Embed Size (px)
Citation preview

Middle East Journal of Applied Sciences ISSN 2077-4613
Volume : 07 | Issue :02 | April-June| 2017 Pages: 280-288
Corresponding Author: Mohammed Hasan Ibraheem, Department of Zoology, Faculty of Science, Minia University, El-Minia 61519, Egypt. E-mail: [email protected]
288
On the Morphology of the Oxyurid Nematode Aplectana macintoshii (Stewart, 1914) Travassos, 1931 (Ascaridida: Cosmocercidae) from the Toad Bufo regularis Reuss in Egypt
Mohammed Hasan Ibraheem Department of Zoology, Faculty of Science, Minia University, El-Minia 61519, Egypt. E-mail :[email protected]
Bahaa Kenawy Abuel-Hussien Abdel-Salam Department of Zoology, Faculty of Science, Minia University, El-Minia 61519, Egypt. Department of Biology, Quwiayha College of Science and Humanities, 11961, Shaqra University, Saudi Arabia. E-mail :[email protected]
Al-Shimaa Mahmoud El-Morsi Department of Zoology, Faculty of Science, Minia University, El-Minia 61519, Egypt. E-mail : [email protected]
Received: 20 November 2016 / Accepted: 25 December 2016 / Publication date: 27 April 2017
ABSTRACT
The aim of the present study was to characterize the morphological features of Aplectana macintoshii using light and scanning electron microscopy. Morphometric measurements of adult worms are given. Scanning electron microscopy revealed a transversely striated body and a triangular mouth opening. The latter is surrounded by three large oral lips and two pairs of thick cuticular flaps. The cephalic sense organs consist of a circle of 12 cephalic papillae (4 internal and 8 external) and a pair of large amphids. The excretory pore, the vulva and the anal opening of the female appeared as transverse slit-like openings transversal to the longitudinal axis of the body. The lateral alae extend along the length of the body from the cephalic extremity to the anal region and become disapparent beyond the anus. The body of both sexes is covered by minute somatic papillae arranged in six longitudinal rows. The posterior sense organs of the female are in the form of three post-anal sessile papillae, while those of the male are at least ten pairs of papillae arranged in post-cloacal, ad-cloacal and pre-cloacal positions. A pair of long, equal, sharply pointed spicules projects from the cloacal opening. The posterior end of the male is sharply bent ventrally and ends with a tapering spike. The latter bears variable number of randomly distributed caudal papillae. Deirids on the anterior end, as well as plectanes, rosette papillae, spines, caudal lateral alae or vesiculated papillae on the posterior end, were not observable in the present study. Key words: Aplectana macintoshii, Ascaridae, Bufo regularis, Cosmocercinae, Egypt, Nematoda, Oxyuridea.
Introduction
Nematode species of the genus Aplectana Railliet & Henry, 1916b are intestinal parasites of reptiles and amphibians (Yorke & Maplestone 1926, Travassos 1931, Baker 1980, 1987). They are cosmopolitan in geographical distribution, being encountered in Africa, China, Europe, Japan, India, South America and Malaysia (Baker 1987, Goldberg et al. 1993, Rago & Maharaj 2003, Yildirimhan et al. 2005, Bursey & Goldberg 2007, Holmes et al. 2008 and Ramalho et al. 2009). Aplectana macintoshii is the most widespread species of the genus and occurs in Africa, China, Europe, India, Malaysia and South America. It is now known from 37 species of frogs and toads, two species of lizards and one species of snakes (see Baker 1987). In Africa it has been reported from Cameroon (Rasheed, 1965) and Egypt (Baylis 1923). To our knowledge, there are few reports concerning A.

