Embed Size (px)
Citation preview

MA07CH03-Lowe ARI 29 October 2014 6:57
Oceanic Forcing of Coral ReefsRyan J. Lowe and James L. FalterARC Centre of Excellence for Coral Reef Studies, School of Earth and Environment,and UWA Oceans Institute, University of Western Australia, Crawley 6009, Australia;email: [email protected], [email protected]
Annu. Rev. Mar. Sci. 2015. 7:43–66
First published online as a Review in Advance onSeptember 17, 2014
The Annual Review of Marine Science is online atmarine.annualreviews.org
This article’s doi:10.1146/annurev-marine-010814-015834
Copyright c© 2015 by Annual Reviews.All rights reserved
Keywords
coral reefs, waves, tides, circulation, temperature, nutrients
Abstract
Although the oceans play a fundamental role in shaping the distribution andfunction of coral reefs worldwide, a modern understanding of the complexinteractions between ocean and reef processes is still only emerging. Thesedynamics are especially challenging owing to both the broad range of spatialscales (less than a meter to hundreds of kilometers) and the complex physicaland biological feedbacks involved. Here, we review recent advances in ourunderstanding of these processes, ranging from the small-scale mechanicsof flow around coral communities and their influence on nutrient exchangeto larger, reef-scale patterns of wave- and tide-driven circulation and theireffects on reef water quality and perceived rates of metabolism. We alsoexamine regional-scale drivers of reefs such as coastal upwelling, internalwaves, and extreme disturbances such as cyclones. Our goal is to show howa wide range of ocean-driven processes ultimately shape the growth andmetabolism of coral reefs.
43
Ann
u. R
ev. M
arin
e. S
ci. 2
015.
7:43
-66.
Dow
nloa
ded
from
ww
w.a
nnua
lrev
iew
s.or
g A
cces
s pr
ovid
ed b
y U
nive
rsity
of
Wes
tern
Aus
tral
ia o
n 01
/30/
15. F
or p
erso
nal u
se o
nly.

MA07CH03-Lowe ARI 29 October 2014 6:57
Upwelling: theupward movement ofwater resulting fromwind-driven Ekmantransport or flowcurvature in thevicinity of islands
Internal waves:wave motions thatpropagate within theinterior of adensity-stratifiedocean
1. INTRODUCTION
The physics, chemistry, and biology of the world’s oceans have long been known to play a criticalrole in shaping the growth, diversity, and distribution of coral reefs. Darwin (1842) first hypoth-esized that the surrounding oceans provide a major source of nutrition to coral reef communitiesand that reefs therefore grow best at their most seaward edges. More than a century later, Munk &Sargent (1954) proposed that the energy dissipated by waves on shallow fore reefs and reef crestsfacilitates the rapid exchange of nutrients and other metabolites necessary to support the robustgrowth of reef communities in otherwise nutrient-poor environments. This basic idea would stim-ulate modern research aimed at understanding how hydrodynamic processes in benthic boundarylayers control the uptake of dissolved nutrients by reef communities (Atkinson & Bilger 1992,Falter et al. 2004, Hearn et al. 2001, Lowe et al. 2005b, Thomas & Atkinson 1997). Additionalwork has since provided more direct evidence of how reefs also feed off the living plankton washedover them from the surrounding ocean (Ayukai 1995, Charpy & Blanchot 1999, Fabricius et al.1998, Wyatt et al. 2010). Nevertheless, the same ocean waves driving the circulation and materialexchange that are beneficial to the growth of reef organisms can also act as a destructive forcewhen large enough, especially when they are generated by large storms such as tropical cyclones(Massel & Done 1993, Woodley et al. 1981). Extreme wave events can act as an additional sourceof physical stress on reef systems already contending with other forms of environmental distur-bance (De’ath et al. 2012) and may also lead to long-term changes in reef community structureand diversity (Dollar & Tribble 1993, Madin & Connolly 2006).
The environment in which reefs live is further shaped by ever-changing ocean conditions atscales much greater than the reef itself. At the shelf or island scale, coastal upwelling caused bywind-driven Ekman transport or generated in the lee of islands can bring up cooler, more nutrient-and particle-rich water. Such environments can support higher rates of nutrient acquisition (Goveet al. 2006, Wyatt et al. 2010), create adverse chemical conditions for the formation of biogenic car-bonate minerals (Manzello 2010), and offer potential refugia against ocean warming (Karnauskas& Cohen 2012). Nonlinear internal waves propagating up continental shelf and island slopesprovide another potential mechanism for transporting cool water and nutrients to shallow coralreefs (Leichter et al. 1996, Wang et al. 2007). At even larger scales, regional circulation patternscontrol the widespread dispersal of reef larvae and, therefore, the biogeographical distribution ofreef organisms (e.g., Richmond 1987, Shulman & Bermingham 1995).
In the future, the oceans will continue to play an important role in determining how globalclimate trends such as increased warming and the dissolution of CO2 will manifest themselves, or“downscale,” at the regional, coastal, shelf, reef, community, and ultimately organism scales. Forexample, although mean global temperatures are expected to increase by ∼4◦C by the end of thecentury under a business-as-usual scenario (Meinshausen et al. 2009), seasonal warming eventsdriving sea surface temperature anomalies of several degrees are already a regular occurrence. Suchintense ocean warming events occurred across the western Pacific and Caribbean in 1998 owing toparticularly intense El Nino conditions (e.g., Bruno et al. 2001, McClanahan et al. 2004) and alongWestern Australia in 2010–2011 owing to the intensification of a warm eastern boundary currentduring an intense La Nina period (Depczynski et al. 2013, Moore et al. 2012). Within the reefsystems themselves, benthic production, respiration, and net calcification can alter the chemistryof circulating water (Kinsey 1978, Odum & Odum 1955, Sargent & Austin 1949, Smith 1973);however, the magnitudes of these changes depend just as much on the degree of hydrodynamicforcing and the morphologies of the reef systems as they do on rates of benthic metabolism(Falter et al. 2013, Venti et al. 2012, Zhang et al. 2012). Thus, changes in the global wave andstorm climate that are expected to occur over the next century (Young et al. 2011) will no doubt
44 Lowe · Falter
Ann
u. R
ev. M
arin
e. S
ci. 2
015.
7:43
-66.
Dow
nloa
ded
from
ww
w.a
nnua
lrev
iew
s.or
g A
cces
s pr
ovid
ed b
y U
nive
rsity
of
Wes
tern
Aus
tral
ia o
n 01
/30/
15. F
or p
erso
nal u
se o
nly.

MA07CH03-Lowe ARI 29 October 2014 6:57
Canopy flow:a region of flow insidethe bottom roughnesslayer created by reeforganisms
lead to concomitant changes to many of the above biogeochemical and ecological processes onreefs.
It is clear that the growth, metabolism, and community structure of coral reefs are heavily influ-enced by the oceanic environment in which they live. Unfortunately, it is not possible for a singlereview to cover all important aspects of the coupling between physical and biological processes oncoral reefs. There have been some prior efforts to summarize the influence of oceanic processes oncoral reefs, although these either were published several decades ago (Hamner & Wolanski 1988)or focused mainly on the details of the prevalent hydrodynamic processes (Monismith 2007). Toprovide what we think is a good balance between breadth and detail, the present review focusesfirst on the small- and reef-scale hydrodynamic processes affecting the transport and transfer ofmass (nutrients and carbon, dissolved and particulate), heat (temperature), and momentum (dragand dissipation) on reefs, and second on how offshore ocean processes affect the physical andchemical properties of nearshore reef waters. The review is thus organized by scale, starting withthe smallest scales (individual reef organisms and canopies; Section 2), moving to intermediatescales (reef communities and systems; Section 3), and concluding with the largest scales that in-fluence the growth and development of reefs along the continental margins and oceanic islandgroups (regional and global; Section 4). For reference, Table 1 summarizes the variables usedthroughout the review.
2. ORGANISM TO REEF CANOPY SCALES (0.1–10 m)
The flow structure in and around reef organisms depends on the complex interactions between theoverlying water motion and the typically large, three-dimensional bottom roughness (or canopies)formed by reef organisms (Figure 1a,b). The flow dynamics throughout reef canopies are highlyanalogous to other, more well-studied canopy flows, particularly those through seagrasses andother aquatic vegetation (for a recent review, see Nepf 2012). The common challenge for under-standing the flow dynamics in all natural canopies is properly accounting for the highly variablespatial flow structure that arises within even the simplest morphologies. There have been someattempts to directly observe or numerically simulate the full three-dimensional turbulent flowstructure through individual branching coral colonies (Chang et al. 2009, Chindapol et al. 2013,Kaandorp et al. 2003); however, such efforts are extremely expensive computationally, requiringflows to be resolved down to scales on the order of millimeters. Thus, it remains impractical toresolve the roughness geometries of reef organisms at the scale of entire reef communities orsystems.
To account for how such unresolved small-scale hydrodynamic processes influence the larger-scale macroscopic properties of a canopy flow (e.g., mean velocity profiles), the traditionalReynolds-averaged Navier-Stokes momentum equations are spatially averaged over a horizon-tal plane that excludes the solid canopy elements (Raupach & Shaw 1982):
D〈u〉Dt
= − 1ρ
∂〈P〉∂x
+ 1ρ
∂τxy
∂z− fx, (1)
where x is the streamwise direction, z is the vertical direction, ρ is the seawater density, u is thestreamwise velocity, P is the pressure, τxy is the shear stress, and the brackets indicate spatiallyaveraged variables. Importantly, the term fx represents a spatially averaged canopy resistance termthat arises from forces exerted by the canopy onto the surrounding flow (Lowe et al. 2005a).
Ultimately, the fundamental challenge in predicting flow through coral reef canopies via Equa-tion 1 remains how to most accurately relate fx to the complicated three-dimensional structure ofreef communities, especially given their abundant variation in morphologies and roughness scales.
www.annualreviews.org • Oceanic Forcing of Coral Reefs 45
Ann
u. R
ev. M
arin
e. S
ci. 2
015.
7:43
-66.
Dow
nloa
ded
from
ww
w.a
nnua
lrev
iew
s.or
g A
cces
s pr
ovid
ed b
y U
nive
rsity
of
Wes
tern
Aus
tral
ia o
n 01
/30/
15. F
or p
erso
nal u
se o
nly.

MA07CH03-Lowe ARI 29 October 2014 6:57
Table 1 Summary of variables used in this review
Variable Unit DescriptionA∞ m Wave orbital excursion amplitudeC mol m−3 Water column species concentrationCd,c — Canopy element drag coefficientCD — Total bed-form drag coefficientCM — Canopy element inertial force coefficientfx m s−1 Spatially averaged canopy resistanceh m Water depthhc m Canopy heighthr m Depth of the reef flatH0 m Offshore wave heightHs m Significant wave heightJ mol m−2 h−1
W m−2Net flux of heat or mass across the benthic and/or air-sea interface
K m2 s−1 Horizontal dispersion coefficientLr m Length of the reef flat from crest to backreefMTR m Mean tidal rangeP kg m−1 s−2 Pressureq m2 s−1 Depth-integrated transportq m2 s−1 Two-dimensional depth-integrated transport vectorRek — Roughness Reynolds numberS m d−1 First-order nutrient uptake rate coefficientSc — Schmidt numberT ◦C TemperatureTA μeq kg−1 Total alkalinity〈u〉 m s−1 Horizontal spatially averaged streamwise velocityU m s−1 Depth-averaged current velocityU ∞ m s−1 Free stream velocityU m s−1 Canopy depth-averaged streamwise velocityα — Canopy flow attenuation parameterβ m−1 Porous media drag parameterη m Dynamic sea level or wave setupλf — Frontal area canopy geometry parameterτcan kg m−1 s−2 Shear stress at the top of the canopyτw s Wave periodτRT h Residence time of water within the reef domain
For natural canopies with simple morphologies (e.g., seagrasses), these form-drag forces can bereadily parameterized using a quadratic drag law that depends on the local in-canopy velocity, theintegrated frontal area of the canopy elements per unit area of the seafloor (λf ), and an empiricaldrag coefficient (Cd,c) (for details, see Nepf 2012). Such approaches have been applied to coral reefcanopies with well-defined branch geometries (e.g., Figure 1a) (Lowe et al. 2005a); however, thecomplex morphologies of most reef canopies (e.g., Figure 1b) make assigning a representative λf
difficult or even impossible because of the challenge of identifying singular, well-defined geomet-ric parameters. As an alternative approach to address this issue, Lowe et al. (2008) used turbulent
46 Lowe · Falter
Ann
u. R
ev. M
arin
e. S
ci. 2
015.
7:43
-66.
Dow
nloa
ded
from
ww
w.a
nnua
lrev
iew
s.or
g A
cces
s pr
ovid
ed b
y U
nive
rsity
of
Wes
tern
Aus
tral
ia o
n 01
/30/
15. F
or p
erso
nal u
se o
nly.

