Embed Size (px)
Citation preview

GENETIC MODULATION OF RNA METABOLISM IN DROSOPHILA. I. INCREASED RATE OF RIBOSOMAL RNA SYNTHESIS
STEPHEN H. CLARK*, LINDA D. STRAUSBAUGH AND BARRY I. KIEFER
Department of Biology, Wesleyan Uniuersity, Middletown, Connecticut 06457
Manuscript received February 25, 1977 Revised copy received April 27, 1977
ABSTRACT
It has been suggested that a particular Y chromosome which is rDNA- deficient ( Y b b S u V a r - 5 ) may be associated with an increased utilization of rDNA template in adult testes (SHEXMOEN and KIEFER 1975). To extend the observa- tions on this chromosome, experiments were designed to determine if the chromosome has an effect on rRNA synthesis in bobbed adults and on classic bobbed phenotypes (shortened and thinner scutellar bristles and delayed development). Specific activity measurements were made on rRNA extracted from adult males of the genotypes car bb/Ybb- and car bb/YbbSuVar-J, which are rDNA-deficient to the same extent, and from Samarkand+ isogenic (Sam+ iso), which is a wild-type stock. The resulting data demonstrated that the pres- ence of the YbbSuVar-5 chromosome increases the rate of ribosomal RNA syn- thesis in adult flies. In addition, it was found that the presence of this particular Y chromosome restores wild-type bristle phenotype and development time. Appropriate genetic crosses indicate that the observed effects (increased rRNA synthesis, restoration of wild-type phenotype) are a function of this par- ticular Y chromosome, and are not due to autosomal factors. The results of these experiments suggest that the rate of rRNA accumulation is under genetic control.
HE present approach to an understanding of the mechanisms involved in Tcell differentiation has at its base the premise that gene activity is selectively and sequentially regulated. The ultimate level at which this selectivity can be achieved is in the restricted synthesis of an RNA product from a DNA template. With respect to the transcription of a particular species of RNA in a given cell at a given time, there are essentially four types of regulatory events possible: (1 ) initiation of transcription, (2) increase in rate of transcription, (3) decrease in rate of transcription, and (4) termination of transcription. These could either represent different manifestations of a single regulatory mechanism, or require as many as four different control systems. While there is a considerable amount of both direct and indirect experimental support for the existence of events (1) and (4), there is surprisingly little evidence that the rate of transcription of an active gene can be altered by genetic or developmental circumstances.
In Drosophila melanogaster the synthesis of ribosomal RNA (rRNA) and the
* Present address: Genetics and Cell Biology Section, Biological Sciences Group, University of Connecticut, Storrs. Connecticut 06268.
Genetics 86: 789-800 August, 1977.

790 S. H. CLARK, L. D. STRAUSBAUGH AND B. I . KIEFER
bobbed (bb) phenotype associated with partial deletions of the genes coding for rRNA (rDNA) has been the subject of several studies beginning with the pioneering work of RITOSSA and SPIEGELMAN (1965). The bobbed mutants are typically characterized as having shorter and thinner bristles, and a longer period of development when compared to wild type. We have utilized this system to investigate the relation of the rate and amounts of rRNA synthesis to the amount of rDNA present (SHERMOEN and KIEFER 1975), and the effects of rDNA dele- tions on a specific differentiation process: spermatogenesis ( KIEFER and SHER- MOEN 1973). In the previous investigation (SHERMOEN and KIEFER 1975). data were presented that suggested that a particular Y chromosome, designated YbbSuVar--li, may be associated with an increased utilization of rDNA template in adult testes, and with the restoration of wild-type bristle phenotype to a bb genotype. The purpose of this communication is to report additional observations on the effects of this chromosome. The experiments described here have demon- strated that: (1 ) the restoration of wild-type bristle phenotype in bb genotypes is a function of the presence of the YbbsuVar-5 chromosome; (2) the increase in rates of transcription due to the YbhSuVar--li chromosome can be demonstrated in whole flies as well as in testes; ( 3 ) the increase in rate of transcription in whole flies containing YbbSuVar--li cannot be attributed to elevation of gene number; and (4) the presence of the YbbSuVar-5 chromosome can restore wild-type development patterns in bb genotypes.
