Embed Size (px)
Citation preview

Chapter
OPTIMIZATION OF CONDITIONS FOR THE PRODUCTION
OF ALKALINE PROTEASES

~ t h the objective of obtaining high yield of alkaline proteases, factors
liM influencing the production by the selected strains were studied. The
strains selected for production by submerged and solid state fermentation were
Bacillus sp. K 25 and BaciMus purnilus K 242 respectively. The factors
influencing the production were studied one by one, examining one factor at a
time, keeping the other factors constant. Once the optimization has been
done with respect to a factor it was incorporated in the experiment for the
optimization of the next factor. Unless otherwise specified, this was the
strategy followed for designing all the experiments described under this
chapter. Experiments were done in triplicate.
Section A
OPTIMIZATION OF CONDITIONS FOR THE PRODUCTION OF ALKALINE PROTEASE
BY BACILLUS SP. K 25 BY THE SUBMERGED FERMENTATION METHOD
MATERIALS AND METHODS
The method of preparation of media, inoculation and incubation
followed for performing the experiments under this chapter were same as
described under section A of Chapter 3, except for the alterations or
modifications mentioned under each experiment.

Growth phase
Gm-acellular alkaline protease accumulation at various phases of
growth of Bacillus sp. K 25 was studied. The medium (pH 8.0) which
contained (per litre) 10 g of peptone, 10 g of beef extract, 5 g of sodium
chloride, 1 g of potassium dihydrogen phosphate, 2 g of dipotassium
hydrogen phosphate, 0.2 g of magnesium sulphate and 0.5 g calcium chloride
was inoculated with the bacterium and the culture samples were taken at
regular intervals during the incubation. The relative cell concentrations in the
samples (ODm was taken in a Systronics spectrocolorirneter 103, path
length: 1 cm) and the alkaline protease activity in the culture supernatants
were determined.
Temperature of Incubation
The effect of temperature on the alkaline protease production was
studied using the same medium as in the previous experiment. The medium
was inoculated, incubated at different temperatures and the alkaline protease
activity was determined.
The medium as in the previous experiments, prepared with different
pH was used for studying the effect of pH on the alkaline protease production.
Carbon and nitrogen sources
The effect of different carbon sources on alkaline protease production
by the strain was studied, using the media containing (gil) ammonium
sulphate, 5; sodium chloride, 5; calcium chloride, 0.5; magnesium sulphate,
0.2; dipotassium hydrogen phosphate, 2; potassium dihydrogen phosphate, 1,
and carbon source, 10.

The effect of different nitrogen sources on the enzyme production was
studied using the media containing (gA) starch, 10; sodium chloride, 5;
calcium chloride, 0.5; magnesium sulphate, 0.2; dipotassium hydrogen
phosphate, 2; potassium dihydrogen phosphate, 1, and each of the different
nitrogen sources, 10.
Starch and soya bean meal which were found to be the best carbon
and nitrogen sources respectively were used together at different
concentrations and the effects on the production were studied. The media
used contained (911) sodium chloride, 5; calcium chloride, 0.5; magnesium
sulphate, 0.2; dipotassium hydrogen phosphate, 2; potassium dihydrogen
phosphate, 1 and different concentrations of starch and soya bean meal.
The effect of using different concentrations of sodium chloride in the
medium, on alkaline protease production was studied using media containing
(911) soya bean meal, 20; starch, 20; dipotassium hydrogen phosphate, 2;
potassium dihydrogen phosphate, 1, and different concentrations of sodium
chloride.
The effect of using other salts in addition to sodium chloride was
studied. The media used for this experiment contained (gA) soya bean meal,
20; starch, 20; dipotassium hydrogen phosphate, 2; potassium dihydrogen
phosphate, 1; sodium chloride, 10, and each of the various salts, 0.5.
The effect of using calcium chloride at different levels in addition to
sodium chloride was studied by using media containing (gll) soya bean meal,
20; starch, 20; dipotassium hydrogen phosphate, 2; potassium dihydrogen

phosphate, 1; sodium chloride, 10, and different concentrations of calcium
chloride.
Age of lnoculum
The effect of age of inoculum on alkaline protease production by the
strain was studied using inocula aged 12, 24, 36 and 48 h. The production
medium used (starch-soya bean meal medium) contained (dl) soya bean
meal, 20; starch, 20; dipotassium hydrogen phosphate, 2; potassium
dihydrogen phosphate, 1; sodium chloride, 10, and calcium chloride, 0.2.
Size of inoculum
Starch-soya bean meal medium mentioned under the previous
experiment was inoculated with different volumes of 24 h grown nutrient
broth culture so that the final level of inoculum varied from 0.5-12%.
Agitation
The alkaline protease production by the unagitated culture and the
culture agitated at different rates was determined.
Incubation period
The starch-soya bean meal medium was inoculated with 24 h grown
nutrient broth culture at a level of two percent and the yield of alkaline
protease was determined after incubation for different periods.

RESULTS
Effect of growth phase
Alkaline protease production by the strain Bacillus sp. K 25, as a
function of growth at room temperature is shown in Figure 1.
Figure
Time (h)
1. Time course of growth and alkaline protease production by Bacillus sp. K 25
The production of enzyme by the strain could be noticed from the early
exponential phase of growth. The bacterium was producing only small
quantities of the enzyme in the early stages. A steady increase in production
a u l d be seen as the growth progressed from early exponential to early
stationaly phase. The maximum enzyme accumulation was seen in the early
stationay phase. Thereafter a decline in activity was observed.

Effect of temperature of incubation
The effect of temperature of incubation on the alkaline protease
production by Bacillussp. K 25 is shown in Figure 2.
I 25 30 35 40 45 50 55 60
Temperature of incubation PC)
Figure 2. Effect of temperature of incubation on alkaline protease production by Bacillus sp. K 25
The maximum alkaline protease production was seen when the culture
was incubated at 45°C.
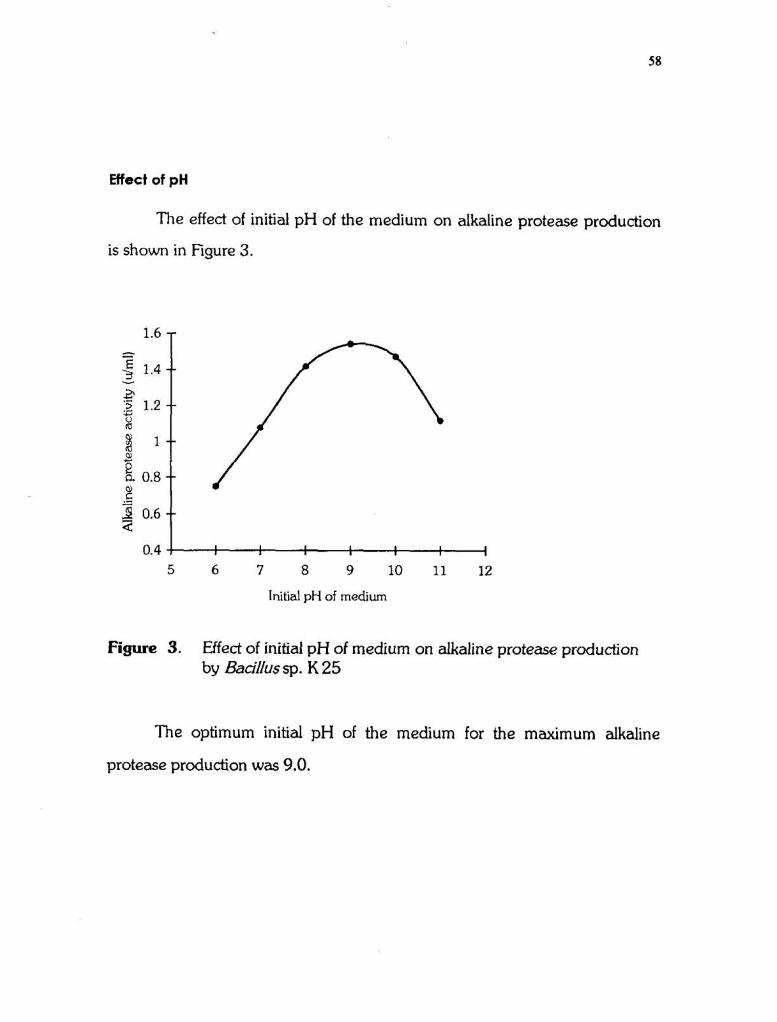
Effect of pH
The effect of initial p H of the medium on alkaline protease production
is shown in Fgure 3.
0.4 4 I 5 6 7 8 9 10 11 12
Initial pH of medium
Figure 3. Effect of initial p H of medium on alkaline protease production by Bacillussp. K 25
The optimum initial p H of the medium for the maximum alkaline
protease production was 9.0.

