Embed Size (px)
Citation preview

THE JOURNAL OF COMPARATIVE NEUROLOGY 370551-562 (1996)
Olivocochlear Innervation of Inner and Outer Hair Cells During Postnatal
Maturation: Evidence for a Waiting Period
D.D. SIMMONS, N.B. MANSDORF, AND J.H. KIM Department of Physiological Science (D.D.S., N.B.M., J.H.K.) and the Brain Research
Institute (D.D.S.), UCLA, Laboratory of Electron Microscopy (D.D.S.), House Ear Institute, Los Angeles, California 90095
ABSTRACT Reconstructions of the efferent innervation of the hamster (Mesocricetus auratus) cochlea
were done during postnatal development. Efferent neurons were labeled via injections of biocytin and horseradish peroxidase into the crossed olivocochlear (OC) bundles using an in vitro brainstem technique. Such injections retrogradely labeled cell bodies in ventral periolivary regions of the superior olive consistent with their being medial OC neurons. Anterogradely labeled axons were traced to the cochlea, where they terminated on or below inner hair cells (IHCs) prior to postnatal day 5 (P5). After P5, labeled axons terminated on IHCs and outer hair cells (OHCs) and after P10, the majority of labeled axons terminated on the OHCs. In the electron microscope, small labeled terminals containing densely packed synaptic vesicles were found both adjacent to IHCs (axosomatic) as well as apposed to afferent and efferent fibers below IHCs prior to P5. By P10, large labeled terminals were axosomatic to OHCs and no longer found on IHCs. Consistent with previous reports, these data suggest that medial OC axons form part of an early primary innervation on and below IHCs before terminating on OHCs. This raises the possibility that OC neurons demonstrate a period of waiting below an intermediate target similar to that described in the development of thalamocortical projections. o 1996 Wiley-Liss, Inc.
Indexing terms: efferents, synaptogenesis, development, inner ear, hamster
During the last stages of neuronal development, many neurons undergo a stereotyped pattern of establishing connections and then modifying those connections to achieve an adult pattern of morphology and function. Contact between axons and appropriate target cells is generally referred to as synaptogenesis, which has been studied most extensively at the neuromuscular junction and in the cortex (Lund and Mustari, 1977; Dodd and Jessell, 1988; Licht- man and Balice-Gordon, 1990; Shatz et al., 1990). To form the appropriate pattern of connection, axons branch as they reach their targets and typically contact more target cells than they do in the adult. Some of the initial axonal branches are subsequently modified by being either elimi- nated or redistributed. The underlying mechanism for this modification is believed to be dependent in part on a competition between the axon terminals (Lichtman and Balice-Gordon, 1990). When there is a temporal mismatch between target maturation and incoming axonal projec- tions, it is believed that axons “wait” and accumulate within some intermediate area (Shatz et al., 1990). Little is known about the developmental mechanisms of either synaptogenesis or synaptic modification in efferent olivoco- chlear (OC) neurons as they project to hair cells in the
mammalian cochlea. In adult animals, the brainstem OC system is composed of three populations of neurons that reside mostly within medial and lateral divisions of the superior olivary complex (Rasmussen, 1960; White and Warr, 1983; Lu et al., 1987; Moore and Moore, 1987; Vetter et al., 1991). These OC populations differ in their neuro- transmitters and their synaptic targets within the cochlea. Medial OC neurons contain acetylcholine, project mostly to the contralateral inner ear via the crossed OC bundles, and terminate on the cell bodies of outer hair cells (OHCs), whereas lateral OC neurons contain either acetylcholine or y-aminobutyric acid (GABA), project mostly to the ipsilat- era1 inner ear, and terminate on afferent fibers below the inner hair cells (IHCs; Takeda et al., 1986; Aschoff and Ostwald, 1987; Vetter et al., 1991). In all studies to date, cholinergic lateral OC neurons co-contain calcitonin gene- related peptide (CGRP) which further differentiates them
Accepted February 17,1996 Address reprint request to Dr. Dwayne D. Simmons, UCLA, Physiological
Science, 621 Campus Circle Drive, South Los Angeles, CA 90095. E-mail: [email protected]
O 1996 WILEY-LISS, INC.

552 D.D. SIMMONS ET AL.
from the medial OC population (Kawai et al., 1985; Lu et al., 1987; Kruger et al., 1988; Vetter et al., 1991).
Although several studies present a great deal of informa- tion about the structure of the immature efferent innerva- tion, few studies have attempted to investigate systemati- cally the arrival of the various subpopulations of efferent fibers in the cochlea. Importantly, there have been few challenges to the assumption that the lateral OC efferents are the first to establish synapses. This assumption has arisen on the basis of light- and electron-microscopic obser- vations in adult animals, where lateral OC terminals are found mostly under IHCs, and the medial OC terminals are located mostly, if not exclusively, on OHCs. Thus, several ultrastructural studies in neonatal animals report that since the first efferent synapses observed in the organ of Corti are under IHCs, the lateral OC efferents arrive first (Ginzberg and Morest, 1984; Pujol and Carlier, 1982; Pujol et al., 1979; Pujol and Lavigne-Rebillard, 1985). Obviously, the presence of an efferent terminal making synaptic contact on to an IHC during development does not necessar- ily define its origin. This assumption regarding the origin of the first efferent synapses has been challenged by recent anterograde tracer studies (Simmons et al., 1990; Cole and Robertson, 1992).
The present study focuses on specific postnatal changes of medial OC terminals as they innervate the cochlea of the hamster. The hamster serves as an excellent model for developmental processes because of its short gestation and its immaturity at birth (Hoffman et al., 1968; Stephens, 1972; Pujol and Abonnenc, 1977; Simmons et al., 1990, 1991). Since the cochlea exhibits differential maturation along its spiral length such that regions in the basal cochlea appear more advanced than regions near the apex (Pujol and Abonnenc, 1977), data were analyzed only in the basal portions of the cochlea to minimize maturational variations within a given postnatal age. This study provides evidence that there is an early “waiting” period on and below IHCs, followed by a period of growth and termination on OHCs. Parts of this study have been presented previously in abstract form (Mansdorf et al., 1992).
MATERIALS AND METHODS Data were obtained from hamsters (Mesocricetus aura-
tus) 0 to 20 days old, where the day of birth represents postnatal day 0 (PO). Animals were examined to insure normal (i.e., nonpathological) inner ears before experimen- tal use. Hamsters were anesthetized with near-lethal injec- tions of sodium pentobarbital (Nembutal, 100 mg/kg, i.p.1 prior to experiment.
