Embed Size (px)
Citation preview

THE JOURNAL OF BIOLOGICAL CHEMISTRY 0 1992 by The American Society for Biochemistry and Molecular Biology, Inc.
Vol. 267, No. 29, Issue of October 15, pp. 21172-21178,1992 Printed in U.S.A.
Biochemical Properties of the 75-kDa Tumor Necrosis Factor Receptor CHARACTERIZATION OF LIGAND BINDING, INTERNALIZATION, AND RECEPTOR PHOSPHORYLATION*
(Received for publication, March 23, 1992)
Diane PennicaS, Van T. Lam, Nancy K. Mize, Richard F. Weber, Martyn Lewis, Brian M. Fendlyf, Michael T. Liparill, and David V. Goeddel From the DeDartments of Molecular BioloPv. SMedicinal and Analytical Chemistry and IICardiouascular Research, Genentech, Incorporated: South San’Francisco, Ca1ifo&zau94080
An expression plasmid encoding the human 75-kDa tumor necrosis factor (TNF) type 2 receptor (TNF-RP) was constructed and used to generate a stable human cell line (293/TNF-R2) overexpressing TNF-RP. Li- gand binding analysis revealed high affinity binding (& = 0.2 nM) with approximately 94,000 f 7,500 sites/cell for 1251-TNF-a and approximately &fold lower affinity for TNF-/3 (& = 1.1 nM) with 264,000 f 2,000 sites/cell. Cross-linking of ‘2SI-TNF-a and lZ6I- TNF-@ to 293/TNF-R2 cells yielded predominant com- plexes with apparent molecular weights of 2 11,000 for TNF-a and 206,000 and 244,000 for TNF-B, suggest- ing these complexes contain two or three TNF-RP mol- ecules. Immunoprecipitation of TNF-R2 from 32P-la- beled 293/TNF-R2 cells demonstrated that the recep- tor is phosphorylated. The majority (97%) of 32Pi incorporation was found in serine residues with a very low level of incorporation (3%) in threonine residues. TNF-a treatment of 293/TNF-R2 cells did not signifi- cantly affect the degree or pattern of phosphorylation. Cell surface-bound 12’I-TNF-a was slowly internalized by the 293/TNF-R2 cell line with a tlI2 = 25 min. Shedding of the extracellular domain of TNF-R2 was induced by 4B-phorbol12-myristate 13-acetate but not by TNF-a or TNF-@.
TNF-a’ and TNF-P are two structurally and functionally related cytokines produced mainly by macrophages and lymphoid cells, respectively (1). They have now been shown to mediate a wide spectrum of activities including cell prolif- eration, cytotoxicity, antiviral responses, and activation of many cellular genes, transcription factors, and kinases (1-4). Although the molecular mechanisms underlying TNF action have not yet been elucidated, many responses have been shown to be initiated by the binding of TNF-a or TNF-P to specific cell surface receptors. Receptors for both TNF-a and TNF-/3, with dissociation constants in the nanomolar range, have been reported on the majority of somatic cell types
* The costs of publication of this article were defrayed in part by the payment of page charges. This article must therefore be hereby marked “advertisement” in accordance with 18 U.S.C. Section 1734 solely to indicate this fact.
$ To whom correspondence should be addressed Dept. of Molecu- lar Biology, Genentech, Inc., 460 Point San Bruno Blvd., South San Francisco, CA 94080.
‘The abbreviations used are: TNF-a, tumor necrosis factor-a; TNF-@, tumor necrosis factor-@; TNF-R1, TNF receptor 1; TNF-R2, TNF receptor 2; PMA, 4@-phorbol 12-myristate 13-acetate; PBS, phosphate-buffered saline; SDS, sodium dodecyl sulfate; PAGE, poly- acrylamide gel electrophoresis; PMSF, phenylmethylsulfonyl fluo- ride; EGS, ethylene glycolbis (succinimidylsuccinate); BSA, bovine serum albumin; PE, phycoerytherin.
analyzed (5-10). Crystallization studies have shown that TNF-a and TNF-P exist as closely packed trimers (11, 12), however, the oligomeric state of biologically active TNF that triggers a response through receptor binding is still somewhat unclear.
Two distinct types of TNF receptors have been identified, and their cDNAs cloned and expressed (13-22). A 55-kDa molecule designated TNF-R1 and a 75-kDa molecule desig- nated TNF-R2 bind both TNF-a and TNF-P although it has been reported that TNF-R2 binds TNF-a with higher affinity than TNF-/3 (16). The extracellular regions of TNF-R1 and TNF-R2 share approximately 30% amino acid identity and are characterized by 4 cysteine-rich repeat units that are presumed to form the ligand-binding domain. The cysteine- rich extracellular domains of the TNF receptors also show significant homology to the extracellular regions of the nerve growth factor receptor (23) and Fas (24), CDw40 (25), CD27 (26), and OX40 antigens (27), as well as open reading frames of the Shope fibroma virus (28), malignant rabbit fibroma, and myxoma viruses (29). Interestingly, the two TNF receptor molecules are unrelated in their cytoplasmic domains, and with one exception (a low level of homology between a 45- amino-acid portion of the cytoplasmic domains of TNF-R1 and the Fas antigen) are not homologous to other known receptors. The lack of homology between the two TNF recep- tor intracellular regions suggests that these receptors are involved in distinct signal transduction pathways.
Since TNFs induce a wide range of biological activities, it is likely that some of these responses are mediated by TNF- R1, others by TNF-R2. Recent studies using receptor agonist antibodies support this view of nonredundant signaling path- ways mediated by the two TNF receptors. These studies have shown for example that TNF-R1 initiates signals for cytotox- icity, induction of several genes, fibroblast proliferation, re- sistance to chlamydiae, and synthesis of prostaglandin E2 (30-32), whereas TNF-R2 initiates signals for thymocyte and cytotoxic T cell proliferation (32). Thus, both TNF-R1 and TNF-R2 appear to be active in initiating signal transduction but are functionally distinct.
In order to further characterize TNF-R2 and gain addi- tional insight into its role(s) in signal transduction, a human cell line was generated that overexpresses the full-length TNF-R2. We have examined the biochemical properties of TNF-R2 including the binding, cross-linking, internalization, and degradation of ligand, TNF-R2 phosphorylation, and the proteolytic generation of soluble TNF-R2 extracellular do- main.
MATERIALS AND METHODS
Reagents-Recombinant human TNF-a (4.75 X lo7 units/mg) and TNF-@ (2.1 X 10’ units/mg) were provided by the Genentech, Inc.
21172

75-kDa TNF Receptor Characterization 21173
manufacturing group. "'I-TNF-a (specific activity, 25.8 pCi/pg) and '"I-TNF-D (specific activity, 37.2 pCi/pg) were prepared using the lactoperoxidase method (33) or purchased from Amersham Corp. (specific activity, 46.0 pCi/pg). Rabbit anti-human TNF-R2 antibod- ies were generated against the soluble extracellular domain of TNF- R2. The titer of this antisera was quantitated by a direct antigen- coated enzyme-linked immunosorbant assay. The dilution of anti- TNF-R2 giving 50% binding to the purified soluble receptor was 1:68,000. Anti-TNF-R1 monoclonal antibody 4E4 (IgG1 isotype) and anti-TNF-R2 monoclonal antibodies 9H9 and 2E8 (IgG2a and IgG2b isotypes, respectively) were produced as described previously (34). Phycoerytherin (PE)-conjugated goat anti-mouse immunoglobulin G (IgG) F(ab'), was purchased from Caltag, South San Francisco. 48- Phorbol 12-myristate 13-acetate (PMA) and phenylmethylsulfonyl fluoride (PMSF) were purchased from Sigma. Ethylene glycolbis (succinimidylsuccinate) (EGS) was purchased from Pierce Chemical Co. The ECL Western blotting detection reagent was from Amersham Corp. Protein A-Sepharose was purchased from Pharmacia LKB Biotechnology Inc. [%3]Cysteine, [35S]methionine, and [32P]ortho- phosphate were purchased from Amersham Corp. Okadaic acid was purchased from LC Services Corp.
