Embed Size (px)
Citation preview

ON THE REGULATION OF ADENYL CYCLASE ACTIVITY IN THE RAT PINEAL GLAND
Benjamin Weiss
Laboratory of Preclinical Pharmacology National Institute of Mental Health
Saint Elizabeth's Hospital Washington, D.C. 20032
The reader might justifiably ask why one should be concerned with the fac- tors controlling the activity of adenyl cyclase of the pineal gland. The answer is twofold. The main reason is to use the pineal adenyl cyclase system as a model for studying the adrenergic receptor. As such, the rat pineal offers certain advantages over other tissues. First, it is innervated largely, if not exclusively, by sympathetic neurons originating from the superior cervical ganglia,l and there- fore one can readily study the effects of sympathetic nerve activity on the cyclic AMP system. Second, the physiological and biochemical activity of this gland can be altered by administering various biogenic amines or by subjecting the animal to different environmental conditions.2-5 Thus, gland activity can be acutely or chronically altered, and these changes may be correlated with changes in the adenyl cyclase-cyclic AMP system.
Another reason for studying the pineal is for its own sake. It is now clear that the pineal gland influences gonadal activity,2-6 and perhaps cyclic AMP is responsible for the formation of the antigonadotropic principle of the pineal.'?
The total intracellular concentration of cyclic AMP is dependent upon the relative rates of its formation and uptake on the one hand and its hydrolysis and exit from the cells on the other hand. However, the biological activity of cyclic AMP is probably determined by the concentration of free nucleotide available to its receptor sites. Therefore, one should study not only how the total concentration of cyclic AMP changes under different conditions, but also how these conditions affect the binding and release of cyclic AMP. Some of the factors controlling the intracellular concentrations of cyclic AMP are depicted diagrammatically in FIGURE 1.
In this review I will discuss the factors controlling the concentration of cyclic AMP in the pineal gland and will try to relate this to the physiological function of the gland. Moreover, using the pineal gland as a model, I will attempt to show that the organism can control the formation of cyclic AMP both acutely and chronically-acutely by activating adenyl cyclase and chron- ically by changing the amount of adenyl cyclase capable of being activated.
Acute Regulation of Adenyl Cyclase Activity
Adenyl cyclase activity of rat pineal gland homogenates is increased by the pharmacologically active catecholamines. This activatlon is specific in that other biogenic amines or polypeptide hormones that activate adenyl cyclase in other tissues are totally ineffective in stimulating pineal adenyl cyclase. The catecholc- mine-induced activation of adenyl cyclase is inhibited by beta-adrenergic block-
507

508 Annals New York Academy of Sciences
HORMONES AND NEUROHORMONES
ENZYME k T N l l Y \ ~ IHORMONE SENSIT- \
/
FIGURE 1. Regulation of the intracellular concentration of cyclic AMP. The forma- tion of cyclic AMP is dependent upon the activity of adenyl cyclase, which in turn is determined by the availability of substrate, the degree of activation of adenyl cyclase and the quantity of adenyl cyclase (synthesis and destruction). The loss of free cyclic AMP may be due to enzymatic hydrolysis by phosphodiesterase, exit of cyclic AMP from the cell, and binding of the cyclic nucleotide to intracellular proteins.
ing agents, such as propranolol and dichloroisoproteranol (DCI), but not by alpha-blo~kers.~~ l o Indeed the alpha-blocking agent, phentolamine, actually increases the response to norepinephrine.'O This suggests that the pineal adenyl cyclase system is related to the beta-adrenergic receptor and that perhaps the alpha-stimulating component of norepinephrine decreases adenyl cyclase activ- ity. This general concept is not unique to this system, but has been proposed by other investigators using other tissue preparations.11-14
Sodium fluoride also stimulates enzyme activity, but at a different site from that of norepinephrine, since its effect is not inhibited by propranolol or DCI.lo
Besides the beta-adrenergic blocking agents, there are other compounds, both naturally occurring and synthetically produced, which inhibit pineal adenyl cyclase. For example, the biogenic amine, ~ero tonin ,~ and nucleotides such as adenosine diphosphate,15 inhibit enzyme activity. Moreover, the phenothiazine derivatives, chlorpromazine and trifluoperazine-but not their sulfoxide metab- olites-also inhibit pineal adenyl cyclase activity (FIGURE 2) .I5
Although the adrenergic neurotransmitter, norepinephrine, increases adenyl cyclase activity when added to pineal homogenates l6 and increases the endog- enous levels of cyclic AMP when added to pineal glands in organ culture,l' there is no direct evidence showing that sympathetic nerve activity acutely

Weiss: Rat Pineal Gland 509
increases either the activity of adenyl cyclase or the concentration of cyclic AMP in the pineal. Nevertheless, sympathetic nerve activity has been shown to chronically change the catecholamine-sensitive adenyl cyclase system of rat pineal gland. These effects and other long-term factors that influence adenyl cyclase activity of pineal gland will be the subject of the next section.