In vitro labeling Hamsters were perfused intracardially with an oxygen-
ated solution of sterile-filtered Eagle’s minimum essential media (MEM; Sigma Chemical) modified with dextrose (2.7 g/L) and sodium bicarbonate (2.2 g/L). After perfusion, hamsters were decapitated and the dorsal cranium overly- ing the cerebellum removed. This preparation was immedi- ately immersed in a constant temperature (20°C) organ chamber (Radnotti) and bathed continuously with fresh oxygenated media. The central vermis of the cerebellum was ablated to expose the floor of the fourth ventricle. A schematic view of the injection is presented in Figure 1. Once the genu of the facial tracts were visualized, an incision approximately 1-2 mm in depth was made along
the midline from the aqueduct of Sylvius (rostrally) to the facial genu (caudally). Immediately afterwards, a micropi- pette containing a tracer solution (2% biocytin (Sigma Chemical) and 15% horseradish peroxidase (HRP; Type IV, Sigma Chemical) made in Tris buffer, pH 7.4) was inserted into the incision and injected at a rate of 20-50 nl per min for 5-15 min using a nanoliter pump. Four to six hours after injection, brainstems were blocked and immersed in cold 4% paraformaldehyde-lysine-periodate in phosphate buffer (PB) solution. Cochleae were isolated, detached, and immersed in a solution of 2.5% glutaraldehyde and 1% paraformaldehyde in 0.1 M PB (pH 7.4) overnight. Cochleae from hamsters P5 or older were also decalcified in 0.1 M EDTA at 10°C. Cochlear tissue was embedded in either a gelatin-agarose or gelatin-albumin mixture and then sec- tioned perpendicular to the cochlear nerve axis on a Vibra- tome (80-100 pm). Brainstems were cryoprotected in 30% sucrose in PB saline (pH 7.4), immersed in OCT, frozen, and then serially sectioned on a cryostat. Sections were processed free floating for histochemical detection of label as described below. After processing, the cochlear spiral was reconstructed and its length normalized as percent distance from the basal extreme.
Histochemical processing Extracellularly injected fibers and their cell bodies in the
brainstem or terminals in the cochlea were visualized via an avidin-biotin-complex peroxidase method (Vectastain Elite ABC kit, Vector Laboratories, Burlingame, CAI. Sections were incubated for 2 hours with an avidin-conjugated peroxidase solution containing 0.1 M Tris-buffered saline, 1% bovine serum albumin, and 0.3% Triton X-100, followed by extensive washings in Tris-buffered saline. Sections were treated with a diaminobenzidine-glucose oxidase solu- tion (0.025% diaminobenzidine, 1% glucose oxidase, and 0.02% glucose) using a heavy metal intensification proce- dure (Adams, 1981). Sections were then dehydrated, cleared and coverslipped with D.P.X. (Aldrich Chemical, Milwau- kee, WI). Either for thin-section light microscopy or for electron microscopy, sections from basal regions of cochleae were further fixed with a solution of 1% osmium tetroxide, stained en bloc with a saturated solution of uranyl acetate, dehydrated, and embedded in plastic resin (Eponate, Ted Pella, Redding, CA). Once cured, 1.5-pm-thick sections were taken on a microtome. Ultrathin sections (80 nm) were taken on an ultramicrotome, placed on Formvar- coated slot grids that were stained with Reynold’s lead and viewed in a Zeiss 902 transmission electron microscope.
Microscopic analysis Labeled efferent fibers were observed and counted in the
light microscope using either a x50 APO or a x 100 APO (oil immersion) objective lens (Zeiss Aus Jena). Mean values were evaluated by one-way analysis of variance using both the Student’s t-test and the Tukey-Kramer HSD test at an alpha ( P ) level of 0.05.
RESULTS In an attempt to study specifically medial OC neurons,
extracellular injections of tracer were made into the crossed OC bundles located near the floor of the fourth ventricle in the rostra1 medulla. In adult animals, these bundles carry predominantly axons originating from medial OC cell bod- ies (White and Warr, 1983). To verify that medial OC

POSTNATAL OLIVOCOCHLEAR EFFERENTS 553
INJECTION PIPET
n. Genu LATERAL
Fig. 1. Semischematic view of injection and anterograde labeling. Central labeling of the crossed projections of the olivocochlear bundle was achieved by injecting biocytin and horseradish peroxidase (HHP) into a cut along the floor of the IVth ventricle midway between the facial genu (VII n. Genu). Rincytin injections resulted in labeled efferent axons in both cochleae. These injections did not result in labeling of the
facial motor neurons. Labeled axons exited the brainstem with the vestibular portion of the auditory-vestibular nerve (VIII n.) before crossing into the auditory nerve. The following nuclei are listed for point of reference: medial nucleus of the trapezoid body (MNTB), medial superior olivav (MSO), lateral superior ulivary (LSO), trigemi- nal nucleus (VN), cochlear nucleus (CN), and ventral periolivary (WO).
TABLE 1. Anternmade Labeline Data Summarvl
Percent Number of distance Number Number Number Number Number
Postnatal Experiments reconstructed label of OSL of IHC of PC of OHCl of OHC2 age with label experiments traveled fibers swellings projections swellings swellings
P I P3
P4 P5 P6 P7
P9 P11
1 54.9 2 55.3, 92.2
1 63.2 1 82.9 1 58.6 2 69.2, 61.2
1 73.8 1 84.8
60 103 171.5 336
(S.D. = 31.6) (S.D. = 22.6) 78 290
117 292 125 383 140.5 414.5
(S.D. = 37.5) (S.D. = 91.2) 36 163 76 260
0 22
20 36 0 0
0 0
0 0 1 0
0 0 2 0
44 15 110 7
125 43 266 130
(S.D. = 25.5) (S.D. = 7.1)
'The number of cochleae with anterograde label as well as the number of reconstructed cochlea are given. In each experiment, anterogradely labeled fibers were typically found in basal regions, but the extent of apically labeled fibers varied across ages. The cochlear extent over which labeled terminals could be found is given. The remamder of the table gives data on the number of labeled fibers traversing the osseous spiral lamina (OSL), the number of swellings (both terminal and en passant) below inner hair cells (IHCs), pillar cell (PC) projections, and outer hair cell ( O H 0 swellings.
neurons projected through the injection site, brainstems were analyzed for retrograde labeling of OC cell bodies at various postnatal ages. Brainstem nuclei within the supe- rior olive were classified on the basis of a previous report (Simmons and Raji-Kubba, 1993). For anterograde labeling studies, 41 cochleae from 31 hamsters age PO to P20 were used. Although qualitative observations were made from all of these cochleae, only cochleae in which there was no spiral ganglion cell label were used for quantitative analysis. Postnatal day 0 animals, which all displayed spiral ganglion label, were not used for numerical analysis, although light-microscopic observations are included. Since cochlear lengths varied within age groups as well as between ages, complete cochlear reconstructions were performed and
used to define percent distance locations from the basal extremity (0%) of the cochlea. Quantitative analysis was restricted to the basal half (50%) of the cochlear spiral for two reasons. Although anterograde label was always found in the basal 50% of the cochlea, the extent to which labeled fibers were found varied between 55% and 92% of cochlear length, as indicated in Table 1. Additionally, the matura- tional variation along the cochlear spiral was minimized by limiting analysis to the basal half. Of the 41 cochleae studied qualitatively, 10 were reconstructed for quantita- tive analysis (Table 1). This represented a total of 1,116 fibers within the osseous spiral lamina, 2,992 fiber swell- ings below the inner hair cells (IHCs), and 978 swellings below the three rows of outer hair cells (OHCs).