Plasmid Construction and Transfection-The human histiocytic lymphoma cell line U-937 was used as a source of mRNA for construc- tion of a cDNA library. Double-stranded cDNA was synthesized using an Invitrogen cDNA kit and inserted into the mammalian expression vector pCDM8 (35) to generate a library of approximately 2 X lo6 recombinants. A TNF-R2 clone was isolated from the cDNA library by screening pools of clones using the polymerase chain reaction and two 22-base long synthetic oligonucleotides corresponding to amino acid numbers 22-28 and 167-173 of the published TNF-R2 cDNA sequence (16). Full-length clones were identified by restriction en- donuclease mapping and sequence analysis. All screening, recloning, and nucleotide sequencing was by standard protocols (36). The full- length coding region for human TNF-R2, contained on a 2.4-kilobase XhoI fragment, was inserted into the pcisNEBO expression vector' between the cytomegalovirus immediate-early promoter and the SV40 termination and polyadenylation signal. This vector also contains the Epstein-Barr virus nuclear antigen 1 gene and oriP region from Epstein-Barr virus and the neomycin resistance gene. The resulting plasmid called p.NEBO.hR2 was transfected into the human embry- onic kidney cell line 293 (37) using the LIPOFECTINTM Reagent (Bethesda Research Laboratories) according to the manufacturer's instructions (5 pg of DNA/lO-cm dish) and analyzed for TNF-R2 expression 36 h after transfection using fluorescence-activated cell sorting.
Isolation of 293 Cells Overexpressing Human TNF-R2-Trans- fected cells were pooled and incubated for 1 h at 4 "C with approxi- mately 10 pg of TNF-R2 monoclonal antibody 9H9/106 cells, washed, then incubated for 45 min at 4 "C with 5 pg of PE-conjugated goat anti-mouse IgG/106 cells. Cells were washed and analyzed on a Coulter Elite flow cytometer equipped with a single 15-milliwatt argon laser tuned to 488 nm. Forward and 90" angle light scatter, propidium iodide exclusion for viable cell gating, and integrated log PE fluores- cence signals (with a band pass filter (575 + 25 nm)) were collected and analyzed. TNF-R2-positive cells were collected by sorting the brightest 10% of viable cells in the sample, washed with medium, and plated on a 35-mm dish in medium containing 600 pg/ml of G418 (GIBCO). After expansion, this sorting procedure was repeated two additional times. The final cell population named 293/TNF-R2 was maintained in low glucose Dulbecco's modified Eagle's media con- taining G418.
Immunoprecipitation of 35S-Labeled TNF-R2 and sTNF-R2-1.2- 1.5 X lo6 293/TNF-R2 cells were seeded in 3 ml of media in a 60-mm plate. Twenty-four h later, the cells were washed twice with PBS then incubated with 1 ml of low glucose Dulbecco's modified Eagle's media without cysteine or methionine for 30 min. This media was replaced with 2 ml of fresh Dulbecco's modified Eagle's media (with- out cysteine or methionine) containing [35S]cysteine and [35S]methi- onine (100 pCi/ml of each). The cells were incubated for an additional 16-18 h at 37 "C. The conditioned media was harvested and centri- fuged at 800 X g at 4 "C for 4 min to remove cells and debris before immunoprecipitation. The cells were washed once with cold PBS, and 350 pl of scrape buffer (25 mM Tris, pH 7.5,250 mM NaC1) containing 0.2 mM PMSF and 20 pg/ml aprotinin was added to each plate and the cell mixture was transferred to an Eppendorf tube containing 350
' G. Cachianes, C. Ho, R. F. Weber, S. R. Williams, D. V. Goeddel, and D. W. Leung, submitted for publication.
pl of 2 X lysis buffer (2% Triton X-100, 1% deoxycholate, 10 mM EDTA, 250 mM NaCl, and 25 mM Tris, pH 7.5). After vortexing, the cells were lysed for 10 min at 4 "C. Nuclear and cell debris were removed by centrifugation at 10,000 X g for 10 min at 4 "C. SDS was added to the supernatant to a final concentration of 0.1%. One ml of conditioned medium or 350 pl of cell lysate was immunoprecipitated by adding 2 pl of anti-TNF-R2 polyclonal antiserum and incubating for 1 h at 4 "C. Fifty p1 of Protein A-Sepharose beads were added and incubation continued for 45 min at 4 "C. The reactions were centri- fuged for 3 min at 15,800 X g at 4 "C and the supernatants discarded. The Protein A-Sepharose pellet was washed four times with 500 pl of 1 X lysis buffer. The final pellet was resuspended in 25 pl of 2 X SDS-PAGE sample buffer, heated at 100 "C for 10 min, and then centrifuged for 2 min at 15,800 X g. The supernatant was electropho- resed on a 4-20% Tris/glycine polyacrylamide gel. The gel was fixed, incubated in Amplify (Amersham Corp.), dried, and exposed to film.
Binding Analysis-293/TNF-R2 cells were harvested with PBS containing 1 mM EDTA (PBS/EDTA) and resuspended in PBS, 0.1% BSA, 0.02% sodium azide (PBSA buffer). Saturation isotherm exper- iments were performed in duplicate by incubating 2 x lo5 cells in 0.2 ml of PBSA with increasing concentrations of "'I-TNF-a or "'I- TNF-D (6.9 PM to 8.3 nM of each labeled ligand). Nonspecific binding was determined by the addition of 100-fold molar excess of unlabeled TNF-a or TNF-8 to an identical set of reactions. Reaction mixes were incubated at 4 "C for 1 h. Reactions were stopped by centrifu- gation at 12,400 X g for 1 min at 4 "C. Unbound '2SI-labeled ligand in the supernatant was removed by aspiration. The pellets were washed once with 0.2 ml of cold PBSA, and the amount of '2sI-TNF-a or lz5I- TNF-P bound to the cells was determined by counting the cell pellets in an Isodata gamma counter. The data were plotted with the Scat- chard equation using nonlinear least-squared regression.
Cross-linking of TNF-a and TNF-D to TNF-R2-1 X lo6 293/TNF- R2 cells were harvested and resuspended in 0.2 ml of PBSA. '"1- TNF-a (5.2 nM, 15.9 pCi/pg) or lZI-TNF-@ (0.4 nM, 32.8 pCi/pg) was added and the cells incubated at room temperature for 1 h in the presence or absence of 0.1 p~ unlabeled TNF-a or TNF-8. The cross- linking reagent, EGS (8.1 mM), was added to each reaction and incubation continued for an additional 30 min at room temperature. The reactions were stopped by adding 20 pl of 1 M glycine (0.1 M final concentration). The pellets were washed three times by resus- pending in 0.2 ml of cold PBS and spinning for 1 min at 12,400 X g at 4 "C. To the final cell pellet, 20 pl of a solution containing 1% Triton X-100, 1 p~ PMSF, was added and the cells lysed by incubat- ing at 4 "C for 1 h. After lysis, the cell debris was removed by centrifuging at 15,800 X g for 5 min at 4 "C. The supernatants were mixed with 2X SDS-PAGE sample buffer, boiled for 10 min, and analyzed on a 4-12% Tris/glycine polyacrylamide gel (Novex).