Chronic Regulation of Adenyl Cyclase Activity
The rate of formation of cyclic AMP is dependent upon the activity of adenyl cyclase. Adenyl cyclase activity, in turn, is dependent upon the availa- bility of substrate (ATP), the degree of activation of the enzyme and the actual quantity of enzyme. Since the concentration of ATP in cells is well above the k,,, for adenyl cyclase (in pineal gland the apparent k,, is about 2 x lo-' M) , I"
it is unlikely that the availability of substrate is the rate limiting factor for en- zyme activity, although it is recognized that this statement may have to be revised when the compartmentation of ATP and adenyl cyclase is finally resolved. The activation of adenyl cyclase has been the subject of extensive investigations over the past decade. However, relatively few studies 18-21 have considered the question of how the organism can regulate the quantity of
100-
90- H TRIFWCPERAZINE &--A CHU)RPROMAZINE O--C TRIFWOPERAZINE
80 - SULFCXIDE
coNCENTRp1K)N ImM)
FIGURE 2. Inhibition of pineal adenyl cyclase activity by phenothiazine derivatives. Adenyl cyclase activity of rat pineal gland homogenates was determined', '' in the presence of [-norepinephrine (lo-' M). (From Weiss?' Reprinted by permission of Prentice-Hall, Inc.)

510 Annals New York Academy of Sciences
adenyl cyclase and, in particular, the amount of enzyme that can be activated by hormones.
Studies on the pineal adenyl cyclase system have provided several lines of evidence suggesting that there exists at least two distinct types of adenyl cyclase: one which is insensitive to hormonal activation and one which can be stimulated by hormonal treatment. This is analogous to the existence in adipose tissue of two lipolytic enzymes: a lipoprotein lipase that is not activated by catechola- mines and an intracellular lipase that is hormone sensitive. The two enzymes are affected differentially by hormonal treatment or by changing environmental conditions.22-20 Our data suggest that the hormone-insensitive adenyl cyclase system remains fairly constant throughout the animals’ life and changes little even in the face of fluctuating environmental conditions. The hormone-sensitive adenyl cyclase system, on the other hand, increases with age and adapts to changes in its exogenous or endogenous environment.
Early studies of the pineal adenyl cyclase system of rats showed not only that norepinephrine activates adenyl cyclase, but also that chronic sympathetic denervation of the pineal gland can increase the activation of adenyl cyclase by norepinephrine.lS Subsequent dose-response studies showed that the stimu- lation of adenyl cyclase by maximal as well as by submaximal concentrations of the catecholamine was increased by chronic denervation. The response to sodium fluoride was also increased by denervating the Since fluoride acts at a different site from that of norepinephrine,1° these results suggested that denervation resulted in a greater quantity of enzyme rather than an in- creased affinity of adenyl cyclase for norepinephrine.
Exposure of rats to continuous light for long periods also increased the norepinephrine and sodium fluoride-responsive adenyl cyclase system. This
E 5 2 250- \
: e \
0 - a2 200-
!-NOREPINEPHRINE (Kl-4 M)
/.
u -1 u > u 1 - 1 I I I I I
0 5 10 15 20 25 30 35 AGE (days)
FIGURE 3. Ontogenetic development of adenyl cyclase of rat pineal gland. Adenyl cyclase activity of rat pineal gland homogenates was determined ’. Is in the absence ( G O ) and in the presence (A--- A) of /-norepinephrine (lo-’ M ) . Each point represents the mean of six experiments. Brackets indicate the standard error. (From Weiss.” Reprinted by permission of the Journal of Neurochemistry.)

Weiss: Rat Pineal Gland 51 1
FIGURE 4. Ontogenetic development of adenyl cyclase of rat pineal gland. Adenyl cyclase activity of rat pineal gland homogenates was determined ', a in the presence of NaF (10.' M ) . Each point represents the mean of ten experiments. Brackets indicate the standard error (From Weiss." Reprinted by permission of the Journal of Neuro- chemistry.)
