Embed Size (px)
Citation preview

APPLIED AND ENVIRONMENTAL MICROBIOLOGY, Dec. 1991, p. 3629-36360099-2240/91/123629-08$02.00/0Copyright C) 1991, American Society for Microbiology
Vol. 57, No. 12
Occurrence and Ultrastructural Characterization ofBacteria in Association with and Isolated from
Azolla carolinianaSANDRA A. NIERZWICKI-BAUER* AND HEDY AULFINGER
Plant Research Group, Department of Biology, Rensselaer Polytechnic Institute,Troy, New York 12180-3590
Received 25 January 1991/Accepted 12 June 1991
The occurrence and ultrastructure of bacteria in leaf cavities of symbiotic Azolla caroliniana were examinedby transmission electron microscopy. Bacteria were observed in all leaf cavities of Azolla cultures. Fiveultrastructurally distinct types of bacteria were observed in each individual leaf cavity. Features used tocharacterize the bacteria included morphology, cell wall structure, and cytoplasmic organization. At least onegram-positive and as many as four gram-negative types of bacteria reside in leaf cavities ofA. caroliniana. Themorphological and ultrastructural characteristics of the gram-positive bacterium suggest that it is an
Arthrobacter sp. The gram-negative bacteria could not be cultured; therefore, they have not been classifiedfurther. Bacterial cell shape and cell wall structure were similar in leaf cavities of different ages, but cell sizeand cytoplasmic composition varied. The relative contributions of each bacterial type to the total communitywithin individual leaves was determined. Ultrastructural characteristics of bacterial isolates cultured fromA. caroliniana in a free-living state were also examined.
The Azolla-Anabaena symbiotic association consists ofsmall water ferns of the genus Azolla, prokaryotic cyano-bacteria tentatively identified as Anabaena azollae, and avariety of bacteria. In this association, specific functions areperformed by each of the partners. The cyanobiont fixesenough atmospheric N2 to satisfy both its own requirementfor combined nitrogen and that of its host (19-22, 27), andthe fern provides the endosymbionts with a carbon source inthe form of sucrose (26). Both Azolla caroliniana and A.azollae carry out photosynthesis of the higher-plant type(18), thereby enabling each to fix atmospheric CO2 via theCalvin cycle (26).As mentioned above, in addition to the cyanobionts,
bacteria are present in dorsal leaf cavities of Azolla spp. (11,12, 15, 16, 24). However, the role of bacteria in Azolla-Anabaena symbioses, if any, is unknown. A number ofinvestigators have classified these bacteria after isolationand subsequent characterization by biochemical and physi-ological tests (15, 24, 33). Although it is agreed that rod- tococcus-shaped bacteria are present in Azolla leaf cavities,the taxonomic classification of these bacteria has varied. Thebacterial isolates have been identified as Pseudomonas spp.(4), Alcaligenes faecalis, Caulobacter fusiformis (15), andmost recently Arthrobacter spp. (12, 24, 33). Eubacteriaidentified as Arthrobacter spp. (24) have been isolated andcultivated from Azolla caroliniana, A. filiculoides, A. mexi-cana, and A. pinnata (12, 24).
In ultrastructural studies ofA. caroliniana (6-8, 10, 23), A.filiculoides (6), and A. mexicana (16), bacteria can be ob-served in transmission electron micrographs of sections ofdorsal leaf cavities. However, detailed ultrastructural char-acterization of bacteria within Azolla leaf cavities has beenreported only for symbiotic and cyanobiont-free A. mexi-cana (17).
* Corresponding author.
The primary objective of the present study was to examinethe occurrence and ultrastructure of bacteria residing in leafcavities of different ages of laboratory-grown A. carolinianacultures. A secondary objective was to elucidate whether thesymbiotic bacterial floras are similar in laboratory-grownand greenhouse-grown cultures. Last, the ultrastructuralcharacteristics of bacterial isolates cultured from A. carolin-iana in a free-living state were examined.
MATERIALS AND METHODS
Organisms. Symbiotic cultures of A. caroliniana Willde-now (International Rice Research Institute [IRRI] Collectionno. 3001) were used in this study.The dorsal lobes from A. caroliniana leaves 5 to 8 (count-
ed from the apical meristem on the main axis) were removedby dissection and used for bacterial isolations according tothe procedure of Petro and Gates (24). In brief, leaves weresurface sterilized in 3% sodium hypochlorite, washed withsterile 5% sodium thiosulfate, and finally washed with sterilebuffered water. Examination of leaf cavities to determinewhether any had ruptured during the sterilization procedureand to check the efficacy of the procedure for removingsurface contaminants was also carried out according to themethod of Petro and Gates (24). Ten surface-sterilized leaveswere homogenized in 2 ml of sterile buffered water. Toobtain bacterial isolates, dilutions of the homogenate wereplated on Difco plate count agar. Bacterial isolations wereperformed on three separate occasions. Two bacterial iso-lates obtained independently of each other were used in thisstudy. These bacterial isolates were subcultured, as de-scribed below, numerous times over at least 1 to 2 monthsbefore preparation for electron microscopy studies.Growth conditions. A. caroliniana cultures were grown
under growth room conditions as described previously (6).
