eight-element sequence.Each rest epoch, during which a black screen was displayed, was
followed by a preparation and an execution phase. The time toprepare for the execution of a sequence (i.e., planning phase) wasindicated to the subject by the brief appearance (500 ms) of ayellow square at the center of the black screen. Subjects wereasked to prepare for sequence execution for a few seconds andthen to feel free to start typing it at the moment of their choice, withno further external cuing, in order to promote self-initiation of theseries of movements. Results revealed that the preparation phaselasted 4.24 s (SD0.91 s) on average (data for the preparationphase are further explored in a separate paper). As soon as thefirst of the eight sequential finger movements was initiated, a bluesquare was displayed for 500 ms at the center of the black screen,confirming the beginning of the execution phase. Subjects wererequired to produce the motor sequence only once on each trial,as fast and accurately as possible.
Material and data analysis
We used a custom-made MR compatible keypad allowing to mea-sure precise finger movement kinematics (Fig. 1). The responsebox was built using four industrial push button switches fixated ona plastic enclosure. All electrical contacts were removed to keeponly the actuator and spring mechanisms. Total displacement of the actuator was 5 mm. Pressing down a button moved a linear encoder strip in front of an optical reading module that sensedmotion and direction, and that was used to send the correspond-ing digital impulses to the interface unit located outside the MRroom. The linear encoder strip was a thin Mylar strip with alter-nating lines and transparent areas (250 lines per inch). The read-ing head used two optical sensors in phase quadrature, meaningthat four impulses were sent to the interface (one count for every
P. Orban et al. / Neuroscience 179 (2011) 94 –103 95

7/18/2019 Orban Et Al., 2011
http://slidepdf.com/reader/full/orban-et-al-2011 3/10
transition) for every full cycle. Motion resolution was 1000 countsper inch (one count every 25 m). Position counts received by theinterface module were stored in a 12 bits counter and those valueswere continuously fed to a digital-to-analog converter. The result-ing signal was then digitized using a 12 bits digitizer module(National Instruments USB-6008, National Instruments Inc., Aus-tin, TX, USA) at a sampling rate of 400 Hz per channel.
Data recorded for each key press were initially saved asarbitrary voltage values, which were directly proportional to thedisplacement of each button. They were temporally smoothed with
a triangular smoothing function to reduce noise. Minimum andmaximum voltage measures obtained for each of the four keyswere normalized to respective values of 0 and 100 (% of verticaldisplacement) such that reliable comparisons of movement kine-matics between fingers could be made. Several time points of interest on each trial were then computed (see Fig. 1): initiation,completion, and release of each button press. Button press dura-tion (i.e., velocity [V]) was obtained by calculating the difference induration between the initiation and completion of each individualmovement (yellow markers in Fig. 1). Although velocity is typicallydescribed as a measure of distance per time unit, time alone wasconsidered here as a measure of velocity because the amplitudeof finger movements remained constant throughout training. Theduration between movements (i.e., transition [T]) was calculated,for each pair of individual movements, by measuring time differ-ences between the release of a given finger movement and theinitiation of the next one (blue markers in Fig. 1). Incorrect se-quences of finger presses were identified as errors and wererejected from the analyses (see below).
FMRI procedures
Brain imaging data were acquired on a 3.0 Tesla MRI scanner (Magnetom Tim Trio, Siemens) with a 12-channel head coil. Func-tional T2*weighted volumes were obtained using a blood-oxy-gen-level-dependent (BOLD) sensitive, single-shot echo planar sequence (TR1000 ms; TE30 ms; FA70°; FoV220220mm2; matrix size6464; voxel size3.43.47 mm3; gap25%; 16 slices). Structural T1*-weighted MRI scans were alsoacquired, using a turbo flash sequence with an inversion pulse(TR2300 ms; TE2.91 ms; FA09°; FoV256256 mm2; ma-
trix size256256; voxel size111.2 mm3; gap50%; 160slices).
Processing and statistical analyses of brain images wereperformed using SPM2 (Wellcome Department of Cognitive Neu-rology, London, UK) implemented in Matlab 7 (Mathworks Inc.,Sherbom, MA, USA). Spatial preprocessing included realignmentand adjustment for in-scanner head movement related effects,coregistration of functional and anatomical images, spatial nor-malization into the stereotactic Montreal Neurological Institute(MNI) space, and spatial smoothing using a Gaussian kernel of 7mm full width at half maximum (FWHM). Statistics were performedusing the linear general model (Friston et al., 1995). At the intra-individual level, the design matrix included two regressors thatmodelled the preparation and execution epochs, respectively. Activation effects related to the preparation phase of the task werenot investigated in the present study, and thus the correspondingregressor was modelled as a covariate of no interest. Variables of interest were related to motor execution and practice-related ef-fects linked to performance on specific kinematic variables (i.e.velocity and transitions) measured during the execution of themotor sequence. Hence, the design matrix included two additionallinear parametric regressors associated with the execution ep-
ochs, modulated respectively by the trial-averaged duration of individual finger button presses (V) and the trial-averaged durationof transitions between finger movements (T). Linear contraststesting for the main effect of execution, as well as for the effects of execution modulated by performance (V or T), were convolvedwith a canonical hemodynamic response function (HRF), gener-ating fixed-effects statistical maps (Friston et al., 2002). It shouldbe noted that the parametric contrasts that tested for perfor-mance-related effects took into account the heterogeneity of thekinematic learning curves observed across subjects, as theseanalyses were performed at the individual level.
Summary statistic maps obtained for each contrast were sub-sequently entered into random-effects level models to allow infer-ences at the population level. Given the a priori knowledge basedon existing literature, loci of activations for performance-related
effects were identified within the striato-cortical and cerebello-cortical motor loops. The brain network of interest encompassedthe striatum and cerebellum, their related cortical motor and pre-motor regions as well as the thalamus. An inclusive mask delin-eating this brain network of interest (size30,000 voxels) wasthus created using the WFU PickAtlas software toolbox ( Maldjianet al., 2003). For the main effect of motor sequence execution,activation maps were thresholded and considered significant atP 0.05 after false discovery rate (FDR) correction for multiplecomparisons (Genovese et al., 2002). Activation maps obtainedfor parametric modulation effects were thresholded at P 0.005(uncorrected) to reveal the full extent of activation, but only brainregions that showed peak voxels surviving a threshold of P 0.001(uncorrected) were considered to be significantly activated.
