Embed Size (px)
Citation preview

Therapeutics, Targets, and Chemical Biology
Oncolytic Adenoviral Delivery of anEGFR-Targeting T-cell Engager ImprovesAntitumor EfficacyCarlos Alberto Fajardo1, Sonia Guedan2, Luis Alfonso Rojas1, Rafael Moreno1,Marcel Arias-Badia1, Jana de Sostoa1, Carl H. June2, and Ramon Alemany1
Abstract
Antiviral immune responses present a major hurdle to theefficacious use of oncolytic adenoviruses as cancer treatments.Despite the existence of a highly immunosuppressive tumorenvironment, adenovirus-infected cells can nonetheless be effi-ciently cleared by infiltrating cytotoxic T lymphocytes (CTL)without compromising tumor burden. In this study, we testedthe hypothesis that tumor-infiltrating T cells could be moreeffectively activated and redirected by oncolytic adenovirusesthat were armed with bispecific T-cell–engager (BiTE) antibo-dies. The oncolytic adenovirus ICOVIR-15K was engineered toexpress an EGFR-targeting BiTE (cBiTE) antibody under thecontrol of the major late promoter, leading to generation ofICOVIR-15K-cBiTE, which retained its oncolytic properties invitro. cBiTE expression and secretion was detected in super-
natants from ICOVIR-15K-cBiTE–infected cells, and the secret-ed BiTEs bound specifically to both CD3þ and EGFRþ cells. Incell coculture assays, ICOVIR-15K-cBiTE–mediated oncolysisresulted in robust T-cell activation, proliferation, and bystandercell-mediated cytotoxicity. Notably, intratumoral injection ofthis cBiTE-expressing adenovirus increased the persistence andaccumulation of tumor-infiltrating T cells in vivo, comparedwith the parental virus lacking such effects. Moreover, in twodistinct tumor xenograft models, combined delivery ofICOVIR-15K-cBiTE with peripheral blood mononuclear cellsor T cells enhanced the antitumor efficacy achieved by theparental counterpart. Overall, our results show how armingoncolytic adenoviruses with BiTE can overcome key limitationsin oncolytic virotherapy. Cancer Res; 77(8); 2052–63. �2017 AACR.
IntroductionOncolytic adenoviruses have gained considerable attention as
anticancer agents. Their attractiveness relies on a multimodalmechanism to kill cancer cells that include direct oncolysis,bystander effect when armed with therapeutic transgenes, andimmunogenic cell death–mediated antitumor immune responses(1). Clinical trials have demonstrated the safety and potential ofoncolytic viruses (2, 3) and highlighted the importance of theimmune system for their success in cancer patients. Tumorsdevelop numerous mechanisms to evade antitumor immuneresponses (4), but oncolytic viruses can revert such immunesuppression (5). However, virus replication in the tumor alsotriggers a potent immune response against viral epitopes, whichhas been shown to be dominant over tumor-associated epitopes(6, 7). Hence, oncolytic virus-infected cells are often cleared fromthe tumor microenvironment by infiltrating virus-specific cyto-
toxic T lymphocytes (CTL) without altering the tumor burden (8).Many efforts to improve oncolytic viruses rely on encodingimmunostimulatory molecules (e.g., GM-CSF, IL12, or CD40L)in the virus genome (9, 10). However, these strategies do notaddress either the virus immunodominance or the infiltratingantivirus CTL.
Bispecific T-cell engagers (BiTE) are novel immunotherapeuticmolecules composed of two single-chain antibodies (scFV) con-nected through a flexible peptide linker. One of the scFV is specificfor a tumor-associated antigen on target cells, whereas the secondscFV is specific for the CD3 T-cell coreceptor. This format allowsthe transient binding of BiTE molecules to T cells and target cellssimultaneously, leading to T-cell activation and specific target celllysis (11). The potential of BiTEs has been demonstrated with theCD19-targeting BiTE blinatumomab, approvedby the FDA for thetreatment of acute lymphoblastic leukemia (12). Other BiTEs areunder investigation for the treatment of solid tumors (13). Addi-tionally, an oncolytic vaccinia virus armed with a BiTE targetingthe EphA2 receptor has shown improved bystander effect andincreased antitumor efficacy compared with its parental counter-part (14).
Among oncolytic viruses, adenoviruses hold features thatmakethem good virotherapy candidates. Adenovirus replication incancer cells can be tightly regulated, and placing transgenes underthe control of the major late promoter secures replication-depen-dent expression (15). Furthermore, oncolytic adenoviruses havefavorable toxicity profiles after systemic administration in cancerpatients, and their potential is exemplified by the virus DNX-2401, which has received fast-track status and orphan drugdesignation by the FDA for the treatment of malignant glioma(16). We hypothesize that oncolytic adenoviruses are good
1ProCure Program, IDIBELL-Institut Catal�a d'Oncologia, L'Hospitalet de Llobre-gat, Spain. 2Abramson Cancer Center and the Department of Pathology andLaboratory Medicine, Perelman School of Medicine, University of Pennsylvania,Philadelphia, Pennsylvania.
Note: Supplementary data for this article are available at Cancer ResearchOnline (http://cancerres.aacrjournals.org/).
Corresponding Author: Ramon Alemany, IDIBELL-Institut Catal�a d'Oncologia,Duran i Reynals Oncology Hospital, Av Gran Via de l'Hospitalet 199-203,L'Hospitalet de Llobregat, 08907 Barcelona, Spain. Phone: 349-3260-7462;Fax: 349-3260-7466; E-mail: [email protected]
doi: 10.1158/0008-5472.CAN-16-1708
�2017 American Association for Cancer Research.
CancerResearch
Cancer Res; 77(8) April 15, 20172052
on March 1, 2021. © 2017 American Association for Cancer Research. cancerres.aacrjournals.org Downloaded from
Published OnlineFirst January 31, 2017; DOI: 10.1158/0008-5472.CAN-16-1708

candidates to deliver BiTEs to specifically redirect immuneresponses toward cancer cells.
In this study, we have engineered an oncolytic adenovirus tosecrete an EGFR-targeting BiTE upon viral replication in cancercells.We show that theBiTE-expressing adenovirus induces robustand specific T-cell activation and proliferation upon infection ofcancer cells in vitro and in vivo. This specific T-cell activationtranslates into the redirected killing of cancer cells, enhancingthe antitumor efficacy of the virus in mouse xenograft models ofcancer. These results demonstrate that BiTE-expressing oncolyticadenoviruses have favorable properties that can overcome majorlimitations in oncolytic virotherapy.
Materials and MethodsCell lines
Cancer cell lines A431 (epidermoid carcinoma), MDA-MB-453 (breast cancer), A549 (human lung carcinoma), HCT116(colorectal carcinoma), FaDu (pharynx squamous cell carcino-ma), Jurkat (T cell leukemia), and HEK-293 (human embryonickidney) were obtained and authenticated by STR profiling bythe ATCC. The A431-GFPLuc cell line was generated by sortingof A431 cells transduced with a lentiviral vector encodingGFP and luciferase.