554 D.D. SIMMONS ET AL.
Labeling of the crossed olivocochlear bundle Extracellular injections into the crossed OC bundle re-
sulted in anterogradely labeled efferent fibers terminating in the cochlea and retrogradely labeled cell bodies in the superior olivary complex. Figure 2A shows retrogradely labeled periolivary cells from one such injection in a P5 hamster brainstem. The majority of labeled cell bodies were found in middle to rostral regions of the superior olive, specifically, in periolivary nuclei. As indicated by histo- grams of labeled cells bodies, typically 90% of the labeled cell bodies were found in ventrally located periolivary regions (Fig. 2B). Regardless of the postnatal age, the lateral superior olive contained few (6% or less) labeled cell bodies. Such data was a positive indication that the major- ity of anterogradely labeled fibers and terminals in the cochlea were from medial OC neurons.
The crossed OC bundle labeled fibers could be traced through the genu of the facial nerve tract and exiting the brainstem with the vestibular nerve root (Fig. 1). Labeled fibers were traced through the vestibular-cochlear anasto- mosis as they entered the cochlear nerve. These fibers were typically grouped in fascicles which coalesced as they ran in an anterior direction forming the intraganglionic spiral bundles. As reported previously by Cole and Robertson (1992), spiral ganglion cell bodies and their fibers were consistently labeled in all PO animals and in three out of seven P1 animals. Fibers from labeled spiral ganglion somata were granular in appearance, unlike crossed OC bundle fibers, which had a homogeneous brown-black pre- cipitate. Brainstem sections showed no apparent contamina- tion of the cochlear nucleus in any such cases. Unique to PO and P1 cases, labeled crossed OC bundle fascicles had branches that traversed the spiral ganglion. From P1 to P3, crossed OC bundle injections labeled fibers terminating either on or below IHCs. Some efferent terminal fibers ramified underneath the inner hair cells before contacting them directly, while others coursed further below IHCs giving the impression of at least two spiral bundles. An example of label found in a P 3 cochlea is shown in Figure 3. In this 2-pm cross section of the organ of Corti, labeled terminals are seen contacting the basolateral sides of the
Fig. 2. Retrogradely labeled neurons in the superior olivary com- plex. A A photomicrograph of periolivary cells labeled after crossed olivocochlear injections. Labeled cells are present in the ventral perioli- vary (VPO) region and absent in the lateral superior olive (LSO), where the majority of the lateral olivocochlear neurons are believed to originate. Dorsal (D) and lateral (L) directions are as indicated and the midline is to the right. Scale bar = 50 pm. B: A three-dimensional reconstruction of 13 consecutive sections through the superior olive of a P12 hamster, as discussed in Materials and Methods. Filled circles represent the positions of retrogradely labeled somata. Greater than 90% of the labeled somata are in the ventral periolovary regions. A few somata were labeled in the capsular region outside the lateral superior olive (LSO). Both the nucleus of the trapezoid body (NTB) and the LSO are shown in partial reconstruction. Dorsal (D) and lateral (L) direc- tions are as indicated and the midline (M) is to the left. Scale bar = 1,000 pm. C: A histogram of retrogradely labeled cells in various regions of the superior olive after a crossed olivocochlear injection. The regions are the LSO, VPO, rostral periolivary (RPO), and other periolivary (PO). The data are taken from three postnatal ages and the counts were made from one side (typically contralateral to the right inner ear) of the brainstem. Sections were cut at 50-pm thickness, and, therefore, were not corrected for double-counting errors. Such counts show that the vast majority of the labeled cells were found within the VPO regions in an area commonly referred to as the ventral nucleus of the trapezoid body.
IHC. In general, labeled endings appeared to segregate along either the pillar cell side of the IHC or along the modiolar side (i.e., the opposite side closer to the cochlear nerve) of the IHC. As shown in Figure 4A, although occasional fibers projected past IHCs in the P3 cochlea, the vast majority of efferent fibers ended on or below IHCs and no labeled fibers were seen terminating on or below the OHCs. Prior to P5. numerous efferent terminations and
0
D
1.000 pm
5 12 20
POSTNATAL AGE (DAYS)

POSTNATAL OLIVOCOCHLEAR EFFERENTS 555
Fig. 3. Photomicrograph of crossed olivocochlear (OC) bundle termi- nals at P3. A photomicrograph of a cross section (plastic section, 2-pm thick) of the organ of Corti taken at roughly the 19% distance location. This preparation shows labeled efferent axons that terminate exclu- sively underneath inner hair cells (IHCs) at P3. Arrowheads mark labeled terminations on the modiolar and pillar sides of the IHC as well as labeled fibers below in the neuropil. OHC, outer hair cell; PC, pillar cell. Scale bar = 10 pm.
fibers were found among the pillar cells. From P5 to P7, theFe was a clear decrease in such pillar cell projections and an increase in OHC terminations. A t P6, numerous labeled efferent axons sent thin collaterals past pillar cells that terminated on or below OHCs. Typically, these collaterals terminated under 1 or 2 OHCs in the first row. Figure 4B shows labeled terminals in a P7 cochlea where there are labeled fibers terminating below IHCs and OHCs. Already the terminal swellings are much larger than the tunnel crossing fibers. After P7, there was a dramatic increase in the number of labeled efferent terminations that were found clearly under OHCs. Efferent fibers also exhibited morphological diversity in their hair cell terminations during this transition. Figures 5 and 6 show representative terminal morphology found between P1 and P8. Similar to descriptions by Simmons et al. (19901, single efferent fibers either had terminations on both IHCs and OHCs or termi- nated solely on IHCs or OHCs. Although not quantitatively assessed, such dual innervations of inner and outer hair cells were commonly observed. After P12, large labeled endings were seen mostly on the OHCs as found in the adult cochlea.
