Embed Size (px)
Citation preview

Anat Embryol (1984) 169:209 218 Anatomy and Embryology �9 Springer-Verlag 1984
On the development of the coronary arteries in human embryos, stages 14-19 Giuseppe Conte and Antonio Pellegrini Istituto di Anatomia Umana Normale dell'Universit/t di Pisa, Pisa, Italy
Summary. Each coronary artery in humans develops, ini- tially, from two anlagen, one distal and the other proximal. The distal anlage, which is forerunner of the subepicardial branches of the coronary arteries, develops as subepicardiat vascular networks on the atrioventricular and interventricu- lar sulci and on the walls of the ventricles and bulbus; these networks are the right-posterior and left-anterior ones. The proximal anlage, which is forerunner of the truncus of the right and left coronary arteries, develops as several endothelial buds of the truncus arteriosus. Normally, only two buds, right and left, hollow out, increase in length and connect with the right and the left vascular networks, re- spectively, so that the coronary arteries are formed. The cardiac veins appear together with the coronary arteries, but as independent vessels. The authors advance a number of hypotheses as to the origin of certain variations and malformations of the coronary arteries.
Key words: Embryology Heart deve lopmen t - Coronary arteries development - Coronary arteries - Heart
Introduction
The descriptions given of the development of the human coronary arteries are incomplete and incorrect (Licata 1954, 1955, 1956; Hackensellner 1956; Vernall 1962; O'Rahilly 1971; Cooper and O'Rahil ly 1971; Hirakow 1983). This is due in part to the fact that, often, only a few human embryos have been studied and sometimes these were iso- lated specimens and did not represent continuous stages of development.
Moreover, not many embryological textbooks deal with this subject (Evans 1911; Bremer 1957; Patten 1960, 1970; Licata 1962; Goerttler 1963; Roberts 1961 ; Goor and Lille- hei 1975; Van Mierop 1979) and, it so, often refer to the development of coronary arteries in other mammalia.
As an exact knowledge of the time and modalities of the development of the coronary arteries is important for understanding the genesis of the variations and malforma- tions of these vessels, we considered it worthwhile to study their development in humans.
Offprint requests to. Prof. Dr. Giuseppe Conte, Istituto di Anato- mia Umana Normale, FacoltA di Medicina e Chirurgia detl'Uni- versit/~ di Pisa, Via Roma, 55, 56100 Pisa, Italy
Material and methods
The embryos studied were selected from the specimens in the embryological collection of the Institute of Normal Hu- man Anatomy, Faculty of Medicine, University of Pisa. Every embryo was the result of a spontaneous abortion and was well preserved. The embryos were fixed in Zenker's fluid, stained in toto with carmalum, embedded in paraffin wax and serially sectioned, in a sagittal or in a transverse direction, at a thickness of 10 gm. The 33 embryos (in the 4 tll to 9 th week of gestation) that we studied correspond to the developmental stages from 11 to 23 following the description of O'Rahilly (1971) and O'Rahil ly et al. (1981), stages which were previously termed "hor izons" by Streeter (1942, 1945, 1948). Among the embryos examined, we con- sidered it worthwhile to describe in detail only those in- cluded in stages 14 to 19 (Table 1) because these are, in
Table 1. Sequence of stages in human heart development
Stage Length Age Cata- mm days logue
n o
3 C 11 ca 24
3 V2
12 3.5 ca 26 C1
3.8 C3 13 ca 28
4 C21
5 C5 14 ca 32
6 V3
7 C2 7 C4 7 M14
15 ca 33 7.5 M6 8 C6
10.5 C22
9 C7 9 M15
16 ca 37 10 CIO 10 C12
Stage Length Age Cata- mm days logue
n o
17 11.5 ca 41 C8
12 M13 13 C13 13.2 C14 14 C9
18 14 ca 44 C19 14 VI 15 MI7 15 M2 16 M7
17 Cll 19 19 ca 47 Mll
20 M16 21 M12
34 M8 23 ca 57
35 MIO

210
our opinion, most suitable for observing the sequence of events in development of the coronary arteries. A wax re- construction was made of two embryonic hearts (7 and 14 mm long) by means of the method devised by Born- Schaper-Peter (Carazzi and Levi 1911) and by Laane and Bourier (1980).