MA07CH03-Lowe ARI 29 October 2014 6:57
‹u›(z)
a b
c ‹u›rms(z)d
Currents Waves
τcan τcan
dP/dxdP/dx
Figure 1(a,b) Examples of bottom roughness on reefs, showing a homogeneous community of Porites compressa forming a canopy of regularcylindrical branches (panel a; photograph by R.J. Lowe) and a more complex reef canopy of plating and branching Acropora spp. (panelb; photograph by the Western Australia Department of Parks and Wildlife). (c,d ) Flow structure under different conditions. Underunidirectional flow (panel c), the shear stress at the top of the canopy (τcan) transfers momentum from the overlying flow field into thecanopy and is opposed by the canopy drag forces that attenuate the flow (〈u〉) inside the canopy. Under wave-driven oscillatory flow(panel d ), the unsteady wave-induced pressure gradient (dP/dx) provides an additional source of momentum that substantiallyincreases the rms flow (〈u〉rms) inside the canopy for a given flow speed.
porous media flow theory to parameterize fx. The advantage of the porous media approach is thatit has already been applied to a number of complex geometrical arrays and tested with a wealthof empirical data (Macdonald et al. 1979). Although this approach has worked well for predictingflow through a community of Porites compressa coral (Lowe et al. 2008), further work is needed toparameterize flow resistance across a broader spectrum of natural reef canopy types.
2.1. Unidirectional Flow
For a unidirectional current over a submerged reef canopy, the discontinuity in form drag fx nearthe top of the canopy generates a region of strong shear in the mean velocity profile (Equation 1),
www.annualreviews.org • Oceanic Forcing of Coral Reefs 47
Ann
u. R
ev. M
arin
e. S
ci. 2
015.
7:43
-66.
Dow
nloa
ded
from
ww
w.a
nnua
lrev
iew
s.or
g A
cces
s pr
ovid
ed b
y U
nive
rsity
of
Wes
tern
Aus
tral
ia o
n 01
/30/
15. F
or p
erso
nal u
se o
nly.

MA07CH03-Lowe ARI 29 October 2014 6:57
Unconfined flow: theflow that occurs whenthe water depth ismuch greater than thecanopy height; thecanopy flow is drivenby a shear layer
Depth-limited flow:the flow that occurswhen the water depthis comparable to thecanopy height; thecanopy flow isenhanced by thepressure gradient
Emergent flow: theflow that occurs whenthe canopy reaches thewater surface; thecanopy flow is drivensolely by the pressuregradient
resulting in a local peak in the turbulent shear stress τcan (Ghisalberti 2009) (Figure 1c). The shearlayer transfers momentum from the overlying water column down into the canopy, thus drivingflow inside the canopy. This is the dominant form of momentum transfer that occurs when theflow is unconfined, or when the canopy height hc occupies only a small fraction of the total waterdepth h. When hc becomes an appreciable fraction of the water depth, the underlying canopyflow becomes depth limited, and the background pressure gradient driving the overlying flow alsostarts to directly drive flow within the canopy itself (Nepf & Vivoni 2000). As hc/h increases, thepressure gradient term continues to increase the in-canopy flow, reaching a limit where the reefcanopy becomes emergent (hc ∼ h) such that the shear layer is no longer present and flow is drivenentirely through the canopy by the external pressure gradient (Nepf 2012).
At typical reef canopy heights (ranging from tens of centimeters to meters), many or mostflows on coral reef flats (h typically 1–2 m) must experience some degree of depth limitation.This seemingly local effect can also have important consequences for the broader circulationpatterns on reefs (hundreds to thousands of meters), as the larger drag forces that arise from morewater being driven through a canopy under depth-limited conditions increases the resistanceexperienced by the overlying flow. This was well illustrated by McDonald et al. (2006), whofound that the community bottom-drag coefficient CD, which relates the total drag force to theoverlying flow speed (not to be confused with the canopy element drag coefficient Cd,c, describedabove), increased by roughly two orders of magnitude (from ∼0.01 to 1) as the depth over acanopy of P. compressa coral approached the emergent limit (see figure 3 in McDonald et al. 2006).Furthermore, there appears to be a relatively abrupt transition to high (order 0.1–1) values of CD
when hc/h increases above ∼0.3 (Lowe et al. 2008, McDonald et al. 2006). Collectively, thesestudies emphasize that careful consideration of the in-canopy momentum dynamics—a factor thathas been largely ignored in traditional coastal ocean models—is critical to accurately parameterizethe effects of bottom drag on reefs (Rosman & Hench 2011).
2.2. Wave-Driven Flow
The dynamics of wave-driven flows through canopies have historically received limited attention,owing largely to the lack of analogous terrestrial canopy models on which to build. Although themomentum equations describing water motion in canopies under oscillatory and unidirectionalflows are fundamentally similar, there are two key differences: (a) Surface waves can generatelarge oscillatory pressure gradients at the dominant wave period T (Figure 1d ), and (b) flowaccelerations inside the canopy can add an inertial force that increases the overall flow resistancefx in Equation 1. To account for these effects, Lowe et al. (2005a, 2008) developed a simple modeldescribing the momentum dynamics within submerged canopies under wave-driven oscillatoryflow by depth integrating Equation 1 over the canopy region:
dUdt
acceleration
= − 1ρ
dPdx
pressure gradient
+ Cf
2hc|U ∞|U ∞
shear stress
− β|U |Uform drag
− CM(1 − φ)φ
dUdt
inertial force
. (2)
Here, U denotes the spatially averaged in-canopy velocity, Cf is a friction coefficient that relatesthe magnitude of the shear stress at the canopy interface to the free stream velocity U ∞, β is adimensional drag parameter based on a porous media formulation that depends on the internalbed geometry, and CM is an inertial force coefficient (for details, see Lowe et al. 2008). Equation 2predicts that the acceleration of the in-canopy flow (term 1) depends on the pressure gradientand shear stress terms responsible for driving the flow (terms 2 and 3) and the form drag andinertial forces that oppose it (terms 4 and 5). Lowe et al. (2005a) further defined a flow attenuation
48 Lowe · Falter
Ann
u. R
ev. M
arin
e. S
ci. 2
015.
7:43
-66.
Dow
nloa
ded
from
ww
w.a
nnua
lrev
iew
s.or
g A
cces
s pr
ovid
ed b
y U
nive
rsity
of
Wes
tern
Aus
tral
ia o
n 01
/30/
15. F
or p
erso
nal u
se o
nly.

MA07CH03-Lowe ARI 29 October 2014 6:57
Mass-transferlimited: describesconditions underwhich the rate ofuptake or release of acompound is limitedby convectivetransport across aconcentrationboundary layer
parameter α ≡ U rms/U rms∞ to be the ratio of the rms wave velocity inside the canopy (U rms) to
that in the free stream (U rms∞ ). Above the canopy, where the wave-driven flow is assumed to be
inviscid (i.e., above the wave boundary layer), the wave-driven pressure gradient is related solelyto the acceleration of the free stream oscillatory flow. Simple scaling arguments show that theimportance of this pressure gradient term to the shear stress term increases with the ratio of thewave orbital excursion amplitude (Arms
∞ ) to the roughness height of the canopy (Lowe et al. 2005a):
shear stresspressure
∼ U rms∞ Thc
∼ Arms∞hc
. (3)
Thus, similar to the case of severely depth-limited unidirectional flow, the pressure gradientimposed by waves substantially enhances flow inside a reef canopy over a pure unidirectional flowof the same magnitude (Figure 1d ). This theory has been supported by experimental studies usingboth idealized reef canopies and experimental assemblages of real branching coral reef canopies(Lowe et al. 2005a, 2008; Reidenbach et al. 2007), which have all shown that unsteady wave motionsubstantially enhances the in-canopy flow (by a factor of 5–10 in these particular studies).
2.3. Nutrient Uptake
Pioneering work by Atkinson and others demonstrated that the biological demand for nutrientson reefs is generally so high that rates of nutrient uptake by reef communities are limited bytheir convective transfer across turbulent boundary layers formed on the surfaces of benthic reefautotrophs (Atkinson & Bilger 1992, Baird & Atkinson 1997, Thomas & Atkinson 1997). Bilger &Atkinson (1992) described these conditions as being “mass-transfer limited,” and adapted a theoryoriginally developed to describe heat and mass transfer to rough surfaces (from the engineeringliterature) to instead describe nutrient uptake by coral reef communities. Under conditions ofmass-transfer limitation, the maximum mass-transfer-limited nutrient uptake rate (Jmax, in mmolm−2 d−1) is first order with respect to the concentration of nutrients in the water column (C,in mmol m−3), i.e., Jmax = SmaxC , where Smax (in m d−1) is the first-order nutrient uptake ratecoefficient. On coral reefs, the conditions of mass-transfer limitation tend to break down onlywhen dissolved nutrient concentrations are more than two orders of magnitude greater than thosenaturally occurring in tropical reef waters (Atkinson & Falter 2003).
The application of these mass-transfer formulations to the problem of nutrient uptake kineticsin reef communities was an important step because (a) it approached the problem at the morehydrodynamically relevant spatial scale of entire reef communities acting as canopies (see above),rather than just that of individual organisms, and (b) it provided for the first time some predictivecapability for how water motion controls nutrient uptake by reef communities. Subsequent ex-perimental work demonstrated the utility of these mass-transfer relationships to predict rates ofnutrient uptake for natural reef communities in laboratory flumes simulating pure unidirectionalflow (Baird & Atkinson 1997, Thomas & Atkinson 1997). Falter et al. (2004) later simplified Bil-ger & Atkinson’s (1992) original nutrient uptake formulations, which also incorporated additionalexperimental data sets:
Smax =
√CDU ∞
Re0.2k Sc 0.58
, (4)
where U ∞ is the free stream flow speed, Sc is the Schmidt number of the dissolved nutrient, Rek
is the roughness Reynolds number of the flow, and is an empirical constant equal to ∼4 × 104.It is important to emphasize that CD in Equation 4 is a bulk drag coefficient that relates the totalflow resistance exerted by a reef community to the free stream flow speed U ∞ (see above). Owing
www.annualreviews.org • Oceanic Forcing of Coral Reefs 49
Ann
u. R
ev. M
arin
e. S
ci. 2
015.
7:43
-66.
Dow
nloa
ded
from
ww
w.a
nnua
lrev
iew
s.or
g A
cces
s pr
ovid
ed b
y U
nive
rsity
of
Wes
tern
Aus
tral
ia o
n 01
/30/
15. F
or p
erso
nal u
se o
nly.