Effect of carbon and nltrogen sources
Results of studies on the effect of different carbon sources (1% w/v) on
alkaline protease production by the strain is shown in Table 7.
Table 7
Effect of different carbon sources on alkaline protease production by Bacillussp. K 25
Carbon source Alkaline protease production (u mi-' It SEM)
Glucose 0.135 + 0.007
Galactose 0.120 + 0.004
Fructose 0.066 + 0.003
Lactose 0.045 + 0.001
Maltose 0.116 i 0.002
Sucrose 0.123 i 0.004
Mannose 0.060 + 0.002
Mannitol 0.057 + 0.001
Inulin 0.176 + 0.008
Dexbin 0.120 + 0.003
Starch 0.192 + 0.012
Glycerol 0.082 + 0.002 &
The best carbon source for the production was found to be starch
followed by inulin.

The results of studies on the effect of different nitrogen sources on the
alkaline protease production is shown in Table 8.
Table 8
Effect of various nitrogen sources on alkaline protease production by Bacillus sp. K 25
Nitrogen source Alkaline protease production ( u ml-' * SEM)
Ammonium sulphate 0.203 1. 0.006
Ammonium chloride 0.168 kO.011
Potassium nitrate 0.062 +0.003
Beef extract 0.820 5 0.370
Casein 2.608 k 0.084
Peptone 2.191 1.0.117
Tryptone 1.660 -t 0.042
Soya bean meal 3.028 50.126
Yeast extract 1.405 +_ 0.035
The production was better with organic nitrogen sources than with the
inorganic sources. The highest production was obtained when soya bean meal
was used as the nitrogen source.

The effect of using starch and soya bean meal at different
concentrations on the enzyme production is shown in Table 9.
Table 9
Effect of using starch and soya bean meal at different concentrations on alkaline protease production by Bacillus sp. K 25
Concentration of Concentration of Alkaline protease starch (%, wlv) soya bean meal (%, wlv) production (u ml-' f SEM)
1 1 3.054 * 0.131
1 2 4.440 + 0.231
1 3 4.213 + 0.147
2 1 3.427 + 0.082
2 2 4.650 * 0.220
2 3 4.102 k 0.172
3 1 3.690 it 0.215
3 2 4.449 + 0.186
3 3 3.714 + 0.256
The maximum production was obtained when both starch and soya
bean meal were used at 2% (w/v) level.
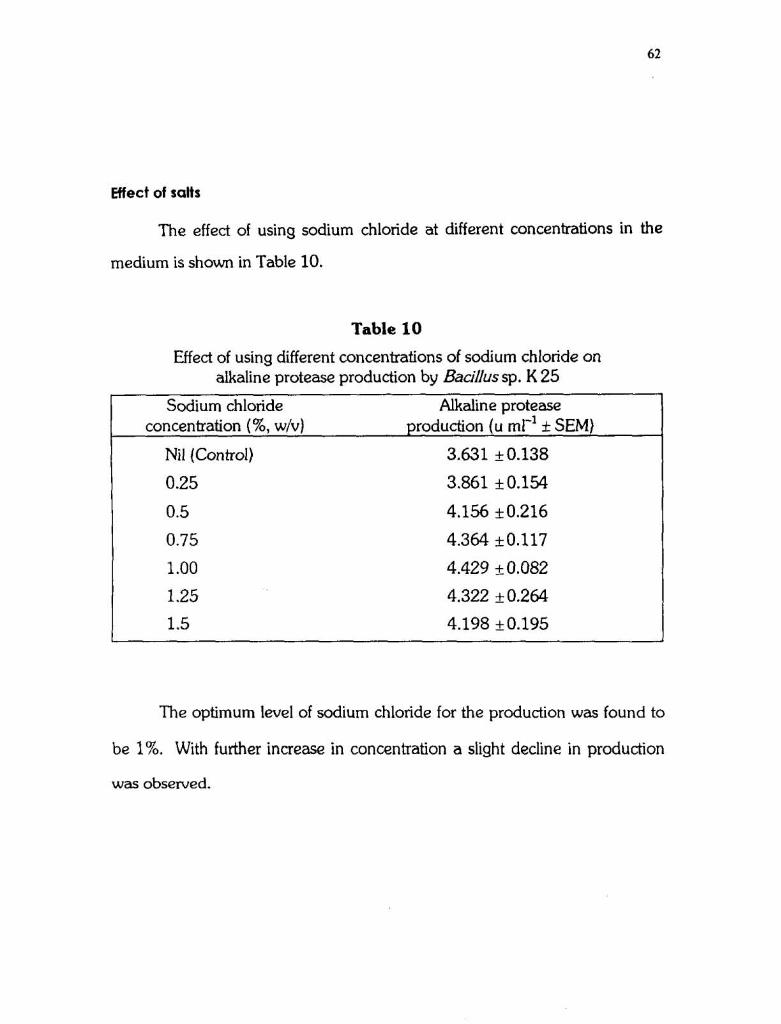
Effect of salts
The effect of using sodium chloride at different concentrations in the
medium is shown in Table 10.
Table 10
Effect of using different concentrations of sodium chloride on alkaline protease production by Bacillussp. K 25
Sodium chloride Alkaline protease concentration (%, w/v) production (u rnl-' + SEM)
Nil (Control) 3.631 k0.138
0.25 3.861 k0.154
0.5 4.156 k0.216
0.75 4.364 50.117
1.00 4.429 +_ 0.082
1.25 4.322 + 0.264
1.5 4.198 +0.195
The optimum level of sodium chloride for the production was found to
be 1%. With further increase in concentration a slight decline in production
was observed.