Direct comparisons, using classical inferences, between theparametric effects for the two kinematic parameters did not yield
significant results. The absence of results likely comes from thefact that parametric effects are typically weaker than main condi-tion effects because the former is orthogonalized with respect tothe latter (Büchel et al., 1998). In addition, the learning curves for the velocity and transition variables, which were entered as cova-riates with identical onsets in the design matrix, correlated to someextent. Thus this may have lowered the likelihood of detectingsignificant differences in parametric activation effects betweenkinematic variables, although these were nonetheless found tobenefit from practice in significantly different proportions (seeResults). In order to partly address this concern, we computedposterior probability maps (PPMs) that enabled conditional or Bayesian inferences about regionally specific effects (Friston andPenny, 2003). This type of analysis allows one to determinewhether a lack of significant statistical effect in a given contrast is
Fig. 1. Experimental task and material. The finger tapping task re-quired the self-initiated execution of an 8-element sequence (3 4 1 3 21 4 2) with the left hand. A custom-made keypad allowed recording,through sensing motion modules, the precise displacement of four button switches. In the right-sided graph, the grey curve (four channelsare shown as one) depicts the sequential finger motions over time (ms)for the eight button presses of one early representative sequence. Anindividual finger movement was considered to be initiated, completedand released at respectively 10%, 70%, and 10% of the maximumbutton switch press. Button press duration (velocity, V) was obtainedby calculating the duration difference between the initiation and com-
pletion of each individual movement (yellow dots illustrate the secondkey press). Transition duration (transition, T) was calculated, for each pair of individual movements, through the time differencebetween the release of a given finger movement and the initiation of the next one (blue dots illustrate the second transition).
P. Orban et al. / Neuroscience 179 (2011) 94–10396
7/18/2019 Orban Et Al., 2011
http://slidepdf.com/reader/full/orban-et-al-2011 4/10
merely due to a failure to detect it (i.e., error of type II/falsenegative). After PPMs were computed for the contrasts of interest(parametric effects for the transition and velocity parameters), theprobability (P %) that a voxel activation would exceed the thresholddefined based on the variance in parameters over all voxels wasidentified. The interpretation of the result is thus that there is less
than P % probability that an area significantly activated in a givencontrast (e.g. transition) would also be activated in the other contrast (e.g. velocity).
RESULTS
Behavioral results
The number of erroneous sequences over the entire train-ing period was very low (3.25 out of 60 trials, SD3.67). Inaddition, the error rate did not change with practice, asindicated by the absence of significant difference betweenthe first (1.67, SD2.53) and second half (1.58, SD1.62)of the training trials [t (11)0.13; P 0.9]. Incorrect trials
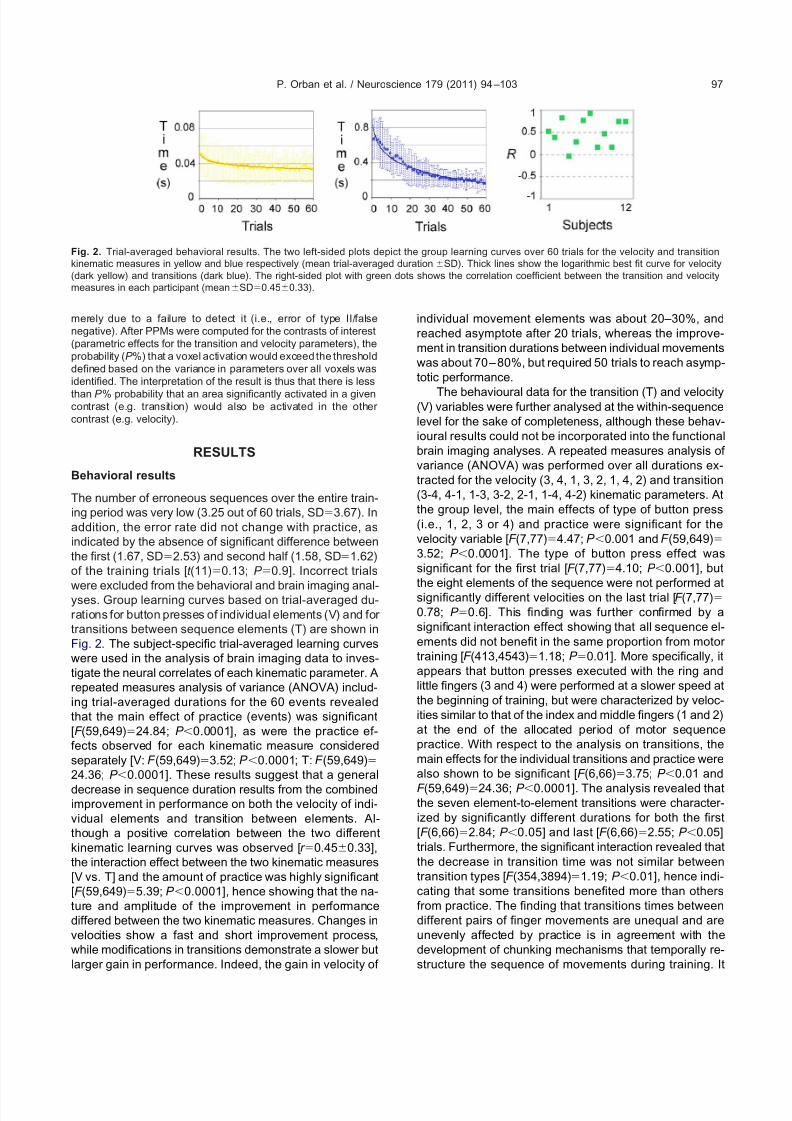
were excluded from the behavioral and brain imaging anal-yses. Group learning curves based on trial-averaged du-rations for button presses of individual elements (V) and for transitions between sequence elements (T) are shown inFig. 2. The subject-specific trial-averaged learning curveswere used in the analysis of brain imaging data to inves-tigate the neural correlates of each kinematic parameter. Arepeated measures analysis of variance (ANOVA) includ-ing trial-averaged durations for the 60 events revealedthat the main effect of practice (events) was significant[F (59,649)24.84; P 0.0001], as were the practice ef-fects observed for each kinematic measure consideredseparately [V: F (59,649)3.52; P 0.0001; T: F (59,649)24.36; P 0.0001]. These results suggest that a general
decrease in sequence duration results from the combinedimprovement in performance on both the velocity of indi-vidual elements and transition between elements. Al-though a positive correlation between the two differentkinematic learning curves was observed [r 0.450.33],the interaction effect between the two kinematic measures[V vs. T] and the amount of practice was highly significant[F (59,649)5.39; P 0.0001], hence showing that the na-ture and amplitude of the improvement in performancediffered between the two kinematic measures. Changes invelocities show a fast and short improvement process,while modifications in transitions demonstrate a slower butlarger gain in performance. Indeed, the gain in velocity of
individual movement elements was about 20–30%, andreached asymptote after 20 trials, whereas the improve-ment in transition durations between individual movementswas about 70– 80%, but required 50 trials to reach asymp-totic performance.
