Embed Size (px)
Citation preview

Institute of Crop Science, Nutritional Crop Physiology
University of Hohenheim
Prof. Dr. Günter Neumann
Nutrient seed priming improves abiotic stress tolerance in
Zea mays L. and Glycine max L.
Dissertation
Submitted in fulfilment of the requirement of the degree “ Doktor der
Agrarwissenschaften”
(Dr. sc. agr. /Ph.D. in Agricultural Sciences)
to the
Faculty of Agriculture Sciences
Presented by
Muhammad Imran
2012!

!!!!!!!!!!!!!!!!!!!!!!!!!!This thesis was accepted as a doctoral dissertation in fulfillment of the
requirements for the degree “ Doktor der Agrarwissenscheften” (Dr. sc.
Agr. / Ph.D in Agricultural Science) by the Faculty of Agricultural
Sciences of the University of Hohenheim.
Date of Examination: 14.02.2013
Examination Committee
Dean and Head of the Committee: Prof. Dr. R. Mosenthin
Supervisor and Reviewer: Prof. Dr. Günter Neumann
Co- Reviewer: Prof. Dr. Michael Kruse
Co-Examiner: Prof. Dr. Uwe Ludewig

Dedicated
to
My loving mother and father

Eidesstattliche Erklärung
Ich erkläre hiermit an Eides statt, dass ich die vorliegende Dissertation
selbständig angefertigt, nur die angegebenen Quellen und Hilfsmittel
benutzt und inhaltlich oder wörtlich übernommene Stellen als solche
gekennzeichnet habe. Ich boch noch keinen weiteren Promotionsversuch
unternommen.
DECLARATION OF ORIGINALITY
I declare that this thesis is description of my own research and this work
has not previously been submitted for the doctoral degree at any other
institution.
Stuttgart, den Muhammad Imran

Table of Contents Summary……………….…………………………..………….………...……1 Zusammenfassung……………………….…….…………...……………….3 General introduction……………………………….…………………….…..6 Impact of nutrient seed priming on germination, seedling development, nutritional status and grain yield of maize ..............................................................................................................46 Nutrient seed priming improves seedling development and increases grain yield of maize exposed to low root zone temperatures during early growth ..............................................................................................................70 Accumulation and distribution of Zn and Mn in soybean seeds after nutrient seed priming and its contribution to plant growth under Zn and Mn deficient conditions ………………………………………………………………………..……...94 General discussion…………………….………………………………….118 References………………………….……………………………….…….141 Curriculum vitae………………..…………………………………………179 Acknowledgements...…………………………………………………….183

List of Figures
Figure Title Page No.
2.1 Schematic diagram of normal germination and seed priming process.
8
2.2 Time course of major events associated with germination and subsequent post germination growth (Bewley, 1997).
9
2.3 A conceptual diagram depicting relationships between plant and soil temperature for nutrient uptake. Soil temperature affects nutrient uptake directly by altering root growth, morphology and uptake kinetics. Indirect effects include altered rates of decomposition and nutrient mineralization, mineral weathering and nutrient transport processes (mass flow and diffusion).
13
2.4 Schematic diagram of the pore system of the apparent space. DFS= Donan free space, WFS = water free space.
17
2.5 Primary effects of a short chill in the light and the dark on photosynthesis in thermophilic plants.
20
2.6 Effect of high and low seed P reserves on shoot and root biomass accumulation in wheat.
41
3.1 Effect of water priming for different time periods on germination rate (a) and seedling growth (b) of maize. Data points represent means and SE of 4 replicates. Different characters indicate significant differences between treatments.
54
3.2 Shoot (a) and root (b) dry biomass of maize plants, grown in nutrient solution after nutrient seed priming with sequential harvests at 2, 3 and 4 weeks after sowing. Data represent means and SE of 4 replicates at each harvest. Different characters indicate significant differences between treatments.
56
3.3 Nutrient concentrations in shoots of maize plants, grown in nutrient solution after nutrient seed priming with sequential harvests at 2, 3 and 4 weeks after sowing. Data represent means and SE of 4 replicates at each harvest. Different characters indicate significant differences between treatments.
57
3.4 Nutrient concentrations in shoots of maize plants, grown in nutrient solution after nutrient seed priming with sequential harvests at 2, 3 and 4 weeks after sowing. Data represent means and SE of 4 replicates at each harvest. Different characters indicate significant differences between treatments.
58

Figure Title Page No.
3.5 Shoot (a) and root (b) dry biomass of 4 weeks old maize plants grown in soil culture after nutrient seed priming. Data represent means and SE of 4 replicates. Different characters indicate significant differences between treatments.
60
3.6 Minimum and maximum temperature at research farm (°C) recorded in April-May, 2010.
68
4.1 Five weeks old maize plants grown in nutrient solution under low RZT stress.
82
4.2 Roots of maize plants grown along the root windows in rhizo-boxes after 3 weeks of low RZT stress.
83
4.3 Root lengths of maize plant roots grown along the root windows after 1 and 3 weeks of low RZT stress. Data represent means and SE of 4 replicates. Different characters indicate significant differences between treatments.
85
4.4 Minimum temperature in Hohenheim (°C) recorded in April-May, 2010 and 2011.
86
4.5 Shoot fresh and dry biomass of maize plants harvested in the field 5 weeks after sowing (year 2011). Data represent means and SE of 4 replicates. Different characters indicate significant differences between treatments.
88
5.1 (a) Radical emergence of soybean seeds, water primed between layers of moist filter paper for different time duration. Data points represent means of 4 replicates. (b) Abnormal seedlings in the germination test after water priming with different methods and time durations. Bars show means and SE of 4 replicates. Different characters indicate significant differences between treatments.
103
5.2 Soybean seedlings grown for 7 days on peat culture substrate after water priming with different methods and priming durations. AC-Method = volume of priming solution according to water absorption capacity of seeds; FP-Method = priming between moist filter paper; SP-Method = submerged priming.
104
5.3 (a) Shoot and (b) root dry biomass production of 5 weeks old soybean plants grown in nutrient solution. Bars show means and standard errors of four replicates. Significant differences (Duncan's Multiple Range Test, p= 0.05) are indicated by different characters. (NS = Nutrient solution; WP = water priming; NS – Zn = nutrient solution without Zn; NS – Mn = nutrient solution without Mn; NS – (Zn+Mn) = nutrient solution without Zn and Mn).
107
5.4 Shoot Zn and Mn (a) concentrations and (b) contents in 5 weeks old soybean plants grown in nutrient solution. Deficiency thresholds for Zn and Mn are marked by horizontal lines..Bars show means and standard errors of four replicates.
108

Figure Title Page No.
5.5 (a) Shoot and (b) root dry biomass of 5 weeks old soybean plants grown in soil culture. Deficiency thresholds for Zn and Mn are marked by horizontal lines..Bars show means and standard errors of four replicates. Significant differences (Duncan's Multiple Range Test., p= 0.05) are indicated by different characters.
110
5.6 Shoot Zn and Mn (a) concentrations and (b) contents in 5 weeks old soybean plants grown in soil culture. A line across the bars indicates critical deficient levels of Zn and Mn. Deficiency thresholds for Zn and Mn are marked by horizontal lines. Bars show means and standard errors of four replicates. Significant differences (Duncan's Multiple Range Test., p= 0.05) are indicated by different characters.
111
6.1 Effect of B seed priming on 2 weeks old wheat seedlings grown under salt stress (8dSm-1).
128
6.2 Effect of B seed priming on root growth of 4 weeks old wheat seedlings grown under salt stress (8dSm-1).
128
6.3 Effect of B seed priming on shoot and root fresh biomass of 4 weeks old wheat plants grown under salt stress (8 dSm-1). Bars show the means and SE of 4 replicates. Different characters show the significant differences between the treatments.
129
6.4 Effect of nutrient seed priming on stomatal conductance (left) and leaf area (right) of 1st fully developed leaf of 5 weeks old maize plants grown in nutrient solution at low root zone temperature (10°C for water, Fe and Zn). Bars represent the means and SE of 4 replicates. Different characters show the significant differences between the treatments.
133
6.5 Effect of nutrient seed priming on rate of photosynthesis (left) and net photosynthesis (right) in 1st fully developed leaf of 5 weeks old maize plants grown in nutrient solution at low root zone temperature (10°C for water, Fe and Zn). Bars represent the means and SE of 4 replicates. Different characters show the significant differences between the treatments.
133
6.6 Effect of nutrient seed priming on shoot Zn concentration of 4 weeks old wheat plants grown under salt stress (8dSm-1). Bars show the means and SE of 4 replicates. Different characters show the significant differences between the treatments.
134
6.7 Effect of P and micronutrients (Zn, Mn and B) seed priming on shoot growth of 4 weeks old soybean plants.
139

List of table
Table Title Page No.
2.1 Effect of high and low seed P (HSP and LSP) reserves and P fertilization on P accumulation in shoot and whole wheat plant.
41
3.1 Concentrations and salts used for maize nutrient seed priming treatments.
51
3.2 Mineral nutrient contents of maize seeds after nutrient seed priming. Data represent means of 4 replicates. Different characters indicate significant differences between treatments.
55
3.3 Nutrient concentrations and contents in young leaves and the remaining shoot of maize plants grown in soil culture. Data represent means of 4 replicates +SE. Different characters indicate significant differences between treatments in similar shoot portion.
61
3.4 Final grain yield of maize grown (kg ha-1). Data represent means +SE of 4 replicates. Different characters indicate significant difference between treatments.
62
4.1 Seed mineral nutrient concentrations and contents after nutrient seed priming. Data represent means and SE of 4 replicates. Different characters indicate significant differences between treatments.
76
4.2 Seed priming treatments and root zone temperature regimes of maize seedlings grown in nutrient solution and rhizo-boxes.
80
4.3 Fresh shoot and root biomass, and root/shoot ratio of maize plants grown in nutrient solution and rhizo-boxes after nutrient seed priming. Data represent means and SE of 4 replicates. Different characters indicate significant differences between treatments.
81
4.4 Total root lengths (cm plant-1) and root lengths in different diameter lasses of maize plants grown in nutrient solution and rhizo-boxes after nutrient seed priming. Data represent means and SE of 4 replicates. Different characters indicate significant differences between treatments.
84
4.5 Shoot mineral concentrations of maize plants grown in nutrient solution and rhizo-boxes under low RZT stress. Data represent means and SE of 4 replicates. Different characters indicate significant differences between treatments.
87
4.6 Shoot mineral contents of maize plants grown in nutrient solution and rhizo-boxes under low RZT stress. Data represent means and SE of 4 replicates. Different characters indicate significant differences between treatments.
87

Table Title Page No.
4.7 Shoot mineral concentrations and contents of maize plants grown in the field for 5 weeks. Data represent means of 4 replicates and SE Different characters indicate significant differences between treatments.
89
4.8 Final grain yields of maize grown in the field (2010-2011). Data represent means and SE of 4 replicates. Different characters indicate significant differences between treatments.
89
5.1 Zinc and manganese contents in complete soybean seeds after nutrient seed priming. Data represent means and SE of 4 replicates. Different characters indicate significant differences between priming treatments.
100
5.2 Effect of seed priming with different Zn and Mn concentrations on abnormal seedling development. Data represent means and standard errors of four replicates. Significant differences (Duncan's Multiple Range Test, p= 0.05) are indicated by different characters.
103
5.3 Distribution of Zn and Mn in soybean seeds after nutrient seed priming. Data represent means and standard errors of four replicates. Column values followed by different letter are significantly different by Duncan's Multiple Range Test, p= 0.05).
104
6.1 Effect of nutrient seed priming on total phenolic compounds and proline in the roots of 4 weeks old maize plants grown in soil culture at low zone temperature (10°C for water, Fe and Zn+Mn treatments). Data represents the means and SE of 4 replicates. Different chracters show the significant differences between the treatments.
136

Chapter(1((((((((((((((((((((((((((((((((((((((((((((((((((((((((((((((((((((((((((((((((((((((((((((((((((((((((((((((((((((((((((((((((((((((((((((((((((((((Summary(
( 1(
Chapter 1
Summary
Seed reserves are the primary source of mineral nutrients for early
seedling development. ‘Nutrient seed priming’ is a technique in which
seeds are soaked in nutrient solution and subsequently dried back to
initial moisture content for storage. It is an efficient approach to increase
seed nutrient contents along with priming effects to improve seed
quality, germination speed and seedling establishment. Various abiotic
stresses, such as sub-optimal temperature, drought, submergence and
soil pH extremes can seriously affect seedling establishment and
nutrient acquisition at early growth stages. This thesis focused on the
role and contribution of nutrient seed priming in plant growth and
nutritional status in maize and soybean under conditions of limited
nutrient availability and low root-zone temperature.
Protocols for nutrient seed priming with Zn, Mn, Zn+Mn, B and P were
optimised for application in maize and soybean seeds (B and P priming
in maize only). Optimum priming durations of 24 h (maize) and 12 h
(soybean) were identified for both plant species but in instead of
submerging seeds in priming solutions slow imbibition between filter
papers was essential for soybean to minimise development of abnormal
seedlings to avoid imbibition damage. Nutrient concentrations were
calculated according to water uptake to double the natural seed
reserves of the respective micronutrients and 50% increase in
phosphorus. However, final uptake of the micronutrients was generally
much higher (+500-1000%) while it was lower for P (+20%). In case of
soybean this could be attributed to a high Zn and Mn binding capacity of
the seed coat, which adsorbed up to 60% of the primed nutrients.
Particularly, Zn and Zn+Mn priming stimulated plant growth in
hydroponic culture systems and to a lower extent also on a soil with low
availability of P, Zn and Fe. This was associated with a high shoot
translocation of the primed nutrients (Zn and Mn), which was most

Chapter(1((((((((((((((((((((((((((((((((((((((((((((((((((((((((((((((((((((((((((((((((((((((((((((((((((((((((((((((((((((((((((((((((((((((((((((((((((((((Summary(
( 2(
expressed in the hydroponic culture system. Combined priming
treatments with Zn and Mn were usually less effective than Zn priming
alone, suggesting an antagonistic interaction. By contrast, mobility of
primed B was extremely low and B priming was completely ineffective.
In soil culture also P priming moderately increased shoot biomass
production by 10-20 %. However the efficiency of P priming was largely
limited by the high P demand of the plants.
Low root zone temperature (RZT) at early spring is a limiting factor for
maize production in Central and Northern Europe. Nutrient acquisition,
nutrient uptake and particularly root growth are severely affected at low
RZT and the consequences of these growth depressions are often not
completely compensated until final harvest. Model experiments in
hydroponics and soil culture revealed that maize nutrient seed priming
with Zn, Mn and Fe is a promising strategy to diminish the deficiency of
specific nutrients, such as Zn, Mn and also P and to maintain plant
growth under low RZT stress. This was mainly attributed to significantly
increased root growth and particularly fine root production in plants
grown from nutrient-primed seeds. Improved net photosynthesis of
primed plants was mainly related with increased leaf area and
preliminary results suggest a higher tolerance to oxidative damage due
to increased production of protective phenolics.
Two independent field experiments under conditions of suboptimal
temperatures during germination and early growth revealed an increase
in grain yield of 10 – 15 % for plants derived from Zn+Mn and Fe primed
seeds. This finding demonstrates long-lasting persistence of priming
effects. The molecular and physiological mechanisms behind require
further investigation.

Chapter(1((((((((((((((((((((((((((((((((((((((((((((((((((((((((((((((((((((((((((((((((((((((((((((((((((((((((((((((((((((((((((((((((((((((((((((((((((((((Summary(
( 3(
Zusammenfassung
Samenreserven sind die primäre Quelle für die Mineralstoffversorgung
während der frühen Keimlingsentwicklung. Nährstoffpriming ist ein
methodischer Ansatz, bei dem Saatgut in Mineralstofflösungen
vorgequollen und anschließend zur weiteren Lagerung auf Ausgangs-
feuchte zurückgetrocknet wird. Hierdurch ist es möglich, neben
vorquellungsbedingten Primingeffekten auch die Mineralstoffreserven
zur Verbesserung der Saatgutqualität zu erhöhen sowie die Auflauf-
Geschwindigkeit und die Keimlingsentwicklung zu verbessern, die durch
verschiedenste Stressfaktoren, wie suboptimale
Temperaturbedingungen, Trockenheit, Staunässe und pH-Extreme
beeinträchtigt werden kann.
Die vorliegende Arbeit beschäftigt sich mit der Wirkung von
Nährstoffpriming auf das Wachstum und den Ernährungsstatus von
Mais und Soja unter Bedingungen eingeschränkter
Nährstoffverfügbarkeit und niedriger Wurzelraumtemperatur.
Für die Saatgutapplikation von Zink (Zn), Mangan (Mn), Mn+Zn, Bor
(B), Eisen (Fe) und Phosphat (P) bei Mais und Soja wurden
Primingprotokolle entwickelt. Bei beiden Pflanzenarten ergab sich eine
optimale Behandlungsdauer von 24 h. Jedoch war es notwendig, das
Soja-Saatgut anstelle von submerser Inkubation in den Nährlösungen,
langsam in Nährlösungsgetränktem Filterpapier einzuquellen, um die
Entwicklung abnormaler Keimlinge aufgrund von Quellungsschäden zu
vermeiden. Anhand der Wasseraufnahme der Keinlinge wurden die
Nährlösungskonzentrationen so berechnet, das sich nach Einquellen
eine Verdopplung der natürlichen Mineralstoffreserven im Samen
ergeben sollte. Jedoch wurden für die Mikronährstoffe erheblich höhere
Aufnahmeraten gemessen (+500 - 1000%), während die Aufnahme von
P geringer war (+ 20%). Bei Soja konnte gezeigt werden, das die
erhöhten Mikronährstoffgehalte auf eine hohe Bindungskapazität der

Chapter(1((((((((((((((((((((((((((((((((((((((((((((((((((((((((((((((((((((((((((((((((((((((((((((((((((((((((((((((((((((((((((((((((((((((((((((((((((((((Summary(
( 4(
Samenschale zurückzuführen waren, die bis zu 60 % der
aufgenommenen Mineral-stoffe adsorbierte.
Das Pflanzenwachstum wurde besonders durch Zn und Zn+Mn-Priming
stimuliert, sowohl in Nährlösungskultur und in geringerem Umfang auch
auf einem Boden mit geringer P-, Zn-, und Fe-Verfügbarkeit, was mit
einer besonders ausgeprägten Translokation der zugesetzten
Nährstoffe ins Sprossgewebe, besonders in Nährlösungskultur
verbunden war. Dabei zeigten Kombinationsbehandlungen (Zn+Mn)
geringere Effekte als alleiniges Zn-Priming, was auf antagonistische
Interaktionen hinweist. Im Gegensatz dazu ergab sich für B eine
besonders geringe Mobilität innerhalb der Pflanze und entsprechend
war B-Priming wirkungslos. In Bodenkultur wurde auch durch P-Priming
ein moderater Wachstums-effekt (+10-20 % Biomasseproduktion)
erzielt. Die Effizienz des P-Priming wurde jedoch stark durch den hohen
P-Bedarf der Pflanzen eingeschränkt.
Niedrige Wurzelraumtemperaturen (NWT) im zeitigen Frühjahr limitieren
den Maisanbau in Zentral und Nordeuropa. Die Nährstoff-aneignung,
Nährstoffaufnahme und besonders das Wurzelwachstum werden durch
NWT stark beeinträchtigt und die Folgen können häufig bis zur Ernte
nicht mehr kompensiert werden. Modellexperimente in Nährlösungs-,
und Bodenkultur ergaben für Saatgutpriming mit Zn, Mn und Fe
deutliche Hinweise darauf, das bei NWT Mangelerscheinungen
bestimmter Nährstoffe, wie Zn, Mn und auch P vermindert werden
können. Dies konnte in erster Linie auf ein signifikant verbessertes
Wurzelwachstum und besonders auf vermehrte Feinwurzelbildung
zurückgeführt werden. Eine erhöhte Nettophotosyntheserate ergab sich
hauptsächlich aus einer vergrößerten Blattfläche und erste Vor-
untersuchungen weisen auch auf eine höhere Toleranz gegenüber
oxidative Schädigungen durch erhöhte Produktion von Phenolen mit
Schutzfunktion hin.

Chapter(1((((((((((((((((((((((((((((((((((((((((((((((((((((((((((((((((((((((((((((((((((((((((((((((((((((((((((((((((((((((((((((((((((((((((((((((((((((((Summary(
( 5(
Zwei unabhängige Feldversuche bei suboptimaler Temperatur während
der Keimphase und des Jugendwachstums ergaben Kornertrags-
steigerungen von 10-15 % bei Mais nach Saatgutpriming mit Zn+Mn
oder Fe. Diese Ergebnisse belegen die Langzeitwirkung der Priming-
effekte. Die molekularen und physiologischen Grundlagen erfordern
allerdings noch weitere Untersuchungen.

Chapter(2( ((((((((((((((((((((((((((((((((((((((((((((((((((((((((((((((((((((((((((((((((((((((((((((((((((((((((((((((((((((((((((((((((General(Introduction(
( 6(
Chapter 2
General introduction
Early and healthy seedling development is essential for optimal growth and
maximum yield in agricultural crops. Strong and well-developed seedlings can
tolerate several biotic and abiotic stresses during various growth stages.
Healthy and vigorous seedlings are also important to overcome the adverse
environmental effects on crops grown under temperate climate conditions
(Lütke-Entrup and Oehmichen, 2000).
The germination process is the crucial step determining the vigor and success
of the life cycle of a crop plant. It includes the events that start with imbibition
by, water uptake, quiescent dry seeds and ends with the elongation of the
embryonic axis (Bewley and Black, 1994). Accelerated and homogeneous
germination is necessary for better crop establishment, efficient usage of seed
reserves and ultimately increased final yields (Harris, 1996). Different methods
and strategies are used to improve seed quality and germination e.g. use of
biotechnology, hybrid seeds, mechanical seed treatments i.e. polishing off or
removal of the seed coat (testa), sorting the seeds into different seed size
classes and various seed treatments like seed coating, pelleting or soaking in
different solutions (Kolotelo et al., 2001). All these various techniques are
applied to reduce the time between seed sowing and seedling emergence and
also to avoid biotic and abiotic stresses during germination phase. Soaking of
seeds into water, also termed as, seed priming is getting very popular and a
successful method to improve seed germination and crop establishment in
developing countries. It is an easy to use and cost effective technique for
resource poor farmers and farming systems (Harris, 1996).

Chapter(2( ((((((((((((((((((((((((((((((((((((((((((((((((((((((((((((((((((((((((((((((((((((((((((((((((((((((((((((((((((((((((((((((((General(Introduction(
( 7(
2.1 Seed priming
Seed priming is a technique in which seeds are soaked in water for a certain
period of time and subsequently dried back to storage moisture contents
before sowing. Seedling emergence from water-primed seeds is faster and
more vigorous compared to emergence from unprimed seeds (Rashid et al.,
2002; Arif et al., 2005; Miraj, 2005). Soaking of seeds into water stimulates a
variety of biochemical changes in the seed that are essential to initiate the
germination process e.g. water imbibition, breaking of dormancy, activation of
certain enzyme etc. (Ajouri et al., 2004). A number of processes stimulating
germination are activated by seed priming and persist following the re-
desiccation of the seed (Asgedom and Becker, 2001). Therefore, upon sowing,
the primed seeds can rapidly imbibe and restore the seed metabolism,
resulting in an increased germination rate, decreased physiological
germination heterogeneity and better seedling development (Rowse, 1995).
The germination process can be divided into three phases: (i) initial fast
imbibition (ii) starting of metabolic processes in the seed and (iii) subsequent
radicle emergence and resumption of growth. In the process of seed priming,
seeds are soaked in water for the estimated time period to complete the first
two phases of the germination process. Drying the seed after the initiation of
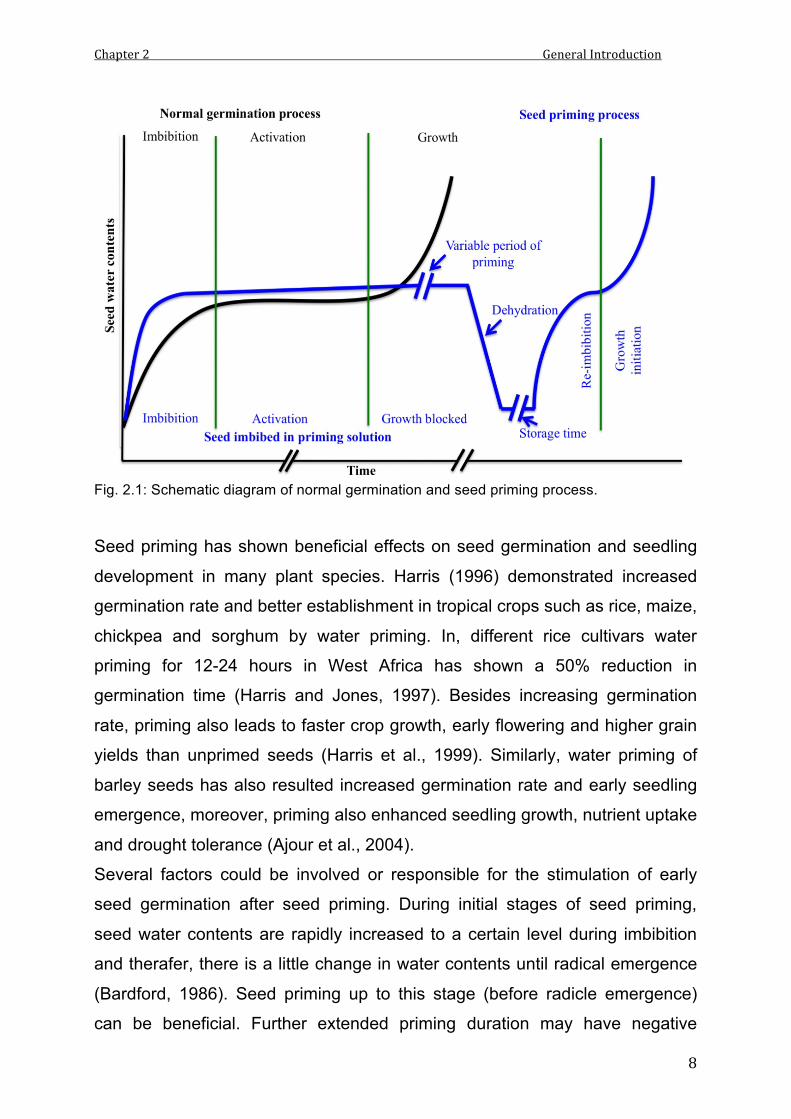
radicle emergence causes damage to seed vigor and viability. Figure 2.1
shows the diagrammatic processes of normal seed germination and seed
priming.
Chapter(2( ((((((((((((((((((((((((((((((((((((((((((((((((((((((((((((((((((((((((((((((((((((((((((((((((((((((((((((((((((((((((((((((((General(Introduction(
( 8(
Fig. 2.1: Schematic diagram of normal germination and seed priming process.
Seed priming has shown beneficial effects on seed germination and seedling
development in many plant species. Harris (1996) demonstrated increased
germination rate and better establishment in tropical crops such as rice, maize,
chickpea and sorghum by water priming. In, different rice cultivars water
priming for 12-24 hours in West Africa has shown a 50% reduction in
germination time (Harris and Jones, 1997). Besides increasing germination
rate, priming also leads to faster crop growth, early flowering and higher grain
yields than unprimed seeds (Harris et al., 1999). Similarly, water priming of
barley seeds has also resulted increased germination rate and early seedling
emergence, moreover, priming also enhanced seedling growth, nutrient uptake
and drought tolerance (Ajour et al., 2004).
Several factors could be involved or responsible for the stimulation of early
seed germination after seed priming. During initial stages of seed priming,
seed water contents are rapidly increased to a certain level during imbibition
and therafer, there is a little change in water contents until radical emergence
(Bardford, 1986). Seed priming up to this stage (before radicle emergence)
can be beneficial. Further extended priming duration may have negative
Chapter(2( ((((((((((((((((((((((((((((((((((((((((((((((((((((((((((((((((((((((((((((((((((((((((((((((((((((((((((((((((((((((((((((((((General(Introduction(
( 9(
effects on seed germination due to death of the seedling on drying (Ajouri et
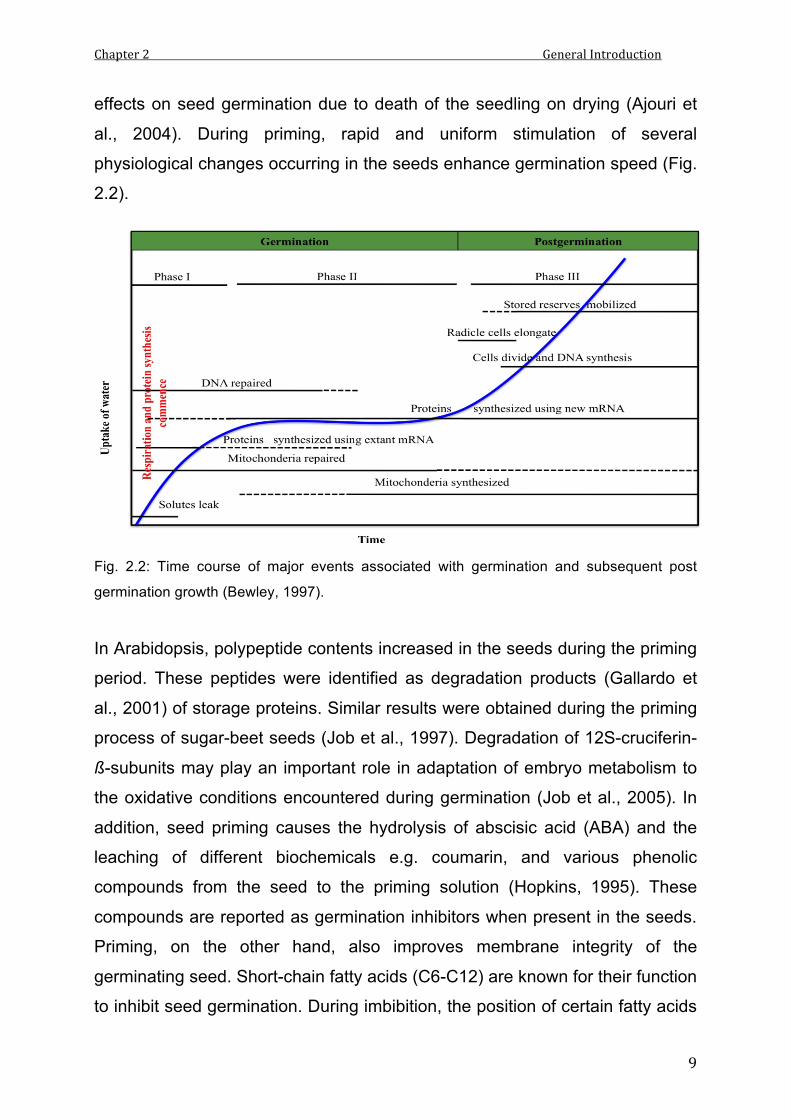
al., 2004). During priming, rapid and uniform stimulation of several
physiological changes occurring in the seeds enhance germination speed (Fig.
2.2).
Fig. 2.2: Time course of major events associated with germination and subsequent post
germination growth (Bewley, 1997).
In Arabidopsis, polypeptide contents increased in the seeds during the priming
period. These peptides were identified as degradation products (Gallardo et
al., 2001) of storage proteins. Similar results were obtained during the priming
process of sugar-beet seeds (Job et al., 1997). Degradation of 12S-cruciferin-
ß-subunits may play an important role in adaptation of embryo metabolism to
the oxidative conditions encountered during germination (Job et al., 2005). In
addition, seed priming causes the hydrolysis of abscisic acid (ABA) and the
leaching of different biochemicals e.g. coumarin, and various phenolic
compounds from the seed to the priming solution (Hopkins, 1995). These
compounds are reported as germination inhibitors when present in the seeds.
Priming, on the other hand, also improves membrane integrity of the
germinating seed. Short-chain fatty acids (C6-C12) are known for their function
to inhibit seed germination. During imbibition, the position of certain fatty acids

Chapter(2( ((((((((((((((((((((((((((((((((((((((((((((((((((((((((((((((((((((((((((((((((((((((((((((((((((((((((((((((((((((((((((((((((General(Introduction(
( 10(
associated with seed dormancy is changed and does not revert upon drying
(Bewley and Black, 1982). Hence, after seed priming, all the necessary
physiological and biochemical changes occurring during the priming process
persist upon drying. Therefore, when a primed seed is sown, it avoids phase
(i) & (ii), as shown above, and a little absorption of water can initiate the radicle
emergence. Instead, an unprimed seed has to go through all phases of seed
germination after sowing, which can be a possible reason for reduced
germination rate and seedling growth compared to primed seeds.
Seed nutrient contents play an important role in germination and early seedling
development. During early establishment stages, seedlings obtain mineral
nutrients partly from seed reserves and partly from soil (Rengel and Graham,
1995). During the germination process, nutrients stored in the seeds are
degraded by enzymes and transported to the growing embryo (Fincher, 1989).
A young seedling passes through various stages in nutrient uptake during
growth, based on the internal and external nutrient supplies and on crop
nutrient requirements (Scaife and Smith, 1973). In the beginning, the plant
utilizes nutrients from the seed reserves; it is the stage when external sources
have a partial effect, and afterwards nutrient uptake by roots is responsible for
plant growth with no further contribution from seed reserves (Whalley et al.
1966). Therefore, sufficient mineral nutrient reserves in the seed are
necessary to maintain seedling growth until the root system starts supplying
the nutrients. This is particularly important for crops grown in soils with low
available nutrients for plant growth (Asher, 1987). In rice, plants from seeds
with high Zn contents were better able to grow under Zn deficient conditions
compared to those grown from low Zn seeds (Rengel and Graham, 1995;
Hacisalihoglu and Kochian, 2003; Tehrani et al., 2003). Previous reports have
shown that wheat seeds with low Mn contents produced less vigorous
seedlings and ultimately resulted in low yields at harvest (Marcar and Graham,
1986; Singh and Bharti, 1985). Delayed germination and seedling
development was also observed in barley and lupin with low Mn seed contents
(Genc et al., 2000; Longnecker et al., 1991; Crosbie et al., 1994). It has been
shown that the percentage of abnormal seedlings increased with decreasing

Chapter(2( ((((((((((((((((((((((((((((((((((((((((((((((((((((((((((((((((((((((((((((((((((((((((((((((((((((((((((((((((((((((((((((((((General(Introduction(
( 11(
level of Boron (B) in soybean seeds (Rerkasem et al., 1993) and in green and
black grams (chickpea) (Rerkasem et al., 1989; Bell et al., 1989). High seed
Phosphorous (P) contents have improved early seedling growth and dry matter
production in clover (Thomson and Bolger, 1993), barley (Zhang et al., 1990),
spring wheat (Bolland and Baker, 1988) and rice (Ros et al., 1997). Seed P
contents also affected the number and mass of the nodules in bean plants
(Teixeria et al., 1999). Zhu and Smith (2001) have shown that wheat plants
grown from high seed P contents were able to accumulate more P in the shoot
compared to plants grown from low P seeds.
Different strategies are used for the continuous supply of micronutrients to the
crops, like direct soil application, foliar sprays and/or seed treatments. Direct
soil application of fertilizers is a widely adopted method but finding a good
quality micronutrient fertilizers and uniform application in the field is a difficult
and cost in-effective method. Foliar sprays of micronutrients have been used
as an effective technique to overcome micronutrient deficiencies in crop plants
(Wilhelm et al., 1988; Savithri et al., 1999) but it is expensive, time consuming
and more laborous (Johnson et al., 2005). Seed treatment with mineral
nutrients, on the other hand, is an easy, cost-effective and less time
consuming method (Robert, 1973; Ros et al., 2000). In case of barley
production in China on calcareous soils, seed soaking in P solution stimulated
seedling growth, and plants used the added P as early as three days after
application (Zhang et al., 1998). Yilmaz et al., (1998) have reported that Zn
seed dressing significantly increased wheat yield on Zn deficient soils.
2.2 Nutrient seed priming ‘Nutrient seed priming’ is a technique in which seeds are soaked in nutrient
solutions instead of water. Nutrient priming is an approach to increase seed
nutrient contents along with priming effects to improve seed quality for better
germination and seedling establishment. Priming seeds in solutions of macro
or micronutrients has been shown to improve yield of rice (Peeran and
Natanasabapathy, 1980), wheat (Khalid and Malik, 1982; Marcar and Graham,
1986; Wilhelm et al., 1988) and forage legumes (Sherrell, 1984), but the

Chapter(2( ((((((((((((((((((((((((((((((((((((((((((((((((((((((((((((((((((((((((((((((((((((((((((((((((((((((((((((((((((((((((((((((((General(Introduction(
( 12(
potential to damage the seed and inhibit germination by priming at high
nutrient concentrations has also been reported (Roberts, 1948; Ajouri et al.,
2004). In barley, nutrient priming of seeds with P and Zn enhanced
germination, seedling growth and uptake of these elements by plants (Ajouri et
al., 2004). Priming of maize seeds with 1% ZnSO4 solution improved plant
growth, final grain yield and increased seed Zn contents in the harvested
grains on soils with limited or deficient Zn supply (Harris et al., 2007). Soybean
seed priming with micronutrients i.e. Zn, Mn and B significantly increased
germination rate and concentrations of primed nutrients in young seedlings
grown under salt stress conditions (Imran, 2006).
2.3 Soil temperature
Soil temperature is an important abiotic factor involved in plant growth, as it
constantly affects other biotic and abiotic components of the soil–plant system
(Pregitzer et al., 2000). Soil temperature has a vital role in determining the soil
moisture contents, rate constants of different chemical reactions as well as
nutrient transport and availability in the soil. Besides that, it also affects plant
physiological processes involved in nutrient uptake, root growth and the soil
microbial activities (Pregitzer and King, 2005). In fact, nearly all processes
taking place in soil, from weathering of primary minerals to release of mineral
nutrients into soil solution, accumulation of soil organic carbon, are strongly
predicted by soil temperature. A number of chemicals, physical and biological
processes involved in the acquisition of mineral nutrients by plants are all
sensitive to temperature. Thus, soil temperature has a large influence on the
plant growth (figure 1.3).
There is a great variation in soil temperature with depth and over time. Air
temperature, irradiation and radiant heat transfer are the key factors
responsible for the large fluctuations in the temperature of surface soil layers
compared to more stable temperature in the deeper soil layers (Marschner,
2011). Root growth is severely affected by soil temperatures (Kasper and
Bland, 1992), and the optimum temperature for better root growth is mainly
under genetic control (Klepper, 1987). Optimum temperature for root growth is

Chapter(2( ((((((((((((((((((((((((((((((((((((((((((((((((((((((((((((((((((((((((((((((((((((((((((((((((((((((((((((((((((((((((((((((((General(Introduction(
( 13(
usually lower than required for shoot growth and also varies among the
species (Sattelmacher et al., 1990c, Cooper, 1973). For example, roots of oats
can grow normally at a temperature as low as 5 °C, but in maize at
temperature below 15 °C root growth already starts declining (Singh, 1998) for
maximum biomass production.
Fig. 2.3: A conceptual diagram depicting relationships between plant and soil temperature
for nutrient uptake. Soil temperature affects nutrient uptake directly by altering root growth, morphology and uptake kinetics. Indirect effects include altered rates of decomposition and nutrient mineralization, mineral weathering and nutrient transport processes (mass flow and diffusion). (Pregitzer and King, 2005).
Low or suboptimal temperature is the temperature at which plant growth is
inhibited. According to Larcher and Baur (1981) low temperature stress can be
defined as “any decline in temperature that can induce reversible or
irreversible functional disturbances or fatal injuries to plant resulting in growth
inhibition”. Therefore, low root zone temperature (RZT) stress can be
associated with decrease in soil temperature responsible for inhibited root
growth, reduced availability and transport of mineral nutrient in the soil for
optimal plant growth.

