Embed Size (px)
Citation preview

REVIEW / SYNTHÈSE
Nutritional aspects of epigenetic inheritance
Shaan E. Alam, R.B. Singh, Siddharth Gupta, Parinita Dherange,Fabien De Meester, Agnieszka Wilczynska, Suniti Dharwadkar, Douglas Wilson, andPali Hungin
Abstract: The impact of diet and environmental factors on genes concerned with epigenetic inheritance and the mechanismof evolution has grown significantly beyond the Modern Synthesis period. Epigenetic inheritance is the passing of pheno-typic change to subsequent generations in ways that are outside the genetic code of DNA. Recently, polymorphisms of thehuman Delta-5 (fatty acid desaturase, FADS1) and Delta-6 (FADS2) desaturase genes have been described as being associ-ated with the level of several long-chain n-3 and n-6 polyunsaturated fatty acids (PUFAs) in serum phospholipids. Increasedconsumption of refined starches and sugar increases the generation of superoxide anion in the tissues and free fatty acids(FFA) in the blood. There is an increased amount and activity of nuclear factor-kB (NF-kB), a transcriptional factor regulat-ing the activity of at least 125 genes, most of which are pro-inflammatory. The consumption of glucose may be associatedwith an increase in 2 other pro-inflammatory transcription factors: activating protein-1 (AP-1), and early growth responseprotein-1 (Egr-1). AP-1 regulates the transcription of matrix metallo-proteinases and Egr-1 modulates the transcription of tis-sue factor and plasminogen activator inhibitor-1. It is possible that a complex set of factors, including nutritional factors,come into play during epigenetic inheritance.
Key words: epigenetic inheritance, TsimTsoum concept, evolution, diet.
Résumé : L’impact de la diète et des facteurs environnementaux sur les gènes concernant l’hérédité épigénétique et les mé-canismes d’évolution, dépasse ce qui avait été envisagé dans Modern Synthesis. L’hérédité épigénétique définit la transmis-sion d’un changement phénotypique à des générations subséquentes de manière externe au code génétique d’ADN.Récemment, les polymorphismes des gènes codant les désaturases Delta-5 (désaturase d’acide gras, FADS1) et Delta-6(FADS2) ont été décrits pour être associés au niveau de plusieurs acides gras polyinsaturés à longue chaine n-3 et n-6 dansles phospholipides sériques. L’augmentation de la consommation de farines et de sucres raffinés augmente la générationd’anion superoxyde dans les tissus et des acides gras libres dans le sang. Il s’en suit une augmentation de la quantité et del’activité du facteur NF-kB, un facteur de transcription qui régule l’activité d’au moins 125 gènes dont la majorité est pro-inflammatoire. La consommation de glucose peut être associée à une augmentation de deux autres facteurs de transcriptionpro-inflammatoires : AP-1 (activating protein-1) et Egr-1 (early growth response protein-1). AP-1 régule la transcription desmétalloprotéases de la matrice et Egr-1 module la transcription du facteur tissulaire TF et de l’inhibiteur de l’activateur duplasminogène PAI-1. Il est possible qu’un jeu complexe de facteurs, incluant des facteurs nutritionnels, soit mis en jeu lorsde l’hérédité épigénétique.
Mots‐clés : hérédité épigénétique, concept TsimTsoum, évolution, diète.
[Traduit par la Rédaction]
Received 1 December 2011. Accepted 12 April 2012. Published at www.nrcresearchpress.com/cjpp on 17 July 2012.
S.E. Alam. University College of Medical Sciences, GTB Hospital, Delhi, India.R.B. Singh, S. Gupta, P. Dherange, F. De Meester, and A. Wilczynska. The TsimTsoum Institute, Golebia Street, 31-007 Krakow,Poland.S. Dharwadkar. S.B. College of Science, Aurangabad, Maharashtra, India.D. Wilson and P. Hungin. School of Medicine and Health, Wolfson Research Institute, Queen’s Campus, Stockton-on-Tees, DurhamTS17 6BH, UK.
Corresponding author: Ram B. Singh (e-mail: [email protected]).
This Invited Review is one of a number of papers published in the Special Issue entitled “Heart Health and Care,” which focuses on newknowledge of the physiology of cardiovascular functions in health, and pathophysiology of cardiovascular dysfunctions.
989
Can. J. Physiol. Pharmacol. 90: 989–994 (2012) doi:10.1139/Y2012-105 Published by NRC Research Press
Can
. J. P
hysi
ol. P
harm
acol
. Dow
nloa
ded
from
ww
w.n
rcre
sear
chpr
ess.
com
by
120.
117.
138.
77 o
n 11
/27/
14Fo
r pe
rson
al u
se o
nly.