To demonstrate the numerical shifts in the number of crossed OC bundle terminations, Figure 7A plots the percentage of labeled swellings (both terminal swellings and en passant swellings) underneath IHCs to the total number of labeled swellings in the organ of Corti. Although the number of labeled fibers in any cochlea is going to be a function of several injection parameters, consistently a greater number of fibers as well as terminals were found at older postnatal ages (Table 1). Labeled endings under IHCs comprised greater than 95% of the total number of labeled endings from P1 through P5 (Fig. 7A). During this same period, the number of labeled fibers traversing the basal osseous spiral lamina doubled while the number of labeled endings underneath IHCs nearly tripled. For example, a P1 cochlea had 60 labeled fibers in the osseous spiral lamina and 103 labeled endings underneath IHCs whereas a P5 cochlea had 117 fibers in the osseous spiral lamina and 292 endings underneath the IHCs. From P6 to P11, although
Fig. 4. Cochlear efferent terminals at P5 and P7. A In the P5 cochlea, efferent terminations are found among the pillar cells. This photomicrograph shows a labeled “pillar cell projection” (asterisk) at approximately the 6% distance location. The termination appears to be on the outer row of pillar cells (OPCs). The inner hair cells (IHCs), inner pillar cells (IPCs), and row one outer hair cells (OHC1) are as labeled. Arrowheads mark en passant swellings commonly observed around the IHCs. Scale bar = 10 pm. B: This photomicrograph of labeled “OHC 1” projections (arrows) was taken at approximately the 12% distance location in the P7 cochlea. Scale bar = 10 pm.
the total number of labeled endings rose from 442 to nearly 800 in individual cochleae (Table l), the percentage of labeled endings underneath IHCs precipitously decreased

D.D. SIMMONS ET AL. 556
P1
I OCB efferent
I OCB efferent
OCB efferent
2 30 pm
I OCB efferent
Fig. 5. Reconstructions of the efferent innervation at PI and P3. These drawings illustrate the typical pattern of labeled terminals found in the cochlea at P1 and P3. During this period, single efferent fibers were injected via the olivocochlear bundle (OCB) and traced from the osseous spiral lamina terminated on or below IHCs, Reconstructions were done using a drawing tube attached to a light microscope. OCB, olivochochlear bundle. Scale bar = 30 wm.
40 Ccm
Fig. 6. Reconstructions of the efferent innervation at P8. These drawings represent the variety of labeled terminal fibers found in the cochlea. During this period, many single olivocochlear bundle (OCB) efferent fibers traced from the osseous spiral lamina terminated on either IHCs (not shown) or OHCs, whereas others had swellings on both IHCs and OHCs. Such dual innervations were commonly ob- served. Reconstructions were done using a drawing tube attached to a light microscope. Scale bar = 40 km.
from 87% to less than 33% (Fig. 7A). Also during this period there was a tremendous increase in the number of en passant swellings and a decrease in the number of terminal swellings under the IHCs.
To show that the decreases in IHC efferent innervation paralleled increases in the OHC innervation, both the number of pillar cell projections and OHC endings were counted (Table 1). In the P1 cochlea, no pillar cell projec- tions were found. In a P3 cochlea, there were 22 pillar cell projections that all terminated well before the OHC region. As the number of pillar cell projections decreased, more terminals were found on OHCs. For example, in the P3 cochlea only one terminal was found well below an OHC (at the 40% distance location), whereas in a P6 cochlea, there were no pillar cell projections but 53 fibers terminated in 59 swellings on or below the OHCs. After P7 the number of OHC terminations increased dramatically. Figure 7B plots the percentage of labeled terminals found on OHCs as a function of postnatal age. At P6 roughly 13% of the total number of swellings were found below OHCs and there was a steady increase to 66% by P11. Not only did more swellings appear under OHCs, but the majority of these crossed OC bundle endings terminated on OHC row 1, as seen in adult animals. As shown in Figure 8, a total of 75% of OHCs swellings were under row 1 OHCs at P6 and 53% at P11. It appears that the first projections and termina-
tions to the OHCs were directed to OHCs in row 1 as previously suggested (Simmons et al., 1990).
Electron microscopy Electron-microscopic investigations within the organ of
Corti further suggest that the early postnatal crossed OC bundle terminations actually make contact with IHC so- mata. In in vitro preparations that were not processed for peroxidase reaction product, efferent terminals can be readily identified contacting IHC somata, as shown by Figure 9A. Similar to efferent terminals on OHCs, these IHC-efferent contacts are characterized by densely packed vesicles, few mitochondria, asymmetric membrane density (thickening), and within the hair cell, a subsynaptic cister- nae. In P1 and P3 animals, labeled crossed OC bundle terminals contacted fibers below IHCs as well as terminat- ing on IHC somata. Nearly all such terminals were ob- served to have a dense population of synaptic vesicles. Contact with IHCs was frequently found on the basolateral portion of the hair cell soma as shown in Figure 9B. In nearly all cases, the presence of membrane specializations was difficult to visualize because of the dark reaction product; however, subsynaptic cisternae were occasionally seen (not shown in Fig. 9B). As seen in the light microscope, the majority of labeled endings contacted the pillar cell side of the IHC (see Fig. 3). By P7, labeled endings contacting IHCs became increasingly more difficult to find. By P11, labeled contacts with the IHC were not found.

POSTNATAL OLIVOCOCHLEAR EFFERENTS
80
U z 70 g 2 60 c ) w g 3 50 5: 40 $J$ 30
m u
= g 20
557
B
1 3 4 5 6 7 9 11
POSTNATAL AGE (DAYS)
Fig. 7. Percentages of swellings on IHCs and OHCs as a function of postnatal age. A The percentage of labeled swellings (both en passant and terminal swellings) found underneath IHCs as a function of postnatal age. The total number of swellings found in the basal 50% of the organ of Corti were counted in at least one cochlea per age. B: The percentage of labeled swellings found underneath OHCs as a function of postnatal age. Same cochlea used as in A.
E W
OHC 1 Swellings
OHC 2 Swellings
- OHC3Swellings
250-
200-
150-
PO P1 P3 P4 P5 P6 P7 P9 P11
POSTNATAL AGE (DAYS) Fig. 8. The number of labeled swellings found as a function of OHC
row and postnatal age in the basal 50% of the cochlea.
As in our light-microscopic observations, the decline in crossed OC bundle fibers contacting IHCs was paralleled by an increase in the OHC innervation. By P7, labeled, vesicu-
lated terminals were readily identified contacting OHC somata in the first OHC row. Such contacts were always along the basal portion of the OHC and differed from the IHC-efferent contact in their much larger size. By P9, labeled crossed OC bundle contacts were found on OHCs from all three OHC rows. Figure 10 depicts efferent contacts on OHCs at P10 and P11 from in vitro preparations. In nonhistochemically processed preparations, synaptic special- izations were clearly visible (Fig. 10A). Even though labeled cases during the second postnatal week were subject to anoxic conditions, clearly labeled terminals were visible (Fig. 10B); however, as seen under IHCs, these labeled OHC terminals lacked the synaptic specializations seen in cases without histochemical processing.