MA07CH03-Lowe ARI 29 October 2014 6:57
to the practical size limitations of the experimental flumes that were used to develop Equation 4,most of the experimental reef communities reported in the literature occupied up to 50% of thewater depth. Thus, these nutrient relationships have been developed almost exclusively underconditions of substantial depth limitation. This explains the high CD values of ∼0.1–0.2 reportedin those studies (see summary in McDonald et al. 2006), suggesting that flow, and perhaps nutrientuptake, would be enhanced within the interior regions of those experimental communities.
There are still substantial gaps in our understanding of dissolved nutrient uptake by reef com-munities under oscillatory flow conditions. However, a growing number of reports—includingseveral direct flume observations (Lowe et al. 2005b, Reidenbach et al. 2006, Thomas &Cornelisen 2003, Weitzman et al. 2013) and inferences based on field studies (Bilger & Atkinson1992, Falter et al. 2004)—have shown that wave-driven oscillatory flows substantially enhancemass-transfer rates, increasing them by factors of 3 or more. Lowe et al. (2005b) asserted that therelevant velocity scale to describe mass transfer should be the in-canopy flow velocity U ratherthan the free stream U ∞ because this is the velocity that more directly interacts with the surfacesof organisms within the interior of the canopy. Thus, the enhancement of nutrient uptake underoscillatory flow can be explained by the higher flow speeds induced within coral reef canopiesunder these conditions. A similar argument can be made for the enhancement of nutrient uptakeunder depth-limited flow. Nevertheless, further data are needed, especially for live natural coralreef communities, to develop better nutrient uptake relationships that can be universally appliedacross a broad range of flow regimes.
2.4. Particle Uptake
The uptake of organic particles by reef heterotrophs can constitute a source of allochthonousnutrients to coral reefs that is as important as the uptake of dissolved nutrients by reef primaryproducers (Houlbreque & Ferrier-Pages 2009), especially for corals recovering from a recentbleaching event (Ferrier-Pages et al. 2003, Houlbreque & Ferrier-Pages 2009, Houlbreque et al.2004, Hughes & Grottoli 2013). There is now abundant evidence of living phytoplankton beingactively grazed by whole communities of reef organisms as water flows over reefs (Genin et al. 2009;Monismith et al. 2010; Wyatt et al. 2010, 2012; Yahel et al. 1998); however, the phytoplanktonbiomass within the oligotrophic tropical waters of most coral reefs is generally dominated bythe smallest size fraction (i.e., picoplankton <2 μm in size). Several studies have thus suggestedthat most of these small phytoplankton are consumed by cryptic filter feeders such as sponges,bivalves, and ascidians rather than by passive suspension feeders such as coral (Ribes & Atkinson2007, Ribes et al. 2005).
Picoplankton feeding by scleractinian corals has been controversial, in part because the tentaclesizes of these corals (0.1–1 mm) tend to be two to three orders of magnitude larger than thesecells, thus raising questions about whether such small particles can be physically captured (S.Humphries, personal communication). However, recent studies have confirmed that at least somespecies of scleractinian coral are capable of feeding on phytoplankton (Leal et al. 2014) and insome cases even feeding on detrital fragments of macrophytes such as seagrass (Lai et al. 2013).Theory suggests that very small suspended particles such as picoplankton should move with someBrownian diffusion, implying that these particles can be assigned a diffusivity analogous to thatof a dissolved tracer (Espinosa-Gayosso et al. 2012). In this particle diffusion regime, it maythus be reasonable to assume that the same mass-transfer formulations for predicting dissolvednutrient uptake (e.g., Equation 4) could serve as a useful foundation for predicting the uptake ofsmall particles by coral reef communities. Although only very limited data are available to assessthese dynamics, some observations suggest that the uptake rate coefficients (S) for picoplankton,
50 Lowe · Falter
Ann
u. R
ev. M
arin
e. S
ci. 2
015.
7:43
-66.
Dow
nloa
ded
from
ww
w.a
nnua
lrev
iew
s.or
g A
cces
s pr
ovid
ed b
y U
nive
rsity
of
Wes
tern
Aus
tral
ia o
n 01
/30/
15. F
or p
erso
nal u
se o
nly.

MA07CH03-Lowe ARI 29 October 2014 6:57
Residence time: thelength of time that awater parcel remainswithin a domain (thecomplement of waterage)
bacteria, and viruses are of magnitudes similar to those for dissolved nutrients (Patten et al. 2011,Ribes & Atkinson 2007, Wyatt et al. 2012), which may lend some support to this hypothesis.
Larger particles (e.g., zooplankton and larger-sized phytoplankton) are fed on by active con-sumers (e.g., planktivorous fish; Yahel et al. 2005) yet are also large enough to be directly in-tercepted by passive suspension feeders such as corals (Humphries 2009). Several studies haveexamined the particular mechanics of zooplankton feeding by coral polyps in unidirectional flow-ing water (e.g., Sebens & Johnson 1991; Sebens et al. 1997, 1998). Although zooplankton feedingrates (often referred to as capture efficiency rates) vary with coral species, coral colony morphol-ogy, and type of zooplankton prey, most studies have shown that capture rates are dependent onflow speed. In many cases, these particle capture rates initially increase with higher flow as a resultof higher particle encounter rates but may eventually decrease at higher flow speeds when tentaclesare no longer able to fully extend against the strong flow (Sebens et al. 1997). Although particlecapture rates should also depend on the magnitude of a wave-driven oscillatory flow, there is a lackof similar detailed studies under wave-dominated flow that are necessary to accurately quantifythese rates. Finally, an important fraction of suspended organic matter crossing through reef sys-tems is remineralized in permeable reef sediments, whose pore-water chemistry and reaction ratesare also driven as much by hydrodynamics (i.e., wave and tidal pumping) as by infaunal metabolism(Cyronak et al. 2013, Falter & Sansone 2000, Huettel & Rusch 2000, Santos et al. 2010).
3. COMMUNITY TO REEF SYSTEM SCALES (10 m–10 km)
3.1. Reef Circulation
One of the most direct mechanisms by which physical ocean processes can influence the biologyand chemistry of coral reefs is through the circulation and residence time of reef waters (Figure 2).For many reefs across the Indo-Pacific and Caribbean, circulation is driven mainly by breakingsurface waves (Hearn 1999, Hench et al. 2008, Lowe et al. 2009b, Munk & Sargent 1948,Symonds et al. 1995). As open-ocean waves approach a reef, they break when the wave heightbecomes a significant fraction of the water depth (Nelson 1994), although the exact break point is
Figure 2Wave-driven circulation of a coastal reef-lagoon system. Waves approach the reef, shoal, and break near the reef crest. This generates arise in the dynamic sea level (η) relative to the offshore mean sea level (msl), referred to as wave setup, as well as an associated shorewardflow of water (�qr) across the shallow reef. The flow across the reef is balanced by the seaward flow of water (�qc) through narrowchannels that perforate the reef structure.
www.annualreviews.org • Oceanic Forcing of Coral Reefs 51
ηcrη(x)
msl
qc
qrqr
Ann
u. R
ev. M
arin
e. S
ci. 2
015.
7:43
-66.
Dow
nloa
ded
from
ww
w.a
nnua
lrev
iew
s.or
g A
cces
s pr
ovid
ed b
y U
nive
rsity
of
Wes
tern
Aus
tral
ia o
n 01
/30/
15. F
or p
erso
nal u
se o
nly.

MA07CH03-Lowe ARI 29 October 2014 6:57
Wave setup: a localelevation in mean sealevel that arises tobalance wave radiationstress gradients
further affected by the nonlinearity of the waves and local bottom steepness (Buckley et al. 2014,Massel & Gourlay 2000). The loss of wave energy via breaking through the surf zone leads to asubsequent reduction in the radiation stress, or the excess momentum flux carried by the incidentwaves (Longuet-Higgins & Stewart 1962). The resulting wave force generated by the radiationstress gradient through the surf zone is opposed in part by an increase, or setup, of mean sea levelthrough the surf zone that raises the water level over the reef.
Shoreward of the surf zone, where wave dissipation becomes comparatively small, the dominantmomentum balance instead tends to be between the wave setup gradient (resulting from a decreasein mean water level across the reef ) and the opposing bottom stress associated with the mean flowof water driven across the reef (Gourlay & Colleter 2005, Lowe et al. 2009b, Symonds et al. 1995).Although many one-dimensional analytical models have been developed to predict wave setup andwave-driven flows on reefs, Gourlay & Colleter (2005) showed that the amount of wave setup atthe reef crest ηcr tends to be roughly proportional to the onshore deepwater wave energy flux and,therefore, proportional to the offshore wave height H0 and period τw according to ηcr ∝ H 2
0 τw.Assuming a quadratic dependency of bottom drag on flow speed with drag coefficient CD (seeSection 2), this implies that the cross-reef transport q is proportional to the square root of theincident wave energy flux, or
q ≡∫ η
−hU (z)dz ≡ U h ∝ H 0
√τw√
CD, (5)
where h is the local water depth, η is the local free-surface height relative to the mean offshorewater level, and U is the depth-averaged flow speed. Therefore, the strength of the wave-drivencirculation tends to increase roughly linearly with the offshore wave height but also depends moreweakly on the wave period and the bottom-drag coefficient. Observations tend to generally supportthe scaling of q in Equation 5 (e.g., Lowe et al. 2009b, Monismith et al. 2013), although the exactmagnitude of q that is generated in response to incident wave forcing can also vary substantiallyamong reef systems owing to the particular morphology of a reef-lagoon system and the degreeto which its channels restrict the return flow of water back to the surrounding ocean (Lowe et al.2010, Monismith 2013, Taebi et al. 2011).
Tides can also play a secondary role influencing wave-driven circulation on wave-exposedreef systems by modulating how and where waves break (Becker et al. 2014, Taebi et al. 2011).Changes in tidal elevation can alter the position and width of the surf zone and therefore the rateat which wave energy is dissipated, which in turn influences the magnitude of the wave setup. Theresistance experienced by cross-reef currents can also be substantially enhanced at lower phases ofthe tide when flow becomes depth limited, significantly increasing the bottom-drag coefficients(see Section 2) (McDonald et al. 2006, Pomeroy et al. 2012, Rosman & Hench 2011). Tidal forcingmay play a more direct role in driving reef circulation when the tidal range is large (e.g., >3 m)and local tidal water level gradients can exceed those induced by wave setup fields.
In addition, tides can play a more direct role in driving circulation in larger and more enclosedlagoons where the channels connecting the lagoon with the open ocean are relatively narrow. Theconstricted exchange of water between these lagoons and the open ocean can cause significantphase lags between a lagoon and offshore water levels (e.g., Dumas et al. 2012). Historically, mostreef studies have found that wind stresses often play only a minor role in driving the circulationof shallow reefs; however, wind forcing can be important or even dominant in the circulation ofdeeper and more isolated lagoons (Atkinson et al. 1981, Delesalle & Sournia 1992, Douillet et al.2001, Lowe et al. 2009a). Unfortunately, the extent to which buoyancy forcing helps drive reefcirculation through either temperature- or salinity-driven stratification has received only limited
52 Lowe · Falter
Ann
u. R
ev. M
arin
e. S
ci. 2
015.
7:43
-66.
Dow
nloa
ded
from
ww
w.a
nnua
lrev
iew
s.or
g A
cces
s pr
ovid
ed b
y U
nive
rsity
of
Wes
tern
Aus
tral
ia o
n 01
/30/
15. F
or p
erso
nal u
se o
nly.