MATERIALS AND METHODS
Genetics: Drosophila cultures were maintained at 25" 2 1" on modified instant Drosophila medium (Carolina). Most of the F, males used in these experiments were obtained by crossing Samarkand isogenic wild type (Sam+ iso), wmA/Ybb-, or wm4/YbbSuVa7-5 males with the follow- ing females: d1-49/y sc4-sc8; y w bb/y w bb; car bb/car bb; gt b b l / C l B ; Sam+ iso. The details of these chromosomes are given in LINDSLEY and GRELL (1968). F, males are used in all experi- ments to avoid potential problems with rDNA magnification (RITOSSA 1968). In addition, the Y chromosomes used in these studies were placed in homogeneous and additional heterogeneous genetic backgrounds for comparison. All mutant Y chromosomes were routinely checked against a sc4-sc8 chromosome.
Development time: Culture bottles were started with 3040 pairs of adult flies and raised at 25" & 1". After four days these parents were transferred to fresh medium and the females were allowed to lay eggs for 12 hours before cultures were cleared of all adults. Ten days after this transfer the second set of bottles was scored for the appearance of adults. Bottles were scored every eight hours until no further adults emerged.
Bristle measurements: Scutellar bristles of adult flies were measured with a Bausch and Lomb dissecting microscope equipped with a 2>< supplementary lens allowing magnifica- tion of 50x. An ocular micrometer disc with 20 pM divisions was used for the measurements.
RNA quantification: RNA was extracted from adult flies using a modified Schmidt-Thann- hauser (1945) protocol. Thirty adult flies were homogenized in 5 ml of 10% perchloric acid, using a Teflon-glass homogenizer. The homogenate was allowed to sit on ice for 30 minutes to precipitate the macromolecules. A pellet was collected by centrifugation in a table-top I.E.C. centrifuge. The pellet was then washed with 5 ml of 10% perchloric acid and placed for another 10 minutes on ice before collecting the pellet by centrifugation. The pellet was next washed twice with 2 ml of 2:l methano1:chloroform solution. The resulting pellet was then suspended in 1 ml of 0.3 M KOH and incubated overnight at 37". To this hydrolysate was added 0.1 ml of 5 N HC1 and 0.9 ml 20% perchloric acid, and the mixture placed on ice for 30 minutes. This

INCREASED RATE OF rRNA SYNTHESIS 79 1 precipitated the DNA and proteins and left the hydrolyzed RNA in solution. The mixture was then centrifuged, and the supernatant carefully removed for UV determination of RNA. RNA optical density measurements were converted to pg quantities, using the formula described by LEVY, SIMPSON and SOBER (1972).
Incorporation time course studies: To determine the kinetics of incorporation of RNA pre- cursors into nucleic acid, flies were injected with labeled precursor (3H-uridine, 1 pC/pl in Ringer's, approximately 0.25 &fly) and allowed to incorporate for fixed periods of time. After designated incubation times, groups of 10 flies were homogenized in 1 ml of RNA extraction buffer (0.1 M NaC1, 0.1 M NaAc, 0.01 M EDTA, 1% SDS, and 5 mg/ml purified Bentonite). Two 100 pl duplicates were precipitated with 5% ice-cold trichloroacetic acid for 30 minutes on ice, and precipitates were collected on 2.5 cm glass fiber filters (Reeve Angel, grade 934 AH). Filters were then dried for at least two hours at 50". Dried filters were placed in scintillation ~ i a l s to which 5 ml of scintillation cocktail (100 mg POPOP, 4 g PPO to 1 liter with scintillation grade toluene) was added. Samples were chilled and counted in a Packard liquid-scintillation counter.
Rate studies: The technique for the injection of isotope, extraction of RNA, and gel electro- phoresis of testis RNA is presented elsewhere (SHERMOEN and KIEFER 1975). For measurements of whole fly RNA synthesis, tritiated RNA precursors in sterile Drosophila Ringer's were injected into the abdomen (3H-uridine, 5 pC/pl, approximately 0.25 fi/fly) and allowed to incorporate for approximately two hours.
R N A extraction: RNA was extracted from whole flies using a modification of the phenol: chloroform technique described by PALMITER (1974). Approximately 25-30 flies were homo- genized in 1 ml RNA extraction buffer in a Dual glass on glass homogenizer. To the homogenate was quickly added 1 ml phenol (redistilled) saturated with RNA extraction buffer. The mixture was vigorously shaken for a few seconds and then allowed to stand for approximately two minutes. To this mixture was then added 1 ml of chloroform and the resulting mixture was again shaken vigorously for a few seconds and allowed to stand for approximately two minutes. The aqueous phase was then separated by three minute centrifugation at 12K x g. The organic phase was removed by carefully slipping a Pasteur pipette down the side of the test tube. The flocculent interphase was left with the aqueous phase. The aqueous phase was then re-extracted with 2 ml of chloroform in the same manner as the first extraction. After centrifugation the lower chloroform layer was removed and a third chloroform extraction was performed. The aqueous was then removed to a clean tube and extracted twice with 2 ml of chloroform. The resulting aqueous was transferred to another clean test tube and extracted again with chloroform. The resulting aqueous was then made 2% with potassium acetate and adjusted to pH 5.5 with glacial acetic acid. The RNA was then precipitated by adding 2.5 volumes of ice cold 95% ethanol and storing a minimum of six hours at -20". The precipitated RNA was collected by centrifugation (top speed in the table top I.E.C. for five minutes) and washed twice with 95% ethanol. The pellet was then dried with filtered air and dissolved in electrophoresis buffer with 10% sucrose.