Preparation of peripheral blood mononuclear and T cellsAll experiments were approved by the ethics committees of the
University Hospital of Bellvitge and the Blood and Tissue Bank(BST) fromCatalonia. Blood samples were obtained from the BSTfrom Catalonia and the Human Immunology Core of the Uni-versity of Pennsylvania. Peripheral blood mononuclear cells(PBMC) of healthy donors were isolated by ficoll density gradientcentrifugation. T cells were negatively isolated using the Rosette-Sep Human T Cell Enrichment Cocktail (STEMCELL Technolo-gies). For stimulation, T cells were cultured with CD3/CD28-activating Dynabeads (Thermo Fisher Scientific) at a bead-to-cellratio of 3. For biodistribution experiments, T cells were trans-duced with a lentiviral vector expressing the click beetle greenluciferase [multiplicity of infection (MOI) ¼ 5] 24 hours afteractivation (17). Preactivated T cellswere expanded andhandled aspreviously described (18).
cBiTE and adenovirusesThe anti-EGFR scFV (C225) was derived from the cetuximab
sequence obtained from publicly available sources. The anti-CD3scFV sequence of the Blinatumomab BiTE was obtained frompatent application WO2004106381. The C225 and anti-CD3variable regions were connected by a (G4S1)3 and a (G2S1)4GGlinker, respectively, and both scFV were connected to each otherby a GGGS flexible linker. The cBiTE was arranged VLC225–VHC225–VHCD3–VLCD3 and included the peptide signal from themouse Ig heavy chain and a FLAG tag at the N- and C-terminus ofthe protein, respectively. The cBiTE construct was optimized forhuman codon usage and synthesized by Genscript (Genscript).Viruses ICOVIR-15K (abbreviated ICO15K) and AdTLRGDKhavebeen previously described (19, 20). The cBiTE gene was incorpo-rated into the ICO15K genomeby recombineering in bacteria (21,22). ICO15K-cBiTE was rescued after transfection of the resultingplasmid into 293 cells. Replicating viruses were propagated inA549 cells and double purified by cesium chloride gradientcentrifugation. Functional [transducing units (TU)/mL] andphys-
ical [viral particles (VP)/mL] titers of purified viruses were deter-mined by anti-hexon staining (23) and by optical absorbance at260 nm (24), respectively.
Antibodies and flow cytometryFlow cytometry was performed with a Gallios cytometer (Beck-
man Coulter). Expression of EGFR was detected with the mousemonoclonal antibody Clone 528 (Merck Millipore) or its IgG2a
isotype control (Santa Cruz Biotechnology), followed by incuba-tion with an Alexa Fluor 488–coupled goat anti-mouse IgGantibody (Thermo Fisher Scientific). For phenotyping of T-cellsubpopulations, the followingmonoclonal antibodies coupled todifferent fluorochromes were used: CD3, CD4, CD8, CD69, andCD25 (Biolegend). Flow cytometry data were analyzed with theFlowJo software v7.6.5 (Tree Star).
Production of supernatantsA549 cells (1 � 107) were infected with ICO15K or ICO15K-
cBiTE (MOI¼ 20) and supernatants were harvested 72 hours afterinfection. For binding assays with effector cells, supernatants wereconcentrated (approximately 20�) with Amicon Ultra-15 filterunits with amolecular weight cutoff of 30 kDa (MerckMillipore).Supernatants from uninfected cells were used as a negative mockcontrol.
Binding assaysTarget (2� 105) or effector (1� 105) cells were incubated with
the supernatants for 1 hour on ice. Cells were stained by using themonoclonal M2 anti-FLAG antibody (Sigma Aldrich) or its cor-responding IgG1 isotype control (Santa Cruz Biotechnology) asprimary antibodies and goat anti-mouse IgG as secondary anti-body (Thermo Fisher Scientific).
In vitro coculture experimentsAll coculture experiments were performed as follows, unless
indicated otherwise. A total of 3 � 104 target cells/well and 1.5 �105 PBMCs/well (E:T¼ 5) were seeded in 96-well plates in 100-mLmedium. Toassess T-cell activationby the supernatants fromvirus-infected cells, cocultures were mixed with 100 mL of the super-natants and incubated for the indicated times. For the oncolysis-mediated T-cell assays, cocultures were infected with ICO15K orICO15K-cBiTE (MOI¼20). For T-cell activation assays, cocultureswere incubated for 48 hours or 5 days as described above and thencells were stained for cell viability with LIVE/DEAD (ThermoFisher Scientific) followed by incubation with antibodies specificfor CD8, CD4, and CD69 or CD25. For cytokine productionassays, supernatants were obtained 48 hours after coculture andassessed for cytokines with the human IFNg and TNFa ELISA(Peprotech). For proliferation assays, PBMCs were labeled with 1mmol/L CFSE (Sigma Aldrich) and cocultured as described abovefor 6 or 7 days. Cells were then stained for cell viability with LIVE/DEAD followed by incubation with antibodies for CD4 and CD8.
Virus- and cell-mediated cytotoxicity assaysVirus cytotoxicity assays were performed as previously
described (25). The inhibitory concentration 50 (IC50) was cal-culated with GraphPad Prism v6.02 (GraphPad Software Inc.) bya dose–response nonlinear regression with a variable slope.
CFSE-labeled target cells (A431 or MDA-MB-453, 1.5 � 104)were cocultured with 1.5 � 105 PBMCs (E:T ¼ 10) in 96-well
T-cell Engager-Armed Oncolytic Adenovirus
www.aacrjournals.org Cancer Res; 77(8) April 15, 2017 2053
on March 1, 2021. © 2017 American Association for Cancer Research. cancerres.aacrjournals.org Downloaded from
Published OnlineFirst January 31, 2017; DOI: 10.1158/0008-5472.CAN-16-1708

plates. Cocultures weremixedwith the supernatants (100 mL) andincubated for 24 hours. Cocultures were trypsinized and stainedwith 10 mg/mL 7-amino-actinomycin D (7-AAD; Enzo LifeSciences). Cells were analyzed by flow cytometry, and the per-centage of CFSEþ/7-AADþ cells was determined.
For bystander killing assays, A549 cells in suspension wereinfected with ICO15K or ICO15K-cBiTE (MOI ¼ 20) for 4hours. Excess virus was washed with PBS. 2.5 � 104 virus-infected A549 cells per well, and 2.5 � 104 A431-GFPLuc perwell were cocultured with 1.5 � 105 PBMCs (E:T ¼ 6 withrespect to A431-GFPLuc cells) in 96-well plates. Cocultureswithout PBMCs were used as a control for virus-mediated cellcytotoxicity. Five days after infection, cocultures were stainedwith 7-AAD as described above. Cells were analyzed by flowcytometry and the absolute number of live A431-GFPLuc wasdetermined using CountBright Absolute Counting Beads(Thermo Fisher Scientific). The percentage of live A431-GFPLucwas obtained by normalizing the absolute count of the samplesto a culture of untreated A431-GFPLuc cells.
Oncolysis-mediated enhanced cell-mediated cytotoxicity wasassessed with the xCELLigence Real-Time Cell Analyzer (ACEABiosciences). A total of 1 � 104 target cells per well (A549 orHCT116) were seeded and infected with ICO15K or ICO15K-cBiTE at an MOI of 1. After overnight cell adherence, effector cells(preactivated T cells)were added (E:T¼5).Cell index (i.e., relativecell impedance) values were monitored every 20 minutes for 120hours and normalized to the maximal cell index value immedi-ately prior to effector cell plating.
In vivo studiesAll animal experiments were approved by the Ethics Com-
mittee for Animal Experimentation from the BiomedicalResearch Institute of Bellvitge (IDIBELL). Animals werehoused in the IDIBELL Animal Core Facility (AAALAC Unit1155). Subcutaneous A549 and HCT116 were established byinjecting 5 � 106 and 2.5 � 106 cells, respectively, into bothflanks of 8-week-old female SCID/beige mice (Envigo). Whentumors reached �100 mm3, mice were randomized andtreated as described below.