Chapter(2( ((((((((((((((((((((((((((((((((((((((((((((((((((((((((((((((((((((((((((((((((((((((((((((((((((((((((((((((((((((((((((((((((General(Introduction(
( 14(
2.3.1 Effect of root zone temperature on nutrient transport in the soil
In soil, most of the chemical reactions and nutrient transport take place in soil
solution; therefore, soil temperature directly affects the nutrient uptake in
plants. Soil nutrients are transported to the root surface by diffusion and mass
flow. Viscosity of the soil solution is inversely related to soil temperature,
almost, in a linear fashion (r2 = 0.94) in a wide range of soils worldwide (0 to
~70 °C), from 1.15x10–3 to 0.41x10–3 N s m–2 (Brown and LeMay, 1981).
Increased viscosity of soil water at low temperature is responsible for the
decreased water uptake rate by roots and transport within the plants (Johnson
and Thornley, 1985; Kramer and Boyer, 1995; Wan et al., 2001). This leads to
the reduced transport of nutrients to the roots by mass flow. Similarly, low root
zone temperature reduces the transport of nutrients from sites of high to low
concentration by the process of diffusion (Pregitzer and King, 2005). Hillel
(1998) has discussed the diffusion of gases in the soil atmosphere or solutes
in the soil solution by Fick’s first law:
Jd =−Ds θdc/dx
Where Jd is the flux rate of ion d, Ds is the diffusion coefficient of that ion
accounting for the tortuosity of the particular soil pore space, θ is the fractional
water volume, and dc/dx is the concentration gradient of species d. The
magnitude of the concentration gradient is considered as the main driving
force of the flux rate in the process of diffusion, but temperature has significant
influence on Jd because Ds increases linearly with increasing absolute
temperature and vice versa (Weast, 1978). Therefore, it can be concluded that
nutrient transport by the process of diffusion will be reduced at low root zone
temperature compared to warm soil conditions.
2.3.2 Effect of low soil temperature on root growth and functions
Under low temperature conditions growth of primary and secondary roots is
severely inhibited (Pahlavanian and Silk, 1988; Gladish and Rost, 1993) but
shoot growth is usually more inhibited than root growth, resulting in a high
root/shoot dry weight ratio (Huang et al., 1991). Exposure of plant roots to low

Chapter(2( ((((((((((((((((((((((((((((((((((((((((((((((((((((((((((((((((((((((((((((((((((((((((((((((((((((((((((((((((((((((((((((((((General(Introduction(
( 15(
soil temperatures usually causes chilling-induced drought stress with following
leaf wilting due to reduced root hydraulic conductance, limiting water uptake
(Allen and Ort, 2001, Bloom et al., 2004). Particularly, impaired stomatal
function is observed under low temperature stress since the hydraulic
conductivity of the roots is decreased due to the higher viscosity of water at
low temperature and changes in the physiological activity of roots (Melkonian
et al., 2004). The root physiological activities of chill-sensitive genotypes are
characterized by stronger inhibition of water uptake and root respiration than in
chill-tolerant genotypes when exposed to low temperature (Sowinski and
Maleszewski, 1989).
The rate of root respiration decreases at low soil temperature conditions as the
enzyme activity is remarkably reduced in cold soil (Covey-Crump et al., 2002).
Sink capacity declines under low root zone temperature conditions, which
could be responsible for inhibited root growth at early spring in temperate
climate region and at high altitudes (Alvarez-Uria and Korner, 2007). At low
soil temperatures, extensibility of cell walls is reduced in the extension zone, -
a possible reason for a decreased rate of root elongation under cold stress
(Marschner, 2011). In maize, cell wall extensibility in the root elongation zone
is reduced by 25 % by lowering the temperature from 30 to 15°C (Pritchard et
al., 1990). Root anatomy is also greatly affected by low root zone
temperatures.
Shoot and leaf elongation rate is inhibited at low root zone temperature, even
without disturbing leaf water potential (Milligan and Dale, 1988), most probably
due to an increase in ABA concentration in the leaves (Smith and Dale, 1988).
Atkin et al., (1973) have reported an almost two fold increase in ABA
concentration in the xylem exudates of maize plants when soil temperature
was shifted from 28 to 8°C. Production of cytokinins in the roots and its export
is also depressed at low root zone temperature. In maize plant, there was 15%
decline of cytokinin production at 18°C compared to 28°C (Atkin et al., 1973).
Although, lipid composition differs between cellular membranes (Yoshida and
Uemura, 1986), plant tissues and plant species (Staehelin and Newcomb,
2000), extreme growth conditions also strongly influence their composition in

Chapter(2( ((((((((((((((((((((((((((((((((((((((((((((((((((((((((((((((((((((((((((((((((((((((((((((((((((((((((((((((((((((((((((((((((General(Introduction(
( 16(
different plant parts. For example, clear changes in sterol concentrations in the
leaves under low temperature stress have ben reporte by Westerman and
Roddick (1981). Low temperature causes changes in the membrane lipid
composition (Welti et al., 2002; Penfield, 2008), and phospholipids are
replaced by di-galatosyl-di-acyglycerol (DGDG) and sulphoquinovosyl-di-
acylglycerol (SQDG) in leaves of P deficient plants (Hölzl and Dörmann, 2007;
White and Hammond, 2008). Low root zone temperature also causes changes
of similar nature in the root membranes (Cakmak and Marschner, 1988c).
Usually, the changes in lipid composition are considered for the adaption of a
plant to growing conditions. Plants growing at low soil temperature have
changes in their membranes; have more phospholipids with charged head
groups and low saturated shorter fatty acid chains, and higher sterol content
compared with plants growing at higher temperatures (Staehelin and
Newcomb, 2000; Wallis and Browse, 2002; Penfield, 2008). These kinds of
changes are necessary for the alteration of the freezing point (i.e. the transition
temperature) of membranes to a lower temperature and could be important for
the maintenance of membrane functions at low root zone temperatures
(Marschner, 2011).
2.3.3 Effect of low root zone temperature on Nutrient uptake
In the plant cell walls the ‘apparent free space’ consists of (i) the ‘water free
space’ (WFS) available to individual ions, charged and uncharged molecules,
and (ii) the ‘Donnan free space’ (DFS) for cation exchange and anion repulsion
(Fig 1.4). The term ‘apparent free space’ (AFS) was introduced by Hope and
Stevens (1952).

Chapter(2( ((((((((((((((((((((((((((((((((((((((((((((((((((((((((((((((((((((((((((((((((((((((((((((((((((((((((((((((((((((((((((((((((General(Introduction(
( 17(
Fig. 2.4: Schematic diagram of the pore system of the apparent space in plant cell walls.
DFS= Donan free space, WFS = water free space. (Marschner, 2012)
Temperature has a little effect on physical processes e.g. exchange adsorption
of cations in the AFS (Q10 = 1.1–1.2, where Q10 refers to the change in a
reaction or process forced by a change in temperature by 10 °C) (Marschner,
2011). On the other hand, temperature has much greater effect on chemical
and biochemical reactions with Q10 = 2 and Q10 > 2, respectively. In the
physiological temperature range, the value of Q10 often raises beyond 2 for the
uptake of ions from low concentration solutions (Clarkson et al., 1988; Wang et
al., 1993). In maize, it has been observed that the process of ion uptake is
more severely inhibited by low temperature than respiration, especially at
temperatures below 10°C (Bravo and Uribe, 1981).
Soil temperature affects ion uptake in plants in two different ways: (i) the short-
term effects of a sudden change in root zone temperature, which take place
instantly and show the direct effects of temperature on the ion uptake system,
and (ii) the long-term effects, where plants are exposed to particular root
temperature for a longer growth period. The long term effects of low
temperature can be observed after several days or weeks of growth at low root

Chapter(2( ((((((((((((((((((((((((((((((((((((((((((((((((((((((((((((((((((((((((((((((((((((((((((((((((((((((((((((((((((((((((((((((((General(Introduction(
( 18(
zone temperature and involve adaptive responses e.g. changes in the
properties of root membranes. Low root zone temperature influences cell
division and cell elongation in the shoots (Marschner, 1995).
Effect of low root zone temperature on nutrient uptake differs between
nutrients. For example, P uptake at low root zone temperature is drastically
reduced as compared to other nutrients (Engels and Marschner, 1992a;
Engels, 1993). In cold-sensitive plants at low root zone temperature, uptake
rate of nitrate is strongly reduced compared to ammonium (Tachibana, 1987),
but in cold-tolerant species, such as barley and ryegrass, low root zone
temperature does not affect the strong preference for ammonium compared
with nitrate uptake (Macduff and Jackson, 1991; Clarkson et al., 1992).
Potassium uptake is more affected by low root zone temperature as compared
to Ca and Mg.
At low root zone temperature, Zn uptake and translocation from root to the
shoot is usually more depressed than growth, resulting in lower Zn
concentration in the shoot compared to roots (Ellis et al., 1964; Sharma et al.,
1968; Schwartz et al., 1987). Higher supply of P fertilizers at low root zone
temperature may also enhance the Zn deficiency (Martin et al., 1965).
Similarly, Mn concentration in the shoot is also decreased at low root zone
temperature (Moraghan, 1985; Moraghan et al., 1986). Furthermore, Fe
uptake and translocation rate was decreased with decreasing root zone
temperature in tomato plants (Riekels and Lingle, 1966).
The roots of soil-grown plants have to explore for many immobile soil nutrients
(Engels and Marschner, 1990; Rengel, 2001; Lynch, 2007; White and
Hammond, 2008; White and Broadley, 2009). In plants grown at low root zone
temperature, therefore, nutrient uptake is not only affected by low nutrient
availability but also by detrimental effects on root growth rate, root system
morphology and functionality.
2.3.4 Effect of low temperature on plant growth
Low temperature affects a number of metabolic (physiological and
biochemical) and structural (anatomical and morphological) changes in the

Chapter(2( ((((((((((((((((((((((((((((((((((((((((((((((((((((((((((((((((((((((((((((((((((((((((((((((((((((((((((((((((((((((((((((((((General(Introduction(
( 19(
plants (Cooper, 1973; Levitt, 1980; Pollock and Eagles, 1988). Effects of low
temperature on physiological and biochemical processes may include:
reactions in the rate of cell division, photosynthesis and assimilate transport,
rate of respiration, synthesis and translocation of hormones, lipid synthesis,
production and activities of proteins and enzymes (Berry and Raison, 1981;
Graham and Patterson, 1982; Long and Woodward, 1988; Howarth and
Ougham, 1993, Engles and Marschner, 1992). Structural changes may affect
branching of roots, elongation, and cell wall properties (Cooper, 1973; Bowen,
1991; Marschner 1995 and 2012; McMichael and Burke, 1998; Rapacz, 1998;
Equiza et al., 2001). Under the conditions of low temperature stress, the plant
nutrient demand is also altered by effects on root to shoot ratio and other
biochemical processes (Clarkson et al., 1988).
The sensitivity of different plant growth processes to low temperature is
variable. Some are more sensitive to low temperature than the others, and the
sensitivity of the same process varies between and even within plant species
(Miedema, 1982; Paul et al., 1990). For example, in sunflower, leaf expansion
is more severely inhibited than cell division by low temperature (Granier and
Tardied, 1998). There was a significant decrease in shoot dry matter
production in barley plants grown at 5°C due to reduced water uptake rather
than to lower photosynthesis (Sharratt, 1991). Low root zone temperature
causes chill-induced drought stress with subsequent leaf wilting due to
inhibited root hydraulic conductance limiting water uptake (Allen and Ort, 2001;
Bloom et al., 2004, Melkonian et al., 2004).
Photosynthetic capacity and efficiency is decreased in plants grown at
suboptimal growth temperature (Nie et al., 1992), particularly, in chilling
sensitive genotypes (Fracheboud et al., 1999). The reduced photosynthetic
activity of the leaves developed under low temperature stress does not resume
to its optimal level, even after shifting plants to normal growth temperature
(Haldimann, 1996; Nie et al., 1992), which indicates structural changes of the
photosynthetic machinery. In contrast to photosynthetic activity, chlorophyll
content of the cold-stressed leaves recovers immediately after shifting to
optimal temperature but chlorophyll pigment composition analyses have

Chapter(2( ((((((((((((((((((((((((((((((((((((((((((((((((((((((((((((((((((((((((((((((((((((((((((((((((((((((((((((((((((((((((((((((((General(Introduction(
( 20(
shown a great reduction in the chlorophyll a/b ratio at suboptimal growth
temperature, especially in cold-tolerant species (Haldimann et al., 1995;
Haldimann, 1998). Effects of chilling on photosynthesis are illustrated in fig.
2.5.
Fig. 2.5: Primary effects of a short-term chilling periods in the light and the dark on
photosynthesis in thermophilic plants. (Allen and Ort, 2011)
The ratio of LHC-II (light-harvesting (or antenna) complex) to the reaction
centre (RC, a complex of several proteins, pigments and other co-factors
assembled together to execute the primary energy conversion reactions of
photosynthesis) increases at suboptimal growth temperature (Caffarri et al.,
2005). Furthermore, the number of minor antenna (predominantly contains
chlorophyll b, xanthophylls, and carotenoids) complexes decreased in
comparison to LHCII when maize seedlings were grown at suboptimal
compared to optimal temperature but to a lesser extent than RCII (Leipner and
Stamp, 2009).
Particularly, when seedlings are grown at low temperature conditions, plastid-

Chapter(2( ((((((((((((((((((((((((((((((((((((((((((((((((((((((((((((((((((((((((((((((((((((((((((((((((((((((((((((((((((((((((((((((((General(Introduction(
( 21(
encoded proteins of the RC fail to accumulate in the mesophyll thylakoids (Nie
and Baker, 1991; Robertson et al., 1993). A typical ultra structure of the
mesophyll chloroplasts reflects the reduced accumulation of certain thylakoid
proteins. Mesophyll chloroplasts in leaves developed at chilling temperature
are a little bigger and more round in shape (Kutík et al., 2004; Robertson et al.,
1993). Low temperature causes disruption in the granal arry (mesophyll
chloroplast) due to extensive vesiculation, particularly in maize genotypes
sensitive to chilling (Pinhero et al., 1999). Furthermore, growth at low
temperature disturbs the fatty acid composition of the chloroplast membranes
by increasing the unsaturated fatty acids (Fracheboud, 1999).
Regarding the activity of enzymes from the C4 cycle and the Calvin cycle,
contents of rubisco and phosphoenolpyruvate carboxylase were reduced in
leaves that developed at suboptimal temperature, whereas the NADP malate
dehydrogenase activity increased compared to leaves grown at optimal
temperature (Kingston-Smith et al., 1999). Chilling stress significantly reduces
the amount of Rubisco in chilling-sensitive plant compared to chilling-tolerant
plants (Pietrini et al., 1999). However, an increase in the activation state of
rubisco may compensate the decline in its activity caused by chilling stress
(Kingston-Smith et al., 1999).
In the C4 photosynthetic system, assimilates are transported between the
mesophyll and the bundle sheath cells. Low temperature stress, in the leaves,
causes the development of a larger area between mesophyll cells and the
bundle sheath and increases the number of plasmodesmata between these
cell types (Sowinski et al., 2003), particularly in cold- tolerant genotypes. This
increase in plasmodesmata density plays an important role in the efficient
exchange of photosynthetic metabolites as well as export of sucrose at low
temperatures responsible for reduced diffusion rate from cell to cell (Sowinski
et al., 2003).
Older or fully developed leaves are less sensitive to chilling than newly
developing leaves (Leipner and Stamp, 2009). The shoot apex is very
sensitive to low root zone temperature; furthermore, low root zone temperature
also adversely affects growth rate and leaf expansion (Engels, 1994; Stone et

Chapter(2( ((((((((((((((((((((((((((((((((((((((((((((((((((((((((((((((((((((((((((((((((((((((((((((((((((((((((((((((((((((((((((((((((General(Introduction(
( 22(
al., 1999). Photosynthetic performance of maize seedlings is strongly inhibited
at low soil temperature, especially in chill-sensitive genotypes (Chassot, 2000;
Hund, 2003), that could be due to delay in leaf development (Sowinski et al.,
2005). Cold-induced growth inhibition of the leaves is caused by a strong
prolongation of cell cycle and by a reduction of cell production (Rymen et al.,
2007). Spatial analysis of maize leaf blades, exposed to low temperature
stress (6°C, 400 µmol m-2 s-1) for four days, showed two zones of chlorosis.
These cholrotic areas were tracked on the leaf meristem and the leaf surface
which were just exposed to light (Leipner and Stamp, 2009). Therefore, it is
concluded that (i) chilling induced damages during cell production limits the
development of fully functional cells, and (ii) the combined effect of high light
intensity and low temperature leads to the impaired development of
chloroplast.
2.4 Functions of Mineral nutrients in plants
2.4.1 Zinc (Zn) in plants
2.4.1 .1 Zinc uptake and transport in plants
Zinc is an essential micronutrient vital for plant growth and development.
Under normal conditions, zinc is taken up as a divalent cation (Zn2+) whereas
uptake in the monovalent form (ZnOH+) is preferred under high pH conditions.
During long-distance transport in xylem, Zn is either bound to organic acids or
may be translocated as free divalent cations (Marschner, 2012). Zinc
concentrations are fairly high in phloem sap, most probably due to its binding
to low-molecular-weight organic solutes (Kochian, 1991). Zinc does not take
part in redox reactions as it exists only as ZnII in plants and other biological
systems. The metabolic functions of Zn depend on its strong tendency to form
tetrahedral complexes with (nitrogen anions) N-, (oxygen anions) O-, and
particularly Sulfur S-ligands for its catalytic and structural role in enzymatic
reactions (Marschner, 1995).
The functions of Zn and its deficiency symptoms in plants have been

Chapter(2( ((((((((((((((((((((((((((((((((((((((((((((((((((((((((((((((((((((((((((((((((((((((((((((((((((((((((((((((((((((((((((((((((General(Introduction(
( 23(
comprehensively discussed and reviewed (Thorne, 1957; Cakmak and
Marschner, 1988; Welch and Norvell, 1993). Zinc has a critical role in
maintaining plasma membrane structure and its proper functioning in plant
cells (Welch and Norvell, 1993). Cakmak (2000) has discussed importance of
Zn in protecting leaf tissues from oxidative stress.
2.4.1 .2 Morphological effects of Zn deficiency in plants
In dicotyledonous plant species, Zn deficiency causes reduction in leaf size,
shortening of internodal distance leading to stunted plant growth. In oil seed
rape, total number and size of leaves was depressed by Zn deficiency (Hedley
et al., 1982a, b). In dicots generally, Zn deficiency causes intervenial chlorosis
of young leaves starting at tips and margins of leaves, followed by
development of irregular intervenial chlorotic spots (Roberth and Francis,
1983), associated with reduced plant growth and decreased dry biomass
production (Hewitt, 1984). Zinc deficiency symptoms may vary depending on
the plant species, age and growing conditions (Snowball and Robson, 1983;
Grundon et al., 1987).
In monocots, Zn deficiency causes short internodes, which results in stunted
plant growth, delayed flowering and maturity (Grundon et al., 1987). In cereals,
like in wheat, Zn deficiency causes reduced shoot growth and development of
whitish-brown necrotic patches on older leaves while young leaves becomes
yellowish green in colour but now showing necrotic lesions (Cakmak, 1996a).
Chlorosis and necrosis in older leaves of Zn-deficient plants could be
secondary effects due to P or B toxicity, or by photo-oxidation because of
impaired translocation of photosynthates (Cakmak, 2006). Generally, under Zn
deficiency, both, in mono and dicots, shoot growth is more inhibited compared
to root growth (Zahng et al., 1991a), which may be a consequence of
increased translocation of photosynthates to roots due to a shift in sink
location from developing shoot tissues to growing roots (Cumbus, 1985;
Cakmak, 1996b). Abiotic stress factors such as low soil water contents and
low soil temperature have been reported to increase in Zn deficiency

Chapter(2( ((((((((((((((((((((((((((((((((((((((((((((((((((((((((((((((((((((((((((((((((((((((((((((((((((((((((((((((((((((((((((((((((General(Introduction(
( 24(
symptoms (Lindsay, 1972). Under conditions of reduced soil moisture
contents, Zn uptake is limited due to reduced diffusion rate of Zn to the root
surface (Marschner, 1993). Low soil temperature impairs root growth, root
activity and reduces Zn translocation from root to the shoot (Moraghan and
Mascagani, 1991; Engels and Marschner, 1992). Moreover, high light intensity
also increases the severity of Zn deficiency symptoms in leaves (Cakmak et
al., 2000).
2.4.1 .3 Physiological and biochemical effects of Zn
Zn is involved in a vast range of physiological and biochemical processes in
plants. It is associated with metabolism of carbohydrates, proteins, enzyme
activities, structural and functional integrity of cell membranes (Römheld and
Marschner, 1991; Cakmak, 2000; Marschner, 2011).
i. Role of Zn in enzymatic functions
Zinc is the only metal, with functions in all six enzyme classes including oxido-
reductases, transferases, hydrolases, lyases, isomerases and ligases, in
biological systems (Sousa et al., 2009). Four different functions of Zn have
been identified in these enzymes: (i) catalytic, (ii) structural, (iii) co-catalytic
and (iv) protein synthesis in determining the biological activity of various
enzymes (Marschner, 2012). The involvement of Zn in structural and functional
activities of such a wide range of enzymes suggests that Zn deficiency may
inhibit the processes involved in the metabolism of carbohydrate, protein,
auxin and reproduction (Römheld and Marschner, 1991; Brown et al., 1993).
The role of Zn in function of some of the key enzymes is reviewed below.
Alcohal dehydrogenase
Alcohol dehydrogenase (ADH) is an exceptional Zn enzyme with 2 atoms per
molecule, one with catalytic and the other with structural functions, while
mostly Zn enzymes have only 1 Zn atom in their molecules (Colman, 1992;

Chapter(2( ((((((((((((((((((((((((((((((((((((((((((((((((((((((((((((((((((((((((((((((((((((((((((((((((((((((((((((((((((((((((((((((((General(Introduction(
( 25(
Auld and Bergman, 2008). ADH is one of the two proteins responsible for the
reduction of cytotoxic acetylaldehyde to ethenol in the ethanolic fermentation
pathway, resulting in continuous regeneration of NAD+ in the cytoplasm
(Chung and Ferl, 1999). Thus, induction of ADH can improve plant growth
under submerged conditions (Johnson et al., 1994). Proper root functions
could be impaired as a consequence of reduced ADH activity due to Zn
deficiency (Moore and Patrick, 1988).
Carbonic anhydrase
Carbonic anhydrase (CA) is a zinc-containing metallo-enzyme that catalyzes
the interconversion of CO2 and HCO3- in many organisms. In plants, both,
chloroplasts and cytoplasm contain CA, which catalyses the reversible
hydration of CO2 :
CO2 + H2O → HCO3- + H+
In C3 plants, plant Zn nutritional status does not have a direct relationship
between CA activity and photosynthetic CO2 assimilation (Atkin et al., 1972;
Brown et al., 1993). CA activity is completely inhibited under extreme Zn
deficiency, but maximum photosynthetic rate may be attained even when CA
activity is low. However, in C4 plants, it is evident that high CA activity is
required in the mesophyll chloroplasts to convert CO2 to bicarbonate (HCO3−).
Consequently, HCO3− is fixed via phosphoenolpyruvate carboxylase (PEPC)
into C4 compounds (e.g. malate) for the shuttle into the bundle sheath
chloroplasts for decarboxylation (Kanai and Edwards, 1999). Hence, Zn
deficiency may impair the photosynthetic rate in C4 plant as compared to C3
plants (Burnell et al., 1990).
Superoxide dismutase (SOD)
The most abundant superoxide dismutase enzyme found in plant cells is CuZn
superoxide dismutase (CuZnSOD); where Cu (copper) is the catalytic metal
component while Zn is part of the structural unit. Localization and functions of

Chapter(2( ((((((((((((((((((((((((((((((((((((((((((((((((((((((((((((((((((((((((((((((((((((((((((((((((((((((((((((((((((((((((((((((((General(Introduction(
( 26(
CuZnSOD have been discussed in details by Cakmak (2000). In different plant
species, under Zn deficiency, CuZnSOD activity is significantly reduced
(Marschner, 1995), and enzyme activity is immediately restored after Zn re-
supply (Cakmak and Marschner, 1993). Yu and Rengel (1999) have reported
that Zn deficiency in lupin decreased the total SOD and CuZnSOD activity, but
MnSOD activity was unaffected. Under Zn deficiency, decreased SOD activity
is responsible for the increase in the rate O2− generation and leads to the
membrane lipids peroxidation and increase membrane permeability (Cakmak
and Marschner, 1988; Cakmak and Marschner, 1993). Hence, abiotic stress
tolerance in transgenic plants can be increased by the over expression of
CuZnSOD (Cakmak, 2010; Kim et al., 2010).
ii. Role of Zn in carbohydrate metabolism
A number of Zn depending enzymes are involved in the carbohydrate
metabolism in plants at various levels (Marschner, 1995). The association of
Zn in carbohydrate metabolism is linked to its role in photosynthesis and sugar
transformation. In different plant species, net photosynthesis could be
depressed up to 70%, depending on the level of Zn deficiency (Brown et al.,
1993). Under Zn deficiency, activity of fructose-1,6-bisphospahte declines
quite rapidly, while the activities of other enzymes is effected to lesser extent
(Marschner, 2011). Sugars and starch often accumulate in the leaves of Zn
deficient plants (Sharma et al., 1982). Cakmak et al., (1989) found that re-
supply of Zn could restore the phloem loading of sucrose. Most probably, the
restoration of phloem loading of sucrose by Zn supply can be attributed to the
role of Zn in the structural integrity of bio-membranes (Welch et al., 1982).
iii. Role of Zinc in protein metabolism
Zinc is an integral component of the ribosomes and for their structural integrity.
Protein synthesis is dramatically impaired in Zn deficient plants but the
composition is almost unchanged (Obata et al., 1999). This decrease in

Chapter(2( ((((((((((((((((((((((((((((((((((((((((((((((((((((((((((((((((((((((((((((((((((((((((((((((((((((((((((((((((((((((((((((((((General(Introduction(
( 27(
protein synthesis can be associated to increase in concentration of free amino
acids under Zn deficiency, as the concentration of free amino acids rapidly
decreases after Zn resupply (Cakmak, 1989). Under Zn deficiency, level of
free RNA and ribosomes is depressed as a result of reduced transcription
(Kitagishi and Obata, 1986; Kitagishi et al., 1987). RNase activity is
significantly increased under zinc deficiency (Sharma et al., 1982). There is an
inverse relationship between Zn supply and RNase activity, and also between
RNase activity and protein content (Johnson and Simons, 1979). Zn is
essential for RNA polymerase activity and prevents ribosomal degradation by
ribonuclease ( Fachuk et al., 1978; Jendrisk and Burgess, 1975). Under Zn
deficiency activity of RNase increase, resulting in stunted plant growth and
changes in leafe anatomy (Dwivedi and Takkar, 1974).
2.4.1.4 Zinc in membrane integrity
Zinc has a critical role in the maintaining integrity and functions of the bio-
membranes (Bettger and O’Dell, 1981; Pinton et al., 1994). Zn deficiency in
plant cells causes damages in plasma membrane structure and increases the
permeability of inorganic ions (Welch et al., 1982). Role of Zn in membrane
stability is not coupled with the function of Ca, as increased external supply of
Ca could not reduce the increased membrane permeability caused by Zn
deficiency (Cakmak and Marschner, 1988c). Zinc protects membrane lipids
and proteins from oxidative damages by binding to phospholipids and
sulfhydryl (Powel, 2000) or form tetrahedral complexes with cysteine residues
of polypeptide chains (Cakmak and Marschner, 1988c). Zinc interference with
NADPH oxidation and also as a metal component of CuZnSOD controls the
generation of toxic oxygen radicals and prevents the membrane damages
against reactive oxygen species (ROS) (Cakmak and Marschner, 1988a,b;
Cakmak, 2000). Redox status of the reactive –SH (sulfhydryl) groups present
in the ion channel proteins controls the activity of the cannel (Cakmak, 2000).
In isolated vacuoles from sugar beet, a shortly induced Zn deficiency strongly
reduced the activity of Ca-activated slow vacuole –type anion channels

Chapter(2( ((((((((((((((((((((((((((((((((((((((((((((((((((((((((((((((((((((((((((((((((((((((((((((((((((((((((((((((((((((((((((((((((General(Introduction(
( 28(
(Hedrich and Kurkdjian, 1988).
2.4.1.5 Zinc localization in seeds
Zinc, in seeds and grains, is mostly localized in protein bodies in the form of
special particles, the globoid crystals (Lott and Buttrose, 1978; Welch, 1986).
Phytate (salts of phytic acid) is the main constituent of these globoids. In
wheat seeds, Zn is highly concentrated (600 µgg-1dry weight) in the scutellum
(Mazzolini et al., 1985). Mostly, Zn, Fe and proteins are co-localized in the
seed tissues (Cakmak et al., 2010b) and concentrations of Zn, Fe and proteins
are highly correlated to each other in seeds of a number of germplasms
(Peterson et al., 1986; Morgounov et al., 2007; Zhao et al., 2009; Cakmak et
al., 2010b). It could be suggested that proteins act as a sink for Zn and Fe.
Phytic acid has a high affinity to bind divalent cations such as Zn, due to the
negative charged phosphate residues, and form insoluble or unavailable Zn-
phytate complexes in the seeds (Lönnerdal, 2002; Schlemmer et al., 2009)
causing major problems by reducing the bioavailability of Zn for monogastric
animals and humans.
2.4.2 Iron (Fe) in plants
2.4.2.1 Iron uptake in plants
Iron (Fe) is one of the most abundant metal elements present on earth.
Dominant forms of soluble Fe present in soils and nutrient solutions are
chelates of Fe (III) and Fe (II) (Marschner, 2012). Under alkaline conditions,
the presence of Fe ions in the soil solution is severely reduced due to the
formation of Fe hydroxides, oxyhydroxides and oxides (Lemanceau et al.,
2007). Depending on the plant species (strategies I and II), uptake of Fe (II) is
favored as compared to Fe (III) in strategy I plants. There is a predominance
of Fe (III) complexes in the long distance transport in xylem. Iron as a
transition element, easily changes its oxidation state and also forms octahedral
complexes with various ligands. Redox potential of Fe (II or III) widely depends

Chapter(2( ((((((((((((((((((((((((((((((((((((((((((((((((((((((((((((((((((((((((((((((((((((((((((((((((((((((((((((((((((((((((((((((((General(Introduction(
( 29(
on the complexing ligands. Due to this character, Fe plays a significant role in
biological redox systems. Presence of low molecular weight Fe chelates and
free iron (Fe3+ or Fe2+) in aerobic systems produces reactive oxygen species
(ROS), such as superoxide and hydroxide radicals and other related
compounds (Halliwell and Gutteridge, 1986; Halliwell, 2009). These extremely
lethal radicals are responsible for membrane lipid and protein peroxidation.
Binding or incorporation of Fe into structures (e.g., heme and non-heme
proteins) is necessary for controlled reversible oxidation–reduction reactions,
to prevent cell damages from ROS.
2.4.2.2 Iron in plant proteins.
Cytochromes are well known heme proteins containing heme Fe-porphyrin
comlex as a prosthetic group. Cytochromes are essential compenents of the
redox systems in chloroplasts (Hil and Scarisbrick, 1951; Hil 1954), in the
electron transport in the respiratory chain of mitochondria and also involved in
the redox chain in nitrate reductase (Cava et al., 2008).
Catalases and peroxidases are other heme enzymes; the activity of these
enzymes declines under the low Fe supply (Marsh et al., 1963; Chouliaras et
al., 2005). Catalases play an important role in the detoxification of
photorespiratory Hydrogen peroxide (H2O2) to water and CO2 (Kendall et al.,
1983; Willekens et al., 1997; Corpas, Barroso and del Río, 2001;
Vandenabeele et al., 2004). Additionaly catalases also have other essential
functions in plant tissues at different developmental stages: catalases take part
in the signal transduction pathways of various stress responses (Hu et al.,
2010; Fukamatsu, Yabe and Hasunuma 2003; Verslues et al., 2007).
Peroxidases exists in the plants in various forms (Shanon, 1948). They mainly
catalyse the following reactions:
XH2 +H2O2 →X+2H2O
and
XH+XH+H2O2 → X---X+2H2O

Chapter(2( ((((((((((((((((((((((((((((((((((((((((((((((((((((((((((((((((((((((((((((((((((((((((((((((((((((((((((((((((((((((((((((((((General(Introduction(
( 30(
For example, scavenging of H2O2 in chloroplasts catalysed by ascorbate
peroxidase (Foyer and Halliwell, 1976; 1977) represents the first type of
reaction, while the polymerization of phenols to lignin is catalysed by cell-wall
bound peroxidase is related to the second type. Impaired peroxidase activity
may alter cell wall formation, as the presence of peroxidase in the cell walls of
the epidermis (Hendricks and Van Loon, 1990) and rhizodermis (Codignola et
al., 1989) is vital for the biosynthesis of lignin and suberin. Phenolic
compounds and H2O2 are required as substrates for the synthesis of both
compunds.
Peroxidase activity is severely reduced in Fe-deficient roots (Sijmons et al.,
1985; Ranieri et al., 2001). This leads to an increase in the production of H2O2
and accumulation of phenolic compounds in the root cell (Ranieri et al., 2001;
Römheld and Marschner, 1981a). It is reported that phenolics are also
released at higher rates from the roots of Fe-deficient compared with Fe-
sufficient plants (Hether et al., 1984; Marschner et al., 1986a; Jin et al., 2007).
It has been proposed that release of high amounts of phenolics contribute to
the utilization and remobilization of apoplastic Fe from the roots (Jin et al.,
2007).
Ferredoxin is an example of non-heme Fe-S proteins; these proteins are
characterized by the presence of Fe-S clusters containing sulfide-linked di-, tri-
, and tetra Fe centers in variable oxidation states. In plants, ferredoxins
function mainly in the process of photosynthesis; they transmit electrons from
photoreduced Photosystem I to ferredoxin NADP+ reductase, where NADPH is
generated for CO2 assimilation. Moreover, ferredoxins also act as electron
transmitters in various ferredoxin-dependent enzymes for the assimilation of
inorganic N, S, N2 and also regulate the CO2 assimilation cycle (Fukuyama,
2004). Iron-superoxide dismutase (Fe SOD) is another example of Fe-S
proteins. Superoxide dismutases degrade superoxide anion free radicals (O2-)
by formation of H2O2 and may contain Cu, Zn, Mn or Fe as metal components
(Fridovich, 1983; Sevilla et al., 1984). FeSOD is mainly present in the
chloroplast (Kwiatowsky et al., 1985) and could be also present in the

Chapter(2( ((((((((((((((((((((((((((((((((((((((((((((((((((((((((((((((((((((((((((((((((((((((((((((((((((((((((((((((((((((((((((((((((General(Introduction(
( 31(
mitochondria and peroxisomes in the cytoplasm (Droillard and Paulin, 1990).
In Fe-deficient plants reduced activity of FeSOD (Iturbe-Ormaetxe, 1995)
causes increase in the activity of CuZnSOD, which increases H2O2 production
(Tewari et al., 2005).
There are also a number of other enzymes where Fe takes part as a metal
component in the redox reactions or as bridging element between enzyme and
substrate (Marschner, 2012). Reduced activity of these enzymes due to Fe
deficiency in plants leads to vital changes in the metabolic processes.
In the process of ethylene biosynthesis, methionine acts as a primary
precursor. Iron(II) catalyses the two step 1 oxidation, during the biosynthetic
pathway in the conversion of 1-aminocyclopropane-1-carboxylic acid (ACC) to
ethylene. In Fe-deficient cells, formation of ethylene is strongly inhibited, but
immediately restored after resupply of Fe, involving protein synthesis
(Bouzayen et al., 1991).
Lipoxygenases enzymes contain a single Fe atom per molecule (Hildebrand,
1989), catalyse the peroxidation of linolic and linolenic acid, i.e. of long chain
polyunsaturated fatty acids. Hence, high lipo-oxygenase activity is essential for
fast growing plant tissues and organs, and could be vital for membrane
stability.
2.4.2.3 Iron in chloroplast Development and Photosynthesis
Size of the chloroplast and protein contents per chloroplast are severely
affected by Fe deficiency in plants as compared to leaf growth, cell number per
unit area, or number of chloroplasts per cell (Terry, 1988). Fe status of the leaf
cells has a vital role in determining the number of ribsomes and protein
synthesis (Lin and Stocking, 1978). Iron deficiency in maize leaves, causes a
25% reduction in total protein contents, while chloroplast production was
decreased by 82% (Perur et al., 1961), as there is a high Fe requirement of
chloroplastic mRNA and rRNA (Spiller et al., 1987). Iron is required for RNA

Chapter(2( ((((((((((((((((((((((((((((((((((((((((((((((((((((((((((((((((((((((((((((((((((((((((((((((((((((((((((((((((((((((((((((((((General(Introduction(
( 32(
synthesis, in sugar beet leaves, a strong inhibition of protein synthesis is linked
to Fe deficiency in the leaves (Nishio et al., 1985). Decrease in leaf protein
contents under Fe deficiency are attributed to the Rubisco protein, which is
approximately 50% of the chloroplast soluble proteins (Ellis, 1979).
There are about 20 Fe atoms directly involved in the electron transport chain in
the thylakoid membranes. Because of higher Fe contents (12 atoms of Fe per
complex) Photosystem (PS) I is a major sink for Fe compared to PS II (3
atoms of Fe per complex) and the Cyt bf complex (5 atoms of Fe per complex)
(Raven et al., 1999). Iron is required in high amounts for the structural and
functional integrity of the thylakoid membranes. Fe deficiency in the leaves
does not decrease, equally, all the photosynthetic pigments and different
components of the electron transport chain (Pushnik and Miller, 1989). Under
Fe deficiency, the activity of PS I is more depressed than that of PS II (Pushnik
and Miller, 1989). The electron-transmitting ability of the PS I is strongly
increased upon re-supplying Fe to chlorotic leaves than that of PS II. Under
severe Fe deficiency, the activity of PS II also decreases and is more difficult
to restore (Terry, 1980; Morales et al., 1991).
Photosynthetic activity in the leaves is generally low under Fe deficiency but
light energy absorbed per chlorophyll molecule increases than the required for
photosynthesis under Fe deficiency, particularly under high radiation (Abadía
et al., 1999). This increases the risk of photoinhibitory and photooxidative
damages in Fe-deficient leaves.
Low concentrations of starch and sugars are a typical characteristic of Fe
deficient leaves (Arulanathan et al., 1990). This is usually attributed to the
reduced chlorophyll and ferredoxin concentrations, impairment of
photosynthetic electron transport and the decreased regeneration of reduced
ferredoxin (Marshner, 2012). Reduced photosynthetic activity is a typical
physiological response of plants to Fe deficiency. Re-supply of Fe to Fe-
deficient plants rapidly increases the photosynthesis (Larbi et al., 2004). Under
Fe deficiency, reduction in photosynthetic activity is attributed to reduced
photosynthetic electron transport and inhibition of carboxylation process due to

Chapter(2( ((((((((((((((((((((((((((((((((((((((((((((((((((((((((((((((((((((((((((((((((((((((((((((((((((((((((((((((((((((((((((((((((General(Introduction(
( 33(
decreased availability of ATP and NADPH for the Calvin cycle (Larbi, 2006).
Another important reason for the low photosynthesis is low concentration of
rubisco proteins (Larbi et al., 2004; Timperio et al., 2007).
2.4.2.4 Iron in seeds
Iron is mainly stored as phytoferritin (plant ferritin) in plants. The structure
resembles a hollow protein shell, where up to 5000 atoms of Fe (III) can be
stored. Phytoferritin has a proposed formula (FeO. OH)8. (FeO.OPO3H2),
which has usually a well-defined crystalline structure (Seckbach, 1982).
Ferritin is present in high abundance in seeds. In pea plants, more than 90%
of the total Fe present in the seed embryo is in the form of ferritin-bound Fe,
indicating that ferritin could be a major form of Fe storage in the seeds
(Marentes and Grusak, 1998). However, Fe concentration in seed as ferritin-
bound Fe largely varies among the plant species. For example, in legume
species, ferritin-Fe concentration in kidney beans ranges from 15% of the total
Fe up to 69% in lentils (Hoppler et al., 2009). Microanalysis of wheat seeds
has shown that Fe is present in high concentration in the aleurone and
scutellum layer of the embryo, while very low concentration in the endosperm
(Mazzolini et al., 1985). In barley sesds, approximately, 70% of the total Fe is
located in the endosperm and aleurone while 7 - 8% is found in the embryo
(Duffus and Rosie, 1976). In monocots, most of the Fe is deposited in the
protein storage vacuole (PSV) of the embryo and the aleurone layer and forms
specialized structures ‘globoids’ after complex formation with phytate (Søren et
al., 2009). The protein storage vacuoles are complex structures, consisting of
a protein matrix with globoid inclusions containing phytate, niacin,
carbohydrates and phenolics (Becraft, 2007). Phytic acid is a P storage
compound in most seeds. It has a strong chelating ability that readily binds
metal cations such as Ca, Zn, Mg, Fe, and Mn, making them insoluble and
decrease in their bioavailability in food (Lönnerdal, 2000; Urbano et al., 2000;
Jiang et al., 2001; Ficco et al., 2009).