effect of light was mediated by the sympathetic nervous system since superior cervical ganglionectomy abolished the increased responsiveness of pineal adenyl cyclase to the catecholamine. In neither case (denervation or exposure to light) was there any significant change in the basal adenyl cyclase activity, i.e., enzyme activity measured in the absence of any stimulant.27
Studies of the ontogenetic development of pineal adenyl cyclase also pro- vided evidence for two adenyl cyclase systems. The enzyme system that was responsible for the basal adenyl cyclase activity remained constant from birth to 60 days of age, but the hormone or fluoride-sensitive enzyme changed dramatically postnatally. Pineal adenyl cyclase of one-day-old rats failed to respond to norepinephrine, whereas in 20-day-old animals the catecholamine increased enzyme activity about threefold (FIGURE 3) .** The response to sodium fluoride also increased with age suggesting that the ontogenetic changes were not at a site specific for norepinephrine (FIGURE 4) .2x
Sex hormones also influence the catecholamine-sensitive adenyl cyclase system of rat pineal gland. For example, we found that norepinephrine stimu- lated pineal adenyl cyclase activity to a greater extent in male than in female rats. Ovariectomy increased the response to norepinephrine and estrogen treat- ment reduced the norepinephrine-induced activation of adenyl cyclase. Once more the basal enzyme activity was unaffected (TABLE 1).29 These results sug- gested that the well-known reciprocal relationship that exists between a gland (pineal) and its target organ (ovary) may involve the adenyl cyclase-cyclic AMP system. Thus the pineal gland influences ovarian activity, and the ovaries, in turn, secrete hormones that regulate the activity of pineal adenyl cyclase.
The experiments described suggest that the organism can change the intra- cellular concentration of cyclic AMP by altering the quantity of adenyl cyclase

512 Annals New York Academy of Sciences
TABLE 1
INDUCED ACTIVATION OF ADENYL CYCLASE OF RAT PINEAL GLAND EFFECT OF OVARIECTOMY AND ESTRADIOL TREATMENT ON NOREPINEPHRINE-
Cyclic 3’,5’AMP formed (prnoleshg proteidmin k SE)
Experimental Subject Control Norepinephrine (lo-’ M) Male 159 -+ 14 444 c 42 Female 1 5 9 k 9 258 -+ 41 Ovariectornized 123 -+ 19 321 2 36 Ovariectornized + Estrogen * 145 k 16 176 & 39
* Estradiol benzoate (0.05 mg/kg s.c., in oil) administered 53, 29, and 5 hours before sacrifice. (From Weiss & Crayton.=)
available for activation. This then offers us the opportunity of finding specific compounds that can produce changes in the synthesis of specific adenyl cyclases. Thus far, studies of the long term effects of pharmacological agents on adenyl cyclase systems have been totally ignored.
Regulation of Phosphodiesterase Activity
About ten years ago, Butcher and Sutherland30 reported that the methyl xanthines inhibit phosphodiesterase activity. Since then, several reports have shown that these compounds can either mimic or potentiate the effects of catecholamines and cyclic AMP in several t i s s ~ e s , ~ l - ~ ~ including the pineal gland.8 However, there are conflicting reports on the effects of methyl xanthines on the intracellular concentrations of cyclic AMP. For example, Brodie and associates lR showed that theophylline increases the concentration of cyclic AMP in adipose tissue. This is in consonance with the reports that theophylline stim- ulates lipolysis when added in vitro to adipose tissue 32, 34 and potentiates the lypolysis induced by electrically stimulating rat epididymal fat pads.35 On the other hand, Kakiuchi and colleagues showed that theophylline blocks the rise in cyclic AMP induced by electrically stimulating slices of cerebral cortex. The problem in correlating the effects of phosphodiesterase inhibition with the con- centration of cyclic AMP may be due to the presence of more than one type of phosphodiesterase 37, 38 or to the possible inhibitory action of these com- pounds on adenyl cyclase activity. More specific and potent phosphodiesterase inhibitors will have to be found (possibly papaverine derivatives 38) before we can adequately investigate the role of phosphodiesterase in regulating the con- centration of cyclic AMP.
Regulation of Cyclic AMP
Obviously, the intracellular concentration of cyclic AMP depends largely on the activities of the enzymes that synthesize and hydrolyze it, and in general, good correlations exist between the concentration of cyclic AMP and the rela-

Weiss: Rat Pineal Gland 513
tive activities of adenyl cyclase and phosphodiesterase. For example, the pineal gland and cerebellum contain particularly high activities of adenyl cyclase com- pared with that of phosphodiesterase suggesting that the concentration of cyclic AMP in these tissues may be higher than in other areas of the brain. This proved to be the case:" However, one should not neglect the possibility that the binding of cyclic AMP to intracellular proteins also can regulate its con- centration.
We are presently studying the effects of catecholamines and the influence of the environment on the concentration of cyclic AMP in pineal gland with the hope of correlating changes in cyclic AMP with other biochemical and physio- logical events seen in the pineal. Cyclic AMP was assayed by converting it to ATP and then determining ATP by the firefly luciferin-luciferase technique as described by Ebadi, Weiss and Costa:" FIGURE 5 outlines the general principle of the assay. Briefly, the samples were homogenized in HCIO, and neutralized with KHCO,. Tritiated cyclic AMP was added to monitor the recovery of the cyclic nucleotide. Cyclic AMP was separated from other nucleotides, nucleo- sides, and purine bases by selective precipitation with barium hydroxide and zinc sulfate solutions, and then passing the barium-zinc supernatant fluid over a cation exchange resin..13 The purified cyclic AMP fraction was converted to ATP by adding phosphodiesterase, myokinase, and pyruvate k ina~e .~" Samples with boiled phosphodiesterase served as a blank. The concentration of ATP was determined in a liquid scintillation counter after adding firefly luciferin-luci- ferase.O3 Liquid scintillation fluid was then added and the samples were recounted to obtain the recovery of 3H-cyclic AMP.