The effed of using other salts along with the sodium chloride is shown
in Table 11.
Table 11
Effect of different salts on alkaline protease production by Bacillus sp. K 25
Salts present Alkaline protease
in the medium production (U ml-' k SEM)
NaCI (1 %, wlv) only 4.495 k0.161
NaCI (I%, w/v)+ Calcium chloride (0.05%, wlv) 4.843 k0.219
NaCl (I%, w/v)+ Magnesium sulphate (0.05%, wlv) 4.587 k0.230
NaCI (I%, w/v)+ Ferric chloride (0.05%, wlv) 4.505 k0.181
NaC1 (I%, w/v)+ Zinc sulphate (0.05%, wlv) 3.444 k0.094
NaCl (I%, w/v)+ Cobalt chloride (0.05%, w/v) 1.799 k0.113
NaCI (I%, w/v)+ Manganese chloride (0.05%, wlv) 3.176 k0.141
NaCI (1%. w/v)+ Potassium chloride (0.05%, wlv) 4.592 k0.244
A slight increase in production could be observed in the presence of
calcium chloride. Presence of magnesium sulphate, femc chloride and
potassium chloride were having little or no effed on the enzyme production.
A decline in production was seen in the presence of zinc sulphate, cobalt
chloride and manganese chloride.

Calcium chloride which was found to be favouring the alkaline
protease production was tested at different concentrations. Results are shown
in Table 12.
Table 12
Effed of different concentrations of calcium chloride on alkaline protease production by Bacillussp. K 25
Calcium chloride Alkaline protease concentration in the medium production (u ml-' i- SEM)
(%, w/v)
0.02 5.006 k0.118
0.05 5.044 i- 0.283
There was no considerable difference in the yield of alkaline protease
with the use of calcium chloride in the range 0.02-0.2% (wlv). At higher
concentration (0.3%, w/v), a slight decline in production was seen.
Effect of age of inoculum
The effect of age of inoculum on the production is shown in Table 13.
Table 13
Effed of age of inoculum on alkaline protease production by Baci/us sp. K 25
1 Age of inoculum (h) Alkaline protease production (u mi-' i SEM)

The age of inoculum was found to be having little or no effect on the
production.
Enect of slze of inoculum
The effect of inoculum size on alkaline protease production by the
strain is shown in Table 14.
Table 14
Effect of inoculum size on alkaline protease production by Bacillus sp. K 25
lnoculum level (%) Alkaline protease production (u ml-' + SEMI
0.5 4.430 k 0.223
1.0 4.712 +0.164
2.0 4.875 k0.186
4.0 4.665 i0.219
8.0 4.864 i0.257
12.0 4.532 i0.208
The optimum level of inoculum for the enzyme production was found
to be 1-8%.

Elfect of agitation
The effect of agitation of culture on alkaline protease production is
shown in Table 15.
Table 15
Effect of agitating the culture on alkaline protease production
Agitation rate Alkaline protease (r.p.m) production (u ml-' * SEM) Unagitated 0.603 ~0.026
100 3.802 k0.203
'200 5.325 k0.276
300 5.562 k 0.250
In the unagitated culture alkaline protease was produced only in v e y
low level. The culture showed an increase in production with the increase in
the agitation rate. Vey high levels of alkaline protease could be achieved by
agitating the culture at 200-300 r.p.m.
Effect of period of incubation
The effect of varying the incubation period on alkaline protease
accumulation is shown in Table 16.
Table 16
Effect of incubation period on alkaline protease accumulation in the culture of Bacillus sp. K 25
incubation period (h) Alkaline protease production (u ml-' _+ SEM)
48 2.905 k0.191
72 5.429 k 0.292
96 6.631 k0.347
120 6.086 +0.181

The maximum accumulation of alkaline protease was seen when the
culture was incubated for 96 h. A slight decline in the activity was seen on
further incubation.
DISCUSSION
The influence of various factors on alkaline protease production by
Bacillus sp. K 25 was studied.
The alkaline protease production profiles of the strain as a function of
growth was examined in a complex medium, containing peptone and beef
extract. The production could be seen from the early exponential phase
onwards. It was very low during the early stages of exponential phase. The
alkaline protease production beginning from the early stages of exponential
phase has been reported only rarely (Fogarty and Griffin, 1973; Deane eta/.,
1986; Kaur eta/., 1998). A possible reason for the early secretion of protease
can be the absence of easily metabolizable carbon sources such as sugar in the
medium. It was suggested by Mc Donald and Chambers (1966) that, it was
the primary function of extracellular protease to ensure a supply of carbon for
growth rather than to supply amino acids for the synthetic process in the
absence of easily metabolizable carbon sources. In this study, the complete
absence of easily metabolizable carbon sources might have created a situation
where the peptide bonds in peptone or proteins had to be cleaved for
obtaining carbon for growth and metabolism. The observation of lag in
protease production in Pseudomonas fluorescens in presence of easily
metabolizable carbon source (Mc Keller, 1982) is supportive of this argument.

The culture of Bacillus sp. K 25 was showing a steady increase in
alkaline protease production with the progression of growth from early
exponential to early stationary phase. The production was maximum in the
early stationary phase. Reports are many, on the maximum extracellular
protease production occurring during the later stages of growth. Different
Bacillus species have been reported to be producing the maximum enzyme
during the late exponential (Atalo and Gashe, 1993), post exponential (Ikeda
et al., 1974; Kitada and Horikoshi, 1976; Debabov, 1982; Ward, 1983;
Manachini et a/., 1988) and the stationary (Durham, 1987; Durham ef a/.,
1987; Purva et dl., 1998), phases of growth. The exact reason for the
increased production of protease during the later stages of growth is not
known. A coincidence of reaching of extracellular protease production at the
maximum level with sporulation, the event occurring mainly during the later
stages of growth, has been reported by some workers (Debabov, 1982; Sinha
and Satyanarayana, 1991). The possibility for the existence of a relationship
between the triggering of protease production and sporulation, as observed by
Debabov (1982) in Bacillusspp., cannot be ruled out in this case also. Such a
relationship if any is there can be the reason for the increased production
during the later stages of growth. Detailed studies are required to anive at a
conclusion.
The optimum temperature for alkaline protease production by the
strain was found to be 45°C. Temperatures at or around 45°C have been
reported for the production by bacteria such as Bacillus sp. P-001A (Atalo and
Gashe, 1993) and Bacillus lichenifomis S40 (Sen and Satyanarayana,
1993).

The effed of initial pH of medium on alkaline protease production was
studied. The maximum production was seen at pH 8-10. The most optimum
initial pH of the medium was 9.0. Alkaline protease production using media
with alkaline pH has been reported by Honan Scientific Research Institute for
Leather Industry (1975), Kitada and Horikoshi (1976), Manachini et al.
(1988), Qiu et a/. (1990a, 1990b), Takii et a/. (1990), Sinha and
Satyanarayana (1991), Cheong et af. (1993), Sen and Satyanarayana (1993)
and Putva eta/. (1998).
Results of study on the effect of carbon source on alkaline protease
production shows starch to be the best carbon source followed by inulin.
Starch or starch hydrolysates have been reported as good carbon sources for
alkaline protease production by different Bacillus species (Emtseva, 1975;
Sinha and Satyanarayana, 1991; Sen and Satyanarayana, 1993; Ferrero
eta/., 1996; Purva eta/., 1998). Compared to starch and inulin, glucose and
other easily metabolizable carbon sources were not so good for alkaline
protease production by Bacillus sp. K 25. Sen and Satyanarayana (1993)
who studied alkaline protease production by Bacillus lichenifomis S40 have
also reported similar observation. The repressing effect of glucose on alkaline
protease production has been reported in Wbrio dginol@cus (Long et a/.,
1981) also.
Results of study on the effect of various nitrogen sources on alkaline
protease production by Bacillus sp. K 25 shows that the organic nitrogen
sources are better than the inorganic ones for the production. This
observation conforms with the earlier reports on the repressing effed of