3629
on July 7, 2018 by guesthttp://aem
.asm.org/
Dow
nloaded from

3630 NIERZWICKI-BAUER AND AULFINGER
Cultures were maintained by frequent transfer and harvestedwhile growing actively.
In addition, A. caroliniana cultures were grown understandard greenhouse conditions without supplemental light-ing in trays containing IRRI medium (35) lacking combinednitrogen. In some instances, IRRI medium was supple-mented with garden soil. The plants used in the presentstudy were collected in April 1988.
Bacterial isolates obtained from A. caroliniana weregrown in a broth medium that was prepared so as to benutritionally equivalent to Difco plate count agar (25) in ashaking incubator (25°C; 200 rpm). Exponentially growingbacteria were harvested for transmission electron micros-copy studies.
Preparation of samples for transmission electron micros-copy. A. caroliniana fronds were prepared for thin sectioningand electron microscopy by double fixation with glutaralde-hyde and osmium tetroxide (OS04) as described previously(31). The bacterial isolates were prefixed in 3% glutaralde-hyde in 0.1 M sodium cacodylate buffer (pH 7.2) for 2 h atroom temperature, and then the cells were concentrated bycentrifugation and washed with sodium cacodylate buffer.The resulting cell pellet was resuspended in a small amountof molten 2% Noble agar (Difco) at 50°C. This cell-agarsuspension was transferred to a glass microscope slide,allowed to solidify, and cut into small blocks (<1 mm oneach side). The blocks were postfixed overnight at roomtemperature in 1% OS04 in 0.1 M sodium cacodylate buffer.Postfixed samples were washed twice with cacodylatebuffer. Azolla fronds and bacterial isolates were dehydratedthrough a graded ethanol series and embedded in Spurr'slow-viscosity epoxy resin (30). Thin sections were cut with adiamond knife and retrieved on 200-mesh nickel specimengrids. The sections were poststained for 15 min with 0.5%uranyl acetate in water and 3 min with 0.4% aqueous leadcitrate (28) or for 10 min with 4.0% aqueous lead citrate and2 h with 2% aqueous uranyl acetate, which is a slightmodification of the method of Ward and Claus (34). The thinsections were viewed and photographed with a JEOL JEM-100CX transmission electron microscope operating at anaccelerating potential of 80 kV. Leaf cavities of all ages fromover 20 individual fronds were thin sectioned and analyzed.Quantitative data on the frequencies of morphologicallydistinct bacteria were obtained from cell counts whichincluded a minimum of 200 cells. All of the bacterial cellspresent within electron micrographs of random fields wereincluded in these counts.
RESULTSThe occurrence and ultrastructure of bacteria in leaf
cavities of different ages were examined with A. carolinianacultures. Bacteria were present in all leaf cavities, and the