When used in the liquid scintillation spectrometer, a purified luciferin- luciferase system can detect as little as lo-" moles of ATP. However, until the tissue blank and contaminating nucleotides present in the myokinase and pyruvate kinase can be reduced further, the assay cannot approach this level of sensitivity. Nevertheless, under the present conditions, the cyclic AMP in about 1 mg of cerebellum (or in one pineal gland) can readily be detected (FIGURE 6) . The entire procedure can be completed in one day, and as many as 100 samples may be run simultaneously.
The preliminary results showed that the in vitro addition of norepinephrine ( l0- l M) to pineal gland in organ cultures produced almost a tenfold increase in the concentration of cyclic AMP within ten minutes after the addition of the catecholamine." Highly significant increases were also seen with M concentrations of norepinephrine. Propranolol, the beta-adrenergic blocking agent which inhibits the norepinephrine-induced activation of adenyl cyclase in
Phosphod l e s t e r a s e CYCLIC 3 ' ,5 ' -AMP + H20
5'AMP + ATP. nyok'nase \ 2 ADP
2 ADP + 2 PHOSPHOENDLPYRUVATE, Pyruvate
> 5'AMP
' 2 ATP + 2 PYRWATE
ATP + LUCIFERIN,-LUCIFERYL AOENYLATE COMPLEX + PYROPHOSPHATE I
5'AMP + LIGHT
FIGURE 5 . Assay of cyclic 3',5'-AMP by the firefly luciferin-luciferase technique. (From Ebadi et a/.")

514 Annals New York Academy of Sciences
80-
0 2 4 6 8 10 12 mg CEREBELLUM
FIGURE 6. Cyclic 3',5'-AMP in rat cerebellum as a function of tissue concentration. Rats were decapitated and brains were homogenized in 0.4 N HCIOl one minute after decapitation. Cyclic AMP was determined by the method described by Ebadi ef al."
pineal gland,g* lo also inhibited the norepinephrine-induced rise in cyclic AMP of pineal gland.'? The physiological relevance of these findings as well as the relationship between the cyclic AMP system and the metabolism of pineal indoles will be the subject of the next section.
Function of Cyclic AMP in Pineal Gland
The physiological effects of the pineal gland on gonadal activity are well known.2-e Moreover, it is clear that adenyl cyclase is present in the pineal gland in particularly high concentrations compared with other brain areas 40 and that it is activated by catech~lamines.~~ However, it was only recently that investi- gators have conclusively shown that cyclic AMP has marked biochemical effects on pineal tissue and that the activation by cyclic AMP of specific enzymes in the pineal may be responsible for its physiological
Some of the interrelationships between physiological and biochemical events in the pineal are easily interpretable while others remain obscure. Part of the problem stems from trying to relate in vitro biochemical findings with in vivo physiological events. The effects of environmental lighting are a case in point.
Pineal glands of rats maintained in darkness have an increased activity of hydroxyindole-0-methyltransferase (HIOMT) 49-51 and an increased concentra- tion of melatonin,52 the product of HIOMT activity.62 Darkness also increases the activity of N-acetyltransferase in pineal gland.53 (N-acetyltransferase cata- lyzes the N-acetylation of serotonin to form N-acetyl serotonin, the precursor for melatonin synthesis.) Thus, in darkness there should be a greater synthesis of N-acetylated and, perhaps, 0-methylated indoles. Since exposure of animals to darkness causes gonadal atrophy and since this is prevented by pinealectomy, 8, 5 4 , 55 it has been suggested that melatonin may be responsible for the inhibitory action of the pineal gland on gonadal development. In support of this Wurtman and associates showed that the administration of melatonin delayed the onset of estrus in the rat.