Chapter(2( ((((((((((((((((((((((((((((((((((((((((((((((((((((((((((((((((((((((((((((((((((((((((((((((((((((((((((((((((((((((((((((((((General(Introduction(
( 34(
2.4.3 Manganese (Mn) in plants
Manganese ions have a high capacity to change the redox state. Mn2+ is the
more dominant form and easily oxidized to Mn3+ or Mn4+, and therefore, in the
plant Mn plays an important role in redox reactions. In biological systems,
Mn2+ and Mn4+ ions are fairly stable while Mn3+ is unstable (Hughes and
Williams, 1988).
2.4.3.1 Role of Mn in enzyme structure and activation
Mn2+ is required for the activation of a large number of enzymes but there are
only a small number of Mn-containing enzymes, namely the Mn-protein in PS
II, the Mn-containing superoxide dismutase (MnSOD) and oxalate oxidase
(Marschner, 2011). Oxalate oxidase is a Mn-containing multimeric
glycosylated enzyme (Kanauchi et al., 2009). Oxalate oxidase catalyses the
conversion of oxalate and dioxygen to CO2 and H2O2 (Lane, 1994).
Superoxide dismutases (SOD) are produced in all aerobic life systems and
play a vital role in protecting against reactive oxygen species (Elstner, 1982;
Fridovich, 1983). The Mn SOD is an important member of SOD isoenzymes
along with others i.e. FeSOD and CuZnSOD. MnSOD is not commonly found
in higher plants (Sandmann and Böger, 1983) and largely located in
mitochondria and peroxisomes. Production of MnSOD in the chloroplast of
numerous transgenic plants has increased abiotic stress tolerance e.g., salinity
(Tanaka et al., 1999) drought (Wang et al., 2005a) and Mn deficiency (Yu et
al., 1999a).
Polypeptide (33kDa) of the water-splitting system in PS II is an important Mn-
containing enzyme. Water splitting system contains six components of the
Mn4Ca, in which four Mn atoms arranged as a cluster, that are very important
for oxidation of 2 water molecules (Nikos et al., 1971).
There are almost 35 enzymes where Mn acts as a cofactor (Burnell, 1988).
For example enzymes which catalyse oxidation-reduction, decarboxylation and
hydrolytic reactions (Marschner, 1995). Manganese is essential for the PEP

Chapter(2( ((((((((((((((((((((((((((((((((((((((((((((((((((((((((((((((((((((((((((((((((((((((((((((((((((((((((((((((((((((((((((((((((General(Introduction(
( 35(
carboxykinase enzyme in the bundle sheath chloroplasts of C4 plants, where it
catalyses the process of decarboxylation. Manganese is required for the
activation of several enzymes of the shikimic acid pathway, and subsequent
pathways, resulting in the biosynthesis of aromatic amino acids (such as
tyrosine), different secondary products (such as lignin, flavonoids) and IAA
(Burnell, 1988; Hughes and Williams, 1988). For example, Mn by stimulating
and combining with peroxidase work as diffusible redox shuttle in lignin
biosynthesis (Önnerud et al., 2002), stabilizes the activity of phenylalanine
ammonia lyase (PAL) (Wall et al., 2008). Indole acetic acid (IAA) oxidase
activity is increased in the leaves suffering from deficient or toxic levels of Mn
(Morgan et al., 1976).
2.4.3.2 Role of Mn in Photosynthesis
In higher plants, photosynthesis is severely depressed under Mn deficiency.
Particularly, Mn deficiency causes a strong inhibition in photosynthetic O2
evolution in PS II (Shenker et al., 2004; Husted et al., 2010). In subterranean
clover, Mn deficiency has little effect on chlorophyll concentration or leaf dry
biomass, but there was more than 50% reduction in Photosynthetic O2
evolution (Nable et al., 1984). Photosynthetic O2 evolution was restored to
normal rate with one day after resupplying the Mn to deficient leaves
(Marscher, 2011). Manganese deficiency causes the changes in the
ultrastructure of thylakoid membranes, responsible for the loss of PS II
functional units in the stacked areas of thylakoid membranes (Husted et al.,
2010). Number of the PS II protein- pigment units in the thylakoid membranes
is restored upon resupplying the Mn (Simpson and Robinson, 1984; Gong et
al., 2010).
Under severe Mn deficiency, concentration and contents of chlorophyll is
significantly low in the leaves and ultrastructure of thylakoids is considerably
changed (Kroengier et al., 1993). Restoration or reversibility of these
ultrastructural changes of thylakoids is very difficult, and are most probably a
consequence of biosynthetic inhibition of lipids and carotenoids, and not of
enhanced photooxidation (lipid per- oxidation) of the thylakoids and chlorophyll

Chapter(2( ((((((((((((((((((((((((((((((((((((((((((((((((((((((((((((((((((((((((((((((((((((((((((((((((((((((((((((((((((((((((((((((((General(Introduction(
( 36(
(Marschner, 2011).
2.4.3.3 Role of manganese in cell division and elongation
In Mn-deficient plants, shortage of carbohydrates and also direct requirement
of Mn for growth is responsible for limited root growth (Campbell and Nable,
1988; Sadana et al., 2002). It is observed that the rate of cell elongation is
more suppressed under Mn deficiency than the rate of cell division (Abbott,
1967). Under normal Mn deficiency, the growth rate is rapidly restored on re-
supply of Mn. Formation and development of lateral root is strongly inhibited
under Mn deficiency (Abbott, 1967). In the roots of Mn-deficient plants,
development of a large number of small non-vacuolated cells in the roots
compared to Mn sufficient plants indicates Mn deficiency hinders the cell
elongation more strongly than cell division (Neumann and Steward, 1968).
2.4.4 Boron (B) in plants
Boron (B) is found in all subcellular compartments of the plant cells e.g. the
apoplasm, cell walls, cytosol and vacuoles, whereas greater part of the cellular
B is bound in the cell walls (Dannel et al., 2002). In the cells, B is present in
water-soluble and water insoluble forms (Matoh, 1997). Water-soluble B is
present in the apoplastic space in the form of boric acid and its concentration
decreases under limited supply of B to the plants (Skok and McIlrath, 1958;
Shive and Barnett, 1973). In some plant species, another water-soluble B
fraction is phloem-mobile B present in the form of B-polyol complexes (Hu et
al., 1997; Hu and Brown, 1997b). Although, size of the mobile B pool is much
smaller relative to total B, but it has vital role in plant growth (Brown et al.,
1999).
The water insoluble B fraction is largely present as a complex with
rhamnogalacturonan-II (RG-II) in higher plants and mostly situated in cell walls
(O’Neill et al., 1996). Mostly, B in the cell walls is coupled with pectin via the B-
RG-II complex, a borate diester (Kobayashi et al., 1996). It has been shown

Chapter(2( ((((((((((((((((((((((((((((((((((((((((((((((((((((((((((((((((((((((((((((((((((((((((((((((((((((((((((((((((((((((((((((((((General(Introduction(
( 37(
that B-RG-II and homogalacturonan (HG) are covalently bound together in
pectin (Ishii and Matsunaga, 2001). Accordingly, RG-II is the sole B containing
polysaccharide found in the biological materials (O’Neill et al., 1996) and most
likely the unique B form structurally complexed within the cell walls (Matoh,
1997).
Parr and Loughman (1983) have postulated 10 important roles of B in plants,
such as involvement in (i) transport of sugar compounds, (ii) synthesis of cell
wall, (iii) lignification, (iv) cell wall structure (v) carbohydrate metabolism, (vi)
RNA metabolism, (vii) respiration, (viii) indole acetic acid (IAA) metabolism, (ix)
phenol metabolism, and (x) functions of membranes. This long list shows the
immense role of B in plants but it still needs more research to fully understand
the functions of B in these processes. Functions of B in higher plants have
been reviewed by many authors (Goldbach, 1997; Blevins and Lukaszewski,
1998; Dannel et al., 2002; Brown et al., 2002). The role of B in cell wall
structure is well described (Matoh, 1997; Matoh and Kobayashi, 1998).
Although, the B function in plasma membrane is also well explained (Cakmak
and Römheld, 1997; Mühling et al., 1998; Kobayashi et al., 1999).
2.4.4.1 Role of B in cell wall structure
Boron is an important constituent of the cell wall structure. Durst (1988) stated
that 96 to 99% of the total B in cultured carrot cells is localized in cell walls,
while only 0.7 to 4.1% is present in the protoplasts. According to the Loomis
and Durst model (1992) for the role of B in cell wall structure, borate is cross-
linked in the middle lamella, probably associated with calcium (Ca) cross-
linking. Boron and Ca are vital to maintain the integrity of cell walls through
binding to the RG-II (rhamnogalacturaon II) (Matoh and Kobayashi, 1998).
Borate and Ca2+ cross-linking in the RG-II region (B-RG-II complex contains
two molecules of boric acid, two molecules of Ca2+ and two chains of
monomeric RG-II) play important role in retaining the chelator-soluble pectic
polysaccharides in cell walls (Kobayashi et al., 1999).
There is no evidence that B is directly involved in cell wall synthesis. However,

Chapter(2( ((((((((((((((((((((((((((((((((((((((((((((((((((((((((((((((((((((((((((((((((((((((((((((((((((((((((((((((((((((((((((((((((General(Introduction(
( 38(
B may influence the assimilation of proteins, pectins and/or their precursors
into the present and extending cell wall (Brown et al., 2002). Boron deficiency
rapidly increases the cell wall pore size, which leads to cell death upon
entering into the elongation phase of growth (Fleischer et al. 1999). The ability
of cells to form a pectin network with proper pore size is impaired under B
deficiency, which may influence important physiological processes, e.g.
polymer incorporation in the cell wall and transportation of wall-modifying
proteins or enzymes to their substrates and also the transport of polymers
from the protoplast into the cell wall (Fleischer et al., 1999; Brown et al., 2002,
Marschner, 2011). Boron could be crucial for cell-to-wall adhesion and to
organise the architectural integrity of the cell (Bassil et al., 2004). 2.4.4.2 Role in Boron Cellular membranes
Boron plays an important role in membrane structure and functional integrity.
The effect of B on plasma membranes may be similar to that in cell walls
(Brown et al., 2002). Accordingly, B is a key element in maintaining proper
structure of the membranes by forming cross-links with glyco-proteins and
glyco-lipids in the membrane “rafts”, membrane sub-domains rich in these
compounds (Brown et al., 2002).
The amount of B is located in the membranes is smaller relative to the total B
pool in the cell (Pollard et al., 1977; Torchia and Hirsh, 1982; Parr and
Loughman, 1983; Tanada, 1983). The B form in membranes is probably the
same as present in the cell walls (Cakmak and Römheld, 1997; Blevins and
Lukaszewski, 1998). Restoration of membrane functions inhibited due to B
deficiency and of B-deficient tissues after B re-supply is fast, and normally
occurs within a few minutes (Goldbach et al., 1990; Schon et al., 1990; Barr et
al., 1993). These functions may include membrane potential, release of
ferricyanide-dependent H+, activity of adenosine triphosphatase (ATPase),
limited nicotinamide adenine dinucleotide (NADH) oxidase activity, transport of
ions, and membrane permeability (Cakmak and Römheld, 1997; Blevins and
Lukaszewski, 1998).

Chapter(2( ((((((((((((((((((((((((((((((((((((((((((((((((((((((((((((((((((((((((((((((((((((((((((((((((((((((((((((((((((((((((((((((((General(Introduction(
( 39(
Boron may have direct functional roles in the cellular membranes, probably by
regulating the activity of membrane-related enzymes (Pfeffer et al., 1998).
Boron deficiency reduces the uptake rate of ions across the plasma membrane
(Pollard et al., 1977). Direct or indirect effects of B on membrane structure
control effects of B on ion uptake and also the functions of different membrane
transport processes, including plasma membrane-bound H+-pumping ATPase
(Goldbach and Wimmer, 2007). In suspension-cultured tobacco cells, indole
acetic acid (IAA) is required for the effect of B on the H+-ATPase, and IAA
induced enhanced H+ excretion requires B (Goldbach et al., 1990). In several
species, typical effects of B on plasma membrane integrity and H+ pumping
activity has been observed in vitro with membrane vesicles from B-sufficient
and B-deficient roots (Goldbach and Wimmer, 2007).
2.4.4.3 Role of B in root elongation and shoot growth
Boron plays a critical role in shoot growth and root elongation. Boron
deficiency causes a fast inhibition or termination of root elongation and shoot-
meristematic growth (Marschner, 1995). Boron deficiency results in stubby or
bushy appearance of the root, while in shoots it may lead to complete
inhibition of meristematic growth or even cause meristem death in different
species (Marschner, 2012). In squash plants, interrupting the B supply for 24 h
completely ceased the root elongation and 12 h after B re-supply, root
elongation again started to increase (Bohnsack and Albert, 1977).
Furthermore, stopping the supply of B increased the activity of IAA oxidase in
the roots, which decreased quickly on B re-supply.
Shoot growth inhibition is characteristic symptom of B deficiency. This growth
inhibition may lead to tissue death in some species and re-supply of B results
in a bushy shoot development as lateral shoots emerge (Marschner, 1995). In
root tips, B deficiency causes alterations in cell division from a normal
longitudinal to a radial direction, which results in a decrease of elongation
growth (Robertson and Loughman, 1974). Enhanced cell division in a radial
direction with a proliferation of cambial cells and impaired xylem differentiation

Chapter(2( ((((((((((((((((((((((((((((((((((((((((((((((((((((((((((((((((((((((((((((((((((((((((((((((((((((((((((((((((((((((((((((((((General(Introduction(
( 40(
are also features typical of the subapical shoot tissue of B-deficient plants
(Marschner, 2012).
2.4.5 Phosphorus (P) in plants
Phosphorus is an essential mineral nutrient, which is an important constituent
of many key structural compounds and also catalyzing a number of various
biochemical reactions taking place in plants. Phosphorus is very important for
some specific growth factors such as, stimulated shoot and root growth,
flowering and seed production, uniform and early crop maturity, biological N-
fixing capacity of legumes and increased disease resistance in crop plants.
Phosphorus is a major structural unit of nucleic acids, amino acids and various
proteins. Both DNA and RNA structures are linked together by phosphate to
form macromolecules. The structural form of P is also high in phospholipids of
biomembranes, where it links different molecules like amino acids, amines,
and alcohols to diglyceride (Marschner, 2012).
In seeds, phytic acid is the main storage form of P (also known as phytate)
(Lott et al. 1995; Park et al. 2006), and its concentration varies from 0.5% to
5.0% in different cereal and legume seeds (Greiner and Egli 2003; Laboure et
al. 1993). In seeds and grains, phytate is the main storage form of different
mineral nutrients like K, Mg, Ca, Mn, Zn and Fe (Lott et al., 1985; Raboy et al.
1989; Ravindran et al. 1994). Phosphorus as a component of different cellular
membranes, DNA and RNA is also found in the seeds (Raboy et al. 2001). In
maize seeds, more than 90% phytate is accumulated in the embryo and
aleurone layers (Lott et al. 1995; Raboy et al. 2000). Phytate plays two
important roles in the seeds (i) it serves as a pool of inositol phosphate and
cations and (ii) it controls the homeostasis of inorganic P during seed
development and also during germination and seedling development (Lott et
al. 1995). Concentration of P and phytic acid in the seeds largely depends on
cultivar, plant nutrient status and climatic condition during seed developmental
stage (Horvatic and Balint 1996; Miller et al. 1980; Raboy et al. 1991). High
seed P reserves have been shown to improve early seedling development and
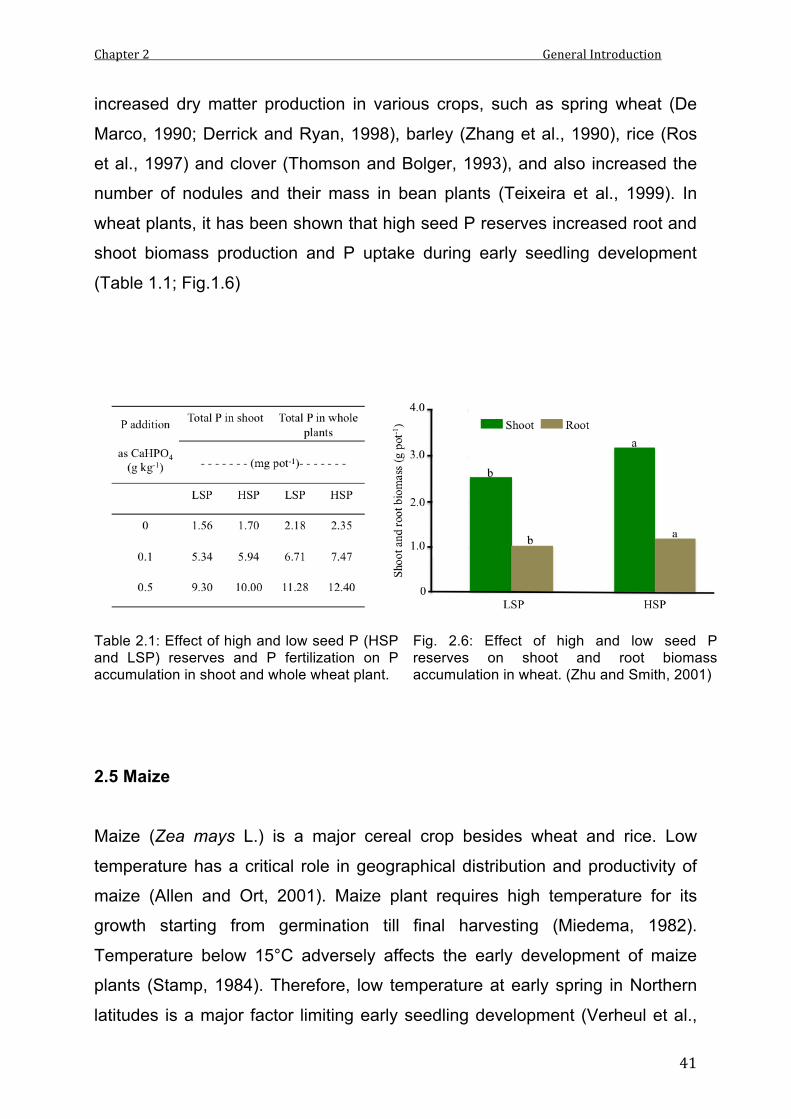
Chapter(2( ((((((((((((((((((((((((((((((((((((((((((((((((((((((((((((((((((((((((((((((((((((((((((((((((((((((((((((((((((((((((((((((((General(Introduction(
( 41(
increased dry matter production in various crops, such as spring wheat (De
Marco, 1990; Derrick and Ryan, 1998), barley (Zhang et al., 1990), rice (Ros
et al., 1997) and clover (Thomson and Bolger, 1993), and also increased the
number of nodules and their mass in bean plants (Teixeira et al., 1999). In
wheat plants, it has been shown that high seed P reserves increased root and
shoot biomass production and P uptake during early seedling development
(Table 1.1; Fig.1.6)
Table 2.1: Effect of high and low seed P (HSP and LSP) reserves and P fertilization on P accumulation in shoot and whole wheat plant.
Fig. 2.6: Effect of high and low seed P reserves on shoot and root biomass accumulation in wheat. (Zhu and Smith, 2001)
2.5 Maize
Maize (Zea mays L.) is a major cereal crop besides wheat and rice. Low
temperature has a critical role in geographical distribution and productivity of
maize (Allen and Ort, 2001). Maize plant requires high temperature for its
growth starting from germination till final harvesting (Miedema, 1982).
Temperature below 15°C adversely affects the early development of maize
plants (Stamp, 1984). Therefore, low temperature at early spring in Northern
latitudes is a major factor limiting early seedling development (Verheul et al.,

Chapter(2( ((((((((((((((((((((((((((((((((((((((((((((((((((((((((((((((((((((((((((((((((((((((((((((((((((((((((((((((((((((((((((((((((General(Introduction(
( 42(
1995). Low temperature adversely affects plant growth rate causing
development of weak seedlings and prolonged growth duration.
Vigorous and well-developed maize seedlings are required to tolerate the cold
and wet conditions of Central and Northern Europe and also for its integration
to no till or conservation tillage systems of temperate region where soil
warming is relatively slow (Tollner et al., 1984; Arshad and Azooz, 1996). In
zero tillage cultivation systems, low soil temperature is the most dominant
physical stress factor which limits the early development of maize seedlings
(Chassot, 2000), where the shoot apex of the maize seedlings which remains
below the soil surface upto the 6-leaf stage (Blacklow, 1972; Erbach et al.,
1986; Stone et al., 1999). Therefore, soil temperature has a direct effect on
both, the shoot and root meristems. Shoot development is impaired by a direct
effect of soil temperature on shoot meristems or by limited supply of mineral
nutrients e.g. P, Zn, Mn etc. through roots (Engels and Marschner, 1990). It
has been found that soil temperature directly controls the early leaf
development of maize plants grown under conditions climatic (Beauchamp and
Lathwell, 1966) and also in the field experiments (Giauffret et al., 1995; Stone
et al., 1999). Existing maize genotypes for cold tolerance are differentiated on
the basis of shoot development (Lee et al., 2002) and root development
(Stamp, 1984; Richner et al., 1997). In a study on roots related traits to low
temperature tolerance, there was a strong link between the length of the lateral
roots on the primary roots and the traits associated with photosynthesis and to
the dry biomass (Hund et al., 2007).
Uptake and translocation of mineral nutrients is markedly reduced in maize at
low soil temperature (Engels et al., 1992). Furthermore, maize has a low
morphological ability for adaptation to low root zone temperature due to limited
biomass allocation to the root. It was further suggested that reduced allocation
of biomass to roots could be responsible for greater susceptibility of maize to
nutrient deficiency at low root zone temperature. Engels and Marschner (1996)
found that in maize plants soil temperature inhibits the translocation of Zn and
Mn form root to shoot irrespective of the chemical nutrient availability in the
rhizosphere or by shoot meristem temperature, while net translocation of Fe is

Chapter(2( ((((((((((((((((((((((((((((((((((((((((((((((((((((((((((((((((((((((((((((((((((((((((((((((((((((((((((((((((((((((((((((((((General(Introduction(
( 43(
regulated according to the internal growth related shoot demand per unit of
roots.
2.6 Soybean
Soybean (Glycine max L.) is an important crop for protein and oil production
worldwide. In 2009, soybean was cultivated on more than 90 million hectares
of agricultural lands. It is grown all over the world in varying climatic and soil
conditions. Zinc deficiency in agricultural soils is a world wide problem,
especially in calcareous soils (Takkar and Walker, 1993; White and Zasoski,
1999) and limits the crop yields (Grewal et al., 1997; Cakmak, 2000). Global
survey conducted by FAO and other studies have shown that about 50% of the
cultivated soils were either Zn deficient (Sillanpa, 1982) or low in available Zn
for plant growth (Graham et al., 1992; Welch, 1993).
Manganese (Mn) deficiency in soils is also another serious problem for
soybean production in many areas throughout the world (Reuter et al., 1988)
and responsible for decreased crop yields. Manganese deficiency is
particularly more prominent in soils with low inherent Mn availability, relatively
high pH levels and under dry soil conditions (drought).
2.7 Aims and objectives
Previous studies have reported positive effects of nutrient seed priming in
different crop plants, but knowledge on the specific contribution of primed
nutrients in plant growth and nutritional status is limited. This thesis focused on
investigating the specific role of nutrient seed priming in maize and soybean
plants grown under different abiotic stress conditions. Main objectives of this
thesis were as follows:
Objective 1.

Chapter(2( ((((((((((((((((((((((((((((((((((((((((((((((((((((((((((((((((((((((((((((((((((((((((((((((((((((((((((((((((((((((((((((((((General(Introduction(
( 44(
To investigate the effect of nutrient seed priming on early seedling
development, plant growth, nutrient uptake and final yield of maize at low root
zone temperature.
Following study questions were addressed to achieve the objective 1:
i. What is the optimum priming duration for maize seeds?
ii. How much Zn, Mn, Fe, B and P can be added into maize seed
via nutrient seed priming?
iii. What are effects of nutrient seed priming on early seedling
development and how long primed nutrients can support plant
growth at early stages?
iv. Can nutrient seed priming improve shoot and root growth of
maize at low root zone temperature stress?
v. Does nutrient seed priming improve the nutrient status of maize
plants under various nutrient availability conditions e.g.
deficient, low availability and low root zone temperature stress
conditions?
vi. Can nutrient seed priming improve plant growth, nutrient
uptake and the grain yield of maize grown in the temperate
climate of Germany?
Objective 2.
To study the effect of nutrient seed priming on soybean seedling development
and plant nutrient status under low Zn and Mn available conditions.
Following study questions were established to achieve the objective 2.
i. What is the optimum priming duration for soybean?
ii. How soybean seed should be primed to avoid damage during the
priming process.
iii. How does Zn and Mn priming effect shoot and root growth of
soybean seedlings under Zn and Mn deficient conditions?
iv. Can Zn and Mn seed priming improve plant nutrient status under
nutrient deficient conditions?

Chapter(2( ((((((((((((((((((((((((((((((((((((((((((((((((((((((((((((((((((((((((((((((((((((((((((((((((((((((((((((((((((((((((((((((((General(Introduction(
( 45(

Chapter(3(((((((((((((((((((((((((((((((((((((((((((((((((((((((((((((((((((((((((((((((((((((((((((((((((((((((((((((((((Maize(Nutrient(Seed(Priming(
( 46(
Chapter 3
Impact of nutrient seed priming on germination, seedling
development, nutritional status and grain yield of maize (Journal of Plant Nutriton: Accepted, 2012)
Authors
Imran Muhammad, Maria Kolla, Römheld Volker, Neumann Günter
Institute of Crop Science, Nutritional Crop Physiology (340h),
Universität Hohenheim, 70593 Stuttgart, Germany
Corresponding author: Imran Muhammad
Institute of Crop Science, Nutritional Crop Physiology (340h)
Universität Hohenheim, 70599, Stuttgart, Germany
Phone: 0045 71536841 Fax: 0049 711 459 23295
E-mail: [email protected]
Contribution in the manuscript:
Planning of experiments, Preparation of experimental materials,
Nutrient analysis, Root length measurements, Conducting field trial,
Statistical analysis, Manuscript writing.

Chapter(3(((((((((((((((((((((((((((((((((((((((((((((((((((((((((((((((((((((((((((((((((((((((((((((((((((((((((((((((((Maize(Nutrient(Seed(Priming(
( 47(
3.1. Abstract
Effects of seed priming with zinc (Zn) plus manganese (Mn), boron (B)
and phosphate (P) on growth and nutritional status of maize were
studied. Nutrient seed priming significantly increased seed contents of
primed nutrients. In nutrient solution (NS) lacking Zn and Mn, growth of
maize plants primed with Zn+Mn increased by more than 50% and
100%, respectively, as compared to control treatment. The primed
nutrients were efficiently translocated to the growing shoot and could
maintain Zn and Mn supply for at least three weeks of the culture
period.
In soil culture, plants suffered from P and Zn deficiency, which was
mitigated to some extent by P and Zn+Mn priming. Particularly,
translocation of Zn seed reserves to the shoot tissue was negatively
affected by the highly calcareous soil. In the field experiment, Zn+Mn
seed priming increased grain yield by 15 %, demonstrating the potential
for long-lasting effects of nutrient seed priming.
Keywords: Zea mays L., nutrient seed priming, Zinc, Boron,
Phosphorus, maize grain yield

Chapter(3(((((((((((((((((((((((((((((((((((((((((((((((((((((((((((((((((((((((((((((((((((((((((((((((((((((((((((((((((Maize(Nutrient(Seed(Priming(
( 48(
3.2. Introduction
Early seedling development and uniform crop stand establishment is an
important determinant for crop yield (Harris, 1996). Different abiotic
stresses such as drought, flooding, low temperatures salinity and other
toxicities can delay germination and increase abnormal seedling
development (Bourgne et al., 2000). Poor crop stand can promote weed
populations causing competition for water, light and limiting nutrients
(Kropff and van Laar, 1993).
Sufficient mineral nutrient reserves in the seed are necessary to
maintain seedling growth until root uptake starts supplying soil nutrients.
This is particularly important for crops grown on soils with low nutrient
availability (Asher, 1987) as demonstrated by numerous studies. Rice
plants obtained from seeds with high Zn contents could better cope with
limited Zn availability in soils as compared with those grown from seeds
with low seed reserves (Rengel and Graham 1995; Hacisalihoglu and
Kochian 2003; Tehrani et al., 2003). Zhu and Smith (2001) have shown
that wheat plants grown from seeds with high P contents were able to
accumulate more P in the shoot compared to plants from low P seeds.
Also, vigor and normal seedling development is strongly determined by
mineral nutrient seed reserves. Seeds with low Mn contents produced
low-vigor wheat seedlings and low yields at harvest (Marcar and
Graham, 1986; Singh and Bharti, 1985). Delayed germination and
seedling development was observed in barley and lupin with low Mn
seed contents (Genc et al., 2000; Longenecker et al., 1991; Crosbie et
al., 1994). It has been shown that percentage of abnormal seedlings
increased with decreasing level of B in the soybean seeds (Rerkasem
et al., 1997) and in green and black grams (Rerkasem et al., 1990; Bell
et al., 1989). High seed P contents have improved early seedling growth
and dry matter production in clover (Thomson and Bolger, 1993), barley
(Zhang et al., 1990), spring wheat (Bolland and Baker, 1988) and rice
(Ros et al., 1997). Seed P contents also affected the number and

Chapter(3(((((((((((((((((((((((((((((((((((((((((((((((((((((((((((((((((((((((((((((((((((((((((((((((((((((((((((((((((Maize(Nutrient(Seed(Priming(
( 49(
biomass of nodules in common bean (Teixeira et al., 1999).
Various techniques are employed to improve seed quality, to reduce the
time between sowing and seedling emergence and also to minimize
biotic and abiotic stresses during germination, e.g. use of
biotechnology, hybrid seeds, mechanical seed treatments such as
polishing off or removal of seed coat (testa), sorting the seeds into
different seed size classes and various seed treatments like seed
coating, pelleting or soaking in different solutions (seed priming)
(Farooq et al., 2012).
In general, seed priming is a pre-sowing seed treatment in which seeds
are soaked in water and dried back to storage moisture contents until
further use. Seed priming in water has been shown to decrease time
between sowing and emergence and to improve seedling vigour (Harris,
1996; Parera and Cantliffe, 1994). ‘Nutrient seed priming’ is a technique
in which seeds are soaked in nutrient solution instead of simple water.
Nutrient seed priming increases the seed nutrient contents along with
the priming effect to improve seed quality for better germination and
seedling establishment. Priming seeds in solutions of macro- or micro-
nutrients has been shown to improve yield of rice (Peeran and
Natanasabapathy, 1980), wheat (Khalid and Malik, 1982; Marcar and
Graham, 1986; Wilhelm et al., 1988), and forage legumes (Sherrell,
1984), but a risk of seed damage and impairment of germination by
priming at high nutrient concentrations has been also reported (Roberts,
1948; Ajouri et al., 2004).
The present study investigates perspectives for seed priming with water
and various nutrients (P, Zn, Mn, and B) in maize. After establishing and
optimising the priming conditions, hydroponic experiments were
conducted to characterise the nutritional potential and the persistence of
priming effects. Perspectives of seed priming to overcome problems of
limited soil nutrient availability were investigated in pot experiments and

Chapter(3(((((((((((((((((((((((((((((((((((((((((((((((((((((((((((((((((((((((((((((((((((((((((((((((((((((((((((((((((Maize(Nutrient(Seed(Priming(
( 50(
a field experiment was conducted to investigate the effects of seed
priming on maize yield formation under temperate climatic conditions in
South West Germany.
3.3. Materials and methods
3.3.1. Water priming
To determine the optimal time duration for maize seed priming, 100
seeds were soaked in 200 ml distilled water for 0, 8, 16, 24 and 32
hours (h). Soaking treatments were started from longer to shorter
incubation periods and seeds were finally removed from water at the
same time. Thereafter, seeds were rinsed with distilled water for 1
minute to remove any leached compounds from the seed surface, and
dried for 2 hours at room temperature before starting the germination
test. For germination, 10 seeds were incubated in rolls of 4 layers moist
filter paper (MN 710, Macchery & Nagel, Düren, Grmany) for 5 days at
24°C, using 6 replicates per treatment. Germination speed, shoot and
root lengths of the seedlings were recorded.
3.3.2. Nutrient seed priming
Nutrient seed priming is a technique, in which seeds are soaked in
mineral nutrient solution for a particular time duration, dried back to
initial moisture contents and stored for further use. Maize seeds were
primed for 24 h (pre-determined for water priming) by soaking in distilled
water and nutrient solutions containing Zn+Mn, B and P, respectively.
Based on the seed water absorbing capacity (approx. 0.08 ml seed-1),
nutrient solution concentrations were calculated to double the contents
of Zn, Mn, B and 50% of the seed P reserves. Concentrations of
nutrients in the priming solutions are shown in Table 3.1. The priming
and germination procedure was further conducted as described for
water priming. Un-primed seeds and water-primed seeds were used as
control treatments.