For the T-cell biodistribution study, tumors were injectedwith 2 � 109 VP of the indicated viruses or PBS. After 5 days,mice received an intravenous injection of 1 � 107 cell prepara-tions containing 59% preactivated CBG luciferase-expressing Tcells, followed by an intraperitoneal dose of 1500 IU IL2(Peprotech). Mice were imaged daily until day 9 with the IVISLumina XRMS Imaging System (PerkinElmer) after adminis-tering intraperitoneal injections of a 15 mg/mL D-luciferinfirefly potassium salt solution (Biosynth AG). Tumor radiancewas measured by drawing a region of interest around the tumorcontour.
For antitumor efficacy, A549 tumors were injected with 2 �109 VP of the indicated viruses or PBS. After 4 and 18 days, PBSor 1 � 107 unstimulated human PBMCs were administeredto the mice by intravenous injection. Mice bearing HCT116tumors received an intravenous injection of 1 � 1010 VP of theindicated viruses or PBS. After 4, 8, and 11 days, mice receivedan intravenous injection of 1 � 107 preactivated T cellsfollowed by an intraperitoneal injection of 1,500 IU IL2. Inboth models, tumors were measured every 2 to 4 days, and thevolume was calculated as V (mm3) ¼ p/6 � W2 � L, where Wand L are the width and the length of the tumor, respectively.
Immunohistofluorescence of OCT-embedded sections of A549tumors at the end of the study was used to evaluate theexpression of the E1a protein as described (26).
mRNA extraction and real-time PCRFrozen tumor samples were disrupted using a mortar and
pestle under liquid nitrogen. Approximately 25 mg of tissuewas homogenized with Qiashredder homogenizers, and RNAwas isolated with the RNeasy kit (Qiagen), with DNase Idigestion to remove genomic DNA. RNA (1 mg) was retro-transcribed with the High-Capacity cDNA Reverse Trancriptionkit (Thermo Fisher Scientific).
Real-time analysis was performed using a LightCycler 480Instrument II (Roche) in the presence of SYBR Green I Master(Roche). PCR conditions were: 95�C 10 minutes, 40� cycles of95�C 15 seconds, 60�C 1 minute and 72�C 7 seconds. Hexonprimers were Ad18852 50-CTTCGATGATGCCGCAGTG-3 andAd19047R 50-ATGAACCGCAGCGTCAAACG-30. cBiTE primerswere qBiTEF 50-CGGCGAGAAAGTGACAATGAC-30 and qBiTER50-TTGGTGAGGTGCCACTTTTC-30. Standard curves for cBiTEand hexon were prepared by serial dilutions of known copynumbers of pUC57-cBiTE or purified ICO15K genomes, respec-tively. Non-retrotranscribed RNA samples, equivalent to theamount cDNA loaded in the PCR, were run to discard genomicDNA contamination.
Statistical analysisFor comparisons of two groups, two-tailed unpaired t tests were
used. For comparison of more than two groups, one-way ANOVAwith Tukey post hoc tests was used. Statistical significance wasestablished as P < 0.05. Data are presented as the mean � SD orSEM. All statistical analyses were calculated with the GraphPadPrism software v6.02.
ResultsGeneration and characterization of a BiTE-armed oncolyticadenovirus targeting the EGFR
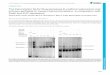
We have reported the generation of ICOVIR-15K (ICO15K),an E1a-D24–based oncolytic adenovirus with palindromic E2Fbinding sites in the E1a promoter and an RGDKmotif replacingthe KKTK heparan sulfate glycosaminoglycan-binding domainin the fiber shaft (19). This virus has shown favorable toxicityprofiles and increased tumor targeting in vivo. We engineeredICO15K to express an EGFR-targeting bispecific T-cell engager(cBiTE) under the control of the adenovirus major later pro-moter (Fig. 1A). This location was chosen to avoid potentialBiTE-mediated interference with virus replication. The virusICO15K-cBiTE was successfully rescued and it retained onco-lytic properties in vitro, despite a decrease in the IC50 valuescompared with its parental counterpart in dose–response cyto-toxicity assays (Fig. 1B).
To evaluate the secretion of the cBiTE by infected cells,flow cytometry–based binding assays were performed byexploiting the FLAG tag encoded in the transgene. For theseassays, a panel of cancer cell lines with varying EGFR expressionlevels, and CD3þ Jurkat and human PBMCs were used (Sup-plementary Fig. S1). cBiTE antibodies were detected only inthe supernatants of ICO15K-cBiTE–infected cells and theyspecifically bound to EGFRþ (A431, A549, HCT116, and FaDu)but not EGFR� (MDA-MB-453) cancer cells (Fig. 2A and
Fajardo et al.
Cancer Res; 77(8) April 15, 2017 Cancer Research2054
on March 1, 2021. © 2017 American Association for Cancer Research. cancerres.aacrjournals.org Downloaded from
Published OnlineFirst January 31, 2017; DOI: 10.1158/0008-5472.CAN-16-1708

Supplementary Fig. S2). cBiTE molecules also bound to CD3þ
Jurkat cells and human PBMCs, and this binding was morepronounced when supernatants were concentrated approxi-mately 20� (Fig. 2B). Furthermore, the binding of the cBiTEto CD4þ and CD8þ T cells within PBMCs was confirmed(Fig. 2C).
Supernatants from ICO15K-cBiTE–infected cells enhance T-cellfunction
We then evaluated the functionality of the secreted cBiTE incoculture experiments of cancer cells with unstimulated humanPBMCs. ICO15K-cBiTE supernatants specifically induced CD8þ
and CD4þ T-cell activation, as indicated by an increase in the
Figure 1.
Generation of a cBiTE-expressing adenovirus and its oncolytic properties in vitro.A, Schematic representation of the genome of the viruses used in the study. L/RITR,left/right inverted terminal repeats; SA, splicing acceptor; K, kozac sequence; pA, polyadenylation signal; S, signal peptide; F, flag tag. B, A549 and HCT116cell lines were incubated with serial dilutions of ICO15K or ICO15K-cBiTE. Six days after infection, cell viability was determined and the IC50 for each virus wascalculated. The mean � SD of triplicates is shown.
T-cell Engager-Armed Oncolytic Adenovirus
www.aacrjournals.org Cancer Res; 77(8) April 15, 2017 2055
on March 1, 2021. © 2017 American Association for Cancer Research. cancerres.aacrjournals.org Downloaded from
Published OnlineFirst January 31, 2017; DOI: 10.1158/0008-5472.CAN-16-1708

expression of the activation markers CD25 and CD69 whenPBMCs were cocultured with EGFRþ cell lines (Fig. 3A). Toconfirm this activation, we assessed cytokine production byPBMCs in coculture assays with a panel of cancer cell lines. IFNgand TNFa were detected at high levels only when ICO15K-cBiTEsupernatants were cocultured with PBMCs and EGFRþ cell lines(Fig. 3B). Importantly, the cBiTE-containing supernatants did notinduce T-cell activation when cocultured alone or with the EGFR-negative cell line MDA-MB-453.
Another important indicator of T-cell activation is their prolif-erative capacity. CD3þ T cells underwent multiple rounds ofproliferation only when cocultured with EGFRþ A431 cells andICO15K-cBiTE supernatants, as evidenced by the dilution of CFSEafter 5 days of incubation (Fig. 3C).