IntroductionThere is evidence that primary risk factors such as diet,
physical inactivity, tobacco, stress, pollutants, radiation, anddrugs can influence genetic structure and function. There hasbeen much discussion over the last 2 decades concerningwhich environmental factors, like nutrition, could be impor-tant in the patho-biology of epigenetic inheritance (Eaton etal. 1998; De Meester 2009; Eaton 2009). Although JulianHuxley, who published his landmark Monograph Evolution:The Modern Synthesis in 1942 did not comment about therole of nutrition in evolution, this monograph brought Dar-win’s ideas into the 20th century and incorporated a knowl-edge of genes that was emerging in this century. GregorMendel’s experiments, at Hynčice (Vražné) in the (now)Czech Republic, on inheritance in this context were quite in-teresting (monograph: Experiments with Plant Hybrids). Inthe mid-20th century, Barbara McClintock discovered trans-posable elements, where parts of the genome can jumparound and cause mutations or alter the gene expression,skewing Mendelian ratios and inheritance patterns. Darwinbelieved that organisms gradually adapt to their environmentsvia minute physical adjustments. Biologists have since foundevidence that dramatic adaptations can also occur, for in-stance in the form of major morphological changes. Theseadaptations appear to be the cause of changes in structurefrom apes to man and thence to various human races. Suchadaptations may be responsible for the development of dis-eases, or health and longevity, or for the evolution of prehis-toric man to modern humans.The lifespan of some organisms can be extended by muta-
tions that alter how DNA is packaged in their cells. Recentstudies indicate that chromatin modifiers regulate lifespan inseveral organisms, raising the question as to whether changesin chromatin states in the parental generation could be in-completely reprogrammed in the next generation, and therebyaffect the lifespan of descendants (Greer et al. 2010, 2011). Itis possible that epigenetic inheritance may be the key to solv-ing many unexplained phenomena in humans, related tohealth and disease as well as with behavior. The concept ofcellular memory passed onto offspring may be mediated bythe character of the spermatozoon, its biochemistry and biol-ogy, and its interaction with environmental factors such asdiet, stress, tobacco, alcohol, radiation, and drugs.
VariabilityAny physical, biological, etc., variable can be studied in a
general, as well as specific diagnostic or therapeutic medicalcontext. In the general context, a variable can be informativewith respect to many human and environmental conditions(Halberg et al. 2003). Such a broad view is particularly war-ranted in the case of blood pressure and heart rate and its re-lated biomarkers, which may have transgenerationalepigenetic inheritance potential. Some people may inherit ta-chycardia and variability in blood pressure and other bio-markers similar to familial history of hypertension, diabetes,and coronary artery disease. Professor Franz Halberg firstlearned about the ranges of variability in the counts of certainblood cells that stain with a pink dye (eosin) (Halberg et al.1958). Next, he mapped the variability of epileptic seizuresand electroencephalograms, and then after solving several
puzzles, had the opportunity to be the first in documentingthe 24 h rhythms of RNA and DNA synthesis (Halberg et al.1958), at a time when nucleic acid was believed to be THEconstant feature of organisms (Halberg et al. 2003). Whatstarted with blood cell counts resulted in scientific tools, ifnot disciplines, as branches of a unified science. It would beinteresting to know the role of variability according to timestructure, and the role of clock genes in transgenerational ep-igenetic inheritance that has been reported for a variety oftraits across a number of species, and which may have verymany confounders.
Epigenetic inheritanceA higher incidence of hypertension, type-2 diabetes, coro-
nary artery disease, and obesity in the offspring of patientssuffering from these problems is well established. A longerlifespan in the offspring of parents also having a longer life-span has also been observed. The color of flowers in plants,eye color in Drosophila, and fur color in mice are all knownto be epigenetically inherited. The exact mechanism of trans-generational epigenetic inheritance has been a mystery, and itis quite interesting that for the first time this has been linkedto longevity, which may be due to the interaction of geneswith environmental factors as well as due to specific altera-tion in the chromatin state and mutations thereof. It is alsopossible that amino acids and u-6/u-3 fatty acid ratio of thecell can influence DNA and RNA as well as chromatins (Fer-nandes et al. 1990; Jolly et al. 2001; Hulbert et al. 2006; Fe-dacko et al. 2010; Takahashi et al. 2011).Studies on Caenorhabditis elegans, a microscopic round-
worm that typically lives to a ripe old age of 2–3 weeks, sug-gested that a single gene called daf-2 could dramaticallyregulate how long an organism lived (Adams 2008). Thegene acts by controlling a lot of other genes that just happento coordinate the survival system within worms. The proteincoded for by this gene (designated DAF-2) looked much likethe receptor protein within humans that responds to the hor-mone insulin. In other words, the worm protein is simply aprimitive form of our own insulin receptors. The gene isknown to regulate a number of factors in addition to aging,including stress resistance, metabolism, and development inworms and flies; the gene codes for a receptor protein that isactivated by an “insulin-like” growth factor. This signalingpathway is analogous to the mammalian insulin pathway.The genes that show the most change fall into several differ-ent classes, some of which line up nicely with existing hy-potheses about the mechanisms of aging in other organisms;this includes the belief that various genes encode for proteinsthat extend life by acting as antioxidants, regulating metabo-lism, and exerting an antibacterial effect (Adams 2008; Fig. 1)One particular gene affected by daf-2 is daf-16; this gene
encodes a transcription factor, or a protein that determineswhen and where hundreds of other genes are turned on. Nor-mally, the DAF-2 protein (which is an insulin receptor) exertsa dampening effect on the DAF-16 protein through phosphor-ylation. Experiments have determined that this activation ofDAF-16 (caused by the absence of a phosphate group) is anecessary step toward lifespan extension (Fig. 1). Transcrip-tion factor DAF-16 controls the expression of a battery ofgenes, many of which have effects on lifespan — promoting
990 Can. J. Physiol. Pharmacol. Vol. 90, 2012
Published by NRC Research Press
Can
. J. P
hysi
ol. P
harm
acol
. Dow
nloa
ded
from
ww
w.n
rcre
sear
chpr
ess.