3ZP-Labeling and Immunoprecipitation of TNF-R2-293/TNF-R2 cells were seeded in 60-mm dishes a t 1.5 X lo6 cells/plate. After 24 h, the cells were washed once with PBS and then incubated in 2 ml of phosphate-free media for 30 min at 37 "C. This media was replaced with 1 ml of phosphate-free media containing 0.5 mCi of ["PI orthophosphate and the cells incubated for 4 h at 37 "C. The labeled cells were harvested with 0.4 ml of PBS/EDTA and the plates rinsed with 0.8 ml of PBSA. The cell mixture was transferred to an Eppen- dorf tube and the cells washed once with 1 ml of cold PBSA by centrifuging at 800 X g for 5 min at 4 "C. The cell pellet was lysed by adding 200 p1 of lysis buffer (0.2 M NaC1, 1 mM EDTA, 50 mM Tris, pH 8.0, 1% Nonidet P-40) containing the phosphatase inhibitors phenyl phosphate (30 mM), sodium orthovanadate (2 mM), ZnClZ (1 mM), and okadaic acid (1 p ~ ) , and the protease inhibitors, leupeptin (0.5 mg/ml), EDTA (1 mM), pepstatin (0.7 mg/ml), and PMSF (0.2 mM), and incubating for 1 h at 4 "C. After lysis, the cell debris was removed by centrifuging for 5 min at 15,800 X g at 4 "C. Immunopre- cipitation of the supernatant using 2 pl of rabbit anti-human sTNF- R2 antiserum was performed as described previously (38).
Phosphoamino Acid Analysis of TNF-R2-The immunoprecipi- tated 32P-labeled TNF-R2 bands from control or TNF-a-treated 293/ TNF-R2 cells were resolved by SDS-PAGE and identified by exposing the gel to film. The TNF-R2 bands were excised and analyzed for phosphoamino acid content by two-dimensional electrophoresis as described (39). Following this, the thin layer plates were exposed to Kodak X-Omat AR film, and spots corresponding to phosphoamino acids were scraped and quantified for 32P.
Internalization of TNF-(u-293/TNF-R2 cells were harvested and washed once with PBS, 0.1% BSA. 1 x lo7 cells were incubated in 5.0 ml of PBS, 0.1% BSA with either 0.25 nM '251-TNF-a (46.2 pCi/ pg, Amersham Corp.) for 1 h at 4 "C or preincubated with anti-TNF-
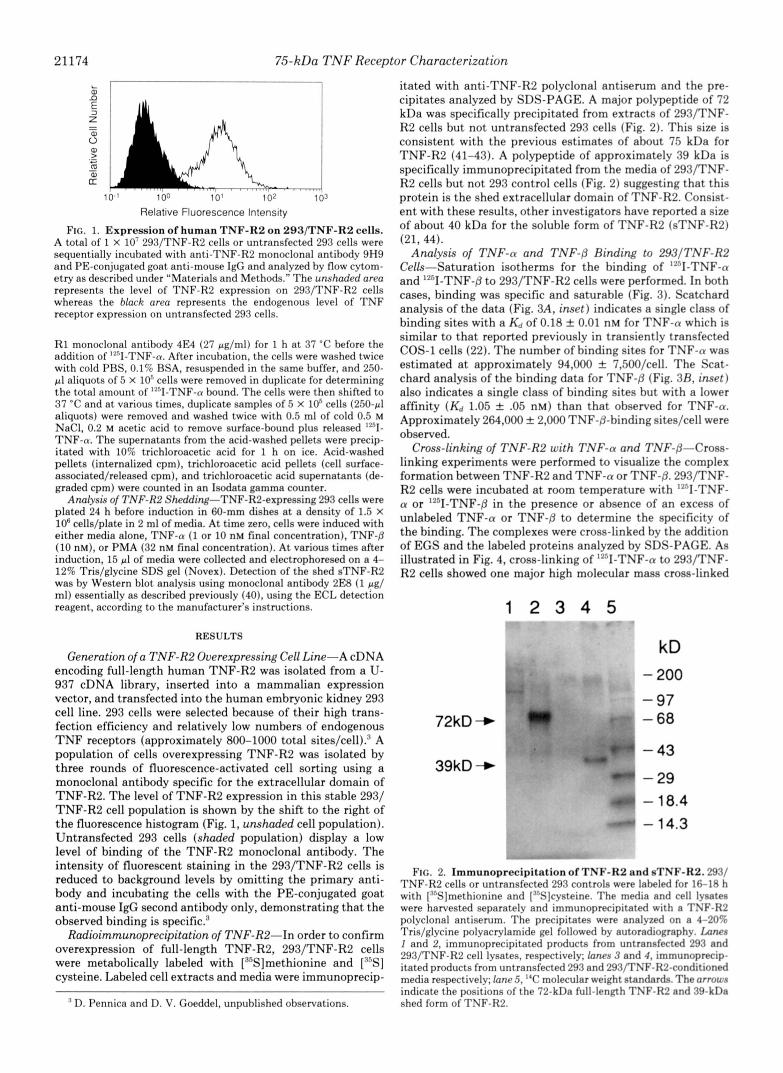
21174 75-kDa TNF Receptor Characterization
L % I I
1 0 ' 10 10' 102 103
Relative Fluorescence Intensity FIG. 1. Expression of human TNF-R2 on 293/TNF-R2 cells.
A total of 1 X 10' 293/TNF-R2 cells or untransfected 293 cells were sequentially incubated with anti-TNF-R2 monoclonal antibody 9H9 and PE-conjugated goat anti-mouse IgG and analyzed by flow cytom- etry as described under "Materials and Methods." The unshaded area represents the level of TNF-R2 expression on 293/TNF-R2 cells whereas the black area represents the endogenous level of TNF receptor expression on untransfected 293 cells.
R1 monoclonal antibody 4E4 (27 pg/ml) for 1 h at 37 "C before the addition of ""I-TNF-n. After incubation, the cells were washed twice with cold PBS, 0.1% BSA, resuspended in the same buffer, and 250- pl aliquots of 5 x IO6 cells were removed in duplicate for determining the total amount of '2sI-TNF-a bound. The cells were then shifted to 37 "C and at various times, duplicate samples of 5 X lo5 cells (250-pl aliquots) were removed and washed twice with 0.5 ml of cold 0.5 M NaCI, 0.2 M acetic acid to remove surface-bound plus released '"I- TNF-a. The supernatants from the acid-washed pellets were precip- itated with 10% trichloroacetic acid for 1 h on ice. Acid-washed pellets (internalized cpm), trichloroacetic acid pellets (cell surface- associated/released cpm), and trichloroacetic acid supernatants (de- graded cpm) were counted in an Isodata gamma counter.