Results
At stage 14, we observed groups of cells and hollow pericar- dial buttons in the subepicardial space on the atrioventricu- lar and interventricular sulci and on the ventricular and bulbar walls, as described by Martin (1894) in rabbit em- bryos; by Licata (1962), Obrucnick et al. (1972) and Hir- akow (1983) in human embryos.
At stage 15, we observed a vascular endothelial network in the subepicardial space at different points: in the posteri- or and right-anterior parts of the atrioventricular sulcus; on the posterior interventricular sulcus; on the diaphrag- matic surface of the left ventricle, near the interventricular sulcus; on the diaphragmatic and right-ventral surfaces of the right ventricle and bulbus (Figs. 1, 2, 7, 8, 9, 10). This
Figs. 1-8. Human embryo (7ram long; C4; stage 15) heart reconstruction ( x 50). Fig. 1, ventral view; Fig. 2, dorsocaudal view
Figs. 3-8 are transversal sections of the reconstruction. On Figs. 3, 4, 8 the microphotographs are superimposed. Fig. 3, at the cranial level; Figs. 4, 5, at the level of the venous sinus (vs), cranially to the great (gv) and small cardiac veins ; Fig. 6 shows both the great (gv) and small (sv) cardiac veins (black). Figs. 7, 8 show, caudally to the venous sinus, the subepicardial coronary network, which is the primordium of the coronary artery branches, ra, la right and left atria; rv, Iv right and left ventricles; t truncus; b bulbus; lh left sinus horn; rn, In right- posterior and left-anterior subepicardial vascular networks (white); Pa Pulmonary artery; Ao Aorta; ts truncus septnm; psv, asv primordia of the pulmonary and aortic valve cusps; ec the atrioventricular endocardial cushions joined together; lavc left atrioventricular canal; cd caudo-dorsal atrioventricular cushion; A pars caudalis A of bulbus-truncus ridge A- 1 ; B pars caudalis B of bulbus-truncus ridge B-3

211
Figs. 9, 10. Human embryo (7 mm long; C4; stage 15), heart sections ( x 120). These sections, dorsal view, correspond to Figs. 7, 8 of the reconstruction. Fig. 9 is 80 btm caudal to Fig. 12; Fig. 10 is 40 gm caudal to Fig. 9; Fig. 10a is a high magnification of Fig. 10 ( x 320). lv left ventricle; b bulbus; ra right atrium; rn right- posterior subepicardial vascular network; cd caudo-dorsal atrioventricular cushion; B pars caudalis B of bulbus truncus ridge B-3
right-posterior subepicardial vascular network (rn) is not connected with the venous sinus (vs). We also observed that, at intervals, branches of the network led off into the underlying myocardium of the left and right ventricles and bulbus. These branches came into contact with the capillary bed and the sinusoids of the left ventricle, in agreement with the reports by Licata (1955, 1956) and Patten (1960, 1970) concerning humans, by Dbal~ et al. (1968) and Vo- bo~il and Schiebler (1969) concerning rats, and by Virfigh and Challice (1981) concerning mice. A less developed vas- cular network was also observed in the subepicardial space at the level of the anterior interventricular sulcus and the left-anterior part of the atrioventricular sulcus (Figs. 1, 7, 8). From this left-anterior subepicardial vascular network (ln), too, branches led off into the underlying myocardium of both ventricles, near the anterior interventricular sul- CUS.
At stage 15, we also observed thin subepicardial vessels (gv, sv) near to and connected with the venous sinus; these
were bloodless and had few ramifications (Figs. 2, 4-6, 11, 12)
At stages 15-16, multiple endothelial solid anlagen, (buds, sprouts) develop from the endothelium of the pulmo- nary and aortic walls, above the primordial semilunar valves (Figs. 13, 13a-b, 14a-b, 15a-b, 16-19); this agrees with Hackensellner's descriptions (1956).
At stage 17, only two hollow endothelial anlagen (rct, Ict) are seen to communicate with the aorta, on the right and left sides, but they are not yet connected with the re- spectively right-posterior and left-anterior subepicardial vascular networks (Figs. 20-25).