com
by
120.
117.
138.
77 o
n 11
/27/
14Fo
r pe
rson
al u
se o
nly.
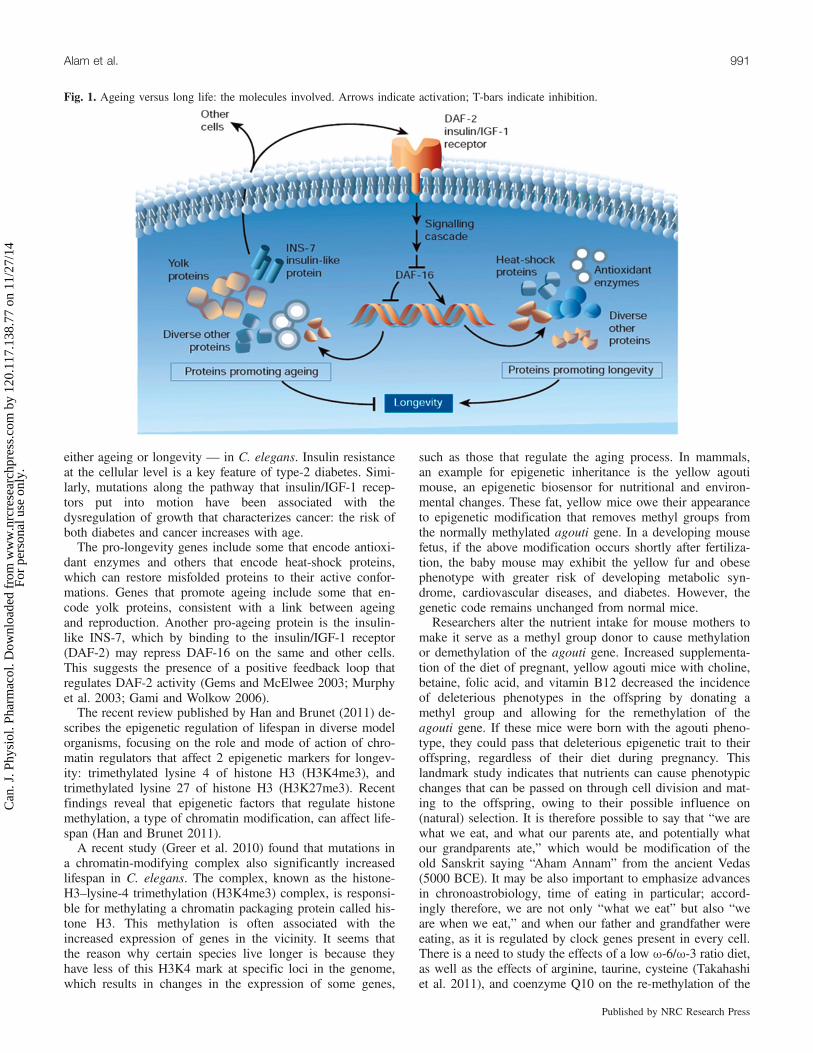
either ageing or longevity — in C. elegans. Insulin resistanceat the cellular level is a key feature of type-2 diabetes. Simi-larly, mutations along the pathway that insulin/IGF-1 recep-tors put into motion have been associated with thedysregulation of growth that characterizes cancer: the risk ofboth diabetes and cancer increases with age.The pro-longevity genes include some that encode antioxi-
dant enzymes and others that encode heat-shock proteins,which can restore misfolded proteins to their active confor-mations. Genes that promote ageing include some that en-code yolk proteins, consistent with a link between ageingand reproduction. Another pro-ageing protein is the insulin-like INS-7, which by binding to the insulin/IGF-1 receptor(DAF-2) may repress DAF-16 on the same and other cells.This suggests the presence of a positive feedback loop thatregulates DAF-2 activity (Gems and McElwee 2003; Murphyet al. 2003; Gami and Wolkow 2006).The recent review published by Han and Brunet (2011) de-
scribes the epigenetic regulation of lifespan in diverse modelorganisms, focusing on the role and mode of action of chro-matin regulators that affect 2 epigenetic markers for longev-ity: trimethylated lysine 4 of histone H3 (H3K4me3), andtrimethylated lysine 27 of histone H3 (H3K27me3). Recentfindings reveal that epigenetic factors that regulate histonemethylation, a type of chromatin modification, can affect life-span (Han and Brunet 2011).A recent study (Greer et al. 2010) found that mutations in
a chromatin-modifying complex also significantly increasedlifespan in C. elegans. The complex, known as the histone-H3–lysine-4 trimethylation (H3K4me3) complex, is responsi-ble for methylating a chromatin packaging protein called his-tone H3. This methylation is often associated with theincreased expression of genes in the vicinity. It seems thatthe reason why certain species live longer is because theyhave less of this H3K4 mark at specific loci in the genome,which results in changes in the expression of some genes,