Analysis of TNF-R2 Sheddin+"NF-R2-expressing 293 cells were plated 24 h before induction in 60-mm dishes at a density of 1.5 X 10' cells/plate in 2 ml of media. A t time zero, cells were induced with either media alone, TNF-n (1 or 10 nM final concentration), TNF-fi (10 nM), or PMA (32 nM final concentration). At various times after induction, 15 pl of media were collected and electrophoresed on a 4- 12% Tris/glycine SDS gel (Novex). Detection of the shed sTNF-R2 was by Western blot analysis using monoclonal antibody 2E8 (1 pg/ ml) essentially as described previously (40), using the ECL detection reagent, according to the manufacturer's instructions.
RESULTS
Generation of a TNF-R2 Overexpressing Cell Line-A cDNA encoding full-length human TNF-R2 was isolated from a U- 937 cDNA library, inserted into a mammalian expression vector, and transfected into the human embryonic kidney 293 cell line. 293 cells were selected because of their high trans- fection efficiency and relatively low numbers of endogenous T N F receptors (approximately 800-1000 total sites/cell).3 A population of cells overexpressing TNF-R2 was isolated by three rounds of fluorescence-activated cell sorting using a monoclonal antibody specific for the extracellular domain of TNF-R2. The level of TNF-RP expression in this stable 293/ TNF-R2 cell population is shown by the shift to the right of the fluorescence histogram (Fig. 1, unshaded cell population). Untransfected 293 cells (shaded population) display a low level of binding of the TNF-R2 monoclonal antibody. The intensity of fluorescent staining in the 293/TNF-R2 cells is reduced to background levels by omitting the primary anti- body and incubating the cells with the PE-conjugated goat anti-mouse IgG second antibody only, demonstrating that the observed binding is specific.:'
Radioimmunoprecipitation of TNF-R2--In order to confirm overexpression of full-length TNF-R2, 293/TNF-R2 cells were metabolically labeled with ["S]methionine and ["S] cysteine. Labeled cell extracts and media were immunoprecip-
D. Pennica and D. V. Goeddel, unpublished observations.
itated with anti-TNF-R2 polyclonal antiserum and the pre- cipitates analyzed by SDS-PAGE. A major polypeptide of 72 kDa was specifically precipitated from extracts of 293/TNF- R2 cells but not untransfected 293 cells (Fig. 2). This size is consistent with the previous estimates of about 75 kDa for TNF-R2 (41-43). A polypeptide of approximately 39 kDa is specifically immunoprecipitated from the media of 293/TNF- R2 cells but not 293 control cells (Fig. 2) suggesting that this protein is the shed extracellular domain of TNF-R2. Consist- ent with these results, other investigators have reported a size of about 40 kDa for the soluble form of TNF-R2 (sTNF-R2) (21, 44).
Analysis of TNF-a and TNF-@ Binding to 293/TNF-R2 Cells-Saturation isotherms for the binding of ""I-TNF-a and '*'I-TNF-@ to 293/TNF-R2 cells were performed. In both cases, binding was specific and saturable (Fig. 3). Scatchard analysis of the data (Fig. 3A, inset) indicates a single class of binding sites with a Kd of 0.18 & 0.01 nM for TNF-a which is similar to that reported previously in transiently transfected COS-1 cells (22). The number of binding sites for TNF-a was estimated a t approximately 94,000 f 7,50O/celI. The Scat- chard analysis of the binding data for TNF-@ (Fig. 3R, inset) also indicates a single class of binding sites but with a lower affinity ( K d 1.05 & .05 nM) than that observed for TNF-a. Approximately 264,000 f 2,000 TNF-P-binding siteslcell were observed.
Cross-linking of TNF-R2 with TNF-a and TNF-@-Cross- linking experiments were performed to visualize the complex formation between TNF-R2 and TNF-a or [email protected]/TNF- R2 cells were incubated a t room temperature with '*'I-TNF- a or ""I-TNF-@ in the presence or absence of an excess of unlabeled TNF-a or TNF-P to determine the specificity of the binding. The complexes were cross-linked by the addition of EGS and the labeled proteins analyzed by SDS-PAGE. As illustrated in Fig. 4, cross-linking of "'I-TNF-a to 293/TNF- R2 cells showed one major high molecular mass cross-linked
1 2 3 4 5 v - * - C"
kD - 200
- 97 72kD + - 68
- 43
- 29 - 18.4 - 14.3
39kD +
FIG. 2. Immunoprecipitation of TNF-R2 and sTNF-R2.293/ TNF-R2 cells or untransfected 293 controls were laheled for 16-18 h with ["'SS]methionine and [""Sjcysteine. The media and cell lysates were harvested separately and immunoprecipitated with a TNF-R2 polyclonal antiserum. The precipitates were analyzed on a 4-20rA Tris/glycine polyacrylamide gel followed by autoradiography. lams 1 and 2, immunoprecipitated products from untransfected 291 and 293/TNF-R2 cell lysates. respectively; lanes 3 and 4. immunoprecip- itated products from untransfected 293 and 293/TNF-H2-conditioned media respectively; lane .5, "C molecular weight standards. The arrous indicate the positions of the 72-kDa full-length TSF-R2 nnd RS-kDa shed form of TNF-R2.

7fj-kDa TNF Receptor Characterization 21175
0 2 4 6 8 10
1251-TNF-a (nM)
i 0
FIG. 3. Binding of 12nI-TNF-a and '*'I-TNF-B to 293/TNF- R2 cells. Saturation isotherms of the specific binding of ""I-TNF-n and ""I-TNF-p were performed by incuhating duplicate samples of 2 X loK 293/TNF-R2 cells with increasing concentrations of ""I-TNF- n or '"'I-TNF-P alone or in the presence of 100-fold molar excess of unlabeled TNF-n or TNF-8, and the specific binding at each concen- tration WRS determined. The inset presents the data transformed by Scatchard analysis. All binding experiments were performed two to three times with similar results. The results from a single represent- ative experiment are shown.
1 2 3 4
200 -
97.4 -
68 -
43 -
29 - 18.4 -
FIG. 4. Cross-linking of ""I-TNF-(r and '2nI-TNF-B to 293/ TNF-R2 cells. 293/TNF-R2 cells were incubated with ""I-TNF-n or ""I-TNF-Lj in the presence or ahsence of unlabeled TNF-n or TNF- 0 and treated with the cross-linking reagent EGS. Cell lysates were resolved on a 4-129; Tris/glycine polyacrylamide gel followed by autoradiography. Ixne I , ""I-TNF-n only; lane 2, ""I-TNF-n + 0.1 pM unlabeled TNF-n; lane 3 , '?"l-TNF-[j only; lane 4, ""I-TNF-8 + 0.1 p~ unlabeled TNF-B. The positions of "C molecular weight standards are indicated.