Chapter(3(((((((((((((((((((((((((((((((((((((((((((((((((((((((((((((((((((((((((((((((((((((((((((((((((((((((((((((((((Maize(Nutrient(Seed(Priming(
( 51(
Table 3.1: Concentrations and salts used for maize nutrient seed priming treatments.
3.3.3.Plant culture
Pot experiments were conducted in a growth chamber under controlled
environmental conditions with average air temperatures of 25 + 3 °C,
15/9 h light/dark periods, light intensity 400 mol m-2 s-1, and a relative air
humidity of 60-80%. For the solution culture experiment, seeds were
germinated for 7 days in filter papers moistened with distilled water as
described for the germination test. Thereafter, seedlings were
transferred into pots filled with 2.5 liters (L) continuously aerated
nutrient solution (6 plants pot-1). As a positive control treatment, plants
were grown in complete nutrient solution, while plants from nutrient
primed seeds were grown in nutrient solution lacking the respective
nutrients. Two plants per pot were harvested 1, 2 and 3 weeks (first
(1st), second (2nd) and third (3rd) harvest) after transfer to nutrient
solution. The nutrient solution was renewed every 2-3 days and had the
following composition: 2 mM Ca(NO3)2, 0.7 mM K2SO4, 0.1mM KCl, 0.5
mM MgSO4, 0.2 mM KH2PO4, 1 µM H3BO3, 0.5 µM MnSO4, 0.2 µM
CuSO4, 0.01 µM (NH4)6Mo7O24, 0.5 µM ZnSO4 and 20 µM Fe-EDTA.
For soil culture, seeds were sown directly at a depth of 2cm and 4
plants were grown in each pot with 1 kg of a highly-buffered calcareous
Loess sub-soil (pH (CaCl2) 7.6; with extremely low nutrient availability
with organic carbon (Corg. <0.3%), CaCO3 -23.3%, available P (CAL
Treatment Seed priming
Control Un-primed
Water Water primed
Zn+Mn 4 mM Zn + 2.5mM Mn solution
as ZnSO4 .H2O and MnSO4
B 5 mM B as H3BO3
P 0.2 M P as K2HPO4

Chapter(3(((((((((((((((((((((((((((((((((((((((((((((((((((((((((((((((((((((((((((((((((((((((((((((((((((((((((((((((((Maize(Nutrient(Seed(Priming(
( 52(
extraction) 5 mg kg-1 soil, water- extractable Ca2+ and Mg2+ (mg kg−1
soil) 59.9 and 11.3, respectively. Calcium chloride–diethylenetriamine
penta-acetic acid (CAT)-extractable micronutrient concentrations (mg
kg−1 soil): Mn=7.4, Fe=369, Zn=0.8, B=0.9, and Cu 0.5 (VDLUFA,
2004). The soil was fertilized once at the start of the experiment with
nitrogen (N), phosphorus (P), potassium (K) and magnesium (Mg).
Nitrogen was applied as NH4NO3 at 100 mg kg-1 soil, P as Ca(H2PO4)2
at 100 mg kg-1 soil, K as K2SO4 at 150 mg kg-1 soil and Mg as MgSO4
at 100 mg kg-1 soil. The different fertilizers were mixed with the soil
substrate and sieved through a 2 mm mesh size before filling the pots.
Soil moisture content was adjusted to 18% on weight (w/w) basis and
kept constant throughout the experiment by gravimetric control.
The field experiment was conducted at the University of Hohenheim,
research station farm “Heidfeldhof”, Stuttgart, Germany. For each of 4
replicates, 80 seeds were sown in 2 rows on an area of 7.5 m2 (20th of
April 2010). Field soil is a silty-clay Luvisol with organic matter 0.97%
and pH (CaCl2) 7.3. After seedling emergence, the number of plants
was reduced to34 in each row (68 plants per replicate). Final harvest
was performed in October 2010. Fertilization was conducted according
to farmers practice with N 167 kg ha-1, P2O5 42 kg ha-1, K2O 42 kg ha-1,
MgO 23 kg ha-1 and sulpher (S) 63 kg ha-1.
3.3.4. Plant analysis
At harvest, shoot and root dry biomass were determined for plants
grown in nutrient solution and soil culture. In the nutrient solution
experiment, three sequential harvests of each 2 plants were performed
at 2, 3 and 4 weeks after sowing. In field-grown plants, final grain yield
was determined. Plant mineral nutrient analysis was conducted for whole shoots of plants
grown in solution culture. In soil-grown plants, 2 young leaves from the
top and rest of the shoot were analyzed separately. For the extraction of

Chapter(3(((((((((((((((((((((((((((((((((((((((((((((((((((((((((((((((((((((((((((((((((((((((((((((((((((((((((((((((((Maize(Nutrient(Seed(Priming(
( 53(
mineral elements of plant and seed samples, microwave wet digestion
in a microwave (MLS 1200 Mega microwave system, Milestonl,
Leutkirch, Germany) was performed. For digestion, 500 mg of dried
plant material from each replicate were transferred into a microwave
digestion vessel (50 mL Teflon bottles). Five mL concentrated HNO3
and 4 mL 30 % H2O2 solution were used as digestion mixture. After
digestion the final volume was adjusted to 25 ml. After addition of 0.1
mL Cs/La buffer to 5 mL of the digested solution, Mn andZn
concentrations were measured by atomic absorption spectrometry
(UNICAM 939, Offenbach/Main Germany) and mass spectrometry (ICP
- ELAN 6000, Perkin Elmer, USA) was used for B analysis. Phosphate
in the ash solution was determined spectrophotometrically
(Spectrophotometer, Hitachi, Japan) by the method of Gericke and
Kurmis (1952). 3.3.5. Statistical analysis
A complete randomized block design with 4 replicates was used in all
experiments. Analysis of variance was performed at a significance level
of p < 0.05 using ‘Sigma Stat version 3.1 software (SYSTAT Software
Inc., Erkrath, Germany).
3.4 Results 3.4.1. Effect of water priming on seed germination
Water priming of maize seeds for 24 and 36 h accelerated germination
and growth of young seedlings. Germination rate of 100 percent (%)
was detectable already at 3 days after sowing, - more than 2 days
before seeds of the remaining treatments which reached approximately
90 % germination (Fig. 3.1a). Shoot and root lengths of 5 days old
maize seedlings were also increased by more than 300% by 24 and 32
h water priming as compared with unprimed control (Fig. 3.1b). Similar
results were obtained when nutrient solutions (Table 3.1) instead of
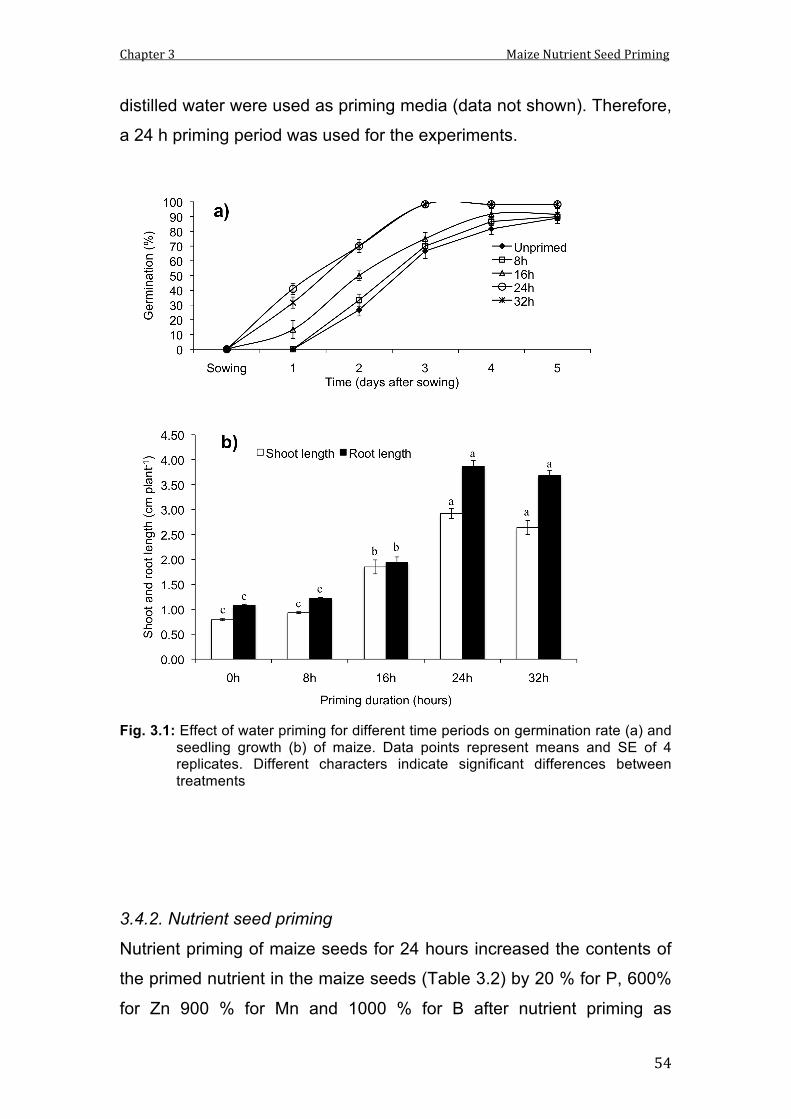
Chapter(3(((((((((((((((((((((((((((((((((((((((((((((((((((((((((((((((((((((((((((((((((((((((((((((((((((((((((((((((((Maize(Nutrient(Seed(Priming(
( 54(
distilled water were used as priming media (data not shown). Therefore,
a 24 h priming period was used for the experiments.
Fig. 3.1: Effect of water priming for different time periods on germination rate (a) and
seedling growth (b) of maize. Data points represent means and SE of 4 replicates. Different characters indicate significant differences between treatments
3.4.2. Nutrient seed priming
Nutrient priming of maize seeds for 24 hours increased the contents of
the primed nutrient in the maize seeds (Table 3.2) by 20 % for P, 600%
for Zn 900 % for Mn and 1000 % for B after nutrient priming as

Chapter(3(((((((((((((((((((((((((((((((((((((((((((((((((((((((((((((((((((((((((((((((((((((((((((((((((((((((((((((((((Maize(Nutrient(Seed(Priming(
( 55(
compared to the natural seed reserves, corresponding with an absolute
increase of .185, 45, 16 and 6.5 µg seed-1, respectively. Differences in
nutrient concentrations (not shown) and contents (Table 3.2) between
un-primed and water primed seeds were negligible for the measured
nutrients. Table 3.2: Mineral nutrient contents of maize seeds after nutrient seed priming. Data
represent means of 4 replicates. Different characters indicate significant differences
between treatments.
Nutrient Seed nutrients contents (µg sesd-1)
Treatments Zn Mn B P
Unprimed 8.5 +0.4 b 2.0 +0.1 b 0.72 +0.2 b 965 +23 b
Water 7.9 +0.4 b 1.7 +0.1 b 0.63 +0.11 b 970 +27 b
Zinc + Mn 52.7 +2.5 a 18.1 +1.0 a 0.71 +0.07 b 961 +31 b
Boron 7.8 +0.6 b 1.7 +0.1 b 7.08 +0.07 a 960 +21 b
Phosphorus 7.8 +0.6 b 1.7 +0.1 b 0.62 +0.09 b 1155 +37 a
3.4.3. Hydroponic culture experiment
Effects on biomass production were most expressed after Zn+Mn
priming, particularly at final harvest at 4 weeks after sowing. Plants
primed with Zn+Mn produced ≈50% more shoot biomass and ≈100%
more root biomass as compared with control plants supplied with
complete nutrient solution (Fig. 3.2), despite of the fact that the nutrient
status of control plants supplied with complete NS was always sufficient
throughout the culture period (Fig. 3.3).
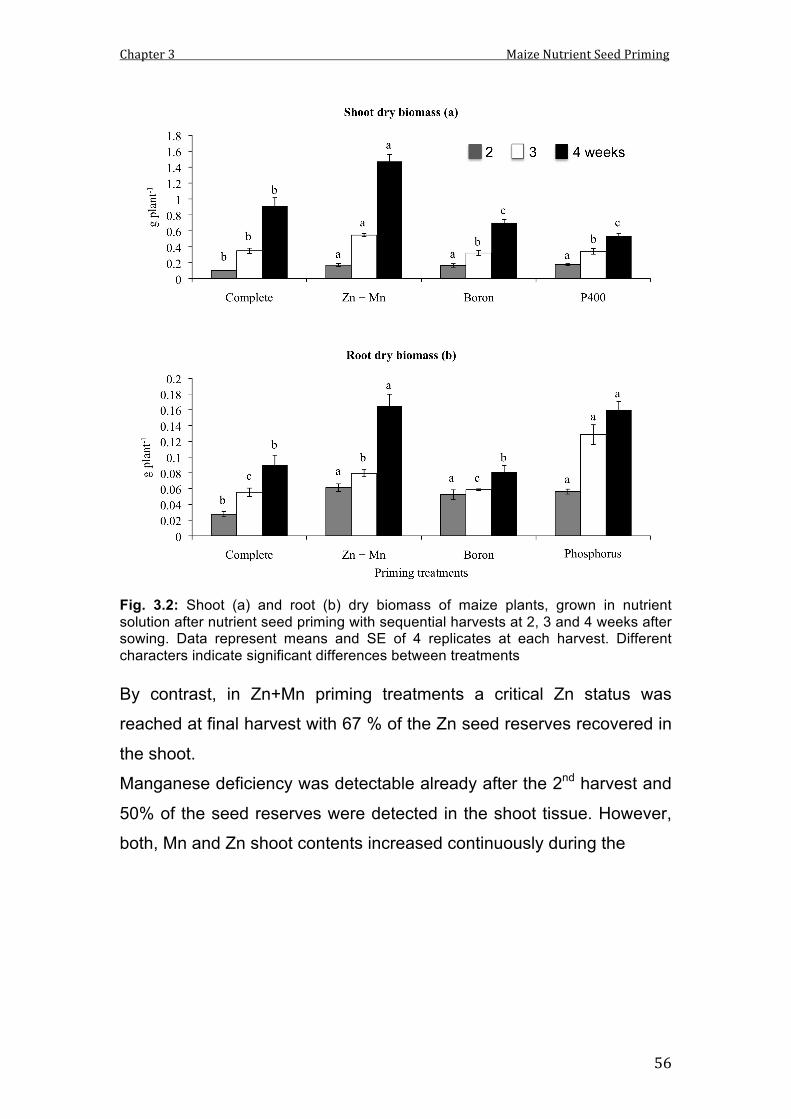
Chapter(3(((((((((((((((((((((((((((((((((((((((((((((((((((((((((((((((((((((((((((((((((((((((((((((((((((((((((((((((((Maize(Nutrient(Seed(Priming(
( 56(
Fig. 3.2: Shoot (a) and root (b) dry biomass of maize plants, grown in nutrient solution after nutrient seed priming with sequential harvests at 2, 3 and 4 weeks after sowing. Data represent means and SE of 4 replicates at each harvest. Different characters indicate significant differences between treatments By contrast, in Zn+Mn priming treatments a critical Zn status was
reached at final harvest with 67 % of the Zn seed reserves recovered in
the shoot.
Manganese deficiency was detectable already after the 2nd harvest and
50% of the seed reserves were detected in the shoot tissue. However,
both, Mn and Zn shoot contents increased continuously during the
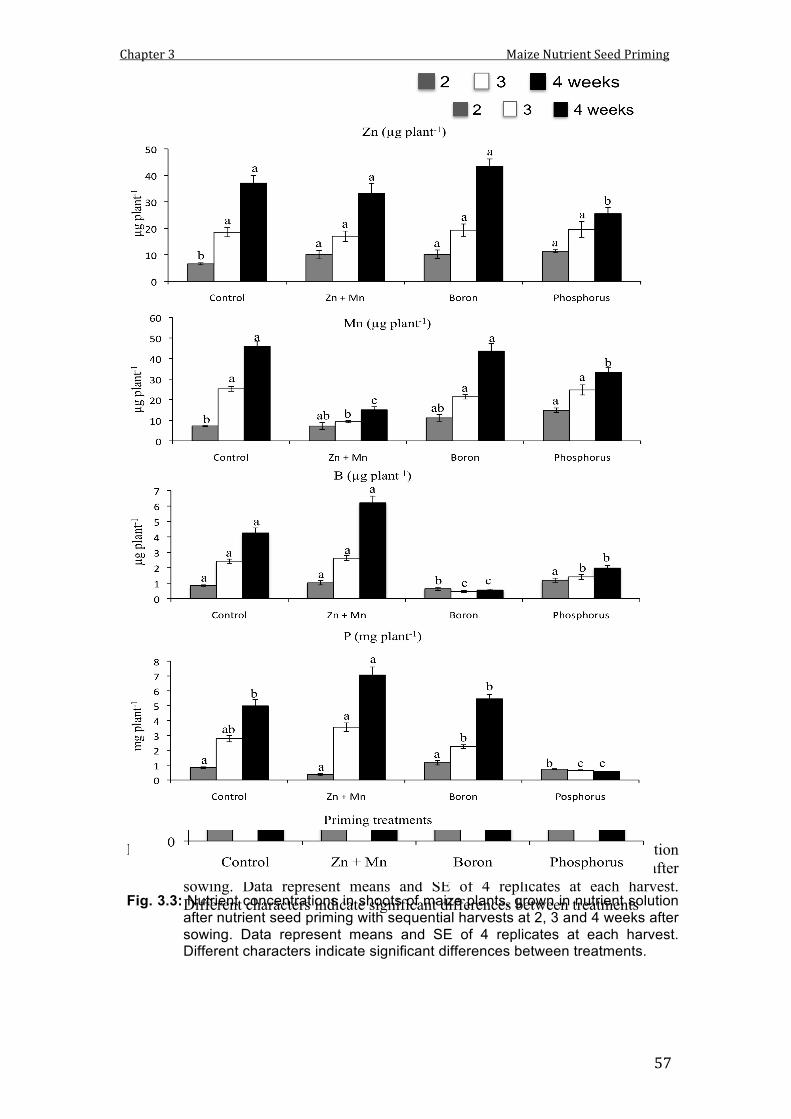
Chapter(3(((((((((((((((((((((((((((((((((((((((((((((((((((((((((((((((((((((((((((((((((((((((((((((((((((((((((((((((((Maize(Nutrient(Seed(Priming(
( 57(
Fig. 3.3: Nutrient concentrations in shoots of maize plants, grown in nutrient solution
after nutrient seed priming with sequential harvests at 2, 3 and 4 weeks after sowing. Data represent means and SE of 4 replicates at each harvest. Different characters indicate significant differences between treatments.
Figure 3.4: Nutrient contents in shoots of maize plants, grown in nutrient solution
after nutrient seed priming with sequential harvests at 2, 3 and 4 weeks after sowing. Data represent means and SE of 4 replicates at each harvest. Different characters indicate significant differences between treatments
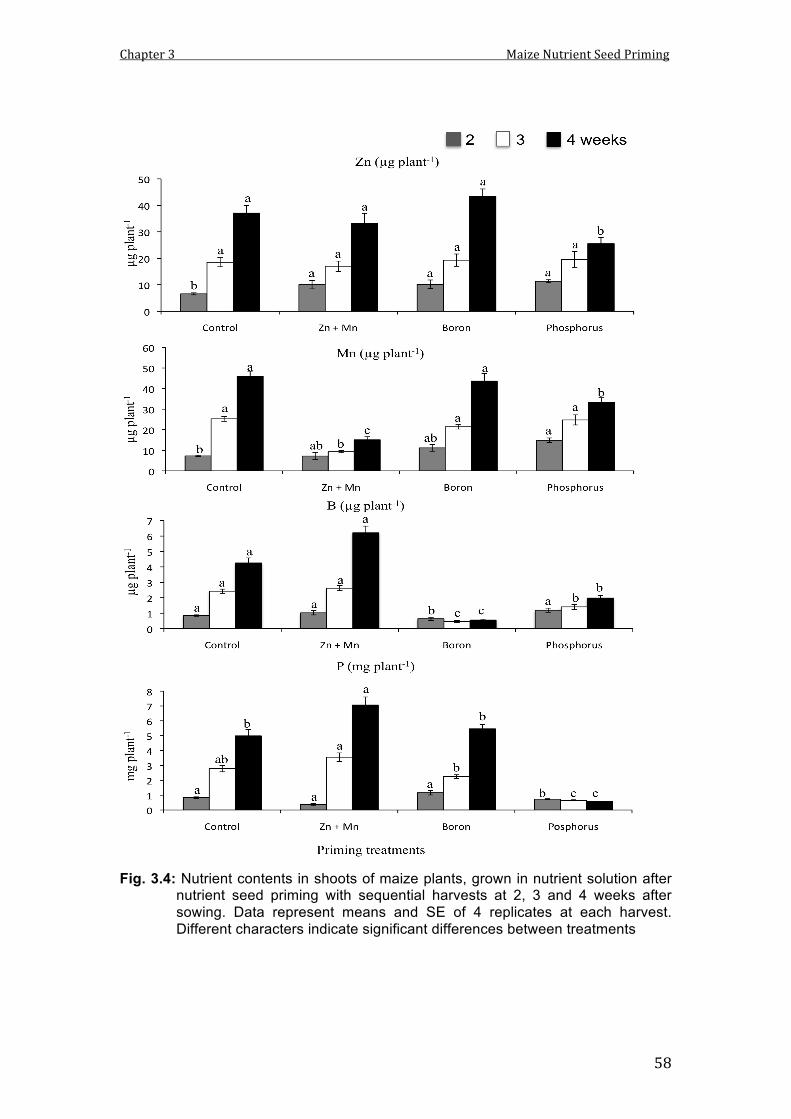
Chapter(3(((((((((((((((((((((((((((((((((((((((((((((((((((((((((((((((((((((((((((((((((((((((((((((((((((((((((((((((((Maize(Nutrient(Seed(Priming(
( 58(
Fig. 3.4: Nutrient contents in shoots of maize plants, grown in nutrient solution after
nutrient seed priming with sequential harvests at 2, 3 and 4 weeks after sowing. Data represent means and SE of 4 replicates at each harvest. Different characters indicate significant differences between treatments

Chapter(3(((((((((((((((((((((((((((((((((((((((((((((((((((((((((((((((((((((((((((((((((((((((((((((((((((((((((((((((((Maize(Nutrient(Seed(Priming(
( 59(
culture period of Zn+Mn-primed plants. Moreover, priming treatments
with Zn+Mn also increased shoot P contents (Fig. 3.4) in shoot by 40%
over the control with complete nutrient solution as soon as Zn shoot
concentrations reached the deficiency threshold.
Shoot growth of plants obtained from B and P-primed seeds did not
reach the biomass level of control plants with complete NS but root
biomass production after P priming exceeded the control treatment at all
harvest dates. This coincided with P limitation already at the 2nd harvest
in P-primed plants.
Boron deficiency in B primed plants was detectable at the 2nd harvest
with marginal effects on biomass production. At the 1st harvest root
biomass of B-primed plants was higher as the control in complete NS.
In contrast to Zn +Mn priming, plants obtained from P and B-primed
seeds did not continuously increase B and P shoot contents during the
culture period. About 50 % of the P reserves in P primed seeds were
translocated to the shoot compared with only 7 % in case of B-primed
plants.
Phosphate priming increased the Zn and Mn contents at the 1st harvest
as compared to control plants supplied with complete nutrient solution.
In contrast, at later stages of plant development P riming reduced Zn
and Mn contents. Only at the 1st harvest, B priming had a positive effect
on Zn and P contents of the plants.
3.4.4. Soil culture experiment
In soil-grown plants, seed priming with Zn+Mn induced only a moderate
stimulation of shoot biomass production (+18%) as compared with the
unprimed and water-primed control, while root biomass was not affected
(Fig. 3.5).
Plants suffered from Zn deficiency in all treatments, although Zn priming
improved the Zn status, particularly, in young leaves. However, even in
Zn-primed plants, only15% of the Zn seed reserves reached the shoot
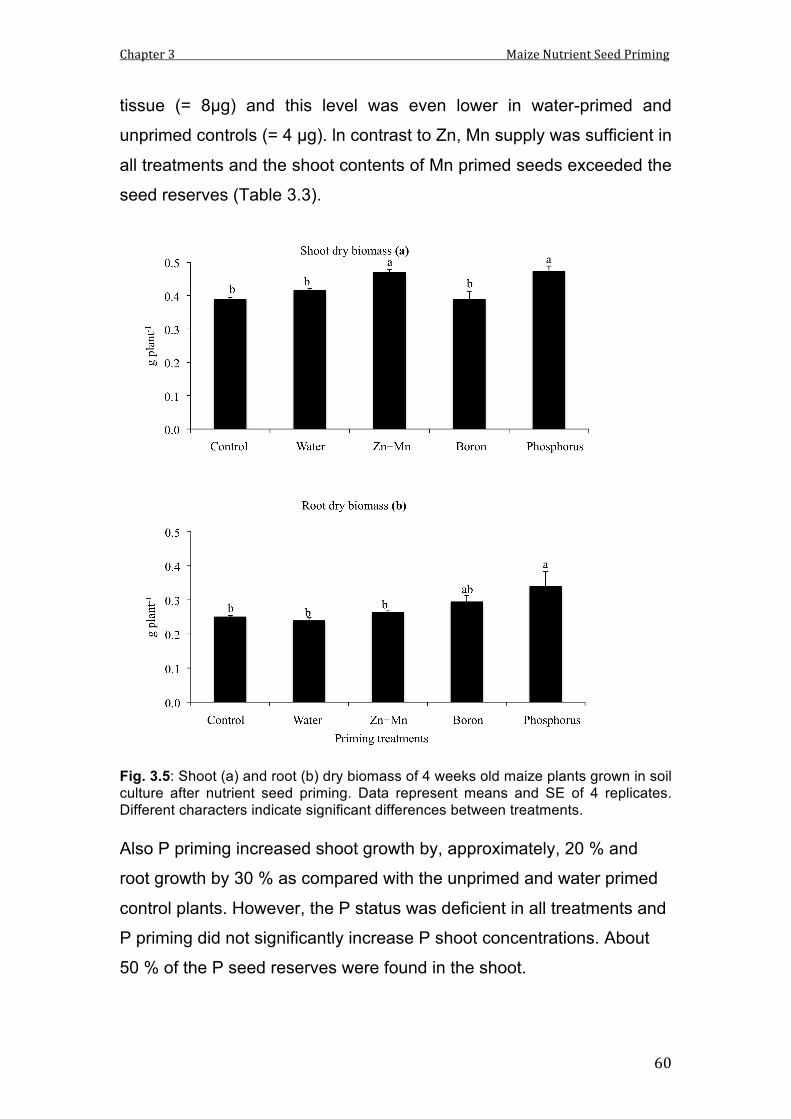
Chapter(3(((((((((((((((((((((((((((((((((((((((((((((((((((((((((((((((((((((((((((((((((((((((((((((((((((((((((((((((((Maize(Nutrient(Seed(Priming(
( 60(
tissue (= 8µg) and this level was even lower in water-primed and
unprimed controls (= 4 µg). ln contrast to Zn, Mn supply was sufficient in
all treatments and the shoot contents of Mn primed seeds exceeded the
seed reserves (Table 3.3).
Fig. 3.5: Shoot (a) and root (b) dry biomass of 4 weeks old maize plants grown in soil culture after nutrient seed priming. Data represent means and SE of 4 replicates. Different characters indicate significant differences between treatments. Also P priming increased shoot growth by, approximately, 20 % and
root growth by 30 % as compared with the unprimed and water primed
control plants. However, the P status was deficient in all treatments and
P priming did not significantly increase P shoot concentrations. About
50 % of the P seed reserves were found in the shoot.
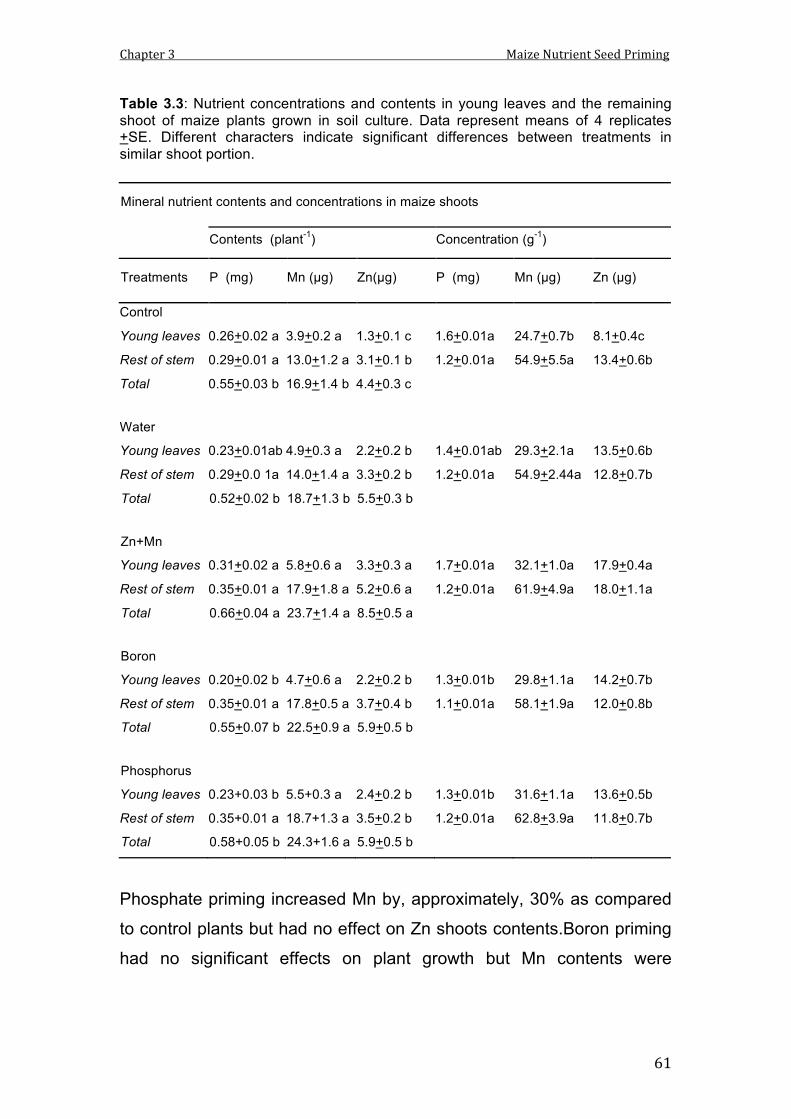
Chapter(3(((((((((((((((((((((((((((((((((((((((((((((((((((((((((((((((((((((((((((((((((((((((((((((((((((((((((((((((((Maize(Nutrient(Seed(Priming(
( 61(
Table 3.3: Nutrient concentrations and contents in young leaves and the remaining shoot of maize plants grown in soil culture. Data represent means of 4 replicates +SE. Different characters indicate significant differences between treatments in similar shoot portion.
Mineral nutrient contents and concentrations in maize shoots
Contents (plant-1) Concentration (g-1)
Treatments P (mg) Mn (µg) Zn(µg) P (mg) Mn (µg) Zn (µg)
Control
Young leaves 0.26+0.02 a 3.9+0.2 a 1.3+0.1 c 1.6+0.01a 24.7+0.7b 8.1+0.4c
Rest of stem 0.29+0.01 a 13.0+1.2 a 3.1+0.1 b 1.2+0.01a 54.9+5.5a 13.4+0.6b
Total 0.55+0.03 b 16.9+1.4 b 4.4+0.3 c
Water
Young leaves 0.23+0.01ab 4.9+0.3 a 2.2+0.2 b 1.4+0.01ab 29.3+2.1a 13.5+0.6b
Rest of stem 0.29+0.0 1a 14.0+1.4 a 3.3+0.2 b 1.2+0.01a 54.9+2.44a 12.8+0.7b
Total 0.52+0.02 b 18.7+1.3 b 5.5+0.3 b
Zn+Mn
Young leaves 0.31+0.02 a 5.8+0.6 a 3.3+0.3 a 1.7+0.01a 32.1+1.0a 17.9+0.4a
Rest of stem 0.35+0.01 a 17.9+1.8 a 5.2+0.6 a 1.2+0.01a 61.9+4.9a 18.0+1.1a
Total 0.66+0.04 a 23.7+1.4 a 8.5+0.5 a
Boron
Young leaves 0.20+0.02 b 4.7+0.6 a 2.2+0.2 b 1.3+0.01b 29.8+1.1a 14.2+0.7b
Rest of stem 0.35+0.01 a 17.8+0.5 a 3.7+0.4 b 1.1+0.01a 58.1+1.9a 12.0+0.8b
Total 0.55+0.07 b 22.5+0.9 a 5.9+0.5 b
Phosphorus
Young leaves 0.23+0.03 b 5.5+0.3 a 2.4+0.2 b 1.3+0.01b 31.6+1.1a 13.6+0.5b
Rest of stem 0.35+0.01 a 18.7+1.3 a 3.5+0.2 b 1.2+0.01a 62.8+3.9a 11.8+0.7b
Total 0.58+0.05 b 24.3+1.6 a 5.9+0.5 b
Phosphate priming increased Mn by, approximately, 30% as compared
to control plants but had no effect on Zn shoots contents.Boron priming
had no significant effects on plant growth but Mn contents were

Chapter(3(((((((((((((((((((((((((((((((((((((((((((((((((((((((((((((((((((((((((((((((((((((((((((((((((((((((((((((((((Maize(Nutrient(Seed(Priming(
( 62(
increased by approximately 20 % as compared with the unprimed
control.
3.4.5. Field experiment
In the field experiment, a significant yield increase was observed in
Zn+Mn-primed plants as compared with unprimed control plants, with
+15 % for fresh, dry and marketable grain yield. Also all water, B and P
priming resulted in slightly higher yield values. However, the differences
were not significant (Table 3.4).
Table 3.4: Final grain yield of maize grown (kg ha-1). Data represent means +SE of 4 replicates. Different characters indicate significant difference between treatments.
Grain yields (kg ha-1)
Treatments Fresh grain yield Dry grain yield Marketable grain yield
Control 11026+341 b 6920+200 b 7889+229 b
Water 11281+184 b 7069+118 b 8059+135 b
Zn+Mn 12700+173 a 7967+144 a 9083+164 a
Boron 11691+260 b 7358+180 b 8388+205 b
Phosphorus 11626+121 b 7295+95 b 8317+108 b
3.5. Discussion
3.5.1. Seed priming
Water priming and nutrient seed priming of maize seeds for 24 hours
accelerated germination and early growth of young seedlings (Fig. 3.1
ab) as previously reported also for other crops, such as rice, sorghum,
pearl millet, finger millet, cotton, beans, and maize (Harris, 1996; Harris
and Jones, 1997; Harris et al., 1999). This has been attributed to pre-
mobilization of seed reserves during the priming period. (Bewley, 1997;
Gallardo et al., 2001; Job et al., 1997), leaching of germination inhibitors
into the priming solution (Hopkins, 1995), hydrolysis of ABA (Ajouri et

Chapter(3(((((((((((((((((((((((((((((((((((((((((((((((((((((((((((((((((((((((((((((((((((((((((((((((((((((((((((((((((Maize(Nutrient(Seed(Priming(
( 63(
al., 2004) and pre-organization of membrane structures (Bewley and
Black, 1982).
Nutrient seed priming increased the seed contents of all applied
micronutrients with the highest levels for P (+195 µg seed-1), Zn (+45
µg seed-1) followed by Mn (+16 µg seed-1) and B (+6 µg seed-1),
corresponding to increases of 20%, 530 %, 800 % and 600% of the
natural seed reserves, respectively (Table 3.2). Similar increases have
been reported in earlier studies after Zn priming of barley (Ajouri et al.,
2004) and maize (Harris et al., 2007). Nutrient concentrations in the
priming solution were calculated to double the natural seed reserves
during imbibition. Much higher uptake rates recorded in case of Zn, Mn
and B suggest that during priming substantial amounts of the primed
nutrients were removed from the priming solution by adsorption to seed
structures and replacement of the removed nutrients via diffusion.
Earlier studies on Zn priming in Vicia faba revealed that nutrient
adsorption to the seed coat may substantially contribute to this process
and only 20.6 % of the primed nutrient entered the cotyledons and the
embryo, while the rest was retained by the seed coat (Polar, 1970). In
contrast, P uptake via seed priming was lower than expected. This may
be attributed to limited uptake capacity in face of P seed contents
exceeding the contents of the investigated micronutrients by 2-3 orders
of magnitude.
3.5.2. Seed priming effects in hydroponic culture
In hydroponic culture, Zn+Mn priming stimulated shoot (+50%) and root
(+100 %) biomass production (Fig. 3.2 ab) as compared with plants
supplied with complete nutrient solution particularly at final harvest at 4
weeks after sowing. This was, particularly, surprising since the nutrient
status of control plants supplied with complete nutrient solution was
always sufficient throughout the culture period.