number of bacterial communities increased concomitantlywith leaf age. Both cyanobacterial and bacterial symbiontswere located peripherally within dorsal leaf cavities andwere separated from the central cavity region by an innerenvelope (data not shown) as described by Nierzwicki-Baueret al. (17).
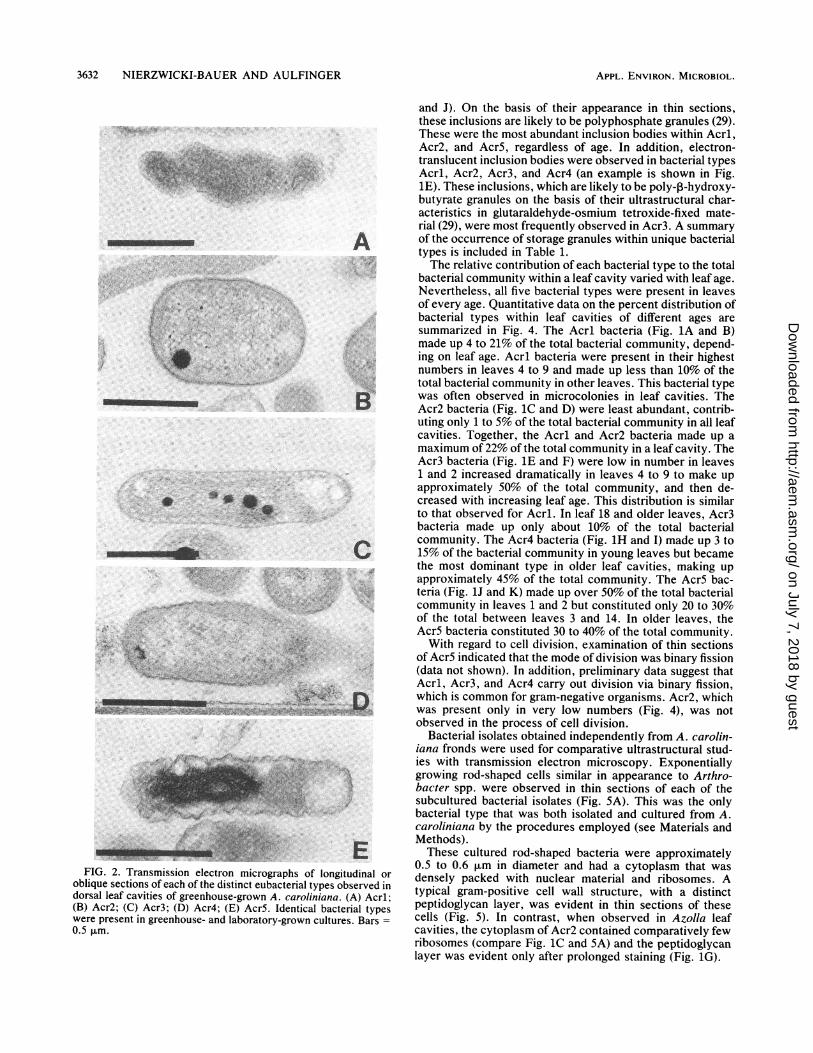
Five ultrastructurally distinct types of bacteria were ob-served within leaf cavities of A. caroliniana grown undergrowth room (Fig. 1) or greenhouse (Fig. 2) conditions. Cellshape, cell wall structure, and cytoplasmic organizationwere used as bases for distinction between types. Longitu-dinal sections and cross-sections of each of the distinctbacterial types found in leaf cavities of laboratory-grown A.caroliniana are presented in Fig. 1. Identical bacterial typeswere observed in greenhouse-grown Azolla cultures (Fig. 2).The five ultrastructurally distinct bacterial types found in
association with A. caroliniana were designated Acrl toAcr5. Characteristic features of these unique bacterial typesincluded morphology, cell diameter, cell wall structure, andaccumulation of storage material. Acr5 was surrounded by atypical gram-negative cell wall. In contrast, a distinct pepti-doglycan layer surrounding Acr2 was evident (Fig. 1G) aftersections were stained for increased times with increasedconcentrations of lead citrate and uranyl acetate (see Mate-rials and Methods). Acrl, Acr3, and Acr4 lacked a distinctpeptidoglycan layer even after extended staining; therefore,these bacteria were classified as gram-negative organisms.Gram-staining reactions confirmed the presence of numer-ous gram-negative bacteria (data not shown). The character-istic features of Acrl to Acr5, as observed in young leafcavities of A. caroliniana, are summarized in Table 1.While it was always possible to distinguish the unique
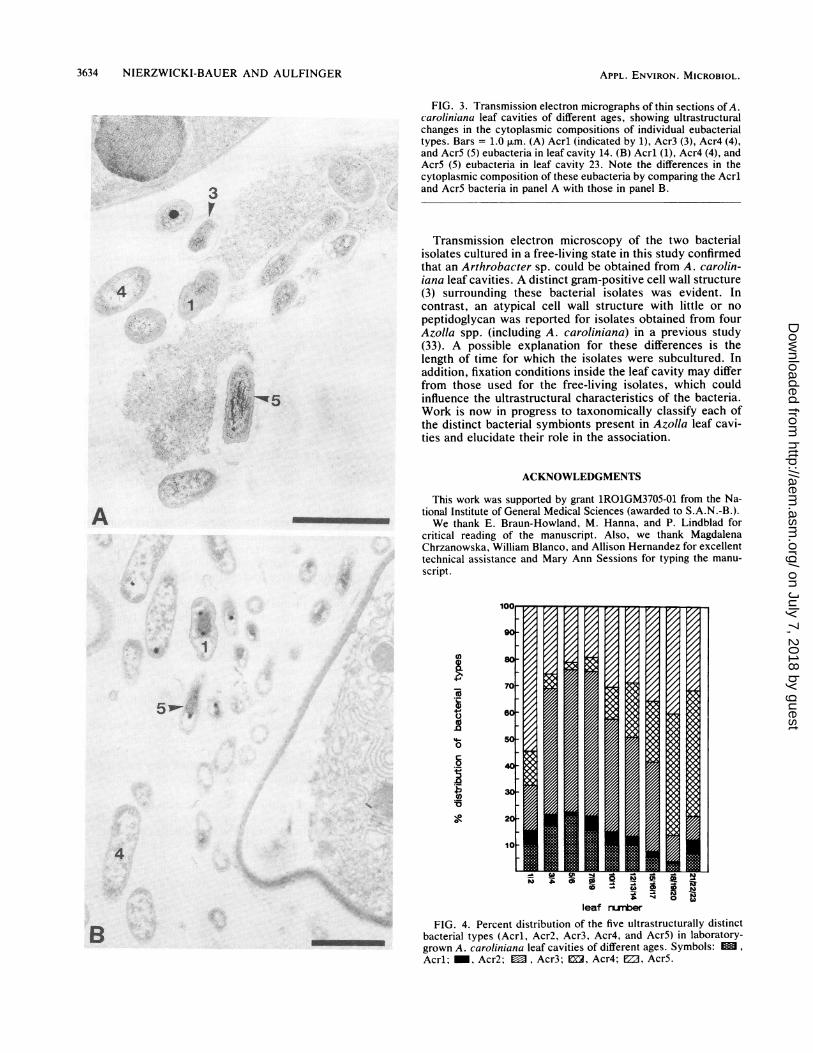
bacterial types from one another on the basis of cell wallstructure and cell shape, all types displayed variations in cellsize and cytoplasmic composition between younger andolder leaf cavities (Fig. 3). For instance, a comparison ofAcr5 present in leaf 14 (Fig. 3A) with Acr5 present in leaf 23(Fig. 3B) demonstrates both of these changes. Increasedelectron-transparent accumulations, which were likely to beglycogen on the basis of their appearance in glutaraldehyde-osmium tetroxide-fixed material (9), were observed in allbacterial types residing in older leaf cavities (Fig. 3B). Theaccumulation of glycogen in cells present in older leafcavities may indicate a microenvironment which is nitrogendeficient but carbohydrate rich. Under these conditions,many bacteria are known to store polysaccharides (36).