Polyacrylamide gel electrophoresis: The technique for gel electrophoresis was that of LOENING (1967). The large ribosomal RNA's were separated on 12 cm compound gels (12 cm in length and 0.6 cm in diameter). The upper 6 cm of the gels was 2.5% acrylamide and the lower 6 cm was either 7.5% or 10% acrylamide (the lower 6 cm higher percentage acrylamide is used to facilitate fractionation in the Gilson fractionator, see below). The preparation and electrophoresis of these gels are described in detail elsewhere (SHERMOEN and KIEFER 1975). After electrophoresis, gels were carefully removed from the tubes and scanned at 260nm on a Gilford recording spectrophotometer. Gels were then carefully returned to the tubes and sliced into 2 mm fractions using a Gilson automatic gel fractionator. Gel fractions were dissolved by overnight incubation in 0.2 ml of 50% H,O, at 60". Vials were then cooled and 10 ml of Biosolve scintillation cocktail (100 mg POPOP, 4 g PPO, 150 ml Biosolve to 1 liter of scintillation grade toluene) was added to each sample and radioactivity was determined using a Packard liquid scintillation counter.
D N A and rRNA preparaiion: The procedure for purification and denaturation of DNA

792 S . H . CLARK, L. D. STRAUSBAUGH AND B. I. KIEFER
from Drosophila has been described in detail by SHERMOEN and KIEFER (1975). Denatured DNA was loaded onto 13 mm Millipore nitrocellulose membrane filters according to the methods of GILLESPIE and SPIEGELMAN (1965).
Schneider’s line Drosophila cells were labeled for five days in medium containing 30-75 UC ?H-uridine/ml. The cells were lysed in extraction buffer (0.1 RI NaCI. 0.01 M NaCH,CO,, 1% SDS, 4 @g/ml polyvinyl sulfate, pH 5.1) and the lysate was extracted by the modification of PALMITER (1974), discussed under rate studies above. The precipitated RNA was collected by centrifugation and resuspended in NETS buffer (0.1 M NaC1, 0.01 M tris, 0.01 M EDTA, 0.2% SDS) and layered over a 10-30% linear sucrose gradient in NETS (SW 41). The gradients were centrifuged at 39K rpm (Spinco L-265 B) for 5 hours at 20” and then collected in 30 drop fractions. The 28s and 18s fractions were pooled and precipitated; the specific actiiity of the RNA was determined and the RNA stored frozen until used for hybridization.
Hybridization procedures: The hybridization and post-hybridization procedures were essen- tially those used by GILLESPIE and SPIEGELMAN (1965). Hybridization between the RNA and DNA was carried out overnight at 60” in 2 x SSC (1 x SSC buffer contains 0.15 M NaCl, 0.015 M sodium citrate, pH 7.0). The filters were washed by the batch method and treated with RNAse to remove any RNA “noise.” Radioactivity retained as hybrid was counted in a Packard Tri-carb scintillation counter and determination of rRNA gene number was made by the same calculations described in detail previously (SHERMOEN and KIEFER 1975).
RESULTS
In Drosophila melanogaster the bobbed phenotype is most easily characterized by shorter scutellar bristles. If the restoration of wild-type bristle pheno- type is a function of the presence of the YbbSuVar-S chromosome, rather than a unique situation due to the specific genotype measured by SHERMOEN and KIEFER (1975), then this chromosome should have the effect of restoring a wild-type phenotype to virtually all rDNA deficient flies. In order to test this hypothesis, males containing different rDNA deficient X chromosomes and different auto- somal backgrounds in combination with Ybbsuvar-s or Ybb- were constructed and their bristles measured. As can be seen in part A of Table 1, the presence of Ybbsuva*-5 restores a wild-type bristle phenotype with a variety of different rDNA- deficient X chromosomes. To eliminate the possibility that the restoration effect was due to differences in autosomal background, both Ybb- and YbbSuVar-5 were placed in homogeneous genetic backgrounds derived from a Samarkand isogenic wild-type stock. Since males used in all experiments were generated by mating sibling car %/car bb virgin females to males containing either Ybb- or YbbSuVa*+, the use of these males with homogeneous backgrounds as parental males elimi- nated the source of genetic variation in the experimental genotypes. Part B of Table 1 shows that the restoration of wild-type bristle lengths occurs with YbbSuVar-5 when compared to Ybb7 even when care has been taken to insure homo- geneity of the genetic elements other than the Y chromosomes. These data indi- cate that the Ybbsuvar-5 chromosome itself is responsible for the restoration of wild-type bristle phenotype.