Norepinephrine and cyclic AMP stimulate melatonin production of pineal
Weiss: Rat Pineal Gland 515
\
0 0) E
\
-
.- a a 16 -
1 4 - -
2 12 -
a 8 -
'" 6 - m u
- a 10- I I
4 -
d 2 - > *
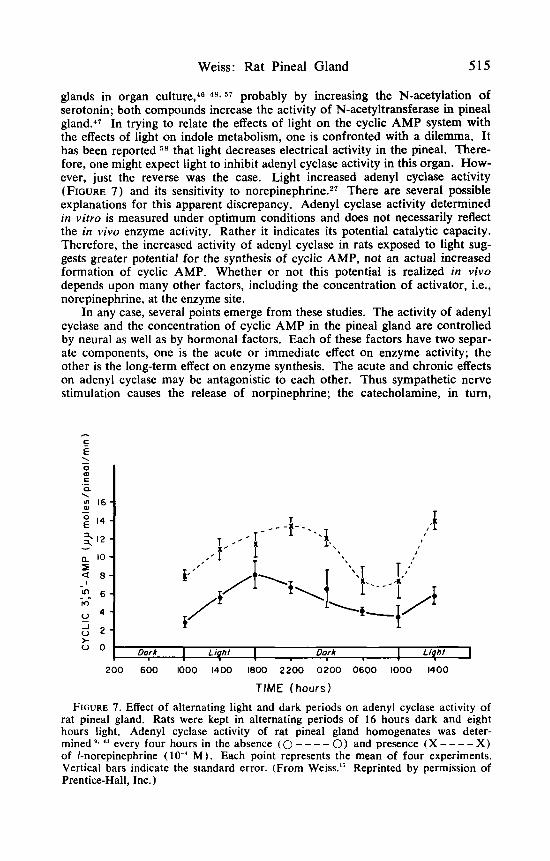
glands in organ culture,"-4X~ S T probably by increasing the N-acetylation of serotonin; both compounds increase the activity of N-acetyltransferase in pineal gland.47 In trying to relate the effects of light on the cyclic AMP system with the effects of light on indole metabolism, one is confronted with a dilemma. It has been reported 5 8 that light decreases electrical activity in the pineal. There- fore, one might expect light to inhibit adenyl cyclase activity in this organ. HOW- ever, just the reverse was the case. Light increased adenyl cyclase activity (FIGURE 7) and its sensitivity to norepinephrine.*? There are several possible explanations for this apparent discrepancy. Adenyl cyclase activity determined in vitro is measured under optimum conditions and does not necessarily reflect the in vivo enzyme activity. Rather it indicates its potential catalytic capacity. Therefore, the increased activity of adenyl cyclase in rats exposed to light sug- gests greater potential for the synthesis of cyclic AMP, not an actual increased formation of cyclic AMP. Whether or not this potential is realized in vivo depends upon many other factors, including the concentration of activator, i.e., norepinephrine, at the enzyme site.
In any case, several points emerge from these studies. The activity of adenyl cyclase and the concentration of cyclic AMP in the pineal gland are controlled by neural as well as by hormonal factors. Each of these factors have two separ- ate components, one is the acute or immediate effect on enzyme activity; the other is the long-term effect on enzyme synthesis. The acute and chronic effects on adenyl cyclase may be antagonistic to each other. Thus sympathetic nerve stimulation causes the release of norpinephrine; the catecholamine, in turn,
T
,

516 Annals New York Academy of Sciences
activates adenyl cyclase 0 and increases the concentration of cyclic AMP.17 However, there are indications that prolonged sympathetic denervation can increase the norepinephrine-sensitive adenyl cyclase ~ystem.~7 Hormones also activate adenyl cyclase of many tissues,I1 but prolonged estrogen treatment causes inhibition of the catecholamine-sensitive adenyl cyclase system of pineal gland.2y When one studies the effects of light, the problem is confounded because light changes both neuronal activity and hormonal secretion. Light entering the eye stimulates the retina. The nerve stimulus reaches the pineal by a pathway involving the retina, inferior accessory optic tract, median forebrain bundle, superior cervical ganglia, and nervi conarii.'. O0 Light also may stimulate the secretion of pituitary hormones by acting on the hypothalamus-median eminance system, and these hormones may affect the pineal adenyl cyclase system. To complicate the situation further, the effects of prolonged exposure to light may be different from those of acute exposure. For example, the light- induced diurnal fluctuations of norepinephrine are prevented by continuous light.6'
NORE P I NE PHR I NE
ADENYL CYCIASE 1 . ATP t C Y C L I C AMP
1 N-ACETYLTRANSFERASE
Y SEROTONIN H I OMT
MELATONIN-S-ADENOSYLMETHIONINE + N-ACETYLSEROTONIN
FIGURE 8. Mechanism of the norepinephrine-induced stimulation of melatonin formation in rat pineal gland. Norepinephrine released from sympathetic nerve endings activates the hormone-sensitive adenyl cyclase system increasing the intracellular concentration of cyclic AMP. Cyclic AMP stimulates N-acetyltransferase activity which causes a greater rate of formation of N-acetylserotonin. N-acetylserotonin is 0-methylated to the antigonadotropic principle, melatonin, under the catalytic influence of hydroxyindole-0-methyltransferase (HIOMT) .