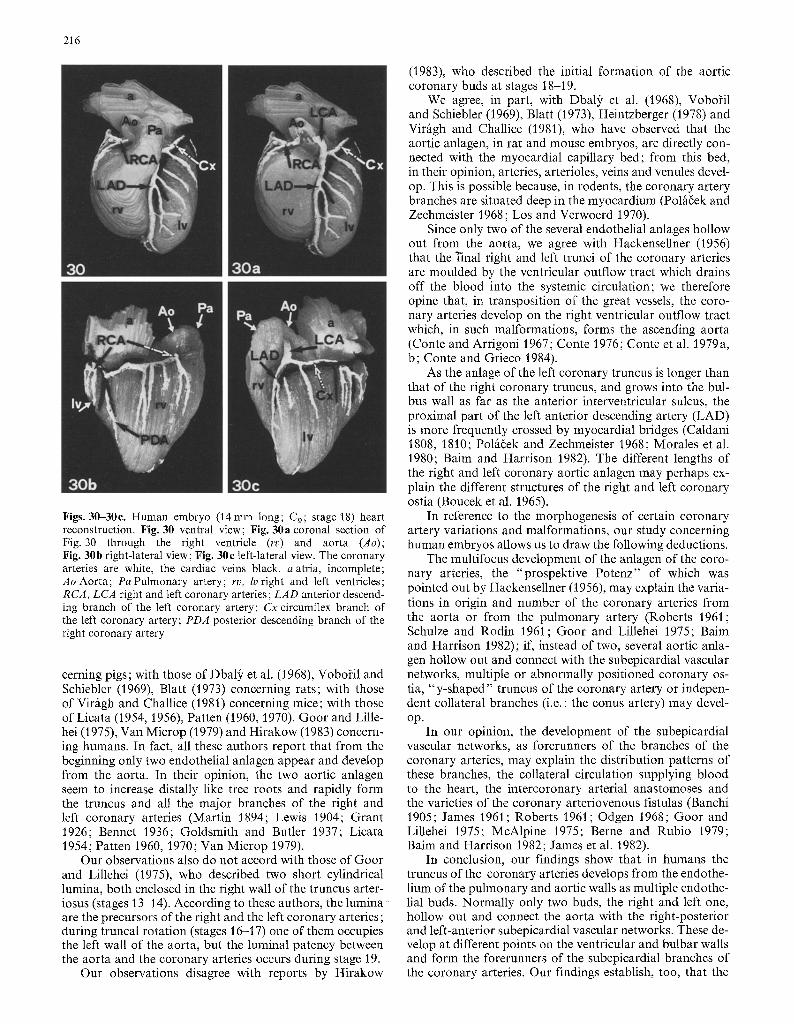
At stage 18, the right anlage is joined to the right-poste- rior subepicardial vascular network and thus the right coro- nary artery (RCA) system is formed (Figs. 26-28, 30, 30a, b). The truncus of the right coronary artery originates from the aortic anlage, while the branches of this artery originate from the subepicardial vascular network. The left anlage is longer (Figs. 21, 26, 27, 29) and runs in the bulbus wall

212
Figs. 11, 12. Human embryo (7 mm long; C4; stage 15), heart sections at the level of the venous sinus (vs), dorsal view ( x 120). Fig. 12 is 40 gm caudal to Fig. 11; Figs. l la , 12a are high magnification details of Figs. 11 and 12 respectively ( x 320). cv, cd cranio-ventral and caudo-dorsal atrioventricular cushions; ravc right atrioventricular canal; lv left ventricle; ra right atrium; rn right-posterior subepicardial vascular network; gv great cardiac vein
as far as the anterior interventricular sulcus. It is joined to the left-anterior subepicardial vascular network and thus the left coronary artery (LCA) system is formed (Figs. 26, 27, 30, 30a, 30c). The truncus of the left coronary artery originates from the aortic anlage as does, perhaps, the prox- imal part of the anterior descending branch of the left coro- nary artery (LAD) (Fig. 30); the branches of this artery, including the circumflex artery, originate from the subepi- cardial vascular network. The aortic anlage of the coronary arteries is more cellular than the corresponding subepicar- dial vascular network (Figs. 28, 29), in agreement with the reports by Boucek et al. (1965) concerning adults.
In this way, at stages 18-19, the right and left coronary system is formed.