The behavioural data for the transition (T) and velocity(V) variables were further analysed at the within-sequencelevel for the sake of completeness, although these behav-ioural results could not be incorporated into the functionalbrain imaging analyses. A repeated measures analysis of variance (ANOVA) was performed over all durations ex-tracted for the velocity (3, 4, 1, 3, 2, 1, 4, 2) and transition(3-4, 4-1, 1-3, 3-2, 2-1, 1-4, 4-2) kinematic parameters. Atthe group level, the main effects of type of button press(i.e., 1, 2, 3 or 4) and practice were significant for thevelocity variable [F (7,77)4.47; P 0.001 and F (59,649)3.52; P 0.0001]. The type of button press effect wassignificant for the first trial [F (7,77)4.10; P 0.001], but
the eight elements of the sequence were not performed atsignificantly different velocities on the last trial [F (7,77)0.78; P 0.6]. This finding was further confirmed by asignificant interaction effect showing that all sequence el-ements did not benefit in the same proportion from motor training [F (413,4543)1.18; P 0.01]. More specifically, itappears that button presses executed with the ring andlittle fingers (3 and 4) were performed at a slower speed atthe beginning of training, but were characterized by veloc-ities similar to that of the index and middle fingers (1 and 2)at the end of the allocated period of motor sequencepractice. With respect to the analysis on transitions, themain effects for the individual transitions and practice werealso shown to be significant [F (6,66)3.75; P 0.01 and
F (59,649)24.36; P 0.0001]. The analysis revealed thatthe seven element-to-element transitions were character-ized by significantly different durations for both the first[F (6,66)2.84; P 0.05] and last [F (6,66)2.55; P 0.05]trials. Furthermore, the significant interaction revealed thatthe decrease in transition time was not similar betweentransition types [F (354,3894)1.19; P 0.01], hence indi-cating that some transitions benefited more than othersfrom practice. The finding that transitions times betweendifferent pairs of finger movements are unequal and areunevenly affected by practice is in agreement with thedevelopment of chunking mechanisms that temporally re-structure the sequence of movements during training. It
Fig. 2. Trial-averaged behavioral results. The two left-sided plots depict the group learning curves over 60 trials for the velocity and transitionkinematic measures in yellow and blue respectively (mean trial-averaged durationSD). Thick lines show the logarithmic best fit curve for velocity(dark yellow) and transitions (dark blue). The right-sided plot with green dots shows the correlation coefficient between the transition and velocitymeasures in each participant (meanSD0.450.33).
P. Orban et al. / Neuroscience 179 (2011) 94 –103 97

7/18/2019 Orban Et Al., 2011
http://slidepdf.com/reader/full/orban-et-al-2011 5/10
should be noted, however, that this group analysis onlyshows effects common to most subjects, and thus likelyovershadows specific differences with regards to the pre-cise improvement profiles observed between participants.
Imaging results
The main effect of execution of the finger tapping se-quence over 60 trials was characterized by a global in-crease in activity in a series of brain areas composing thestriato-cortical and cerebello-cortical motor loops (Table 1,Fig. 3). Large cerebellar activity was detected bilaterally in
lobules 8, 4-5 and 6. Increased activity was also observedin the contralateral putamen and thalamus bilaterally. Atthe cortical level, the primary motor cortex as well as lateraland medial premotor areas showed motor execution-re-lated activity bilaterally.
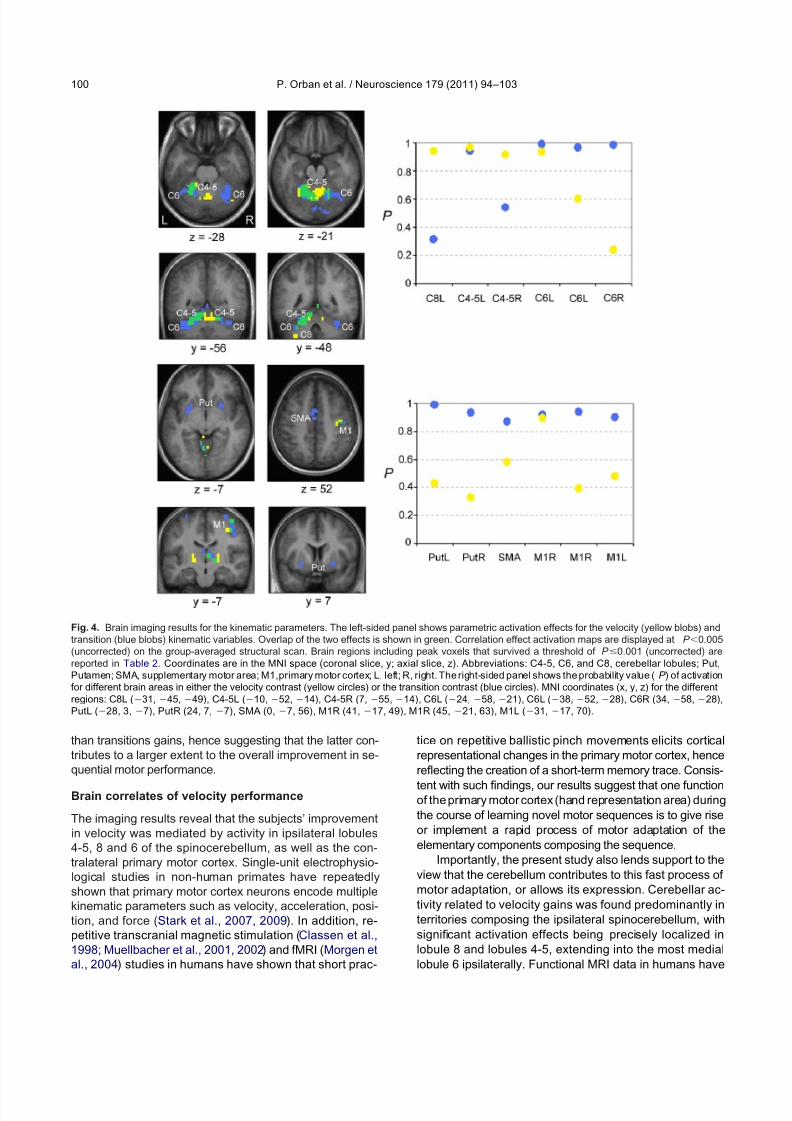
In order to determine the precise role played by thebrain regions reported above during the course of motor sequence learning, additional parametric analyses werecarried out to look for specific correlations between theincreases in brain activity and the practice-related de-creases in duration for button presses of the sequence (V),as well as for delays between them (T) (Table 2, Fig. 4).First, the results revealed that movements’ velocity modu-lated the magnitude of neural responses in the spinocer-
ebellum as significant activations were observed in theipsilateral (left) lobule 8, in lobules 4-5 bilaterally, and in themedial lobule 6 ipsilaterally. A similar modulation effectwas observed in the left thalamus and the contralateral(right) primary motor cortex (encompassing the hand rep-resentation area). Second, improvements in movements’transitions were accompanied by increased responses inboth right and left putamen, as well as in the right thala-mus. Increased transition-related activity in the cerebellumwas mostly localized in lobules 6 of the neocerebellumbilaterally, extending into cerebellar lobule 4-5 on the ipsi-lateral side. Finally, changes in transition performance atthe cortical level correlated with bilateral increases in ac-
tivation levels in the supplementary motor area and pri-mary motor cortex (including and extending outside thehand representation area). The probabilities (P ) that areaswith significant increases in activity as a function of onekinematic parameter would be similarly activated in rela-tion to the other kinematic variable are provided in Fig. 4.