SHERMOEN and KIEFER (1975) measured rates of rRNA synthesis in testes of males of the genotypes y w bbds/Ybb- and y w bbds/Ybbsuvar-5. Despite the fact that these males have equivalent rDNA, there was a dramatic increase in rRNA synthesis in testes with the Ybbsuvar-5 genotype. Because of the known genetic

INCREASED RATE OF rRNA SYNTHESIS
TABLE 1
Scutellar bristle lengths (microns)
793
A. Effect of different X b b chromosomes
X chromosome XSamt is0
car bb yw bb gt bb*l
Y chromosome ySam+ is0 yab- ybbsuvar-5
414 i: 2.0 412 f 1.8 412 f 1.6 - 324 -I 2.0 402 2 2.0 - 340 f 2.0 392 f 2.0 - 326 2 2.0 389 f 2.0
B. Effect of genetic background Bristle lengths of F, males when Yaa- and Ybasuvar-5 are contributed from flies with homogeneous Samarkand+ isogenic X and autosomal backgrounds:
cur bb/Yb” 320 f 2.0 car bb/ybbSuvar-s 396 f 2.0
F, genotype Bristle length
Values are mean f standard errors.
activity of the Y chromosome in testes, it was of interest to see if the modulation of RNA metabolism observed in testes also occurred in somatic tissue. For this reason. experiments were done using whole flies.
Most studies of synthetic rates are actually measurements of rates of accu- mulation, from which transcriptional rates are estimated. These rates are usually expressed as a specific activity, which is generally defined as the amount of radioactive precursor incorporated per unit macromolecule (e.g., dpmJpg RNA). When comparing such rate data between experimental systems, it is first neces- sary to determine the similarity or dissimilarity of the rate formula denominator (e.g., amount of RNA) in the systems being compared. Experiments to measure these parameters show that there is no significant difference in mg/wet weight between the genotypes measured in this study (Table 2). This conclusion insures a constant denominator in the RNA rate formula.
Since a synthetic rate analysis requires incorporation of radioactive precursor, it is important to choose incubation time points which are in the linear portion of the incorporation curve and to know that differences between genotypes are not due to differences in the pools for the radioactive precursor. Studies of
TABLE 2
R N A quantification
Genotype No. determinations pg RNA/fly p g RNA/mg wet weight
Sam+ is0 16 2.35 i: 0.13 3.16 f 0.10 car bb/Ybb- 14 2.16 f 0.08 3.15 f 0.10 cur bb/ybbSuVar-5 7 2.28 f 0.20 3.12 f 0.15
~ ___
Values are the mean f the standard error of the mean. RNA was isolated using the acid extraction procedure of SCHMIDT and THANNHAUSER (1945)
and isolated nucleic acid was quantified by UV absorption (LEVY, SIMPSON and SOBER 1972).

794 S. H. CLARK, L. D. STRAUSBAUGH A N D B. I. KIEFER
3H-ur;dine incorporation into total RNA show that the incorporation increases linearly for at least two hours for all genotypes (Figure 1 ) . In addition, results from measurements of ribonucleoside pools using density labeling demonstrate that any differences of incorporation between car bbJYbbSuVar-5 and car bb/Ybh males at two hours cannot be due to differences in the precursor pools (STRAUS- BAUGH. CLARK and GRAINGER, manuscript in preparation). Furthermore, the biological data argue strongly against the rate increase being an artifact of pre- cursor pools. If the observed differences in rRNA specific activity between the two genotypes were due to differences in precursor pools rather than real dif- ferences in rates of accumulation. then both genotypes should display bobbed bristle lengths and development times. This is clearly not the case. Pool con- siderations and RNA incorporation curves indicate that two hours or less is a suitable time for rate analysis in this system.