Middle East J. Appl. Sci., 7(2): 280-288, 2017 ISSN 2077-4613
281
macintoshii (Stewart, 1914) Travassos, 1931 from a few localities in Egypt. The only existing data on A. macintoshii from Egypt are those of Moravec el al. (1987), Farrag (1994), Mohammed (1996) and Moustafa (1999). Generally, a wide gap exists concerning the morphology of this nematode parasite. The purpose of this study was to throw light on the morphology of this helminth parasite as seen by light and scanning electron microscopy, as well as its prevalence of infection and mean abundance in the alimentary tract of the toad Bufo regularis Reuss in El-Minia Governorate, Egypt. Material and Methods
Two hundred and twenty specimens of the maculated toad Bufo regularis Reuss were captured from Mallawi Farm (27° 43′ 55° 43′ N and 30° 49′ 51° 53′ E), Minia, Egypt. The toads were brought to the laboratory alive and were immediately dissected. Intestinal contents were investigated in 0.75% physiological saline solution under a stereomicroscope and the collected worms were washed several times in saline solution. Twenty-six female and ten male nematodes of the genus Aplectana were obtained from the intestines of thirty-six adults of B. regularis. The nematodes were fixed in 70% ethanol and cleared in lactophenol for morphological study.
For SEM, the worms were fixed in a solution of 2.5% glutaraldehyde made in 0.1 M sodium cacodylate buffer containing calcium acetate (pH 7.2) at 4ºC for at least 2 hours. Subsequently, they were washed three times with the same buffer and post-fixed in 1% osmium tetroxide in 0.1 M sodium cacodylate buffer, pH 7.2, at 4ºC for 3 h. After washing several times in the buffer solution, the specimens were dehydrated through an assending ethanol series and dried in a critical-point drying machine using liquid carbon dioxide as a transitional medium. Later on, the specimens were mounted on aluminium stubs and then coated with gold using a JEOL JFC-1100 E ion sputtering device with a setting of 10 –15 mA, for 20 min. Afterwards, the specimens were examined with a stereoscan JEOL JSM-5400 LV operating at 15 kV. Morphometric measurements stated in the results section were taken from un-stained whole mount preparations of slightly flattened adult specimens and given in millimetres. All measurements are summarized in Table 1 and expressed as mean values, with their standard deviation (SD) and standard error (SE) values. Results Light Microscopy
Aplectana macintoshii is a small, slender nematode, whitish in color and fusiform in shape. The body tapers gradually towards the posterior region in the female, ending in a long tail. In the male the posterior extremity is slightly curved ventrally and the tail is short. The females are of a slightly greater length than the males. The lateral alae are narrow and extend throughout the greater part of the body from a post oesophageal level to the beginning of the tail in both sexes. They are much broader towards the posterior part than in the anterior. The mouth opening is triangular in shape and terminal in position. The esophagus is divided into a short anterior pharyngeal portion, an elongated corpus, a short narrow isthmus and a short subglobular tri-vulvated oesophageal bulb. The excretory pore opens at a short distance above the level of the oesophageal bulb. The esophagus leads to a yellowish tabular intestine that opens into the anus in the females or into the cloacal opening in the males. All morphometric measurements are given in Table (1).
Scanning Electron Microscopy
Scanning electron microscopic observations on both sexes of Aplectana macintoshii collected from the small intestine of Bufo regularis revealed a small slender body with the anterior end tapers gradually towards the posterior end. The cuticle bears transverse striations at regular intervals from the cephalic region to the tip of the tail, most of which are evident on the anterior half of the body. The mouth is triangular in shape (Fig. 1A). It is surrounded by three large lips; one lip is located dorsal and the other two lips are located subventral to the mouth opening. This structure is surrounded by two pairs of thick cuticular flaps; a dorsal pair lies on the dorsal lip and a single cuticular flap located on each sub ventral lip (Fig. 1A).

Middle East J. Appl. Sci., 7(2): 280-288, 2017 ISSN 2077-4613
282
Table 1: Morphometric measurements of male and female Aplectana macintoshii collected from Bufo regularis in Egypt.