Such results indicate that velocity and transition engagedoverlapping networks of brain regions (left cerebellar lob-ules 4-5 and 6 as well as right primary motor cortex), butnonetheless suggest that subsets of brain areas are morespecifically recruited to support gains in velocity (left lobule8 and right lobule 4-5 of the cerebellum) and transitionperformance (lateral cerebellar lobules 6 bilaterally, puta-men, SMA, and primary motor cortex).
DISCUSSION
Practice on a complex series of repeating finger move-ments usually leads to a gradual decrease in the time
required to execute the whole motor sequence (Korman etal., 2003; Walker et al., 2003; Kuriyama et al., 2004; Ho-
Table 1. Brain imaging results for the main effect of motor sequence
execution
Brain region x y z Z
Cerebellum lobule 8 L 17 55 49 3.11
Cerebellum lobule 8 R 14
62
49 3.04Cerebellum lobule 4-5 L 10 52 14 5.13
Cerebellum lobule 4-5 R 7 55 14 3.70
Cerebellum lobule 6 L 21 52 28 4.72
Cerebellum lobule 6 R 17 55 21 4.77
Thalamus R 17 17 7 3.53
Thalamus L 17 24 14 2.81
Putamen R 28 0 7 3.08
Pre-supplementary motor area 3 10 49 3.86
Supplementary motor area 3 0 63 3.52
Primary motor cortex R 45 17 56 4.66
Primary motor cortex L 38 21 63 3.70
Peak voxels from activation maps significant at P 0.05 (FDR-cor-
rected). x, y, and z are stereotactic coordinates in the Montreal Neu-
rological Institute (MNI) space. Z , Z-statistic score; R and L, right and
left.
Fig. 3. Brain imaging results for sequence execution. Main effectactivation maps (red blobs) are displayed at P 0.05 (FDR-corrected)on the group-averaged structural scan. Peak voxels are reported for each brain region in Table 1. Coordinates are in the MNI space(coronal slice, y; axial slice, z). Abbreviations: C4-5, C6, and C8,cerebellar lobules; Put, Putamen; SMA, supplementary motor area;M1, primary motor cortex; L, left; R, right.
P. Orban et al. / Neuroscience 179 (2011) 94–10398

7/18/2019 Orban Et Al., 2011
http://slidepdf.com/reader/full/orban-et-al-2011 6/10
termans et al., 2006; Sheth et al., 2008). This process isknown to be contingent on proper functioning of variousmotor-related brain areas (Hikosaka et al., 1999, 2002;Doyon and Benali, 2005; Ashe et al., 2006; Orban et al.,2008; Doyon et al., 2009). In particular, it has been shownthat the gradual improvement in normalized jerk during thecourse of graphomotor sequence learning is tightly accom-panied by changes in activity in the striatum, cerebellum,and associated motor cortical areas (Swett et al., 2010).The exact pattern of brain activity reorganization that ac-count for the global change in performance however likelydepends on the precise structure of the task and its exactunderlying learning processes. Our findings further indi-cate that the overall improvement in performance arisesfrom combined and simultaneous modifications in distinctbut complementary aspects of sequential movement kine-matics, namely velocity of individual movements and transi-tions between them. Velocity and transitions were found to bemediated by partially overlapping and segregated subnet-works embedded within the striato-cortical and cerebello-cortical motor loops. As will be discussed later, it is worth
noting that our results identifying the brain correlates of either velocity or transition refer to both learning-dependent andlearning-independent neural mechanisms that ultimately sub-tend together the expression of improved performance for velocity and transitions (Orban et al., 2010).
Movement kinematics during motor sequence
learning
The experimental paradigm used here allowed to studymotor sequence learning without superimposing con-straints on the execution of individual movements such aspacing or cueing. This enabled the measurement of bothvelocity and transition kinematic parameters as well as the
description of their respective contribution to the overallimprovement in performance. The results reveal that thesingle finger movements that build up the sequence benefitfrom motor training. This suggests that a fast adaptation inelementary finger motions, reflected through improvedmovement velocity, does not only take place during prac-tice of repetitive ballistic contractions performed with asingle finger (Classen et al., 1998; Muellbacher et al.,2001, 2002), but that it can also occur during learning of sequences that comprise multiple serial finger movements.Previous work has shown that brief practice of pinch move-ments, typically performed with the thumb, results in arapid adaptation of ballistic motor behaviors, as reflectedthrough the optimization of movements’ direction, acceler-ation, and force (Classen et al., 1998; Muellbacher et al.,2001, 2002). The present results show that a similar ex-perience-dependent effect on individual movements pro-duced in sequence contributes to the overall improvementin speed when executing an entire motor sequence.
In addition to changes in velocity, a gradual improve-ment was observed at the level of the element-to-elementtransitions embedded within the sequence, a finding inagreement with studies that reported such an effect usingskill finger tapping tasks similar to ours (Kuriyama et al.,2004; Sheth et al., 2008). As shown by the analysis of thewithin subjects’ trials, the performance improvement wasnot due to a global decrease in all transition times betweenmovements, but rather to a more pronounced positiveeffect of practice on some of the pairs of finger move-ments. The fact that some transitions benefited more frompractice than others suggests that the structure of themotor sequence underwent a learning-dependent tempo-
ral reorganization. Consequently, we interpret the overalldecrease in transition durations as arising from the emer-gence of chunking processes, which enable disparatesubs-sequence memory units to be gradually amalgam-ated into a larger single memory representation with prac-tice (Gobet et al., 2001; Sakai et al., 2003, 2004). Becausethis qualitative reorganization of the hierarchical structureof a motor sequence is tightly coupled to the subject-specific spontaneous emergence of rhythmic performance,the gains in transitions performance are also thought toreflect the optimization of the temporal control of sequen-tial movements (Janata and Grafton, 2003; Ivry and Spen-cer, 2004; Sakai et al., 2004).