product of 211 kDa (lane I), possibly representing two TNF- R2 molecules complexed with one TNF-n trimer. The smaller complexes seen faintly at M , = 101,000 and 134,000 would be consistent with the cross-linking of one TNF-R2 molecule to one TNF-n monomer and trimer, respectively. Cross-linking of 12sII-TNF-P to TNF-R2, on the other hand, showed a slightly different pattern of two major high molecular mass complexes with apparent molecular masses of approximately 205 and 244 kDa (Fig. 4, lane 3 ) . The 205-kDa band could represent two TNF-R2 molecules complexed with one TNF-p trimer, while the 244-kDa band might contain an additional molecule of TNF-R2 or TNF-@. Two less prominent, lower molecular mass complexes of about 98 and 123 kDa were also observed which may represent one TNF-R2 molecule cross-linked with one TNF-P monomer and trimer, respectively. No labeled complexes were observed when cross-linking was performed in the presence of excess unlabeled TNF-n or TNF-/5' (Fig. 4, lanes 2 and 4 ) indicating that the observed binding and complex formation is specific. Immunoprecipitation of the cross-linked products with a TNF-R2-specific polyclonal anti- serum followed by SDS-PAGE analysis yielded similar re- sults.:' Cross-linking analysis on untransfected 293 cells showed no observable complexes when compared under iden- tical conditions with 293/TNF-R2 cells.:'
Phosphorylation of TNF-R2 and Phosphoamino Acid Anal-v- sis-TNF-R2 phosphorylation was examined by metabolically labeling 293/TNF-R2 cells with ["P]orthophosphate in the presence or absence of TNF-n followed by immunoprecipita- tion with TNF-R2 polyclonal antiserum. SDS-PAGE analysis of the precipitated products indicated that TNF-R2 is a phosphoprotein both in the absence (Fig. 5A, lanm I-.?) or presence of TNF-n (Fig. 5A, lanes 4-6) . No significant differ- ence in the amount of ,'"Pi incorporation was seen following TNF-n treatment.
Phosphoamino acid analysis of .'"P-labeled immunoprecip- itated TNF-R2 from unstimulated and TNF-n-treated cells revealed that 97% of the ,'"PI was present as phosphoserine while only about 3% was incorporated in threonine residues (Fig. 5 R ) . TNF-n treatment did not significantly change the amount of serine or threonine phosphorylation.
Internalization of TNF-n by TNF-R2"ln order to follow the fate of TNF-n after binding to TNF-RP, internalization and degradation of ""I-TNF-a were measured. After binding 12'II-TNF-n to 293/TNF-R2 cells at 4 "C for 1 h, the cells were washed to remove excess, free '*'I-TNF-n, and incubated at 37 "C. At various times, samples were analyzed for cell surface bound plus released TNF-n, internalized TNF-n and demaded TNF-a. After 2 h at 37 "C approximately 40% of cell surface- bound TNF-a was accumulated intracellularly in the 293/ TNF-R2 cells (Fig. 6). Degraded TNF-n was first detected after 2 h and continued to slowly increase to about 5% of the total bound cpm, whereas the surface-bound plus released TNF-n slowly decreased to about 50%. In order to investigate whether the very low level of endogenous TNF-R1 in 293 cells contributed to the observed internalization and degradation of TNF-a, 293/TNF-R2 cells were preincubated with an anti- TNF-R1 monoclonal antibody that blocks binding of TNF-n to TNF-R1. This antibody had no effect on internalization. cell surface association, or degradation of '"''I-TNF-n (Fig. 6).
Induction of Soluble TNF-R2 Sheddinx-The shedding of the extracellular portion of TNF-R2 from 293/TNF-R2 cells observed during the radioimmunoprecipitation experiment was further investigated to see if this processing is affected by TNF. 293/TNF-R2 cells were treated with media alone, TNF-a, TNF-0, or the phorbol ester, PMA. At various times after induction, the amount of sTNF-R2 released from the

21176 75-kDa TNF Receptor Characterization
A 1 2 3 4 5 6
kD - 106 - 80
- 49.5 - 32.5
- 18.5 - 27.5
B -TNF-a + TNF-a -*. ~ ~ ..- . . ~ ". . . .I ...
p-se* P-Thr
P-Tyr
FIG. 5. P o n d A , [".'l']orthophosphnte laheling o f TNF-R2. 293/ TNF-R2 cells were laheled with ,'''l', for 4 h as described under "Materials and Methods." Cells were either stimulated with media alone or 2 nM TNF-tu for 15-30 min. Cell lysates were prepared from the labeled cells and immunoprecipitated with TNF-RZ polyclonal antiserum. The immunoprecipitated samples were resolved on a 4- 20"% Tris/glycine polyacrylamide gel and the gel exposed to film. Lanes 1-3, immunoprecipitates from cells treated with media alone; lanes 4-6, immunoprecipitates from cells treated with 2 nM TNF-n. The positions of prestained molecular weight standards are indicated on the right. Pancl H , phosphoamino acid analysis of TNF-R2. 2921 TNF-R2 cells were metabolically labeled with ."Pi and processed as descrihed above. Immunoprecipitated "'P-labeled TNF-R2 was ex- cised from the gel, eluted, and then analyzed for phosphoamino acid content by two-dimensional elect.rophoresis as descrihed previously (39). The thin layer chromatography plates were exposed to film for 13 days. The migration positions of phosphoserine, phosphothreo- nine, and phosphotyrosine are indicat.ed.
cells and appearing in the media was measured by Western analysis. Constitutive shedding of TNF-R2 occurred at low but detectable levels throughout the duration of the experi- ment (Fig. 7). TNF-a a t 1 nM concentration did not affect the amount of sTNF-R2 released. Treatment of cells with 10 nM TNF-a or 10 nM TNF-/3 also had no effect on the rate of shedding.' However, within 5 min of PMA treatment, there was a rapid release of sTNF-R2 from the cells which continued to accumulate over the course of the experiment (Fig. 7).
DISCUSSION
We have constructed a human 293 cell line that expresses the human TNF-R2 (75-kDa T N F receptor) a t levels more than 100-fold above the endogenous level of T N F receptors on 293 cells. This high level of overexpression facilitated an examination of the biochemical characteristics of TNF-R2.
The TNF-R2 expressed on the surface of 293 cells was found to bind both TNF-a (Kd = 0.2 nM) and TNF-/3 (Kd = 1.1 nM) with high affinity. Others (16, 22), using transiently expressing TNF-R2 in COS cells, have reported similar bind- ing constants. The number of binding sites on the 293/TNF- R2 cells for TNF-a was found to be lower than for TNF-/3 (94,000 uersu.7 264,000). All 264,000 TNF-/3 sites can be com- peted effectively by TNF-a:' demonstrating that these sites are not specific for TNF-/3 only. Two possible explanations
2- I / 0 t - C = .
L
0 1 2 3 4 5 Time (hours)
FIG. 6. '*'I-TNF-a internal izat ion and degradat ion by 293/ TNF-R2 cells. 29:3/TNF-H2 cells were incrhntrtl with 0.2.5 nM " . I - TNF-n for 1 h at 4 "C. Cells were washed twice then shifted to 37 "C for 0 , 5, l5,30,60, 120, and 240 min. A t the indicated times. duplicate samples of 5 X 10" cells were processed as described under "Materials and Methods." Cirrles. acid-washed pellet cpm (internalized '."I-'TXVF- n); squares. non-trichloroacetic acid-precipitable cpm (rley.rnded ""I- TNF-tr); and trianKlrs. trichloroacetic acid-precipitated srlprrnatnnt cpm derived from acid-washed cell supernatants (cell surfare-associ- ated ""I-TNF-tr). Oprn synhob represent data ohtained in the pren- ence of anti-TNF-R1 monoclonal antihotly (4E4). ~ ' h s d symbols Rre without antibody.