Morphological parameters
Male Female Mean SD n SE Mean SD n SE
Total body length 2.504 0.591 10 0.187 4.772 1.169 26 0.229 Maximum width 0.240 0.040 10 0.013 0.406 0.110 26 0.022 Total length of esophagus 0.436 0.185 10 0.058 0.477 0.066 26 0.013 Pharynx 0.039 0.018 10 0.006 0.041 0.014 26 0.003 Esophageal corpus length 0.249 0.045 10 0.014 0.310 0.056 26 0.011 Esophageal corpus width 0.033 0.006 10 0.002 0.047 0.011 26 0.002 Esophageal bulb length 0.080 0.024 10 0.008 0.123 0.028 26 0.006 Esophageal bulb width 0.084 0.022 10 0.007 0.128 0.028 26 0.005 Nerve ring from anterior end 0.179 0.036 9 0.012 0.209 0.042 16 0.010 Excretory pore from ant. end 0.300 0.103 6 0.042 0.398 0.071 16 0.018 Vulva from anterior end - - - - 1.970 0.500 10 0.158 Intestine length 1.448 0.463 10 0.146 3.544 1.088 26 0.222 Maximum width of intestine 0.126 0.042 10 0.013 0.204 0.062 26 0.012 Cloaca from posterior end 0.235 0.030 8 0.011 - - - - Copulatory spicules length 0.191 0.051 10 0.016 - - - - Gubernaculum length 0.042 0.015 8 0.005 - - - - Egg length - - - - 0.090 0.062 75 0.007 Egg width - - - - 0.050 0.021 75 0.002 Total length of larva - - - - 0.407 0.184 75 0.021 Maximum width of larva - - - - 0.028 0.010 75 0.001 Anus from posterior end - - - - 0.507 0.092 16 0.023 Tail length 0.211 0.014 8 0.005 0.473 0.093 16 0.023
Fig. 1: Scanning electron micrographs (SEM) of the anterior end of female Aplectana macintoshii.-A. Triangular mouth opening retracted below the level of the lips. Note the complete closure of the amphid openings (2000x).-B. A pair of amphid openings is seen on both sides of the protruded mouth. Black arrows point to the circular groove, where the cephalic region is capable of being retracted into the anterior region of the body (1500x).-C. Partially retracted cephalic region inside the anterior region of the body (1500x) (Asterik refers to the mouth opening; cf, cuticular flap; cp, cephalic papillae; dl, dorsal lip; sl, sub ventral lip; White arrow refers to the amphid opening).
dl
C
cf
10 µm
dl
sl *
cp
10 µm A
cf
sl
*
10 µm
B
Middle East J. Appl. Sci., 7(2): 280-288, 2017 ISSN 2077-4613
283
The cephalic sense organs consist of a circle of eight external labial papillae, three of which are on each sub-ventral lip and two are on the dorsal lip. The sub-median pair of the cephalic papillae is much smaller than those of the ventro-lateral and dorso-lateral papillae (Fig. 1A). A pair of large amphids is present on both sides of the anterior end between the dorsal and the ventral pairs of the cuticular flaps (Figs. 1B, C).
The posterior end of the cephalic region of the worm is often marked by a constriction in the form of a circular groove, separating it from the rest of the body (Fig. 1B). At this level, the cephalic end sometimes appeared retractile, particularly in the female specimens (Fig. 1C).