Chapter(3(((((((((((((((((((((((((((((((((((((((((((((((((((((((((((((((((((((((((((((((((((((((((((((((((((((((((((((((((Maize(Nutrient(Seed(Priming(
( 64(
In Zn+Mn priming treatments, a critical Zn deficiency status was
reached after the 3rd harvest (Fig. 3.3) with 67 % of the Zn seed
reserves recovered in the shoot (Fig. 3.4). Manganese deficiency was
detectable already after the 2nd harvest at 3 weeks after sowing and
50% of the seed reserves were detected in the shoot tissue. However,
both, Mn and Zn shoot contents increased continuously during the
culture period of Zn+Mn primed plants, indicating an intense root to
shoot translocation of these nutrients with increasing Zn and Mn
limitation of the shoot tissue. The mobility was particularly high for Mn,
where more than 80% of the seed reserves were detected in the shoot
at final harvest.
Stimulation of root biomass production in the Zn+Mn primd plants may
be a reaction to Zn limitation (Fig. 3.3) and has been similarly observed
in soybean plants (Imran et al., unpublished). Zinc limitation can induce
inhibition of P re-translocation to the roots, thereby triggering P
deficiency responses in the root tissue, such as increased root/shoot
ratio and up-regulation of high affinity root P uptake systems even in
presence of high external P concentrations (Zhang et al., 1991; Huang
et al., 2000). Accordingly, due to stimulation of P uptake, shoot P
contents in Zn+Mn primed plants increased by 40% over the control
with complete nutrient solution as soon as shoot concentrations
reached the Zn deficiency threshold. However, the large stimulation of
shoot growth even above the control supplied with complete nutrient
solution, despite of critical levels of Zn and Mn, is a yet unexplained
phenomenon.
Shoot growth of plants obtained from B and P-primed seeds did not
reach the biomass level of control plants with complete nutrient solution
(Fig. 3.2a), but root biomass production after P priming exceeded the
control treatment at all harvests (Fig. 3.2b). This coincided with P
limitation already at the 2nd harvest in P-primed plants suggesting that P
deficiency stimulated root growth and inhibited shoot biomass

Chapter(3(((((((((((((((((((((((((((((((((((((((((((((((((((((((((((((((((((((((((((((((((((((((((((((((((((((((((((((((((Maize(Nutrient(Seed(Priming(
( 65(
production. Boron deficiency in B primed plants was detectable at the
2nd harvest but without pronounced effects on biomass production.
In contrast to Zn+Mn priming, plants obtained from P and B-primed
seeds did not continuously increase B and P shoot contents (Fig. 3.4)
during the culture period, suggesting, only a limited distribution from
seed or root reserves to the shoot. About 50 % of the P reserves in P
primed seeds were translocated to the shoot. Since P is a macronutrient
this was obviously not sufficient to cover the P demand of the shoot
tissue at later stages of plant development (2nd and 3rd harvest). Even
more extremely, only 7 % of the B reserves in B-primed seeds reached
the shoot tissue during the culture period. Thus, the success of P and B
seed priming is limited mainly by the high demand in case of P and by
limited mobility in case of B.
3.5.3. Seed priming effects in soil culture
In soil culture, plants suffered from P and Zn deficiency in all treatments
(Table 3.3), despite of additional P fertilisation, demonstrating the high
P fixation capacity of the calcareous subsoil. In contrast to plants grown
in nutrient solution, in Zn+Mn primed soil-grown plants only 15% of the
Zn seed reserves reached the shoot tissue (≈ 8µg) and this level was
even lower in water-primed and unprimed plants (≈4 µg) (Table 3).
These findings suggest also a very low Zn availability of the respective
soil. Similarly, Cenc et al., (2000) reported that plants grown on low Zn
soils from seeds with high Zn contents have shown greater Zn shoot
concentrations than those grown from low Zn seeds.
The soil culture system, however, obviously also affected the Zn
distribution within the plants leading to much lower Zn supply to the
shoots as compared with the hydroponic culture system. In calcareous
soils, plants are facing various stress factors such as P and Fe
limitation, high pH and bicarbonate levels with the potential to induce
the accumulation of carboxylates and phenolics in the root tissue

Chapter(3(((((((((((((((((((((((((((((((((((((((((((((((((((((((((((((((((((((((((((((((((((((((((((((((((((((((((((((((((Maize(Nutrient(Seed(Priming(
( 66(
(Neumann and Römheld, 2007). The chelating properties of these
compounds may promote Zn sequestration in the root tissue and limit
translocation to the shoot as similarly reported by Forno et al., (1975)
and Dogar and Hai, (1980). Therefore, the marginal surplus of Zn
supply to the shoot tissue by Zn+Mn priming in soil-grown plants also
induced only a moderate stimulation of shoot biomass production
(+18%) as compared to plants grown in hydroponics (+50%). An intense
stimulation of root growth as observed in hydroponics was absent in soil
culture. This may be also attributed to additional stress factors in
calcareous soils, e.g. high pH, high bicarbonate levels (Neumann and
Römheld, 2007).
In contrast to Zn, Mn supply was sufficient in all treatments and the
shoot contents of Mn primed seeds exceeded the seed reserves (Table
3.3), demonstrating efficient additional Mn acquisition via the roots. Also
P priming increased shoot growth by approximately 20 % and root
growth by 30 % as compared with the unprimed and water primed
control and similar to nutrient solution culture about 50 % of the P seed
reserves were found in the shoot (Table 3.3). Obviously, on the highly P
fixing soil, P priming had a certain potential to mitigate P limitation,
which was even present after additional P fertilisation. However, the
moderate surplus of primed P was immediately transformed into plant
growth and the P nutritional status at final harvest was not improved.
Similarly Derrick and Ryan, (1998) and Zhu and Smith, (2001) reported
that high P seed contents improved seedling growth under P-deficient
soil conditions and also enhanced the utilization efficiency of fertilizer P.
Phosphorus priming increased Mn, but not Zn shoot contents, by
approximately 30% as compared to control plants. This may be a
consequence of improved Mn acquisition due to the observed
improvement of root growth in P-primed plants. Zinc availability in the
respective soil was obviously too low to observe a significant effect on
Zn acquisition in this case.

Chapter(3(((((((((((((((((((((((((((((((((((((((((((((((((((((((((((((((((((((((((((((((((((((((((((((((((((((((((((((((((Maize(Nutrient(Seed(Priming(
( 67(
3.5.4. Seed priming effects under field conditions
In a field experiment, conducted with maize in 2010 on a silty-clay
Luvisol with pH (CaCl2) 7.3 fertilized according to farmer’s practice,
micronutrient (Zn+Mn) seed priming was efficient enough to produce a
15 % surplus in final grain yield (Table 3.4). The may be attributed to
the stimulation of vegetative plant growth (Fig. 3.2 and 3.5) observed in
the model experiments. On the field soil with higher nutrient availability
as compared to the extremely nutrient-deficient soil used in the model
experiment, stimulatory effects on root growth (Fig. 3.5) may promote
nutrient acquisition. This may act as an advantage not only during early
seedling development but also during reproductive and grain filling
stages. Similar long-lasting effects of seed priming treatments on yield
formation have been reported for maize (Harris et al., (2007), chickpea
and wheat (Arif et al., 2007).
Although fertilisation and sowing was performed according to good
farmers practice, field-grown maize plants in the present study were
facing low temperature stress (Fig. 3.6) during germination and early
growth, a stress factor which can severely impair root growth and
nutrient acquisition of young maize plants in temperate climates (Engels
and Marschner, 1996).
In face of the great importance of Zn and Mn in stress tolerance of
higher plants (Cakmak and Marschner, 1988 a, b, 1992; Cakmak,
2000), it remains to be established whether Zn and Mn seed priming
can exert positive effects to mitigate low temperature stress during
germination and early growth.
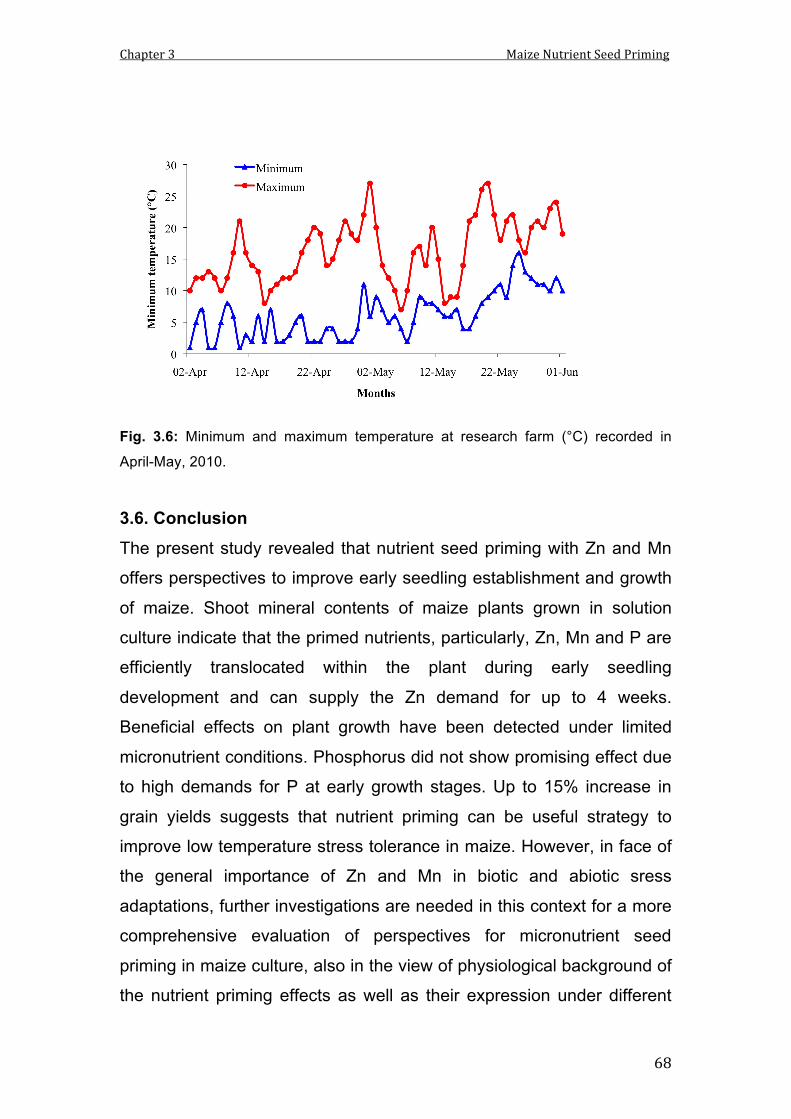
Chapter(3(((((((((((((((((((((((((((((((((((((((((((((((((((((((((((((((((((((((((((((((((((((((((((((((((((((((((((((((((Maize(Nutrient(Seed(Priming(
( 68(
Fig. 3.6: Minimum and maximum temperature at research farm (°C) recorded in
April-May, 2010.
3.6. Conclusion
The present study revealed that nutrient seed priming with Zn and Mn
offers perspectives to improve early seedling establishment and growth
of maize. Shoot mineral contents of maize plants grown in solution
culture indicate that the primed nutrients, particularly, Zn, Mn and P are
efficiently translocated within the plant during early seedling
development and can supply the Zn demand for up to 4 weeks.
Beneficial effects on plant growth have been detected under limited
micronutrient conditions. Phosphorus did not show promising effect due
to high demands for P at early growth stages. Up to 15% increase in
grain yields suggests that nutrient priming can be useful strategy to
improve low temperature stress tolerance in maize. However, in face of
the general importance of Zn and Mn in biotic and abiotic sress
adaptations, further investigations are needed in this context for a more
comprehensive evaluation of perspectives for micronutrient seed
priming in maize culture, also in the view of physiological background of
the nutrient priming effects as well as their expression under different

Chapter(3(((((((((((((((((((((((((((((((((((((((((((((((((((((((((((((((((((((((((((((((((((((((((((((((((((((((((((((((((Maize(Nutrient(Seed(Priming(
( 69(
environmental conditions (e.g. soil type, soil-water and redox status,
fertilisation management, variation in priming treatments and
genotypes).

Chapter(4((((((((((((((((((((((((((((((((((((((((((((((((((((((((((((((((((((((((((((((((((((((((((((((((((((((((((((((((((((((((((((((((((((((((((((Maize(at(Low(RZT(
( 70(
Chapter 4
Nutrient seed priming improves seedling development of maize
exposed to low root zone temperatures during early growth
(European Journal of Agronomy. (2013) 49: 141-148)
Authors
Muhammad Imran, Asim Mahmood, Volker Römheld, Günter Neumann
Institute of Crop Science, Nutritional Crop Physiology (340h),
Universität Hohenheim, 70593 Stuttgart, Germany
Corresponding author: Muhammad Imran
Institute of Crop Science, Nutritional Crop Physiology (340h)
Universität Hohenheim, 70599, Stuttgart, Germany
Phone: 0049 152 11325368 Fax: 0049 711 459 23295
E-mail: [email protected]
Contribution in the manuscript:
Planning of experiments, Preparation of experimental materials, Nutrient
analysis, Root length measurements, Conducting field trials, Statistical
analysis, Manuscript writing.

Chapter(4((((((((((((((((((((((((((((((((((((((((((((((((((((((((((((((((((((((((((((((((((((((((((((((((((((((((((((((((((((((((((((((((((((((((((((Maize(at(Low(RZT(
( 71(
4.1. Abstract
Low root zone temperature (RZT) in early spring is a major constraint for
maize production in Central and Northern Europe. Nutrient acquisition,
nutrient uptake and particularly root growth are severely reduced at low
RZT and the consequences of these growth depressions are frequently
not completely compensated until final harvest. Perspectives to overcome
these limitations by seed priming treatments with different micronutrients
(Fe, Zn, Mn) were studied with maize seedlings exposed to low RZT
(12°C).
Model experiments were performed in nutrient solution and soil culture
using rhizo-boxes with root observation windows under green house
conditions. To observe effects on final grain yield, additionally two field
experiments were conducted in 2010 and 2011. Nutrient seed priming
resulted in a significant increase in seed contents of the respective
nutrients i.e. Fe (25 %), Zn (500%) and Mn (800%). At low RZT, biomass
production and total root length of maize plants were significantly
increased after Fe and Zn+Mn priming treatments, both in nutrient
solution and in rhizo-box culture. There was no prominent difference in
shoot Fe, Zn, Mn and P concentrations but total shoot contents per plant
were significantly increased after nutrient seed priming. Plant growth
promotion and improved micronutrient status was detectable also under
field conditions at 5 weeks after sowing. This offers perspectives for using
micronutrient seed priming for improving early seedling development and
plant nutrient status of maize under low temperature climatic conditions.
Keywords: Maize, nutrient seed priming, low root zone temperature, root
growth, grain yield.

Chapter(4((((((((((((((((((((((((((((((((((((((((((((((((((((((((((((((((((((((((((((((((((((((((((((((((((((((((((((((((((((((((((((((((((((((((((((Maize(at(Low(RZT(
( 72(
4.2. Introduction
Low temperature at early spring is a major yield-limiting factor for maize
grown in Central and Northern Europe. Temperature between 25-30°C is
optimal for maize growth i.e. germination, shoot and root growth, leaf
development and expansion (Miedema et al., 1987; Duncan and Hesketh,
1968; Muldoon, et al., 1984). Foyer et al., (2002) discussed limited
capacity of maize plants to acclimatize to low temperature during early
growth. Maize seed germination is significantly reduced at low soil
temperature (Miedema, 1982). Seedling development largely depends on
soil temperature from the very beginning as the shoot meristem remains
belowground up to the V6 stage (Fortin and Pierce, 1991; Ritchie et al.,
1986; Stone et al., 1999). Maize growth is severely impaired below 15°C
(Miedema, 1982; Verheul et al., 1996). Mozafer et al. (1993) reported
stagnant growth of maize plants at low root zone temperature (9°C) at
varying photoperiods (6, 12 and 18 hours). Air temperature below 10°C
increases necrosis of maize leaves and reduces plant survival drastically
(Janowiak and Markowski, 1994). At temperatures below 15°C
development of secondary root branches and root dry biomass production
significantly declines (Cutforth et al., 1986) but root dry biomass seems to
be less affected as compared to root length (Kaspar and Boland, 1992).
Nutrient acquisition at early developmental stages is particularly limited by
low temperature. Low RZT involves two major problems with respect to
nutrient uptake: (i) limited solubility and availability of certain nutrients
(Mengel and Kirkby, 1987; Salisbury and Ross, 1992) and (ii) limited root
activity (Sowinski and Maleszewski, 1989; Pregitzer and King, 2005). Low
soil temperature is associated with declining solubility of nutrients in the
soil solution and reduction in the speed of diffusion. Therefore, sparingly
soluble nutrients such as P, K, NH4+, Fe, Zn, Mn, and Cu mainly delivered
by diffusion (Barber, 1984; Jungk, 1991; Martin et al., 1965; Giordano and
Mortvedt, 1978) are more strongly affected. Viscosity of water is inversely
related to soil temperature (Brown and LeMay, 1981). This increase in

Chapter(4((((((((((((((((((((((((((((((((((((((((((((((((((((((((((((((((((((((((((((((((((((((((((((((((((((((((((((((((((((((((((((((((((((((((((((Maize(at(Low(RZT(
( 73(
water viscosity reduces the speed of transport processes of mineral
nutrients from soil to the roots also via mass flow (Kramer and Boyer,
1995; Wan et al., 2001). Additionally, the root itself is affected by low RZT
by inhibition of root growth for spatial nutrient acquisition (Chassot and
Richner, 2002; Hund et al., 2007), inhibition of adaptive root exudation for
nutrient mobilisation in soils, e.g. release of phytosiderophores for Fe and
Zn acquisition (Marschner et al., 1987) and inhibition of nutrient uptake
and root to shoot translocation of nutrients. According to Ehdaie et al.
(2010) there is a positive correlation between root biomass production and
uptake of P and K. At low root zone temperature, also Zn uptake and
translocation from root to shoot is impaired, which may result in elevated
Zn root concentrations (Ellis et al., 1964; Sharma et al., 1968; Schwartz
et al., 1987; Moraghan and Mascagni, 1991). Engels and Marschner
(1996) reported decreased net translocation rates of Zn and Mn. Also Fe
uptake is affected under low temperature conditions. Iron concentration in
maize shoots and roots significantly declined with decreasing root zone
temperature, and shoot Fe concentration was further decreased when
plants were exposed to longer photoperiods (Mozafar et al., 1993).
According to Engels and Marschner (1996), the net root to shoot
translocation rate of Fe is markedly enhanced when shoot is grown at
24°C even in presence of low root zone temperature and similar results
have been reported for Cu, Ca, K and N, suggesting a regulation of the
translocation by the shoot demand. By contrast, translocation of Zn, Mn
and P was affected by low RZT but not by the shoot temperature,
indicating a general limitation of acquisition and uptake of these nutrients
under conditions of low RZT (Engels and Marschner, 1996, 1991).
Only strong and vigorous maize seedlings can cope with cold and wet
weather conditions, frequently observed in Central and Northern Europe.
Well-developed shoot and root systems are key features for genotypic
differences in cold tolerance (Stamp, 1984; Richner et al., 1997; Lee et
al., 2002; Hund et al., 2007). According to Cooper and McDonald (1970)

Chapter(4((((((((((((((((((((((((((((((((((((((((((((((((((((((((((((((((((((((((((((((((((((((((((((((((((((((((((((((((((((((((((((((((((((((((((((Maize(at(Low(RZT(
( 74(
maize seed reserves are exhausted, at the 3-4 leaf stage. After depletion
of seed reserves, the seedling relies on nutrient uptake from the soil. A
high density of long lateral roots can improve early vigour of young
seedlings under chilling stress conditions (Hund et al., 2007). Availability
and uptake of the limiting nutrients can be increased by starter
fertilization. In soils with limited P availability, relative efficiency of P
fertilizer was increased when placed near the seed (Fiedler et al., 1983).
Seed dressing or coating of limiting nutrients is a cost effective and very
useful approach to improve early plant growth (Robert, 1973; Ros et al.,
2000). Zinc seed dressing of wheat even increased final grain yield
(Yilmaz et al., 1998). In barley, nutrient priming of seeds with P and Zn
enhanced germination, seedling growth and uptake of these elements
(Ajouri et al., 2004).
In general, seed priming is a pre-sowing seed treatment in which seeds
are soaked in water and dried back to storage moisture levels until further
use. This can help crop plants to cope with stress factors, such as
drought, and pest damage and can increase crop yield (Harris et al.,
1999; Harris et al., 2000). ‘Nutrient seed priming’ is a technique in which
seeds are soaked in nutrient solution instead of pure water as an
approach to increase seed nutrient contents along with the priming effect
to improve seed quality for better germination and seedling establishment.
Maize seed priming with 1% ZnSO4 not only enhanced plant growth but
also increased the final grain yield and seed Zn contents in plants grown
on soil with limited Zn availability (Harris et al., 2007). Recently, Guan et
al. (2009) reported that maize seed priming with ‘chitosan’ improved
germination and seedling growth under low temperature stress conditions.
In face of the multitude of functions reported for micronutrients, such as
Zn, Mn and Fe in stress resistance of higher plants (Marschner, 2012), the
aim of the present study was to investigate the effects of micronutrient
seed priming (seed priming with water, Zn+Mn and Fe) on seedling
development, early growth and yield formation in maize exposed to low

Chapter(4((((((((((((((((((((((((((((((((((((((((((((((((((((((((((((((((((((((((((((((((((((((((((((((((((((((((((((((((((((((((((((((((((((((((((((Maize(at(Low(RZT(
( 75(
root zone temperature during early growth. For this purpose three different
experimental approaches were employed. Model experiments were
performed in nutrient solution (hydroponic study) and as rhizo-box
experiments in soil culture under greenhouse conditions. Additionally two
field experiments were conduced in 2010 and 2011 to evaluate effects on
grain yield.
4.3. Materials and methods
4.3.1. Seed priming
Seeds of Zea mays L. cv F030xF047 (provided by Institute of Plant
Breeding, Seed Science and Population Genetics) were used for all
experiments. Maize seeds were primed for a pre-determined time of 24
hours for water and nutrient seed priming by soaking in distilled water and
nutrient solutions, respectively. Water absorbing capacity of maize seeds
was 0.08 ml seeds-1, about 60% of the absorbed water during priming
was removed during drying the seeds. For water priming, 60 seeds were
soaked in 200 ml of distilled water. Iron, Zn and Mn were used for nutrient
seed priming. Iron was used alone (8.5 mM Fe as Fe-EDTA), while Zn
and Mn were combined together (4mM Zn + 2.5mM Mn solution as ZnSO4
.H2O and as MnSO4 .7H2O). 60 seeds were soaked in 200 ml of
respective solutions in the dark for 24 hours. Thereafter, seeds were
taken out, rinsed with running distilled water for 1 minute to remove the
priming solutions. Subsequently, seeds were air dried at room
temperature for a minimum time period of 1 hour for easy handling and to
facilitate clump-free sowing according to on-farm priming method adopted
by Harris et al. (2007). Optimum priming conditions (priming duration and
nutrient concentrations) were determined in a separate set of experiments
(Imran et al., 2012). Seeds for all model and field experiments were
primed with the same method. Un-primed seeds were used for control
treatments. Table 4.1 summarises the various priming treatments
employed for the experiments.

Chapter(4((((((((((((((((((((((((((((((((((((((((((((((((((((((((((((((((((((((((((((((((((((((((((((((((((((((((((((((((((((((((((((((((((((((((((((Maize(at(Low(RZT(
( 76(
Table 4.1: Seed priming treatments and root zone temperature regimes of maize
seedlings grown in nutrient solution and rhizo-boxes.
4.3.2. Plant culture
The plants were grown under greenhouse conditions with an average air
temperature of 20°C, light intensities during the growing period ranged
between 250- 400 µmol m-2 s-1 PAR and relative humidity from 60-80%.
For the nutrient solution experiment, primed seeds were germinated for 7
days in rolls of filter papers moistened with distilled water before transfer
into pots containing 400 mL of aerated nutrient solution. (1 plant per pot
with 4 replicates per treatment).
Composition of the nutrient solution used in hydroponic study was: 2 mM
Ca(NO3)2, 0.7 mM K2SO4, 0.1mM KCl, 0.5 mM MgSO4, 0.1 mM KH2PO4,
10 µM H3BO3, 0.5 µM MnSO4, 0.2 µM CuSO4, 0.01 µM (NH4)6Mo7O24, 0.5
µM ZnSO4, and 20 µM Fe-EDTA. The pH varied between 6.0 and 6.5. The
nutrient solution was renewed every 2-3 days to avoid nutrient depletion.
For soil culture, plants were grown in rhizo-boxes (35 x 10 x 2 cm)
equipped with root observation windows and filled with 500 g of a silty-
clay Luvisol (organic matter 0.97% and pH (CaCl2) 7.3). The soil was
fertilized once with N, P, K and Mg. Nitrogen was applied as NH4NO3 at
100 mg N kg-1 soil, P as Ca(H2PO4)2 at 100 mg P kg-1 soil, K as K2SO4 at
150 mg K kg-1 soil and Mg as MgSO4 at 50 mg Mg kg-1 soil. Before filling
Treatment Seed priming
Root zone temperature
Control Un-primed 12 + 2°C
Water Water primed 12 + 2°C
Fe Fe priming 12 + 2°C
Zn+Mn Zn+Mn priming 12 + 2°C
Control (20°C) Un-primed 20 + 2°C

Chapter(4((((((((((((((((((((((((((((((((((((((((((((((((((((((((((((((((((((((((((((((((((((((((((((((((((((((((((((((((((((((((((((((((((((((((((((Maize(at(Low(RZT(
( 77(
the rhizo-boxes, the different fertilizers were mixed with the soil substrate
and the soil was sieved through 2 mm mesh size. The soil moisture
content was kept at 18% (w/w) by gravimetric determination. Into each
box, three seeds were sown at a depth of 2 cm and finally one plant was
kept for further cultivation. To stimulate gravitropic root growth along the
root observation window, the boxes were fixed at an angle of 60° with the
observation window on the lower side of the rhizo-box and covered with
black plastic foil.
Two field trials were conducted at the University of Hohenheim, research
station farm “Heid-feldhof”, Stuttgart, Germany in 2010 and 2011. The
field soil had the following properties: pH 6.9, Ctot 1.5%, Ntot 0.14%,
mineral N 14.6 mg kg–1, SOM 2.2%. Concentrations of plant-available
nutrients (CAL: Ca-acetate-lactate extract) were 250 mg kg–1 K, 160
mg kg–1 Mg, 225 mg kg–1 P. The soil texture was silty–loam (FAO-
UNESCO, 1997) consisting of 22.6% clay, 62.9% silt, 14.5% sand.
Mineral fertilizers were applied according to farmers practice with N 167
kg ha-1, P2O5 42 kg ha-1, K2O 42 kg ha-1, MgO 23 kg ha-1 and S 63 kg ha-1
in 2010 and N 147 kg ha-1, P2O5 38 kg ha-1, K2O 33 kg ha-1, MgO 28 kg
ha-1 and S 61 kg ha-1 in 2011. In both years, winter wheat was grown in
experimental fields before setting maize experiments. The experiment
was laid out as a randomized complete block design with four replicates.
Each plot consisted of two rows, and each row was 5 m length with a
distance of 0.75 m between rows (Bhosle et al., 2007). Sowing was
performed in the last week of April in both years. After seedling
emergence thinning was conducted to obtain a plant density of 88 000
plants ha-1. Final harvest was performed in October.
4.3.3. Low RZT stress treatment
After transferring seven days old seedlings into hydroponics, the pots
were transferred to a temperature controlled water bath and cultivated for
4 weeks with a RZT of 12°C in the greenhouse. Similarly, after a culture

Chapter(4((((((((((((((((((((((((((((((((((((((((((((((((((((((((((((((((((((((((((((((((((((((((((((((((((((((((((((((((((((((((((((((((((((((((((((Maize(at(Low(RZT(
( 78(
period of 7 days, rhizo-boxes were sealed in plastic bags and cultivated in
the water bath for 3 weeks at a RZT of 12°C. One set of 4 replicates for
both, solution and soil culture was grown at ambient temperature (Control
20°C).
4.3.4. Plant analysis
At harvest, fresh biomass of shoots and roots, and dry matter of shoots
was determined for plants grown in nutrient solution and soil culture. The
field experiment 2010 was designed as a first pilot trial. In the field
experiment (2011) intermediate sampling was performed. Shoots of 4
plants from each replicate of treatments were collected at random in an
intermediate harvest at 5 weeks after sowing to determine fresh and dry
biomass and mineral analysis. Plant dry matter (500 mg) was ashed at
500 °C for 5 h in a muffle oven. After cooling, the samples were extracted
twice with 2 mL of 3.4 M HNO3 until dryness to precipitate SiO2. The ash
was dissolved in 2 mL of 4 M HCl, subsequently diluted ten times with hot
deionized water, and boiled for 2 min. After addition of 0.1 mL Cs/La
buffer to 4.9 mL ash solution, Fe, Mn and Zn concentrations were
measured by atomic absorption spectrometry (UNICAM 939,
Offenbach/Main Germany). Phosphate in the ash solution was determined
spectro-photpmetrically by the method of Gericke and Kurmis (1952).
Total root lengths of plants grown in the nutrient solution and rhizo-boxes
were calculated. For this purpose, washed roots were stored in 30% (v/v)
ethanol solution until further analysis. For length determination of the
roots grown along the root windows in rhizo-boxes, root sketches were
drawn on transparent plastic foil, before and after the exposure to low
RZT. After digitalization with a flatbed scanner, the root analysis software
WinRhizo (WinRHIZO, Pro V. 2009, Regent Instrument Canada Inc.) was
used to determine the total root length, root diameter of the root samples
washed out from soil and root growth along the root observation windows

Chapter(4((((((((((((((((((((((((((((((((((((((((((((((((((((((((((((((((((((((((((((((((((((((((((((((((((((((((((((((((((((((((((((((((((((((((((((Maize(at(Low(RZT(
( 79(
4.3.5. Statistical analysis
A complete randomized block design with 4 replicates for each treatment
was used in all experiments. Significant differences were tested by
analysis of variance (p < 0.05), using the Sigma Stat version 3.1’software
(SYSTAT Software Inc., Erkrath, Germany).
4.4. Results 4.4.1. Nutrient seed priming effects on seed nutrient levels
Nutrient seed priming of maize seeds with Fe and particularly with Zn+Mn
significantly increased the concentrations and contents of these elements
within the seeds (Table 4.2). No significant differences in nutrient
concentrations and contents were detected between un-primed and water
primed seeds.
4.4.2. Priming effects on seedling growth at low RZT
The effects of priming treatments on seedling growth (shoot and root
fresh biomass production and root length) under low RZT conditions were
studied in nutrient solution and rhizo-box experiments conducted under
green house conditions. Plants grown in nutrient solutions at 20 °C were
more healthy and vigorous as compared to those at low RZT, irrespective
of the priming treatments. Water priming and particularly, Fe and Zn+Mn
seed priming improved seedling growth compared to unprimed seeds at
low RZT (Figure 4.1), both in nutrient solution and rhizo-box experiments
(Table 4.3).
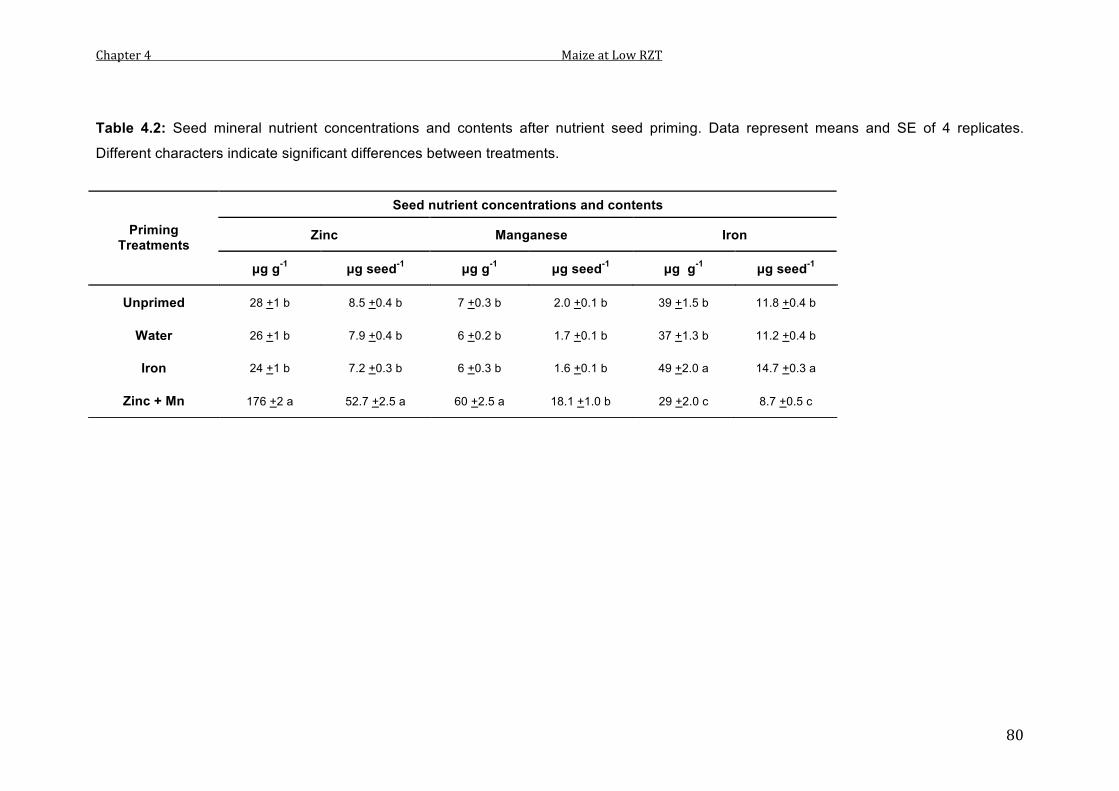
Chapter(4((((((((((((((((((((((((((((((((((((((((((((((((((((((((((((((((((((((((((((((((((((((((((((((((((((((((((((((((((((((((((((((((((((((((((((Maize(at(Low(RZT(
( 80(
Table 4.2: Seed mineral nutrient concentrations and contents after nutrient seed priming. Data represent means and SE of 4 replicates.
Different characters indicate significant differences between treatments.
Seed nutrient concentrations and contents
Zinc Manganese Iron Priming Treatments
µg g-1 µg seed-1 µg g-1 µg seed-1 µg g-1 µg seed-1
Unprimed 28 +1 b 8.5 +0.4 b 7 +0.3 b 2.0 +0.1 b 39 +1.5 b 11.8 +0.4 b
Water 26 +1 b 7.9 +0.4 b 6 +0.2 b 1.7 +0.1 b 37 +1.3 b 11.2 +0.4 b
Iron 24 +1 b 7.2 +0.3 b 6 +0.3 b 1.6 +0.1 b 49 +2.0 a 14.7 +0.3 a
Zinc + Mn 176 +2 a 52.7 +2.5 a 60 +2.5 a 18.1 +1.0 b 29 +2.0 c 8.7 +0.5 c
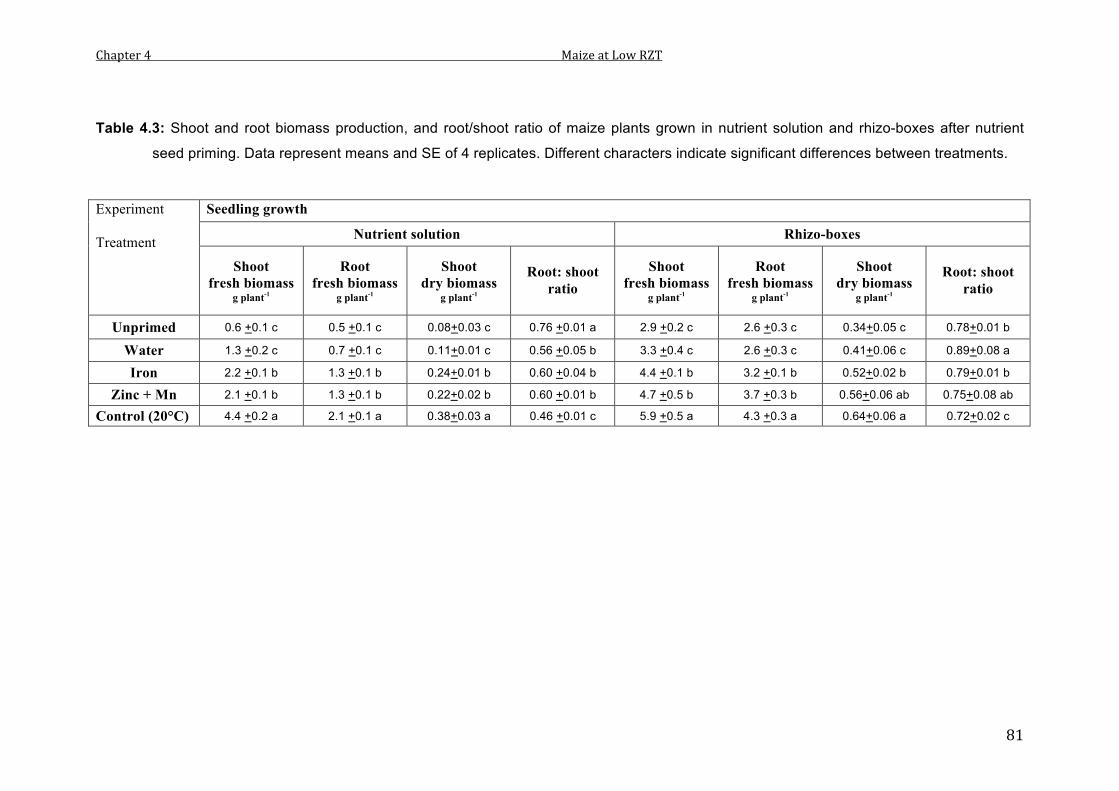
Chapter(4((((((((((((((((((((((((((((((((((((((((((((((((((((((((((((((((((((((((((((((((((((((((((((((((((((((((((((((((((((((((((((((((((((((((((((Maize(at(Low(RZT(
( 81(
Table 4.3: Shoot and root biomass production, and root/shoot ratio of maize plants grown in nutrient solution and rhizo-boxes after nutrient
seed priming. Data represent means and SE of 4 replicates. Different characters indicate significant differences between treatments.
Seedling growth
Nutrient solution Rhizo-boxes
Experiment Treatment
Shoot fresh biomass
g plant-1
Root fresh biomass
g plant-1
Shoot dry biomass
g plant-1
Root: shoot
ratio Shoot
fresh biomass g plant-1
Root fresh biomass
g plant-1
Shoot dry biomass
g plant-1
Root: shoot
ratio
Unprimed 0.6 +0.1 c 0.5 +0.1 c 0.08+0.03 c 0.76 +0.01 a 2.9 +0.2 c 2.6 +0.3 c 0.34+0.05 c 0.78+0.01 b
Water 1.3 +0.2 c 0.7 +0.1 c 0.11+0.01 c 0.56 +0.05 b 3.3 +0.4 c 2.6 +0.3 c 0.41+0.06 c 0.89+0.08 a
Iron 2.2 +0.1 b 1.3 +0.1 b 0.24+0.01 b 0.60 +0.04 b 4.4 +0.1 b 3.2 +0.1 b 0.52+0.02 b 0.79+0.01 b
Zinc + Mn 2.1 +0.1 b 1.3 +0.1 b 0.22+0.02 b 0.60 +0.01 b 4.7 +0.5 b 3.7 +0.3 b 0.56+0.06 ab 0.75+0.08 ab
Control (20°C) 4.4 +0.2 a 2.1 +0.1 a 0.38+0.03 a 0.46 +0.01 c 5.9 +0.5 a 4.3 +0.3 a 0.64+0.06 a 0.72+0.02 c
(