DISCUSSION The results of anterograde labeling in the present study
are consistent with the existence of an efferent waiting period below IHCs in the basal portions of the postnatal hamster cochlea. Table 2 summarizes the percentage of labeled terminals below inner and outer hair cells resulting from crossed OC bundle injections. From P1 to P4, antero- gradely labeled crossed OC bundle efferents form a dense plexus of terminations below IHCs. From P5 to P9, large numbers of anterogradely labeled crossed OC bundle fibers terminate on IHCs as well as OHCs. After P10, the number of labeled crossed OC bundle terminals dramatically de- crease under IHCs and increase under OHCs. Data from a companion study on the immunocytochemical development of proteins associated with growth and synapses generally supports the interpretation of the anterograde data (Sim- mons et al., 1996). Growth and synaptic proteins appear to be expressed below IHCs throughout the first two postnatal weeks (PO-P14). However, below OHCs both growth and synaptic proteins appear only after P5.
Medial versus lateral OC projections This study has attempted to address the origin of efferent
fibers and their terminations in the cochleae of neonatal animals. The results suggest that the majority of fibers initially contacting IHCs in the postnatal cochlea are from the crossed OC bundle, and thus, most likely represent the terminations of medial OC neurons. In contrast, the over- whelming majority of studies in the neonate cochlea as- sume that the lateral OC neurons are the first efferents to establish synapses in the cochlea (Shnerson et al., 1982a,b; Despres et al., 1988; Gil-Loyzaga and Pujol, 1988; Emmer- ling et al., 1990; Merchan-Perez et al., 1990, 1993). This assumption is based almost entirely on early electron- microscopic studies in newborn animals that show efferent synapses specifically on IHCs (Kikuchi and Hilding, 1965; Ginzberg and Morest, 1984; Pujol and Carlier, 1982; Pujol et al., 1979; Pujol and Lavigne-Rebillard, 1985). Since the first efferent fibers to reach the organ of Corti terminate on IHCs, they have been assumed to be from lateral OC neurons as is the case in the adult. At least two other recent studies in rodents agree, however, with the data presented here. In the hamster, Simmons et al. (1990) suggest that there may be multiple fiber types that project into the organ of Corti at different stages of development. Based on extracellular HRP injections into the eighth nerve, a popu- lation of thick nonvaricose efferent fibers made termina- tions directly on to IHCs during the first three postnatal days (Simmons et al., 1990). Similar injections at later postnatal ages showed a progression where thick nonvari-
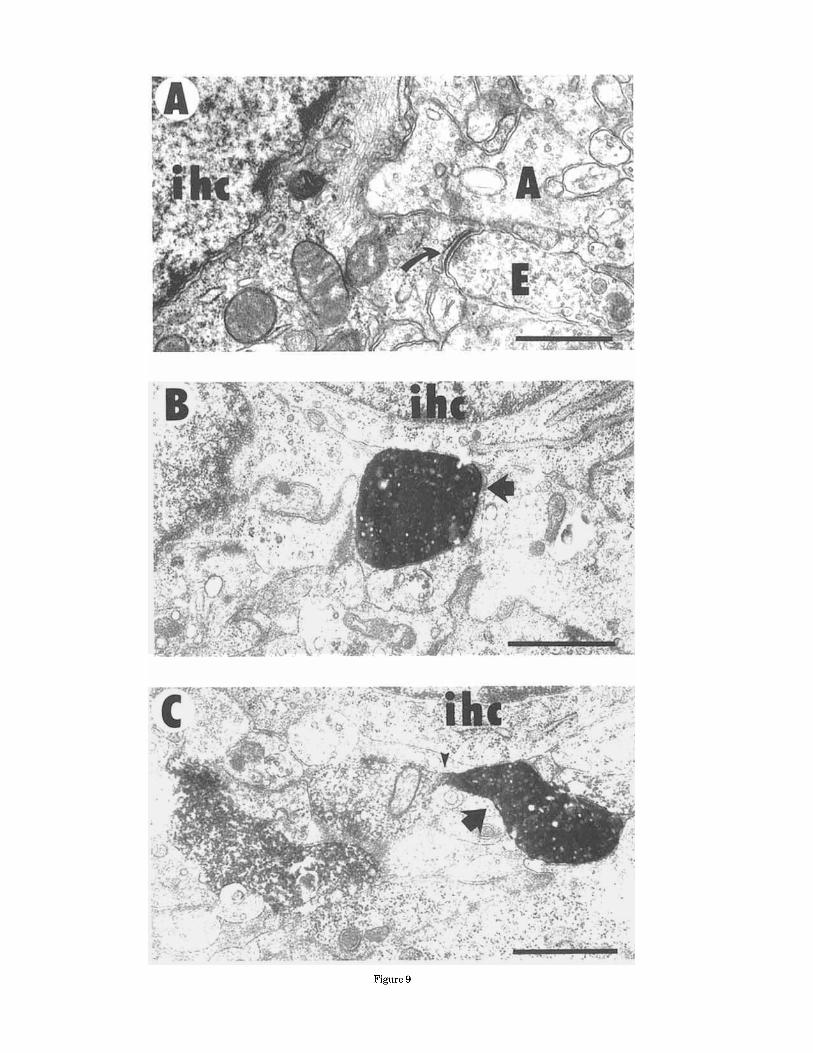
Figure 9

POSTNATAL OLIVOCOCHLEAR EFFERENTS 559
TABLE 2. Chronology of Labeling Below Hair Cells’
P1 P3 P4 P5 P6 P7 P9 P11
LabelunderIHC 100% 99.7% 100% 99.3% 87% 78% 47% 34% LabelunderOHC 0% 0.3% 0% 0.7% 13% 22% 53% 66%
‘The percentage of the total labeled swellings (both terminal and en passant) found below inner and outer hair cells is given in chronological fashion.
cose efferent fibers had dual terminations on IHCs and OHCs, and then at later postnatal ages only on OHCs similar to the adult. Cole and Robertson (1992) applied DiI to the cut central ends of crossed OC bundle axons in neonatal rats. As a result, crossed OC bundle fibers were labeled and their terminations observed in the organ of Corti. Cole and Robertson (1992) found that efferent fibers had massive projections to IHCs first (Pl-P2), and then at later ages (PPP6) projected to OHCs. These studies com- bined with the present study strongly support the notion that postnatal efferent fibers from the crossed OC bundle stop for a period of time below IHCs before going on to terminate on OHCs. Consistent with the anterograde label- ing data, our study also suggests the need to reinterpret the results using neurochemical markers from other studies of cochlear development.