Control (Media only)
+ TNF-a "
- "Y"
FIG. 7. Induction by PMA of TNF-R2 shedding. 29B/TNF- R2 cells were incuhated with media only, "SF-n ( I nM 1 or I'MA ( : E nM) and 15-111 aliquots of the media removed at 0. 5. 15, 30 min and 1, 2, 4, 6, and 19 h after induction. The samples were resolved on a 4-20?; Tris/glycine polyacrylamide gel followed by \Vestern hlot analysis. One ng of purified sTNF-H2 was also resolved on the gel as an internal control and appears on the ripht hand sidr of earh row.
for the discrepancy in number of sites are that TNF-n and TNF-P may hind with different stoichiometries or that not all of the ""I-TNF-8 is active. We favor the latter interpre- tation, since Scatchard analysis, size exclusion chromatogra- phy, and cross-linking suggest that two or three molecules of the recombinant extracellular domain of TNF-R2 (sTNF-R2) can hind to a single trimer of TNF-n or TNF-P.'
A range of sizes for cross-linked T N F receptor-ligand com- plexes using laheled TNF-a and TNF-[j on human and murine cell lines has been reported (45. 46). We observe a single
' D. Pennica. V. T. Lam, R. F. Weher. It'. J. Kohr, I,. J . Hasa. M. W. Spellman, A. Ashkenazi, and D. \I. Coeddel. manuscript in prep- aration.

75-kDa TNF Receptor Characterization 21177
distinct species after cross-linking TNF-R2 with TNF-a and two major bands after cross-linking with TNF-/3. The com- plexes we observe appear to be significantly larger than what has been previously reported (205-244 uers'sus -75-100 kDa). The most likely explanation for this discrepancy is that our cross-linking studies were done at room temperature rather than at 4 "C. When we perform cross-linking at 4 "C, the major species observed3 are smaller sized complexes, with sizes consistent with those reported previously (45, 46). At room temperature the fluidity of the cell membrane could allow lateral movement and ligand-mediated oligomerization of TNF-R2, resulting in higher molecular mass ligand-recep- tor (205-244 kDa) complexes. One can speculate that the major high molecular weight complexes observed are com- posed solely of a single trimer of TNF-a or TNF-/3 bound to two or three TNF-R2 molecules. It is possible that only this one type of complex exists and that the minor products observed at smaller sizes are a result of incomplete cross- linking of receptor molecules with ligand. The complexes might also contain receptors cross-linked to associated regu- latory proteins since the cross-linking reagent used in these studies is membrane permeable. Hayakawa et al. (47) have suggested that TNF receptors purified from human placental membranes may be complexed with a guanine nucleotide- binding protein (47) and other workers have suggested that the Fas antigen may be associated with TNF receptors (48).
The mechanism of signal transduction through TNF-R2 remains obscure. One interesting feature of the cytoplasmic region of TNF-R2 is that it contains an unusually high number of serine, threonine, proline, and glutamic acid resi- dues. Proteins that are rich in these residues are often rapidly degraded intracellularly (49). Whether these residues are im- portant in TNF-R2 degradation, or signaling, however, re- mains to be determined. Another feature of the intracellular domain of TNF-R2 is the lack of any tyrosine residues or of any potential phosphorylation sites for CAMP-dependent ki- nase. There is, however, one potential site for protein kinase C phosphorylation at Ser-415 (16, 50). TNF-R2 isolated from 32P-labeled cells was found to be phosphorylated largely on serine residues with very low levels of phosphorylation on threonine residues. This pattern of phosphorylation appeared to be unchanged upon TNF-a treatment. Others have re- ported the constitutive phosphorylation of natural TNF-R2 in SW480T cells (51). However, the significance of TNF-R2 phosphorylation in 293 or SW 980T cells is unclear since no TNF-R2-specific responses have yet been demonstrated in these cells. A serine-rich region in the cytoplasmic domain of the human interleukin-2 receptor /3 chain was found to be essential for ligand-mediated signal transduction through this receptor (52). CDw40, another member of the TNF receptor superfamily, has been found to be constitutively phosphoryl- ated in the cytoplasmic domain (53). Although the importance of serine and threonine phosphorylation in mediating signal transduction through TNF-R2 is not known, the fact that the serine and threonine residues are highly conserved between mouse and human (19) suggests a possible role in signaling.
Previous reports have indicated that following binding to cell surface receptors, TNF-a is rapidly internalized at tem- peratures above 15 "C and subsequently degraded (9, 10, 54). Electron microscopy studies using gold particle-labeled TNF- a have shown that TNF-a is internalizedin L929 cells through clathrin-coated pits and endosomes and ends up in lysosomes where it is degraded (55). It was not determined in these studies, however, whether internalization occurred through TNF-R1 or TNF-R2. However, Yoshie et al. (10) used Hela cells which have predominantly TNF-R1 and undetectable
TNF-R2 (42). Two other reports investigated internalization of human TNF-a in mouse cells (9, 54). As human TNF-a cannot recognize mouse TNF-R2 (19), these results were also specific for TNF-R1. In all three cases, 80-90% of the TNF- a bound to TNF-R1 was rapidly internalized and degraded. Our results indicate that TNF-a bound to 293/TNF-R2 cells is more slowly internalized and much less efficiently degraded. After 2 h at 37 "C, only about 40% of the "'1-TNF-a initially bound at the cell surface was internalized, and less than 10% was degraded after 4 h. Unlike TNF-R1, TNF-R2 has no tyrosines in its cytoplasmic domain and therefore lacks a consensus sequence for rapid cellular internalization via coated pits (56, 57). This suggests that internalization and degradation of TNF-a proceeds by different mechanisms de- pending on whether it is bound to TNF-R1 or TNF-R2.
The soluble form of TNF-R2 was slowly, but spontaneously, released into the media from 293/TNF-R2 cells, and treat- ment of these cells with the phorbol ester PMA rapidly and dramatically increased the rate of TNF-R2 shedding (from < 0.3 ng/ml/106 cells at time 0 to approximately 20 ng/ml/106 cells after 30 min). Neither TNF-a nor TNF-P induced the formation of soluble TNF-R2. Down-regulation of cell surface TNF-binding sites by phorbol esters and interleukin 1 has been reported previously, although neither the specific TNF receptor involved nor the mechanism of loss of TNF binding were known (58-60). Proteolytic release of many membrane proteins is a specific and regulated event and is enhanced after activation of the host cell. This has been found to be a protein kinase C-dependent process for TNF receptors (58- 61), the Mel-14 antigen (62), CSF-1 receptor (63), and pro- TGF-a (64). It has recently been determined that the protease elastase is responsible for the shedding of the extracellular domain of TNF-R2 from activated neutrophils while having no effect on the shedding of TNF-R1 from endothelial cells (65). Soluble TNF-binding proteins have been associated with certain disease states. For example, increased levels of both soluble TNF-R1 and TNF-R2 have been found in the sera of cancer patients (66). Although the physiological significance of TNF receptor shedding is unknown, the production of soluble receptor forms may be involved in the modulation of TNF activities.
The individual role of TNF-R1 and TNF-R2 in initiating signal transduction is only beginning to be understood. For example, antibodies against TNF-R1 were found to act as specific agonists for this receptor and initiate several TNF responses (30, 67-70). More recently, we have found that rabbit polyclonal antibodies to human and murine TNF-R1 and TNF-R2 each acted as specific receptor agonists and induced a subset of TNF activities (32, 71). Taken together, these studies provide evidence that TNF-R1 and TNF-R2 are not redundant in mediating cellular responses and support the existence of receptor-specific signal transduction path- ways.