MA07CH03-Lowe ARI 29 October 2014 6:57
Eulerian: examining adynamic flow fieldfrom the perspective ofa fixed frame ofreference
Lagrangian:examining a dynamicflow field from theperspective of a waterparcel moving with theflow
study; however, this forcing may also be important in certain reef systems (Hoeke et al. 2013,Monismith et al. 2006).
3.2. Biogeochemical Transformation of Reef Waters
The degree to which any local process occurring within a reef (physical, chemical, or biological)can affect the spatial and temporal variation in water quality is dependent as much on the strengthand pattern of circulation as on the process itself. These relationships can be understood eitherfrom an Eulerian perspective, in which circulation and changing water quality are consideredwithin a fixed reference frame, or from a Lagrangian perspective, in which changes in water qual-ity are observed within a defined water parcel flowing through a reef system. From an Eulerianperspective, these relationships are best illustrated by the depth-integrated nonconservative trans-port equation, which shows the mutual dependence of spatial (∂/∂x,∂/∂y) and temporal (∂/∂t)changes in water quality on both the transport q and the net flux of heat or mass into the watercolumn J (Falter et al. 2008):
Jflux
= h∂C∂t
local
+ q · ∇Cadvection
− K∇2Cdispersion
, (6)
where C is some nonconservative water quality parameter of interest (nutrient concentration,alkalinity, temperature, etc.) and K is a horizontal dispersion coefficient. J can represent the sum ofall biogeochemical processes responsible for the uptake or release of nutrients, particles, dissolvedinorganic carbon, alkalinity, etc. It can also represent the net exchange of heat, moisture, andgasses across the air-sea interface. Limited field measurements have shown that mass transportresulting from advection is much greater than that resulting from horizontal dispersion (i.e.,q·∇C � K∇2C) on shallow, wave-driven reefs (Falter et al. 2008); however, horizontal dispersioncould be more important in reefs with strongly sheared two-dimensional circulation patterns.Furthermore, a fully three-dimensional nonconservative transport equation may be necessary tobudget the uptake, release, and transport of mass in deeper and more stratified sections of a reef(Genin et al. 2009, Monismith et al. 2010, Teneva et al. 2013).
The application of the nonconservative transport equation to model changes in water qualityhas proven robust for inversely calculating benthic fluxes in shallow reef environments for morethan half a century (e.g., Odum & Odum 1955, Smith 1973), albeit through the use of simplifiedversions of Equation 6. Earlier methodologies discriminated between flow respirometry, whichmeasures changes in water chemistry along one axis (generally oriented in the cross-reef direction)and assumes that J ≈ qx∂C/∂x (Barnes & Devereux 1984, Gattuso et al. 1993, Kraines et al. 1997),and the slack-water method, which considers only the time-dependent changes in water chemistryat a specific site and assumes that J ≈ h∂C/∂t (Kinsey 1978, McMahon et al. 2013, Shaw et al.2012). Both approaches constitute selective simplifications of the same Eulerian nonconservativetransport equation shown in Equation 6. The simplification of upstream-downstream applicationsof flow respirometry tends to be most robust on shallow reef flats where the flow is both reason-ably steady and strong (>5 cm s−1) and chemical gradients vary only weakly in time such thatqx∂C/∂x � h∂C/∂t. The slack-water simplification generally requires flow speeds to be on theorder of ∼1 cm s−1 or less so that the time-dependent term dominates over the advective terms(i.e., h∂C/∂t � q ·∇C). This latter condition is particularly difficult to meet for two reasons. First,it requires that the predominant current is very weak yet still strong enough to provide the nec-essary turbulent stirring to vertically mix the water column, a criterion that can easily be violated(Kinsey 1978). Second, it requires that the horizontal gradients in water quality also be relativelysmall, a criterion that is difficult to meet for many reefs given that the spatial distributions of
www.annualreviews.org • Oceanic Forcing of Coral Reefs 53
Ann
u. R
ev. M
arin
e. S
ci. 2
015.
7:43
-66.
Dow
nloa
ded
from
ww
w.a
nnua
lrev
iew
s.or
g A
cces
s pr
ovid
ed b
y U
nive
rsity
of
Wes
tern
Aus
tral
ia o
n 01
/30/
15. F
or p
erso
nal u
se o
nly.
MA07CH03-Lowe ARI 29 October 2014 6:57
Water age: the age ofa water parcel relativeto when it entered adomain (thecomplement ofresidence time)
biomass and metabolism are known to vary by an order of magnitude or more (Andrefouet et al.2001, Atkinson & Falter 2003, Kinsey 1985, Tanaka et al. 2011).
One additional process contributing to transport in reef systems is Stokes drift, or the excessmass transport induced by the nonlinearity of waves (Longuet-Higgins & Stewart 1962). Stokesdrift is a depth-dependent, Lagrangian property of wave motion and thus cannot be directlymeasured by time-averaging fixed-point current meter data; however, it can be estimated fromlocal wave measurements at a specific reef site. Stokes drift within the coastal zone can range fromseveral centimeters per second to tens of centimeters per second depending on the water depth,wave height, and wave period (Monismith & Fong 2004). This can cause the total transport to befar greater than that measured by fixed current meters alone, thus potentially distorting perceivedrates of material exchange (Falter et al. 2008). Which terms within the general transport equationabove (Equation 6) can or cannot be ignored, and whether Stokes drift should be considered, mustbe carefully assessed for each reef environment on a case-by-case basis.
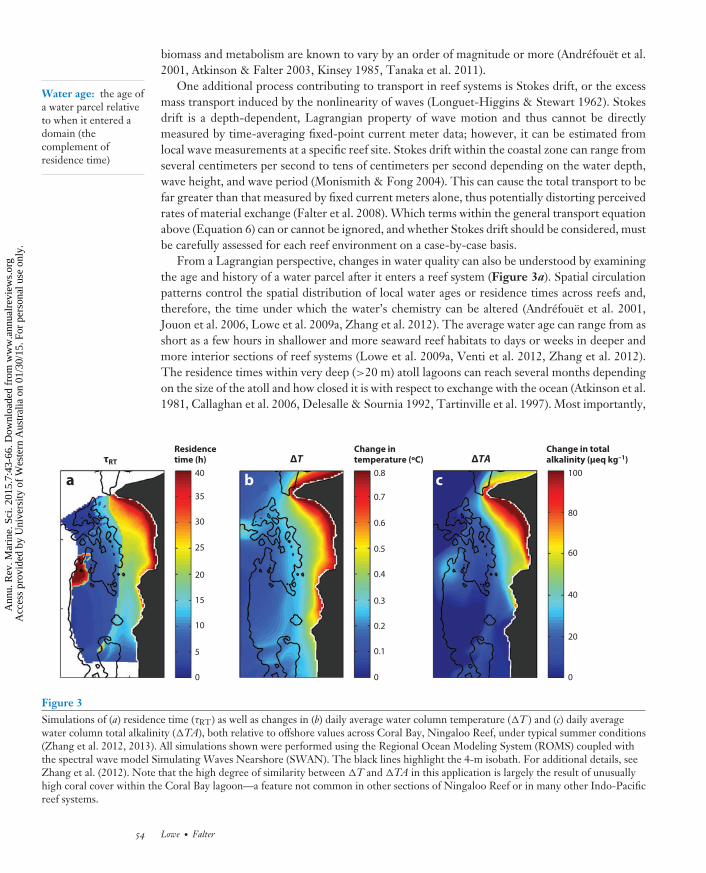
From a Lagrangian perspective, changes in water quality can also be understood by examiningthe age and history of a water parcel after it enters a reef system (Figure 3a). Spatial circulationpatterns control the spatial distribution of local water ages or residence times across reefs and,therefore, the time under which the water’s chemistry can be altered (Andrefouet et al. 2001,Jouon et al. 2006, Lowe et al. 2009a, Zhang et al. 2012). The average water age can range from asshort as a few hours in shallower and more seaward reef habitats to days or weeks in deeper andmore interior sections of reef systems (Lowe et al. 2009a, Venti et al. 2012, Zhang et al. 2012).The residence times within very deep (>20 m) atoll lagoons can reach several months dependingon the size of the atoll and how closed it is with respect to exchange with the ocean (Atkinson et al.1981, Callaghan et al. 2006, Delesalle & Sournia 1992, Tartinville et al. 1997). Most importantly,
τRT ΔT ΔTAResidencetime (h)
Change intemperature (ºC)
Change in totalalkalinity (μeq kg–1)
a b c40
35
30
25
20
15
10
5
0
0.8
0.7
0.6
0.5
0.4
0.3
0.2
0.1
0
100
0
80
40
20
60
Figure 3Simulations of (a) residence time (τRT) as well as changes in (b) daily average water column temperature (�T ) and (c) daily averagewater column total alkalinity (�TA), both relative to offshore values across Coral Bay, Ningaloo Reef, under typical summer conditions(Zhang et al. 2012, 2013). All simulations shown were performed using the Regional Ocean Modeling System (ROMS) coupled withthe spectral wave model Simulating Waves Nearshore (SWAN). The black lines highlight the 4-m isobath. For additional details, seeZhang et al. (2012). Note that the high degree of similarity between �T and �TA in this application is largely the result of unusuallyhigh coral cover within the Coral Bay lagoon—a feature not common in other sections of Ningaloo Reef or in many other Indo-Pacificreef systems.
54 Lowe · Falter
Ann
u. R
ev. M
arin
e. S
ci. 2
015.
7:43
-66.
Dow
nloa
ded
from
ww
w.a
nnua
lrev
iew
s.or
g A
cces
s pr
ovid
ed b
y U
nive
rsity
of
Wes
tern
Aus
tral
ia o
n 01
/30/
15. F
or p
erso
nal u
se o
nly.

MA07CH03-Lowe ARI 29 October 2014 6:57
Albedo: the fractionof solar radiation that asurface or objectreflects
residence times at any reef site can never be considered a static quantity because they are drivenby ever-changing wind, wave, and tide conditions.
Changes to the properties of a flowing water mass depend not only on the simple age of a waterparcel but also on the specific trajectory or streamline that determines the various reef habitatsthe parcel is exposed to. Without explicit knowledge of the flow path that a given water masstook prior to the measurement of its changed chemistry relative to some reference point (typicallyoffshore waters), it is not possible to know which sections or areas of reef habitat to assign theseobserved changes to. This is particularly problematic when attempting to calculate benthic fluxesand reaction rates for entire reef systems using bulk estimates of water residence time and localmeasurements of water chemistry made at specific reef sites (Falter et al. 2013, Venti et al. 2012,Zhang et al. 2012). Nonetheless, changes in the chemistry of offshore waters are generally mostrapid over the most seaward several hundred meters of shallow reef flat. These changes can be welldescribed by a simple analytical equation that depends on offshore wave forcing, reef morphology,and the net benthic and surface mass fluxes (Falter et al. 2013):
�C ∝ JH 0
√τw
(L2
r
hr
)3/4
, (7)
where Lr and hr are the length and depth of the reef flat, respectively, and advection is assumed tobe the dominant transport term (q · ∇C � ∂C/∂t).
3.3. Temperature Variations in Reef Waters
The dynamics driving spatial and temporal changes in water temperature across reefs(Figure 3b) are analogous to those driving changes in water chemistry (Figure 3c) because theyare both governed by the same fundamental nonconservative transport equation (Equation 6)(Davis et al. 2011, Falter et al. 2014, McCabe et al. 2010, Zhang et al. 2013). The additional heatgained by reef waters from atmospheric exchange can have profound effects on local temperatureanomalies and cumulative thermal stresses on reefs (Davis et al. 2011, Falter et al. 2014, McCabeet al. 2010, Pineda et al. 2013, Zhang et al. 2013). In contrast to rates of benthic reef metabolism,which can vary by more than an order of magnitude (Andersson & Gledhill 2013, Kinsey 1985),net heat fluxes are far less spatially variable across reefs. First, spatial variations in atmosphericconditions (e.g., air temperatures, humidity, and wind speeds) that drive net surface and bottomheat fluxes (J ) are limited over the scale of an individual reef (McGowan et al. 2010, Welleret al. 2008). Second, the capacities of different reef communities to absorb shortwave radiation(dependent on their albedo) generally vary by less than a factor of 2 (Hochberg et al. 2003).Third, a substantial fraction of shortwave radiation is absorbed by the water column itself, andthis proportion increases with water depth. The end result is that spatial variation in bottomalbedo resulting from a changing benthic reef habitat usually has only a secondary effect on thenet heating or cooling of reef waters, even for very shallow reef systems (Zhang et al. 2013).
4. REGIONAL TO GLOBAL SCALES (>10 km)
4.1. Global Distribution of Wave and Tidal Forcing
With only a few exceptions, studies have found that the circulation of reefs is dominantly forcedby waves, or in some cases by tides (Andrefouet et al. 2006, Falter et al. 2013, Monismith 2007).Although the three-dimensional circulation of reef slopes and lagoons can be strongly influenced bylocal wind forcing (Atkinson et al. 1981, Wolanski & Thomson 1984) as well as by buoyancy forcingfrom thermal gradients (Monismith et al. 2006) or freshwater discharge (Hoeke et al. 2013), these
www.annualreviews.org • Oceanic Forcing of Coral Reefs 55
Ann
u. R
ev. M
arin
e. S
ci. 2
015.
7:43
-66.
Dow
nloa
ded
from
ww
w.a
nnua
lrev
iew
s.or
g A
cces
s pr
ovid
ed b
y U
nive
rsity
of
Wes
tern
Aus
tral
ia o
n 01
/30/
15. F
or p
erso
nal u
se o
nly.

MA07CH03-Lowe ARI 29 October 2014 6:57
Longitude
La
titu
de
a
b
c
e
d
f
Hs (m)
MTR (m)
MTR/Hs
MTR/Hs
MTR (m)
Hs (m)
Fre
qu
en
cy (
%)
20°N
0°
20
1020°S
20°N
0°
20°S
20°N
0°
20°S
0° 60°E 120E° 180° 120°W 60°W 0°
0° 60°E 120E° 180° 120°W 60°W 0°
0° 60°E 120E° 180° 120°W 60°W 0°
3
2
1
0
3
2
1
0
3
2
1
0
0
20
10
0
20
10
0
0 1 2 3
0 1 2 3
0 1 2 3
Figure 4Global spatial distributions and frequency distributions of (a,b) annual mean significant wave height (Hs), (c,d ) mean tidal range (MTR),and (e,f ) relative tidal range (MTR/Hs) for warm-water coral reef systems. Global coral reef distributions are from the 1-km-resolutiondata set from UNEP-WCMC (2003) and were joined with global wave height and tide distributions. Annual significant wave heightswere derived using data from Young et al. (2011). Mean tidal ranges were computed from tidal harmonics derived from the OregonState University Tidal Inversion Software (OTIS) (Egbert & Erofeeva 2002). Note that the values in all panels are capped at amaximum of 3, so some reefs may experience larger mean values.
Macrotidal reefsystem: a reef systemwhere the tidal rangeexceeds 3 m
effects tend to be dominant only on reefs protected from ocean swell or within the relatively deeplagoons of barrier reefs and atolls. There are strong regional differences in the global distribution ofsignificant wave heights Hs within the ± 30◦ latitude band where warm-water coral reefs are found(Figure 4). Reefs in the Pacific and Indian Oceans tend to be dominantly wave forced (annual meanHs > 1.5 m), whereas reefs in the Caribbean, Southeast Asia, and northern parts of Australia tendto experience only weak to moderate wave forcing (typical annual mean Hs < 1 m). These patternslikely drive substantial regional-scale differences in the wave-driven flows and residence timeswithin reefs, depending, of course, on the morphologies of individual reefs within each region (seeSection 3).
Despite the dominance of wave-driven reef systems globally, there are still numerous reefsystems worldwide that can be considered tide dominated, defined here simply as locations wherethe ratio of the mean tidal range (MTR) to the annual mean Hs is greater than 1 (Figure 4).Such systems have for the most part remained neglected in the literature; they include someextreme examples of macrotidal reef systems (MTR > 3 m) in regions off northern Australia, offeast Africa, and off Central and South America (Figure 4). For example, in the Kimberley regionof northwestern Australia, where numerous reefs are subject to tidal ranges greater than 10 m,water frequently drains off reef terraces as “waterfalls” during ebb phases of the tide (Figure 5).The dynamics of these tide-dominated reef systems, where the strongly varying flows lead to acritical flow point at the reef terrace that restricts flow, are fundamentally different from those oftraditional wave-driven coral reef flows, in which flow resistance is due solely to bottom drag. A
56 Lowe · Falter
Ann
u. R
ev. M
arin
e. S
ci. 2
015.
7:43
-66.
Dow
nloa
ded
from
ww
w.a
nnua
lrev
iew
s.or
g A
cces
s pr
ovid
ed b
y U
nive
rsity
of
Wes
tern
Aus
tral
ia o
n 01
/30/
15. F
or p
erso
nal u
se o
nly.