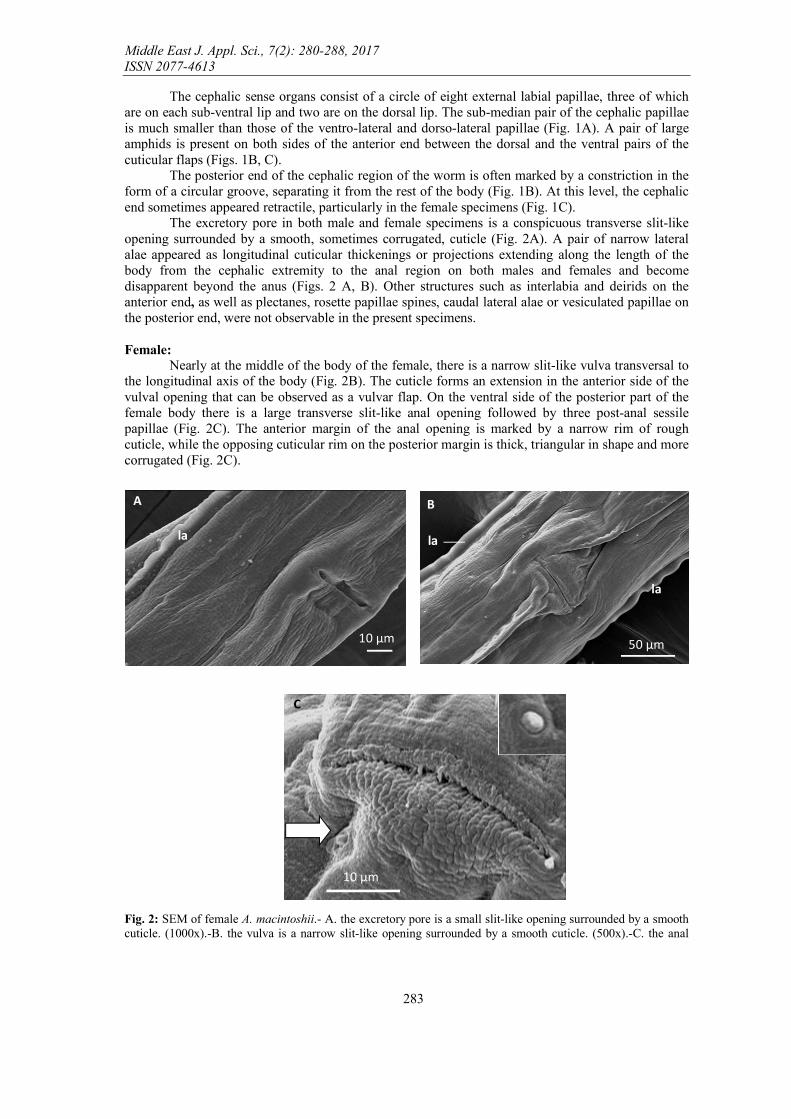
The excretory pore in both male and female specimens is a conspicuous transverse slit-like opening surrounded by a smooth, sometimes corrugated, cuticle (Fig. 2A). A pair of narrow lateral alae appeared as longitudinal cuticular thickenings or projections extending along the length of the body from the cephalic extremity to the anal region on both males and females and become disapparent beyond the anus (Figs. 2 A, B). Other structures such as interlabia and deirids on the anterior end, as well as plectanes, rosette papillae spines, caudal lateral alae or vesiculated papillae on the posterior end, were not observable in the present specimens. Female:
Nearly at the middle of the body of the female, there is a narrow slit-like vulva transversal to the longitudinal axis of the body (Fig. 2B). The cuticle forms an extension in the anterior side of the vulval opening that can be observed as a vulvar flap. On the ventral side of the posterior part of the female body there is a large transverse slit-like anal opening followed by three post-anal sessile papillae (Fig. 2C). The anterior margin of the anal opening is marked by a narrow rim of rough cuticle, while the opposing cuticular rim on the posterior margin is thick, triangular in shape and more corrugated (Fig. 2C).
Fig. 2: SEM of female A. macintoshii.- A. the excretory pore is a small slit-like opening surrounded by a smooth cuticle. (1000x).-B. the vulva is a narrow slit-like opening surrounded by a smooth cuticle. (500x).-C. the anal
10 µm
la
A B
la
la
50 µm
C
10 µm

Middle East J. Appl. Sci., 7(2): 280-288, 2017 ISSN 2077-4613
284
opening is a large crescent-shaped opening (inset is one of the three post-anal papillae). (2000x) (la, lateral alae; White arrow refers to the post-anal papilla)
Fig. 3: SEM of male A. macintoshii.-A. Shallow body striations and somatic papillae on the ventral surface. (2000x).-B. the posterior end showing slit-like cloacal opening with a single papilla at its anterior rim, pre-cloacal papillae, ad-cloacal papillae, a single papilla at anterior rim of the cloacal opening , spicules and spike. (750x).-C. cloacal opening with a single papilla (White arrow) at its anterior rim, pre-cloacal papillae, ad-cloacal papillae, a single papilla at anterior rim of the cloacal opening, spicules and spike. (2000x).-D. numerous caudal papillae randomly distributed on the spike tip. (2000x) (ap, ad-cloacal papillae; cl, cloacal opening; la, lateral alae; p, papillae; pp, pre-cloacal papillae; s, spicules; sk, spike).