Altogether, such findings complement those from inde-
pendent investigations showing that different aspects of motor learning can develop in parallel, presumably withoutinterfering with one another (Overduin et al., 2008; Moi-sello et al., 2009). The latter studies indicate that learningto respond to the spatial order of a series of arm-reachingmovements may occur simultaneously to motor adaptation of these movements. Our results extend such findings by show-ing that the gains in speed of movements on a skilled finger tapping task arise not only from the development of chunkingprocesses but also from the concomitant motor adaptation of individual movements. It is worth noting that the improvementprofiles related to those two kinematic measures show thatvelocity gains are smaller and occur on a shorter time-scale
Table 2. Brain imaging results for the velocity and transition kinematic
variables
Brain region x y z Z C
Velocity
Cerebellum lobule 8 L
31
45
49 3.46 8Cerebellum lobule 4-5 L 10 52 14 3.66
67Cerebellum lobule 4-5 R 7 55 14 3.32
Cerebellum lobule 6 L 24 58 21 3.30 77
Thalamus L 21 21 7 3.26 12
Primary motor cortex R 41 17 49 3.49 12
Transition
Cerebellum lobule 4-5 L 7 55 14 3.53 44
Cerebellum lobule 6 L 38 52 28 3.51120
Cerebellum lobule 6 R 34 58 28 3.66
Thalamus R 14 21 0 3.22 27
Putamen R 24 7 7 3.28 9
Putamen L 28 3 7 3.97 9
Supplementary motor area 0 7 56 3.05 17
Primary motor cortex R 45 21 63 3.65 20
Primary motor cortex L 31 17 70 3.09 4
Peak voxels are significant at P 0.001 (uncorrected). x, y, and z are
stereotactic coordinates in the Montreal Neurological Institute (MNI)
space. Z , Z-statistic score (Z ); C, cluster size that shows the full extent
of activation at P 0.005 (uncorrected); R and L, right and left.
P. Orban et al. / Neuroscience 179 (2011) 94 –103 99
7/18/2019 Orban Et Al., 2011
http://slidepdf.com/reader/full/orban-et-al-2011 7/10
than transitions gains, hence suggesting that the latter con-tributes to a larger extent to the overall improvement in se-quential motor performance.
Brain correlates of velocity performance
The imaging results reveal that the subjects’ improvementin velocity was mediated by activity in ipsilateral lobules4-5, 8 and 6 of the spinocerebellum, as well as the con-tralateral primary motor cortex. Single-unit electrophysio-logical studies in non-human primates have repeatedlyshown that primary motor cortex neurons encode multiplekinematic parameters such as velocity, acceleration, posi-tion, and force (Stark et al., 2007, 2009). In addition, re-petitive transcranial magnetic stimulation (Classen et al.,1998; Muellbacher et al., 2001, 2002) and fMRI (Morgen etal., 2004) studies in humans have shown that short prac-
tice on repetitive ballistic pinch movements elicits corticalrepresentational changes in the primary motor cortex, hencereflecting the creation of a short-term memory trace. Consis-
tent with such findings, our results suggest that one functionof the primary motor cortex (hand representation area) duringthe course of learning novel motor sequences is to give riseor implement a rapid process of motor adaptation of theelementary components composing the sequence.
Importantly, the present study also lends support to theview that the cerebellum contributes to this fast process of motor adaptation, or allows its expression. Cerebellar ac-tivity related to velocity gains was found predominantly interritories composing the ipsilateral spinocerebellum, withsignificant activation effects being precisely localized inlobule 8 and lobules 4-5, extending into the most mediallobule 6 ipsilaterally. Functional MRI data in humans have
Fig. 4. Brain imaging results for the kinematic parameters. The left-sided panel shows parametric activation effects for the velocity (yellow blobs) andtransition (blue blobs) kinematic variables. Overlap of the two effects is shown in green. Correlation effect activation maps are displayed at P 0.005(uncorrected) on the group-averaged structural scan. Brain regions including peak voxels that survived a threshold of P 0.001 (uncorrected) arereported in Table 2. Coordinates are in the MNI space (coronal slice, y; axial slice, z). Abbreviations: C4-5, C6, and C8, cerebellar lobules; Put,Putamen; SMA, supplementary motor area; M1,primary motor cortex; L, left; R, right. The right-sided panel shows the probability value (P ) of activationfor different brain areas in either the velocity contrast (yellow circles) or the transition contrast (blue circles). MNI coordinates (x, y, z) for the differentregions: C8L (31, 45, 49), C4-5L (10, 52, 14), C4-5R (7, 55, 14), C6L (24, 58, 21), C6L (38, 52, 28), C6R (34, 58, 28),PutL (28, 3, 7), PutR (24, 7, 7), SMA (0, 7, 56), M1R (41, 17, 49), M1R (45, 21, 63), M1L (31, 17, 70).
P. Orban et al. / Neuroscience 179 (2011) 94–103100

7/18/2019 Orban Et Al., 2011
http://slidepdf.com/reader/full/orban-et-al-2011 8/10
shown that these cerebellar territories contain somatotopicrepresentations of the forelimb, notably encompassing thefingers and wrist areas (Grodd et al., 2001; Manni andPetrosini, 2004). This suggests that these cerebellar re-gions may cooperate with the primary motor cortex inmediating gains in velocity performance on individual fin-
ger movements. This hypothesis is consistent with recentresting-state functional connectivity work that points to theexistence of segregated functional cerebello-cerebral net-works in humans, including a sensorimotor network encom-passing the cerebellar lobules 8 and 4-5 and the primarymotor cortex (Krienen and Buckner, 2009; O’Reilly et al.,2009). Moreover, research using transneuronal tracers inmonkeys has demonstrated the existence of anatomical pro-
jections between the arm areas of both cerebral and cerebel-lar motor regions (Kelly and Strick, 2003), hence providing ananatomical substrate for the occurrence of interactive pro-cesses between these brain regions.
Brain correlates of transition performanceThe most prominent locus of transition-related activity waslocated in the lobule 6 of the neocerebellum bilaterally.