The ultimate goal of BiTE antibodies is to retarget T-cell–mediated cytotoxicity toward cancer cells. To confirm this, weperformed cell-mediated cytotoxicity assays by coculturing A431and MDA-MB-453 CFSE-labeled cells with PBMCs and the dif-ferent supernatants for 24 hours. The coculture of A431 but not ofMDA-MB-453 cells with the cBiTE-containing supernatants andPBMCs led to a significant increase in cell cytotoxicity comparedwith ICO15K and the mock control (Fig. 3D).
ICO15K-cBiTE–mediated oncolysis enhances T-cell functionand induces a T-cell–mediated bystander effect
The experiments described above were performed with super-natants of infected cells that contained the cBiTE. We next wantedto evaluate the cBiTE-expressing virus in a setting thatmore closelyresembles the oncolytic process. For this, cocultures of humanPBMCs with either A549 or HCT116 EGFRþ cell lines wereinfected at an MOI of 20. Cells were incubated for 5 and 7 daysto address activation and proliferation, respectively. These incu-bation times were chosen to allow a full replication cycle of thevirus and the proper actionof the cBiTE. ICO15K-cBiTE–mediatedoncolysis led to an increase in CD25þ and CD69þ T cells and toextensive proliferation of both T-cell subsets only when cancercells were present (Fig. 4A and B).
Another important feature of a secreted BiTE is its bystandereffect. To test this without the interfering cytotoxicity of the virus,we chose the EGFRþ A431-GFPLuc cell line, which expresses lowlevels of the coxsackie- and adenovirus receptor. This cell line ispartly resistant to adenovirus infection and shows low adenovi-rus-mediated cytotoxicity and no cBiTE production at high MOIs(Supplementary Fig. S3). A431-GFPLuc cells were coculturedwithICO15K- or ICO15K-cBiTE–infected A549 cells in the presence or
Figure 2.
cBiTE antibodies are expressed from ICO15K-cBiTE–infected cells and specifically bind to target and effector cells. A and B, A431 and MDA-MB-453 cells (A) orPBMC and Jurkat cells (B) were incubated with the indicated supernatants, and cBiTE binding was detected by flow cytometry using an anti-FLAG antibody.C, Human PBMCs were incubated with concentrated supernatants from uninfected or ICO15K/ICO15K-cBiTE–infected A549 cells. cBiTE binding to CD4þ andCD8þ T cells was detected by flow cytometry.
Fajardo et al.
Cancer Res; 77(8) April 15, 2017 Cancer Research2056
on March 1, 2021. © 2017 American Association for Cancer Research. cancerres.aacrjournals.org Downloaded from
Published OnlineFirst January 31, 2017; DOI: 10.1158/0008-5472.CAN-16-1708

absence of human PBMCs for 5 days. Thus, in this setting, A549cells act as BiTE producers, while A431-GFPLuc cells represent thetargets of the T cells. ICO15K-cBiTE–infected A549 cells induced asignificant decrease in the percentage of live A431-GFPLuc cellscompared with ICO15K-infected cells when cocultured withPBMCs from two different donors (Fig. 4C). This increasedcytotoxicity was dependent on the presence of PBMCs, as ICO15Kand ICO15K-cBiTE induced similar levels of cell death in theabsence of PBMCs.
We then tested whether the infection of cancer cells withICO15K-cBiTE at low MOIs could provide an advantage duringoncolysis over the nonmodified virus when cocultured withpreactivated T cells. Preactivated T cells have a phenotype thatmore closely resembles that of adenovirus-specific CTLs, themainT-cell population expected to infiltrate tumors during oncolysis.We found that preactivated T cells show enhanced cytotoxicpotential compared to na€�ve T cells (Supplementary Fig. S4A)and produce high amounts of proinflammatory cytokines
Figure 3.
cBiTE antibodies expressed from ICO15K-cBiTE–infected cells enhance T-cell functions. PBMCs were cocultured with the indicated cell lines in the presence ofthe different supernatants. A, Forty-eight hours after coculture, the expression levels of CD25 and CD69 on CD4þ and CD8þ T cells were assessed by flowcytometry. Representative results from one of two healthy donors are shown. B, Forty-eight hours after cocultures, supernatants were harvested and cytokineproduction was evaluated by ELISA. Representative results from one of two healthy donors are shown. Bars, mean � SD of triplicates. C, Six days after coculture,the CFSE content (i.e., cell proliferation) on CD3þ T cells was evaluated by flow cytometry. Representative results from one of two healthy donors areshown. D, Twenty-four hours after coculture the percentage of cytotoxicity of target cells (CFSEþ/7AADþ) was assessed by flow cytometry. Representativeresults from one of three healthy donors are shown. Bars, mean � SD of triplicates.
T-cell Engager-Armed Oncolytic Adenovirus
www.aacrjournals.org Cancer Res; 77(8) April 15, 2017 2057
on March 1, 2021. © 2017 American Association for Cancer Research. cancerres.aacrjournals.org Downloaded from
Published OnlineFirst January 31, 2017; DOI: 10.1158/0008-5472.CAN-16-1708

(Supplementary Fig. S4B) when engaged by cBiTE antibodies. Tcells were added to the cultures of HCT116- or A549-infected cellsand cell death was assessed using the xCELLigence system. Thecombination T cells and ICO15K-cBiTE showed a remarkableadditive effect by reducing by half the time to complete death ofA549 and HCT116 cells when compared with any of the othervirus treatments (Fig. 4D).
ICO15K-cBiTE increases the persistence and accumulation oftumor-infiltrating T cells
To address the potential of ICO15K-cBiTE in vivo, we firstevaluated the persistence and biodistribution of human T cellsin SCID/beige mice. To this end, human T cells were preactivatedand transduced with a lentiviral vector encoding the Click BeetleGreen (CBG) luciferase and GFP. T-cell preparations had 59%
Figure 4.
ICO15K-cBiTE–mediated oncolysis enhances T-cell function and induces a T-cell–mediated bystander effect. A549 andHCT116 cells infectedwith ICO15K and ICO15K-cBiTE were coculturedwith PBMCs. Uninfected (mock) cells were used a negative control.A, Five days after coculture CD8þ and CD4þ T cells were assessed by flowcytometry for the expression of CD25 and CD69. The mean � SD of triplicates is shown. B, Seven days after coculture, the CFSE content in CD4þ and CD8 Tcellswasdeterminedbyflowcytometry. A representative result from triplicates is shown.C, ICO15K- or ICO15K-cBiTE–infectedA549cellswere coculturedwithA431-GFPLuc cells and PBMCs from two different donors. Cocultures without PBMCs were used as a negative control. Five days after infection, cocultures were stainedwith 7-AAD, and the absolute count of live A431-GFPLuc cells was determined by flow cytometry. The mean � SD of triplicates is shown. D, A real-timexCELLigence-based cytotoxicity assay was used to evaluate the lysis of A549 and HCT116 cells over a 120-hour period. Tumor cells were seeded and infected withICO15K or ICO15K-cBiTE at t0. Preactivated T cellswere added 24 hours later. Impedance atwell bottomswasmeasured every 20minutes and normalized to baselineimpedance values with medium only. The mean � SD of duplicates is shown. One representative experiment from three independent T-cell preparations fromdifferent healthy donors is shown. �� , P < 0.01; ��� , P < 0.001 by one-way ANOVA with Tukey post hoc test.
Fajardo et al.