Discussion
Aplectana macintoshii was originally described by Stewart (1914) as Oxysoma macintoshii for specimens taken from the intestines of Rana tigrina (currently Hoplobatrachus tigerinus) and Bufo stomaticus in Luck now, India. Railliet and Henry (1916a) assumed Aplecta as the type species of Ascaris acuminata (Schrank, 1788) during their review for oxyurid nematodes. Railliet and Henry (1916b) established the generic name Aplectana as a replacement name for Aplecta. Railliet and Henry (1916 b) assumed that Aplectana has had a somewhat turbulent history and eliminated them from Cosmocerca because those forms have no plectanes. Travassos (1931) reassigned the genus Oxysoma to Aplectana after a comprehensive review of the Cosmocercidae. He also accepted the 13 valid species so far and assigned them to the genus Aplectana. In addition, he described A. brumpti, A. schneideri and A. stromi to bring the total number of species assigned to the genus to 22.
A
10 µm
la
10 µm
B
sk s
cl
C
5 µm S S
pp
ap
cl
D
10 µm
p

Middle East J. Appl. Sci., 7(2): 280-288, 2017 ISSN 2077-4613
285
A. macintoshii has been reported by Rasheed (1965) from the Mascarene grassland frog, ptychadena mascareniensis (= Rana ptychadena mascareniensis) in Cameroon. Baker (1980) revised the genus and provided a key to 32 species. Later, Baker (1987) assigned 39 species to the genus. A. macintoshii was previously reported in the gecko Cyrtodactylus louisiadensis from New Guinea by Bursey et al. (2005). McAllister et al. (2010) reported a new host, Scotobleps gabonics for this cosmocercoid nematode.
In comparison to other Aplectana spp., it is observed that the female of A. macintoshii in the present study is slightly larger in size than that of other species like A. degraafi (Baker, 1981), A. herediaensis (Bursey et al., 2006) and A. krausi (Bursey and Goldberg, 2007). It has also been demonstrated that it is smaller than that of A. artigasi (Puga and Torres, 1997) and A. Capensis (Baker, 1981). Furthermore, the male of A. macintoshii is smaller in size than that of other species such as A. artigasi, A. capensis, A. degraafi and A. Krausi. The male of A. macintoshii in the present study most closely resembles that of A. herediaensis as recorded by Bursey et al., (2006), Puga and Torres (1997) and Bursey and Goldberg (2007). Barker (1981) reported that A. capensis most closely resembles A. macintoshii and distinguished these species from each other by the capitiulum of spicules in A. capensis that is not present in A. macintoshii. Furthermore, the posterior half of the male tail in A. capensis lacks papillae, whereas in A. macintoshii the papillae occur on the anterior three quarters of the tail. However, Bursey and Goldberg (2007) concluded that many specimens of A. macintoshii recovered from frogs in Australo-Papuan region have irregular patterns.
Scanning electron microscopic examinations of adult A. macintoshii in the present study revealed a transversely annulated cuticle with regular annuli. These annulations extend over the whole body, except for the tips of the head and tail. Annuli are not divided into sub annuli. Kozek and Marroquin (2002) considered the main function of the cuticle is to protect the nematodes from environmental influences. Smyth (1994) concluded that it maintains the shape, serves as antagonistic system for the somatic muscles and protects against inflammatory cells. Finally, Frantova (2002) assumed that it supplies the nutrients.
The mouth opening of the present material is originally surrounded by three lips and 12 sensory papillae but no cuticular teeth are observed. In A. artigasi (Puga et al., 1997) has an oral opening surrounded by one dorsal and two subventral lips. Each lip with a cuticular flap on the anterior ridge. cephalic sense organs consist of an inner circle of six minute papillae and an outer circle of four submedian papillae. In A. Krausi (Bursey et al., 2006), the mouth has three lips and four papillae. In A. capensis (Baker, 1981), there is a large oral opening surrounded by three lips and the cephalic extremity with six small inner labial papillae and six outer papillae on the anterior extremity of the oesophagus. Baker (1981) also recorded the presence of three lips, six minute inner labial papillae, six small outer papillae and three toothed like projections covered by a thick ring of cuticle at the anterior extremity of the oesophagus in A. degraaffi. Moreover, Wharton (1987) and Bursey and Goldberg (2007) reported that at the end of the oesophagus (in oesophageal bulb) of Aplectana spp, there is an additional valve that prevents the reflux of ingested materials.