such as those that regulate the aging process. In mammals,an example for epigenetic inheritance is the yellow agoutimouse, an epigenetic biosensor for nutritional and environ-mental changes. These fat, yellow mice owe their appearanceto epigenetic modification that removes methyl groups fromthe normally methylated agouti gene. In a developing mousefetus, if the above modification occurs shortly after fertiliza-tion, the baby mouse may exhibit the yellow fur and obesephenotype with greater risk of developing metabolic syn-drome, cardiovascular diseases, and diabetes. However, thegenetic code remains unchanged from normal mice.Researchers alter the nutrient intake for mouse mothers to
make it serve as a methyl group donor to cause methylationor demethylation of the agouti gene. Increased supplementa-tion of the diet of pregnant, yellow agouti mice with choline,betaine, folic acid, and vitamin B12 decreased the incidenceof deleterious phenotypes in the offspring by donating amethyl group and allowing for the remethylation of theagouti gene. If these mice were born with the agouti pheno-type, they could pass that deleterious epigenetic trait to theiroffspring, regardless of their diet during pregnancy. Thislandmark study indicates that nutrients can cause phenotypicchanges that can be passed on through cell division and mat-ing to the offspring, owing to their possible influence on(natural) selection. It is therefore possible to say that “we arewhat we eat, and what our parents ate, and potentially whatour grandparents ate,” which would be modification of theold Sanskrit saying “Aham Annam” from the ancient Vedas(5000 BCE). It may be also important to emphasize advancesin chronoastrobiology, time of eating in particular; accord-ingly therefore, we are not only “what we eat” but also “weare when we eat,” and when our father and grandfather wereeating, as it is regulated by clock genes present in every cell.There is a need to study the effects of a low u-6/u-3 ratio diet,as well as the effects of arginine, taurine, cysteine (Takahashiet al. 2011), and coenzyme Q10 on the re-methylation of the
Fig. 1. Ageing versus long life: the molecules involved. Arrows indicate activation; T-bars indicate inhibition.
Alam et al. 991
Published by NRC Research Press
Can
. J. P
hysi
ol. P
harm
acol
. Dow
nloa
ded
from
ww
w.n
rcre
sear
chpr
ess.
com
by
120.
117.
138.
77 o
n 11
/27/
14Fo
r pe
rson
al u
se o
nly.