Discussion
Before the coronary arteries develop, the myocardium of the embryonic heart is nourished by means of the sinusoids through the heart cavities (Licata 1956; Patten 1956, 1970).
At stages 14-15, two subepicardial vascular networks develop: one of these is wider and is situated on the posteri- or and right-anterior part of the atrioventricular sulcus, on the interventricular posterior sulcus, on the diaphrag- matic surface of the ventricles and on the right-anterior surface of the right ventricle and bulbus. The name we have given to this subepicardial vascular network is r i g h t - p o s t e r i o r , because the right coronary artery branches origi- nate from it. The other network is less developed and is

213
Figs. 13-15b. Human embryo (7 mm long; C4; stage 15), heart sections. The figures, dorsal view, show the pulmonary artery (Pa) and aorta (Ao) and the buds of the truncus of the coronary arteries (arrows). Fig. 13 (x 125); Fig. 13a (pulmonary artery) and 13b (aorta) are a high magnification ( x 250) of Fig. 13 Figs. 14a, 15a show sections subsequent to the one in Fig. t3a ( x 250) Figs. 14b, lgb show sections prior to the one in Fig. 13b ( x 250)
situated on the anterior interventricular sulcus, on the adja- cent wall of both the ventricles and on the left-anterior part of the atrioventricular sulcus. The name we have given to this subepicardial vascular network is left-anterior be- cause the left coronary artery branches originate from it. These two vascular networks are not connected with the venous sinus.
Moreover, at stage 15, we also observed thin subepicar- dial vessels connected with the venous sinus, which corre- spond to the vessels proviously interpreted as cardiac veins in rabbit, pig and mouse embryos by Martin (1894), Lewis (1904), Grant (1926), Bennet (1936), Goldsmith and Butler (1937), Halpern (1953), Vobof'il and Schiebler (1969) and Virfigh and Challice (1981).
Our observations, therefore, disagree with the conclu- sions of the above-mentioned authors who believe that the cardiac veins develop before the coronary arteries. In fact, in rabbit (from 12 to 14 days p.c.) and bat (Vesperugo pip- istrellus from 5.5 to 7 mm long) embryos, too, we have seen (unpublished observations) small branched venous ves- sels near to, and connected with, the venous sinus. We have also observed other, less branched, vessels: one of these is on the right-anterior part of the atrioventricular sulcus and on the right wall of the bulbus; the other is on the anterior interventricular sulcus and on the left-anterior part of the atrioventricular sulcus; these latter vessels, which have been observed in mouse embryos (11-12 days p.c.) by Vir~gh and Challice (1981), are not connected with the

Figs. 16-18. Human embryo (9 mm long; Cv; stage 16), heart sections, ventral view ( x 180). Fig. 16 is a coronal section of the aorta, at the level of the posterior valve cusps (D). Figs. 17, 18 are coronal sections of the pulmonary artery, at the level of the semilunar valve cusps
Fig. 19. Human embryo (10 mm long; C12; stage 16), heart section, ventral view ( x 100). Coronal section of the aorta (Ao) and pulmonary artery (Pa), cranially to the primordia of the aortic and pulmonary valve cusps. The figures show the buds of the truncus of the coronary arteries (arrows). Vventral valve cusp; rP right- posterior valve cusp ; lP left-posterior valve cusp ; asv, psv aortic and pulmonary semilunar valve cusps
Figs. 20-25. Human embryo (11.5 mm long; Cs; stage 17), heart sections, ventral view ( x 130). Sections of the aorta at the level of the origin of the left (lct) (Figs. 20, 21, 22) and right (rct) (Figs. 23, 24, 25) coronary artery trunci. The figures show sections of the left- anterior (ln) and right-posterior (rn) coronary subepicardial vascular networks, not yet connected to the corresponding trunci of the left and right coronary arteries

215
Figs. 26, 27. Human embryo (14 mm long; C9; stage 18), heart sections, ventral view ( x 90). The figures show the completion of the right (RCA) and the left (LCA) coronary artery
Fig. 28. High magnification ( x 330) of Fig. 26
Fig. 29. High magnification ( x 180) of Fig. 21. The figures show the right (rct) and the left (lct) coronary trunci, which are more cellular than the subepicardial coronary vascular networks (rn, ln). gv great cardiac vein; Ao Aorta
venous sinus and correspond to the human right-posterior and left-anterior subepicardial vascular networks. In hu- mans these develop at the same time (stages 14-15) as the ductus venosus (Keith 1921; Chiarugi 1944; Patten 1968; Puerta Fonoll/t and Jim6nez Collado 1980).