Additionally, the bacteria associated with A. carolinianacontained inclusions of two different electron densities andvarious sizes. There was no apparent correlation betweenthe number and/or type of inclusions and leaf age. Electron-dense spherical inclusion bodies ranging from 0.02 to 0.15,um in diameter were observed in thin sections of Acrl,Acr2, Acr3, and Acr5 (examples are shown in Fig. 1A, B, G,
FIG. 1. Transmission electron micrographs of thin sections of the five ultrastructurally distinct bacterial types observed in leaf cavities ofA. caroliniana cultures that were grown in a controlled-environment room (see Materials and Methods). Longitudinal sections of bacteria areshown on the left, and cross-sections are shown on the right. Intracellular inclusion bodies resembling polyphosphate (PP) granules andpoly-,-hydroxybutyrate (PHB) granules were occasionally observed. Bars = 0.5 p.m. (A and B) Helix-shaped bacteria, designated Acrl,containing a densely packed cytoplasm with nuclear material, ribosomes, and PP granules. (C and D) Rod- to coccus-shaped bacteria,designated Acr2, with a cytoplasmic region containing relatively few ribosomes. (E and F) Irregularly rod-shaped bacteria, designated Acr3,containing a dense cytoplasmic region (c), electron-transparent glycogen accumulations (G) which are generally 25 to 30 nm in diameter, andlarger electron-transparent PHB granules. (G) Cross-section of Acr2 and Acr3. Sections were stained with 4.0% lead citrate for 10 min and2% uranyl acetate for 2 h to detect a densely stained gram-positive cell wall surrounding Acr2 (arrowhead). This wall structure is characteristicof Arthrobacter spp. (H and I) Thick rod-shaped bacteria, designated Acr4. (J and K) Rod-shaped bacteria, designated Acr5, similar inappearance to Acr4 except for having a more uniform size and shape.
APPL. ENVIRON. MICROBIOL.
on July 7, 2018 by guesthttp://aem
.asm.org/
Dow
nloaded from

BACTERIA ASSOCIATED WITH A. CAROLlNIANA 3631
.s
X,#
,0., *_s .7. fX,.: .::R . . -w fs
f } v DDllc M < iLlilr#' '8 A 1. | r
w wSr. -s_. t . r :{ t > 't i asERS 0 _ 01 ;'f *5 w 9 Y
-s 8, e ft
/Wa: s
_ v
C-
rw.0, t_
t 1B5:F =oWf ei 8l );
46OFt t s. s *
ns . .w
teJ to .
O 0 ts * < \d-_ pXf s | L'V0'v:*t Dl E Si k L00.ev R 0k fx' * l s'
'' 8^'i^G____ *;VOL. 57, 1991
.. I
-1
I"I 1.1
1. k itt '.T.
k0*.6L,.,
'A G.
11
I. 'Amlokobt.-
-I"
r
A6
-1
".- I
F7-
iw-icvrI
*1 I
4_L.. " K
on July 7, 2018 by guesthttp://aem
.asm.org/
Dow
nloaded from
3632 NIERZWICKI-BAUER AND AULFINGER
:1
FIG. 2. Transmission electron micrographs of longitudinal oroblique sections of each of the distinct eubacterial types observed indorsal leaf cavities of greenhouse-grown A. caroliniana. (A) Acrl;(B) Acr2; (C) Acr3; (D) Acr4; (E) Acr5. Identical bacterial typeswere present in greenhouse- and laboratory-grown cultures. Bars =
0.5 ,um.