Adult males of the genotypes car bb/YbbSuVar-5, car bbJYbb-, and Sam+iso were injected with iH-uridine and allowed to incorporate label for two hours. The RNA was then extracted and run on polyacrylamide gels. The results of a typical gel scan and fractionation are shown in Figure 2. Data from graphs such as these are used to calculate the rates of rRNA synthesis for each genotype and the results are shown in Table 3. As can be seen from this table, car bb/Ybbsuvar-s has an approximately two-fold increase in the rate of rRNA synthesis as com- pared to car bb/Ybb-. Although very deficient in rDNA, the Ybbsuvar-5 genotype achieves a level of rRNA synthesis which is equivalent to wild type.
~ 4 . 0 - --I LL
I
\ 0
3.0- != x
a
4
I 20 40 S'O 80 100 120 140
T I M E (minutes)
FIGURE 1 .-Time course of 3H-uridine incorporation into nucleic acid. Flies were injected with 0.25 pl of 3H-uridine (1 aC/p1) and allowed to incorporate label. At appropriate intervals groups of flies (10 flies/group) were homogenized and TCA precipitable radioactivity determined.
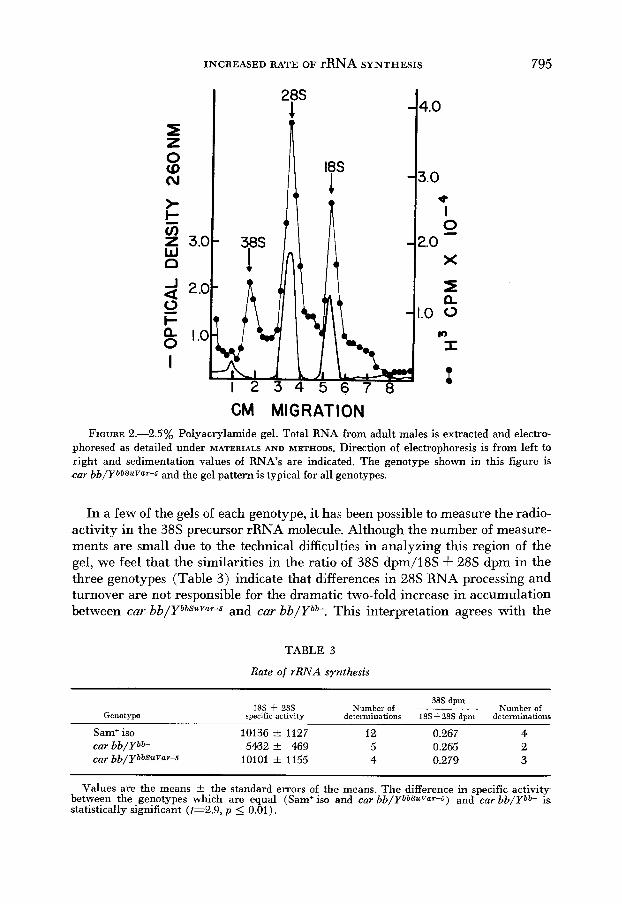
INCREASED RATE OF rRNA SYNTHESIS 795
28s
t 4.0
3.0 d I 0
x I e
.o 0
I
- LO
In
1 CM MIGRATION
FIGURE 2.-2.5% Polyacrylamide gel. Total RNA from adult males is extracted and electro- phoresed as detailed under MATERIALS AND METHODS. Direction of electrophoresis is from left to right and sedimentation values of RNA’s are indicated. The genotype shown in this figure is car bb/YbbSuVaT-s and the gel pattern is typical for all genotypes.
In a few of the gels of each genotype, it has been possible to measure the radio- activity in the 38s precursor rRNA molecule. Although the number of measure- ments are small due to the technical difficulties in analyzing this region of the gel, we feel that the similarities in the ratio of 38s dpm/l8S -I- 28s dpm in the three genotypes (Table 3 ) indicate that differences in 28s RNA processing and turnover are not responsible for the dramatic two-fold increase in accumulation between car bb/YbbSuVar-5 and car bbJYb”. This interpretation agrees with the
TABLE 3
Rate of rRNA synthesis
38s dpm ‘8s. + 28s Number of Number of
Genotype specific activity determinations 18Sf28S dpm determinations
Sam+ is0 10136 rtl 1127 12 0.267 4 car b b / Y b b - 5432 f 469 5 0.265 2 car bb /YbbSuVas- s 10101 f 1155 4 0.279 3
Values are the means f the standard errors of the means. The difference in specific activity between the genotypes which are equal (Sam+ is0 and car bb /YbbSuVar-5 ) and car b b / Y b b - is statistically significant (t=2.9, p 5 0.01).