Several species of Aplectana exhibit irregular distribution patterns of male caudal papillae (Skrjabin et al., 1961). A. macintoshii (Stewart, 1914), particularly, can be easily separated from both A. krausi (Bursey et al., 2007) and A. zweifeli (Moravec and Sey, 1986) which have defined patterns. In A. macintoshii the number of caudal papillae range from 10 to 32 pairs and some postanal papillae are not paired. Moreover, they cannot be distinguished from somatic papillae (Baker, 1980, Baker and Vaucher, 1986, Moravec and Sey, 1985 and Yuen, 1965). In the present study, the postanal papillae are not paired and the caudal papillae are variable in number and are randomly distributed.
As being concluded by Skrjabin et al. (1961) it can also be distinguished from other species of Aplectana by lacking the gubernaculum. The spicules are formed in a dorsal sac of the cloaca called the spicular pouch (Jamur, 1967). The spicules can be moved back and forth by accessory muscles and during copulation, they are inserted in the female's vulva. A thickening of the dorsal wall of spicular pouch, the gubernaculums, stabilizes the protruded spicule (Smyth, 1994). Aplectana spp., are characterized by equal spicules (Barker, 1981, Bursey & Golbderg, 2007 and Ramalho et al., 2008). In the present study the spicules are also equal in length and gubernaculum was detected. The characteristics by which Baylis and Daubney (1926) were able to distinguish between Aplectana sp. and Oxysomatium sp. are the relative length of the gubernaculum and the spicules, the position of the vulva and the condition of the ova when deposited. According to the results of Baylis and Daubney

Middle East J. Appl. Sci., 7(2): 280-288, 2017 ISSN 2077-4613
286
(1926) the position of the vulva in both Aplectana sp. and Oxysomatium sp. is median or anterior to the middle of the body, the ova hatch in utero and the gubernaculum is small.
Baylis (1923) recorded a nematode designated as A. acuminata from both Bufo regularis and Rana mascareniensis in Cairo and Alexandria (Egypt). Anyway, owing to insufficient data concerning the European specimens of this species, the identification of this material was uncertain. It has been concluded from the results of this study that A. macintoshii is the most abundant parasite of Bufo regularis in Cairo and its vicinity. Myers et al. (1962) considered Bufo regularis and Rana mascareniensis are the most frequently recorded host species of A. macintoshii in Africa. A. macintoshii has also been recorded from Morocoo by Baker 1980 in Rana ridibunda (Moravec et al., 1987). Moravec el al. (1987) described A. macintoshii from Imbaba in Cairo. This species has also been described by Farrag (1994) from Dakahlya Governorate, Mohammed (1996) from Sohag Governorate and Moustafa (1999) from Aswan Governorate.
Some minor differences in morphometric measurements between the present material and those of other authors are explained in Tables 1 & 2. These differences may be attributed to differences in the methods and techniques used and can be summarized as follows: 1. Farrag (1994) concluded that the males are longer and thicker than the males; a character which
was not confirmed in any previously described specimens.The males described by Farrag (1994) were longer than the females, while it is known that generally in nematodes, the males are usually shorter than the stouter females.
2. The vulva was mostly found post equatorial, but in the present material and in the material of Travassos (1931) and Mohammed (1996), it was found in an equatorial position. However, the position of the vulva may be subjected to minor variations depending on the state of the worm at the beginning of fixation.