Although several unanswered questions remain, particularly with regard to the effects of light, it is clear from in vitro studies that the norepinephrine- induced activation of pineal adenyl cyclase is responsible for the alterations of pineal metabolism. This scheme is shown in FIGURE 8. Norepinephrine activates adenyl cyclase and increases the concentration of cyclic AMP. Cyclic AMP increases N-acetyl transferase thereby increasing the N-acetylation of serotonin. N-acetylserotonin is 0-methylated to form melatonin under the catalytic influ- ence of hydroxyindole-0-methyltransferase, S-adenosylmethionine serving as the methyl donor. Thus, the physiological response of sympathetic nerve activity through the adenyl cyclase-cyclic AMP system increases the synthesis of the methoxyindole, melatonin. Melatonin inhibits gonadal activity. Gonadal hor- mones can feed back to the pineal suppressing the norepinephrine sensitive adenyl cyclase system.

Weiss: Rat Pineal Gland 517
References
1.
6.
7.
8.
9.
10.
1 1 .
12.
13.
14.
15.
16.
17. 18.
19.
20.
KAPPERS, J. A. 1960. The development, topographical relations and innervation of the epiphysis cerebri in the albino rat. Z. Zellforschung. Mikrosk. Anat. 52: 163-215.
MESS, B. 1968. Endocrine and neurochemical aspects of pineal function. Intern. Rev. Neurobiol. 11: 171-198.
WURTMAN, R. J., J. AXELROD & D. E. KELLY. 1968. The Pineal. Academic Press. New York, N.Y.
WURTMAN, R. J. & F. ANTON-TAY. 1969 The mammalian pineal as a neuroendo- crine transducer. Recent Progr. Hormone Res. 25: 493-514.
COSTA, E. 1970. Function and diseases of the epiphysis cerebri. I n Cellular and Molecular Basis of Neurologic Diseases. E. S. Goldensohn & S. H. Appel, Eds. Lea and Febiger. Philadelphia, Pa.
REITER, R. J. 1967. The effect of pineal grafts, pinealectomy, and denervation of the pineal gland on the reproductive organs of male hamsters. Neuroendo- crinol. 2: 138-146.
W~rss , B. & J. CRAYTON. 1970. Neural and hormonal regulation of pineal adenyl cyclase activity. I n Advances in Biochemical Psychopharmacology. Vol. 3 . P. Greengard & E. Costa, Eds. : 217-239. Raven Press. New York, N. Y.
KLEIN, D. C. & G . R. BERG. 1970. Pineal gland: stimulation of melatonin pro- duction by norepinephrine involves cyclic AMP mediated stimulation of N- acetyltransferase. I n Advances in Biochemical Psychopharmacology. Vol. 3. P. Greengard & E. Costa, Eds. : 241-263. Raven Press. New York, N. Y.
WEISS, B. & E. COSTA, 1968. Selective stimulation of adenyl cyclase of rat pineal gland by pharmacologically-active catecholamines. J. Pharmacol. Exp. Therap. 161: 310-319.
WEISS, B. 1969. Similarities and differences in the norepinephrine- and sodium fluoride-sensitive adenyl cyclase system. J. Pharmacol. Exp. Therap. 166: 330- 338.
ROBISON, G. A., R. W. BUTCHER & E. W. SUTHERLAND. 1967. Adenyl cyclase as an adrenergic receptor. Ann. N. Y. Acad. Sci. 139: 703-723.
TURTLE, J . R. & D. M. KIPNIS. 1967. An adrenergic receptor mechanism for the control of cyclic 3‘,5’-adenosine monophosphate synthesis in tissues. Biochem. Biophys. Res. Comm. 28: 797-802.
HANDLER, J. S., R. BENSINGER & J. ORLOFF. 1968. Effect of adrenergic agents on toad bladder response to ADH, 3’,5’-AMP and theophylline. Am. J. Physiol. 215: 1024-1031.
ROBISON, G. A., A. ARNOLD & R. C. HARTMANN. 1969. Divergent effects of epinephrine and prostaglandin E, on the level of cyclic AMP in human blood platelets. Pharmacol. Res. Comm. 1: 325-332.
WEISS, B. 1970. Factors affecting adenyl cyclase activity and its sensitivity to biogenic amines. I n Biogenic Amines as Physiological Regulators. J. J . Blum, Ed. : 35-73. Prentice Hall, Inc. Englewood Cliffs, N. J.
WEISS, B. & E. COSTA. 1967. Adenyl cyclase activity in rat pineal gland: Effects of chronic denervation and norepinephrine. Science 156: 1750-1752.
STRADA, S., D. C. KLEIN & B. WEISS. Unpublished observations. BRODIE, B. B., J . I. DAVIES, S. HYNIE, G. KRISHNA & B. W~rss . 1966. Interrela-
lationships of catecholamines with other endocrine systems. Pharmacol. Rev. 18: 273-289.
KRISHNA, G., S. HYNIE & B. B. BRODIE. 1968. Effects of thyroid hormones on adenyl cyclase in adipose tissue and on free fatty acid mobilization. Proc. Nat. Acad. Sci. U.S. 59: 884-889.
ROSEN, 0. M. & S. Mi ROSEN. 1968. The effect of catecholamines on the adenyl cyclase of frog and tadpole hemolysates. Biochem. Biophys. Res. Comm. 31: 82-9 1 .