Activations in this cerebellar area have been repeatedlyobserved in motor sequence learning tasks (see Desmondand Fiez, 1998, for a review). Over the last decades, oneof the core functions of the cerebellum has been linked tothe representation of temporal information (Janata andGrafton, 2003; Ivry and Spencer, 2004). The cerebellar hemispheres, in particular the lobules 6, have been shownusing fMRI to be engaged during tasks requiring complexprocessing of timing needed for rhythm elaboration ( Aso etal., 2010; Bengtsson et al., 2004, 2005; Thaut et al., 2008),
while patients with cerebellar lesions fail to accurately master the timing component of tasks that involve movement pro-duction (Spencer et al., 2007). Thus, the integration of sen-sory-motor inputs in specific areas of the cerebellar cortex intasks with high degrees of timing complexity may be anessential part of the process leading to rhythm formation andoptimized chunking during motor sequence learning (Janataand Grafton, 2003; Sakai et al., 2004).
Bilateral activity in the putamen was linked to transitionperformance, thus arguing for a complementary functionfor this subcortical area in processing timing and rhythmsprocesses. Accordingly, data from neuroimaging studies inhealthy subjects and reports of Parkinson’s disease pa-tients highlight a role for the putamen in the internal gen-
eration of timing and beat during rhythmic performance of sequential movements (Grahn, 2009; Grahn and Rowe,2009). Such findings are also in agreement with reports of strong basal ganglia activation during learning of timedmotor sequences (Penhune and Doyon, 2002, 2005). Inaddition, basal ganglia dysfunction is accompanied withmotor learning deficits thought to be due, in part, to theinability to construct or express chunked responses (Gray-biel, 1995, 1998). Accordingly, individuals that suffer basalganglia stroke fail to hierarchically organize movementsequences into chunks as efficiently as neurologically in-tact controls (Boyd et al., 2009). In monkeys, blockade of striatal type-2 dopamine receptors prevents from chunking
movements into a smooth sequence of motor responses(Levesque et al., 2007). Altogether, findings from the pres-ent study thus suggest that both the lateral cerebellum andputamen mediate the optimization of the temporal struc-ture of the motor sequence during learning, although it isunclear how their precise functions may differ in subtend-
ing or expressing the improved timing, rhythmic andchunking processes.
At the cortical level, a role for the supplementary motor area and the primary motor cortex in supporting improvedtransitions during motor sequence learning is consistentwith previous findings highlighting their roles in the expres-sion of sequential movements. Medial premotor areas areknown to process temporal complexity in timing tasks ( Asoet al., 2010; Bengtsson et al., 2004, 2005) and to supportmultiple functions linked to the organization of movementsproduced in sequence, like coding for rank-order effects,the ordinal position of elements, and the intervals betweenelements or chunks (Nakamura et al., 1998; Shima and
Tanji, 2000; Tanji, 2001; Kennerley et al., 2004). Withrespect to the primary motor cortex, a distinct activationassociated with transition gains was observed in additionto that related to velocity performance, outside the handrepresentation area. Single-cell recording experimentsconducted in monkeys have previously revealed that theprimary motor cortex contains neuronal populations thatmay code for serial knowledge (Carpenter et al., 1999) andsequential movements (Lu and Ashe, 2005), thus support-ing the view that the role of neuronal populations withinand outside the hand representation area in the primarymotor cortex may extend beyond processing basic motor parameters such as velocity.
Limitations and perspectives
The present findings provide evidence for the existence of partly different quantitative relationships between two as-pects of movement kinematics and neural activity in specificbrain regions during practice of novel motor sequences. How-ever, it should be stressed that the present study does notallow to dissociate between the brain regions that undergo agenuine learning-dependent functional plasticity, which ac-tively induces behavioral changes, from those that merelyimplement and express the increase in performance oc-curring as a by-product of learning (Orban et al., 2010). Inother words, our findings do not allow to determinewhether neural activity within separate subareas of the two
kinematic-related brain networks is primary or secondaryto the observed behavioural change. Yet, the present re-sults confirm the presence of partially distinct brain net-works supporting velocity and transition movement kine-matics. Both aspects of sequential movements concomi-tantly improve during the course of motor sequencelearning, although they follow different temporal dynamics.In addition, the relatively low temporal resolution of fMRIprevented us from directly investigating the neural mech-anisms that may more specifically take place within eachpractice trial. Consequently, the exact nature of the brainsubstrates that dynamically subtend chunking during mo-tor sequence learning could not be investigated in detail
P. Orban et al. / Neuroscience 179 (2011) 94 –103 101

7/18/2019 Orban Et Al., 2011
http://slidepdf.com/reader/full/orban-et-al-2011 9/10
here. A more refined understanding could be provided byother brain imaging techniques that would allow comparingthe neural correlates of fast transitions (observed withinchunked segments) and slow transitions (i.e. the so-called“problem points” within a motor sequence) (Kuriyama etal., 2004; Sheth et al., 2008). For the same reason, it is
unclear how the brain could implement the optimization of specific individual elements of the motor sequence in acontext-dependent manner, based on the fact that coar-ticulation does not only have an impact on between-ele-ments segments, but also on the individual profiles of eachsequence element as a function of the preceding andsucceeding motor units (Jerde et al., 2003).
CONCLUSION
The present findings provide a dynamic perspective onmotor sequence learning, a property of the brain deemedessential for humans and other mammalians. Our resultssuggest that distinct but intertwined processes that simul-
taneously develop with motor practice exhibit differentlearning shapes, and that their expression are subtendedby partially segregated brain networks. Specifically, our results lend support to the view that the primary motor cortex and anterior spinocerebellum preferentially contrib-ute to performance changes in velocity with which elemen-tary motor units that compose the sequence are produced,whereas brain areas encompassing the anterior neocere-bellum, the putamen, and a larger extent of the frontalmotor cortex mediates more specifically the improvementsin transition performance. These results thus help to teaseapart the neural underpinnings of the expression of chunk-ing and rhythm, which are viewed as key mechanisms
involved in motor sequence learning, from those related tomore basic motor adaptation processes. These two kine-matic features of motor sequence production contributetogether to the reorganization of brain activity usually ob-served with practice on novel motor sequences.
Acknowledgments—The authors are most grateful to André Cyr
for building the MR-compatible keypad and for technical assis-
tance, to Estelle Breton and Vo An Nguyen for skillful help with
data acquisition and analysis, and to Maria Korman for early
discussions on this topic. This work was funded through a grant
from the Natural Sciences and Engineering Research Council
(NSERC, Canada) to JD. PO and PM are supported by the
National Funds for Scientific Research (FNRS, Belgium). OL is
supported by a fellowship from the Ministère du développement
économique, de l’innovation et de l’exportation (MDEIE, Canada).