Chapter(4((((((((((((((((((((((((((((((((((((((((((((((((((((((((((((((((((((((((((((((((((((((((((((((((((((((((((((((((((((((((((((((((((((((((((((Maize(at(Low(RZT(
( 82(
(Fig. 4.1: Five weeks old maize plants grown in nutrient solution (hydroponic
experiment) under low RZT stress.
1Under low RZT conditions, seedlings grown in rhizo-boxes were higher in
shoot and root fresh biomass and root length than the seedlings grown in
nutrient solution. In the nutrient solution experiment, there was almost 60-
70% increase in shoot and root fresh biomass and root length with Fe and
Zn+Mn priming as compared to water priming alone, whereas for the
seedlings grown in the rhizo-boxes, this difference was reduced to 30%
for shoot and root fresh biomass production (Table 4.3). Root to shoot
ratio was significantly increased for the plants grown at low RZT
compared to 20°C control plants, both, in solution and soil culture (Table
4.3). In both culture systems, Fe and Zn+Mn priming resulted in an
approximately 100% increase in total root length after 3-4 weeks exposure
to low RZT (Table 4.4). Root length in different diameter classes (0.2, 0.4
and 0.6 mm) revealed that nutrient seed priming particularly promoted the
growth of fine roots, as compared to un-primed and water primed
treatments at low RZT. There was more than 80% increase in root length
of the diameter class ≤ 0.2 mm after nutrient seed priming (Table 4.4). In ((((((((((((((((((((((((((((((((((((((((((((((((((((((((1((
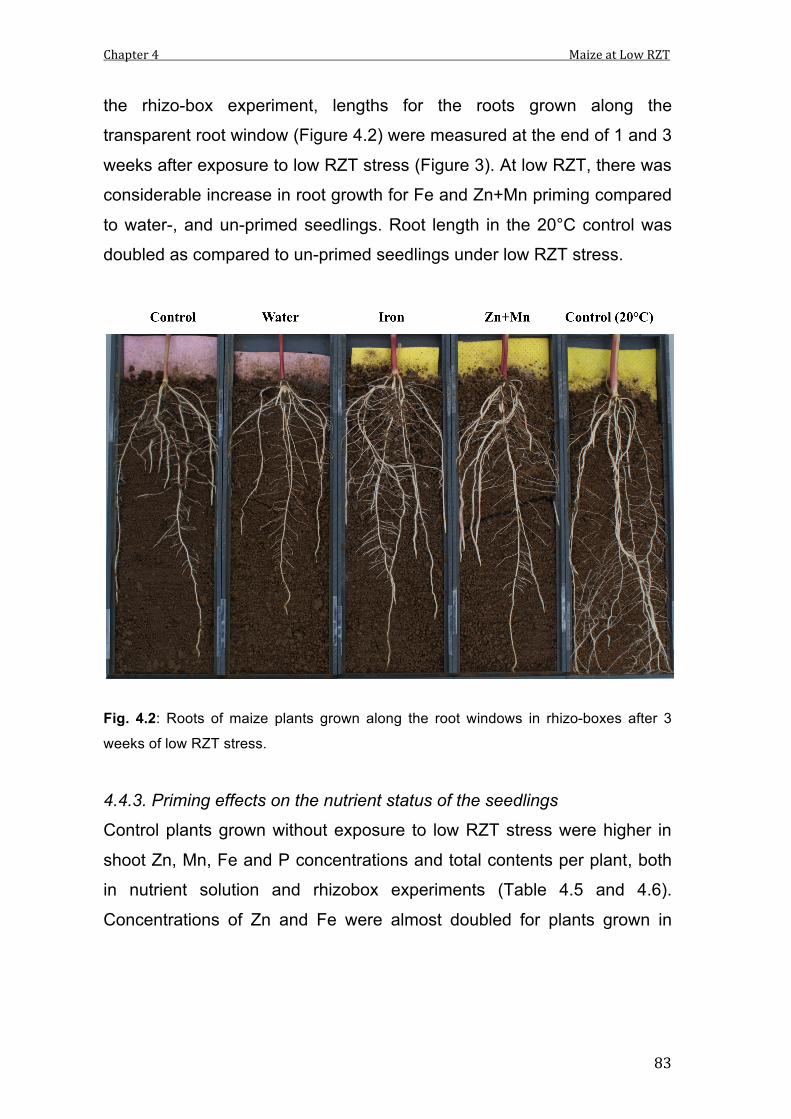
Chapter(4((((((((((((((((((((((((((((((((((((((((((((((((((((((((((((((((((((((((((((((((((((((((((((((((((((((((((((((((((((((((((((((((((((((((((((Maize(at(Low(RZT(
( 83(
the rhizo-box experiment, lengths for the roots grown along the
transparent root window (Figure 4.2) were measured at the end of 1 and 3
weeks after exposure to low RZT stress (Figure 3). At low RZT, there was
considerable increase in root growth for Fe and Zn+Mn priming compared
to water-, and un-primed seedlings. Root length in the 20°C control was
doubled as compared to un-primed seedlings under low RZT stress.
(
Fig. 4.2: Roots of maize plants grown along the root windows in rhizo-boxes after 3
weeks of low RZT stress.
4.4.3. Priming effects on the nutrient status of the seedlings
Control plants grown without exposure to low RZT stress were higher in
shoot Zn, Mn, Fe and P concentrations and total contents per plant, both
in nutrient solution and rhizobox experiments (Table 4.5 and 4.6).
Concentrations of Zn and Fe were almost doubled for plants grown in
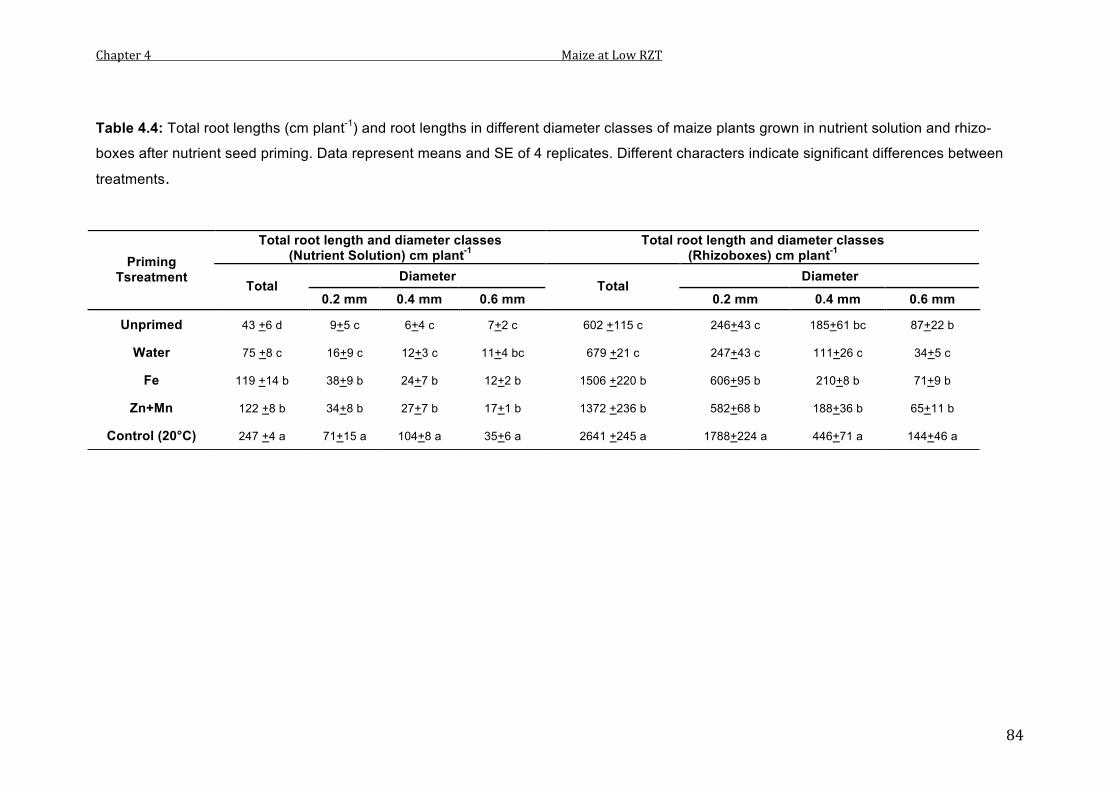
Chapter(4((((((((((((((((((((((((((((((((((((((((((((((((((((((((((((((((((((((((((((((((((((((((((((((((((((((((((((((((((((((((((((((((((((((((((((Maize(at(Low(RZT(
( 84(
Table 4.4: Total root lengths (cm plant-1) and root lengths in different diameter classes of maize plants grown in nutrient solution and rhizo-
boxes after nutrient seed priming. Data represent means and SE of 4 replicates. Different characters indicate significant differences between
treatments.
Total root length and diameter classes (Nutrient Solution) cm plant-1
Total root length and diameter classes (Rhizoboxes) cm plant-1
Diameter Diameter Priming
Tsreatment Total
0.2 mm 0.4 mm 0.6 mm Total
0.2 mm 0.4 mm 0.6 mm
Unprimed 43 +6 d 9+5 c 6+4 c 7+2 c 602 +115 c 246+43 c 185+61 bc 87+22 b
Water 75 +8 c 16+9 c 12+3 c 11+4 bc 679 +21 c 247+43 c 111+26 c 34+5 c
Fe 119 +14 b 38+9 b 24+7 b 12+2 b 1506 +220 b 606+95 b 210+8 b 71+9 b
Zn+Mn 122 +8 b 34+8 b 27+7 b 17+1 b 1372 +236 b 582+68 b 188+36 b 65+11 b
Control (20°C) 247 +4 a 71+15 a 104+8 a 35+6 a 2641 +245 a 1788+224 a 446+71 a 144+46 a
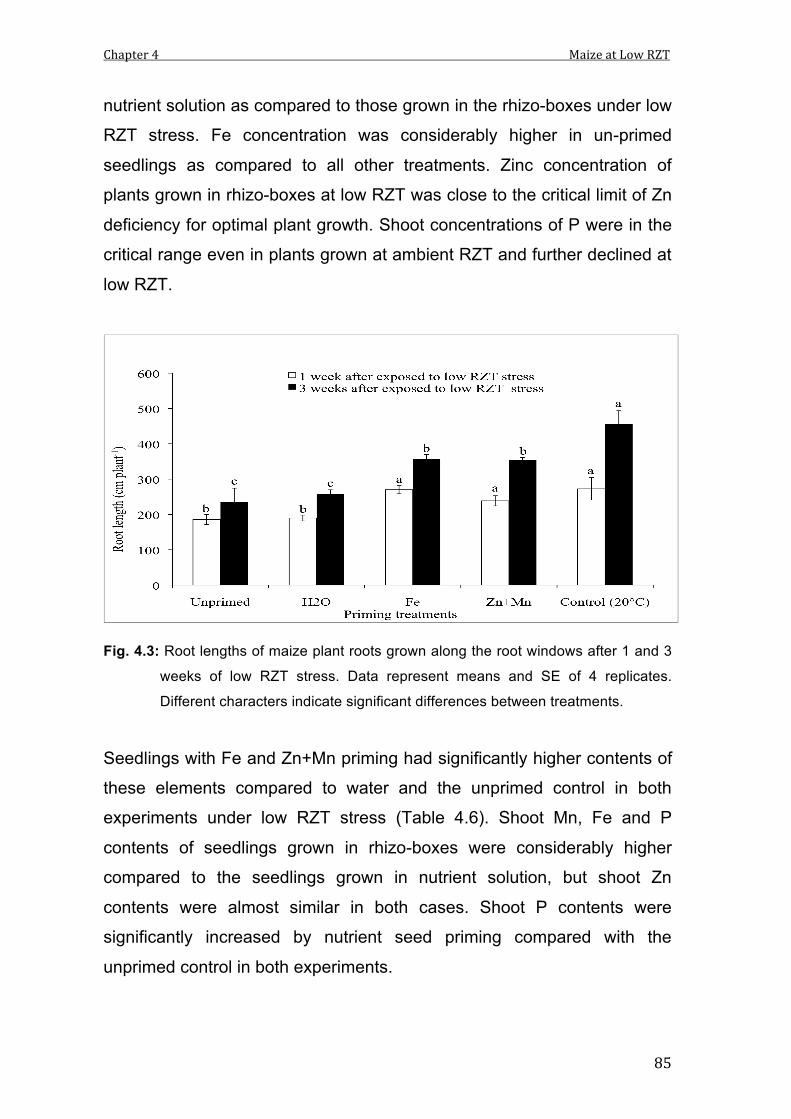
Chapter(4((((((((((((((((((((((((((((((((((((((((((((((((((((((((((((((((((((((((((((((((((((((((((((((((((((((((((((((((((((((((((((((((((((((((((((Maize(at(Low(RZT(
( 85(
nutrient solution as compared to those grown in the rhizo-boxes under low
RZT stress. Fe concentration was considerably higher in un-primed
seedlings as compared to all other treatments. Zinc concentration of
plants grown in rhizo-boxes at low RZT was close to the critical limit of Zn
deficiency for optimal plant growth. Shoot concentrations of P were in the
critical range even in plants grown at ambient RZT and further declined at
low RZT.
(
(Fig. 4.3: Root lengths of maize plant roots grown along the root windows after 1 and 3
weeks of low RZT stress. Data represent means and SE of 4 replicates.
Different characters indicate significant differences between treatments.
Seedlings with Fe and Zn+Mn priming had significantly higher contents of
these elements compared to water and the unprimed control in both
experiments under low RZT stress (Table 4.6). Shoot Mn, Fe and P
contents of seedlings grown in rhizo-boxes were considerably higher
compared to the seedlings grown in nutrient solution, but shoot Zn
contents were almost similar in both cases. Shoot P contents were
significantly increased by nutrient seed priming compared with the
unprimed control in both experiments.
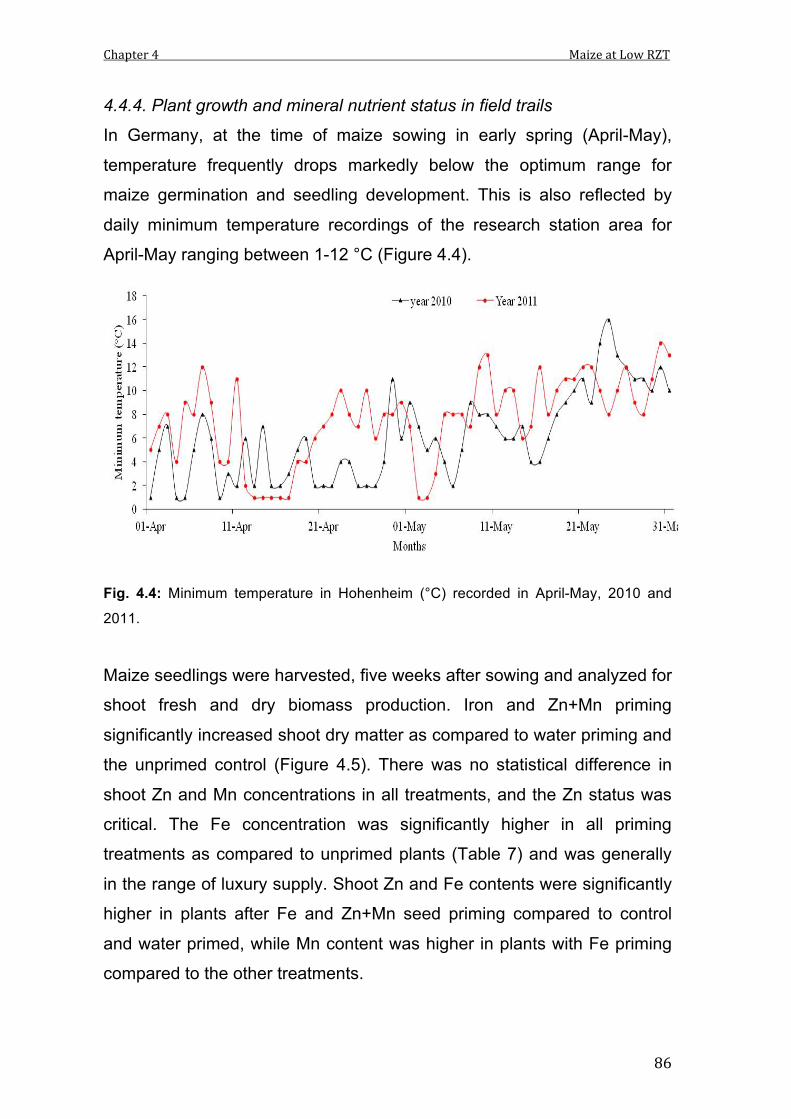
Chapter(4((((((((((((((((((((((((((((((((((((((((((((((((((((((((((((((((((((((((((((((((((((((((((((((((((((((((((((((((((((((((((((((((((((((((((((Maize(at(Low(RZT(
( 86(
4.4.4. Plant growth and mineral nutrient status in field trails In Germany, at the time of maize sowing in early spring (April-May),
temperature frequently drops markedly below the optimum range for
maize germination and seedling development. This is also reflected by
daily minimum temperature recordings of the research station area for
April-May ranging between 1-12 °C (Figure 4.4).
Fig. 4.4: Minimum temperature in Hohenheim (°C) recorded in April-May, 2010 and
2011.
Maize seedlings were harvested, five weeks after sowing and analyzed for
shoot fresh and dry biomass production. Iron and Zn+Mn priming
significantly increased shoot dry matter as compared to water priming and
the unprimed control (Figure 4.5). There was no statistical difference in
shoot Zn and Mn concentrations in all treatments, and the Zn status was
critical. The Fe concentration was significantly higher in all priming
treatments as compared to unprimed plants (Table 7) and was generally
in the range of luxury supply. Shoot Zn and Fe contents were significantly
higher in plants after Fe and Zn+Mn seed priming compared to control
and water primed, while Mn content was higher in plants with Fe priming
compared to the other treatments.
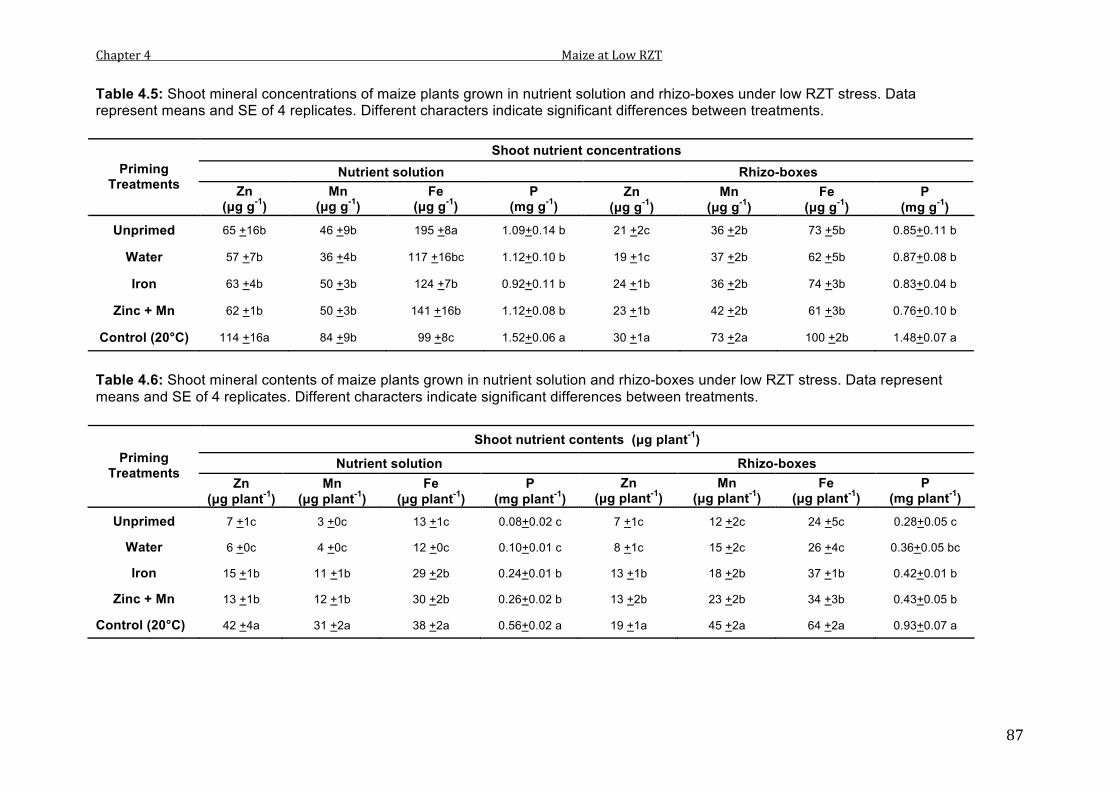
�� ����������������������������������������������������������������������������������������������������������������������������������������������������������������
� ��
Table 4.5: Shoot mineral concentrations of maize plants grown in nutrient solution and rhizo-boxes under low RZT stress. Data represent means and SE of 4 replicates. Different characters indicate significant differences between treatments.
Shoot nutrient concentrations Nutrient solution Rhizo-boxes Priming
Treatments Zn (µg g-1)
Mn (µg g-1)
Fe (µg g-1)
P (mg g-1)
Zn (µg g-1)
Mn (µg g-1)
Fe (µg g-1)
P (mg g-1)
Unprimed 65 +16b 46 +9b 195 +8a 1.09+0.14 b 21 +2c 36 +2b 73 +5b 0.85+0.11 b
Water 57 +7b 36 +4b 117 +16bc 1.12+0.10 b 19 +1c 37 +2b 62 +5b 0.87+0.08 b
Iron 63 +4b 50 +3b 124 +7b 0.92+0.11 b 24 +1b 36 +2b 74 +3b 0.83+0.04 b
Zinc + Mn 62 +1b 50 +3b 141 +16b 1.12+0.08 b 23 +1b 42 +2b 61 +3b 0.76+0.10 b
Control (20°C) 114 +16a 84 +9b 99 +8c 1.52+0.06 a 30 +1a 73 +2a 100 +2b 1.48+0.07 a
Table 4.6: Shoot mineral contents of maize plants grown in nutrient solution and rhizo-boxes under low RZT stress. Data represent means and SE of 4 replicates. Different characters indicate significant differences between treatments.
Shoot nutrient contents (µg plant-1)
Nutrient solution Rhizo-boxes Priming Treatments
Zn (µg plant-1)
Mn (µg plant-1)
Fe (µg plant-1)
P (mg plant-1)
Zn (µg plant-1)
Mn (µg plant-1)
Fe (µg plant-1)
P (mg plant-1)
Unprimed 7 +1c 3 +0c 13 +1c 0.08+0.02 c 7 +1c 12 +2c 24 +5c 0.28+0.05 c
Water 6 +0c 4 +0c 12 +0c 0.10+0.01 c 8 +1c 15 +2c 26 +4c 0.36+0.05 bc
Iron 15 +1b 11 +1b 29 +2b 0.24+0.01 b 13 +1b 18 +2b 37 +1b 0.42+0.01 b
Zinc + Mn 13 +1b 12 +1b 30 +2b 0.26+0.02 b 13 +2b 23 +2b 34 +3b 0.43+0.05 b
Control (20°C) 42 +4a 31 +2a 38 +2a 0.56+0.02 a 19 +1a 45 +2a 64 +2a 0.93+0.07 a
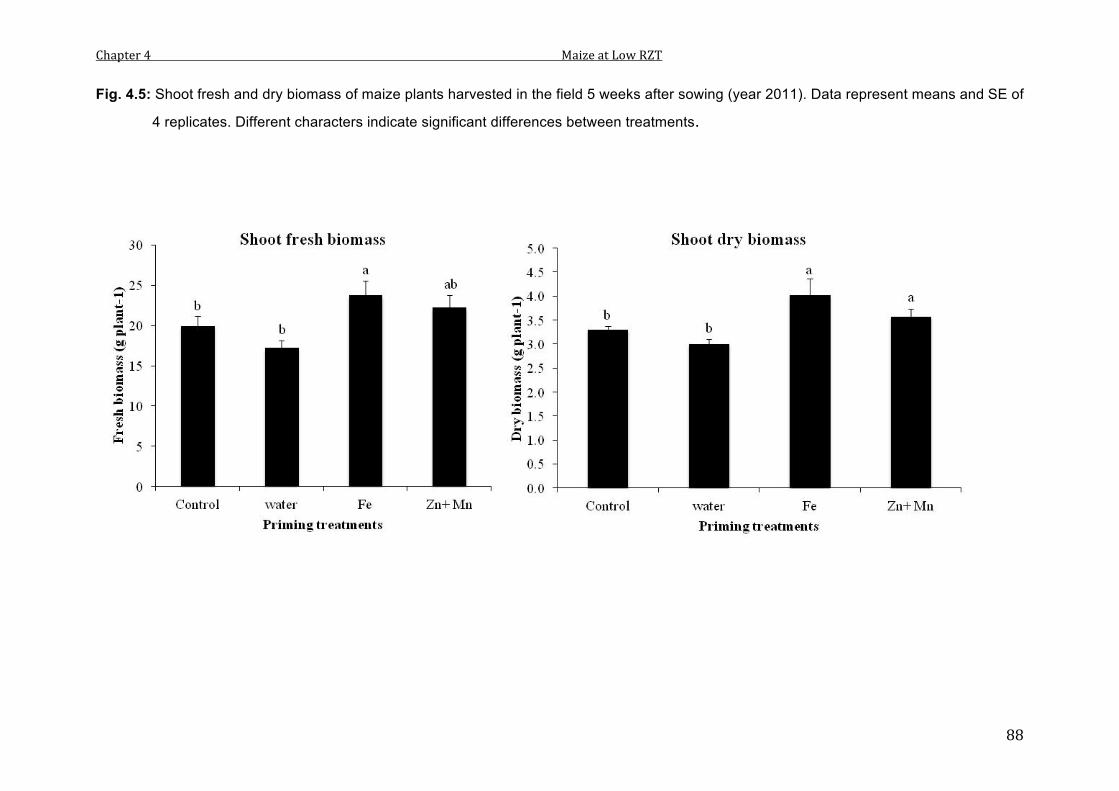
�� ����������������������������������������������������������������������������������������������������������������������������������������������������������������
� ��
Fig. 4.5: Shoot fresh and dry biomass of maize plants harvested in the field 5 weeks after sowing (year 2011). Data represent means and SE of
4 replicates. Different characters indicate significant differences between treatments. �
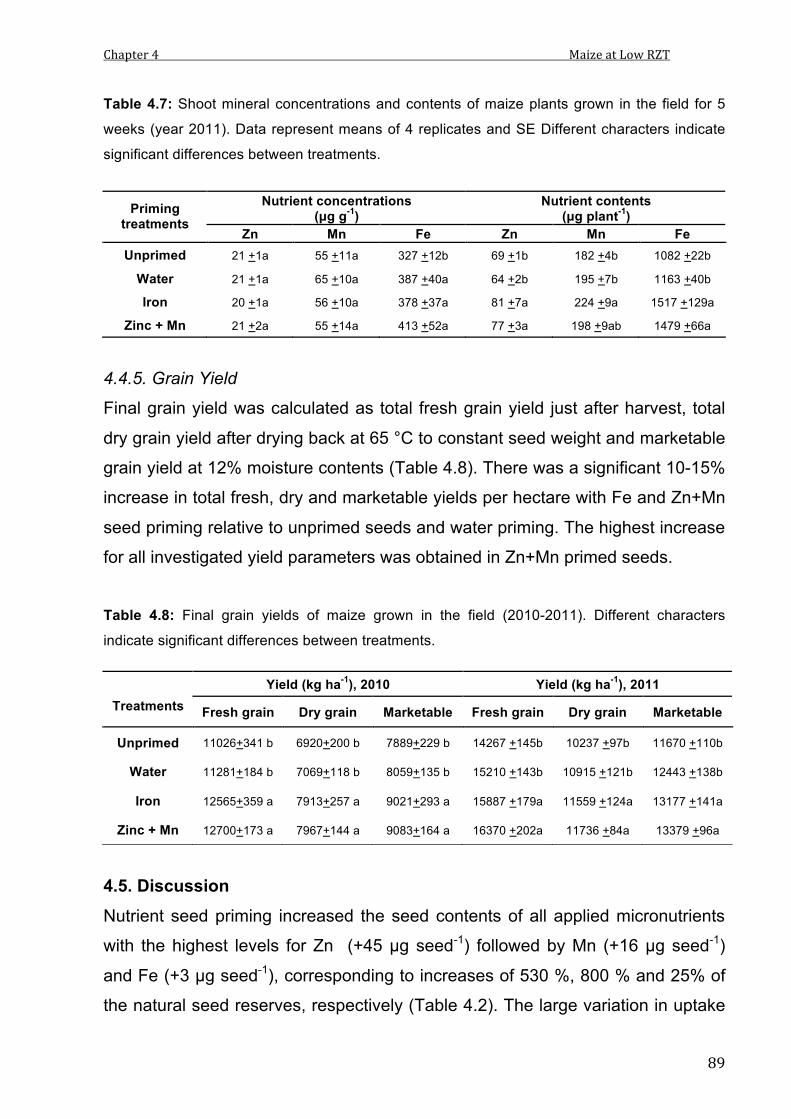
Chapter(4((((((((((((((((((((((((((((((((((((((((((((((((((((((((((((((((((((((((((((((((((((((((((((((((((((((((((((((((((((((((((((((((((((((((((((Maize(at(Low(RZT(
( 89(
Table 4.7: Shoot mineral concentrations and contents of maize plants grown in the field for 5
weeks (year 2011). Data represent means of 4 replicates and SE Different characters indicate
significant differences between treatments.
4.4.5. Grain Yield
Final grain yield was calculated as total fresh grain yield just after harvest, total
dry grain yield after drying back at 65 °C to constant seed weight and marketable
grain yield at 12% moisture contents (Table 4.8). There was a significant 10-15%
increase in total fresh, dry and marketable yields per hectare with Fe and Zn+Mn
seed priming relative to unprimed seeds and water priming. The highest increase
for all investigated yield parameters was obtained in Zn+Mn primed seeds.
Table 4.8: Final grain yields of maize grown in the field (2010-2011). Different characters
indicate significant differences between treatments.
4.5. Discussion
Nutrient seed priming increased the seed contents of all applied micronutrients
with the highest levels for Zn (+45 µg seed-1) followed by Mn (+16 µg seed-1)
and Fe (+3 µg seed-1), corresponding to increases of 530 %, 800 % and 25% of
the natural seed reserves, respectively (Table 4.2). The large variation in uptake
Nutrient concentrations (µg g-1)
Nutrient contents (µg plant-1) Priming
treatments Zn Mn Fe Zn Mn Fe
Unprimed 21 +1a 55 +11a 327 +12b 69 +1b 182 +4b 1082 +22b
Water 21 +1a 65 +10a 387 +40a 64 +2b 195 +7b 1163 +40b
Iron 20 +1a 56 +10a 378 +37a 81 +7a 224 +9a 1517 +129a
Zinc + Mn 21 +2a 55 +14a 413 +52a 77 +3a 198 +9ab 1479 +66a
Yield (kg ha-1), 2010 Yield (kg ha-1), 2011 Treatments Fresh grain Dry grain Marketable Fresh grain Dry grain Marketable
Unprimed 11026+341 b 6920+200 b 7889+229 b 14267 +145b 10237 +97b 11670 +110b
Water 11281+184 b 7069+118 b 8059+135 b 15210 +143b 10915 +121b 12443 +138b
Iron 12565+359 a 7913+257 a 9021+293 a 15887 +179a 11559 +124a 13177 +141a
Zinc + Mn 12700+173 a 7967+144 a 9083+164 a 16370 +202a 11736 +84a 13379 +96a

Chapter(4((((((((((((((((((((((((((((((((((((((((((((((((((((((((((((((((((((((((((((((((((((((((((((((((((((((((((((((((((((((((((((((((((((((((((((Maize(at(Low(RZT(
( 90(
within the different applied nutrients despite of similar concentrations of
micronutrients in the priming solutions (2.5 - 8.5 mM) requires further
investigations on uptake kinetics and compartmentation within the seeds. Based
on calculations of nutrient concentrations (Table 2), similar increases have been
reported in earlier studies after Zn priming of barley (Ajouri et al., 2004) and
maize (Harris et al., 2007).
Both, in hydroponics and in soil culture, plant exposure to low RZT without
priming treatments resulted in a severe reduction of biomass production by 75-
85% and 40-50% respectively (Table 4.3). Particularly root length was reduced
by approximately 80% in both culture systems. Due to stronger inhibition of shoot
growth as compared to root growth, the root/shoot biomass ratio increased under
low RZT in comparison with plants gown under ambient temperature. Probably
due to a generally better nutrient availability in hydroponics, the root/shoot ratio
was generally lower than in soil culture.
The strong inhibition of shoot growth has been related to hormonal imbalances,
such as reduced biosynthesis and root to shoot translocation of cytokinins (Atkin
et al., 1973), as a typical response also to N and P deficiency. Accordingly,
Marschner and Engels (1991) reported reduced root to shoot translocation of N
and particularly of P in maize plants exposed to low RZT. However, cytokinin
biosynthesis may be also impaired by a general reduction of metabolic activity in
the root meristems at lower soil temperatures (Covey-Crump et al., 2002). At the
same time, root to shoot translocation of the cytokinin antagonist abscisic acid
(ABA), counteracting the biosynthesis of cytokinins and vice versa, increased
with inhibitory effects on shoot growth (Atkin et al.1973). Increased ABA
translocation to the shoots is also a common response to water limitation and
may therefore also be associated with a reduced hydraulic conductivity of the
roots and limited water supply to the shoots frequently reported in response to
low RZT (Huang et al., 1991).
All priming treatments, but particularly micronutrient seed priming alleviated the
detrimental effects of low RZT on plant growth. Biomass production of primed
seedlings reached 50 % of the control plants grown at ambient temperature in

Chapter(4((((((((((((((((((((((((((((((((((((((((((((((((((((((((((((((((((((((((((((((((((((((((((((((((((((((((((((((((((((((((((((((((((((((((((((Maize(at(Low(RZT(
( 91(
the hydroponic culture system and 75-85 % in soil culture (Table 4.3). Particularly
in the soil culture system, root length development was much stronger stimulated
by micronutrient seed priming (120-150 %, Table 4.4) than root biomass
production (20-40%, Table 3). This indicates the formation of longer and finer
roots in response to the priming treatments (Table 4.4), a feature promoting
nutrient acquisition under low RZT (Hundt et al., 2007) but also after recovery
from stress conditions. Figure 2 shows that this was particularly attributed to
stimulation of lateral root formation.
Inhibition of plant growth due to low RZT, associated with reduced uptake and
root to shoot translocation of micronutrients has been previously reported by
Engels and Marschner (1996a) for Zn, Mn, Cu and Fe. At the same time also
assimilate translocation to the roots was inhibited (Engels and Marschner,
1996b). This may indicate a limitation of photosynthesis, which strongly depends
on the supply with micronutrients, such as Fe, Mn, Zn. and Cu (Marschner, 1995)
with growth inhibition as a final consequence. Accordingly, Strauss et al. (2007)
reported a functional limitation of the photosystem II (PSII) in plants exposed to
low RZT, while Nagasuga et al. (2011) stated that reduced growth and biomass
production of rice plants, grown under low RZT was mainly attributed to the
reduction of shoot growth and leaf area.
Almost all types of plant stresses, including suboptimal temperatures, involve
oxidative stress as one of the rapid responses (Blokhina et al., 2003). This is
associated with increased production of active oxygen species (AOS), such as
superoxide, hydrogen peroxide and the hydroxyl radical, involved in membrane
damage by lipid peroxidation, protein degradation, enzyme inactivation and
disruption of DNA strands (Allen 1995). Micronutrients, such as Zn and Mn are
important co-factors of various enzymes involved in the detoxification of AOS,
such as superoxide dismutases (SODs, Cakmak and Marschner, 1988 a, b,
1992; Cakmak, 2000). Deficiency of Zn, Mn and Cu alters the activities of SOD
forms and these changes depend on the type and the duration of the deficiency
stress (Yu and Rengel, 1999). Moreover, Mn is a cofactor of various enzymes
involved in the biosynthesis of phenolic compounds with anti-oxidative properties

Chapter(4((((((((((((((((((((((((((((((((((((((((((((((((((((((((((((((((((((((((((((((((((((((((((((((((((((((((((((((((((((((((((((((((((((((((((((Maize(at(Low(RZT(
( 92(
(Thompson and Huber, 2007). Zinc has functions in direct membrane
stabilization, in biosynthesis of auxins (Salami and Kenefick, 1970) and
gibberellins (Sekimoto et al., 1997) involved in plant growth regulation and in
protein synthesis in general. Therefore, limited supply of these micronutrients
under conditions of low RZT (Engels, 1994) may increase oxidative damage of
root cells and induce disturbances in plant growth, which may be alleviated by
supplementation via micronutrient seed priming.
Both in hydroponics and in soil culture, micronutrient shoot concentrations were
not substantially elevated by the priming treatments (Table 4.5). However
significant increases were observed for the contents of the respective nutrients
(Table 4.6). This increase was still detectable even in field grown plants at 5
weeks after sowing (Table 4.7). Obviously, the surplus in micronutrient supply in
the priming treatments was immediately transformed into plant growth. For Mn
and particularly for Zn, the amount of micronutrients supplemented by seed
priming (Table 4.2) was sufficiently high to explain the increase in micronutrient
contents in the plants at 3-5 weeks after sowing (Tables 4.6 and 4.7). However,
the amount of Fe taken up by the seeds (3 µg seed-1, Table 2) was much lower
than the observed increases in Fe shoot contents (Table 4) of up to 500 µg
seedling-1 in field grown plants (Table 4.7). This finding demonstrates that
improvements of the micronutrient status by seed priming are not only a direct
consequence of nutrient supplementation but most probably also of improved
nutrient acquisition from the soil via stimulation of root growth (Table 4.4). This is
further underlined by the observation that iron seed priming simultaneously
increased the Zn and Mn status of the plants and vice versa (Table 6) and that
also P content, as an important sparingly soluble macronutrient, was increased
by the priming treatments.
Micronutrient seed priming treatments were efficient enough to produce a 10-15
% increase in final grain yield in two independent field experiments conducted in
2010 and 2011 (Table 4.8). The extraordinary persistence of the priming effects
may be attributed particularly to the enormous stimulation of root growth
observed in the model experiments (Tables 4.3 and 4.4). This may act as an

Chapter(4((((((((((((((((((((((((((((((((((((((((((((((((((((((((((((((((((((((((((((((((((((((((((((((((((((((((((((((((((((((((((((((((((((((((((((Maize(at(Low(RZT(
( 93(
advantage not only during the stress treatments but may further promote plant
growth during and even after the recovery phase. Moreover, several other
studies have demonstrated long-lasting positive effect of micronutrient seed
priming also on the alleviation of other stress factors. Similarly, Harris et al.,
(2007) have shown beneficial effects of Zn seed priming on maize by 27%
increase in grain yield and almost 3 mg kg-1 increase in seed Zn concentration on
Zn deficient soils in Pakistan.
4.6. Conclusion
Many previous investigations have demonstrated declining nutrient supply to
sensitive plant species exposed to low RZT during germination and early growth.
However, this is the first report showing that micronutrient (Fe, Zn, Mn) supply
acts as a specific growth limiting factor under these conditions, which can be
supplemented by micronutrient seed priming with long-lasting effects even until
final harvest. The physiological background of these effects as well as their
expression under different environmental conditions (e.g. soil type, soil-water and
redox status, fertilisation management, variation in priming treatments and
genotypes etc.) need to be further elucidated. Nevertheless, the present study
demonstrates that micronutrient seed priming may offer a realistic perspective to
mitigate problems of plant performance under low RZT. The seed priming
technique is simple, inexpensive and can be conducted directly prior to sowing
e.g. by small-scale farmers but allows also combinations with industrial seed
dressing techniques due to persistence of the effects after drying-back the
primed seeds.