518
21. BRODIE, B. B., G. KNSHNA & S. HYNIE. 1969. On the role of adenyl cyclase in the regulation of lipolysis in fasting. Biochem. Pharmac. 18: 1129-1 134.
22. BJORNTORP, P. & R. H. FURMAN. 1962. Lipolytic activity in rat epididymal fat pads. Am. J. Physiol. 203: 316-322.
23. MALLOV, S. 1963. Cold effects in rat: Plasma and adipose tissue free fatty acids adipose lipase. Am. J. Physiol. 204: 157-164.
24. GOODMAN, H. M. & E. KNOBIL. 1964. Hypophysectomy and the lipolytic action of epinephrine in vitro. Proc. Soc. Exp. Biol. Med. 115: 849-852.
25. GORIN, E. & E. SHAFRIR. 1964. Lipolytic activity in adipose tissue homogenate toward tri-, di-, and monoglyceride substrates. Biochem. Biophys. Acta 84: 24- 34.
26. GOODMAN, H. M. 1969. The effects of epinephrine on glycerol production in seg- ments of adipose tissue preincubated with dexamethasone and growth hormone. Proc. SOC. Exp. Biol. Med. 130: 909-912.
27. WEISS, B. 1969. Effects of environmental lighting and chronic denervation on the response of adenyl cyclase of rat pineal gland to norepinephrine and sodium fluoride. J. Pharm. Exp. Therap. 168: 146-152.
28. WEISS, B. 197 1. Ontogenetic development of adenyl cyclase and phosphodiester- ase in rat brain. J. Neurochem. 18: 469-478.
29. WEISS, B. & J. CRAYTON. 1970. Gonadal hormones as regulators of pineal adenyl cyclase activity. Endocrinology 87: 527-533.
30. BUTCHER, R. W. & E. W. SUTHERLAND. 1962. Adenosine 3’,5’-phosphate in bio- logical materials. I. Purification and properties of cyclic 3’,5’-nucleotide phosphodiesterase and use of this enzyme to characterize adenosine 3’,5’- phosphate in human urine. J. Biol. Chem. 237: 1244-1250.
31. RALL, T. W. & T. C. WEST. 1963. The potentiation of cardiac inotropic responses to norepinephrine by theophylline. J. Pharmacol. Exp. Therap. 139 269-274.
32. WEISS, B., J. I. DAVIES & B. B. BRODIE. 1966. Evidence for a role of adenosine 3’,5’-monophosphate in adipose tissue lipolysis. Biochem. Pharmacol. 15: 1553- 1561.
33. SIGGINS, G. R., B. J. HOFFER & F. E. BLOOM. 1969. Cyclic adenosine mono- phosphate: possible mediator for norepinephrine effects on cerebellar Purkinje cells. Science 165: 1018-1020.
34. HYNIE, S., G. KRISHNA & B. B. BRODIE. 1966. Theophylline as a tool in studies of the role of cyclic adenosine 3‘,5’-monophosphate in hormone-induced lipoly- sis. J. Pharmacol. Exp. Therap. 153: 90-96.
35. WEISS, B. & R. P. MAICKEL. 1968. Sympathetic nervous control of adipose tissue lipolysis. Intern. J. Neuropharm. 7: 395-403.
36. KAKIUCHI, S., T. W. RALL & H. MCILWAIN. 1969. The effect of electrical stimu- lation upon the accumulation of adenosine 3‘,5’-phosphate in isolated cerebral tissue. J. Neurochem. 16: 485-491.
37. HARDMAN, J . G. & E. W. SUTHERLAND. 1965. A cyclic 3’,5’-nucleotide phos- phodiesterase from heart with specificity for uridine 3’,5’-phosphate. J. Biol. Chem. 240: PC3704-PC3705.
38. BROOKER, G., L. J. THOMAS, JR. & M. M. APPLEMAN. 1968. The assay of adenosine 3’,5’-cyclic monophosphate and guanosine 3’,5‘-cyclic monophosphate in biological materials by enzymatic radioisotopic displacement. Biochemistry 7: 4177-4181.
39. MARKWARDT, F. & A. HOFFMAN. 1970. Effects of papaverine derivatives on cyclic AMP phosphodiesterase of human platelets. Biochem. Pharmacol. 19: 25 19- 2520.
40. WEISS, B. & E. COSTA. 1968. Regional and subcellular distribution of adenyl cyclase and 3’,5’-cyclic nucleotide phosphodiesterase in brain and pineal gland. Biochem. Pharmacol. 17: 2107-21 16.
41. EBADI, M. S., B. WEIS & E. COSTA. 1971. Distribution of cyclic adenosine mono- phosphate in rat brain. Arch. Neurol. 24: 353-357.
Annals New York Academy of Sciences

Weiss: Rat Pineal Gland 519
42.