Cancer Res; 77(8) April 15, 2017 Cancer Research2058
on March 1, 2021. © 2017 American Association for Cancer Research. cancerres.aacrjournals.org Downloaded from
Published OnlineFirst January 31, 2017; DOI: 10.1158/0008-5472.CAN-16-1708

transduced (i.e., GFPþ) cells and retained the ability to proliferatein vitro (Supplementary Fig. S5). To test the biodistribution andpersistence of these Luc-T cells in vivo, SCID/beige mice bearingsubcutaneous HCT116 tumors were injected intratumorally withPBS, ICO15K-cBiTE, or the parental virus. Five days after virusadministration, mice received an intravenous injection of theCBG-Luc T-cell preparations and tumor luminescence was mea-sured daily by in vivo imaging. T cells were mainly distributed inthe lymphoid organs 1 day after infusion and the signal generallydecreased within 3 days. Tumor radiance was measured from day3 on to avoid background signal coming from the initial engraft-ment in the lymphoid organs. Tumors treated with ICO15K-cBiTE–induced T-cell accumulation from day 3 with a peak atday 4, and with the signal lasting for 9 days until reaching thelevels of PBS- or ICO15K-treated tumors (Fig. 5A). Half of thetumors treatedwith ICO15K-cBiTE, but noneof those treatedwithPBS or ICO15K, showed a detectable increase in bioluminescentsignal after 3 days of adoptive T-cell transfer (Fig. 5B). In a similarindependent experiment, virus and cBiTE transcripts weredetected in tumors upon intratumoral administration of virusand subsequent systemic administration of T cells (Fig. 5C).
ICO15K-cBiTE enhances antitumor efficacy in vivoWe then assessed the antitumor efficacy of ICO15K-cBiTE in
mouse xenograft models of cancer. SCID/beige mice bearingsubcutaneous A549 tumors were injected intratumorally withPBS, ICO15K-cBiTE, or the parental virus. At 4 and 18 days aftervirus administration, mice received intravenous injections ofeither unstimulated human PBMCs or PBS. The PBS groupsshowed the fastest tumor growth and the cBiTE-expressing ade-novirus showed a similar antitumor efficacy as the parental virusin the absence of PBMCs. Notably, the administration of humanPBMCs significantly enhanced the antitumor efficacy of ICO15K-cBiTE but not that of ICO15K (Fig. 6A). Histologic analysis of thetumors revealed the expression of the E1a protein at day 49 in allgroups treated with any of the adenoviruses in the study, indi-cating that PBMC-mediated cytotoxicity had no effect on thepersistence of the virus in the tumor (Fig. 6B).
We also tested the effect of delivering ICO15K-cBiTE intrave-nously, a setting that is more therapeutically relevant. SCID/beigemice bearing subcutaneous HCT116 tumors were injected intra-venously with PBS, ICO15K, or ICO15K-cBiTE. On days 4, 8, and11 after virus administration, mice received an intravenous injec-tion of preactivated T cells followed by an intraperitoneal injec-tion of IL2. This regime was chosen based on the peak ofpersistence at day 4 (Fig. 5A), and it aimed at maximizing thepresence of T cells in the tumor during the first weeks after virustreatment. Adenovirus-treatedmice had a smaller tumor size thanthose treated with PBS, with ICO15K-cBiTE–treated ones havingthe smallest tumor volume after T-cell administration (Fig. 6C).Although these differences were not statistically significant fortumor volume, ICO15K-cBiTE showed significant reduction intumor growth compared with PBS and ICO15K from day 7 aftervirus administration (Fig. 6D). The presence of virus and cBiTE intumors was confirmed at the end of the study by immunohisto-logic analysis (Supplementary Fig. S6).
DiscussionIn this study, we armed the oncolytic adenovirus ICO15K with
an EGFR-targeting BiTE (cBiTE). cBiTEs secreted from infected
cells retained key features of BiTEs including target cell–depen-dent T-cell activation and proliferation and redirected lysis ofcancer cells (27, 28). The anti-EGFR scFV in the cBiTE was derivedfrom the monoclonal antibody cetuximab, which is used inpatients with colorectal and head-and-neck squamous cell cancer(29, 30).One of themajormechanisms of resistance to cetuximabin colorectal cancer is the mutation of downstream signalinggenes such as BRAF, KRAS, PIK3CA, and PTEN (31). A cetuxi-mab-derived BiTE overcomes this resistance by successfully redir-ecting T cells to kill BRAF- and KRAS-mutated colorectal cancercells (32). Here, we also demonstrate that ICO15K-cBiTE inducesa T-cell–mediated killing of KRAS-mutated HCT116 cells in vitroand in vivo.
The potential of BiTE-armed oncolytic viruses has been previ-ously demonstrated using an oncolytic vaccinia virus with anEphA2-targeting BiTE (vv-EphA2; ref. 14). Whereas vv-EphA2induced T-cell activation and PBMCs-mediated bystander killingof cancer cells, no T-cell proliferation was observed in the absenceof exogenous IL2 in vitro and in vivo. In contrast, we observed thatICO15K-cBiTE induced remarkable proliferation of T cells in vitro,without the addition of IL2 to the cocultures. This difference couldbe related to the BiTE design or the oncolytic virus used. Anothernoteworthyfinding fromour in vivo experimentswas the increasedT-cell infiltration and expansion observed in subcutaneoustumors treated with the cBiTE-expressing virus after intravenousT-cell administration. With regard to antitumor efficacy, vv-EphA2 prevented tumor growth of subcutaneous A549 tumorsthat were coimplanted with PBMCs immediately followed by anintraperitoneal injection of the virus, and delayed growth of 7-dayestablished lung A549 tumors after the coadministration of vv-EphA2 and PBMCS. In our antitumor efficacy experiments, wechose to use tumors established for longer periods (three and twoweeks for A549 and HCT116, respectively), and efficacy wasobserved even in this setting. Altogether, our work confirms thepotential of arming oncolytic viruseswith BiTEs. Leaving aside thedifferent tumor targets used in these approaches (EphA2 orEGFR), vaccinia could offer a faster and more immunogeniconcolysis, whereas adenovirus a higher selectivity based on pRBpathway constitutively activated in tumor cells.
The strategy of arming oncolytic adenoviruses with BiTEsexploits the best features of both therapies simultaneously, whileovercoming many of the hurdles encountered by both as singleagents. From the point of view of the BiTE, its localized expressionfrom infected cancer cells would reduce the adverse effects com-monly observed with BiTEs, some of which can be fatal if nottreated appropriately (33). Additionally, the continuous produc-tion of BiTEs from infected cancer cells would increase its avail-ability at the tumor site. This would be advantageous given theshort half-life of BiTEs in the serum, which requires continuousdrug infusion (i.e., 4–6 weeks; ref. 34). Conversely, BiTEs couldhelp to overcome some of the limitations in oncolytic virother-apy. Although oncolytic adenoviruses have been shown to elicitspecific T-cell responses to tumor neoepitopes in vivo (35), theimmunodominance of adenoviral epitopes masks immuneresponses to delivered transgenes or capsid-displayed tumorantigens (36, 37). We and others have successfully developedapproaches to favor MHC-I–restricted antitumor rather thanantiviral immune responses (7, 38). However, these strategiesrely onMHC-I expression, which is downregulated on cancer cellsas onemechanism to escape from immune responses (4, 39). TheMHC-I–independent and polyclonal mode of action of BiTEs
T-cell Engager-Armed Oncolytic Adenovirus
www.aacrjournals.org Cancer Res; 77(8) April 15, 2017 2059
on March 1, 2021. © 2017 American Association for Cancer Research. cancerres.aacrjournals.org Downloaded from
Published OnlineFirst January 31, 2017; DOI: 10.1158/0008-5472.CAN-16-1708

(40) offers a unique opportunity to theoretically activate andredirect any tumor infiltrating T cell, including adenovirus-specificCTLs, to cancer cells. Interestingly, it has been recently reportedthat BiTE antibodies can engage cytomegalovirus-specific CTLs tokill cancers cells in vitro (41). Because we have demonstrated thecBiTE-mediated polyclonal engagement of na€�ve and preactivatedCD4þ and CD8þ T cells toward cancer cells, we speculate thatredirecting adenovirus-specific CTLs is also feasible. In addition tothe antiviral immune responses, limited virus spread within thetumor due to stromal barriers is another limitation for oncolyticadenoviruses (1, 42). The small size of BiTEs (�55 kDa) isadvantageous for penetration and distribution throughout thetumor as demonstrated by imaging studies (43). Thiswould allowimproving the overall bystander effect of oncolytic adenoviruseseven if their spread is limited. Furthermore, this strategy can beexpanded to attack the tumor stroma by using BiTEs targeting, forexample, the fibroblast activation protein alpha on cancer-asso-ciated fibroblasts. Such a BiTE has been validated in vitro (44), andits expression from infected cells would focus the bystander effect
to stromal cells, which are normally resistant to the oncolyticeffect of the virus.