References Baker, M. R., 1980. Revision of Old World species of the genus Aplectana Railliet & Henry, 1916
(Nematoda, Cosmocercidae). Bull. Mus. Natn. Hist. Nat. Ser. 4, Sect A, 2: 955 – 998. Baker, M. R., 1981. Cosmocercoid nematode parasites from frogs of southern Africa. Koedoe, 24: 25
– 32. Baker, M. R., 1987. Synopsis of the Nematoda parasitic in amphibians and reptiles. Memorial
University of Newfoundland Occasional Papers in Biology, 11: 1 – 325. Baker M. R. and C.Vaucher, 1986. Parasitic helminths from Paraguay XII: Aplectana Railliet and
Henry, 1916 (Nematoda: Cosmocercoidea) from frogs. Revue Suisse de Zoologie, 93: 607 – 616.
Baylis H. A., 1923. Report on a collection of parasitic nematodes, mainly from Egypt. Parasitology, 15: 14 – 23.
Baylis, H. A. and R.Daubney, 1926. A Synopsis of the families and genera of Nematoda. British Museum (Natural History), London, 277 pp.
Bursey, C. R. and S. R. Goldberg, 2007. A new species of Aplectana (Nematoda, Cosmocercidae) in Platymantis boulengeri (Anura, Ceratobatrachidea) from pupa New Guinea. Acta Parasitologica, 52 (4): 368 – 370.
Bursey, C. R., S. R.Goldberg and F.Kraus, 2005. New genus, new species of Cestoda (Anoplocephalidae), new species of Nematoda (Cosmocercidae) and other helminths in Cyrtodactylus louisiadensis (Sauria: Gekkonidae) from Papua New Guinea. J. Parasitol., 91: 882 – 889.
Bursey, C. R., S. R. Goldberg and S. R. Telford, 2006. New species of Aplectana (Nematoda: Cosmocercidae) and Mesocoelium monas (Digenea: Brachycoeliidae) in Lepidophyma flavimaculatum (Squmata: Xantusiidae). From Costa Rico. Carib. J. Sci., 42 (2): 164 – 170.
Farrage, H., 1994. Studies on certain parasitic fauna of Amphibia in Dakahlyia Governorate. M. Sc. Thesis, Faculty of science, Mansoura University.
Frantová, D., 2002. On the morphology and surface ultrastructure of some parasitic nematodes (Nematoda) of birds (Aves). Acta Societatis Zoologicae Bohemoslove–nicae, 66: 85 – 97.

Middle East J. Appl. Sci., 7(2): 280-288, 2017 ISSN 2077-4613
287
Goldberg, S. R., C. R. Bursey, and R.Tawil, 1993. Aplectana macintoshii (Nematoda: Cosmocercidae) in Eumeces latiscutatus (Sauria: Scincidae) from Japan. J. Helminthol. Soc. Wash., 60: 283 - 284.
Holmes, R. M., A. Bocchiglieri, F. R. De Araujo, and R. J. De Silva, 2008. New records of endoparasites infecting Hypsiboas albopunctatus in a savanna area in Brasilia. Barazil. Rasitol. Res., 102: 621 - 623.
Jamur, M. P., 1967. Studies of spermatogenesis in nematode Nippostrongylus brasliensis. Cell. Biol., 31: 38 – 396.
Kozek, J. W. and F. H. Marroquin, 2002. Electron microscopy of Onchocerca volvulus and its nodule. Proceeding of 10th International Congress of Parasitol., 4 – 9 August, Vancouver, Canda.
McAllister, C.T., C. R. Bursey, and P. S. Freed, 2010. Helminth parasites (Cestoidea, Nematoda, Pentastomida) of selected herpetofauna from Cameroon, West Africa. Acta Parasitologica, 55 (1): 90 – 93.
Mohammed, A. H., 1996. Studies on endoparasites in amphibian hosts collected from Sohag Governorate. M. Sc. Thesis, Sohag Faculty of science, South Valley University.
Moravec F. and O.Sey, 1985. Some nematode parasites of frogs (Rana spp) from North Viet Nam. Parasitologia Hungarica, 18: 63 – 77.
Moravec, F. and O.Sey, 1986. Three new nematode species from Phrynomantis spp. (Amphibia: Microhylidae) from Papua New Guinea. Folia. Parasitol. (Prague), 33: 343 – 351.