Our observations in the electron microscope show not only that labeled crossed OC bundle terminal profiles are vesiculated but that they also directly contact IHC somata. This data is consistent with light-microscopic observations that show distinct en passant and terminal swellings contact- ing IHC somata. However, whether crossed OC bundle terminals synapse with IHCs remains an open question since we could not definitively identify synaptic specializa- tion. More extensive investigation at the electron-micro- scopic level is needed to demonstrate that crossed OC bundle terminals actually give rise to the early efferent synapses seen in normal preparations of the same age as well as reported previously in neonatal ultrastructural studies. It is quite plausible that some crossed OC bundle fibers project to the IHC region without making synaptic specializations. The lack of synaptic specialization could imply that their terminals “wait” beneath IHCs for several days before progressing to their final OHC targets. Unsus- tained “waiting periods” are not likely. Although axonal waiting periods of different durations have been described in the development of thalamocortical projections, it is believed that these waiting periods all involve synaptic connections with intermediate targets before synapses are made onto final targets (Lund and Mustari, 1977; Rakic,
Fig. 9. Electron micrographs of terminals contacting IHCs At P3, A: afferent (A) and efferent (E) terminals contacting an IHC. The efferent terminal has densely packed vesicles and the curved arrow identifies the synaptic specializations typically associated with efferent synapses: membrane specializations and within the hair cell a cistern. This control preparation was not processed for peroxidase histochemis- try but was otherwise treated similarly to other in vitro preparations. Scale bar = 0.5 pm. B: At P1, labeled efferent terminals contacted IHCs, In this micrograph, a labeled efferent terminal is filled with translucent vesicles and forms a contact with the inner hair cell although no junctional specializaion can be seen. The arrow indicates labeled ending. Scale bar = 1 pm. C: Afferent and efferent (e) terminals contact this IHC from the P3 cochlea. The labeled efferent terminals are filled with translucent vesicles. Arrows indicate labeled endings. The large arrowhead identifies a constriction in the efferent terminal and demonstrates the range ofvariability in the quality of the label even within a single terminal. Scale bar = 1 pm.
1977; Shatz et al., 1990). Currently, it is not known why ingrowing axons sometimes wait before innervating their final synaptic targets, it has been hypothesized that the “waiting period” is necessary for the final maturational processes of the target cells.
If crossed OC bundle terminals make functional synaptic junctions with IHCs, then this could explain otherwise puzzling data regarding the effects of early efferent stimula- tion in the kitten. During a period when efferent fibers should be terminating almost exclusively on IHCs in neona- tal cats (Pujol et al., 19801, Carlier and Pujol (1976) were able to stimulate electrically the crossed OC bundle in the brainstem and observe short latency increases in the co- chlear potentials similar to the adult. This result was originally interpreted as due to the axodendritic lateral efferent synapses that are the first efferent connections established in the cat organ of Corti (Carlier and Pujol, 1976; Merchan-Perez et al., 1990). However, in adult animals, such responses are due to the direct termination of crossed OC bundle fibers on OHCs. In light of the present data, it raises the possibility that the crossed OC bundle might be capable of transmitting information directly to IHCs and afferent fibers which in turn are capable of generating early postnatal responses.
Lateral OC terminals The presence of labeled terminals under IHCs from
crossed OC bundle injections does not preclude the pres- ence also of ipsilaterally projecting terminations from lat- eral OC neurons. Previous studies indicate that lateral OC neurons may project to the cochlea during the same time as medial OC neurons (Robertson et al., 1989; Simmons et al., 1996). In a study by Robertson et al. (1989), fluorescent tracers were injected into cochleae from rats ranging in age from PO to adult. Not only were the total number of labeled OC cell bodies not statistically different in the newborn rat compared to the adult, but there was also a clear separation of OC cells into lateral and medial efferent groups. These results indicate that both sets of axons are at least present in the rat cochlea beginning at PO. A study by Simmons et al. (1990) also suggests at least two populations of efferent fibers exist at birth in the hamster cochlea. Although they were only able to trace large-diameter axons to their terminations on hair cells, Simmons et al. (1990) routinely labeled both thick and thin diameter axons in the vestibular- cochlear anastomosis and the intraganglionic spiral bundle. Presumably, the thin efferents were lateral OC fibers. Whether or not they terminated below or on IHCs remains to be documented.
Thus, there could be several populations of efferent terminals found under IHCs in the present study: terminal fibers that are consistent with growing axons under IHCs, and terminal fibers that have characteristics of mature synapses under IHCs.
Model for innervation by medial OC axons Based on our observations using anterograde tracer
techniques, we hypothesize that the medial OC neurons are present underneath IHCs and then make terminations on OHCs. A proposed innervation scheme by medial OC efferents in the basal cochlea is presented in Figure 11 that combines the results of this study as well as earlier investi- gations (Simmons et al., 1990; Cole and Robertson, 1992). In this scenario there are three phases: 1) medial OC neurons innervate IHCs initially, 2) go through a transi-

Fig.
10.
E
lect
ron
mic
rogr
aphs
of t
erm
inal
s co
ntac
ting
OH
Cs
at P
10. A
An
elec
tron
mic
ro-
grap
h of
a ro
w 1
OH
C w
ith e
ffer
ent (
arro
whe
ad) a
nd a
ffer
ent t
erm
inal
s. A
s is t
ypic
al o
f mos
t hai
r ce
ll-ef
fere
nt s
ynap
ses,
the
re is
an
obvi
ous
subs
ynap
tic c
iste
rna
(ast
eris
k) a
djac
ent t
o th
e O
HC
m
embr
ane.
Thi
s m
icro
grap
h is
from
an
in v
itro
cont
rol c
ase
that
was
sub
ject
ed to
roug
hly
4 h
surv
ival
in v
itro,
then
imm
edia
tely
imm
erse
d in
fixa
tive,
and
pre
pare
d fo
r ele
ctro
n m
icro
scop
y
with
out h
isto
chem
ical
pro
cess
ing.
Sca
le b
ar re
pres
ents
0.5
pm
. B: A
n e
lect
ron
mic
rogr
aph
of a
ro
w 1
OH
C w
ith a
labe
led
effe
rent
term
inal
(ar
row
head
). A
s de
pict
ed b
y th
e as
teri
sks,
ther
e is
so
me
dem
onst
ratio
n of
pre
syna
ptic
mem
bran
e sp
ecia
lizat
ion.
How
ever
, the
re is
no
subs
ynap
tic
cist
erna
seen
in th
e O
HC
. Sca
le b
ar =
0.5
pm
.