inorganic nitrogen sources on bacterial alkaline protease production (Long
eta/., 1981; Fujiwara and Yamamoto, 1987; Giesecke et al., 1991; Sen and
Satyanarayana, 1993). The reason for the better production with organic
nitrogen sources can be supposed to be their ability to induce protease
production. The inducing effect of organic nitrogen sources on bacterial
alkaline protease production has been reported by Lasure (19801, Ferrero
eta/. (1996) and Kaur eta/. (1998). Of the various nitrogen sources tested
soya bean meal was found to be the best for production. It has been reported
as a suitable nitrogen source for the alkaline protease production by many
bacteria (Honan Scientific Research Institute of Leather Industry, 1975;
Chandrasekaran and Dhar, 1983; Nihete et a/., 1986; Na and Yu, 1988;
Takami et a/., 1989; Lee and Chang, 1990; Purva eta/., 1998).
Starch and soya bean meal found to be the best carbon and nitrogen
sources respectively, were tested together at different concentrations. The
optimal level of both these ingredients for the maximum production was found
to be 2% (wlv).
The results of study on the effect of sodium chloride show that the
presence of sodium chloride can enhance the alkaline protease production by
the strain. The optimum lwel of sodium chloride for the production was
1% w/v. The enhancing effect of sodium chloride on bacterial alkaline
protease production has been reported only rarely. Chandrasekaran and
Dhar (1983) who studied the alkaline protease production by Sb-eptomyces
moderatus NRRL 3150 have observed a beneficial effect of sodium chloride
on the production. The exact reason for the increased production in the

presence of sodium chloride is not known. But it is well-known that sodium
chloride at its optimum level can provide a conducive osmotic environment
for the growth of bacterial cells. The enhanced production of alkaline
protease in the presence of sodium chloride can be supposed to be an
outcome of such an effect.
The effect of using other salts in addition to sodium chloride was
studied. Of the different salts tested only calcium chloride was found to be
enhancing the production. Though calcium chloride is a very common
ingredient of production media for bacterial alkaline proteases, its role in
enhancing the production has not yet been studied. Salts such as magnesium
sulphate, ferric chloride and potassium chloride were found to be having no
effect on the production. The other salts tested, zinc sulphate, cobalt chloride
and manganese chloride were having inhibitory effects.
The effect of using calcium chloride at different concentrations was also
studied. 0.02-0.2% (wlv) level of it was found to be favouring the production.
With the use of higher concentration (0.3% w/v) a slight decline in production
could be noticed. The calcium chloride concentrations used by earlier workers
in the production media for different bacteria range from 0.006% as for a
strain of Bacillus subtihs (Massuco et al., 1980) to 0.3% as for a strain of
Baci//us pumi/us (Honan Scientific Research Institute of Leather Indushy,
1975). In fad the level of calcium chloride required in the medium depends
not only upon the bacterium used, but also upon the presence of other salts
and their concentrations in the medium. This may account for the wide

differences in calcium chloride concentration requirements by the different
bacterial SmF systems.
Studies showed that the alkaline protease production by Bacillus sp.
K 25 was independent of the age of inoculum. Similar observations have
been reported by Miusawa et d. (1969) and Sen and Satyanarayana (1993)
who studied the alkaline protease production by Streptomyces rectus var.
proteolytcus and Bacillus lichenifomis S40 respectively.
The optimum level of inoculum for alkaline protease production by the
strain was found to be 1-8%. This observation is in conformity with the
reports by Sinha and Satyanarayana (1991), Sen and Satyanarayana (1993)
and Gajju et a/. (1996) who studied the alkaline protease production by
Bacillus lichenifonnis N3, Bacillus lichenifomis S40 and Bacillus coagulans
PB 77 respectively.
Agitation of culture was found to be essential for the high production of
alkaline protease by Bacillus sp. K 25. The unagitated culture of this
bacterium was characterized by diminished growth due to pellicle fonation
over the surface of the culture. Only very low levels of alkaline protease was
produced in the unagitated culture. The culture showed an increase in
production with the increase in the agitation rate. Very high yield of alkaline
protease was obtained by agitating the culture at 200-300 r.p.m. Similar
observations have been made on the submerged fermentation systems for the
production of alkaline proteases by Bacillus lichenifomis S40 (Sen and
Satyanarayana, 1993), Bacillussp. (Takami etal., 1989) and Bacillus sp. IS-3
(Purva eta/., 1998).

The incubation period required for obtaining the maximum yield, with
starch-soya bean meal medium providing the optimum conditions was found
to be 96 h. Requirement for such a long incubation period for the maximum
alkaline protease production is not so common for bacterial SmF systems.
The long incubation period observed in the present study can be supposed to
be due to the presence in high concentrations of carbon and nitrogen sources
which are slowly metabolizable. Gajju et a/. (1996) have reported an
incubation period of 96 h for the maximum accumulation of alkaline protease
by Bacillus coagulans PB 77 in a medium containing casein.
As a result of optimization studies, the yield of alkaline protease by the
strain could be increased approximately six-fold. After optimization the
activity was more than 6.0 u ml-'. A precise comparison of the yield obtained
by Bacillus sp. K 25 with the yield in most of the earlier reports is difficult,
because different methodologies have been adopted by different workers for
the assay of alkaline proteases. The composition and pH of the buffer systems
used for assay, temperature of incubation, substrate etc. are different in
different works. Moreover the unit definitions followed by different workers for
expressing the activity were also different. So the yield of alkaline protease
obtained by Bacillus sp. K 25 could be compared only with a few earlier
reports where the methodology of assay and unit definitions were similar or
comparable. The yield by Bacillus sp. K 25 could be found to be better than
the yield reported by Chandrasekaran and Dhar (1983), Tsujibo eta/. (1990),
Sinha and Satyanarayana (1991), Sen and Satyanarayana (1993) and
Gessesse and Gashe (1997) in different bacteria.

Section B
OPTIMIZATION OF CONDITIONS FOR THE PRODUCTION OF ALKALINE PROTEASE
BY BACILLUS PUMlLUS K242 BY THE SOLID STATE FERMENTATION METHOD
MATERIALS AND METHODS
Various factors influencing the production of alkaline protease by
Bacilluspumilus K 242, by solid state fermentation were studied. Except for
the alterations or modifications described under each experiment, the
procedures followed for performing SSF were same as in the preliminaty SSF
studies described under section B of chapter 3.
Solid substrate
Suitability of different commercially available substrates such as wheat
bran, rice bran, green gram bran, black gram bran, coconut oil cake and
ground nut oil cake for use in SSF was studied. SSF was performed using
these substrates in place of wheat bran and the yield of alkaline protease was
determined.
Particle size
Wheat bran which was found to be the most suitable commercially
available solid substrate was sieved and graded based on the particle size.
SSF was performed using the graded wheat bran and the alkaline protease
yield was determined.