Moravec F., V.Baruš and B.Ryšavý, 1987. Some parasitic nematodes, excluding Heterakidae and Pharyngodonidae, from amphibians and reptiles in Egypt. Folia Parasitologica, 34: 255 – 267.
Moustafa, N., 1999. Studies on parasitic helminths of amphibian in Aswan Governorate. M. Sc. Thesis, Aswan Faculty of Ccience, South Valley University.
Myers B. J., R. E. Kuntz and W. H. Wells, 1962. Helminth parasites of reptiles, birds and mammals in Egypt VII. Check list of nematodes collected from 1948 – 1955. Can. J. Zool., 40: 531 – 538.
Puga, S. and P.Torres, 1997. Aplectana artigasi n.sp. (Nematoda: Cosmocercidae) from the frog Eupsophus calcaratus (Anura: Lelptodactylidae) in southern Chile. Mem. Inst. Oswaldo Cruz, 92: 767 – 770.
Railliet, A. and A.Henry, 1916a. Nouvelles remarques sur les oxyuridés. Ibid., 79 (7): 247 – 250. Railliet, A. and Henry, A. (1916b): Untitled footnote, in Railliet, A. 1916, L’évolution des
Schistosomes ou Bilharzies, D’après MM. Leiper, Atkinson et autres. Rec. Med. Vet., 92: 426. Rago, M. R. and I. R. Maharaj, 2003. Helminthes of the cane toad Bufo marinus from Trinidacl West
Indies. Caribbean. Sci., 39 (2): 242 - 245. Ramalho, G., C. R. Bursey and S. R. Goldberg, 2008. New Species of Aplectana (Ascaridida:
Cosmocercidae) in the toads, Rhinella granulose and Rhinella schneideri (Anura: Bufonidae) from northern Argentina. Parsitol., 94 (6):1357 – 1360.
Ramalho, A. C., R. J.da Silva, H.Schwartz and A. K. Peres, 2009. Helminthes from an introduced species (Tupinambia merianae) and two endemic species (Trachylepis atiantica and Amphisbaena ridlyi) from Fernando de Noronha Archipelago, Brazil. Parasitol. 13: 1 – 5.
Rasheed S., 1965. Some parasitic nematodes from the Cameroons (W. Africa). Journal of Helminthology, 39: 67 – 100.
Schrank, F. von P., 1788. Verzeichniss der bisher hinlänglich bekannten Eingeweidewürmer, nebst einer Abhandlung über ihre Anverwandtschaften. KgI. Swenska vetensk. Akad, Stockholm, 116 pp.
Skrjabin, K. I., N. P.Shikkhobalova, and E. A. Lagodovskaya, 1961. Oxyurata of animals and man Part 2. Israel Program for Scientific Translations, Keter Publishing House, Jerusalem, Israel (1974), 460 pp.
Smyth, J. A., 1994. Animal parasitology. 3rdedit. Combridge Univ. New York. Stewart, F. H., 1914. Studies on Indian Helminthology, No. I. Records of the Indian Museum, 10 (3):
165 – 193. Travassos, L., 1931. Pesquizas helminthologicas realizadas em Hamburgo. IX. Ensaio monographico
da familia Cosmocercidae Travassos, 1925 (Nematoda). Memorias do Instituto Oswaldo Cruz., 25 (3): 237 – 298.
Wharton, D. A., 1987. A functional biology of nematodes. Croom Helm., London.

Middle East J. Appl. Sci., 7(2): 280-288, 2017 ISSN 2077-4613
288
Yildirimhan, H. S., C. R. Bursey and S. R. Goldberg, 2005. Helminth parasites of the Caucasian salamander, Mertensiella caucasica from Turkey. Comp Parasitol., 72: 75 - 87.
Yorke, W. and P. A. Maplestone, 1926. The nematode parasites of vertebrates. J and A. Churchill, London, UK, 536 pp.
Yuen P. H., 1965. Some studies on the taxonomy and development of some rhabdisoid and cosmocercoid nematodes from Malayan amphibians. Zoologischer Anzeiger, 174: 275 – 298.





