Althoughwewere able to amplify ICO15K-cBiTE, its onocolyticproperties were reduced approximately 2-fold compared with thenonmodified virus. This loss in cytotoxicity may be the result ofthe competition between the cBiTE and viral genes for transcrip-tion and translation. Despite this, the cBiTE gene gives a remark-able advantage to the virus in the presence of T cells even at lowMOI in vitro. Importantly, the loss in in vitro cytotoxicity of thecBiTE-expressing virus did not translate into a loss in antitumorefficacy in vivo in the absence of PBMCs.
We evaluated ICO15K-cBiTE systemically, which is the pre-ferred route of administration for oncolytic adenoviruses tomaximize the delivery of the virus to metastatic tumors. Further-more, we have described that replacing the KKTK domain in theshaft of the fiber with an RGD motif enhances the systemicantitumor efficacy of ICO15K by reducing liver targeting andincreasing virus bioavailability in the blood (19). In our study, thesystemic administration of ICO15K to SCID/beige mice led to a
Figure 5.
ICO15K-cBiTE induces accumulation and increased persistence of tumor-infiltrating T cells in vivo. A, SCID/beige mice bearing subcutaneous HCT116 tumors onboth flanks were intratumorally injected with PBS, ICO15K, or ICO15K-cBiTE (n ¼ 10 per group). Five days later, mice received an intravenous injection ofpreactivated T cells expressing the click beetle green luciferase (Luc-T cells), followed by an intraperitoneal injection of IL2. Body luminescence wasmonitored by invivo imaging for 9 days. The mean tumor radiance� SEM is shown from day 3 onward. � , P < 0.05, ICO15K-cBiTE versus ICO15K by the Student t test. B,Radiance from individual tumors from the experiment described inA.C, Presence of hexon and cBiTEmRNAupon similar treatment conditions as inA. T cellswithoutluciferase were administered systemically 8 days after intratumoral virus injection, and RNA was extracted 4 days later for qPCR analysis.
Fajardo et al.
Cancer Res; 77(8) April 15, 2017 Cancer Research2060
on March 1, 2021. © 2017 American Association for Cancer Research. cancerres.aacrjournals.org Downloaded from
Published OnlineFirst January 31, 2017; DOI: 10.1158/0008-5472.CAN-16-1708

Figure 6.
ICO15K-cBiTE shows improved antitumor efficacy in vivo.A, SCID/beige mice bearing subcutaneous A549 tumors were intratumorally injected with PBS, ICO15K, orICO15K-cBiTE. Four and 18 days after virus administration, mice received either an intravenous injection of PBMCs (n ¼ 9–10 per group) or PBS (n ¼ 10 per group).The mean tumor volume � SEM is shown. � , P < 0.05, ICO15K-cBiTE þ PBMCs versus all groups by the Student t test. B, OCT-embedded cryosections fromthe tumors described inAwere stained for the adenovirus E1a protein (green) and for nuclei with DAPI (blue). Representative composite images from each group areshown. C, SCID/beige mice subcutaneous HCT116 tumors were intravenously injected with PBS or ICO15K or ICO15K-cBiTE. Four, 8, and 11 days after virusadministration,mice received an intravenous injection of preactivated T cells, followedby an intraperitoneal injection of IL2. Themean tumor volume� SEM is shown.D, Tumor growth of the experiment described in C. The mean tumor growth � SEM is shown. � , P < 0.05, ICO15K-cBiTE versus all groups.
T-cell Engager-Armed Oncolytic Adenovirus
www.aacrjournals.org Cancer Res; 77(8) April 15, 2017 2061
on March 1, 2021. © 2017 American Association for Cancer Research. cancerres.aacrjournals.org Downloaded from
Published OnlineFirst January 31, 2017; DOI: 10.1158/0008-5472.CAN-16-1708

modest decrease in tumorgrowth comparedwithPBS. Thismodestantitumor efficacy ismost likely explained by the low dose of virusadministered. It has been demonstrated that SCID/beige mice aremore sensitive to oncolytic adenovirus infection thanBALB/c nudemice due to their increased immunodeficiency (45). Based on ourpreliminary studies, we determined that the highest systemictolerated dose of our viruses in SCID/beige mice was 1 � 1010
VP/mouse (data not shown). This dose is five times lower than theone used for efficacy studies of ICO15K in nude mice, in which aclear advantage was observed in several mouse models of cancer.Note, however, that this low dose was enough to observe theadvantage of ICO15K-cBiTE in the tumor model we evaluated.
Because the EGFR is expressed on cancer cells targeted by thevirus, we were concerned about the potential interference of thecBiTE-mediated cell killing with virus replication and persistencein vivo. E1a expression was detected in all virus-treated tumors atthe end of the study and it was independent of PBMC adminis-tration. These results show that, at least in our model, the cBiTE-mediated cancer cell death does not compromise the persistenceof the virus in the tumor. It is worth mentioning the limitation ofour model due to the transient persistence of human T cells inSCID/beige mice. An ideal setting to prove the persistence of thevirus in the tumor would involve the use of immunocompetentmousemodels. Furthermore, such amodel would enable to studythe ability of virus-delivered BiTEs to activate infiltrating T cellsunder the highly immunosuppressive environment of the tumor.However, the lack of adenovirus replication inmurine cells limitsthe study of adenovirus-mediated oncolysis in such models.Despite the cumbersomemodels of PBMC transfer in SCID/Beigemice with human tumor xenografts, it is worth highlighting that aBiTE-armed oncolytic adenovirus represents a single-agent sys-temic treatment easy to translate in patients where the immuneresponse against the virus provides a continuous source of endog-enous T cells to the tumors, enhancing the potential efficacy.
The marketing approval in the United States of the oncolyticvirus T-VEC for the treatment of advanced melanoma encouragesthe development of oncolytic viruses with improved immuno-therapeutic potential. Our data demonstrate that BiTE-armedoncolytic adenoviruses hold the unique properties of inducing
specific and redirected antitumor immune responses. This strategyhas the potential to solve key limitations in oncolytic virotherapyand encourages its further evaluation and development.
Disclosure of Potential Conflicts of InterestC.H. June reports receiving a commercial research grant from Novartis. No
potential conflicts of interest were disclosed by the other authors.