Moisture level, Temperature of incubation and Incubation period
Solid substrate media with different moisture levels were prepared
using different volumes of salt solution for moistening wheat bran (particle size
500-710 p) and the initial moisture content of the media was determined.
The yield of alkaline protease was determined after incubation of inoculated
media for different periods at different temperatures.
The effect of pH on alkaline protease production was studied
performing SSF using moistening solution adjusted to different pH (7.510.5).
The moistening solution used contained (dl) dipotassium hydrogen
phosphate, 2; potassium dihydrogen phosphate, 1; magnesium sulphate, 0.1;
calcium chloride, 0.1 and zinc sulphate, 0.01.
Supplementation with carbon sources
Effect of supplementation of solid substrate medium with different
carbon sources on the alkaline protease production was studied. Different
carbon sources were incorporated into the moistening solution so that their
final levels in the moistened solid substrate media were 1, 2 and 3% (w/w). In
addition to the various carbon sources at different concentrations, the
moistening solution contained (d) dipotassium hydrogen phosphate, 2;
potassium dihydrogen phosphate, 1; magnesium sulphate, 0.1; calcium
chloride, 0.1 and zinc sulphate, 0.01. The pH of the moistening solution was
adjusted to 9.0.

Supplementation with nitrogen sources
Effect of supplementation of solid substrate medium with different
nitrogen sources on alkaline protease production was studied. Different
nitrogen sources were incorporated into moistening solution so that their final
levels in the moistened solid substrate media were 1 and 2% (wlw).
In addition to the various nitrogen sources at different concentrations, the
moistening solution contained (911) dipotassium hydrogen phosphate, 2;
potassium dihydrogen phosphate, 1; magnesium sulphate, 0.1; calcium
chloride, 0.1; zinc sulphate, 0.01 and glucose at a level so as to get the final
concentration of 2% (wlw) in the moistened substrate. pH of the moistening
solution was adjusted to 9.0.
Supplementation with sodium chloride
Effect of supplementing the moistening solution with different
concentrations of sodium chloride, on alkaline protease production was
studied. The moistening solution used contained (gA) dipotassium hydrogen
phosphate, 2; potassium dihydrogen phosphate, 1; magnesium sulphate, 0.1;
calcium chloride, 0.1; zinc sulphate, 0.01, glucose at a level so as to get the
final concentration of 2% (wlw) in the moistened substrate and the different
concentrations (0.1,0.2,0.5 and 1% w/v) of sodium chloride.
Age of inoculum
Effect of age of inoculum on alkaline protease production was studied
using inocula aged 24,48, 72 and 96 h.

Size of inoculum
Effect of size of inoculum on alkaline protease production was studied
vaying the inoculum Levels from 2 to 12% v/w of the moistened substrate.
Medium volume:Flask volume
In order to study the effect of varying the ratio medium vo1ume:flask
volume on the yield, SSF was performed with the different volumes of
moistened medium taken in 250 ml Erlenmeyer flasks.
RESULTS
Effect of different solid substrates
Results of studies on the alkaline protease production by Bacillus
pumilus K 242 by SSF using different commercially available substrates are
given in Table 17.
Table 17
Alkaline protease production by Bacilluspumilus K 242 by SSF, using different commercially available substrates
I Substrates Alkaline protease production (u/g DBB * SEM) ] Wheat bran 50.60 + 2.85'
Rice bran 28.38 + 2.27
Green gram bran 37.72 + 1.03
Black gram bran 28.31 _+ 2.37
Coconut oil cake 8.46 * 0.39
Groundnut oil cake 15.66 * 1.07 'As obtained in the earlier experiment.
Of the various substrates tested wheat bran was found to be the best.
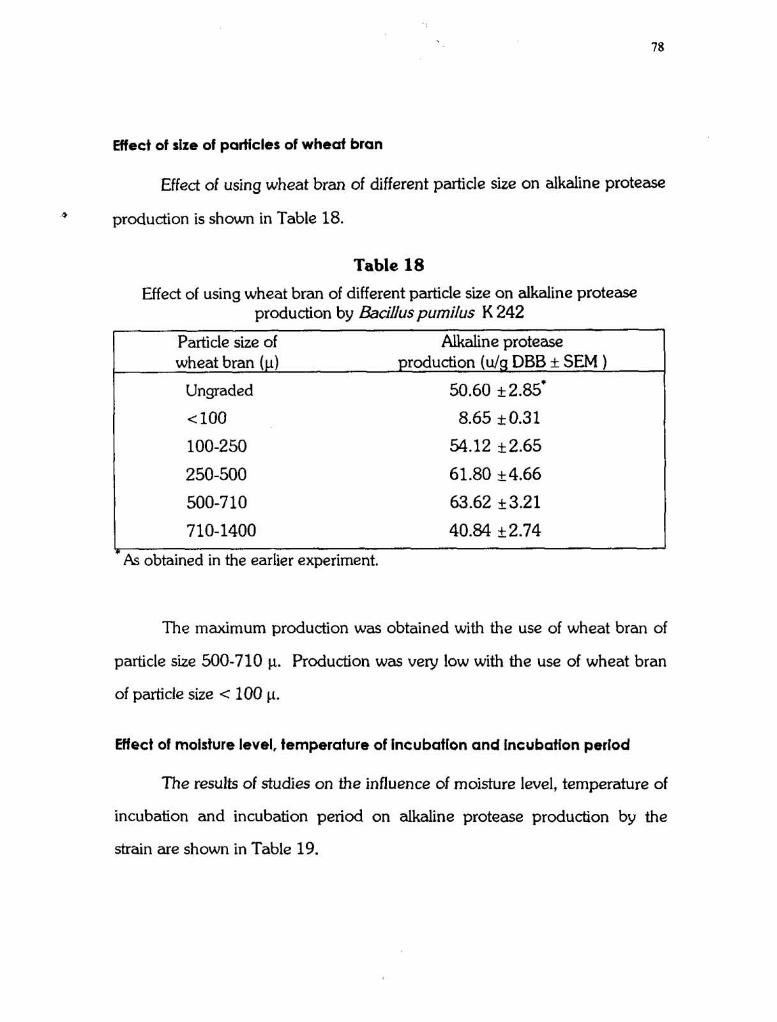
Effect of size of particles of wheat bran
Effed of using wheat bran of different particle size on alkaline protease
1 production is shown in Table 18.
Table 18 Effed of using wheat bran of different particle size on alkaline protease
production by Bacilluspumilus K 242
Particle size of Alkaline protease wheat bran (p) production (ulg DBB * SEM )
Ungraded 50.60 k2.85'
< 100 8.65 k 0.31
100-250 54.12 k2.65
-As obtained in the earlier experiment.
The maximum production was obtained with the use of wheat bran of
particle size 500-710 p. Production was very low with the use of wheat bran
of particle size < 100 p.
Effect of moisture level, temperature of incubation and incubation period
The results of studies on the influence of moisture level, temperature of
incubation and incubation period on alkaline protease production by the
strain are shown in Table 19.