agouti gene and their effects on phenotypic variations. How-ever this mode of inheritance needs to penetrate more than afew generations before it earns a place in evolutionary concept.Greer et al. (2010) proposed that the plasticity of aging
suggests that longevity may be controlled epigenetically byspecific alterations in chromatin state. The link between chro-matin and aging has mostly focused on histone deacetylationby the Sir2 family. However, less is known about the role ofother histone modifications in longevity. Histone methylationhas a crucial role in development and in maintaining stemcell pluripotency in mammals. Regulators of histone methyl-ation have been associated with aging in worms and flies, butcharacterization of their role and mechanism of action hasbeen limited. Here we identify the ASH-2 trithorax complex,which trimethylates histone H3 at lysine-4 (H3K4), as a reg-ulator of lifespan in C. elegans in a directed RNA interfer-ence (RNAi) screen in fertile worms. Deficiencies inmembers of the ASH-2 complex, ASH-2 itself, WDR-5, andthe H3K4 methyltransferase SET-2, extend worm lifespan.Conversely, the H3K4 demethylase RBR-2 is required fornormal lifespan, consistent with the idea that an excess ofH3K4 trimethylation — a mark associated with active chro-matin — is detrimental for longevity. Lifespan extension in-duced by ASH-2 complex deficiency requires the presenceof an intact adult germline and the continuous production ofmature eggs. ASH-2 and RBR-2 act in the germline, at leastin part, to regulate lifespan and to control a set of genes in-volved in lifespan determination. These results indicate thatthe longevity of the soma is regulated by an H3K4 methyl-transferase–demethylase complex acting in the C. elegansgermline. There is a need to study the effect of the u-6/u-3ratio of fatty acids, amino acids, and energy restriction in thecells for which H3K4 regulates the lifespan (Fernandes et al.1990; Jolly et al. 2001; Hulbert et al. 2006; Takahashi et al.2011).Findings imply that overexpression of superoxide dismu-
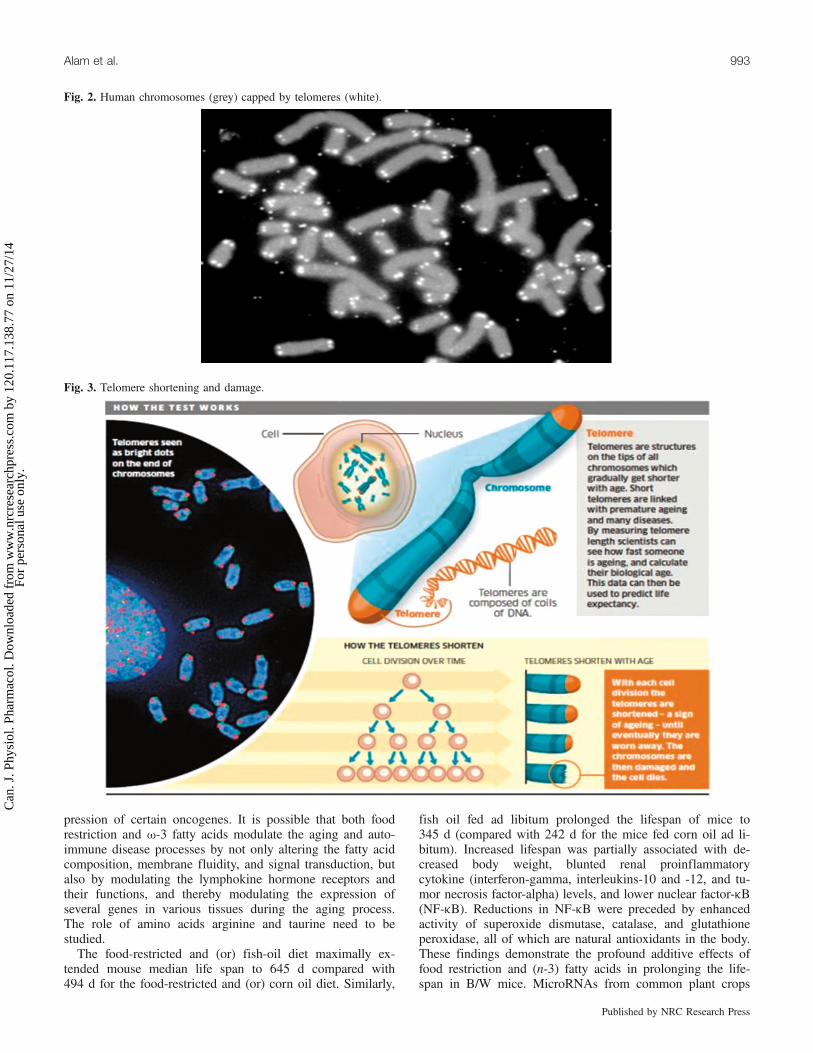
tase increases the lifespan of C. elegans by activating longevity-promoting transcription factors (Cabreiro et al. 2011). Telo-meres are structures at the end of a chromosome that areinvolved in the replication and stability of the chromosome.Genetic factors and environmental stressors can shorten thelength of the telomere, with telomere length becoming anemerging marker of biological age (McElligott and Wellinger1997; Figs. 2 and 3).Telomere lengths in humans are largely genetically deter-
mined and contained within upper and lower limits. Telo-meres that are too short lead to premature aging andincreased risk of aging-associated diseases. Telomeres thatare too long may have negative impacts through an increasedrisk of proliferative-related disorders. Both situations wouldincrease vulnerability and reduce the chances of living avery long life (Joeng et al. 2004). Several studies have shownincreased survival rates among individuals with establishedcardiovascular disease that have a high dietary intake of ma-rine u-3 fatty acids.When multivariable models were used to examine the as-
sociation of baseline levels of u-3 fatty acids (docosahexae-noic acid [DHA] and eicosapentaenoic acid [EPA]) withsubsequent change in telomere length, Farzaneh-Far et al.(2010) found that individuals in the lowest quartile of DHA+EPA experienced the most rapid rate of telomere shortening,
whereas those in the highest quartile experienced the slow-est rate of telomere shortening. Among the patients withstable coronary artery disease, there was an inverse relation-ship between baseline blood levels of marine u-3 fattyacids and the rate of telomere shortening over 5 years.Omega-3 fatty acids may protect against oxidative stress, orincrease the activity of the telomerase enzyme, which maythen decrease telomere shortening by creating more accuratetelomere copies. These findings raise the possibility that u-3 fatty acids may protect against cellular aging in patientswith coronary heart disease (Farzaneh-Far et al. 2010). Te-lomere length is an indicator of biological aging, and dys-function of telomeres is linked to age-related pathologiessuch as cardiovascular disease, Parkinson’s disease, Alz-heimer’s disease, and cancer. Telomere length has beenshown to be positively associated with nutritional status inhuman and animal studies. Various nutrients influence telo-mere length, potentially through mechanisms that reflecttheir role in cellular functions including inflammation, oxi-dative stress, DNA integrity, DNA methylation, and the ac-tivity of telomerase, the enzyme that adds the telomericrepeats to the ends of the newly synthesized DNA. Ahealthy lifestyle and diet are positively correlated with telo-mere length.Twelve different vitamins, minerals, and bioactives have