Before the development of the ductus venosus, the oxy- genated placental blood of the umbilical veins nourishes the ventricular walls through the ventricular sinusoids (Li- cata 1956; Patten 1956, 1970); the myocardial capillary bed develops during this stage (Licata 1956; Obrucnick et al. 1972).
When the right umbilical vein disappears and the ductus venosus develops (stages 14-15), most of the oxygenated blood returnig from the placenta by way of the left umbili- cal vein passes into the developing inferior vena cava by way of the ductus venosus. This blood, on reaching the right atrium, is usually shunted - through the foramen ovale
- to the left atrium and from this latter passes into the left ventricle. As a consequence, the right ventricular cavity receives mainly the non-oxygenated blood from the cardinal veins through the developing superior vena cava and the right atrium. Therefore, the right-posterior and left-anterior subepicardial vascular networks, which connect the left ven- tricular cavity with the wall of the right ventricle and bulbus
would, in our opinion, distribute a considerable supply of 0 2 to the myocardium of the right ventricle and bulbus. Furthermore, the same subepicardial vascular networks are conceivably the inductors of the numerous endothelial buds from the truncus arteriosus.
In fact, in human embryos the initial growth of the anlagen of the truncus of the coronary arteries may be seen in the pulmonary artery and in the aorta at stages 15 16; these anlagen are in the form of multiple endothelial sprouts from the pulmonary and aortic walls, on the primordial semilunar valve cusps. At stage 17, only two of the multiple endothelial sprouts hollow out, on the right and left walls of the aorta; at stage 18 they connect the aorta with the subepicardial right-posterior and left-anterior vascular net- works, respectively, to complete the bimodal development (Conte 1982; Hirakow 1983) of the coronary arteries. The right anlage forms the proximal part (truncus) of the right coronary artery; the left anlage, which is longer than the right one, forms the proximal part of the left coronary ar- tery and it may actually extend into the first portion of the LAD (see Fig. 30).
These observations are at variance with those of Martin (1894), Lewis (1904), Grant (1926) concerning rabbits; with those of Bennet (1936), Goldsmith and Butler (1937) con-
216
Figs. 30-30c. Human embryo (14ram long; C9; stage 18) heart reconstruction. Fig. 30 ventral view; Fig. 30a coronal section of Fig. 30 through the right ventricle (rv) and aorta (Ao); Fig. 30b right-lateral view; Fig. 30e left-lateral view. The coronary arteries are white, the cardiac veins black, a atria, incomplete; Ao Aorta; Pa Pulmonary artery; rv, lv right and left ventricles; RCA, LCA right and left coronary arteries; LAD anterior descend- ing branch of the left coronary artery; Cx circumflex branch of the left coronary artery; PDA posterior descending branch of the right coronary artery
cerning pigs; with those of Dbal) et al. (1968), Vobofil and Schiebler (1969), Blatt (1973) concerning rats; with those of Vir/tgh and Challice (1981) concerning mice; with those of Licata (1954, 1956), Patten (1960, 1970), Goor and Lille- hei (1975), Van Mierop (1979) and Hirakow (1983) concern- ing humans. In fact, all these authors report that from the beginning only two endothelial anlagen appear and develop from the aorta. In their opinion, the two aortic anlagen seem to increase distally like tree roots and rapidly form the truncus and all the major branches of the right and left coronary arteries (Martin 1894; Lewis 1904; Grant 1926; Bennet 1936; Goldsmith and Butler 1937; Licata 1954; Patten 1960, 1970; Van Mierop 1979).
Our observations also do not accord with those of Goor and Lillehei (1975), who described two short cylindrical lumina, both enclosed in the right wall of the truncus arter- iosus (stages 13-14). According to these authors, the lumina are the precursors of the right and the left coronary arteries; during truncal rotation (stages 16-17) one of them occupies the left wall of the aorta, but the luminal patency between the aorta and the coronary arteries occurs during stage 19.
Our observations disagree with reports by Hirakow
(1983), who described the initial formation of the aortic coronary buds at stages 18-19.