Table 19 Effect of moisture level, temperature of incubation and incubation period on
alkaline protease production by Bacilluspumilus K 242
Percentage moisture level*
60.8
(1:1.3)
64
(1:1.5)
70
(1:2)
Temperature of incubation (OC)
* Wheat bran:moistening solution ratios are given in brackets.
30
37
45
30
37
45
30
37
45
Alkaline protease production (u/g DBB + SEM) when incubated for different periods
28.18 + 0.83
41.15 + 2.45
35.24 + 2.51
37.26 + 2.80
49.68 + 3.12
42.80 + 3.24
40.01 + 3.68
55.40 + 3.72
38.46 + 2.09
96 h 48 h 72 h
43.18 + 2.53
55.54 + 3.44
40.80 + 2.92
48.88 + 3.88
65.02 + 4.87
47.16 + 3.53
45.65 + 0.90
62.14 + 3.10
44.82 + 2.12
51.10 + 3.98
48.32 + 3.90
30.73 k 3.08
58.14 + 4.05
59.84 + 2.83
34.68 + 1.70
46.57 + 1.34
51.22 k 3.66
33.43 + 2.34

The highest production was seen in solid substrate medium with 64%
initial moisture level, after incubation for 72 h at 37°C.
Effect of pH of moistening solution
The effect of pH of the moistening solution on alkaline protease
production is shown in Figure 4.
pH of moistening solution
Figure 4. Effect of pH of moistening solution on alkaline protease production by Bacil/uspumilus K 242
The highest production was obtained when the moistening solution of
pH 9.0 was used.
Effect of extra carbon sources
Effect of supplementation of the wheat bran medium with different
carbon sources, on alkaline protease production is shown in Table 20.
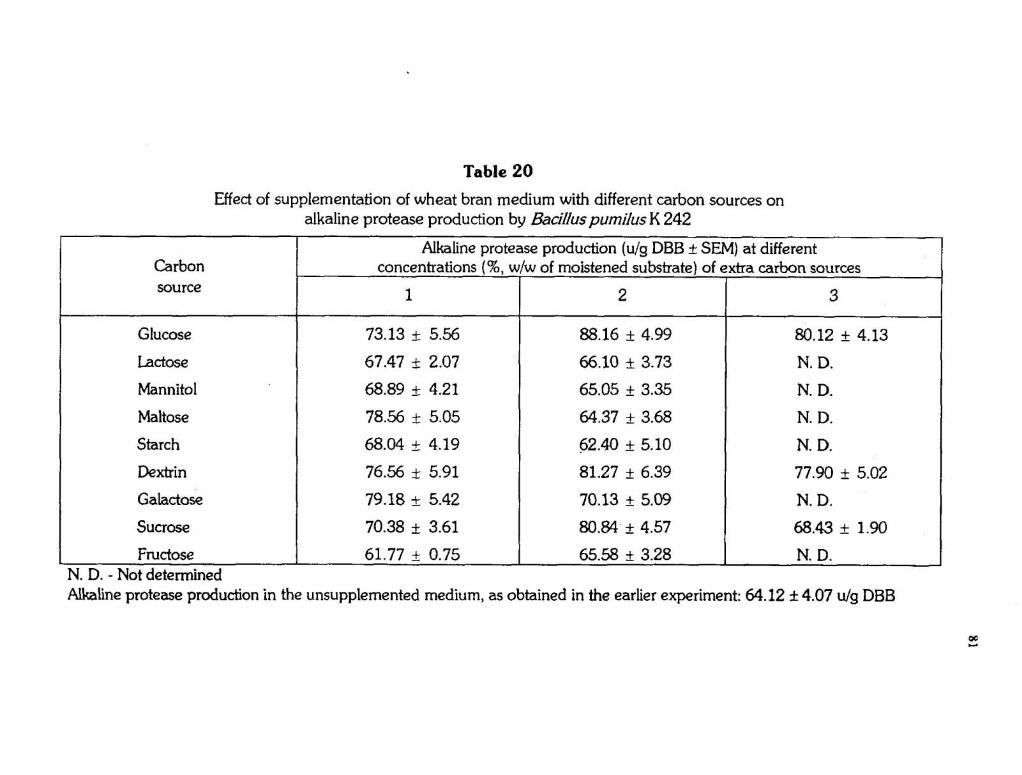
Table 20
Glucose
Lactose
Mannitol
Maltose
Starch
Dextrin
Galactose
Sucrose
Effect of supplementation of wheat bran medium with different carbon sources on alkaline protease production by Bacillus pumilus K 242
Fructose 61.77 + 0.75 65.58 + 3.28 I N. D. - Not determined
Carbon source
Alkaline protease production in the unsupplemented medium, as obtained in the earlier experiment: 64.12 f 4.07 u/g DBB
Alkaline protease production (u/g DBB 4 SEM) at different concentrations (%, wlw of moistened substrate) of extra carbon sources
1 2 3
A considerable increase in production could be noticed by
supplementation of wheat bran medium with many of the carbon sources
tested especially when used at their optimal levels. Carbon sources such as
glucose, dextrin and sucrose at their optimal level of 2% (w/w of the
moistened substrate) were enhancing the production considerably. Maltose
and galactose when used at a level of 1% were also enhancing the
production. The other carbon sources when tested upto a level of 2% were
having little or no effect on the production. Of the different carbon source
supplements tested, glucose at 2% w/w of the moistened substrate was found
to be the best.
Effect of extra nitrogen sources
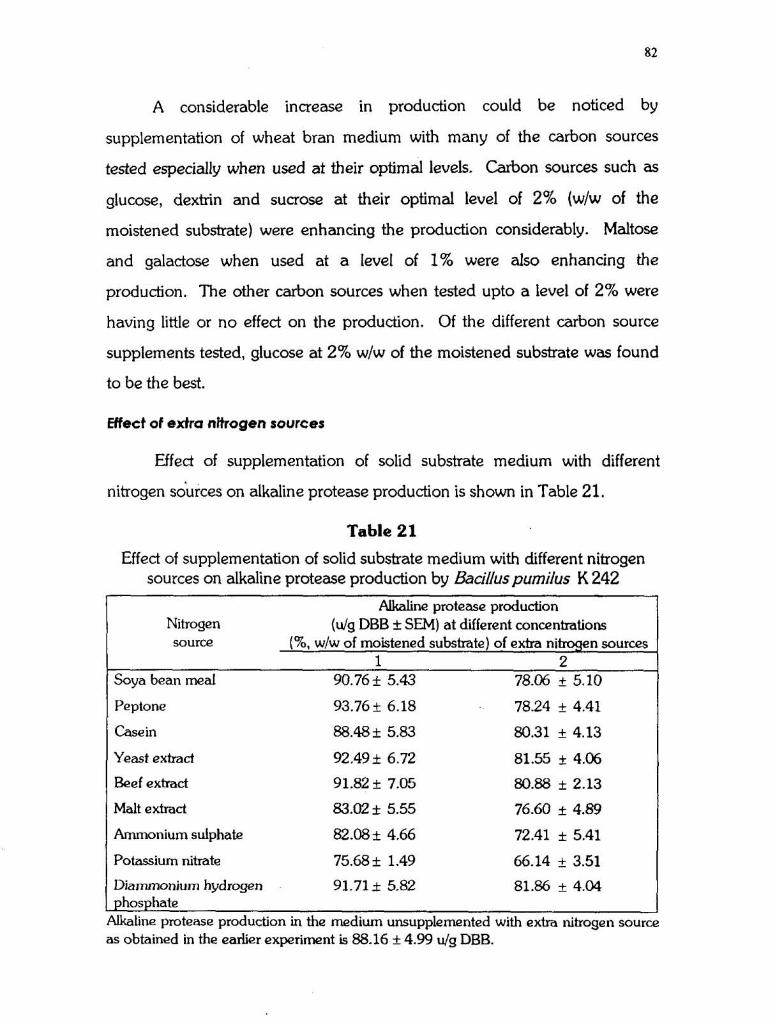
Effect of supplementation of solid substrate medium with different
nitrogen sources on alkaline protease production is shown in Table 21.
Table 21
Effect of supplementation of solid substrate medium with different nitrogen sources on alkaline protease production by Bacilluspurnilus K 242
Alkaline protease production Nitrogen (ulg DBB + SEM) at different concentrations source (%, W/W of moistened substrate) of extra nitrogen sources
1 2 Soya bean meal 90.76 1t 5.43 78.06 + 5.10
Peptone 93.76+ 6.18 78.24 + 4.41
Casein 88.48+ 5.83 80.31 it 4.13
Yeast extract
Beef extract
Malt extract
Ammonium sulphate 82.08 + 4.66 72.41 i 5.41
Potassium nitrate 75.68 + 1.49 66.14 + 3.51
Diammoniurn hydrogen 91.71 i 5.82 81.86 + 4.04 I phosphate Alkaline protease production in the medium unsupplemented with extra nitrogen source - as obtained in the earlier experiment is 88.16 + 4.99 ulg DBB.