POSTNATAL OLIVOCOCHLEAR EFFERENTS 561
SUPERIOR OLIVARY COMPLEX
ORGAN OF CORTI
P1- P4
P 5 - P 9
HCs
P 10+
Fig. 11. Proposed postnatal innervation of scheme. In this scenario there are three phases: 1) medial OC neurons innervate the IHC region where they accumulate over the period from P1 to P4; 2) medial OC neurons go through a transitional stage innervating both IHCs and OHCs during the period from P5 to P9; and 3) medial OC neurons become restricted to OHCs after P10. Depicted are the nuclei of the superiur ulivary complex: nucleus of the trapezoid body (NTBj, medial superior olive (MSO), and lateral superior olive (LSO). Medial OC
tional stage where they innervate both IHCs and OHCs, and 3) then become restricted to OHCs. If this scenario is correct, it raises several questions. For example, what leads to the first synaptogenic event, what induces medial OC fibers to cross the pillar cells and seek OHC targets, what induces synaptic modifications, and what roles do the cholinergic and GABA-ergic lateral efferents have? To understand the mechanism(s) responsible for this behavior, it is essential to learn more about the cellular environment in which the OC axons wait below IHCs.
The efferent OC system may adhere to a pattern of development found in other neurons, that is, exuberant axonal branching as they reach their initial targets, and subsequent terminal modification either via elimination or redistribution. Axonal competition has been suggested as the underlying mechanism for such modification of axon terminals. Competition for synaptic sites among like neu- rons is a well-known mechanism used by developing motor neurons (Perry and Maffei, 1988; Balice-Gordon and Licht- man, 1993). It is tempting to speculate that there may be interaction between medial and lateral efferent branches or terminals that somehow results in efferent synaptic remod- eling. Whether such competition actually exists and whether it occurs between medial and lateral or lateral and lateral OC neurons would be worth further investigation.
In vitro retrograde labeling studies and glutamate toxic- ity studies of spiral ganglion neurons suggest that there may be transient terminal modifications among some affer- ent fibers that overlap with the efferent transition from IHCs to the OHCs (Gil-Loyzaga and Pujol, 1990; Simmons, 1994). For example, there is evidence that between P4 and P6, radial afferent fibers also terminated on OHCs. These
Tunnei\of Corti
neurons (MOCj are located in periolivary regions as illustrated and project via the crossed OC bundle (COCB) to the organ of Corti. The relative size of the LSO dramatically increases as a function of postnatal age (Simmons and Raji-Kubba, 1993). From P1 to P4, there is no tunnel of Corti and the inner and outer pillar cells are apposed as shown. From P5 to P9, the tunnel of Corti begins to form and is adult-like by P10. Also there is a general increase in the size of' hair cells as well as intercellular spaces between OHCs from P1 to P10.
radial afferent terminations are believed to be transient since they were not observed either before or after this postnatal period. Not only does the transient OHC afferent fiber period overlap with the massive innervation of OHCs by the crossed OC system, but they both appear to termi- nate preferentially on OHCs in row 1. Additional evidence for transient afferent projections comes from degeneration studies of radial afferent fibers by Pujol and colleagues (Pujol and Lavigne-Rebillard, 1985; Gil-Loyzaga and Pujol, 1990). Their work suggests that glutamate-sensitive afTer- ent fibers exists only beneath IHCs in adult animals but during development are also found beneath OHCs. To- gether these observations raise the possibility that tran- sient afferent projections may somehow interact with the efferent innervation. It was originally proposed that there must be competition between afferent and efferent fibers (Pujol et al., 1978). This idea was further supported by studies in kittens in which the OC efferents were lesioned at birth and the normal decrease in afferent terminals below OHCs did not occur (Pujol and Carlier, 1982). However, whether a competition is actually the mechanism used to displace the OHC afferent fibers is questionable since such efferent-afferent competition is not presently known to exist in other parts of the nervous (Perry and Maffei, 1988; Lichtman and Balice-Gordon, 1990). An alternative to efferent-afferent competition is that the afferent axons somehow serve to guide efferent axons, as has been pro- posed in the development of thalamocortical projections initially to subplate neurons and then to layer 4 cortical neurons (Shatz et al., 1990). The presence of axonal processes that temporarily connect to OHCs in row 1 suggests that afferent axons could establish a neural scaf-

562 D.D. SIMMONS ET AL.
fold or framework by which efferent axons complete their development (Simmons, 1994).
ACKNOWLEDGMENTS The authors thank Ms. J. Kubba and Dr. C . Bertolotto for
outstanding technical contributions as well as Dr. J.K. Moore for valuable comments and criticisms. This research was funded in part by grants from the NIDCD (R29 DC01777) and the UCLA Academic Senate.
LITERATURE CITED Adams, J.C. (1981) Heavy metal intensification of DAB-based HRP reaction
product. J. Histochem. Cytochem. 29t775. Aschoff, A. and J. Ostwald (1987) Different origins of cochlear efferents in
some bat species, rats, and guinea pigs. J. Comp. Neurol. 264t56-72. Balice-Gordon, R.J. and J.W. Lichtman (1993) In vivo observations of pre-
and postsynaptic changes during the transition from multiple to single innervation at developing neuromuscular junctions. J. Neurosci. 13.434- 855.
Carlier, E. and R. Pujol (1976) Early effects of efferent stimulation on the kitten cochlea. Neurosci. Letts. 3.21-27.
Cole, K.S. and D. Robertson (1992) Early efferent innervation of the developing rat cochlea studied with a carbocyanine dye. Brain Res. 575.223-230.
Despres, G., N. Giry, and R. Romand (1988) Immunohistochemical localisa- tion of nerve growth factor-like protein in the organ of Corti of the developing rat. Neurosci. Letts. 85t5-8.
Dodd, J. and T.M. Jessell (1988) Axon guidance and the patterning of neuronal projections in vertebrates. Science 242:692499.
Emmerling, M.R., H.M. Sobkowicz, C.V. Levenick, G.L. Scott, S.M. Slapnick, and J.E. Rose (1990) Biochemical and morphological differentiation of acetylcholinesterase-positive efferent fibers in the mouse cochlea. J. Electr. Micros. Tech. 15:123-143.
Gil-Loyzaga, P. and R. Pujol(1988) Synaptophysin in the developingcochlea. Int. J. Devl. Neurosci. 6,155-160.
Gil-Loyzaga, P. and R. Pujol (1990) Neurotoxicity of kainic acid in the rat cochlea during early developmental stages. Eur. Arch. Otorhinolaryngol. 248t4048.
Ginzberg, R.D. and D.K. Morest (1984) Fine structure of cochlear innerva- tion in the cat. Hearing Res. 14t109-127.