and J). On the basis of their appearance in thin sections,these inclusions are likely to be polyphosphate granules (29).These were the most abundant inclusion bodies within Acrl,Acr2, and Acr5, regardless of age. In addition, electron-translucent inclusion bodies were observed in bacterial typesAcrl, Acr2, Acr3, and Acr4 (an example is shown in Fig.1E). These inclusions, which are likely to be poly-p-hydroxy-butyrate granules on the basis of their ultrastructural char-acteristics in glutaraldehyde-osmium tetroxide-fixed mate-rial (29), were most frequently observed in Acr3. A summaryof the occurrence of storage granules within unique bacterialtypes is included in Table 1.The relative contribution of each bacterial type to the total
bacterial community within a leaf cavity varied with leaf age.Nevertheless, all five bacterial types were present in leavesof every age. Quantitative data on the percent distribution ofbacterial types within leaf cavities of different ages aresummarized in Fig. 4. The Acrl bacteria (Fig. 1A and B)made up 4 to 21% of the total bacterial community, depend-ing on leaf age. Acrl bacteria were present in their highestnumbers in leaves 4 to 9 and made up less than 10% of thetotal bacterial community in other leaves. This bacterial typewas often observed in microcolonies in leaf cavities. TheAcr2 bacteria (Fig. 1C and D) were least abundant, contrib-uting only 1 to 5% of the total bacterial community in all leafcavities. Together, the Acrl and Acr2 bacteria made up amaximum of 22% of the total community in a leaf cavity. TheAcr3 bacteria (Fig. 1E and F) were low in number in leaves1 and 2 increased dramatically in leaves 4 to 9 to make upapproximately 50% of the total community, and then de-creased with increasing leaf age. This distribution is similarto that observed for Acrl. In leaf 18 and older leaves, Acr3bacteria made up only about 10% of the total bacterialcommunity. The Acr4 bacteria (Fig. 1H and I) made up 3 to15% of the bacterial community in young leaves but becamethe most dominant type in older leaf cavities, making upapproximately 45% of the total community. The Acr5 bac-teria (Fig. 1J and K) made up over 50% of the total bacterialcommunity in leaves 1 and 2 but constituted only 20 to 30%of the total between leaves 3 and 14. In older leaves, theAcr5 bacteria constituted 30 to 40% of the total community.With regard to cell division, examination of thin sections
of Acr5 indicated that the mode of division was binary fission(data not shown). In addition, preliminary data suggest thatAcrl, Acr3, and Acr4 carry out division via binary fission,which is common for gram-negative organisms. Acr2, whichwas present only in very low numbers (Fig. 4), was notobserved in the process of cell division.
Bacterial isolates obtained independently from A. carolin-iana fronds were used for comparative ultrastructural stud-ies with transmission electron microscopy. Exponentiallygrowing rod-shaped cells similar in appearance to Arthro-bacter spp. were observed in thin sections of each of thesubcultured bacterial isolates (Fig. 5A). This was the onlybacterial type that was both isolated and cultured from A.caroliniana by the procedures employed (see Materials andMethods).These cultured rod-shaped bacteria were approximately
0.5 to 0.6 ,um in diameter and had a cytoplasm that wasdensely packed with nuclear material and ribosomes. Atypical gram-positive cell wall structure, with a distinctpeptidoglycan layer, was evident in thin sections of thesecells (Fig. 5). In contrast, when observed in Azolla leafcavities, the cytoplasm of Acr2 contained comparatively fewribosomes (compare Fig. 1C and SA) and the peptidoglycanlayer was evident only after prolonged staining (Fig. 1G).
APPL. ENVIRON. MICROBIOL.
on July 7, 2018 by guesthttp://aem
.asm.org/
Dow
nloaded from

BACTERIA ASSOCIATED WITH A. CAROLINIANA 3633
TABLE 1. Properties of bacterial cell types residing in young leaf cavities (leaves 1 to 5) of symbiotic A. caroliniana
Type Morphology (Dim)a structureb Storage granules" Other features
Acrl Helix 0.30-0.41 (G-) PP (abundant), PHB (rare) Occurred in microcoloniesAcr2 Rod to coccus 0.50-0.55 (G+) PP (abundant), PHB (rare) Least abundant typeAcr3 Long, irregularly 0.30-0.42 (G-) G (abundant), PP (abundant),
shaped rod PHB (abundant)Acr4 Thick rod 0.354.44 G- PP (rare) PHB (rare) Usually lacked inclusion bodiesAcr5 Thin rod 0.24-0.30 G- PP (abundant) PP frequently located at poles or ends of
electron-dense nucleoplasmic region
a Diameter as measured from cross-sections.b (G-), probable G- (gram-negative) cell wall structure; (G+), G+ (gram-positive) cell wall structure, based on peptidoglycan layer observed in overstained
sections and absence of evident lipopolysaccharide layer.I G, glycogen; PHB, poly-,3-hydroxybutyrate; PP, polyphosphate.
The cell division process of the bacterial isolates obtainedfrom A. caroliniana (shown in Fig. 5) was quite similar, if notidentical, to that reported for Arthrobacter spp. (13, 32, 34).Cell division was initiated by simultaneous invagination ofthe cytoplasmic membrane and the peptidoglycan layer toform a division septum (Fig. 5B). Septum formation contin-ued until the cytoplasm was transected by a cross-septum.At this time, the cells were still attached and had a commonouter membrane layer. Later in the process of cell division,daughter cells with a ruptured outer membrane were ob-served (Fig. 5C). Dividing cells showing scars where theouter membrane of the cell wall had ruptured were seen insome instances (Fig. 5D). This cell division mechanism wasnever observed in situ for eubacteria in the symbiotic state.However, as indicated above, the one bacterium that wasclassified as a gram-positive organism (Acr2) was not ob-served in the process of cell division.
DISCUSSION
Previous studies have characterized bacteria isolated fromAzolla spp. and cultured in a free-living state (11, 12, 24, 33).However, these studies have excluded the examination ofsymbiotic bacteria that were unable to grow under thespecific isolation conditions employed. In contrast, thepresent study used direct electron microscopic examinationof thin sections of Azolla leaf cavities for the preliminaryclassification of all bacterial symbionts found in associationwith A. caroliniana.