Chapter(5((((((((((((((((((((((((((((((((((((((((((((((((((((((((((((((((((((((((((((((((((((((((((((((((((((((((((((Soybean(Nutrient(Seed(Priming(
( 94(
Chapter 5
Accumulation and distribution of Zn and Mn in soybean seeds after
nutrient seed priming and its contribution to plant growth under Zn
and Mn deficient conditions (Journal of Plant Nutrition (submitted))
Authors
Imran Muhammad, Römheld Volker, Neumann Günter
Institute of Crop Science, Nutritional Crop Physiology (340h),
Universität Hohenheim, 70593 Stuttgart, Germany
Corresponding author: Imran Muhammad
Institute of Crop Science, Nutritional Crop Physiology (340h)
Universität Hohenheim, 70599, Stuttgart, Germany
Phone: 0049 152 13172241 Fax: 0049 711 459 23295
E-mail: [email protected]
Contribution in the manuscript:
Planning of experiments, Preparation of experimental materials, Nutrient
analysis, Root length measurements, Conducting field trials, Statistical
analysis, Manuscript writing.

Chapter(5((((((((((((((((((((((((((((((((((((((((((((((((((((((((((((((((((((((((((((((((((((((((((((((((((((((((((((Soybean(Nutrient(Seed(Priming(
( 95(
5.1. Abstract
Nutrient seed priming is a strategy to increase the seed reserves of mineral
nutrients as primary source for mineral nutrition during seedling
development and early growth. The present study investigates effects of
zinc (Zn) and manganese (Mn) seed priming on growth and nutritional
status of soybean under conditions of Zn and Mn limitation. Soybean
seeds were primed with distilled water and nutrient solutions containing Zn
(10mM), Mn (5mM) and Zn+Mn (10mM + 5mM) for 12 hours between
layers of moist filter paper to minimize imbibition damage. Nutrient seed
priming increased the natural seed reserves for Zn by, approximately 6 fold
and by 5 fold for Mn. However, 40-60 % of the primed nutrients were
adsorbed to the seed coat.
Soybean plants were grown for 5 weeks in nutrient solution under Zn and
Mn deficiency and on a calcareous Loess subsoil low in plant available Zn
and Mn. Zinc and Zn+Mn priming improved plant growth and increased
shoot biomass production both, in solution culture and in the soil
experiment. Zinc seed priming was able to maintain plant growth for five
weeks in the same way as Zn supply via the nutrient solution, while the
amount of primed Mn was not sufficient to overcome Mn deficiency
symptoms. Antagonistic effects were observed after combined priming with
Zn and Mn.
It is concluded that particularly nutrient seed priming with Zn offers
perspectives to improve seed quality of soybean for early seedling
development under limited nutrient supply or availability and needs further
investigation on performance under various stress conditions.
Keywords: Soybean, nutrient seed priming, Zn and Mn localization,
nutrient uptake

Chapter(5((((((((((((((((((((((((((((((((((((((((((((((((((((((((((((((((((((((((((((((((((((((((((((((((((((((((((((Soybean(Nutrient(Seed(Priming(
( 96(
5.2. Introduction
Seed priming is a pre-sowing seed treatment in which seeds are soaked
in water and dried back to storage moisture contents for further use. In
the recent past, the ‘on-farm seed priming’ method according to Harris et
al., (1997) has become very popular in developing countries. For ‘on-
farm seed priming’, seeds are soaked in water durng a certain time
period and only shortly air-dried (not to initial moisture contents) before
sowing. This can speed up germination, improve the tolerance to various
stress conditions and can increase crop yield (Harris et al., 2000).
‘Nutrient seed priming’ is a technique in which seeds are soaked in
nutrient solutions instead of water. This increases seed nutrient contents
along with the priming effects to improve seed quality for better
germination and seedling growth. Maize seed priming with Zn stimulated
plant growth and increased final grain yield and seed Zn contents on
soils with limited Zn supply (Harris et al., 2007) and under stress of low
root-zone temperature (Imran et al., 2013). In barley, nutrient priming of
seeds with P and Zn stimulated germination, seedling growth and plant
uptake of these elements (Ajouri et al., 2004). In contrast, Zn and B
seed priming had shown no effects on germination rate and grain yield
of chickpea, lentil and cowpea grown in Zn and B deficient soils in Nepal
(Johnson et al., 2005).
Under stress conditions during the sensitive stages of germination and
early seedling development, micronutrients are of outstanding
importance. Zinc and manganese are co-factors of various enzymes
(Superoxide dismutase, SOD) involved in the detoxification of reactive
oxygen species such as O2- (superoxide radical) and H2O2 (hydrogen
peroxide) (Cakmak, 2000). Moreover, Mn is a cofactor of many
enzymes involved in the biosynthesis of phenolic compounds with anti-
oxidative and antibiotic properties (Thompson and Huber, 2007). Zinc
has functions in direct membrane stabilization, in biosynthesis of auxins
(Salami and Kenefick, 1970) and gibberellins (Sekimoto et al., 1997)

Chapter(5((((((((((((((((((((((((((((((((((((((((((((((((((((((((((((((((((((((((((((((((((((((((((((((((((((((((((((Soybean(Nutrient(Seed(Priming(
( 97(
involved in the plant growth regulation and in protein synthesis in
general.
In the recent past, soybean (Glycine max L.) cropping has been largely
extended and its cultivation is rapidly increasing even in countries with
temperate climates, with a shorter growth period for crop development
(from germination to maturity) (Dashti et el., 1998). Under these
conditions, speeding up germination, and early seedling development is
vital for on-time crop maturity and optimal grain yields. Moreover,
soybean cultivation in temperate climates implies additional abiotic and
biotic stress factors, such as sub-optimal air and soil temperature
leading to reduced water uptake, decreased nutrient availability and
short photoperiods as well as disease problems (Ogle et al., 1979;
James, 1998; Osborne, 2006). Based on the promising results of Zn
and Mn seed priming with respect to improvement of plant growth and
stress tolerance in various crop species (Ajouri et al., 2004; Imran et al.,
2012b), the present study investigates perspectives for seed priming
with Zn and Mn on growth of soybean under conditions of limited
nutrient availability, with the major aims:
(i) To develop an optimised priming protocol for soybean seeds.
(ii) To estimate the time frame for a direct nutritional impact of
micronutrient seed priming during plant development.
(iii) Testing the effects of micronutrient seed priming on early
growth of soybean on a soil with limited availability of Zn and
Mn.
5.2. Materials and methods
5.2.1. Seed priming methods
Soybean (Glycine max L. cv. ‘Conquista’) seeds were obtained from
Adriana Farms (Brazil). Seeds were primed using the ‘on-farm seed
priming method’ according to Harris et al., (1997), in which seeds were

Chapter(5((((((((((((((((((((((((((((((((((((((((((((((((((((((((((((((((((((((((((((((((((((((((((((((((((((((((((((Soybean(Nutrient(Seed(Priming(
( 98(
surface dried for 2 hours after the priming treatment to avoid soil
clogging on seed surface during sowing. Optimisation tests for priming
treatments were performed with each 100 seeds in 4 replicates using
distilled water for 6, 12 and 24 hours with three different methods:
(i) Priming according to water uptake capacity of the seeds (absorption
capacity - (AC) - method)
Seeds were soaked in a pre-determined quantity of distilled water
(equal to the water absorption capacity of the seeds) for each time
treatment in glass beakers. To facilitate uniform absorption, seeds were
evenly immersed in distilled water by placing them in a single layer in
the beakers. Beakers were covered with parafilm to avoid evaporation
and incubated at room temperature (22 °C) in the dark.
(ii) Submerged priming (SP-Method)
In this method, seeds were soaked into 100 ml of distilled water in glass
beakers. All seeds were submerged during the priming period and were
incubated at room temperature in the dark in beakers closed with
Parafilm..
(iii) Priming between filter paper layers (FP-Method)
Seeds were incubated in petri dishes between 3 layers of filter paper
(MN 710, Macchery & Nagel, Düren, Germany) with three layers below
and two layers on top of the seeds. A calculated amount of priming
solution equal to the absorbing capacity of the seeds and filter papers
was applied. Petri dishes were covered with lids to minimize
evaporation losses, and incubated at room temperature in the dark.
After the priming treatments, the seeds were rinsed with distilled water
to remove any adhering compounds and nutrients from the seed
surface. Rinsed seeds were air dried at room temperature for 2 hours
before further use .

Chapter(5((((((((((((((((((((((((((((((((((((((((((((((((((((((((((((((((((((((((((((((((((((((((((((((((((((((((((((Soybean(Nutrient(Seed(Priming(
( 99(
5.2.2 Germination test
Primed soybean seeds were germinated in olls prepared of 4 layers of
germination paper (MN 710, Macchery & Nagel, Düren. Germany)
moistened with 60 mL of distilled water in 4 replicates (10 seeds per
replicate) for 7 days. Seed germination was evaluated according to
ISTA (International Seed Testing Association) rules, 1983.
Additionally, early seedling development up to 7 days was also
monitored in peat culture substrate (TKS®1 Instant Plus, Floragard,
Germany). For this purpose, four water primed seeds were sown in 0.5
liter pots filled with moistened peat culture substrate . Development of
normal seedlings was assessed according to ISTA rules.
5.2.3 Nutrient seed priming
To determine the suitable concentrations of Zn and Mn for optimal
effects of nutrient seed priming, soybean seeds were primed with 5, 10
and 15 mM ZnSO4 and MnSO4 solutions. Zinc and Mn concentrations
for nutrient priming were selected based on the development of
abnormal seedlings in the germination test described in section 2.2..
Seeds were primed according to the FP-Method described in section
2.1.
5.2.4 Plant culture
Experiments were conducted in a climate chamber under controlled
environmental conditions. Average air temperature was adjusted to 25+
3 °C with a light period of 15 h, a light intensity of 200 mol m-2 s-1, and a
relative air humidity of 60-80%.
Nutrient solution experiment
For hydroponic culture, primed seeds were germinated in rolls with 4
layers of moist germination paper (MN 710, Macchery & Nagel, Düren.
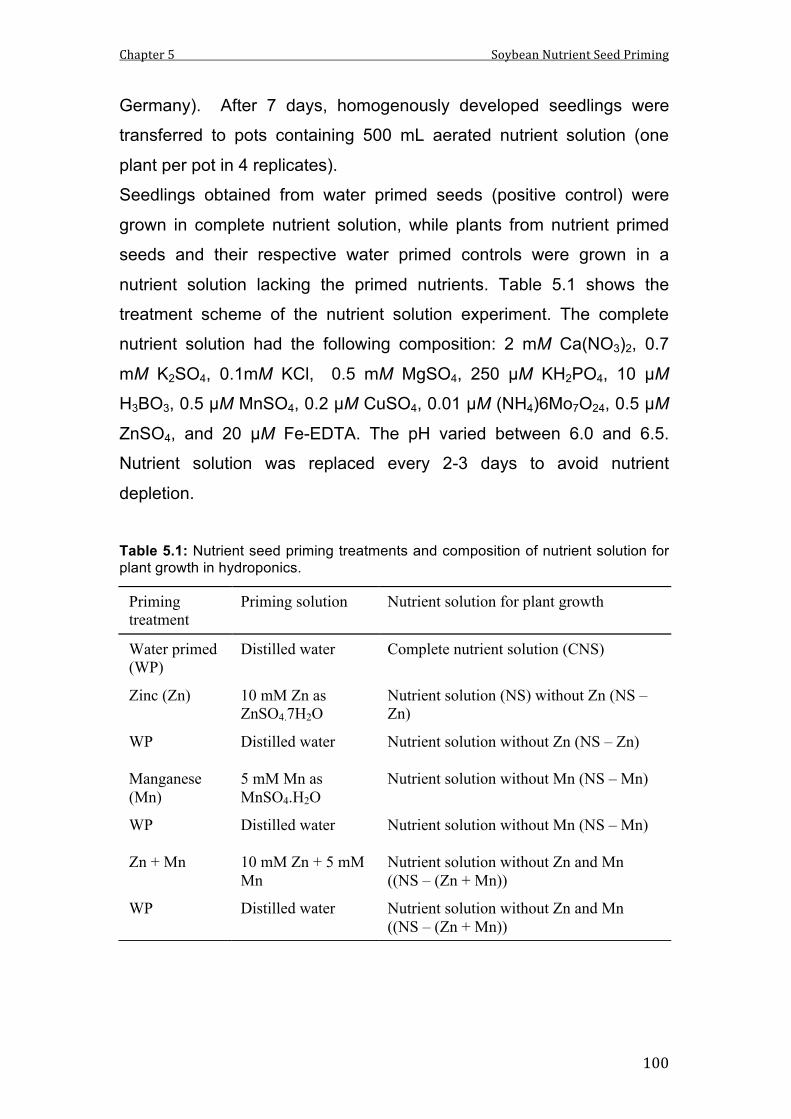
Chapter(5((((((((((((((((((((((((((((((((((((((((((((((((((((((((((((((((((((((((((((((((((((((((((((((((((((((((((((Soybean(Nutrient(Seed(Priming(
( 100(
Germany). After 7 days, homogenously developed seedlings were
transferred to pots containing 500 mL aerated nutrient solution (one
plant per pot in 4 replicates).
Seedlings obtained from water primed seeds (positive control) were
grown in complete nutrient solution, while plants from nutrient primed
seeds and their respective water primed controls were grown in a
nutrient solution lacking the primed nutrients. Table 5.1 shows the
treatment scheme of the nutrient solution experiment. The complete
nutrient solution had the following composition: 2 mM Ca(NO3)2, 0.7
mM K2SO4, 0.1mM KCl, 0.5 mM MgSO4, 250 µM KH2PO4, 10 µM
H3BO3, 0.5 µM MnSO4, 0.2 µM CuSO4, 0.01 µM (NH4)6Mo7O24, 0.5 µM
ZnSO4, and 20 µM Fe-EDTA. The pH varied between 6.0 and 6.5.
Nutrient solution was replaced every 2-3 days to avoid nutrient
depletion.
Table 5.1: Nutrient seed priming treatments and composition of nutrient solution for plant growth in hydroponics.
Priming treatment
Priming solution Nutrient solution for plant growth
Water primed (WP)
Distilled water Complete nutrient solution (CNS)
Zinc (Zn) 10 mM Zn as ZnSO4.7H2O
Nutrient solution (NS) without Zn (NS – Zn)
WP Distilled water Nutrient solution without Zn (NS – Zn)
Manganese (Mn)
5 mM Mn as MnSO4.H2O
Nutrient solution without Mn (NS – Mn)
WP Distilled water Nutrient solution without Mn (NS – Mn)
Zn + Mn 10 mM Zn + 5 mM Mn
Nutrient solution without Zn and Mn ((NS – (Zn + Mn))
WP Distilled water Nutrient solution without Zn and Mn ((NS – (Zn + Mn))

Chapter(5((((((((((((((((((((((((((((((((((((((((((((((((((((((((((((((((((((((((((((((((((((((((((((((((((((((((((((Soybean(Nutrient(Seed(Priming(
( 101(
Soil culture experiment
For the soil culture experiment, four primed soybean seeds (FP-
Method) were sown directly at a depth of 2 cm into pots filled with 500
grams of a calcareous Loess subsoil (pH (CaCl2) 7.6; Corg.<0.3%;
CaCO3 23.3%; water- extractable Ca2+ and Mg2+ (mg kg−1 soil) 59.9 and
11.3, respectively). Calcium chloride–diethylenetriamine penta-acetic
acid (CAT)-extractable micronutrient concentrations (mg kg−1 soil):
Mn=7.4, Fe=369, Zn=0.8, B=0.9, Cu= 0.5 and P (CAL) was 5 mg kg-1
(VDLUFA, 2004).
Before sowing, the soil was fertilized once with N, P, K and Mg.
Nitrogen was supplied as NH4NO3 at 100 mg kg-1 soil, P as Ca(H2PO4)2
at 100 mg kg-1 soil, K as K2SO4 at 150 mg kg-1 soil and Mg as MgSO4
at 100 mg kg-1 soil. The different fertilizers mixed homogenously with
the soil substrate. Before filling the pots, the soil was sieved through 2
mm mesh size. Soil moisture was daily adjusted to 18% (w/w). After 7
days, in each pot two seedlings were removed and two well-developed
seedlings were further cultivated for 4 weeks. Control treatments
comprised unprimed and water-primed seeds.
5.2.5 Plant analysis
Soybean plants grown in hydroponics and in soil culture were
harvested after a growth period of five weeks. Fresh biomass of plant
shoots and roots was determined and the plant material was
subsequently oven-dried at 65°C for determination of dry matter and
mineral nutrient analysis.
For seed mineral nutrient analysis after the priming treatments, seeds
were rinsed with de-ionized water for 1 min to remove any compounds
and nutrients adhering to the seed coat. Afterwards the seed coat was
removed carefully to avoid damage of the remaining parts.
Mineral nutrients were determined in complete unwashed and washed
seeds, in the seed coat, the remaining seed and in shoots of soybean

Chapter(5((((((((((((((((((((((((((((((((((((((((((((((((((((((((((((((((((((((((((((((((((((((((((((((((((((((((((((Soybean(Nutrient(Seed(Priming(
( 102(
plants. After drying at 65 °C, ground samples were ashed in a muffle
furnace at 500°C for 5h. After cooling, the samples were extracted
twice with 2 ml of 3.4 M HNO3 (v/v) and subsequently evaporated to
dryness. The ash was dissolved in 2 mL of 4 M HCl, subsequently
diluted 10-fold with hot de-ionized water, and boiled for 2 min. After
adding 0.1 ml of Cs/La buffer to 4.9 ml ash solution, Zn and Mn
concentrations were measured by atomic absorption spectrometry
(UNICAM 939, Offenbach/Main, Germany).
2.6 Statistics
A completely randomised block design with 4 replicates was used in all
experiments. The data were analysed by analysis of variance with a
significance level of p < 0.05. For mean separation Duncan’s Multiple
Range Test (DMRT) were used. The software packages Microsoft
Excel Professional 2007 and interaction between priming methods and
durations was analysed using the ‘Sigma Stat version 3.1’ software
(SYSTAT Software Inc., Erkrath, Germany).
5.3 Results
5.3.1 Optimisation of seed priming treatments
Soybean seeds, water primed for 6, 12 and 24 hours with three
different methods (submergence in water = SP-Method, submergence
in water according to the water uptake capacity of the seeds = AC-
Method) incubation between layers of moist filter papers = FP-Method))
were tested for germination and seedling development.
Figure 5.1 shows the time course of radicle emergence (germination
speed) after seed water priming between filter paper layers. Water
priming significantly increased germination speed in 12 h priming
treatment but not after priming durations of 6 h and 24 h. (Fig 5.1a).
Within all tested priming treatments, seed priming for 12 h between
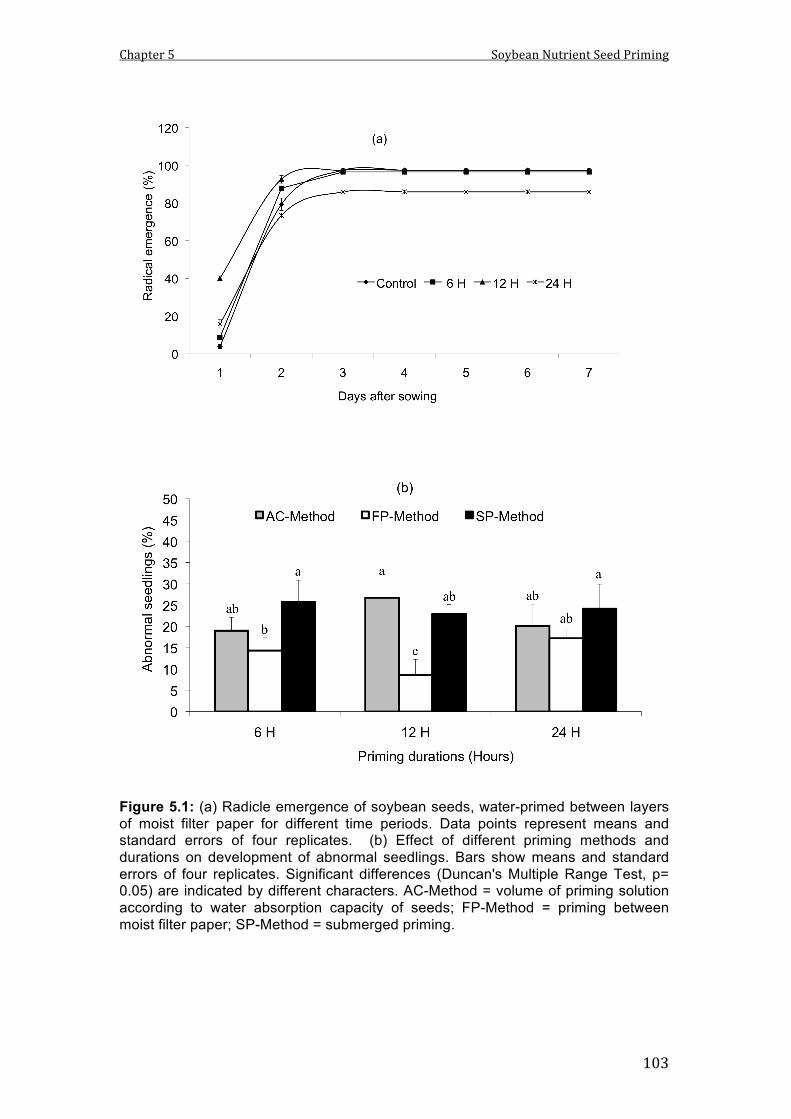
Chapter(5((((((((((((((((((((((((((((((((((((((((((((((((((((((((((((((((((((((((((((((((((((((((((((((((((((((((((((Soybean(Nutrient(Seed(Priming(
( 103(
(
(Figure 5.1: (a) Radicle emergence of soybean seeds, water-primed between layers of moist filter paper for different time periods. Data points represent means and standard errors of four replicates. (b) Effect of different priming methods and durations on development of abnormal seedlings. Bars show means and standard errors of four replicates. Significant differences (Duncan's Multiple Range Test, p= 0.05) are indicated by different characters. AC-Method = volume of priming solution according to water absorption capacity of seeds; FP-Method = priming between moist filter paper; SP-Method = submerged priming.

Chapter(5((((((((((((((((((((((((((((((((((((((((((((((((((((((((((((((((((((((((((((((((((((((((((((((((((((((((((((Soybean(Nutrient(Seed(Priming(
( 104(
layers of moist filter paper resulted in the lowest proportion of abnormal
seedlings (Fig. 5.1b) and the most homogenous seedling emergence
on peat culture substrate (Fig. 5.2) All three methods and priming
durations were significantly different from each other with p values of
0.001 and 0.008, respectively, but the interaction between methods and
priming durations was found to be non-significant with a p value of
0.066.
Figure 5.2: Soybean seedlings grown for 7 days on peat culture substrate after water priming with different methods and priming durations. AC-Method = volume of priming solution according to water absorption capacity of seeds; FP-Method = priming between moist filter paper; SP-Method = submerged priming. Based on these findings, priming treatments for all other experiments
were routinely performed for 12 h between moist layers of filter paper.
In germination tests, the development of abnormal seedlings was
significantly reduced by Zn and Mn seed priming in all treatments
compared to the unprimed control, but no significant differences was
found between different concentration levels for both Zn and Mn (Table
5.2). Therefore, based on the lowest means determined for abnormal
seedling development (Table. 5.2), 10 mM Zn and 5 mM Mn
concentrations and their combination (10 mM Zn + 5mM Mn) were
selected for soybean seed nutrient priming.
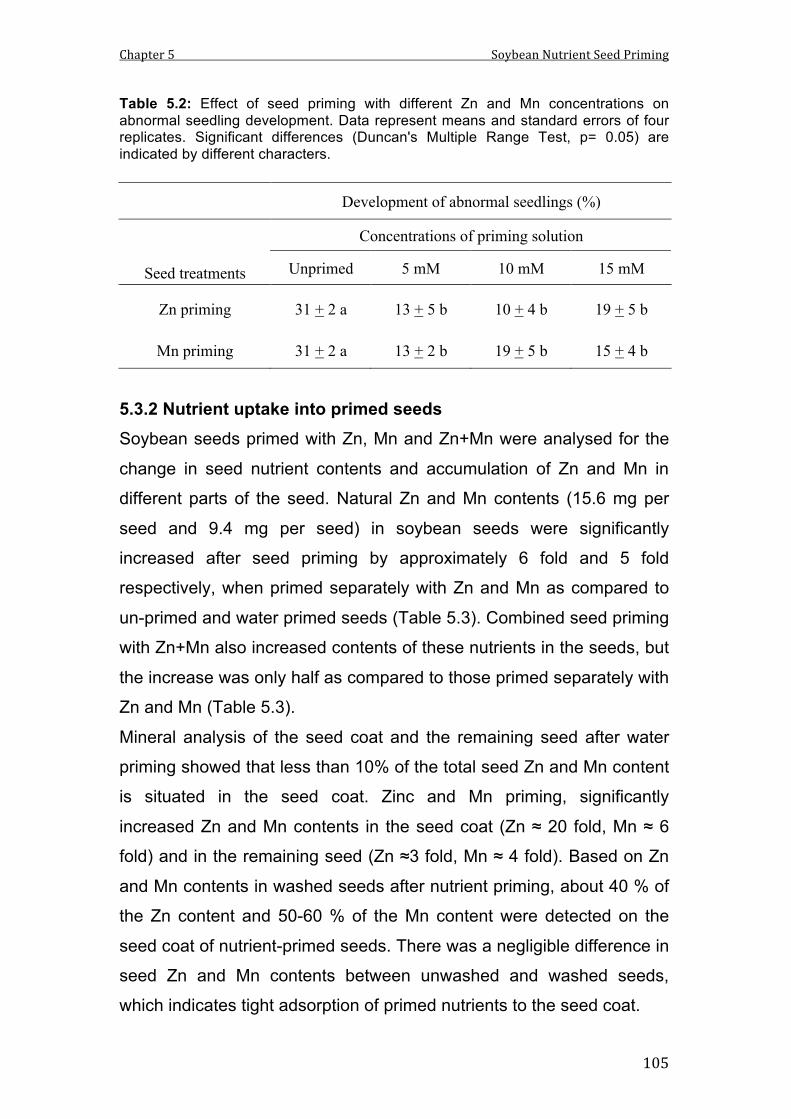
Chapter(5((((((((((((((((((((((((((((((((((((((((((((((((((((((((((((((((((((((((((((((((((((((((((((((((((((((((((((Soybean(Nutrient(Seed(Priming(
( 105(
Table 5.2: Effect of seed priming with different Zn and Mn concentrations on abnormal seedling development. Data represent means and standard errors of four replicates. Significant differences (Duncan's Multiple Range Test, p= 0.05) are indicated by different characters.
Development of abnormal seedlings (%)
Concentrations of priming solution
Seed treatments Unprimed 5 mM 10 mM 15 mM
Zn priming 31 + 2 a 13 + 5 b 10 + 4 b 19 + 5 b
Mn priming 31 + 2 a 13 + 2 b 19 + 5 b 15 + 4 b
5.3.2 Nutrient uptake into primed seeds
Soybean seeds primed with Zn, Mn and Zn+Mn were analysed for the
change in seed nutrient contents and accumulation of Zn and Mn in
different parts of the seed. Natural Zn and Mn contents (15.6 mg per
seed and 9.4 mg per seed) in soybean seeds were significantly
increased after seed priming by approximately 6 fold and 5 fold
respectively, when primed separately with Zn and Mn as compared to
un-primed and water primed seeds (Table 5.3). Combined seed priming
with Zn+Mn also increased contents of these nutrients in the seeds, but
the increase was only half as compared to those primed separately with
Zn and Mn (Table 5.3).
Mineral analysis of the seed coat and the remaining seed after water
priming showed that less than 10% of the total seed Zn and Mn content
is situated in the seed coat. Zinc and Mn priming, significantly
increased Zn and Mn contents in the seed coat (Zn ≈ 20 fold, Mn ≈ 6
fold) and in the remaining seed (Zn ≈3 fold, Mn ≈ 4 fold). Based on Zn
and Mn contents in washed seeds after nutrient priming, about 40 % of
the Zn content and 50-60 % of the Mn content were detected on the
seed coat of nutrient-primed seeds. There was a negligible difference in
seed Zn and Mn contents between unwashed and washed seeds,
which indicates tight adsorption of primed nutrients to the seed coat.
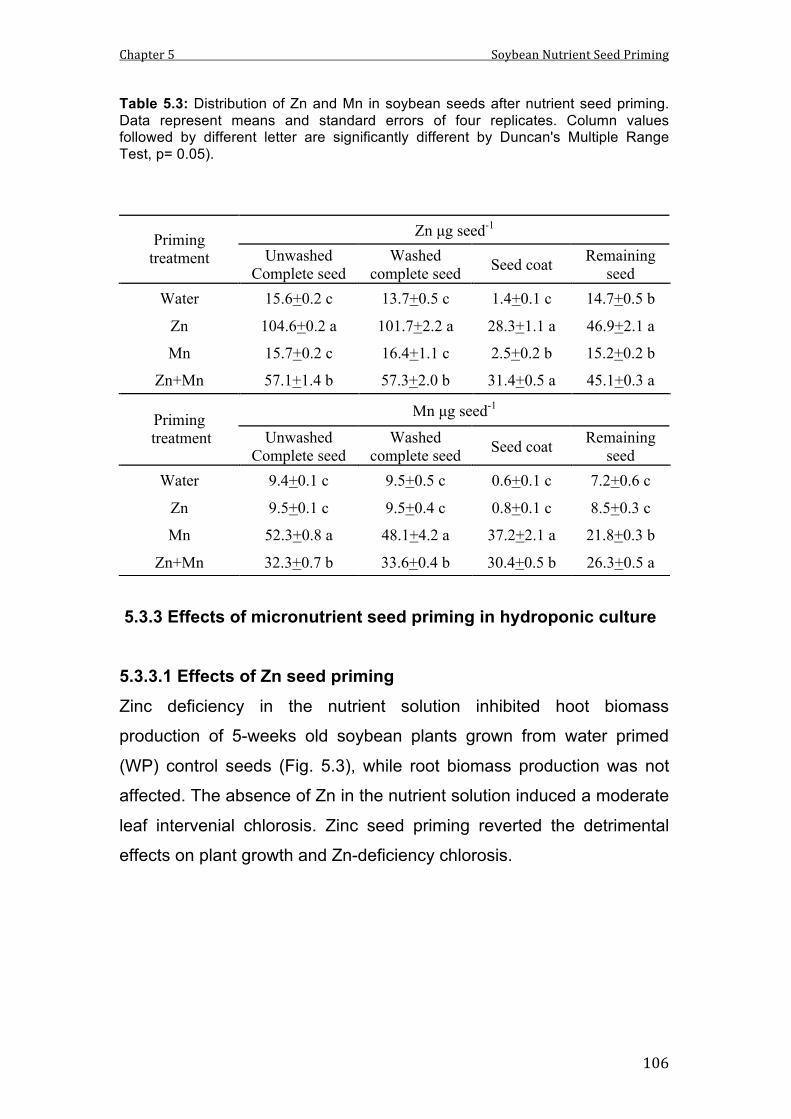
Chapter(5((((((((((((((((((((((((((((((((((((((((((((((((((((((((((((((((((((((((((((((((((((((((((((((((((((((((((((Soybean(Nutrient(Seed(Priming(
( 106(
Table 5.3: Distribution of Zn and Mn in soybean seeds after nutrient seed priming. Data represent means and standard errors of four replicates. Column values followed by different letter are significantly different by Duncan's Multiple Range Test, p= 0.05).
5.3.3 Effects of micronutrient seed priming in hydroponic culture
5.3.3.1 Effects of Zn seed priming
Zinc deficiency in the nutrient solution inhibited hoot biomass
production of 5-weeks old soybean plants grown from water primed
(WP) control seeds (Fig. 5.3), while root biomass production was not
affected. The absence of Zn in the nutrient solution induced a moderate
leaf intervenial chlorosis. Zinc seed priming reverted the detrimental
effects on plant growth and Zn-deficiency chlorosis.
Zn µg seed-1 Priming treatment Unwashed
Complete seed Washed
complete seed Seed coat Remaining seed
Water 15.6+0.2 c 13.7+0.5 c 1.4+0.1 c 14.7+0.5 b
Zn 104.6+0.2 a 101.7+2.2 a 28.3+1.1 a 46.9+2.1 a
Mn 15.7+0.2 c 16.4+1.1 c 2.5+0.2 b 15.2+0.2 b
Zn+Mn 57.1+1.4 b 57.3+2.0 b 31.4+0.5 a 45.1+0.3 a
Mn µg seed-1 Priming treatment Unwashed
Complete seed Washed
complete seed Seed coat Remaining seed
Water 9.4+0.1 c 9.5+0.5 c 0.6+0.1 c 7.2+0.6 c
Zn 9.5+0.1 c 9.5+0.4 c 0.8+0.1 c 8.5+0.3 c
Mn 52.3+0.8 a 48.1+4.2 a 37.2+2.1 a 21.8+0.3 b
Zn+Mn 32.3+0.7 b 33.6+0.4 b 30.4+0.5 b 26.3+0.5 a
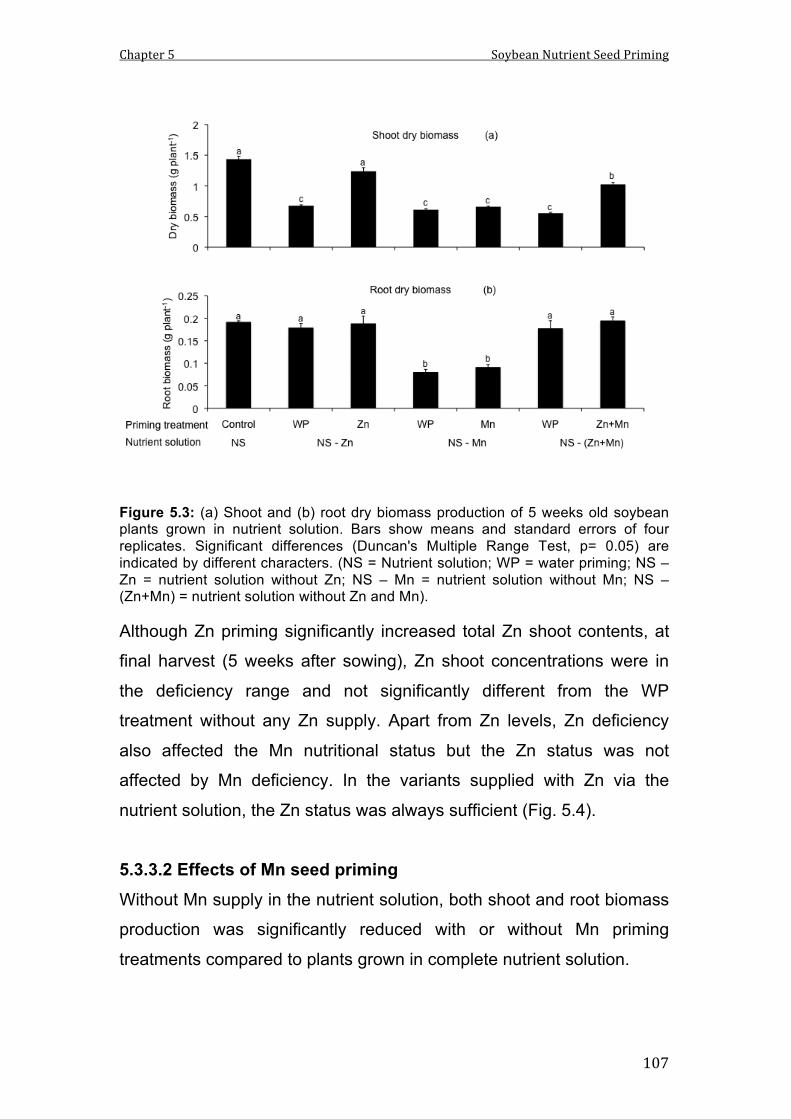
Chapter(5((((((((((((((((((((((((((((((((((((((((((((((((((((((((((((((((((((((((((((((((((((((((((((((((((((((((((((Soybean(Nutrient(Seed(Priming(
( 107(
(Figure 5.3: (a) Shoot and (b) root dry biomass production of 5 weeks old soybean plants grown in nutrient solution. Bars show means and standard errors of four replicates. Significant differences (Duncan's Multiple Range Test, p= 0.05) are indicated by different characters. (NS = Nutrient solution; WP = water priming; NS – Zn = nutrient solution without Zn; NS – Mn = nutrient solution without Mn; NS – (Zn+Mn) = nutrient solution without Zn and Mn).
Although Zn priming significantly increased total Zn shoot contents, at
final harvest (5 weeks after sowing), Zn shoot concentrations were in
the deficiency range and not significantly different from the WP
treatment without any Zn supply. Apart from Zn levels, Zn deficiency
also affected the Mn nutritional status but the Zn status was not
affected by Mn deficiency. In the variants supplied with Zn via the
nutrient solution, the Zn status was always sufficient (Fig. 5.4).
5.3.3.2 Effects of Mn seed priming
Without Mn supply in the nutrient solution, both shoot and root biomass
production was significantly reduced with or without Mn priming
treatments compared to plants grown in complete nutrient solution.
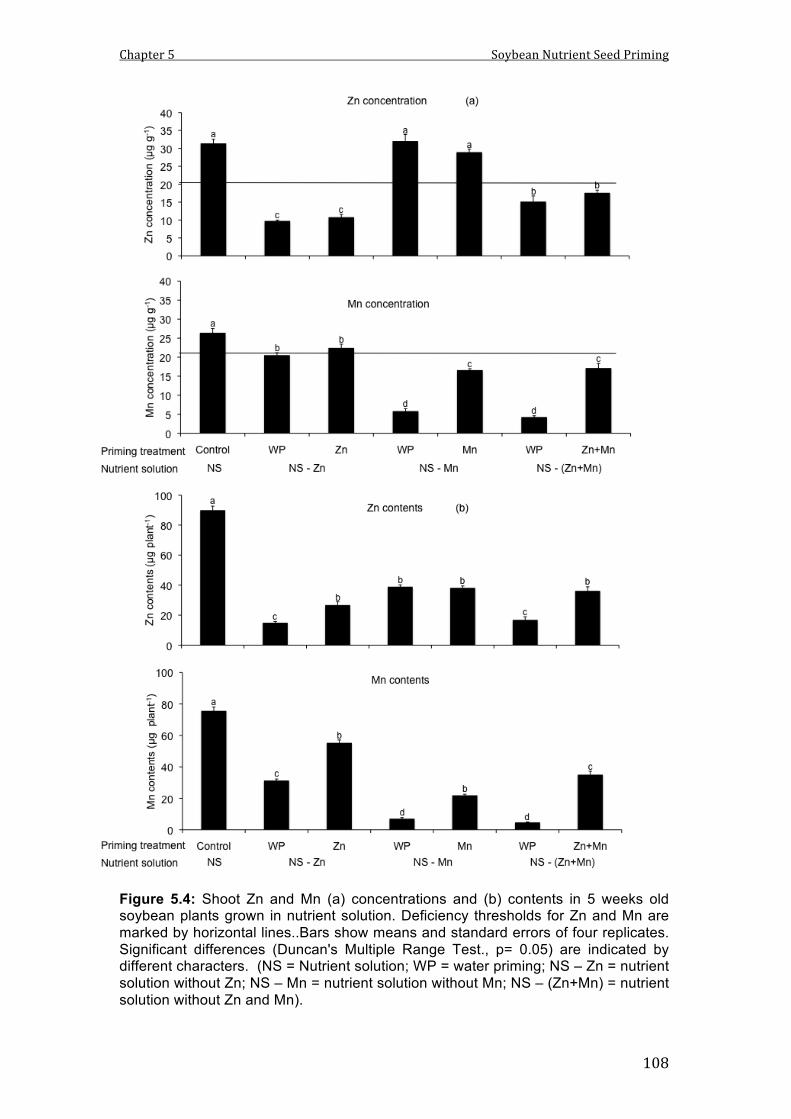
Chapter(5((((((((((((((((((((((((((((((((((((((((((((((((((((((((((((((((((((((((((((((((((((((((((((((((((((((((((((Soybean(Nutrient(Seed(Priming(
( 108(
Figure 5.4: Shoot Zn and Mn (a) concentrations and (b) contents in 5 weeks old soybean plants grown in nutrient solution. Deficiency thresholds for Zn and Mn are marked by horizontal lines..Bars show means and standard errors of four replicates. Significant differences (Duncan's Multiple Range Test., p= 0.05) are indicated by different characters. (NS = Nutrient solution; WP = water priming; NS – Zn = nutrient solution without Zn; NS – Mn = nutrient solution without Mn; NS – (Zn+Mn) = nutrient solution without Zn and Mn).