REFERENCES
Ashe J, Lungu OV, Basford AT, Lu X (2006) C ortical control of motor
sequences. Curr Opin Neurobiol 16:213–221.
Aso K, Hanakawa T, Aso T, Fukuyama H (2010) Cerebro-cerebellar
interactions underlying temporal information processing. J Cogn
Neurosci 22:2913–2925.
Bengtsson SL, Ehrsson HH, Forssberg H, Ullén F (2004) Dissociating
brain regions controlling the temporal and ordinal structure of
learned movement sequences. Eur J Neurosci 19:2591–2602.
Bengtsson SL, Ehrsson HH, Forssberg H, Ullén F (2005) Effector-
independent voluntary timing: behavioural and neuroimaging evi-
dence. Eur J Neurosci 22:3255–3265.
Boyd LA, Edwards JD, Siengsukon CS, Vidoni ED, Wessel BD, Lins-
dell MA (2009) Motor sequence chunking is impaired by basal
ganglia stroke. Neurobiol Learn Mem 92:35–44.
Büchel C, Holmes AP, Rees G, Friston KJ (1998) Characterizing
stimulus-response functions using nonlinear regressors in para-
metric fMRI experiments. Neuroimage 8:140–148.
Carpenter AF, Georgopoulos AP, Pellizzer G (1999) Motor corticalencoding of serial order in a context-recall task. Science 283:
1752–1757.
Classen J, Liepert J, Wise SP, Hallett M, Cohen LG (1998) Rapid
plasticity of human cortical movement representation induced by
practice. J Neurophysiol 79:1117–1123.
Desmond JE, Fiez JA (1998) Neuroimaging studies of the cerebellum:
language, learning and memory. Trends Cogn Sci 2:355–362.
Destrebecqz A, Peigneux P, Laureys S, Degueldre C, Del Fiore G,
Aerts J, Luxen A, Van Der Linden M, Cleeremans A, Maquet P
(2005) The neural correlates of implicit and explicit sequence
learning: interacting networks revealed by the process dissociation
procedure. Learn Mem 12:480– 490.
Doyon J, Bellec P, Amsel R, Penhune V, Monchi O, Carrier J, Lehéricy
S, Benali H (2009) Contributions of the basal ganglia and function-
ally related brain structures to motor learning. Behav Brain Res
199:61–75.
Doyon J, Benali H (2005) Reorganization and plasticity in the adult
brain during learning of motor skills. Curr Opin Neurobiol 15:
161–167.
Doyon J, Song AW, Karni A, Lalonde F, Adams MM, Ungerleider LG
(2002) Experience-dependent changes in cerebellar contributions
to motor sequence learning. Proc Natl Acad Sci U S A 99:
1017–1022.
Friston KJ, Glaser DE, Henson RN, Kiebel S, Phillips C, Ashburner J
(2002) Classical and Bayesian inference in neuroimaging: applica-
tions. Neuroimage 16:484–512.
Friston KJ, Holmes AP, Poline JB, Grasby PJ, Williams SC, Frack-
owiak RS, Turner R (1995) Analysis of fMRI time-series revisited.
Neuroimage 2:45–53.
Friston KJ, Penny W (2003) Posterior probability maps and SPMs.
Neuroimage 19:1240–1249.Genovese CR, Lazar NA, Nichols T (2002) Thresholding of statistical
maps in functional neuroimaging using the false discovery rate.
Neuroimage 15:870– 878.
Gobet F, Lane PC, Croker S, Cheng PC, Jones G, Oliver I, Pine JM
(2001) Chunking mechanisms in human learning. Trends Cogn Sci
5:236–243.
Grahn JA (2009) The role of the basal ganglia in beat perception:
neuroimaging and neuropsychological investigations. Ann N Y
Acad Sci 1169:35–45.
Grahn JA, Rowe JB (2009) Feeling the beat: premotor and striatal
interactions in musicians and nonmusicians during beat percep-
tion. J Neurosci 29:7540–7548.
Graybiel AM (1995) Building action repertoires: memory and learning
functions of the basal ganglia. Curr Opin Neurobiol 5:733–741.
Graybiel AM (1998) The basal ganglia and chunking of action reper-toires. Neurobiol Learn Mem 70:119–136.
Grodd W, Hülsmann E, Lotze M, Wildgruber D, Erb M (2001) Senso-
rimotor mapping of the human cerebellum: fMRI evidence of so-
matotopic organization. Hum Brain Mapp 13:55–73.
Hikosaka O, Nakahara H, Rand MK, Sakai K, Lu X, Nakamura K,
Miyachi S, Doya K (1999) Parallel neural networks for learning
sequential procedures. Trends Neurosci 22:464–471.
Hikosaka O, Nakamura K, Sakai K, Nakahara H (2002) Central mech-
anisms of motor skill learning. Curr Opin Neurobiol 12:217–222.
Hotermans C, Peigneux P, Maertens de Noordhout A, Moonen G,
Maquet P (2006) Early boost and slow consolidation in motor skill
learning. Learn Mem 13:580–583.
Ivry RB, Spencer RM (2004) The neural representation of time. Curr
Opin Neurobiol 14:225–232.
P. Orban et al. / Neuroscience 179 (2011) 94–103102

7/18/2019 Orban Et Al., 2011
http://slidepdf.com/reader/full/orban-et-al-2011 10/10
Janata P, Grafton ST (2003) Swinging in the brain: shared neural
substrates for behaviors related to sequencing and music. Nat
Neurosci 6:682–687.
Jerde TE, Soechting JF, Flanders M (2003) Coarticulation in fluent
fingerspelling. J Neurosci 23:2383–2393.
Jueptner M, Frith CD, Brooks DJ, Frackowiak RS, Passingham RE
(1997a) Anatomy of motor learning. II. Subcortical structures and
learning by trial and error. J Neurophysiol 77:1325–1337.
Jueptner M, Stephan KM, Frith CD, Brooks DJ, Frackowiak RS, Pass-
ingham RE (1997b) Anatomy of motor learning. I. Frontal cortex
and attention to action. J Neurophysiol 77:1313–1324.
Karni A, Meyer G, Jezzard P, Adams MM, Turner R, Ungerleider LG
(1995) Functional MRI evidence for adult motor cortex plasticity
during motor skill learning. Nature 377:155–158.
Kelly RM, Strick PL (2003) Cerebellar loops with motor cortex and
prefrontal cortex of a nonhuman primate. J Neurosci 23:8432–
8444.
Kennerley SW, Sakai K, Rushworth MF (2004) Organization of action
sequences and the role of the pre-SMA. J Neurophysiol 91:
978–993.