In agreement with results previously reported for threeAzolla spp. (24), increased numbers of bacteria were ob-served in A. caroliniana leaf cavities of increasing age. Inaddition, on the basis of quantitative analyses (from countsof bacteria in serial sections of leaf cavities of different ages),it was determined that total bacterial numbers were higher inleaf cavities of comparable ages for A. caroliniana than forA. mexicana. This result is in only partial agreement withdata presented by Petro and Gates (24), in whose study thenumbers of bacteria in individual leaf cavities were higher inA. caroliniana up to leaf 8 and higher in A. mexicana forleaves older than leaf 8. However, the numbers obtained byPetro and Gates (24) include only those bacteria that wereisolated aerobically on plate count agar. Thus, a number ofbacteria present in Azolla leaf cavities may have beenexcluded in the Petro and Gates (24) study. Regardless of thequantitative method used, there is widespread agreementthat in many Azolla species, very large bacterial communi-ties are present in the cavities of older leaves. In fact, insome instances, the number of bacteria can actually equal
the number of cyanobacteria (11). In view of the largenumbers of bacteria in Azolla leaves, it seems likely thatthey have a significant effect and function in the association.However, it remains unclear why A. caroliniana harborsmore types as well as larger numbers of bacteria than A.mexicana.
In the A. caroliniana association, five morphologically andultrastructurally distinct types of bacteria were present ineach Azolla leaf cavity, regardless of age. In addition, thesame five bacterial types could be identified in culturesgrown under greenhouse conditions. These results suggestthat a specific bacterial community is always associated witha specific Azolla species or strain. Supporting this conclu-sion are transmission electron micrographs of sections ofleaf cavities ofA. caroliniana cultures grown under a varietyof conditions in different laboratories. In reports from thoselaboratories, bacteria that are readily discernible as Acr4 andAcrS (12) and Acrl, Acr3, Acr4, and Acr5 (8) are evident. Inother instances, magnifications of the micrographs wereinsufficient for positive bacterial identifications to be made(10, 23).Although the results of this study demonstrate the pres-
ence of five ultrastructurally distinct bacterial types in A.caroliniana leaf cavities, it must be emphasized that thisdoes not necessarily mean that these types represent fivedifferent genera or species of bacteria. Because a singleorganism can assume a variety of morphologies (i.e., bepleomorphic), the possibility remains that there are fewerthan five unique bacterial types in association with A.caroliniana.
It is interesting to note that only one of the ultrastructur-ally distinct bacterial types, the helical bacterium, is ob-served in both A. caroliniana (Acrl) and A. mexicana (16)(Amxl). Therefore, the helical bacterium may be of partic-ular interest for future studies because of its apparentlybroad host range.
Regarding identification and classification of the bacteriafound in association with A. caroliniana, it can be concludedthat only one of the bacterial types identified in this study, onthe basis of morphology, ultrastructure, and cell division, islikely to belong to the genus Arthrobacter. In support of thisstatement are morphological and ultrastructural compari-sons between the symbiotically associated bacteria andfree-living Arthrobacter spp. grown under extremely variedconditions (1, 2, 5, 13, 14, 32). In such comparisons, none ofthe symbiotically associated bacterial types were ultrastruc-turally identical to free-living Arthrobacter spp. However,Acr2 closely resembles Arthrobacter spp., having a similarmorphology and cell wall structure.
VOL. 57, 1991
on July 7, 2018 by guesthttp://aem
.asm.org/
Dow
nloaded from
3634 NIERZWICKI-BAUER AND AULFINGER
FIG. 3. Transmission electron micrographs of thin sections of A.caroliniana leaf cavities of different ages, showing ultrastructuralchanges in the cytoplasmic compositions of individual eubacterialtypes. Bars = 1.0 ,um. (A) Acrl (indicated by 1), Acr3 (3), Acr4 (4),and Acr5 (5) eubacteria in leaf cavity 14. (B) Acrl (1), Acr4 (4), andAcr5 (5) eubacteria in leaf cavity 23. Note the differences in thecytoplasmic composition of these eubacteria by comparing the Acrland Acr5 bacteria in panel A with those in panel B.
A.
d st
54t:fiXAtesr '
/g%,;
4-;i
Transmission electron microscopy of the two bacterialisolates cultured in a free-living state in this study confirmedthat an Arthrobacter sp. could be obtained from A. carolin-iana leaf cavities. A distinct gram-positive cell wall structure(3) surrounding these bacterial isolates was evident. Incontrast, an atypical cell wall structure with little or nopeptidoglycan was reported for isolates obtained from fourAzolla spp. (including A. caroliniana) in a previous study(33). A possible explanation for these differences is thelength of time for which the isolates were subcultured. Inaddition, fixation conditions inside the leaf cavity may differfrom those used for the free-living isolates, which couldinfluence the ultrastructural characteristics of the bacteria.Work is now in progress to taxonomically classify each ofthe distinct bacterial symbionts present in Azolla leaf cavi-ties and elucidate their role in the association.