The mechanism(s) by which TNF-R2 signals these cellular responses, however, has not yet been well defined. Recent evidence suggests that, similar to TNF-R1 (30, 32, 68), the initiation of signal transduction by TNF-R2 involves receptor aggregation. These studies (32) have shown that TNF-R2- specific polyclonal antibodies act as receptor agonists sug- gesting that receptor clustering provides a sufficient signal for certain TNF effects. Data from our cross-linking experi- ments also provide evidence for TNF-R2 aggregation induced by both TNF-a and TNF-P. Studies with a recombinant extracellular domain of TNF-R2 show that one trimer of TNF-a or TNF-/3 can simultaneously bind two to three sTNF- R2 molecule^,^ thereby providing a mechanism for TNF-

21178 75-kDa TNF Receptor Characterization
induced receptor cross-linking. Ligand-induced receptor ag- gregation has been observed for a number of growth factor receptors including epidermal growth factor receptor, platelet- derived growth factor receptor, colony-stimulating factor 1 receptor, and the insulin receptor and is thought to be the initiating event in the signal transduction process (72-74). Although the early stages of signal transduction by TNF-R1 and TNF-RZ may be initiated similarly by receptor aggrega- tion, the wide diversity of cellular responses mediated through the two TNF receptors will probably be shown to be depend- ent on a diversification of post-receptor signaling mecha- nisms.
Acknowledgments-We thank Dr. Avi Ashkenazi for preparation of 1261-TNF-a and 12SI-TNF-fl, Greg Bennett for preparation of the anti-human TNF-R2 polyclonal antiserum, Kimberly Rosenthal for supplying the sTNF-R1 and sTNF-R2 monoclonal antibodies, Dr. Richard Vandlen for help with the phosphoamino acid quantitation, Dr. Louis Tartaglia for helpful suggestions, and Bunnie Jack, Sunita Sohrabji, and Kerrie Andow for preparation of the manuscript and graphics.
REFERENCES 1. Vilcek, J., and Lee, T. H. (1991) J. Biol. Chem. 266,7313-7316 2. Camussi, G., Albano, E., Tetta, C., and Bussolino, F. (1991) Eur. J. Biochem.
3. Fiers, W. (1991) Fed. Eur. Biochem. SOC. 286,199-212 4. Jaattela, M. (1991) Lab. Invest. 6 4 , 724-742 5. Rubin, B. Y., Anderson, S. L., Sullivan, S. A., Williamson, B. D., Carswell,
6. Kull, F. C., Jacobs, S., and Cuatrecasas, P. (1985) Proc. Natl. Acad. Sci.
7. Baglioni, C., McCandless, S., Tavernier, J., and Fiers, W. (1985) J. Biol.
8. Aggarwal, B. B., Eessalu, T. E., and Hass, P. E. (1985) Nature 318 , 665-
202,3-14
E. A., and Old, L. J. (1985) J. Exp. Med. 162,1099-1104
U. S. A. 82,5756-5760
Chem. 260,13395-13397
fw7 9. Tsi&oto, M., Yip, Y. K., and Vilcek, J. (1985) Proc. Natl. Acad. Sci.
U. S. A. 82.7626-7630 10. Yoshie, 0.. Tada, K., and Ishida, N. (1986) J. Biochem. (Tokyo) 100,531-
SA1 11. Jones, E. Y., Stuart, D. I., and Walker, N. P. C. (1989) Nature 338 , 225-
228 12. Eck, M. J., Ultsch, M., Rinderknecht, E., deVos, A. M., and Sprang, S. R.
(1992) J. Biol. Chem. 267,2119-2122 13. Loetscher, H., Pan, Y.-C. E., Lahm, H. W., Gentz, R., Brockhaus, M.,
Tabuchi, H., and Lesslauer, W. (1990) Cell 61,351-359 14. Schall, T. J., Lewis, M., Koller K. J., Lee A., Rice G. C., Wong, G. H. W.,
Gatanaga T., Granger, G. A., Lentz, R., Raab, H., Kohr, W. J., and
15. Gray, P. W., Barrett, K., Chantry, D., Turner, M., and Feldman, M. (1990) Goeddel D. V. (1990) Cell 61,361-370
16. Smith, C. A., Davis. T., Anderson, D., Solam, L., Beckmann, M. P., Jerzy, Proc. Natl. Acad. Sei. U. S. A. 8 7 , 7380-7384
R.. Dower. S. K.. Cosman. D.. and Goodwin. R. G. (1990) Science 248, 1019-1023
17. Himmler, A., Maurer-Fogy, I., Kronke, M., Scheurich, P., Pfizenmaier, K., Lantz. M.. Olsson. I.. HauDtmann. R.. Stratowa. C.. and Adolf. G. R.
. .
(1990)' DNA Cell &i9, 765-715 . . . .
18. Nophar, Y., Kemper, O., Brakebusch, C., Engelmann, H., Zwang, R., Aderka, D., Holtmann, H., and Wallach, D. (1990) EMBO J. 9 , 3269- """
19. Lewis, M., Tartaglia, L. A., Lee, A., Bennett, G. L., Rice, G. C., Wong, G. H. W., Chen, E. Y., and Goeddel, D. V. (1991) Proc. Natl. Acad. Sci. U. S. A. 88,2830-2834
20. Goodwin, R. G., Anderson, D., Jerzy R., Davis, T., Brannan, C. I., Copeland, N. G., Jenkins, N. A., and Smith, C. A. (1991) Mol. Cell. Biol. 11,3020-
si IL)
21
22
23
24
25 26
27. 28.
. Kohno, T., Brewer M. T., Baker S. L., Schwartz, P. E., King, M. W., Hale 3026
Natl. Acad. Sci. U. S. A. 87,8331-8335 K. K., Squires, C. H., Thompson, R. C., and Vannice J. L. (1990) Proc.
. Dembic, Z., Loetscber, H., Gubler, U., Pan, Y.-C. E., Lahm, H. W., Gentz, R., Brockhaus, M., and Lesslauer, W. (1990) Cytokine 2,231-237
. Johnson, D., Lanaham, A., Buck, C. R., Sehgal, A., Morgan, C., Mercer, E., Bothwell, M., and Chao, M. (1986) Cell 47,545-554 . Itoh, N., Yonehara, S., Ishii, A., Yonehara, M., Mizushima, S., Sameshima, M., Hase, A., Seto, Y., and Nagata, S. (1991) Cell 66,233-243
. Stamenkovic, J., Clark, E. A., and Seed, B. (1989) EMBO J. 8,1403-1410
. Camerini, D., Walz, G., Loenen, W. A. M., Borst, J., and Seed, B. (1991) J.