MA07CH03-Lowe ARI 29 October 2014 6:57
Figure 5(a) A macrotidal reef system in the Kimberley region of northwestern Australia, where the spring tidal range reaches up to 10 m(photograph by R.J. Lowe). The photograph shows the reef at low tide with water draining off the coralline algal reef platform as a“waterfall.” (b) A schematic of water draining off an emergent macrotidal reef island at low tide.
much-improved understanding of tide-dominated reef systems is necessary to ultimately developappropriate predictive models of these very distinct coastal flows.
Whereas tidal forcing is predictable and quasi-stationary over long timescales (i.e., governed byset tidal harmonics), wave forcing on reefs is often episodic and subject to extreme events. Extremewave events generated by tropical cyclones may be rare and short lived; however, the large waveheights and storm surges generated can have a profound and long-lasting impact on coral reefecosystems. In addition, large extratropical cyclones in the major ocean basins may also transmitlarge swell across the ocean to remote reef systems thousands of kilometers away—as occurs, forexample, when large waves generated by remote storms impart substantial wave energy to Pacificisland reefs (Hench et al. 2008). Overall, it appears that, for many reefs, these extreme wave events
www.annualreviews.org • Oceanic Forcing of Coral Reefs 57
a
b
Ann
u. R
ev. M
arin
e. S
ci. 2
015.
7:43
-66.
Dow
nloa
ded
from
ww
w.a
nnua
lrev
iew
s.or
g A
cces
s pr
ovid
ed b
y U
nive
rsity
of
Wes
tern
Aus
tral
ia o
n 01
/30/
15. F
or p
erso
nal u
se o
nly.

MA07CH03-Lowe ARI 29 October 2014 6:57
play a primary role in shaping the reef community composition and zonation by establishing themechanical limits of various reef organisms (Dollar & Tribble 1993, Grigg 1998, Lugo-Fernandez& Gravois 2010, Madin & Connolly 2006). Other investigators have also argued that long-termtrends in the frequency of extreme wave events can drive associated declines in coral coverageof reefs. For example, De’ath et al. (2012) argued that one major cause for the decline in coralcoverage across the Great Barrier Reef over the past ∼30 years was the number of unusually strongtropical cyclones.
4.2. Offshore Ocean Interactions with Reefs
Offshore ocean dynamics are gaining increasing recognition for the critical role they play indriving many reef ecosystem processes (Freeman et al. 2012, Gove et al. 2013, Wyatt et al. 2012).Nevertheless, our quantitative understanding of how reefs are connected to the surrounding oceanremains poor owing to both the complex nature of these interactions and the lack of comprehensivestudies that simultaneously consider reef ecosystem dynamics over a broad range of relevant scales(from regional down to community scales). At the broadest scale—that of entire ocean basins—thepresence of major ocean boundary currents has long been known to regulate the distribution ofmany tropical coral reef systems globally. The general lack of major coral reef systems on thecontinental shelves of eastern ocean basins has been attributed largely to persistent upwelling-favorable conditions generated by wind-driven, equatorward-flowing eastern boundary currentsthat can inhibit the formation of oligotrophic tropical reefs (Birkeland 1997). In fact, many reefsacross the globe depend on regional ocean currents to provide thermal conditions favorable forreef growth. Some good examples are the high-latitude coral reefs found between 28◦S and 32◦Soff the coasts of eastern and western Australia (Lord Howe Island and the Houtman-AbrolhosIslands, respectively); these reef systems exist much farther south than those in other parts of theSouthern Hemisphere owing to the poleward transport of heat by the East Australian Currentand the Leeuwin Current. Nevertheless, predictions of how coral reef systems respond to globalclimate variability have tended to neglect how subtle changes in regional ocean circulation patternsmay locally affect reefs in the future, instead tending to assume that reefs will be exposed only toocean-basin-scale changes in temperature and carbonate chemistry (e.g., Hoegh-Guldberg et al.2007, Silverman et al. 2009).
A growing body of literature has described how a wide range of transient hydrodynamic pro-cesses on the shelf or slopes offshore of reefs influence shallow inshore reef systems. Multiplestudies have identified coastal upwelling in particular as an important mechanism driving thecross-shelf exchange of nutrients between the ocean and many reefs; these studies include sev-eral that have examined how wind- or current-driven upwelling contributes substantial fluxes ofsubsurface nutrients to the Great Barrier Reef (Andrews & Gentien 1982, Brinkman et al. 2002,Wolanski et al. 1988) and to Ningaloo Reef off western Australia (Wyatt et al. 2010, 2012) as wellas through the secondary flows generated by island wakes in the Pacific (Gove et al. 2006). Studieshave also demonstrated that coastal upwelling significantly cools the coastal waters surroundingsome coral reefs, suggesting that these regions could provide some refuge from increased oceanwarming (e.g., Chollett & Mumby 2013, Karnauskas & Cohen 2012, Riegl & Piller 2003, VanHooidonk et al. 2013). Indeed, frequent, episodic upwelling can locally reduce water temperaturesby up to several degrees in reef systems ranging from northwestern Australia (Lowe et al. 2012)to the Caribbean, the Red Sea, and the eastern coastlines of Africa (Riegl & Piller 2003).
Higher-frequency internal waves are similarly well known to drive substantial cross-shelf fluxesof nutrients, plankton, and heat both to and from many coral reefs. The most comprehensiveobservations of these dynamics have come from studies of internal waves off the Florida Keys (Davis
58 Lowe · Falter
Ann
u. R
ev. M
arin
e. S
ci. 2
015.
7:43
-66.
Dow
nloa
ded
from
ww
w.a
nnua
lrev
iew
s.or
g A
cces
s pr
ovid
ed b
y U
nive
rsity
of
Wes
tern
Aus
tral
ia o
n 01
/30/
15. F
or p
erso
nal u
se o
nly.

MA07CH03-Lowe ARI 29 October 2014 6:57
et al. 2008; Leichter et al. 1996, 2003) and in French Polynesia (Leichter et al. 2012, Wolanski &Delesalle 1995). Despite the clear importance of internal waves to cross-shelf exchange in theseregions, their specific influence on the nutrient and heat budgets of reefs in other parts of theworld remains poorly known. Ultimately, our understanding of how shallow reef systems respondto the larger-scale dynamics of the surrounding ocean (including those driven by coastal upwelling,internal waves, coastal eddy fields, and episodic extreme events such as tropical cyclone mixing)remains primitive.
SUMMARY POINTS
1. The oceans influence the structure and function of coral reefs through a variety of mech-anisms, ranging from small-scale boundary-layer dynamics occurring on submillimeterscales to regional circulation patterns spanning hundreds of kilometers.
2. Mass and momentum exchange within and above coral reef canopies can differ substan-tially depending on the prevailing flow regime. Under unconfined, unidirectional flow,flow within the canopy is driven primarily by a shear layer at the top of the canopy and isrelatively weak. Under severely depth-limited flow or wave-driven oscillatory flow, flowwithin the canopy is driven primarily by pressure gradients and is much stronger.
3. Rates of dissolved nutrient and particle uptake by reef communities are heavily influencedby the predominant boundary-layer physics; however, mechanisms of particle trappingand feeding are highly varied, and their physics are still not well understood.
4. Circulation across most shallow reef systems is wave driven, with the tides acting as asecondary variable modulating wave-driven flows. However, in some regions the directforcing of reef circulation by tides is much stronger than by waves.
5. Spatial and temporal changes in reef water temperature and chemistry are intrinsicallylinked through a balance between the net fluxes of heat or mass that drive their noncon-servative behavior and the circulation of reef waters. The circulation, residence times,and ages of reef waters are, in turn, dependent on the hydrodynamic forces driving theflow as well as the particular morphology of a given reef system.
6. Episodic upwelling and internal waves can provide short-term (days) and intermediate-scale (kilometers) mechanisms through which coral reefs are exposed to cooler and morenutrient- and/or plankton-rich waters. Longer-term changes in regional circulation pat-terns (weeks to months) can either provide conditions favorable for reef growth or helpfoment large-scale thermal stress events (tens to hundreds of kilometers).
FUTURE ISSUES
1. Predicting flow and drag forces within the complex and varied morphologies of reefcanopies remains a major challenge. New experimental data sets and modeling approachesare needed to further advance predictions of reef canopy flows under both unidirectionaland wave-driven conditions.
www.annualreviews.org • Oceanic Forcing of Coral Reefs 59
Ann
u. R
ev. M
arin
e. S
ci. 2
015.
7:43
-66.
Dow
nloa
ded
from
ww
w.a
nnua
lrev
iew
s.or
g A
cces
s pr
ovid
ed b
y U
nive
rsity
of
Wes
tern
Aus
tral
ia o
n 01
/30/
15. F
or p
erso
nal u
se o
nly.

MA07CH03-Lowe ARI 29 October 2014 6:57
2. Likewise, existing nearshore models neglect how bottom drag under waves and currents isinfluenced by in-canopy momentum dynamics. Improving the parameterizations of thesecanopy-scale dynamics, including coupling them within wave and circulation models,would no doubt substantially improve hydrodynamic models in reef applications.
3. Although existing mass-transfer formulations provide some ability to predict rates ofdissolved nutrient uptake by reef communities, there are still major gaps in our knowledgeof nutrient mass transfer under wave-driven oscillatory flow.
4. Some empirical data on particle uptake rates by reef communities are available, butmechanistic approaches for modeling these dynamics are virtually nonexistent. The de-pendency of uptake rates on the mechanism of particle capture, as well as on the size andcomposition of the particles themselves, makes this a particularly challenging researchendeavor.
5. Little is known about circulation around reefs with relatively large tidal ranges (mean tidalrange of >3 m) and how this affects the spatial and temporal variation in reef metabolismand water quality. Such systems should be given greater attention.
6. Multiple studies over the past two decades have documented the short-term (days) impactof coastal upwelling and internal waves on the temperature, chemistry, and planktoncommunities of reef waters. However, our understanding of how important these episodicevents are to the growth and structure of reef communities over the long term (monthsto years) is very limited, as is our understanding of how they are mechanistically linkedto climate-driven changes in regional ocean dynamics.
DISCLOSURE STATEMENT
The authors are not aware of any affiliations, memberships, funding, or financial holdings thatmight be perceived as affecting the objectivity of this review.
ACKNOWLEDGMENTS
We dedicate this review to Marlin Atkinson (1951–2013), who will be remembered as a pioneerin the study of ocean forcing of reef ecosystems and who helped inspire our own interest inthis subject more than a decade ago. Through the years, we have learned much about coralreefs through collaboration with colleagues and friends in the research community, especiallyStephen Monismith, Jeff Koseff, Eric Hochberg, Graham Symonds, Greg Ivey, Steve Smith,Frank Sansone, Mark Merrifield, Carlos Coimbra, Pascale Cuet, Alex Wyatt, Zhenlin Zhang,Malcolm McCulloch, Richard Brinkman, Chari Pattiaratchi, Dan Schar, Jerome Aucan, ChristinePequignet, Uri Shavit, Geno Pawlak, Serge Andrefouet, Frazer McGregor, Andrew Pomeroy,Mark Buckley, Ap van Dongeren, Dano Roelvink, Stuart Humphries, Ben Radford, Nicole Jones,Curt Storlazzi, Renee Gruber, Mike Cuttler, Gundula Winter, Edwin Drost, Soheila Taebi, CecileRousseaux, Jim Hench, Johanna Rosman, Sam Kahng, Matt Reidenbach, and Sandy Chang. Wealso thank Stefan Zieger and Ian Young for providing the mean global wave height data used inFigure 3. Support for this work was provided by the Australian Research Council (ARC) Centreof Excellence for Coral Reef Studies (CE140100020). R.J.L. is also grateful for support fromARC under both the Future Fellowship (FT110100201) and Discovery Projects (DP140102026)
60 Lowe · Falter
Ann
u. R
ev. M
arin
e. S
ci. 2
015.
7:43
-66.
Dow
nloa
ded
from
ww
w.a
nnua
lrev
iew
s.or
g A
cces
s pr
ovid
ed b
y U
nive
rsity
of
Wes
tern
Aus
tral
ia o
n 01
/30/
15. F
or p
erso
nal u
se o
nly.