been identified to enhance telomere protection and keepaging at bay. They include the B vitamins folate and B12,niacin, vitamin A, vitamin D (which lowers levels of CRP, aharmful protein), vitamins C and E (which limit oxidativestress to telomeres), magnesium (which is a required mineralfor DNA replication and repair), zinc, iron (which is onlygood in small doses), turmeric (spice), and u-3 fatty acids(both of which stimulate antioxidants), and polyphenols(e.g., from grape seeds and green tea, which also protectDNA from oxidative stress) (Paul 2011).Free radicals and lipid peroxides arising from both sponta-
neous and enzymatic oxidation of polyunsaturated fatty acidsare the major source of endogenous DNA damage linked tovarious age-related pathologies and initiating carcinogenesisand atherogenesis. The polyunsaturated fatty acids in biolog-ical membranes serve as both the target and source of oxida-tive damage and can be regarded as the most unstable classof biomolecules in the body. Muscle and liver phospholipidsfrom these long-living mice lines have a reduced amount ofthe highly polyunsaturated u-3 docosahexaenoic acid com-pared with the genetically heterogeneous Dancer (Dc) mice,and consequently their membranes are less prone to peroxida-tive damage. The relationship between maximum longevityand membrane peroxidation index is similar for these mouselines, as previously observed for mammals in general. It issuggested that peroxidation-resistant membranes may be animportant component of extended longevityIn short-lived autoimmune-prone mice, both food restric-
tion and u-3 (n-3) fatty acid diets lowered the severity ofautoimmune disease, both in lupus-prone (New ZealandBlack × New Zealand White (B/W)) F1 mice and in miceprone to develop lymphoproliferative and renal diseases,whereas saturated (n-9) and polyunsaturated (n-6) dietary lip-ids exacerbated autoimmune disease, as well as significantlyenhancing the expression of several oncogenes in lymphoidtissues. Food restriction and u-3 fatty acids decreased the ex-
992 Can. J. Physiol. Pharmacol. Vol. 90, 2012
Published by NRC Research Press
Can
. J. P
hysi
ol. P
harm
acol
. Dow
nloa
ded
from
ww
w.n
rcre
sear
chpr
ess.
com
by
120.
117.
138.
77 o
n 11
/27/
14Fo
r pe
rson
al u
se o
nly.
pression of certain oncogenes. It is possible that both foodrestriction and u-3 fatty acids modulate the aging and auto-immune disease processes by not only altering the fatty acidcomposition, membrane fluidity, and signal transduction, butalso by modulating the lymphokine hormone receptors andtheir functions, and thereby modulating the expression ofseveral genes in various tissues during the aging process.The role of amino acids arginine and taurine need to bestudied.The food-restricted and (or) fish-oil diet maximally ex-
tended mouse median life span to 645 d compared with494 d for the food-restricted and (or) corn oil diet. Similarly,
fish oil fed ad libitum prolonged the lifespan of mice to345 d (compared with 242 d for the mice fed corn oil ad li-bitum). Increased lifespan was partially associated with de-creased body weight, blunted renal proinflammatorycytokine (interferon-gamma, interleukins-10 and -12, and tu-mor necrosis factor-alpha) levels, and lower nuclear factor-kB(NF-kB). Reductions in NF-kB were preceded by enhancedactivity of superoxide dismutase, catalase, and glutathioneperoxidase, all of which are natural antioxidants in the body.These findings demonstrate the profound additive effects offood restriction and (n-3) fatty acids in prolonging the life-span in B/W mice. MicroRNAs from common plant crops
Fig. 2. Human chromosomes (grey) capped by telomeres (white).