None of the nitrogen sources tested was showing an enhancing effect
on the production. At 1% (w/w of moistened substrate) level, all the nitrogen
sources except potassium nitrate were having little or no effect on the
production. At 2% level all the nitrogen sources were inhibitoty.
Effect of sodium chloride
Effects of supplementation of moistening solution with sodium chloride
at different concentrations are shown in Table 22.
Table 22
Effect of incorporating sodium chloride at different concentrations into the moistening solution, on alkaline protease production
by BaciIIus pumilus K 242
Concentration (%, w/v) of Alkaline protease sodium chloride in the
moistening solution production (u/g DBB * SEM)
Nil 87.03 k3.98
0.10 92.07 k3.80
0.20 94.73 + 5.03
0.50 100.51 k5.21
1.00 87.38 k6.19
Incorporation of sodium chloride into the moistening solution was
found to be enhancing the production slightly. The optimum concentration of
sodium chloride for the production was 0.5% (wlv).

Effect of age of lnoculurn
Results of study on the effect of age of inoculum on alkaline protease
production is given in Table 23.
Table 23
Effect of age of inoculum on alkaline protease production by Bacilus pumilus K 242
- Age of Alkaline protease
inoculum (h) production (u/g DBB rt SEM) 24 98.34 k 7.06
48 100.37 k 6.52
72 105.65 k 6.41
96 96.71 k4.29
The age of inoculum was found to be having little or no effect on the
production.
Effect of size of inoculurn
The effect of size of inoculum on the enzyme production is shown in
Table 24.
Table 24
Effect of inoculum size on alkaline protease production by Bacillus pumilus K 242
Sue of inoculum (%, v/w of Alkaline protease 7
moistened substrate) production (ulg DBB * SEM) 2 104.17 rt6.22
4 102.43 k 7.35
8 107.04 k5.84
12 95.78 k5.50
The inoculum at a level of 2-8% v/w of moistened substrate was found
to be optimum for obtaining the high yield.
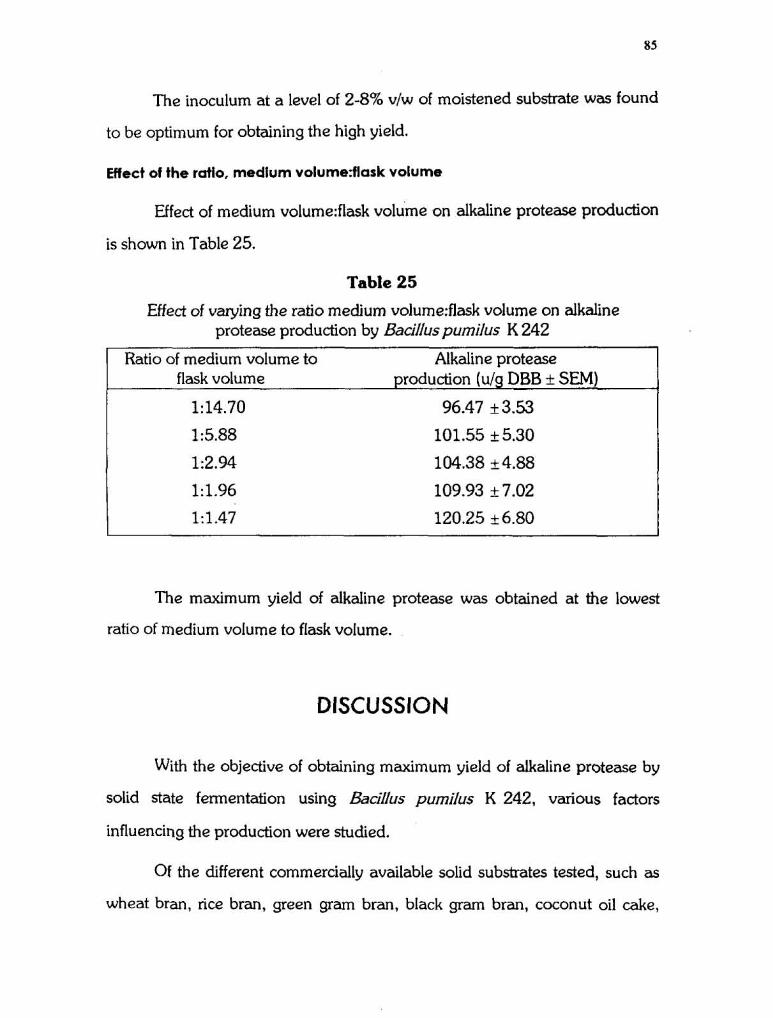
Effect of the ratio, medium volume:flask volume
Effect of medium volume:flask volume on alkaline protease production
is shown in Table 25.
Table 25
Effect of varying the ratio medium volume:flask volume on alkaline protease production by Bacilluspumilus K 242
Ratio of medium volume to Alkaline protease flask volume production (u/g DBB + SEM)
1:14.70 96.47 k3.53
1:5.88 101.55 + 5.30
1:2.94 104.38 k4.88
1:1.96 109.93 + 7.02
1:1.47 120.25 k6.80
The maximum yield of alkaline protease was obtained at the lowest
ratio of medium volume to flask volume.
DISCUSSION
With the objective of obtaining maximum yield of alkaline protease by
solid state fermentation using Bacillus pumilus K 242, various factors
influencing the production were studied.
Of the different commercially available solid substrates tested, such as
wheat bran, rice bran, green gram bran, black gram bran, coconut oil cake,

and groundnut oil cake, wheat bran was found to be the most suitable
followed by green gram bran. The production with coconut and groundnut oil
cakes was found to be v e y low. With rice bran and black gram bran the yield
was intermediate. Earlier workers who developed successful bacterial SSF
systems for the production of proteases, also have reported the use of wheat
bran as the substrate for solid state fermentation (Chakraborty and Srinivasan,
1993; George et a/., 1995; Sen, 1995).
Wheat bran with different particle sizes were tested for the suitability for
SSF process. The highest production was obtained when the wheat bran with
particle size 500-710 p was used. The wheat bran with particle sue 250-500 p
was also good. The production was v e y low with the use of bran of particle
size below 100 p. The difference in production with the use of different grades
of wheat bran can be due to the differences in their nutritional value, porosity,
moisture holding capacity etc. Bran of smallest particle size though
nutritionally better often cause stickiness of the medium after autoclaving.
This can in turn lead to the poor growth of the bacterium resulting in the low
yield. Wheat bran of largest particle size though do not cause stickiness after
autoclaving may not have much nutritional value and moisture holding
capacity. The wheat bran of moderate particle sue (500-710 p) can be
supposed to be satisfying the growth requirements by the strain.
Moisture content of the medium, incubation time and incubation
temperature are three important factors that can influence the enzyme
production by SSF. Since the influences of these fadors are interdependent
the effects of these fadors were studied simultaneously. The maximum
alkaline protease production by Baci/us pumilus K 242 was obtained in the