796 S. H. CLARK, L. D. STRAUSBAUGH A N D B. I. KIEFER
conclusions of MOHAN (1975) that there are no differences in processing of rRNA’s between bobbed and wild-type flies.
The observation that car bb/YbbSuVa7-5 has elevated rRNA synthesis relative to car bb/Ybb- demonstrates modulation of RNA synthesis only if cur bb/YbbSuva7-J has an equal or lesser number of genes €or rRNA than does car bb/Ybb-. Previous measurements of the Ybb- and YbbSuvar-5 chromosomes have shown them to be equivalent in rDNA content (MALVA et al. 1972; SHERMOEN and KIEFER 1975). The genotype cur bb/Ybb- has been measured and has only about 90 genes com- pared to wild type which has about 400 genes (SHERMOEN and KIEFER 1975). Since the genotype car bb/YbbSuva7-5 had not been measured for gene number, it was important to rule out the possibility that its increase in transcription was due to an increased amount of template. Saturation hybridization experiments were done to address this possibility, and the results are shown in Table 4. It is clear from these data that car bb/YbbSuVa7-5 males are in fact rDNA deficient to a degree that is very similar to that for car bb/Ybb- (each contains approximately 25 percent of the rDNA of wild type). The results of this determination eliminate the possibility that the basis for the two-fold increase in rRNA synthesis for car bb/YbbSuVar-5 relative to car bb/Ybb- is some kind of dramatic increase in gene number.
Since cur bb/YbbSuVar-5 has only 25% of the rDNA of Sam+ iso, but the same rate of ribosomal RNA accumulation, the car bb/YbbSuvar-5 flies must be better able to utilize their rDNA: that is, the rate per gene is considerably higher in car bb/YbbSuVa7-5. In addition, cur bbJYbb and car bb/YbbSuVa7-5 have about the same rDNA content, yet cur bb/YbbSuVar-5 has a much higher synthetic rate. These observations are proof for the conclusion that the YbbSuVa7-5 chromosome is able to increase the transcription of rRNA genes.
We have measured the effect of this chromosome on another classic bobbed phenotype, delayed development. Figure 3 shows the adult development times for the same genotypes for which synthetic rates were measured. The data pre- sented in Figure 3 demonstrate that the presence of the Ybbsuva7-5 chromosome significantly decreases the period of development in rDNA-deficient flies. These data are predicted by the assumption that the delayed development in bobbed flies is due to a decreased rate of accumulation of rRNA, and that the Ybbsuvar-5
TABLE 4
SH-rRNA-DNA snturation hybridization levels
Gtnqtvpo
rRNA hybridization x 100% Gene Number of determinations
DNA number done a t saturation
Sam+ is0 0.356 f 0.024 407 8 0.100 c 0.004 114 6 0.078 f 0.012* 89 6
Values are means f the standard errors of the means. * Data from SHERMOEN and KIEFER (1975).

INCREASED RATE OF rRNA SYNTHESIS 79 7
DEVELOPMENTAL TIME STUDY
(Compilation of Data From Four Experiments)
; carbb / Y bb- L bL 0 $ 4 -
2-
I 2 3 4 5 6 7 8 9 1 0 1 1 1 2 1 3 1 4 1 5 DAYS
FIGURE S.-Adult development time. Histograms of male eclosion times of the three geno- types are shown. All cultures were started simultaneously to assure synchronous development.
chromosome restores the wild-type rate of accumulation by increasing the rate of transcription of rRNA genes.
DISCUSSION
The short-bristle phenotype and delayed development of rDNA-deficient (bb) mutants have been explained in terms of inadequate protein synthesis due to a reduced number of ribosomes. While this is a reasonable explanation. neither aspect of it has been experimentally demonstrated: that is, that rDNA-deficient cells have reduced numbers of ribosomes, and that reduced numbers of ribosomes result in quantitative and/or qualitative restrictions of protein synthesis. The findings in this study that increase in bristle length and decrease in development tima occur coincidentally with an increase in rates of rRNA synthesis are con- sistent with this explanation, but still require the yet-to-be-validated assumption that increases in rRNA synthesis result in an increase in the number of func- tional ribosomes. In any event, in agreement with both WEINMANN (1972) and SHERMOEN (1974), it is clear that the bristle phenotype is related to the rate of accumulation of rRNA which, in turn, has at least two components: the rate of transcription per gene and the number of genes being transcribed. I t would appear that both of these components are subject to regulation. KRIDER and

798 S . H . CLARK, L. D. STRAUSBAUGH AND B. I. KIEFER
PLAUT (1972), MEYER and HENNIG (1974) and MCKNIGHT and MILLER (1976) have presented evidence that suggests that not all rRNA genes present are neces- sarily transcriptionally active at any given time, and the evidence presented in this and the following paper and elsewhere (SHERMOEN and KIEFER 1975) is most easily interpreted as a change in the rate of transcription. If neither of these components were variable, then the bristle phenotype could be predicted from the rDNA content. That this is not the case has been demonstrated by SHERMOEN and KIEFER (1975) and can be deduced from the data of TARTOF (1973).