MA07CH03-Lowe ARI 29 October 2014 6:57
schemes. This work was also partially supported by a grant from the Western Australia MarineScience Institution (WAMSI) under the Kimberley Marine Research Program.
LITERATURE CITED
Andersson AJ, Gledhill D. 2013. Ocean acidification and coral reefs: effects on breakdown, dissolution, andnet ecosystem calcification. Annu. Rev. Mar. Sci. 5:321–48
Andrefouet S, Ouillon S, Brinkman R, Falter JL, Douillet P, et al. 2006. Review of solutions for 3D hydrody-namic modeling applied to aquaculture in South Pacific atoll lagoons. Mar. Pollut. Bull. 52:1138–55
Andrefouet S, Pages J, Tartinville B. 2001. Water renewal time for classification of atoll lagoons in the TuamotuArchipelago (French Polynesia). Coral Reefs 20:399–408
Andrews JC, Gentien P. 1982. Upwelling as a source of nutrients for the Great Barrier Reef ecosystems: asolution to Darwin’s question? Mar. Ecol. Prog. Ser. 8:257–69
Atkinson MJ, Bilger RW. 1992. Effect of water velocity on phosphate uptake in coral reef-flat communities.Limnol. Oceanogr. 37:273–79
Atkinson MJ, Falter JL. 2003. Coral reefs. In Biogeochemistry of Marine Systems, ed. KP Black, GB Shimmield,pp. 40–64. Boca Raton, FL: CRC
Atkinson MJ, Smith S, Stroup E. 1981. Circulation in Enewetak atoll lagoon. Limnol. Oceanogr. 26:1074–83Ayukai T. 1995. Retention of phytoplankton and planktonic microbes on coral reefs within the Great Barrier
Reef, Australia. Coral Reefs 14:141–47Baird ME, Atkinson MJ. 1997. Measurement and prediction of mass transfer to experimental coral reef
communities. Limnol. Oceanogr. 42:1685–93Barnes D, Devereux M. 1984. Productivity and calcification on a coral reef: a survey using pH and oxygen
electrode techniques. J. Exp. Mar. Biol. Ecol. 79:213–31Becker J, Merrifield M, Ford M. 2014. Water level effects on breaking wave setup for Pacific Island fringing
reefs. J. Geophys. Res. 119:914–32Bilger RW, Atkinson MJ. 1992. Anomalous mass transfer of phosphate on coral reef flats. Limnol. Oceanogr.
37:261–72Birkeland C, ed. 1997. Life and Death of Coral Reefs. New York: SpringerBrinkman R, Wolanski E, Deleersnijder E, McAllister F, Skirving W. 2002. Oceanic inflow from the Coral
Sea into the Great Barrier Reef. Estuar. Coast. Shelf Sci. 54:655–68Bruno JF, Siddon CE, Witman JD, Colin PL, Toscano MA. 2001. El Nino related coral bleaching in Palau,
Western Caroline Islands. Coral Reefs 20:127–36Buckley M, Lowe RJ, Hansen J. 2014. Evaluation of nearshore wave models in steep reef environments. Ocean
Dyn. 64:847–62Callaghan DP, Nielsen P, Cartwright N, Gourlay MR, Baldock TE. 2006. Atoll lagoon flushing forced by
waves. Coast. Eng. 53:691–704Chang S, Elkins C, Alley M, Eaton J, Monismith SG. 2009. Flow inside a coral colony measured using magnetic
resonance velocimetry. Limnol. Oceanogr. 54:1819Charpy L, Blanchot J. 1999. Picophytoplankton biomass, community structure and productivity in the Great
Astrolabe Lagoon, Fiji. Coral Reefs 18:255–62Chindapol N, Kaandorp JA, Cronemberger C, Mass T, Genin A. 2013. Modelling growth and form of the
scleractinian coral Pocillopora verrucosa and the influence of hydrodynamics. PLoS Comput. Biol. 9:e1002849Chollett I, Mumby PJ. 2013. Reefs of last resort: locating and assessing thermal refugia in the wider Caribbean.
Biol. Conserv. 167:179–86Cyronak T, Santos I, McMahon A, Eyre B. 2013. Carbon cycling hysteresis in permeable carbonate sands
over a diel cycle: Implications for ocean acidification. Limnol. Oceanogr. 58:131–43Darwin C. 1842. The Structure and Distribution of Coral Reefs. London: Smith Elder & Co.Davis KA, Leichter JJ, Hench JL, Monismith SG. 2008. Effects of western boundary current dynamics on the
internal wave field of the Southeast Florida shelf. J. Geophys. Res. 113:C09010Davis KA, Lentz SJ, Pineda J, Farrar JT, Starczak VR, Churchill JH. 2011. Observations of the thermal
environment on Red Sea platform reefs: a heat budget analysis. Coral Reefs 30:25–36
www.annualreviews.org • Oceanic Forcing of Coral Reefs 61
Ann
u. R
ev. M
arin
e. S
ci. 2
015.
7:43
-66.
Dow
nloa
ded
from
ww
w.a
nnua
lrev
iew
s.or
g A
cces
s pr
ovid
ed b
y U
nive
rsity
of
Wes
tern
Aus
tral
ia o
n 01
/30/
15. F
or p
erso
nal u
se o
nly.

MA07CH03-Lowe ARI 29 October 2014 6:57
De’ath G, Fabricius KE, Sweatman H, Puotinen M. 2012. The 27-year decline of coral cover on the GreatBarrier Reef and its causes. Proc. Natl. Acad. Sci. USA 109:17995–99
Delesalle B, Sournia A. 1992. Residence time of water and phytoplankton biomass in coral-reef lagoons. Cont.Shelf Res. 12:939–49
Depczynski M, Gilmour J, Ridgway T, Barnes H, Heyward A, et al. 2013. Bleaching, coral mortality andsubsequent survivorship on a West Australian fringing reef. Coral Reefs 32:233–38
Dollar SJ, Tribble GW. 1993. Recurrent storm disturbance and recovery: a long-term study of coral commu-nities in Hawaii. Coral Reefs 12:223–33
Douillet P, Ouillon S, Cordier E. 2001. A numerical model for fine suspended sediment transport in thesouthwest lagoon of New Caledonia. Coral Reefs 20:361–72
Dumas F, Gendre RL, Thomas Y, Andrefouet S. 2012. Tidal flushing and wind driven circulation of Ahe atolllagoon (Tuamotu Archipelago, French Polynesia) from in situ observations and numerical modelling.Mar. Pollut. Bull. 65:425–40
Egbert GD, Erofeeva SY. 2002. Efficient inverse modeling of barotropic ocean tides. J. Atmos. Ocean. Technol.19:183–204
Espinosa-Gayosso A, Ghisalberti M, Ivey GN, Jones NL. 2012. Particle capture and low-Reynolds-numberflow around a circular cylinder. J. Fluid Mech. 710:362–78
Fabricius KE, Yahel G, Genin A. 1998. In situ depletion of phytoplankton by an azoothanthallae soft coral.Limnol. Oceanogr. 43:354–56
Falter JL, Atkinson MJ, Merrifield MA. 2004. Mass-transfer limitation of nutrient uptake by a wave-dominatedreef flat community. Limnol. Oceanogr. 49:1820–31
Falter JL, Lowe RJ, Atkinson MJ, Monismith SG, Schar DW. 2008. Continuous measurements of net pro-duction over a shallow reef community using a modified Eulerian approach. J. Geophys. Res. 113:C07035
Falter JL, Lowe RJ, Zhang Z, McCulloch M. 2013. Physical and biological controls on the carbonate chemistryof coral reef waters: effects of metabolism, wave forcing, sea level, and geomorphology. PLoS ONE8:e53303
Falter JL, Sansone FJ. 2000. Hydraulic control of pore water geochemistry within the oxic-suboxic zone of apermeable sediment. Limnol. Oceanogr. 45:550–57
Falter JL, Zhang ZL, Lowe RJ, McGregor F, Keesing J, McCulloch M. 2014. Assessing seasonal and spatialtemperature variability within coral reefs through in situ observations and global climate data. Limnol.Oceanogr. 59:1241–55
Ferrier-Pages C, Witting J, Tambutte E, Sebens K. 2003. Effect of natural zooplankton feeding on the tissueand skeletal growth of the scleractinian coral Stylophora pistillata. Coral Reefs 22:229–40
Freeman LA, Miller AJ, Norris RD, Smith JE. 2012. Classification of remote Pacific coral reefs by physicaloceanographic environment. J. Geophys. Res. 117:C02007
Gattuso J-P, Pichon M, Delesalle B, Frankignoulle M. 1993. Community metabolism and air-sea CO2 fluxesin a coral reef ecosystem (Moorea, French Polynesia). Mar. Ecol. Prog. Ser. 96:259–67
Genin A, Monismith SG, Reidenbach MA, Yahel G, Koseff JR. 2009. Intense benthic grazing of phytoplanktonin a coral reef. Limnol. Oceanogr. 54:938–51
Ghisalberti M. 2009. Obstructed shear flows: similarities across systems and scales. J. Fluid Mech. 641:51–61Gourlay M, Colleter G. 2005. Wave-generated flow on coral reefs: an analysis for two-dimensional horizontal
reef-tops with steep faces. Coast. Eng. 52:353–87Gove JM, Merrifield MA, Brainard RE. 2006. Temporal variability of current-driven upwelling at Jarvis Island.
J. Geophys. Res. 111:C12011Gove JM, Williams G, McManus M. 2013. Quantifying climatological ranges and anomalies for Pacific coral
reef ecosystems. PLoS ONE 8:e61974Grigg RW. 1998. Holocene coral reef accretion in Hawaii: a function of wave exposure and sea level history.
Coral Reefs 17:263–72Hamner W, Wolanski E. 1988. Hydrodynamic forcing functions and biological processes on coral reefs: a
status review. In Proceedings of the 6th International Coral Reef Symposium, Vol. 1: Plenary Addresses and StatusReview, ed. JH Choat, D Barnes, MA Borowitzka, JC Coll, PJ Davies, et al., pp. 103–13. Townsville, Aust.:Int. Coral Reef Symp. Exec. Comm.
62 Lowe · Falter
Ann
u. R
ev. M
arin
e. S
ci. 2
015.
7:43
-66.
Dow
nloa
ded
from
ww
w.a
nnua
lrev
iew
s.or
g A
cces
s pr
ovid
ed b
y U
nive
rsity
of
Wes
tern
Aus
tral
ia o
n 01
/30/
15. F
or p
erso
nal u
se o
nly.