medium with 64% initial moisture level, after incubation for 72 h at 37°C.
Production in the medium with 70% moisture level after incubation for 72 h at
37°C was also good. At all temperatures, i.e., 30, 37 and 45"C, the optimum
moisture level for the production was found to be 64%. Moisture level is a
factor determining the level of available water, solubiliiation of nutrients, gas
exchange and oxygen transfer in the solid substrate medium. Since different
bacteria are requiring these factors at different levels, their moisture level
requirements will also be different. Chakmbotty and Srinivasan (1993) have
reported that Pseudomonas sp. B45 was producing alkaline protease
maximally in medium with initial moisture level 74%, incubated at 37°C for
120 h. George ef a/. (1995) who studied protease production by Bacillus
arnyloliquefaciens ATCC 23844 obtained the maximum yield when wheat
bran moistened at 1:2 ratio was used. The incubation temperature and period
were 37°C and 24 h respectively. In the present study the incubation period
required for obtaining the maximum yield at 37 and 45°C was 72 h. A longer
incubation period, i.e., 96 h was required for obtaining the maximum yield
when incubated at 30°C.
Although pH is a critical factor that can influence the enzyme
production, monitoring and control of pH during the SSF process is not
usually attempted (Lonsane et d, 1985). Good buffering capacity of the
sub&ates used in SSF generally help in eliminating the need for pH control
during the process. This advantage was exploited in this study also.
No attempts were made to control the pH during the fermentation process.
The effect of pH on production was studied by using the moistening solutions

with different pH. The maximum production was obtained with the use of
moistening solution having pH 9.0.
The effect of supplementing solid substrate medium with different
carbon and nitrogen sources were studied. A considerable increase in
production could be noticed by supplementation of wheat bran medium with
many of the carbon sources tested, especially at their optimal levels. While
the optimal level of glucose, dextrin and sucrose was 2% (wlw of the
moistened substrate), it was 1% for maltose and galactose. Of the different
carbon sources tested glucose at 2% (wlw of the moistened substrate) level
was found to be the best as supplement. These observations on the effects of
extra carbon sources were different from the observations reported by earlier
workers in this field. Chakraborty and Srinivasan (1993), George et a/. (1995)
and Sen (1995) who studied on the bacterial SSF systems for protease
production have reported the inability of glucose and some other carbon
sources to improve the yield. The differences in the metabolic characters of
different bacteria may not be the sole reason for such a difference in
observations. It has to be taken into account that the methodology followed in
the present study for determining the effect of carbon source was different
from that followed in the earlier reports. In this study, for determining the
effect of extra carbon sources, they were incorporated into moistening
solution, instead of adding them directly into wheat bran. Due to this reason
pH could be easily adjusted thereafter. On the other hand, if these carbon
sources were added directly into wheat bran, there would not have been such
a facility for the adjustment of pH. In such cases the addition of extra carbon
sources into wheat bran may possibly result in the solid substrate medium with

unfavourable pH. The SSF performed with media having unfavoumble pH
can definitely result in the decreased enzyme production. But in this study,
such a situation could be avoided by adjusting the pH of the moistening
solution after the addition of carbon sources.
Of the different nitrogen sources tested none was found to be
enhancing the production. Though most of them were having little or no
effed on production at 1% (w/w of the moistened substrate) level, they were
inhibitory at 2% level. Potassium nitrate was inhibitory even at the 1% level.
Similar observations have been reported by earlier workers also. Chakmbotty
and Srinivasan (1993) who studied the alkaline protease production by
B. amyloliquefaciens ATCC 23844, have reported a decrease in production
on supplementation of wheat bran with extra nitrogen sources. Sen (1995)
has reported that supplementation of wheat bran medium with extra nitrogen
sources was ineffective in improving the yield of alkaline protease by Bacillus
licheniformis S40. Results of the present study indicate that the nitrogen
sources in the unsupplemented medium itself is sufficient to support the
growth and to induce the enzyme production by the strain. This again points
to the universal suitability of wheat bran as the substrate for the SSF
processes.
The effed of supplementing the moistening solution with different
concentrations of sodium chloride, on alkaline protease production was
studied. Results indicate that sodium chloride can be incorporated into the
moistening solution at a level 0.5% (wlv), to improve the yield of alkaline
protease. Such an observation has not been reported by the earlier workers in
this field.

The effects of both age and sue of inoculum on the protease
production were studied. The age of inoculum was found to be having liffle or
no effect on production. The size of inoculum for obtaining the maximum
yield was 23% (vlw of moistened substrate). At a higher level, i.e., 12% there
was a slight decrease in production. The size of inoculum used for the alkaline
protease production of Bacillus lichenifomis 540 has been reported to be
10% (vlw) (Sen, 1995).
The effect of varying the ratio, medium volume:flask volume was also
studied. It was interesting to observe that the activity was increasing with the
use of larger volumes of moistened medium in the conical flask. The highest
activity was obtained at the lowest ratio tested, i.e., 1:1.47. At this ratio the
moistened solid substrate medium in the flask was having a height of 4 cm
and only a liffle air space was left over it. Chakraborty and Srinivasan (1993)
who studied the effect of medium volume:flask volume on alkaline protease
production by Pseudomonas sp. B45, have observed that there was a
decrease in production with the decrease in the ratio. This difference in the
obse~ations can be ascribed to the differences in the physiological and
metabolic characters of the two bacteria. The comparatively higher production
obtained by BaciJJus pumilus K 242 on using thicker layer of'solid substrate
medium, is of much techno-economic importance. Results indicate that SSF
for the large scale production of alkaline protease by B. pumilus K 242 can be
performed in containers leaving minimum air space over the medium. This
can facilitate the maximum utilization of space in the containers and will be
helpful in reducing the cost of production to some extent.

As a result of optimization of conditions, the alkaline protease
production by Bacilluspumilus K 242 could be increased more than two-fold,
resulting in the production of 120 u of alkaline protease per gram dry bacterial
bran. A comparison of the yield obtained by B. pumilus K 242 with the yields
reported in other bacterial SSF systems is difficult, because the assay
procedures and the unit activity definitions followed in these studies were
different. However taking into account the high yielding nature of B. pumilus
K 242 and the ease and economy for the enzyme production, the strain can
be suggested as suitable for alkaline protease production in a large scale. With
the use of specially designed fennentors under better process control, it may
be possible to improve the yield further.













![Versatility of microbial proteases · Extracellular proteases carry out protein hydrolysis in fermented media and enable the cell to absorb and utilize hydrolytic products [12]. Alkaline](https://img.pdfslide.us/doc/110x75/5fd6f3fcc465456e8b0d4ec5/versatility-of-microbial-proteases-extracellular-proteases-carry-out-protein-hydrolysis.jpg)