Fig. 3. Telomere shortening and damage.
Alam et al. 993
Published by NRC Research Press
Can
. J. P
hysi
ol. P
harm
acol
. Dow
nloa
ded
from
ww
w.n
rcre
sear
chpr
ess.
com
by
120.
117.
138.
77 o
n 11
/27/
14Fo
r pe
rson
al u
se o
nly.

such as rice and cabbage can be found in the blood and tis-sues of humans and other plant-eating mammals. In a recentstudy (Fedacko et al. 2012), the score for intake of prudentfoods was significantly greater and the ratio of u-6/u-3 fattyacids of the diet significantly lower for deaths due to “injury”and accidental causes compared with deaths due to non-communicable diseases (NCD). Multivariate logistic regres-sion analysis revealed that after adjustment for age, totalprudent foods (odds ratio (OR; men): 1.11; CI, 1.06–1.18;OR (women): 109; CI, 1.04–1.16) as well as fruits, vegeta-bles, legumes, and nuts (OR (men): 1.07; CI, 1.02–1.12;OR (women): 1.05; CI, 1.99–1.11) were independently, in-versely associated with deaths due to NCDs, whereas West-ern-type foods (OR (men): 1.02; CI, 0.95–1.09; OR(women): 1.00; CI, 0.94–1.06); meat and eggs (OR (men):1.00; CI, 0.94–1.06; OR (women): 0.98; CI, 0.93–1.04),and refined carbohydrates (OR (men): 0.98; CI, 0.91–1.05;OR (women): 0.95; CI, 0.89–1.02), and high u-6/u-3 ratioof fatty acids were positively associated with deaths due toNCDs. It is possible that a low u-6/u-3 ratio diet may havebeneficial effects on telomere length or on genes responsi-ble for longevity, whereas a high u-6/u-3 ratio may haveadverse effects. In brief, these observations may have addi-tional implications in the management of obesity, diabetes,atherogenesis, thrombosis, cancer, and (or) the aging proc-ess. It is possible that the greater life expectancy in Japanand Mediterranean countries may be because of increasedintake of u-3 fatty acids and lower consumption of u-6 fattyacids.
AcknowledgementsWe thank the TsimTsoum Institute for providing the logis-
tic support to write this article. The authors acknowledge thatthere is no conflict of interest associated with this study.
ReferencesAdams, J.U. 2008. Genetic control of aging and life span. Nature
Education, 1(1).Cabreiro, F., Ackerman, D., Doonan, R., Araiz, C., Back, P., Papp, D.,
et al. 2011. Increased life span from overexpression of superoxidedismutase in Caenorhabditis elegans is not caused by decreasedoxidative damage. Free Radic. Biol. Med. 51(8): 1575–1582.doi:10.1016/j.freeradbiomed.2011.07.020. PMID:21839827.
De Meester, F. 2009. Progress in lipid nutrition: the Columbusconcept addressing chronic diseases. World Rev. Nutr. Diet. 100:110–121. doi:10.1159/000235716. PMID:19696532.
Eaton, S.B. 2009. Evolution and cholesterol. World Rev. Nutr. Diet.100: 46–54. doi:10.1159/000235710. PMID:19696526.
Eaton, S.B., Eaton, S.B., III, Sinclair, A.J., Cordain, I., and Mann,N.J. 1998. Dietary intake of long chain polyunsaturated fattyacids during the Paleolithic. World Rev. Nutr. Diet. 83: 12–23.doi:10.1159/000059672. PMID:9648501.
Farzaneh-Far, R., Lin, J., Epel, E.S., Harris, W.S., Blackburn, E.H.,and Whooley, M.A. 2010. Association of marine omega-3 fattyacid levels with telomeric aging in patients with coronary heartdisease. JAMA, 303(3): 250–257. doi:10.1001/jama.2009.2008.PMID:20085953.
Fedacko, J., Vargova, V., Singh, R.B., De Meester, F., Pella, D., andJuneja, L. 2010. How nutrient and nutraceuticals modulate geneticexpressions in cardiovascular diseases. In World Heart ResearchYearbook. Vol. 2. Edited by F. Halberg and R.B. Singh. NovaScience Publishers, Inc., Hauppauge, N.Y.
Fedacko, J., Vargova1, V., Singh, R.B., Anjum, B., Takahash, T.,Tongnuka,, M., Dharwadkar, S., et al. 2012. Association of high u-6/u-3 fatty acid ratio diet with causes of death due tononcommunicable diseases among urban decedents in north India..The Open Nutra. J.5: 113–123. Available from benthamscience.com/open/tonutraj/articles/V005/113TONUTRAJ.pdf.