Mallet, S., Fossum, S., and Barclay, A. N. (1990) EMBO J. 9,1063-1068 Smith, C. A., Davis, T., Wignall, J. M., Din, W. S., Farrah, T., Upton, C.,
Imm~nol . 147,3165-3169
mun. 176,335-342 McFadden, G., and Goodwin R. G. (1991) Biochem. Biophys. Res. Com-
29. Upton, C., Macen, J. L., Schreiber, M., and McFadden, G. (1991) Virology 184,370-382
30. Engelmann, H., Holtmann, H., Brakebusch, C., Avni, Y. S., Sarov, I., Nouhar. Y.. Hadas. E.. Leitner. 0.. and Wallach. D. (1990b) J. Biol. C h m . 266; 14497-'14564
. . , . I
31. Esoevik. T.. Brockhaus. M.. Loetscher. H.. Nonstad. U.. and Shalabv. R. i m o j J. 'ExP. Med. i71,'415-426
I , ~ , ~ I ~~ ~~~~ < , ~
32. Tarta lia, L. A., Weber, R. F., Figari, I. S., Reynolds, C., Palladino, M. A.,
33. Miyata, A., Kangawa, K., Toshimori, T., Hato, T., and Matsuo, H. (1985) anfGoedde1, D. V. (1991) Proc. Natl. Acad. Sei. U. S. A. 88,9292-9296
34. Pennica, D., Kotr, W. J , Fendly, B. M., Shire, S. J., Raab, H. E., Borchardt, Biochem. Bio hy Res Commun. 129,248-255
35. Seed, B., and Aruffo, A. (1987) Proc. Natl. Acad. Sci. U. S. A. 8 4 , 8573- P. E., Lewis, M., and Goeddel, D. V. (1992) Biochemistry 31,1134-1141
36. Sambrook, J., Fritsch, E. F., and Maniatis, T. (1989) Molecular Cloning: A 8577
Luboratory Manual, 2nd ed., Cold Spring Harbor Laboratory, Cold Spring
37. Graham, F. L., Smiley, J., Russell, W. C., and Nairn, R. (1977) J. Gen.
38. Opp:Eann, H., Levinson, A. D., and Varmus H. E. (1981) Virology 108 ,
Harbor, NY
Virol. 3 6 , 59-77
39. Hunter, T., and Sefton, B. M. (1980) Proc. Natl. Acad. Sci. U. S. A. 77 ,
40. Shyjan, A. W., and Levenson, R. (1989) Biochemistry 28,4531-4535 41. Hohmann, H., Remy, R., Brockhaus, M., and van Loon, A. P. G. M. (1989)
J. Biol. Chem. 264,14927-14934 42. Brockhaus, M., Schoenfeld, H. J., Schlaeger, E. J., Hunziker, W., Lesslauer,
W., and Loetscher, H. R. (1990) Proc. Natl. Acad. Sci. U. S. A. 87,3127-
43. Loetscher, H., Schlaeger, E. J., Lahm H. W., Pan, Y.-C. E., Lesslauer, W., 3131
44. Heller, R., Song, K., Onasch, M. A,, Fischer, W. H., Chang, D., and Ringold, and Brockhaus, M. (1990) J. Biol. Chem. 266,20131-20138
45. Smith, R. A., and Baglioni C. (1992) Tumor Necrosis Factors, Structure, G. M. (1990) Proc. Natl. Acad. Sci. U. S. A. 87,6151-6155
Function, and Mechanism ofAction (Ag m a l , B. B., and Vilcek, J., eds) pp. 131-160, Marcel Dekker Inc., New fork
46. Loetscher, H. (1992) Tumor Necrosis Factors, Structure, Function, and Mechanism of Action (Agganval, B. B., and Vilcek, J., eds) pp. 160-187,
47. Ha akawa, M., Hori, T., Shibamoto, S., Tsujimoto, M., Oku, N., and Ito, Marcel Dekker Inc., New York
48. Yonehara, S., Ishii, A., and Yonehara, M., (1989) J. Exp. Med. 169,1747- I? (1991) Arch. Biochem. Biophys. 286,323-329
49. Rogers, S., Wells, R., and Rechsteiner, M. (1986) Science 234,364-368 1756
50. Woodgett, J. R., Gould K. L., and Hunter, T. (1986) Eur. J. Biochem. 161,
4 I - ' IU
1311-1315
51. Hohmann, H-P., Remy, R., Aigner, L., Brockhaus, M., and van Loon, A.
52. Ht+eyama, M., Mori, H., Doi, T., and Taniguchi, T. (1989) Cell 69,837-
177-184
P. G. M. (1992) J. Biol. Chem. 267,2065-2072
53. Inui, S., Kaisho, T., Kikutani, H., Stamenkovic, I., Seed, B., Clark, E. A,, ti45
54. Watanabe, N., Neda, H., Ohtusuka, Y., Sone, H., Yamauchi, N., Maeda, and Kishimoto, T., (1990) Eur. J. Immunol. 20,1747-1753
M., Kuriyama, H., and Niitsu, Y. (1989) Cancer Immunol. Immunother.
55. Mosselmanns, R., Hepburn, A., Dumont, J. E., Fiers, W., and Galand, P. 28,157-163
(1988) J. Immunol. 141,3096-3100 56. Chen. W-J.. Goldstein. J. L., and Brown, M. S. (1990) J. Biol. Chem. 266 ,
3ii6-3123 57. Collawn, J. F., Stangel, M., Kuhn, L. A,, Esekogwu, V., Jing, S., Trowbridge,
58. Holtmann, H., and Wallach D. (1987) J. Immunol. 139,1161-1167 59. Aggarwal B. B., and Eessalu T. E. (1987) J. Biol. Chem. 262,16450-16455 60. Unglaub, R., Maxeiner, B., Thoma, B., Pfizenmaier, K., and Scheurich, P.
61. Lantz, M., Gueberg, U., Nilsson, E., and Olsson, I. (1990) J. Clin. Invest.
62. Kisbimoto, T. K., Jutila, M. A., Berg, E. L., and Butcher, E. C. (1989)
63. Downing. J. R., Roussel, M. F., and Sherr, C. J. (1989) Mol. Cell. Biol. 9,
I. S., and Tainer, J. A. (1990) Cell 63,1061-1072
(1987) J. Ex Med. 166,1788-1797
86,1396-1402
Science 246,1238-1241
2890-%396 64. Pandiella, A., and Massame, J. (1991) Proc. Natl. Acad. Sci. U. S. A. 8 8 ,
1726-1730
(1991) J. Biol. Chem. 266,18846-18853
D. and Kushtai G. (1991) Cancer Res. 51,5602-5607
and van'Loon:'A. P. G. M. (1990b) J. Biol. Chem. 266: 22409-22417
pl990) J. Exp. Med. 171,415-426
and 6spe;ik; T. (1990) J. Exp. Med. 172, 15617-1520
146,3045-3048
65. Porteu, F., Brockhaus, M., Wallach, D., Engelmann, H., and Nathan, C. F.
66. Aderka, D., Engelmann, H., Hornik, V., Skornick, Y., Levo, Y., Wallach,
67. Hohdann H. P Brockhaus M., Baeurerle, P. A., Remy R., Kolbeck, R.,
68. Es evik, T., Brockhaus, M., Loetscher, H., Nonstad, U., and Shalaby, R.
69. Shalab , M R , Sundan, A,, Loetacher, H., Brockhaus, M., Lesslauer, W.,
70. Naume, B., Shalaby, R., Lesslauer, W., and Espevik, T. (1991) J. Immunol.
71. Tarta lia L. A. and Goeddel, D. V. (1992) Immunol. Today 13 , 151-153 72. Boni-Echketzle;, M., and Pilch, P. F. (1987) Proc. Natl. Acad. Sci. U. S. A.
73. Yarden Y. and Schlessinger, J. (1987) Biochemistry 26,1434-1442 74. Yarden: Y.: and Schlessinger, J. (1987) Biochemistry 26,1443-1451
84,7832-7836





