We agree, in part, with Dball) et al. (1968), Vobo~il and Schiebler (1969), Blatt (1973), Heintzberger (1978) and Virfigh and Challice (1981), who have observed that the aortic anlagen, in rat and mouse embryos, are directly con- nected with the myocardial capillary bed; from this bed, in their opinion, arteries, arterioles, veins and venules devel- op. This is possible because, in rodents, the coronary artery branches are situated deep in the myocardium (Pol/t~ek and Zechmeister 1968 ; Los and Verwoerd 1970).
Since only two of the several endothelial anlages hollow out from the aorta, we agree with Hackensellner (1956) that the "final right and left trunci of the coronary arteries are moulded by the ventricular outflow tract which drains off the blood into the systemic circulation; we therefore opine that, in transposition of the great vessels, the coro- nary arteries develop on the right ventricular outflow tract which, in such malformations, forms the ascending aorta (Conte and Arrigoni 1967; Conte 1976; Conte et al. 1979a, b; Conte and Grieco 1984).
As the anlage of the left coronary truncus is longer than that of the right coronary truncus, and grows into the bul- bus wall as far as the anterior interventricular sulcus, the proximal part of the left anterior descending artery (LAD) is more frequently crossed by myocardial bridges (Caldani 1808, 1810; Polgt~ek and Zechmeister 1968 ; Morales et al. 1980; Bairn and Harrison 1982). The different lengths of the right and left coronary aortic anlagen may perhaps ex- plain the different structures of the right and left coronary ostia (Boucek et al. 1965).
In reference to the morphogenesis of certain coronary artery variations and malformations, our study concerning human embryos allows us to draw the following deductions.
The multifocus development of the anlagen of the coro- nary arteries, the "prospektive Potenz" of which was pointed out by Hackensellner (1956), may explain the varia- tions in origin and number of the coronary arteries from the aorta or from the pulmonary artery (Roberts 1961; Schulze and Rodin 1961; Goor and Lillehei 1975; Bairn and Harrison 1982); if, instead of two, several aortic anla- gen hollow out and connect with the subepicardial vascular networks, multiple or abnormally positioned coronary os- tia, "y-shaped" truncus of the coronary artery or indepen- dent collateral branches (i.e. : the conus artery) may devel- op.
In our opinion, the development of the subepicardial vascular networks, as forerunners of the branches of the coronary arteries, may explain the distribution patterns of these branches, the collateral circulation supplying blood to the heart, the intercoronary arterial anastomoses and the varieties of the coronary arteriovenous fistulas (Banchi 1905; James 1961; Roberts 1961; Odgen 1968; Goor and Lillehei 1975; McAlpine 1975; Berne and Rubio 1979; Bairn and Harrison 1982; James et al. 1982).
In conclusion, our findings show that in humans the truncus of the coronary arteries develops from the endothe- lium of the pulmonary and aortic walls as multiple endothe- lial buds. Normally only two buds, the right and left one, hollow out and connect the aorta with the right-posterior and left-anterior subepicardial vascular networks. These de- velop at different points on the ventricular and bulbar walls and form the forerunners of the subepicardial branches of the coronary arteries. Our findings establish, too, that the

217
cardiac veins develop simultaneously with the coronary ar- teries, but as independent vessels.