MA07CH03-Lowe ARI 29 October 2014 6:57
Hearn CJ. 1999. Wave-breaking hydrodynamics within coral reef systems and the effect of changing relativesea level. J. Geophys. Res. 104:30007–19
Hearn CJ, Atkinson MJ, Falter JL. 2001. A physical derivation of nutrient-uptake rates in coral reefs: effectsof roughness and waves. Coral Reefs 20:347–56
Hench JL, Leichter JJ, Monismith SG. 2008. Episodic circulation and exchange in a wave-driven coral reefand lagoon system. Limnol. Oceanogr. 53:2681–94
Hochberg E, Atkinson M, Andrefouet S. 2003. Spectral reflectance of coral reef bottom-types worldwide andimplications for coral reef remote sensing. Remote Sens. Environ. 85:159–73
Hoegh-Guldberg O, Mumby PJ, Hooten AJ, Steneck RS, Greenfield P, et al. 2007. Coral reefs under rapidclimate change and ocean acidification. Science 318:1737–42
Hoeke RK, Storlazzi CD, Ridd PV. 2013. Drivers of circulation in a fringing coral reef embayment: a wave-flowcoupled numerical modeling study of Hanalei Bay, Hawaii. Cont. Shelf Res. 58:79–95
Houlbreque F, Ferrier-Pages C. 2009. Heterotrophy in tropical scleractinian corals. Biol. Rev. Camb. Philos.Soc. 84:1–17
Houlbreque F, Tambutte E, Allemand D, Ferrier-Pages C. 2004. Interactions between zooplankton feeding,photosynthesis and skeletal growth in the scleractinian coral Stylophora pistillata. J. Exp. Biol. 207:1461–69
Huettel M, Rusch A. 2000. Transport and degradation of phytoplankton in permeable sediment. Limnol.Oceanogr. 45:534–49
Hughes AD, Grottoli AG. 2013. Heterotrophic compensation: a possible mechanism for resilience of coralreefs to global warming or a sign of prolonged stress? PLoS ONE 8:e81172
Humphries S. 2009. Filter feeders and plankton increase particle encounter rates through flow regime control.Proc. Natl. Acad. Sci. USA 106:7882–87
Jouon A, Douillet P, Ouillon S, Fraunie P. 2006. Calculations of hydrodynamic time parameters in a semi-opened coastal zone using a 3D hydrodynamic model. Cont. Shelf Res. 26:1395–415
Kaandorp JA, Koopman EA, Sloot PM, Bak RP, Vermeij MJ, Lampmann LE. 2003. Simulation and analysisof flow patterns around the scleractinian coral Madracis mirabilis (Duchassaing and Michelotti). Philos.Trans. R. Soc. Lond. B 358:1551–57
Karnauskas KB, Cohen AL. 2012. Equatorial refuge amid tropical warming. Nat. Clim. Change 2:530–34Kinsey DW. 1978. Productivity and calcification estimates using slack-water periods and field enclosures. In
Coral Reefs: Research Methods, ed. DR Stoddart, RE Johannes, pp. 439–68. Paris: UNESCOKinsey DW. 1985. Metabolism, calcification, and carbon production: I. Systems level studies. In Proceedings of
the Fifth International Coral Reef Congress, Vol. 4: Symposia and Seminars, ed. C Gabrie, B Salvat, pp. 505–26.Moorea, Fr. Polyn.: Antenne Mus.–EPHE
Kraines S, Suzuki Y, Omori T, Shitashima K, Kanahara S, Komiyama H. 1997. Carbonate dynamics of thecoral reef system at Bora Bay, Miyako Island. Mar. Ecol. Prog. Ser. 156:1–16
Lai S, Gillis LG, Mueller C, Bouma TJ, Guest JR, et al. 2013. First experimental evidence of corals feedingon seagrass matter. Coral Reefs 32:1061–64
Leal MC, Ferrier-Pages C, Calado R, Thompson ME, Frischer ME, Nejstgaard JC. 2014. Coral feeding onmicroalgae assessed with molecular trophic markers. Mol. Ecol. 23:3870–76
Leichter JJ, Stewart HL, Miller SL. 2003. Episodic nutrient transport to Florida coral reefs. Limnol. Oceanogr.48:1394–407
Leichter JJ, Stokes MD, Hench JL, Witting J, Washburn L. 2012. The island-scale internal wave climate ofMoorea, French Polynesia. J. Geophys. Res. 117:C06008
Leichter JJ, Wing SR, Miller SL, Denny MW. 1996. Pulsed delivery of subthermocline water to Conch Reef(Florida Keys) by internal tidal bores. Limnol. Oceanogr. 41:1490–501
Longuet-Higgins MS, Stewart RW. 1962. Radiation stress and mass transport in gravity waves, with applicationto surf beats. J. Fluid Mech. 13:481–504
Lowe RJ, Falter JL, Monismith SG, Atkinson MJ. 2009a. A numerical study of circulation in a coastal reef-lagoon system. J. Geophys. Res. 114:C06022
Lowe RJ, Falter JL, Monismith SG, Atkinson MJ. 2009b. Wave-driven circulation of a coastal reef-lagoonsystem. J. Phys. Oceanogr. 39:873–93
Lowe RJ, Hart C, Pattiaratchi CB. 2010. Morphological constraints to wave-driven circulation in coastalreef-lagoon systems: a numerical study. J. Geophys. Res. 115:C09021
www.annualreviews.org • Oceanic Forcing of Coral Reefs 63
Ann
u. R
ev. M
arin
e. S
ci. 2
015.
7:43
-66.
Dow
nloa
ded
from
ww
w.a
nnua
lrev
iew
s.or
g A
cces
s pr
ovid
ed b
y U
nive
rsity
of
Wes
tern
Aus
tral
ia o
n 01
/30/
15. F
or p
erso
nal u
se o
nly.

MA07CH03-Lowe ARI 29 October 2014 6:57
Lowe RJ, Ivey GN, Brinkman RM, Jones NL. 2012. Seasonal circulation and temperature variability near theNorth West Cape of Australia. J. Geophys. Res. 117:C04010
Lowe RJ, Koseff JR, Monismith SG. 2005a. Oscillatory flow through submerged canopies: 1. Velocity struc-ture. J. Geophys. Res. 110:C10016
Lowe RJ, Koseff JR, Monismith SG, Falter JL. 2005b. Oscillatory flow through submerged canopies: 2. Canopymass transfer. J. Geophys. Res. 110:C10017
Lowe RJ, Shavit U, Falter JL, Koseff JR, Monismith SG. 2008. Modeling flow in coral communities with andwithout waves: a synthesis of porous media and canopy flow approaches. Limnol. Oceanogr. 53:2668–80
Lugo-Fernandez A, Gravois M. 2010. Understanding impacts of tropical storms and hurricanes on submergedbank reefs and coral communities in the northwestern Gulf of Mexico. Cont. Shelf Res. 30:1226–40
Macdonald IF, Elsayed MS, Mow K, Dullien FAL. 1979. Flow through porous media—the Ergun equationrevisited. Ind. Eng. Chem. Fundam. 18:199–208
Madin JS, Connolly SR. 2006. Ecological consequences of major hydrodynamic disturbances on coral reefs.Nature 444:477–80
Manzello DP. 2010. Ocean acidification hot spots: spatiotemporal dynamics of the seawater CO2 system ofeastern Pacific coral reefs. Limnol. Oceanogr. 55:239–48
Massel SR, Done TJ. 1993. Effects of cyclone waves on massive coral assemblages on the Great Barrier Reef:meteorology, hydrodynamics and demography. Coral Reefs 12:153–66
Massel SR, Gourlay MR. 2000. On the modelling of wave breaking and set-up on coral reefs. Coast. Eng.39:1–27
McCabe RM, Estrade P, Middleton JH, Melville WK, Roughan M, Lenain L. 2010. Temperature variabilityin a shallow, tidally isolated coral reef lagoon. J. Geophys. Res. 115:C12011
McClanahan TR, Baird AH, Marshall PA, Toscano MA. 2004. Comparing bleaching and mortality responsesof hard corals between southern Kenya and the Great Barrier Reef, Australia. Mar. Pollut. Bull. 48:327–35
McDonald CB, Koseff JR, Monismith SG. 2006. Effects of the depth to coral height ratio on drag coefficientsfor unidirectional flow over coral. Limnol. Oceanogr. 51:1294–301
McGowan HA, Sturman AP, MacKellar MC, Wiebe AH, Neil DT. 2010. Measurements of the local energybalance over a coral reef flat, Heron Island, southern Great Barrier Reef, Australia. J. Geophys. Res.115:D19124
McMahon A, Santos IR, Cyronak T, Eyre BD. 2013. Hysteresis between coral reef calcification and theseawater aragonite saturation state. Geophys. Res. Lett. 40:4675–79
Meinshausen M, Meinshausen N, Hare W, Raper SC, Frieler K, et al. 2009. Greenhouse-gas emission targetsfor limiting global warming to 2◦C. Nature 458:1158–62
Monismith SG. 2007. Hydrodynamics of coral reefs. Annu. Rev. Fluid Mech. 39:37–55Monismith SG. 2013. Flow through a rough, shallow reef. Coral Reefs 33:99–104Monismith SG, Davis KA, Shellenbarger GG, Hench JL, Nidzieko NJ, et al. 2010. Flow effects on benthic
grazing on phytoplankton by a Caribbean reef. Limnol. Oceanogr. 55:1881–92Monismith SG, Fong DA. 2004. A note on the potential transport of scalars and organisms by surface waves.
Limnol. Oceanogr. 49:1214–17Monismith SG, Genin A, Reidenbach MA, Yahel G, Koseff JR. 2006. Thermally driven exchanges between a
coral reef and the adjoining ocean. J. Phys. Oceanogr. 36:1332–47Monismith SG, Herdman LM, Ahmerkamp S, Hench JL. 2013. Wave transformation and wave-driven flow
across a steep coral reef. J. Phys. Oceanogr. 43:1356–79Moore JA, Bellchambers LM, Depczynski MR, Evans RD, Evans SN, et al. 2012. Unprecedented mass
bleaching and loss of coral across 12 of latitude in Western Australia in 2010–11. PLoS ONE 7:e51807Munk WH, Sargent MC. 1948. Adjustment of Bikini Atoll to ocean waves. Trans. Am. Geophys. Union 29:855–
60Munk WH, Sargent MC. 1954. Adjustment of Bikini Atoll to ocean waves. In Bikini and Nearby Atolls: Part
2. Oceanography (Physical), pp. 275–80. US Geol. Surv. Prof. Pap. 260. Washington, DC: US Gov. Print.Off.
Nelson RC. 1994. Depth limited design wave heights in very flat regions. Coast. Eng. 23:43–59Nepf HM. 2012. Flow and transport in regions with aquatic vegetation. Annu. Rev. Fluid Mech. 44:123–42
64 Lowe · Falter
Ann
u. R
ev. M
arin
e. S
ci. 2
015.
7:43
-66.
Dow
nloa
ded
from
ww
w.a
nnua
lrev
iew
s.or
g A
cces
s pr
ovid
ed b
y U
nive
rsity
of
Wes
tern
Aus
tral
ia o
n 01
/30/
15. F
or p
erso
nal u
se o
nly.