Authors' ContributionsConception and design: C.A. Fajardo, S. Guedan, R. AlemanyDevelopment of methodology: C.A. Fajardo, S. Guedan, R. Moreno,R. AlemanyAcquisition of data (provided animals, acquired and managed patients,provided facilities, etc.): C.A. Fajardo, L.A. Rojas, M. Arias-BadiaAnalysis and interpretation of data (e.g., statistical analysis, biostatistics,computational analysis): C.A. Fajardo, L.A. Rojas, C.H. June, R. AlemanyWriting, review, and/or revision of the manuscript: C.A. Fajardo, S. Guedan,C.H. June, R. AlemanyAdministrative, technical, or material support (i.e., reporting or organizingdata, constructing databases): M. Arias-BadiaStudy supervision: R. AlemanyOther (helped carrying out some of the experiments): J. de Sostoa
AcknowledgmentsThe authors thank Brian Keith for helpful comments on the article.
Grant SupportThis work was supported by a BIO2014-57716-C2-1-R grant from the
Ministerio de Economía y Competitividad of Spain and a 2014SGR364research grant from the Generalitat de Catalunya. C.A. Fajardo was sup-ported by the European Commission Marie Curie Initial Training NetworkADenoViruses as novel clinical treatments (ADVance, FP7; project reference:290002). Co-funded by the European Regional Development Fund, a way toBuild Europe. S. Guedan and C.H. June are members of the Parker Institutefor Cancer Immunotherapy, which supported the University of PennsylvaniaCancer Immunotherapy Program.
The costs of publication of this articlewere defrayed inpart by the payment ofpage charges. This article must therefore be hereby marked advertisement inaccordance with 18 U.S.C. Section 1734 solely to indicate this fact.
Received June 28, 2016; revised January 11, 2017; accepted January 24, 2017;published OnlineFirst January 31, 2017.
References1. Russell SJ, Peng K-W, Bell JC. Oncolytic virotherapy. Nat Biotech
2012;30:658–70.2. Buijs PRA, Verhagen JHE, van Eijck CHJ, van den Hoogen BG. Oncolytic
viruses: from bench to bedside with a focus on safety. Hum VaccinImmunother 2015;11:1573–84.
3. Pol J, Bloy N, Obrist F, Eggermont A, Galon J, Cremer I, et al. Trial Watch.OncoImmunology 2014;3:e28694.
4. Drake CG, Jaffee E, Pardoll DM. Mechanisms of Immune Evasion byTumors. Advances in immunology. Volume 90: Academic Press; 2006.p51–81.
5. Lichty BD, Breitbach CJ, Stojdl DF, Bell JC. Going viral with cancerimmunotherapy. Nat Rev Cancer 2014;14:559–67.
6. Galivo F, Diaz RM, Thanarajasingam U, Jevremovic D, Wongthida P,Thompson J, et al. Interference of CD40L-mediated tumor immunotherapyby oncolytic vesicular stomatitis virus. Hum Gene Ther 2010;21:439–50.
7. Rodriguez-Garcia A, Svensson E, Gil-Hoyos R, Fajardo CA, Rojas LA, Arias-Badia M, et al. Insertion of exogenous epitopes in the E3-19K of oncolyticadenoviruses to enhance TAP-independent presentation and immunoge-nicity. Gene Ther 2015;22:596–601.
8. Smith RR, Huebner RJ, RoweWP, SchattenWE, Thomas LB. Studies on theuse of viruses in the treatment of carcinoma of the cervix. Cancer1956;9:1211–18.
9. Cerullo V, Koski A, V€ah€a-Koskela M, Hemminki A. Chapter Eight - Onco-lytic Adenoviruses for Cancer Immunotherapy: Data fromMice, Hamsters,and Humans. In:David TC, Paul BF, editors. Advances in cancer research.Volume 115: Academic Press; 2012. p265–318.
10. de Gruijl TD, Janssen AB, van Beusechem VW. Arming oncolytic viruses toleverage antitumor immunity. Expert Opin Biol Ther 2015;15:959–71.
11. Baeuerle PA, Kufer P, Bargou R. BiTE: Teaching antibodies to engage T-cellsfor cancer therapy. Curr Opin Mol Ther 2009;11:22–30.
12. Klinger M, Benjamin J, Kischel R, Stienen S, Zugmaier G. Harnessing T cellsto fight cancer with BiTE� antibody constructs – past developments andfuture directions. Immunol Rev 2016;270:193–208.
13. Frankel SR, Baeuerle PA. Targeting T cells to tumor cells using bispecificantibodies. Curr Opin Chem Biol 2013;17:385–92.
14. Yu F,Wang X, Guo ZS, Bartlett DL, Gottschalk SM, Song X-T. T-cell engager-armed oncolytic vaccinia virus significantly enhances antitumor therapy.Mol Ther 2014;22:102–11.
15. NettelbeckDM.Cellular genetic tools to control oncolytic adenoviruses forvirotherapy of cancer. J Mol Med 2008;86:363–77.
16. New Drugs/Drug News. Pharmacy and Therapeutics2014;39:733–85.17. Barrett DM, Liu X, Jiang S, June CH, Grupp SA, Zhao Y. Regimen-specific
effects of RNA-modified chimeric antigen receptor T cells in mice withadvanced leukemia. Hum Gene Ther 2013;24:717–27.
Fajardo et al.
Cancer Res; 77(8) April 15, 2017 Cancer Research2062
on March 1, 2021. © 2017 American Association for Cancer Research. cancerres.aacrjournals.org Downloaded from
Published OnlineFirst January 31, 2017; DOI: 10.1158/0008-5472.CAN-16-1708

18. Carpenito C, Milone MC, Hassan R, Simonet JC, Lakhal M, Suhoski MM,et al. Control of large, established tumor xenografts with geneticallyretargeted human T cells containing CD28 and CD137 domains. ProcNatl Acad Sci 2009;106:3360–5.
19. Rojas JJ, Gimenez-Alejandre M, Gil-Hoyos R, Cascallo M, Alemany R.Improved systemic antitumor therapy with oncolytic adenoviruses byreplacing the fiber shaft HSG-binding domain with RGD. Gene Ther2012;19:453–7.
20. Bayo-Puxan N, Gimenez-Alejandre M, Lavilla-Alonso S, Gros A, CascalloM, Hemminki A, et al. Replacement of adenovirus type 5 fiber shaftheparan sulfate proteoglycan-binding domain with RGD for improvedtumor infectivity and targeting. Hum Gene Ther 2009;20:1214–21.
21. Stanton RJ, McSharry BP, Armstrong M, Tomasec P, Wilkinson GW. Re-engineering adenovirus vector systems to enable high-throughput analysesof gene function. Biotechniques 2008;45:659–62, 64–8.
22. Puig-Saus C, Rojas LA, Laborda E, Figueras A, Alba R, Fillat C, et al. iRGDtumor-penetrating peptide-modified oncolytic adenovirus showsenhanced tumor transduction, intratumoral dissemination and antitu-mor efficacy. Gene Ther 2014;21:767–74.
23. Gros A, Martínez-Quintanilla J, Puig C, Guedan S, Molleví DG, AlemanyR, et al. Bioselection of a gain of function mutation that enhancesadenovirus 5 release and improves its antitumoral potency. Cancer Res2008;68:8928–37.
24. Maizel JV, White DO, Scharff MD. The polypeptides of adenovirus. Virol-ogy 1968;36:115–25.
25. Rojas JJ, Guedan S, Searle PF, Martinez-Quintanilla J, Gil-Hoyos R,Alcayaga-Miranda F, et al. Minimal RB-responsive E1A promoter modifi-cation to attain potency, selectivity, and transgene-arming capacity inoncolytic adenoviruses. Mol Ther 2010;18:1960–71.