References
Baim DS, Harrison DC (1982) Nonatherosclerotic coronary heart disease (including coronary artery spasm). In: Hurst JW (ed) The heart. McGraw-Hill Book Company, New York
Banchi A (1905) Morfologia delle arterie coronarie cordis. Arch Ital Anat Embriol 3:87-164
Bennett HS (1936) The development of the blood supply to the heart in the embryo pig. Am J Anat 60:27-53
Berne RM, Rubio R (1979) Coronary circulation. In: Berne RM, Sperelakis N, Geiger SR (eds) Handbook of physiology. Ameri- can Physiological Society, Bethesda, Maryland
Blatt H-J (1973) Uber die Entwicklung der Coronararterien bei der Ratte Licht- und elektronenmikroskopische Untersuchun- gen- Z Anat Entwickl-Gesch 142:53-64
Boucek RL, Takeshita R, Brady A1 H (1965) Microanatomy and intramural physical forces within the coronary arteries (Man). Anat Rec 153:233-242
Bremer JL (1957) Congenital anomalies of the viscera. Their em- bryological basis. Harvard University Press, Cambridge
Caldani F (1808) Iconum anatomicarum Explicatio. PIII , S I, Pi- cotti, Venetiis
Caldani F (1810) Icones anatomicae. VIII, S I, Picotti, Venetiis Carazzi D, Levi G (1911) Tecnica microscopica. SEL, Milano Chiarugi G (I944) Trattato di embriologia. P IV, S II, SEL, Milano Conte G (1976) A further contribution to the ontogenesis of" Com-
plete Transposition" of the great arteries. Atti Soc Ital Anat (XXXIII Conv Catania) 6667
Conte G (1982) Timing and sequence of events in human coronary circulation development. Boll Soc Ital Biol Sper 58:1238-1243
Conte G, Arrigoni P (1967) Precisazioni embriologiche su due alterazioni congenite di prima formazione del cuore: Aorta a cavaliere e trasposizione completa dei grossi vasi. Atti Soc Ital Cardiol, Milano 2:23-14
Conte G, Grieco M (1984) Closure of the interventricular foramen and morphogenesis of the membranous septum and ventricular septal defects in the human heart. Anat Anz 155:39-55 (in press)
Conte G, Grieco M, Giannessi F (1979a) Hemodynamic develop- mental patterns of the cardiac malformations. Atti Soc ItaI Anat (XXXVI Conv Ancona) Suppl Arch Ital Anat Embryol 84:113-114
Conte G, Grieco M, Paparelli A (1979 b) Embryology of the double outlet right ventricle-DORV. Atti Soc Ital Anat (XXXVI Conv Ancona) Suppl Arch Ital Anat Embryol 84:114
Cooper MH, O'Rahilly R (1971) The human heart at seven post- ovulatory weeks. Acta Anat 79:280-299
Dbal) J, Ogtfidal B, Rychter Z (1968) Development of the coronary arteries in rat embryos. Acta Anat 71 : 209-222
Evans HM (1911) Die Entwicklung des Blutgef/igsystems. In: Kei- bel F, Mall FP (eds) Hdb der Entwickl-Gesch des Menschen Bd 2, Hirzel, Leipzig
Goerttler K (1963) Entwicklungsgeschichte des Herzens. In: Barg- mann W, Doerr W (eds) Das Herz des Menschen Bd 1, Thieme, Stuttgart
Goldsmith JB, Butler HW (1937) The development of the cardiac- coronary circulatory system. Am J Anat 60:185-201
Goor DA, Lillehei CW (1975) Congenital malformations of the heart. Grune and Stratton, New York
Grant RT (1926) Development of the cardiac coronary vessels in the rabbit. Heart 13:261-271
Hackensellner HA (1956) Akzessorische Kranzgef/il3anlagen der Arteria pulmonalis unter 63 menschlichen Embryonen-serien mit einer gr613ten L/inge von 12 bis 36 mm. Z Mikrosk Anat Forsch 62:153 164
Halpern MH (1953) Extracoronary cardiac veins in the rat. Am J Anat 92 : 307-327
Heintzberger CFM (1978) DeveIopment of the vessels of the heart in the chicken and rat. Acta Morphol Neerl Scand 16:140-141
Hirakow R (1983) Development of the cardiac blood vessels in staged human embryos. Acta Anat 115:220-230
James TN (1961) Anatomy of the coronary arteries. Hoeber, Medi- cal Division of Harper and Brothers, USA
James TN, Sherf L, Schlant RC, Silverman ME (1982) Anatomy of the Heart. In: Hurst JW (ed) The heart arteries and veins, P I 5 th Ed, McGraw-Hill Book Company, New York
Keith A (192I) Human embryology and morphology, Arnold, Lon- don
Laane HM, Bourier J (1980) A quick three-dimensional reconstruc- tion method. Acta Morphol Neerl Scand 18 : 85 91
Lewis FT (1904) The question of sinusoids. Anat Anz 25:261-279 Licata RH (1954) The human embryonic heart in the ninth week.