MA07CH03-Lowe ARI 29 October 2014 6:57
Nepf HM, Vivoni ER. 2000. Flow structure in depth-limited vegetative flow. J. Geophys. Res. 105:28547–57Odum HT, Odum EP. 1955. Trophic structure and productivity of a windward coral reef community on
Eniwetok atoll. Ecol. Monogr. 25:291–320Patten NL, Wyatt ASJ, Lowe RJ, Waite AM. 2011. Uptake of picophytoplankton, bacterioplankton and
virioplankton by a fringing coral reef community (Ningaloo Reef, Australia). Coral Reefs 30:555–67Pineda J, Starczak V, Tarrant A, Blythe J, Davis K, et al. 2013. Two spatial scales in a bleaching event:
Corals from the mildest and the most extreme thermal environments escape mortality. Limnol. Oceanogr.58:1531–45
Pomeroy A, Lowe RJ, Symonds G, van Dongeren A, Moore C. 2012. The dynamics of infragravity wavetransformation over a fringing reef. J. Geophys. Res. 117:C11022
Raupach MR, Shaw RH. 1982. Averaging procedures for flow within vegetation canopies. Bound. Layer Me-teorol. 22:79–90
Reidenbach MA, Koseff JR, Monismith SG. 2007. Laboratory experiments of fine-scale mixing and masstransport within a coral canopy. Phys. Fluids 19:075107
Reidenbach MA, Koseff JR, Monismith SG, Genin A. 2006. Effects of waves, currents and morphology onmass transfer in branched reef corals. Limnol. Oceanogr. 51:1134–41
Ribes M, Atkinson M. 2007. Effects of water velocity on picoplankton uptake by coral reef communities. CoralReefs 26:413–21
Ribes M, Coma R, Atkinson MJ, Kinzie RA. 2005. Sponges and ascidians control removal of particulate organicnitrogen from coral reef water. Limnol. Oceanogr. 50:1480–89
Richmond R. 1987. Energetics, competency, and long-distance dispersal of planula larvae of the coral Pocilloporadamicornis. Mar. Biol. 93:527–33
Riegl B, Piller WE. 2003. Possible refugia for reefs in times of environmental stress. Int. J. Earth Sci. 92:520–31Rosman JH, Hench JL. 2011. A framework for understanding drag parameterizations for coral reefs. J. Geophys.
Res. 116:C08025Santos IR, Erler D, Tait D, Eyre BD. 2010. Breathing of a coral cay: tracing tidally driven seawater recirculation
in permeable coral reef sediments. J. Geophys. Res. 115:C12010Sargent M, Austin T. 1949. Organic productivity of an atoll. Trans. Am. Geophys. Union 30:245–49Sebens KP, Grace SP, Helmuth B, Maney EJ, Miles JS. 1998. Water flow and prey capture by three scleractinian
corals, Madracis mirabilis, Montastrea cavernosa and Porites porites, in a field enclosure. Mar. Biol. 131:347–60Sebens KP, Johnson A. 1991. Effects of water movement on prey capture and distribution of reef corals.
Hydrobiologia 226:91–101Sebens KP, Witting J, Helmuth B. 1997. Effects of water flow and branch spacing on particle capture by the
reef coral Madraci mirabalis (Duchassaing and Michelotti). J. Exp. Mar. Biol. Ecol. 211:1–28Shaw EC, McNeil BI, Tilbrook B. 2012. Impacts of ocean acidification in naturally variable coral reef flat
ecosystems. J. Geophys. Res. 117:C03038Shulman M, Bermingham E. 1995. Early life histories, ocean currents, and the population genetics of
Caribbean reef fishes. Evolution 49:897–910Silverman J, Lazar B, Cao L, Caldeira K, Erez J. 2009. Coral reefs may start dissolving when atmospheric
CO2 doubles. Geophys. Res. Lett. 36:L05606Smith SV. 1973. Carbon dioxide dynamics: a record of organic carbon production, respiration, and calcification
in the Eniwetok reef flat community. Limnol. Oceanogr. 18:106–20Symonds G, Black KP, Young IR. 1995. Wave-driven flow over shallow reefs. J. Geophys. Res. 100:2639–48Taebi S, Lowe RJ, Pattiaratchi CB, Ivey GN, Symonds G, Brinkman R. 2011. Nearshore circulation in a
tropical fringing reef system. J. Geophys. Res. 116:C02016Tanaka Y, Miyajima T, Watanabe A, Nadaoka K, Yamamoto T, Ogawa H. 2011. Distribution of dissolved
organic carbon and nitrogen in a coral reef. Coral Reefs 30:533–41Tartinville B, Deleersnijder E, Rancher J. 1997. The water residence time in the Mururoa atoll lagoon:
sensitivity analysis of a three-dimensional model. Coral Reefs 16:193–203Teneva L, Dunbar RB, Mucciarone DA, Dunckley JF, Koseff JR. 2013. High-resolution carbon budgets
on a Palau back-reef modulated by interactions between hydrodynamics and reef metabolism. Limnol.Oceanogr. 58:1851–70
www.annualreviews.org • Oceanic Forcing of Coral Reefs 65
Ann
u. R
ev. M
arin
e. S
ci. 2
015.
7:43
-66.
Dow
nloa
ded
from
ww
w.a
nnua
lrev
iew
s.or
g A
cces
s pr
ovid
ed b
y U
nive
rsity
of
Wes
tern
Aus
tral
ia o
n 01
/30/
15. F
or p
erso
nal u
se o
nly.

MA07CH03-Lowe ARI 29 October 2014 6:57
Thomas FIM, Atkinson MJ. 1997. Ammonium uptake by coral reefs: effects of water velocity and surfaceroughness on mass transfer. Limnol. Oceanogr. 42:81–88
Thomas FIM, Cornelisen CD. 2003. Ammonium uptake by seagrass communities: effects of oscillatory versusunidirectional flow. Mar. Ecol. Prog. Ser. 247:51–57
UNEP-WCMC (UN Environ. Programme World Conserv. Monit. Cent.). 2003. Global distribution of coralreefs (version 7). A 1 km resolution grid produced by UNEP-WCMC from an updated version of the datalayer used in Spalding et al. (2001). Data Set, UNEP-WCMC, Cambridge, UK. http://data.unep-wcmc.org/datasets/14
Van Hooidonk R, Maynard J, Planes S. 2013. Temporary refugia for coral reefs in a warming world. Nat.Clim. Change 3:508–11
Venti A, Kadko D, Andersson AJ, Langdon C, Bates NR. 2012. A multi-tracer model approach to estimatereef water residence times. Limnol. Oceanogr. Methods 10:1078–95
Wang YH, Dai CF, Chen YY. 2007. Physical and ecological processes of internal waves on an isolated reefecosystem in the South China Sea. Geophys. Res. Lett. 34:L18609
Weitzman JS, Aveni-Deforge K, Koseff JR, Thomas FIM. 2013. Uptake of dissolved inorganic nitrogen byshallow seagrass communities exposed to wave-driven unsteady flow. Mar. Ecol. Prog. Ser. 475:65–83
Weller E, Nunez M, Meyers G, Masiri I. 2008. A climatology of ocean–atmosphere heat flux estimates over thegreat barrier reef and coral sea: implications for recent mass coral bleaching events. J. Clim. 21:3853–71
Wolanski E, Delesalle B. 1995. Upwelling by internal waves, Tahiti, French Polynesia. Cont. Shelf Res. 15:357–68
Wolanski E, Drew E, Abel KM, O’Brien J. 1988. Tidal jets, nutrient upwelling and their influence on theproductivity of the alga Halimeda in the Ribbon Reefs, Great Barrier Reef. Estuar. Coast. Shelf Sci. 26:169–201
Wolanski E, Thomson R. 1984. Wind-driven circulation on the northern Great Barrier Reef continental shelfin summer. Estuar. Coast. Shelf Sci. 18:271–89
Woodley J, Chornesky E, Cliffo P. 1981. Hurricane Allen’s impact on a Jamaican coral reef. Science 214:749–54Wyatt ASJ, Falter JL, Lowe RJ, Humphries S, Waite AM. 2012. Oceanographic forcing of nutrient uptake
and release over a fringing coral reef. Limnol. Oceanogr. 57:401–19Wyatt ASJ, Lowe RJ, Humphries S, Waite AM. 2010. Particulate nutrient fluxes over a fringing coral reef:
relevant scales of phytoplankton production and mechanisms of supply. Mar. Ecol. Prog. Ser. 405:113–30Yahel G, Post AF, Fabricius K, Marie D, Vaulot D, Genin A. 1998. Phytoplankton distribution and grazing
near coral reefs. Limnol. Oceanogr. 43:551–63Yahel R, Yahel G, Berman T, Jaffe JS, Genin A. 2005. Diel pattern with abrupt crepuscular changes of
zooplankton over a coral reef. Limnol. Oceanogr. 50:930–44Young IR, Zieger S, Babanin AV. 2011. Global trends in wind speed and wave height. Science 332:451–55Zhang Z, Falter JL, Lowe RJ, Ivey G. 2012. The combined influence of hydrodynamic forcing and calcification
on the spatial distribution of alkalinity in a coral reef system. J. Geophys. Res. 117:C04034Zhang Z, Falter JL, Lowe RJ, Ivey G, McCulloch M. 2013. Atmospheric forcing intensifies the effects of
regional ocean warming on reef-scale temperature anomalies during a coral bleaching event. J. Geophys.Res. 118:4600–16
66 Lowe · Falter
Ann
u. R
ev. M
arin
e. S
ci. 2
015.
7:43
-66.
Dow
nloa
ded
from
ww
w.a
nnua
lrev
iew
s.or
g A
cces
s pr
ovid
ed b
y U
nive
rsity
of
Wes
tern
Aus
tral
ia o
n 01
/30/
15. F
or p
erso
nal u
se o
nly.

MA07-FrontMatter ARI 6 November 2014 7:45
Annual Review ofMarine Science
Volume 7, 2015 Contents
Reflections on My Career as a Marine Physical Chemist, and Tales ofthe DeepFrank J. Millero � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � 1
Regional Ocean Data AssimilationChristopher A. Edwards, Andrew M. Moore, Ibrahim Hoteit,
and Bruce D. Cornuelle � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � �21
Oceanic Forcing of Coral ReefsRyan J. Lowe and James L. Falter � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � �43
Construction and Maintenance of the Ganges-Brahmaputra-MeghnaDelta: Linking Process, Morphology, and StratigraphyCarol A. Wilson and Steven L. Goodbred Jr. � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � �67
The Dynamics of Greenland’s Glacial Fjords and Their Role inClimateFiamma Straneo and Claudia Cenedese � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � �89
The Role of the Gulf Stream in European ClimateJaime B. Palter � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � 113
Long-Distance Interactions Regulate the Structure and Resilience ofCoastal EcosystemsJohan van de Koppel, Tjisse van der Heide, Andrew H. Altieri,
Britas Klemens Eriksson, Tjeerd J. Bouma, Han Olff, and Brian R. Silliman � � � � � � � 139
Insights into Particle Cycling from Thorium and Particle DataPhoebe J. Lam and Olivier Marchal � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � 159
The Size-Reactivity Continuum of Major Bioelements in the OceanRonald Benner and Rainer M.W. Amon � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � 185
Subsurface Chlorophyll Maximum Layers: Enduring Enigma orMystery Solved?John J. Cullen � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � 207
vi
Ann
u. R
ev. M
arin
e. S
ci. 2
015.
7:43
-66.
Dow
nloa
ded
from
ww
w.a
nnua
lrev
iew
s.or
g A
cces
s pr
ovid
ed b
y U
nive
rsity
of
Wes
tern
Aus
tral
ia o
n 01
/30/
15. F
or p
erso
nal u
se o
nly.

MA07-FrontMatter ARI 6 November 2014 7:45
Cell Size as a Key Determinant of Phytoplankton Metabolism andCommunity StructureEmilio Maranon � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � 241
Phytoplankton Strategies for Photosynthetic Energy AllocationKimberly H. Halsey and Bethan M. Jones � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � 265
Techniques for Quantifying Phytoplankton BiodiversityZackary I. Johnson and Adam C. Martiny � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � 299
Molecular Mechanisms by Which Marine Phytoplankton Respond toTheir Dynamic Chemical EnvironmentBrian Palenik � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � 325
The Molecular Ecophysiology of Programmed Cell Death in MarinePhytoplanktonKay D. Bidle � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � 341
Microbial Responses to the Deepwater Horizon Oil Spill: From CoastalWetlands to the Deep SeaG.M. King, J.E. Kostka, T.C. Hazen, and P.A. Sobecky � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � 377
Denitrification, Anammox, and N2 Production in Marine SedimentsAllan H. Devol � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � 403
Rethinking Sediment Biogeochemistry After the Discovery of ElectricCurrentsLars Peter Nielsen and Nils Risgaard-Petersen � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � 425
Mussels as a Model System for Integrative EcomechanicsEmily Carrington, J. Herbert Waite, Gianluca Sara, and Kenneth P. Sebens � � � � � � � � � � 443
Infectious Diseases Affect Marine Fisheries andAquaculture EconomicsKevin D. Lafferty, C. Drew Harvell, Jon M. Conrad, Carolyn S. Friedman,
Michael L. Kent, Armand M. Kuris, Eric N. Powell, Daniel Rondeau,and Sonja M. Saksida � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � 471
Diet of Worms Emended: An Update of Polychaete Feeding GuildsPeter A. Jumars, Kelly M. Dorgan, and Sara M. Lindsay � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � 497
Fish Locomotion: Recent Advances and New DirectionsGeorge V. Lauder � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � 521
There and Back Again: A Review of Residency and Return Migrationsin Sharks, with Implications for Population Structure andManagementDemian D. Chapman, Kevin A. Feldheim, Yannis P. Papastamatiou,
and Robert E. Hueter � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � 547
Contents vii
Ann
u. R
ev. M
arin
e. S
ci. 2
015.
7:43
-66.
Dow
nloa
ded
from
ww
w.a
nnua
lrev
iew
s.or
g A
cces
s pr
ovid
ed b
y U
nive
rsity
of
Wes
tern
Aus
tral
ia o
n 01
/30/
15. F
or p
erso
nal u
se o
nly.

MA07-FrontMatter ARI 6 November 2014 7:45
Whale-Fall Ecosystems: Recent Insights into Ecology, Paleoecology,and EvolutionCraig R. Smith, Adrian G. Glover, Tina Treude, Nicholas D. Higgs,
and Diva J. Amon � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � 571
Errata
An online log of corrections to Annual Review of Marine Science articles may be foundat http://www.annualreviews.org/errata/marine
viii Contents
Ann
u. R
ev. M
arin
e. S
ci. 2
015.
7:43
-66.
Dow
nloa
ded
from
ww
w.a
nnua
lrev
iew
s.or
g A
cces
s pr
ovid
ed b
y U
nive
rsity
of
Wes
tern
Aus
tral
ia o
n 01
/30/
15. F
or p
erso
nal u
se o
nly.