Korman M, Raz N, Flash T, Karni A (2003) Multiple shifts in the
representation of a motor sequence during the acquisition of skilled
performance. Proc Natl Acad Sci U S A 100:12492–12497.Krakauer JW, Shadmehr R (2006) Consolidation of motor memory.
Trends Neurosci 29:58– 64.
Krienen FM, Buckner RL (2009) Segregated fronto-cerebellar circuits
revealed by intrinsic functional connectivity. Cereb Cortex 19:
2485–2497.
Kuriyama K, Stickgold R, Walker MP (2004) Sleep-dependent learning
and motor-skill complexity. Learn Mem 11:705–713.
Lehéricy S, Benali H, Van de Moortele PF, Pelegrini-Issac M, Waech-
ter T, Ugurbil K, Doyon J (2005) Distinct basal ganglia territories
are engaged in early and advanced motor sequence learning. Proc
Natl Acad Sci U S A 102:12566–12571.
Levesque M, Bedard MA, Courtemanche R, Tremblay PL, Scherzer P,
Blanchet PJ (2007) Raclopride-induced motor consolidation im-
pairment in primates: role of the dopamine type-2 receptor in
movement chunking into integrated sequences. Exp Brain Res
182:499–508.Lu X, Ashe J (2005) Anticipatory activity in primary motor cortex codes
memorized movement sequences. Neuron 45:967–973.
Maldjian JA, Laurienti PJ, Kraft RA, Burdette JH (2003) An automated
method for neuroanatomic and cytoarchitectonic atlas-based inter-
rogation of fMRI data sets. Neuroimage 19:1233–1239.
Manni E, Petrosini L (2004) A century of cerebellar somatotopy: a
debated representation. Nat Rev Neurosci 5:241–249.
Moisello C, Crupi D, Tunik E, Quartarone A, Bove M, Tononi G,
Ghilardi MF (2009) The serial reaction time task revisited: a study
on motor sequence learning with an arm-reaching task. Exp Brain
Res 194:143–155.
Morgen K, Kadom N, Sawaki L, Tessitore A, Ohayon J, Frank J,
McFarland H, Martin R, Cohen LG (2004) Kinematic specificity of
cortical reorganization associated with motor training. Neuroimage
21:1182–1187.Muellbacher W, Ziemann U, Boroojerdi B, Cohen L, Hallett M (2001)
Role of the human motor cortex in rapid motor learning. Exp Brain
Res 136:431–438.
Muellbacher W, Ziemann U, Wissel J, Dang N, Kofler M, Facchini S,
Boroojerdi B, Poewe W, Hallett M (2002) Early consolidation in
human primary motor cortex. Nature 415:640–644.
Nakamura K, Sakai K, Hikosaka O (1998) Neuronal activity in medial
frontal cortex during learning of sequential procedures. J Neuro-
physiol 80:2671–2687.
O’Reilly JX, Beckmann CF, Tomassini V, Ramnani N, Johansen-Berg
H (2009) Distinct and overlapping functional zones in the cerebel-
lum defined by resting state functional connectivity. Cereb Cortex
20:953–965.
Oldfield RC (1971) The assessment and analysis of handedness: the
Edinburgh inventory. Neuropsychologia 9:97–113.
Orban P, Lungu O, Doyon J (2008) Motor sequence learning and
developmental dyslexia. Ann N Y Acad Sci 1145:151–172.
Orban P, Peigneux P, Lungu O, Albouy G, Breton E, Laberenne F,
Benali H, Maquet P, Doyon J (2010) The multifaceted nature of the
relationship between performance and brain activity in motor se-
quence learning. Neuroimage 49:694–702.
Overduin SA, Richardson AG, Bizzi E, Press DZ (2008) Simultaneous
sensorimotor adaptation and sequence learning. Exp Brain Res
184:451–456.
Peigneux P, Maquet P, Meulemans T, Destrebecqz A, Laureys S,
Degueldre C, Delfiore G, Aerts J, Luxen A, Franck G, Van der
Linden M, Cleeremans A (2000) Striatum forever, despite se-
quence learning variability: a random effect analysis of PET data.
Hum Brain Mapp 10:179–194.Penhune VB, Doyon J (2002) Dynamic cortical and subcortical net-
works in learning and delayed recall of timed motor sequences.
J Neurosci 22:1397–1406.
Penhune VB, Doyon J (2005) Cerebellum and M1 interaction during
early learning of timed motor sequences. Neuroimage 26:801–
812.
Sakai K, Hikosaka O, Nakamura K (2004) Emergence of rhythm during
motor learning. Trends Cogn Sci 8:547–553.
Sakai K, Kitaguchi K, Hikosaka O (2003) Chunking during human
visuomotor sequence learning. Exp Brain Res 152:229–242.
Seidler RD, Purushotham A, Kim SG, Ugurbil K, Willingham D, Ashe J
(2002) Cerebellum activation associated with performance change
but not motor learning. Science 296:2043–2046.
Sheth BR, Janvelyan D, Khan M (2008) Practice makes imperfect:
restorative effects of sleep on motor learning. PLoS One 3:e3190.
Shima K, Tanji J (2000) Neuronal activity in the supplementary andpresupplementary motor areas for temporal organization of multi-
ple movements. J Neurophysiol 84:2148–2160.
Spencer RM, Verstynen T, Brett M, Ivry R (2007) Cerebellar activation
during discrete and not continuous timed movements: an fMRI
study. Neuroimage 36:378–387.
Stark E, Drori R, Abeles M (2009) Motor cortical activity related to
movement kinematics exhibits local spatial organization. Cortex
45:418–431.
Stark E, Drori R, Asher I, Ben-Shaul Y, Abeles M (2007) Distinct
movement parameters are represented by different neurons in the
motor cortex. Eur J Neurosci 26:1055–1066.
Swett BA, Contreras-Vidal JL, Birn R, Braun A (2010) Neural sub-
strates of graphomotor sequence learning: a combined fMRI and
kinematic study. J Neurophysiol 103:3366–3377.
Tanji J (2001) Sequential organization of multiple movements: involve-ment of cortical motor areas. Annu Rev Neurosci 24:631–651.
Thaut MH, Demartin M, Sanes JN (2008) Brain networks for integra-
tive rhythm formation. PLoS One 3:e2312.
Walker MP, Brakefield T, Hobson JA, Stickgold R (2003) Dissociable
stages of human memory consolidation and reconsolidation. Na-
ture 425:616–620.
(Accepted 20 January 2011)(Available online 26 January 2011)
P. Orban et al. / Neuroscience 179 (2011) 94 –103 103





