ACKNOWLEDGMENTS
This work was supported by grant 1ROlGM3705-01 from the Na-tional Institute of General Medical Sciences (awarded to S.A.N.-B.).We thank E. Braun-Howland, M. Hanna, and P. Lindblad for
critical reading of the manuscript. Also, we thank MagdalenaChrzanowska, William Blanco, and Allison Hernandez for excellenttechnical assistance and Mary Ann Sessions for typing the manu-script.
9c
.0(0
~0
so
7C
so
4C
30
20
10
N) Ji a % Q NC
A
-" 0
leaf rumber
u
n
CO
FIG. 4. Percent distribution of the five ultrastructurally distinctbacterial types (Acrl, Acr2, Acr3, Acr4, and Acr5) in laboratory-grown A. caroliniana leaf cavities of different ages. Symbols: ,
Acrl; M, Acr2; 1 , Acr3; R, Acr4; C, Acr5.
'Iihir 1
APPL. ENVIRON. MICROBIOL.
on July 7, 2018 by guesthttp://aem
.asm.org/
Dow
nloaded from

BACTERIA ASSOCIATED WITH A. CAROLINIANA 3635
v
B
r S' ~~DFIG. 5. Transmission electron micrographs of thin-sectioned
bacteria that were isolated from dorsal leaf cavities of A. carolinianaand cultured in a free-living state, showing details of the cell divisionprocess. Bars = 0.5 ,um. (A) Exponentially growing rod-shaped cell.(B) Cell in an early stage of division. Septum formation (arrowhead)via invagination of the cytoplasmic membrane and peptidoglycanlayer of the cell wall is visible. (C) Cells late in the process ofdivision. A separate outer membrane layer surrounds each daughtercell. (D) Dividing cell showing scars (s) where the outer membranelayer of the cell wall has ruptured. This is typical of snappingdivision.
REFERENCES1. Aminov, R. I., V. M. Anan'in, and E. L. Golovlev. 1986. The
dynamics of Arthrobacter globiformis growth under the condi-tions of batch and continuous cultivation. Mikrobiologiya 55:395Z400.
2. Balkwill, D. L., and W. C. Ghiorse. 1985. Characterization ofsubsurface bacteria associated with two shallow aquifers inOklahoma. Appl. Environ. Microbiol. 50:580-588.
3. Beveridge, T. J. 1981. Ultrastructure, chemistry, and function ofthe bacterial wall. Int. Rev. Cytol. 72:229-317.
4. Bottomley, W. B. 1920. The effect of organic matter on thegrowth of various water plants in culture solution. Ann. Bot.(London) 39:353-365.
5. Boylen, C. W., and J. L. Pate. 1973. Fine structure of Arthro-bacter crystallopoietes during long-term starvation of rod andspherical stage cells. Can. J. Microbiol. 19:1-5.
6. Braun-Howland, E. B., P. Lindblad, S. A. Nierzwicki-Bauer, andB. Bergman. 1988. Dinitrogenase reductase (Fe-protein) in thecyanobacterial symbionts of three Azolla species: localizationand sequence of appearance during heterocyst differentiation.Planta 176:319-332.
7. Braun-Howland, E. B., and S. A. Nierzwicki-Bauer. 1990. Oc-currence of t}e 32-kDa Q3-binding protein of photosystem II inve&etative cells, heterocysts and akinetes of Azolla carolinianacyanobionts. Planta 180:361-371.
8. Calvert, H. E., M. K. Pence, and G. A. Peters. 1985. Ultrastruc-tural ontogeny of leaf cavity trichomes in Azolla implies a
functional role in metabolic exchange. Protoplasma 129:10-27.9. Chao, L., and C. C. Bowen. 1971. Purification and properties of
glycogen isolated from a blue-green alga, Nostoc muscorum. J.Bacteriol. 105:331-338.
10. Duckett, J. G., R. Toth, and S. L. Sonia. 1975. An ultrastructuralstudy of the Azolla, Anabaena azollae relationship. New Phy-tol. 75:111-118.
11. Gates, J. E., R. W. Fisher, and R. A. Candler. 1980. Theoccurrence of coryneform bacteria in the leaf cavity of Azolla.Arch. Microbiol. 127:163-165.
12. Grilli Caiola, M., C. Forni, and M. Castagnola. 1988. Bacteria inthe Azolla-Anabaena association. Symbiosis 5:185-198.
13. Krulwich, T. A., and J. L. Pate. 1971. Ultrastructural explana-tion for snapping postfission movements in Arthrobacter crys-tallopoietes. J. Bacteriol. 105:408-412.
14. Luscombe, B. M., and T. R. G. Gray. 1971. Effect of varyinggrowth rate on the morphology of Arthrobacter. J. Gen. Micro-biol. 69:433-434.
15. Newton, J. W., and A. I. Herman. 1979. Isolation of cyanobac-teria from the aquatic fern, Azolla. Arch. Microbiol. 120:161-165.
16. Nierzwicki-Bauer, S. A., and H. Aulfinger. 1990. Ultrastructuralcharacterization of eubacteria residing within leaf cavities ofsymbiotic and cyanobiont-free Azolla mexicana. Curr. Micro-biol. 21:123-129.