Hoffman, R.A., P.F. Robinson, and H. Magalhaes (1968) The Golden Syrian Hamster, Its Biology and Use in Medical Research. Ames, I A Iowa State University Press.
Kawai, Y., K. Takami, S. Shiosaka, P.C. Emson, C.J. Hillyard, S. Girgis, I. MacIntyre, and M. Tohyama (1985) Topographic localization of calcito- nin gene-related peptide in the rat brain: An immunohistochemical analysis. Neuroscience 15747-763.
Kikuchi, K. and D. Hilding (1965) The development of the organ of Corti in the mouse. Acta Otolaryngol. 6Ot207-222.
Kruger, L., P.W. Mantyh, C. Sternini, N.C. Brecha, and C.R. Mantyh (1988) Calcitonin gene-related peptide (CGRP) in the rat central nervous system: Patterns of immunoreactivity and receptor binding sites. Brain. Res. 463.223-244.
Lichtman, J.W. and R.J. Balice-Gordon (1990) Understanding synaptic competition in theory and in practice. J. Neurobiol. 1:99-106.
Lu, S.M., L. Schweitzer, N.B. Cant, and D. Dawbarn (1987) Immunoreactiv- ity to calcitonin gene-related peptide in the superior olivary complex and cochlea of cat and rat. Hearing Res. 31t137-146.
Lund, R.D. and M.J. Mustari (1977) Development of the geniculocortical pathway in rats. J. Comp. Neurol. 173339-306.
Mansdorf, N.B., D.D. Simmons, and K. Bell (1992) Efferent terminations in the neonatal hamster cochlea. SOC. Neurosci. Abstr. 18t1193.
Merchan-Perez, A., P. Gil-Loyzaga, and M. Eybalin (1990) Immunocytochemi- cal detection of calcitonin gene-related peptide in the postnatal develop- ingrat cochlea. Int. J. Devl. Neurosci. 8:603-612.
Merchan-Perez, A,, M.V. Bartolome, M.A. Ibanez, and P. Gil-Loyzaga (1993) Expression of GAP-43 in growing efferent fibers duringcochlear develop- ment. J. Oto-Rhino-laryngol. 55t208-210.
Moore, J.K. and R.Y. Moore (1987) Glutamic acid decarboxylase-like immu- noreactivity in brainstem auditory nuclei of the rat. J. Comp. Neurol. 26Ot157-174.
Perry, V.H. and L. Maffei (1988) Dendritic competition: Competition for what? Dev. Brain Res. 41:195-208.
Pujol, R. and M. Abonnenc (1977) Receptor maturation and synaptogenesis in the golden hamster cochlea. Arch. Oto-Rhino-Laryngol. 21 7tl-12.
Pujol, R., E. Carlier, and C. Devigne (1978) Different patterns of cochlear innervation during the development of kitten. J. Comp. Neurol. 177t529- 536.
Pujol, R., E. Carlier, and C. Devigne (1979) Significance of presynaptic formations in early stages of cochlear synaptogenesis. Neurosci. Letts. 15:97-102.
Pujol, R., E. Carlier, and M. Lenoir (1980) Ontogenetic approach to inner and outer hair cell function. Hearing Res. 2t423-430.
Pujol, R. and E. Carlier (1982) Cochlear synaptogenesis after sectioning the efferent bundle. Dev. Brain Res. 3~151-154.
Pujol, R. and M. Lavigne-Rebillard (1985) Early stages of innervation and sensory cell differentiation in the human fetal organ of corti. Acta Otolaryngol. (Stockh.). Suppl. 423:43-50.
Pujol, R., M. Lenoir, D. Robertson, M. Eybalin, and B.M. Johnstone (1985) Kainic acid selectively alters auditory dendrites connected with cochlear inner hair cells. Hearing Res. 18t145-154.
Rakic, P. (1977) Prenatal genesis of connections subserving ocular domi- nance in the rhesus monkey. Nature 261t467-471.
Rasmussen, G.L. (1960) Efferent fibers of the cochlear nerve and the cochlear nucleus. In G.L. Rasmussen and W.G. William (eds): Neural Mechanisms of the Auditory and Vestibular Systems. Springfield, IL: Charles C. Thomas, pp. 105-115.
Robertson, D., A.R. Harvey, and K.S. Cole (1989) Postnatal development of the efferent innervation of the rat cochlea. Dev. Brain Res. 47:197-207.
Shatz, C.J., A. Ghosh, S.K. McConnell, K.L. Allendoerfer, E. Friauf, and A. Antonini (1990) Pioneer neurons and target selection in cerebral cortical development In: The Brain. Plainview: Cold Spring Harbor Laboratory Press, pp. 469480.
Shnerson, A., C. Devigne, and R. Pujol (1982a) Age-related changes in the C57BL/6J mouse cochlea: 11. Ultrastructural findings. Dev. Brain Res. 2: 77-82.
Shnerson, A,, C. Devigne, and R. Pujol (198213) Age-related changes in the C57BL/6J mouse cochlea: I. Physiological findings. Dev. Brain Res. 265-75.
Simmons, D.D., L. Manson-Gieseke, T.W. Hendrix, and S. McCarter (1990) Reconstructions of efferent fibers in the postnatal hamster cochlea. Hearing Res. 49t127-140.
Simmons, D.D., L. Manson-Gieseke, T.W. Hendrix, K. Morris, and S.J. Williams (1991) Postnatal maturation of spiral ganglion neurons: A horseradish peroxidase study. Hearing Res. 55t81-91.
Simmons, D.D. and J. Raji-Kubba (1993) Postnatal calcitonin gene-related peptide in the superior olivary complex. J. Chem. Neuroanat. 6t407-418.
Simmons, D.D. (1994) A transient afferent innervation of outer hair cells in the postnatal cochlea. NeuroReport 5 no.ll:1309-1312.
Simmons, D.D., H.D. Moulding, and D. Zee (1996) Olivocochlear innervation of inner and outer hair cells duringpostnatal maturation: An immunocy- tochemical study. Dev. Brain Res., (In press).
Stephens, C.B. (1972) Development of the middle and inner ear in the golden hamster. Acta Otolaryngol. (Suppl.) 296:l-51.
Takeda, N., M. Kitajiri, S. Girgis, C.J. Hillyard, I. MacIntyre, P.C. Emson, S. Shiosaka, M. Tohyama, and T. Matsunaga (1986) The presence of a calcitonin gene-related peptide in the olivocochlear bundle in rat. Exp. Brain. Res. 61t575-578.
Vetter, D.E., J.C. Adams, and E. Mugnaini (1991) Chemically distinct rat olivocochlear neurons. Synapse 72.2143.
White, J.S. and W.B. Warr (1983) The dual origins of the olivocochlear bundle in the albino rat. J. Comp. Neurol. 219r203-214.