Whenever specific activity measurements are used to estimate synthetic rates, it is necessary to consider the rate of degradation of the molecules being studied. The present study utilized relatively short incubation periods to circumvent this problem. Over longer periods it is obvious that RNA degradation plays an impor- tant role in controlling the levels of RNA molecules in cells. Since bobbed mutants in general (MOHAN and RITOSSA 1970) and car bb/Ybb- and car bb/ YbbSuVnr-5 adults specifically (Table 2) have the same amount of RNA despite apparent lower synthetic rates in car bb/Yb” flies (Table 3), it is clear that the flies control the level of RNA by regulating rates of RNA degradation. This type of control implies the existence of a feedback mechanism that monitors RNA content within the cells and is capable of maintaining a constant level of ribo- somes. This control of RNA degradation, combined with delayed development, would allow most tissues of car bb/Ybb- flies to achieve a wild-type level of RNA despite a reduced rate of rRNA synthesis. Only those tissues that cannot com- pensate in this manner would have the bobbed phenotype (i.e., bristle cells and spermatids). This explanation is consistent with the data presented here and those observed by others.
The genetic basis for the effects of YbbSuYar-5 remains obscure at this time. Although YbbSuVar-5 and Ybb- are rDNA deficient to the same extent, it is con- ceivable that the deletions in these two Y chromosomes are not identical with respect to chromosomal location or organization within the rDNA region. While the two Y chromosomes may be quantitatively equivalent in rDNA, there may be important qualitative differences in the rDNA regions present. For example, BAKER (1971) has suggested the existence of an adjacent regulatory region for rDNA. and this region could be different between Yb’+ and YbbSuVar-5. Of course, it is also possible that the number of functional genes are different in the two Y chromosomes; since these bobbed Y chromosomes contribute such a small num- ber of genes to the total genotype, it is unlikely that a relative difference in functional activities in these genes could account for the observed difference in rRNA accumulation. An alternate possibility is that the genetic locus responsible for the modulatory effect of YbbSuVar-5 is not located in o r near the rDNA region, but is elsewhere on the Y chromosome. Finally, it may be that the genetic basis for the observed effect is not due to a specific gene, but represents a complex chromosomal interaction (see, for example, LIFSCHYTZ and LINDSLEY 1972).
While the most simple interpretation of the Ybbsuvar-5 mediated increase is increased transcription of a constant number of genes, there are complex situa-

INCREASED RATE OF rRNA SYNTHESIS 799
tions that involve changes in gene number which might also change the results. For example, it is possible that particular tissues in the adult are very active transcriptionally and account for most of the accumulation observed. These tissues could conceivably have undergone an increase in gene number, which might not be detected in whole-fly rDNA measurements. In fact, it is possible that a large percentage of the incorporation in whole flies is accounted for by the accumulation in testes, which has been previously reported ( SHERMOEN and KIEFER 1975). However. the increase in bristle lengths and the decrease in devel- opment times indicate a widespread occurrence of the effect.
In summary, the YbbSuVar-5 chromosome causes an increased rate of transcrip- tion of the DNA coding for 28s and 18s rRNA. The data of SHERMOEN and KIEFER (1975) and those presented here clearly indicate a level of genetic con- trol of rDNA other than the number of genes, that of the utilization of available genes. In this work we have measured the effect of YbbSuVar-5 only on ribosomal RNA synthesis. An important question that bears on the modes of expression and the molecular mechanisms of the “regulator” effect of this chromosome is whether the effect is specific for rRNA or includes other species of RNA as well. This question is addressed in the following paper. Another potentially important aspect of the design of the experiments investigating the effects of YbbSuVar-5 to date has been the use of a bobbed X chromosome. An important consideration in this respect is whether the Ybbsuvar-5 chromosome will have any effect on RNA metabolism in a genotype which contains a nonbobbed X . Experiments designed to investigate this aspect of the YbbSuVar-5 effect are currently being done in our laboratory and will be reported in subsequent papers.