17. Nierzwicki-Bauer, S. A., H. Aulfinger, and E. B. Braun-How-land. 1989. Ultrastructural characterization of an inner envelopethat confines Azolla endosymbionts to the leaf cavity periphery.Can. J. Bot. 67:2711-2719.
18. Peters, G. A. 1977. The Azolla-Anabaena azollae symbiosis, p.231-258. In A. Hollaender (ed.), Genetic engineering for nitro-gen fixation. Plenum Publishing Corp., New York.
19. Peters, G. A., and B. C. Mayne. 1974. The Azolla, Anabaenaazollae relationship. I. Initial characterization of the associa-tion. Plant Physiol. (Bethesda) 53:813-819.
20. Peters, G. A., and B. C. Mayne. 1974. The Azolla, Anabaenaazollae relationship. II. Localization of nitrogenase activity as
assayed by acetylene reduction. Plant Physiol. (Bethesda) 53:820-824.
21. Peters, G. A., T. B. Ray, B. C. Mayne, and R. E. Toia, Jr. 1980.Azolla-Anabaena association: morphological and physiologicalstudies, p. 293-309. In W. E. Newton and W. H. Orme-Johnson(ed.), Nitrogen fixation, vol. 2. University Park Press, Balti-more.
22. Peters, G. A., R. E. Toia, Jr., W. R. Evans, D. K. Crist, B. C.Mayne, and R. E. Poole. 1980. Characterization and compari-
VOL. 57, 1991
on July 7, 2018 by guesthttp://aem
.asm.org/
Dow
nloaded from

3636 NIERZWICKI-BAUER AND AULFINGER
sons of five N2-fixing Azolla-Anabaena associations. I. Optimi-zation of growth conditions for biomass increase and N contentin a controlled environment. Plant Cell Environ. 3:261-269.
23. Peters, G. A., R. E. Toia, Jr., D. Raveed, and N. J. Levine. 1978.The Azolla-Anabaena azollae relationship. VI. Morphologicalaspects of the association. New Phytol. 80:583-593.
24. Petro, M. J., and J. E. Gates. 1987. Distribution of Arthrobactersp. in the leaf cavities of four species of the N-fixing Azolla fern.Symbiosis 3:41-48.
25. Rand, M. C., A. E. Greenberg, and M. J. Taras (ed.). 1976.Standard methods for the examination of water and wastewater,14th ed. American Public Health Association, Washington,D.C.
26. Ray, T. B., B. C. Mayne, R. E. Toia, Jr., and G. A. Peters. 1979.Azolla-Anabaena relationship. VIII. Photosynthetic character-ization of the association and individual partners. Plant Physiol.(Bethesda) 64:791-795.
27. Ray, T. B., G. A. Peters, B. C. Mayne, and R. E. Toia, Jr. 1978.Azolla-Anabaena relationship. VII. Distribution of ammonia-assimilating enzymes, protein, and chlorophyll between hostand symbiont. Plant Physiol. (Bethesda) 62:463-467.
28. Reynolds, E. S. 1963. The use of lead citrate at high pH as anelectron-opaque stain in electron microscopy. J. Cell Biol.17:208-212.
29. Shively, J. M. 1974. Inclusion bodies of prokaryotes. Annu.
Rev. Microbiol. 28:167-187.30. Spurr, A. R. 1969. A low-viscosity epoxy resin embedding
medium for electron microscopy. J. Ultrastruct. Res. 26:31-43.31. Stevens, S. E., Jr., D. A. M. Paone, and D. L. Balkwill. 1981.
Accumulation of cyanophycin granules as a result of phosphatelimitation in Agmenellum quadruplicatum. Plant Physiol. (Be-thesda) 67:716-719.
32. Stevenson, I. L. 1968. The fine structure of Arthrobacter pas-cens and the development of mesosomes during the growthcycle. Can. J. Microbiol. 14:1029-1034.
33. Wallace, W. H., and J. E. Gates. 1986. Identification of eubac-teria isolated from leaf cavities of four species of the N-fixingAzolla fern as Arthrobacter Conn and Dimmick. Appl. Environ.Microbiol. 52:425-429.
34. Ward, C. M., Jr., and G. W. Claus. 1973. Gram characteristicsand wall ultrastructure of Arthrobacter crystallopoietes duringcoccus-rod morphogenesis. J. Bacteriol. 114:378-389.
35. Watanabe, I., C. R. Espihias, N. A. Berja, and B. V. Alimagno.1977. The utilization of the Azolla-Anabaena complex as anitrogen fertilizer for rice. Int. Rice Res. Inst. Res. Pap. Ser.11:1-15.
36. Zevenhuizen, L. P. T. M. 1966. Formation and function of theglycogen-like polysaccharide of Arthrobacter. Antonie vanLeeuwenhoek J. Microbiol. Serol. 32:356-372.
APPL. ENVIRON. MICROBIOL.
on July 7, 2018 by guesthttp://aem
.asm.org/
Dow
nloaded from





















![Wylie%20 familiarization%203630 3629[1]](https://img.pdfslide.us/doc/110x75/55ccb7cdbb61eb0e758b45e7/wylie20-familiarization203630-36291-55cd9dc6144b2.jpg)