26. Puig-Saus C, Laborda E, Rodriguez-Garcia A, Cascallo M, Moreno R,Alemany R. The combination of i-leader truncation and gemcitabineimproves oncolytic adenovirus efficacy in an immunocompetent model.Cancer Gene Ther 2014;21:68–73.
27. Brischwein K, Parr L, Pflanz S, Volkland J, Lumsden J, Klinger M, et al.Strictly target cell-dependent activation of T cells by bispecific single-chainantibody constructs of the BiTE class. J Immunother 2007;30:798–807.
28. Mølhøj M, Crommer S, Brischwein K, Rau D, SriskandarajahM, HoffmannP, et al. CD19-/CD3-bispecific antibody of the BiTE class is far superior totandem diabody with respect to redirected tumor cell lysis. Mol Immunol2007;44:1935–43.
29. Jonker DJ, O'Callaghan CJ, Karapetis CS, Zalcberg JR, Tu D, Au H-J, et al.Cetuximab for the treatment of colorectal cancer. N Engl J Med2007;357:2040–8.
30. Vermorken JB, Trigo J, Hitt R, Koralewski P, Diaz-Rubio E, Rolland F,et al. Open-label, uncontrolled, multicenter phase II study to evaluatethe efficacy and toxicity of cetuximab as a single agent in patients withrecurrent and/or metastatic squamous cell carcinoma of the head andneck who failed to respond to platinum-based therapy. J Clin Oncol2007;25:2171–7.
31. Bardelli A, Siena S. Molecular mechanisms of resistance to cetuximab andpanitumumab in colorectal cancer. J Clin Oncol 2010;28:1254–61.
32. Lutterbuese R, Raum T, Kischel R, Hoffmann P,Mangold S, Rattel B, et al. Tcell-engaging BiTE antibodies specific for EGFR potently eliminate KRAS-
and BRAF-mutated colorectal cancer cells. Proc Natl Acad Sci 2010;107:12605–10.
33. Teachey DT, Rheingold SR, Maude SL, Zugmaier G, Barrett DM, Seif AE,et al. Cytokine release syndrome after blinatumomab treatment related toabnormal macrophage activation and ameliorated with cytokine-directedtherapy. Blood 2013;121:5154–7.
34. Klinger M, Brandl C, Zugmaier G, Hijazi Y, Bargou RC, Topp MS, et al.Immunopharmacologic response of patients with B-lineage acute lym-phoblastic leukemia to continuous infusion of T cell–engaging CD19/CD3-bispecific BiTE antibody blinatumomab. Blood 2012;119:6226–33.
35. Woller N, Gurlevik E, Fleischmann-Mundt B, Schumacher A, Knocke S,Kloos AM, et al. Viral infection of tumors overcomes resistance to PD-1-immunotherapy by broadening neoantigenome-directed T-cell responses.Mol Ther 2015;23:1630–40.
36. Schirmbeck R, Reimann J, Kochanek S, Kreppel F. The immunogenicity ofadenovirus vectors limits the multispecificity of CD8 T-cell responses tovector-encoded transgenic antigens. Mol Ther 2008;16:1609–16.
37. Frahm N, DeCamp AC, Friedrich DP, Carter DK, Defawe OD, Kublin JG,et al. Human adenovirus-specific T cells modulate HIV-specific T cellresponses to an Ad5-vectored HIV-1 vaccine. J Clin Invest 2012;122:359–67.
38. Capasso C, HirvinenM, GarofaloM, RomaniukD, Kuryk L, Sarvela T, et al.Oncolytic adenoviruses coated with MHC-I tumor epitopes increase theantitumor immunity and efficacy against melanoma. OncoImmunology2016;5:e1105429.
39. Carretero R, Romero JM, Ruiz-Cabello F, Maleno I, Rodriguez F, CamachoFM, et al. Analysis of HLA class I expression in progressing and regressingmetastatic melanoma lesions after immunotherapy. Immunogenetics2008;60:439–47.
40. Offner S, Hofmeister R, Romaniuk A, Kufer P, Baeuerle PA. Induction ofregular cytolytic T cell synapses by bispecific single-chain antibodyconstructs on MHC class I-negative tumor cells. Mol Immunol2006;43:763–71.
41. Schmittnaegel M, Levitsky V, Hoffmann E, Georges G, Mundigl O, Klein C,et al. Committing cytomegalovirus-specific CD8 T cells to eliminate tumorcells by bifunctional major histocompatibility class I antibody fusionmolecules. Cancer Immunol Res 2015;3:764–76.
42. Guedan S, Rojas JJ, Gros A, Mercade E, Cascallo M, Alemany R.Hyaluronidase expression by an oncolytic adenovirus enhances itsintratumoral spread and suppresses tumor growth. Mol Ther 2010;18:1275–83.
43. Warnders FJ, Waaijer SJH, Pool M, Lub–de Hooge MN, Friedrich M,Terwisscha van Scheltinga AGT, et al. Biodistribution and PET imaging oflabeled bispecific T cell–engaging antibody targeting EpCAM. J Nucl Med2016;57:812–17.
44. W€uest T, Moosmayer D, Pfizenmaier K. Construction of a bispecific singlechain antibody for recruitment of cytotoxic T cells to the tumour stromaassociated antigen fibroblast activation protein. J Biotechnol 2001;92:159–68.
45. Engler H, Machemer T, Philopena J, Wen S-F, Quijano E, Ramachandra M,et al. Acute hepatotoxicity of oncolytic adenoviruses in mouse models isassociated with expression of wild-type E1a and induction of TNF-a;.Virology 2004;328:52–61.
www.aacrjournals.org Cancer Res; 77(8) April 15, 2017 2063
T-cell Engager-Armed Oncolytic Adenovirus
on March 1, 2021. © 2017 American Association for Cancer Research. cancerres.aacrjournals.org Downloaded from
Published OnlineFirst January 31, 2017; DOI: 10.1158/0008-5472.CAN-16-1708

2017;77:2052-2063. Published OnlineFirst January 31, 2017.Cancer Res Carlos Alberto Fajardo, Sonia Guedan, Luis Alfonso Rojas, et al. Improves Antitumor EfficacyOncolytic Adenoviral Delivery of an EGFR-Targeting T-cell Engager
Updated version
10.1158/0008-5472.CAN-16-1708doi:
Access the most recent version of this article at:
Material
Supplementary
http://cancerres.aacrjournals.org/content/suppl/2017/01/31/0008-5472.CAN-16-1708.DC1
Access the most recent supplemental material at:
Cited articles
http://cancerres.aacrjournals.org/content/77/8/2052.full#ref-list-1
This article cites 44 articles, 9 of which you can access for free at:
Citing articles
http://cancerres.aacrjournals.org/content/77/8/2052.full#related-urls
This article has been cited by 9 HighWire-hosted articles. Access the articles at:
E-mail alerts related to this article or journal.Sign up to receive free email-alerts
Subscriptions
Reprints and
To order reprints of this article or to subscribe to the journal, contact the AACR Publications Department at
Permissions
Rightslink site. Click on "Request Permissions" which will take you to the Copyright Clearance Center's (CCC)
.http://cancerres.aacrjournals.org/content/77/8/2052To request permission to re-use all or part of this article, use this link
on March 1, 2021. © 2017 American Association for Cancer Research. cancerres.aacrjournals.org Downloaded from
Published OnlineFirst January 31, 2017; DOI: 10.1158/0008-5472.CAN-16-1708