We thank JEAN BERTMAN for technical assistance. This research was supported by research grant GM-19130 from the Public Health Service.
LITERATURE CITED
BAKER, W. K., 1971
GILLESPIE, D. and S. SPIEGELMAN, 1965
KIEFER, B. I. and A. W. SHERMOEN, 1973
KRIDER, H. and W. PLAUT, 1972
Evidence for position effect suppression of the ribosomal RNA cistrons in
A quantitative assay for DNA-RNA hybrids with
Defective spermiogenesis in fertile rDNA-deficient Drosophila melanogaster. Genetics 74: s138.
Studies on nucleolar RNA synthesis in Drosophila melanogas- ter. 11. The influence of condition on the rate of synthesis and secondary constriction forma- tion. J. Cell Sci. 11: 675-687.
Fractionation of chromatin components. Bio-
The role of X chromosome inactivation during spermio-
Genetic variation of Drosophila melanogaster. Carnegie
The fractionation of high-molecular-weight ribonucleic acid by poly-
Drosophila melanogaster. Proc. Nat. Acad. Sci. US. 62 : 2472-2476.
DNA immobilized on a membrane. J. Mol. Biol. 12: 829-842.
LEVY, S., R. T. SIMPSON and H. A. SOBER, 1972
LIFSCHYTZ, E. and D. L. LINDSLEY, 1972
LINDSLEY, D. L. and E. H. GRELL, 1968
LOENING, U. E., 1967
chem. 11: 1547-1554.
genesis. Proc. Nat. Acad. Sci. U.S. 1: 182-186.
Inst. Washington Publ. No. 627.
acrylamide gel electrophoresis. Biochem. J. 102 : 251-257.

800 S. H. CLARK, L. D. STRAUSBAUGH AND B. I. KIEFER
MALVA, C . , F. GRAZIANI, E. BONICELLI, L. POLITO and F. RITOSSA, 1972 Check of gene number during the process of rDNA magnification. Nature New Biol. 239: 135-137.
MCKNIGHT, S. and 0. MILLER, 1976 Ultrastructural patterns of RNA synthesis during early embryogenesis of Drosophila melanogaster. Cell 8 : 305-319.
MEYER, G. F. and W. HENNIG, 1974 The nucleolus in primary spermatocytes of Drosophila hydei. Chromosoma (Berlin) 46: 121-144.
MOHAN, J. 1975 The synthesis of 5 s RNA and its relationship to 18s and 28s ribosomal RNA in the bobbed mutants of Drosophila mehogaster . Genetics 81: 723-738.
MOHAN, J. and F. RITOSSA, 1970 Regulation of ribosomal RNA synthesis and its bearing on the bobbed phenotype in Drosophila melanogaster. Devel. Biol. 22 : 495-512.
PALMITER, R. D., 1974 Magnesium precipitation of ribonucleoprotein complexes expedient techniques for the isolation of undegraded polysomes and messenger ribonucleic acid. Bio- chem. 13: 3606-3615.
RITOSSA, F. M., 1968 Unstable redundancy of genes for ribosomal RNA. Proc. Nat. Acad. Sci.
R~TOSSA, F. M. and S. SPIEGELMAN, 1965 Localization of DNA complementary to ribosomal RNA in the nucleolus organizer region of Drosophila melanogaster. Proc. Nat. Acad. Sci. U.S. 53: 737-745.
A method for the determination of deoxyribo- nucleic acid, ribonucleic acid, and phosphoproteins in animal tissues. J. Biol. Chem. 161 :
Ribosomal RNA synthesis and spermatogenesis in rDNA-deficient
Regulation in rDNA-deficient Drosophila melanogas-
Regulation of ribosomal RNA gene number multiplicity in Drosophila
Regulation of ribosomal RNA and 5s RNA synthesis in Drosophila mel-
Corresponding editor: A. CHOVNICK
U.S. 60: 509-516.
SCHMIDT, G. and S. J. THANNHAUSER, 1945
83-89. SHERMOEN, A. W., 1974
SHERMOEN, A. W. and B. I. KIEFER, 1975
TARTOF, K. D., 1973
WEINMANN, R , 1972
Drosophila melanogaster. Ph.D. Thesis, Wesleyan University, Middletown, Connecticut.
ter. Cell 4: 275-280.
me1,anogaster. Genetics 73: 57-64.
anoqaster. I. Bobbed mutants. Genetics 72 : 267-276.





