Am J Anat 94:73-125 Licata RH (1955) The developmental basis of the blood supply
to the human heart. Anat Rec 121:330-331 Licata RH (1956) A continuation study of the development of
the blood supply of the human heart. Part II: the deep or intramural circulation. Anat Rec 124:326
Licata RH (1962) Coronary circulation: embryology. In: Abram- son DI (ed) Blood vessels and lymphatics. Academic Press, New York
Los JA, Verwoerd CDA (1970) The development of a primary venous system from epicardial villi in the cardiac wall of the chicken and the mouse embryo, and the relationship between this venous system and the arterial vascularisation in the mouse. Acta Morphol Neerl Scand 8:233
Martin H (1894) Recherches anatomiques et embryologiques sur les art4res coronaires du coeur chez les vert6br6s. Steinheil G, Paris
McAlpine WA 0975) Heart and coronary arteries. An anatomical atlas for clinical diagnosis, radiological investigation and surgi- cal treatment. Springer, Berlin
Morales AR, Romanelli R, Boucek RJ (1980) The mural left anteri- or descending coronary artery, strenuous exercise and sudden death. Circulation 62: 230-237
Obrucnik M, Malinsky J, Lichnovsky V (1972) The early stages of differentiation of the vascular bed in the ventricular wall of the human embryonic heart as seen in the electron micro- scope. Folia Morphol (N. Y.) 20: 49-51
Odgen JA (1968) The origin of the coronary arteries. Circulation, Suppl VI:37-38
O'Rahilly R (1971) The timing and sequence of events in human cardiogenesis. Acta Anat 79 : 70-75
O'Rahilly R, Bossy J, Mfller F (1981) Introduction fi l'6tude des stades embryonnaires chez l'homme. Bull Assoc Anat 65:139 236
Patten BM (1956) The development of the ventricular myocardium in relation to its blood supply. Anat Rec 124:344
Patten BM (1960) The development of the heart. In: Gould SE (ed) Pathology of the heart, Ch II, Thomas Ch C, Springfield, USA
Patten BM (1968) Human embryology 3 th Ed, McGraw-Hill Book Company, New York
Patten BM (1970) The development of the ventricular wall and its blood supply. In: Jaffee OC (ed) Cardiac development with special reference to congenital heart disease. Proceedings of the 1968 Int Symp, University of Dayton Press, Dayton, Ohio
Polfi~ek P, Zechmeister A (1968) The occurrence and significance of myocardial bridges and loops on coronary arteries. Opuscula Cardiologica; Acta Fac Med Univ Brunensis 36:5-101
Puerta Fonollfi J, Jim6nez Collado J (1980) Malformation of the venous sinus in human embryos. Acta Anat 106:240245
Roberts JT (1961) Arteries, veins and lymphatic vessels of the heart. In : Luisada AA (ed) Development and structure of the cardiovascular system. McGraw-Hill Book Company, New York
Schulze WB, Rodin AE (1961) Anomalous origin of both coronary arteries. Arch Pathol 72:36M6

218
Streeter GL (1942) Developmental horizons in human embryos. Description of age group XI, 13 to 20 Somites, and age group XII, 21 to 29 Somites. Carnegie Contrib Embryol, Wash 30:211-245
Streeter GL (1945) Developmental horizons in human embryos. Description of age group XIII, embryos about 4 or 5 milli- meters long, and age group XIV, period of indentation of the lens vesicle. Carnegie Contrib Embryol, Wash 31:27-63
Streeter GL (1948) Developmental horizons in human embryos. Description of age groups XV, XVI, XVII, and XVIII, being the third issue of a survey of the Carnegie collection. Carnegie Contrib Embryol, Wash 32:133-203
Van Mierop LHS (1979) Morphological development of the heart. In: Berne RM, Sperelakis N, Geiger SR (eds) Handbook of
physiology. American Physiological Society, Bethesda, Mary- land
Vernall DG (1962) The human embryonic heart in the seventh week. Am J Anat 111 : 17-24
Virfigh S, Challice CE (1981) The origin of the epicardium and the embryonic myocardial circulation in the mouse. Anat Rec 201 : 157-168
Vobo~il Z, Schiebler TH (1969) Uber die Entwicklung der Gef/il3- versorgung des Rattenherzens. Z Anat Entwickl-Gesch 129 : 24~40
Accepted February 14, 1984




