43.
44.
45.
46.
47.
48.
49.
50.
51.
52.
53.
54.
55.
56.
57.
58.
59.
60.
61.
62.
63.
EBADI, M. S., B. WEISS & E. COSTA. 1971. Microassay of adenosine 3’,5’-mono- phosphate (cyclic AMP) in brain and other tissues by the luciferin-luciferase system. J. Neurochem. 18 183-192.
KRISHNA, G., B. WEISS & B. B. BRODIE. 1968. A simple, sensitive method for the assay of adenyl cyclase. J. Pharmacol. Exp. Therap. 163: 379-385.
BRECKENRIDGE, B. McL. 1964. The measurement of cyclic adenylate in tissues. Proc. Nat. Acad. Sci. U.S. 52: 1580-1586.
EBADI, M. S., B. WEISS & E. COSTA. 1970. Adenosine 3’,5’-monophosphate in rat pineal gland: increase induced by light. Science 170: 188-190.
SHEIN, H. M. & R. J. WURTMAN. 1969. Cyclic adenosine monophosphate stimu- lation of melatonin and serotonin synthesis in cultured rat pineals. Science 161: 519-520.
KLEIN, D. C., G. R. BERG & J. WELLER. 1970. Melatonin synthesis: adenosine 3’,5’-monophosphate and norepinephrine stimulate N-acetyltransferase. Science
KLEIN, D. C., G. R. BERG, J. WELLER & W. GLINSMANN. 1970. Pineal gland: dibutyryl cyclic adenosine monophosphate stimulation of labeled melatonin production. Science 167: 1738-1740.
WURTMAN, R. J., J. AXELROD & L. S. PHILLIPS. 1963. Melatonin synthesis in the pineal gland: Control by light. Science 142: 1071-1073.
AXELROD, J., R. J. WURTMAN & S. H. SNYDER. 1965. Control of hydroxyindole-0- methyltransferase activity in the rat pineal gland by environmental lighting. J. Biol. Chem. 240: 949-954.
WEISS, B. 1968. Discussion of the formation, metabolism, and physiologic effects of melatonin. Advan. Pharmacol. 6A: 152-155.
TOMATIS, M. E. & R. ORIAS. 1967. Changes in melatonin concentration in pineal gland in rats exposed to continuous light or darkness. Acta Physiol. Latino. Americana 17: 227-233.
KLEIN, D. C. & J. L. WELLER. 1970. Indole metabolism in the pineal gland: A circadian rhythm in N-acetyltransferase. Science 169 1093-1095.
REITER, R. J., S . D. SORRENTINO, J. C. HOFFMANN & P. H. RUBIN. 1968. Pineal, neural and photic control of reproductive organ size in early androgen-treated male rats. Neuroendocrinology 3: 246-255.
REITER, R. J. 1968. The pineal gland and gonadal development in male rats and hamsters. Fert. Steril. 19: 1009-1017.
WURTMAN, R. J., J. AXELROD & E. W. CHU. 1963. Melatonin, a pineal substance: Effect on the rat ovary. Science 141: 277-278.
AXELROD, J., H. M. SHEIN & R. J. WURTMAN. 1969. Stimulation of C“-melatonin synthesis from C”-tryptophan by noradrenaline in rat pineal in organ culture. Proc. Nat. Acad. Sci. US. 62: 544-549.
TAYLOR, A. N. & R. W. WILSON. 1970. Electrophysiological evidence for the action of light on the pineal gland in the rat. Experientia 26: 267-269.
AXELROD, J., S . H. SNYDER, A. HELLER & R. Y . MOORE. 1966. Light-induced changes in pineal hydroxyindole-0-methyltransferase: Abolition by lateral hypothalamic lesions. Science 154: 898-899.
MOORE, R. Y., A. HELLER, R. J. WURTMAN & J. AXELROD. 1967. Visual pathway mediating pineal response to environmental light. Science 155: 220-223.
WURTMAN, R. J., J. AXELROD, G . SEDVALL & R. Y. MOORE. 1967. Photic and neural control of the 24-hour norepinephrine rhythm in the rat pineal gland. J. Pharmacol. Exp. Therap. 157: 487492 .
AXELROD, J. & H. WEISSBACH. 1961. Purification and properties of hydroxy- Iindole-0-methyltransferase. J. Biol. Chem. 236: 21 1-213.
TAL, E., S. DIKSTEIN & F. G. SULMAN. 1964. ATP determination with the tricarb scintillation counter. Experientia 20: 652-653.
168: 979-980.


![Light and electron microscopic examination of pineal gland in ......histological structure of the pineal gland [8–18]. So, in our study it was aimed to examine the pineal gland of](https://img.pdfslide.us/doc/110x75/60c0abd73475835fb9682a43/light-and-electron-microscopic-examination-of-pineal-gland-in-histological.jpg)