Fernandes, G., Venkatraman, J., Khare, A., Horbach, G.J., andFriedrichs, W. 1990. Modulation of gene expression in auto-immune disease and aging by food restriction and dietary lipids.Proc. Soc. Exp. Biol. Med. 193(1): 16–22. PMID:2403672.
Gami, M.S., and Wolkow, C.A. 2006. Studies of Caenorhabditiselegans DAF-2/insulin signaling reveal targets for pharmacologi-cal manipulation of life span. Aging Cell, 5(1): 31–37. doi:10.1111/j.1474-9726.2006.00188.x. PMID:16441841.
Gems, D., and McElwee, J.J. 2003. Ageing: Microarraying mortality.Nature, 424(6946): 259–261. doi:10.1038/424259a. PMID:12867961.
Greer, E.L., Maures, T.J., Hauswirth, A.G., Green, E.M., Leeman,D.S., Maro, G.S., et al. 2010. Members of the H3K4trimethylation complex regulate lifespan in a germline-dependentmanner in C. elegans. Nature, 466(7304): 383–387. doi:10.1038/nature09195. PMID:20555324.
Greer, E.L., Maures, T.J., Ucar, D., Hauswirth, A.G., Mancini, E.,Lim, J.P., et al. 2011. Transgenerational epigenetic inheritance oflongevity in Caenorhabdites elegance. Nature, 479(7373): 365–371. doi:10.1038/nature10572. PMID:22012258.
Halberg, F., Barnum, C.P., Silber, R.H., and Bittner, J.J. 1958. 24-hour rhythms at several levels of integration in mice on differentlighting regimens. Proc. Soc. Exp. Biol. Med. 97(4): 897–900.PMID:13554517.
Halberg, F., Cornelissen, G., Katinas, G., Syutkina, E.V., Sothern,R.B., Zaslavskaya, R., et al. 2003. Transdisciplinary unifyingimplications of circadian findings in the 1950s. J. CircadianRhythms, 1(1): 2. doi:10.1186/1740-3391-1-2. PMID:14728726.
Han, S., and Brunet, A. 2011. Histone methylation makes its mark onlongevity. Trends Cell Biol. 22(1): 42–49. doi:10.1016/j.tcb.2011.11.001. PMID:22177962.
Hulbert, A.J., Faulks, S.C., Harper, J.M., Miller, R.A., andBuffenstein, R. 2006. Extended longevity of wild-derived miceis associated with peroxidation-resistant membrane. Mech. AgeingDev. 127(8): 653–657. doi:10.1016/j.mad.2006.03.002. PMID:16620917.
Joeng, K.S., Song, E.J., Lee, K.J., and Lee, J. 2004. Long lifespan inworms with long telomeric DNA. Nat. Genet. 36(6): 607–611.doi:10.1038/ng1356. PMID:15122256.
Jolly, C.A., Muthukumar, A., Avula, C.P., Troyer, D., and Fernandes,G. 2001. Life span is prolonged in food-restricted autoimmune-prone (NZB × NZW)F(1) mice fed a diet enriched with (n-3) fattyacids. J. Nutr. 131(10): 2753–2760. PMID:11584100.
McElligott, R., and Wellinger, R.J. 1997. The terminal DNA structureof mammalian chromosomes. EMBO J. 16(12): 3705–3714.doi:10.1093/emboj/16.12.3705. PMID:9218811.
Murphy, C.T., McCarroll, S., Bargmann, C.I., Fraser, A., Kamath,R.S., Ahringer, J., et al. 2003. Genes that act downstream ofDAF-16 to influence the life span of Caenorhabditis elegans.Nature, 424(6946): 277–283. doi:10.1038/nature01789. PMID:12845331.
Paul, L. 2011. Diet, nutrition and telomere length. J. Nutr. Biochem.22(10): 895–901. doi:10.1016/j.jnutbio.2010.12.001. PMID:21429730.
Takahashi, T., Toda, E., Singh, R.B., De Meester, F., Wilczynska, A.,and Wilson, D. 2011. Essential and nonessential amino acids inrelation to glutamate. Open Nutraceuticals J. 4(1): 205–212.doi:10.2174/1876396001104010205.
994 Can. J. Physiol. Pharmacol. Vol. 90, 2012
Published by NRC Research Press
Can
. J. P
hysi
ol. P
harm
acol
. Dow
nloa
ded
from
ww
w.n
rcre
sear
chpr
ess.
com
by
120.
117.
138.
77 o
n 11
/27/
14Fo
r pe
rson
al u
se o
nly.





