Chapter(5((((((((((((((((((((((((((((((((((((((((((((((((((((((((((((((((((((((((((((((((((((((((((((((((((((((((((((Soybean(Nutrient(Seed(Priming(
( 109(
(Both, Mn contents and Mn concentrations in the shoot tissue were
significantly increased after Mn seed priming. However, at final harvest
(5 weeks after sowing), Mn concentrations were still below the Mn
deficiency threshold of 20 µg g-1 dry matter, while the Mn status was
sufficient when Mn was supplied via the nutrient solution (Fig. 5.4).
3.3.3 Effects of combined seed priming with Zn and Mn
Surprisingly, in contrast to plants grown under sole Mn deficiency, root
biomass production of plants grown under combined deficiency of Zn
and Mn in the nutrient solution was almost similar to the plants grown in
complete nutrient solution. Shoot biomass was reduced but shoot
elongation was not affected and the leaves showed typical Mn
deficiency chlorosis. Shoot Zn and Mn status was in the deficiency
range but higher shoot concentrations of Zn were detected under
combined deficiency of Zn and Mn as compared to Zn deficiency alone.
The beneficial effect of Zn supply on shoot growth was less expressed
in the Zn+Mn priming treatment as compared with Zn priming alone and
Mn deficiency chlorosis was not reverted by the priming treatment.
Seed priming with Zn+Mn increased both Mn and Zn shoot contents but
concentrations did not increase above the deficiency thresholds.
5.3.4 Effects of micronutrient seed priming in soil culture
In the soil culture experiment on a calcareous Loess sub-soil, both, Mn
and Zn seed priming had a positive effect on shoot elongation and
slightly increased shoot biomass production by approximately 15 - 20
%, while root biomass was not affected (Fig. 5.5).
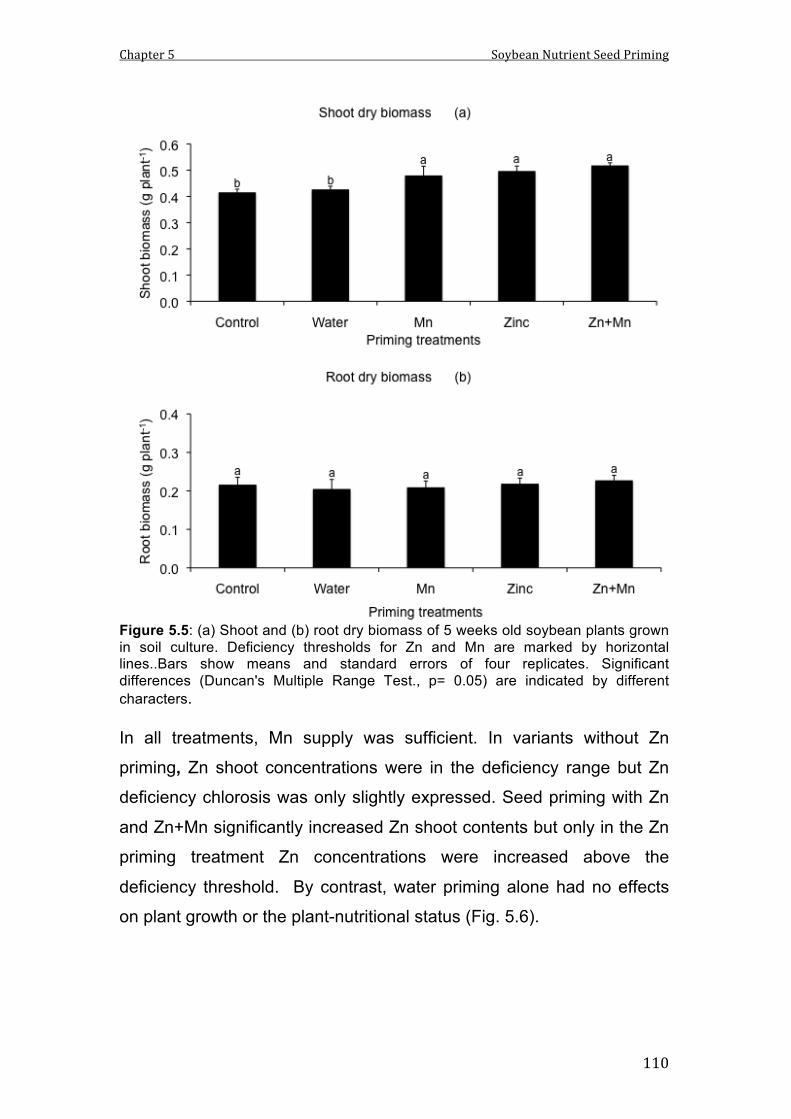
Chapter(5((((((((((((((((((((((((((((((((((((((((((((((((((((((((((((((((((((((((((((((((((((((((((((((((((((((((((((Soybean(Nutrient(Seed(Priming(
( 110(
Figure 5.5: (a) Shoot and (b) root dry biomass of 5 weeks old soybean plants grown in soil culture. Deficiency thresholds for Zn and Mn are marked by horizontal lines..Bars show means and standard errors of four replicates. Significant differences (Duncan's Multiple Range Test., p= 0.05) are indicated by different characters. In all treatments, Mn supply was sufficient. In variants without Zn
priming, Zn shoot concentrations were in the deficiency range but Zn
deficiency chlorosis was only slightly expressed. Seed priming with Zn
and Zn+Mn significantly increased Zn shoot contents but only in the Zn
priming treatment Zn concentrations were increased above the
deficiency threshold. By contrast, water priming alone had no effects
on plant growth or the plant-nutritional status (Fig. 5.6).
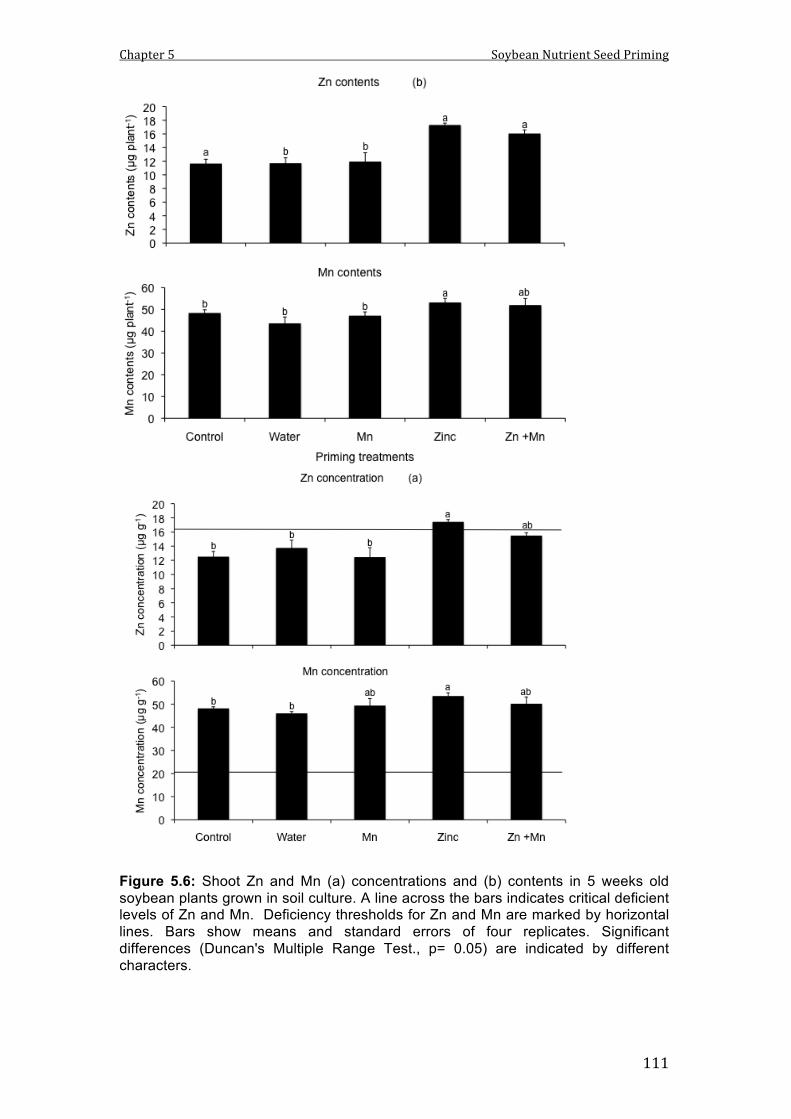
Chapter(5((((((((((((((((((((((((((((((((((((((((((((((((((((((((((((((((((((((((((((((((((((((((((((((((((((((((((((Soybean(Nutrient(Seed(Priming(
( 111(
(((((((((((((((((((((
Figure 5.6: Shoot Zn and Mn (a) concentrations and (b) contents in 5 weeks old soybean plants grown in soil culture. A line across the bars indicates critical deficient levels of Zn and Mn. Deficiency thresholds for Zn and Mn are marked by horizontal lines. Bars show means and standard errors of four replicates. Significant differences (Duncan's Multiple Range Test., p= 0.05) are indicated by different characters.

Chapter(5((((((((((((((((((((((((((((((((((((((((((((((((((((((((((((((((((((((((((((((((((((((((((((((((((((((((((((Soybean(Nutrient(Seed(Priming(
( 112(
5.4 Discussion
5.4.1 Seed priming effects on seedling vigor and micronutrient
status
In the seed priming treatment optimized in this study for soybean
seeds, germination was accelerated during the first 24 h but already
after two days the number of germinated seeds was similar in unprimed
and all water priming treatments (Fig. 5.1a). Accelerated germination
and early seedling development have been similarly reported in maize
and barley as a result of water priming (Ajouri et al., 2004; Harris et al.,
2007). This has been attributed to pre-mobilisation of seed reserves
during the priming period (Bewley, 1997), leaching of germination
inhibitors into the priming solution (Hopkins, 1995), hydrolysis of ABA
(Ajouri et al., 2004) and pre-organisation of membrane structures
(Bewley and Black, 1982). The comparatively moderate effect of seed
priming treatments on germination speed observed in the present study
may be attributed to the fact that priming was performed between
layers of moist filter paper, slowing down the speed of imbibition. This
treatment may also reduce the induction speed for the priming
reactions described above.
However, imbibition and rehydration of dry seeds is a critical process
during germination and particularly large-seeded leguminous plants are
sensitive to imbibition damage. Rapid absorption of water can cause
severe membrane damage leading to leakage of electrolytes, sugars,
and amino acids (Powell and Matthews, 1978). Wuebker et al. (2001).
Bochicchio et al. (1990) reported embryo membrane damage, directly
linked with the speed of imbibition at low seed moisture. Accordingly,
pre-emergence exposure to flooding for 24 hours reduced emergence
of soybean by 50% (Wuebker et al., 2001). Also in the present study,
seed priming by slow imbibition between filter papers for 12 h resulted
in the highest germination rate and the smallest proportion of abnormal

Chapter(5((((((((((((((((((((((((((((((((((((((((((((((((((((((((((((((((((((((((((((((((((((((((((((((((((((((((((((Soybean(Nutrient(Seed(Priming(
( 113(
seedlings (Fig. 5.1b), which was reduced by up to 75 % as compared
with seed soaking directly in priming solutions. Alternatively, osmotic
seed priming has been reported to improve germination rate in Brassica
spp. (Brassica oleracea L.), cabbage (Khan, 1992; Zheng et al., 1994).
During osmotic priming, seeds are slowly moisturized by adjusting the
osmotic potential of the priming solution by using chemically inert but
osmotically active compounds like polyethylene glycol (PEG) for
reducing imbibition damage to seeds during soaking (Khan, 1992).
Nutrient seed priming treatments with Mn and Zn substantially
increased the natural seed content of Zn and Mn by 6 fold and 5 fold,
respectively (Table 5.3). However, 40 – 60 % of the primed nutrients
were found attached to the seed coat as compared to only 10 % in
unprimed seeds. Seed washings with distilled water did not significantly
change Zn and Mn contents suggesting a tight adsorption at the
surface of the seed coat. Similarly an isotope study by Polar (1970)
revealed that only 20.6 % of the total (Zn65) radionuclide absorbed by
the whole seed entered the cotyledons and the embryo of Vicia faba,
while the rest was retained by the seed coat.
Interestingly, the increase of Zn and Mn seed content after combined
seed priming with Zn+Mn was only half as compared with Zn or Mn
priming alone. This finding suggests a competitive interaction of both
nutrients during the priming treatment. However, the differences in seed
contents disappeared when only the remaining seed was analysed after
removal of the seed coat, indicating that competition between Zn and
Mn was mainly restricted to the outer parts of the seeds. Finally,
nutrient seed priming increased Zn and Mn contents in the inner part of
the seeds (embryo + endosperm + cotyledons) by 300 - 400%. Total
seed contents obtained after micronutrient seed priming of approx. 210
mg kg-1 seeds are similar to values reported in earlier studies of Ajouri
et al. (2004) on barley and Harris et al. (2007) on maize.

Chapter(5((((((((((((((((((((((((((((((((((((((((((((((((((((((((((((((((((((((((((((((((((((((((((((((((((((((((((((Soybean(Nutrient(Seed(Priming(
( 114(
5.4.2 Seed priming effects on plant growth
In hydroponics, Zn deficiency reduced shoot growth of 5 weeks old
soybean plants and inhibited shoot-, but not root biomass production
(Fig. 5.3). Inhibition of shoot growth in Zn-deficient plants may be
attributed to increased oxidative degradation or inhibited synthesis of
IAA, as previously suggested by Cakmak et al. (1989). On the other
hand, Zn deficiency can also induce root P deficiency responses, such
as increased expression of high affinity P uptake (Huang et al., 2000),
even in presence of high external P levels as a consequence of Zn
deficiency-induced inhibition of shoot to root re-translocation of P in the
phloem (Zhang et al., 1991). This phenomenon may also be
responsible for the Zn deficiency-induced increase in root/shoot ratio
observed in the present study, which resembles a typical response to P
limitation (Marschner, 2012).
Detrimental effects on early soybean growth due to Zn limitation were
completely reverted by Zn seed priming. However, at final harvest, the
Zn shoot concentration was in the deficiency range, demonstrating that
any surplus of Zn supply was immediately transformed into plant growth
and Zn priming could cover the Zn demand of the plant for a growth
period of maximum 5 weeks (Fig. 5.4a).
Manganese deficiency in the nutrient solution inhibited both, shoot and
root biomass production (Fig. 5.3) associated with intense leaf
intervenal chlorosis. Obviously, Mn deficiency affected photosynthesis
more severely than Zn deficiency (Shenker et al., 2004) as indicated by
stronger expression of leaf chlorosis in Mn-deficient plants. Limited
photosynthesis and limited carbohydrate supply to the roots have been
also reported in earlier studies as main factors limiting root
development and growth of Mn-deficient plants (Campbell and Nable,
1988).
However, in contrast to shoot biomass production, shoot elongation
was not affected by Mn deficiency. This may be attributed to the

Chapter(5((((((((((((((((((((((((((((((((((((((((((((((((((((((((((((((((((((((((((((((((((((((((((((((((((((((((((((Soybean(Nutrient(Seed(Priming(
( 115(
concomitant Zn supply via the nutrient solution, which was sufficient to
maintain IAA production. (Cakmak et al., 1989).
Manganese seed priming was not able to increase shoot Mn
concentrations above the deficiency threshold and the detrimental
effects of Mn deficiency on plant growth were not reverted by the
priming treatments, although the improvement of Mn nutrition was
much stronger as compared with the effect of Zn priming on the Zn
nutritional status (Fig. 5.4a). This may reflect a particularly high Mn
demand of the soybean plants. Moreover, the Mn status of the plants
was negatively affected even by water priming (Fig. 5.4b), an effect that
was not observed in case of Zn nutritional status.
Under combined deficiency of Zn and Mn, the inhibitory effect of Mn
deficiency on root biomass production was not detectable and growth
responses where similar as observed in Zn-deficient plants but with a
stronger expression of leaf chlorosis. This may indicate that the
observed responses to Zn deficiency, such as reduction of shoot
elongation and shoot biomass production but maintenance of root
growth may be induced earlier in the culture period as compared to the
responses to Mn limitation. Therefore, in the absence of both Zn and
Mn supply, the more rapidly induced growth responses to Zn limitation
may override to some extent the negative Mn effect on root growth..
In Zn+Mn priming treatments, about 47 % of primed Zn and 90 % of
primed Mn was recovered in the shoots of 5 weeks old soybean plants
grown in Zn- and Mn-deficient nutrient solutions. Despite of sufficient
translocation of these nutrients to the growing shoot, physiological
mechanisms involved in the translocation of primed nutrients from the
seed coat and other seed components to growing shoot and root during
early plant growth are still unknown. Similarly, seed priming with Zn+Mn
have shown that 50% and 80% of the seed Zn and Mn contents,
respectively, were found in the shoots 4 weeks old maize seedling
grown under Zn and Mn deficient conditions (Imran et al., 2012a).

Chapter(5((((((((((((((((((((((((((((((((((((((((((((((((((((((((((((((((((((((((((((((((((((((((((((((((((((((((((((Soybean(Nutrient(Seed(Priming(
( 116(
In plants, simultaneously primed with Zn and Mn, the beneficial effect of
Zn priming on shoot biomass production was less expressed as
compared with Zn priming alone (Fig. 5.4). This may indicate some
antagonistic interactions between these micronutrients as already
observed for Zn and Mn uptake during the priming treatments. Similar
Zn/Mn antagonisms have been reported for other plant species such as
rice (Ishizuka and Ando, 1968) and maize (Aref, 2012).
In soil culture, both priming treatments with Zn and Mn had a positive
effect on shoot elongation and slightly increased shoot biomass
production while root biomass was not affected (Fig. 5). In all
treatments without additional Zn supply, Zn shoot concentrations were
in the deficiency range. This may reflect an extremely low Zn availability
of the highly calcareous soil (23.3 % CaCO3) used for the experiments,
leading to Zn immobilisation by adsorption to CaCO3 and clay minerals
(Trehan and Sekhon, 1977).
Manganese supply was sufficient in all treatments and due to the
absence of Mn deficiency root growth was not affected. In accordance
with the observed stimulation of shoot growth, seed priming with Zn
increased the shoot concentrations of Zn even above the deficiency
threshold (Fig. 5.6). However, as already observed in the nutrient
solution experiment, this effect was less expressed after combined
priming with Zn and Mn, suggesting the presence of an antagonistic
interaction
5.5 Conclusion
The present study demonstrates that nutrient seed priming, particularly,
with Zn offers perspectives to improve seedling development and early
growth of soybean. Controlled, slow imbibition of soybean seeds
increased beneficial effects of seed priming, demonstrating the
importance of priming conditions avoiding imbibition damage during
priming process. Shoot Zn and Mn contents of soybean plants grown in

Chapter(5((((((((((((((((((((((((((((((((((((((((((((((((((((((((((((((((((((((((((((((((((((((((((((((((((((((((((((Soybean(Nutrient(Seed(Priming(
( 117(
nutrient solution indicate that the primed nutrients are efficiently
translocated within the plant during early seedling development and
could be sufficient to cover the complete Zn demand up to 5 weeks
under limited Zn supply. Beneficial effects of micronutrient seed priming
comprised improvements of early plant growth and micronutrient status.
However, in face of the general importance of Zn and Mn in biotic and
abiotic stress adaptations, further investigations are required for a more
comprehensive assessment of perspectives for micronutrient seed
priming in soybean culture, considering also the physiological
background of the nutrient priming effects as well as their expression
under variable growth conditions.such as. soil types, soil-water and
redox status, fertilisation management, variation in priming treatments
and genotypes.

Chapter(6((((((((((((((((((((((((((((((((((((((((((((((((((((((((((((((((((((((((((((((((((((((((((((((((((((((((((((((((((((((((((((((((((((General(Discussion(
( 118(
Chapter 6
General Discussion
Uniform germination, vigorous seedling development and continuous
nutrient supply to young fast growing seedlings is vital for better crop
establishment and ultimately for maximum yield. A young seedling
passes through various stages for acquiring and utilizing mineral
nutrients during growth, based on the internal and external nutrient
supplies and on crop nutrient requirements (Scaife and Smith 1973). At
initial growth stages, seed mineral reserves are the primary source of
nutrients for young seedling, and only a small part is taken from soil.
During germination process, nutrients stored in the endosperm are
released by the enzymes, synthesized by aleurone and scutellum and
transported to the growing embryo (Fincher, 1989). Therefore, sufficient
mineral nutrient reserves in the seed are crucial to nourish young
seedlings until the development of the root system and its functioning in
supplying the nutrients. This is important for crops, particularly, grown in
soils with low available nutrients for plant growth (Asher, 1987). Seed
mineral contents have been shown to play an important role in the
germination process and early seedling development in different crop
plants (Rengel and Graham, 1995; Hacisalihoglu and Kochian, 2003;
Tehrani et al., 2003, Nadeem et al., 2011).
Application of mineral nutrients to the seeds (seed treatment) prior to
sowing is an efficient and cost-effective approach to improve nutrient
availability to plants at early growth stages (Robert, 1973; Ros et al.,
2000). Farooq et al. (2012) have reported that seed treatment with
different mineral nutrients can improve plant growth and increase grain
yield of field crops. Different approaches are practiced for applying
mineral nutrients to the seed like, such as coating or dressing and
soaking in nutrient solution. Zinc dressing of wheat seeds has shown
significant increase in grain yield (Yilmaz et al., 1998). Zhang et al.

Chapter(6((((((((((((((((((((((((((((((((((((((((((((((((((((((((((((((((((((((((((((((((((((((((((((((((((((((((((((((((((((((((((((((((((((General(Discussion(
( 119(
(1998) have shown that soaking barley seeds in P solution stimulated
seedling growth and plant used the added P as early as three days after
application (Zhang et al., 1998).
In past few years, seed priming has acquired special attention due to its
promising role in improving germination rate and plant growth. Seed
priming is a pre-sowing seed treatment in which seeds are soaked in
water and dried back to storage moisture contents before further use.
Several authors have reported the beneficial affects of seed priming in
improving seed vigour and early seedling development (Taylor et al.,
1998; Harris et al; 1999; McDonald, 2000; Harris, 2006; Farooq et al.,
2009, 2010) for better crop production under optimal and various stress
conditions.
‘Nutrient seed priming’ is a technique in which seeds are soaked in
nutrient solutions instead of simple water. Nutrient seed priming
increases seed nutrient contents along with the priming effects to
improve seed quality for better germination and seedling growth. Maize
seed priming with Zn have shown stimulated plant growth, increase in
grain yield and seed Zn contents on soils with limited Zn supply (Harris
et al., 2007) and under stress of low root-zone temperature (Imran et al.,
2013). In barley, nutrient priming of seeds with P and Zn stimulated
germination, seedling growth and plant uptake of these elements (Ajouri
et al., 2004).
Various abiotic stresses like low or high soil temperature, very low or
high soil pH, flooding and drought conditions are important constraints
for normal seed germination, plant growth and nutrient acquisition. Low
temperature at early spring is a major yield-limiting factor for maize
grown in Central and Northern Europe. Maize seed germination, early
seedling development and plant growth is severely inhibited at low
temperature. Nutrient acquisition, at early developmental stages, is
particularly limited by low temperature due to (i) limited solubility and
availability of certain nutrients and (ii) disrupted root activity.

Chapter(6((((((((((((((((((((((((((((((((((((((((((((((((((((((((((((((((((((((((((((((((((((((((((((((((((((((((((((((((((((((((((((((((((((General(Discussion(
( 120(
Even at optimal environmental conditions, soil properties e.g. soil
structure, pH and soil fertility level play very important role in early
seedling establishment, plant growth, availability and uptake of nutrients
from the soil. Based on the promising results of nutrient seed priming
with respect to improvement of early seedling establishment, plant
growth and stress tolerance, the present thesis investigated the role of
nutrient seed priming in mitigating low root zone temperature stress on
early seedling development and nutrient acquisition in maize. In
addition, effects of nutrient priming were investigated on germination,
plant growth and nutrient uptake in soybean under deficient and limited
nutrient supply. A series of experiments conducted on maize (chapter 3,
4) and soybean (chapter 5) have shown substantial results of nutrient
seed priming in improving germination rate, plant growth and nutrient
acquisition under limited nutrient availability and supply. The following
discussion is focused on the most prominent results of this thesis to a
greater relevance for agricultural crop production under abiotic stress
conditions and use of nutrient seed priming for improvement of plant
growth and nutrient uptake in crop plants.
6.1 Seed water priming improves germination rate and seedling
establishment
Early and healthy seedling establishment is very important for crop
growth in any kind of environmental conditions. Water priming is a
technique in which seeds are soaked in water for a certain time period
and then dried back to storage moisture contents for further use. During
the germination process, initially seed takes up water rapidly to start the
germination process and thereafter, only a small change occurs in seed
water contents until radicle emergence (Bardford, 1986). Seed priming
up to the point, just before the radicle emergence, can be positive for
seed germination, while further extension in priming duration may have
negative effects on seed quality (Ajouri et al., 2004). Water priming has

Chapter(6((((((((((((((((((((((((((((((((((((((((((((((((((((((((((((((((((((((((((((((((((((((((((((((((((((((((((((((((((((((((((((((((((((General(Discussion(
( 121(
been shown to improve germination rate and early seedling
development in some tropical crops, such as rice, chickpea, sorghum
and maize (Harris, 1996). Germination time of different rice cultivars
was reduced up to 50% by water priming in West Africa (Harris and
Jones, 1997). Water priming also showed significant increase in
germination rate and early seedling emergence of barley (Ajour et al.,
2004).
In the present study, water priming of maize seeds for 24 hours by
soaking into distilled water (chapter 4) revealed that germination speed
(Fig. 4.1a) and growth of young seedlings (Fig. 4.1b) was significantly
improved in standard germination tests. The faster germination speed
and improved seedling growth can be attributed to acceleration of a
number of biochemical processes (Fig. 1.2) initiated during controlled
imbibition. It has been shown that water priming of Arabidopsis
(Gallardo et al., 2001) and sugar beet (Job et al., 1997) increased the
polypeptide fraction in the seeds, identified as degradation products of
storage proteins. Abscisic acid (ABA) is involved in maintaining the
seed in the dormant state. Abscisic acid is reported to prevent, radicle
cell wall loosening as a late event during germination (Bewley, 1997),
which leads to the inhibition of radicle extension in the germinating
embryo (Schopfer and Plachy, 1985). Moreover, presence of ABA in the
seed does not affect the osmotic potential and/or water uptake ability of
the embryonic axes (Bewley and Black, 1994). Gibberallin (GA) is
responsible for the synthesis of various hydrolytic enzymes involved in
the mobilization of endosperm reserves (Jacobsen et al., 1995). In
various reports, ABA has shown antagonistic effects on GA activities,
both, in gene expression level and in modulating germination per se
(Constance et al., 2000; Jacobsen et al., 1995; Liu et al., 1994; Steber
et al., 1998). Inhibited activity of GA represses the synthesis of amylase
enzymes in germinating seeds (Gomez-Cadenas et al., 2001). α-
amylase plays an important role in hydrolyzing the endosperm starch

Chapter(6((((((((((((((((((((((((((((((((((((((((((((((((((((((((((((((((((((((((((((((((((((((((((((((((((((((((((((((((((((((((((((((((((((General(Discussion(
( 122(
into sugars as energy source for the newly developing shoot and roots
(Kaneko et al., 2002). Another amylase enzyme, β-1,3-glucanase has
been reported to contribute to loosening of cell wall components for
radicle protrusion (Wu and Bradford, 2003).
It is reported that pre-soaking of seeds causes the hydrolysis of ABA
together with the leaching of other germination inhibitors like coumarin
and phenolic compounds from the seed into solution (Hopkins, 1995).
Therefore, seed priming might result in diminishing the antagonistic
effect of ABA on GA activity during the germination process. Positioning
of short chain fatty acids (C6-C12) is also known to be involved in
inhibition of seed germination. During imbibition, the position of certain
fatty acids associated with seed dormancy is changed and does not
reverse on drying (Bewley and Black, 1982).
Hence, in the process of seed priming, all the necessary physiological
and biochemical changes occurred in the seeds during the priming
period persist upon drying. Therefore, when a primed seed is sown, a
small uptake of water can already stimulates the radicle emergence.
Instead, an unprimed seed has to go through all the phases of seed
germination after sowing under sometimes unfavorable soil conditions,
which is a possible reason for lower germination rate and seedling
growth compared to primed seeds.
In addition, although imbibition is vital for re-constitution of bio-
membranes, activation of enzymes, mobilization of storage compounds
and protein synthesis in the seed, it can also induce imbibition damage,
particularly when water is taken up rapidly.
Powell and Matthews (1978) have reported damages to pea seeds
soaked in free water due to physical damages of cell membranes
leading to impaired cell structure. For soybean seeds cracking of
cotyledons and its vascular bundles has been reported during
uncontrolled imbibition (Kokuryu et al., 2009). Fast absorption of water
increases the leakage of electrolytes, sugars, and amino acids (Powell

Chapter(6((((((((((((((((((((((((((((((((((((((((((((((((((((((((((((((((((((((((((((((((((((((((((((((((((((((((((((((((((((((((((((((((((((General(Discussion(
( 123(
& Matthews, 1978; Egley et al., 1983; Pereira & Andrews, 1985;
Wuebker et al., 2001) and disruption of cell structures affects
germination and seedling growth (Larson, 1968; Bewley and Black,
1978; Hahalis et al., 1996).
Results of the germination tests in soybean (chapter 5) revealed that
water priming between filter paper layers is a suitable method to prime
soybean seeds to reduce imbibition damage to seeds. A significant
decrease in the development of abnormal seedlings (Fig. 5.1) by water
priming between filter paper layers demonstrated that slow uptake of
water during priming not only provides the potential benefits of water
priming, but also decreases the possible damages to the seed during
imbibition. In case of maize, soaking seeds in the water for 24 hours did
not result in any abnormal seedlings, revealing that maize seeds are
more tolerant to imbibition damage as compared with soybean.
In previous studies, it has been reported that various physiological
stress factors including ethanol toxicity, oxygen deprivation, carbon
dioxide accumulation and oxidative damages (Sung, 1995; Sallam and
Scott, 1987; Scott et al., 1989) are affecting seed germination during
and after submerging seeds in water for extended periods of time.
Results of the present study on priming duration suggested that it is
important not only to control the speed of germination but also oxygen
supply and priming duration are important factor in determining the
efficiency of the priming process. Hence, slow germination speed,
development of abnormal seedlings and reduced seedling growth of
soybean and maize occasionally observed in the present study may be
attributed to different reasons (i) disruptive and incomplete activation of
the germination process for shorter priming duration treatments due to
insufficient water uptake in to the seed (ii) anoxic and post anoxic
damages to various seed parts because of longer exposure to
submerged conditions (longer priming durations) in the absence of
oxygen.

Chapter(6((((((((((((((((((((((((((((((((((((((((((((((((((((((((((((((((((((((((((((((((((((((((((((((((((((((((((((((((((((((((((((((((((((General(Discussion(
( 124(
6.2 Increasing seed nutrient contents with nutrient seed priming
Priming maize (chapter 3, 4) and soybean (chapter 5) seeds in nutrient
solutions revealed that nutrient priming is a suitable technique to
increase seed nutrient contents. Concentrations of micronutrients in
priming solutions were calculated to double the amount of seed nutrient
contents based on the seed water absorbing capacity, while for P, it
was calculated to increase 50% in natural seed P content. However,
results of seed nutrient contents in maize (table 3.2, 4.1) and soybean
(Fig. 5.2) showed a relatively higher increase in seed micronutrient
contents than the calculated amount, for example Zn contents were
increased from 500 % (maize) up to 700 % (soybean) in seeds. Uptake
of micronutrients in the seeds higher than the calculated, almost 200 %,
may reflect a high capacity of the seed for micronutrient immobilization
e.g. at the seed coat (since the calculated increase of 200 % refers only
to the soluble micronutrients in the imbibition solution). If micronutrients
were continuously removed from this solution by adsorption to outer
seed structures, this would have induced additional uptake driven by
diffusion. Accordingly, in soybean seeds 40-60 % of primed Zn and Mn
remained at the seed coat. Similarly, in an isotope uptake study (Polar,
1970) about 20.6 % of the total (Zn65) radionuclide absorbed by the
whole seed enters the cotyledons and embryo of Vicia faba, while the
rest was retained by the seed coat. The high binding capacity of seed
coat, may act as filter, for excluding ions occasionally present in
excessive concentrations in the soil solution to avoid toxic effects of
metal ions. This revealed that in case of soybean the seed coat was
the most important sink for removal of micronutrients from the imbibition
solution. In water-primed soybean seed, the natural Zn and Mn contents
were 16 µg seed-1 and 9 µg seed-1, respectively. After Zn and Mn seed
priming, Zn and Mn contents of the remaining seed parts (embryo +
cotyledon) were 47 µg seed-1 and 21 µg seed-1, respectively. Hence,

Chapter(6((((((((((((((((((((((((((((((((((((((((((((((((((((((((((((((((((((((((((((((((((((((((((((((((((((((((((((((((((((((((((((((((((((General(Discussion(
( 125(
seed coat absorbed less than 3% of the total seed water contents,
which reflects that Zn and Mn contents were not much higher than the
calculated amounts based on the seed imbibition capacity, as the
remaining seed parts imbibes most of the water during priming.
After maize seed P priming, seed P contents would have increased by
500 µg seed-1 but increase was 200 µg P seed-1 in natural seed P
contents (965 µg P seed-1), less than 50 % of the calculated amount.
This could be probably due to P uptake already reached the storage
capacity of the seeds or a high osmotic potential of the priming solution
counteracted P uptake. However, the low efficiency of P priming in
terms of plant growth in comparison with micronutrient priming must be
attributed to the high plant demand for P as a macronutrient.
Nutrients stored in the embryo and cotyledons are the primary source of
nutrient supply during germination and early seedling development. As
a result of nutrient seed priming, in addition to an increase in Zn and Mn
contents in the embryo + cotyledons, there was an enormous increase
in seed coat Zn and Mn contents (Table 5.1). The nutrients absorbed in
the seed coat may also be an important source of nutrient supply for
growing plants, as the seed coat remained attached to the cotyledons,
even after seedling emergence. Also shedding and decomposition of
seed coat near the seedling may a be source of nutrient supply as a
localized fertilizer.
Therefore, the results of the present study revealed that increase in
seed nutrient contents via nutrient priming could be a suitable technique
for improving seed quality and nutrient supply for young maize and
soybean seedlings.
6.3 Improving maize and soybean growth under nutrient limitation
via nutrient seed priming.
It is well documented that seed nutrient reserves play an important role
in seedling development and early growth. Increased seed Zn and Mn

Chapter(6((((((((((((((((((((((((((((((((((((((((((((((((((((((((((((((((((((((((((((((((((((((((((((((((((((((((((((((((((((((((((((((((((((General(Discussion(
( 126(
contents via nutrient seed priming have shown pronounced effects on
maize (Chapter 3) and soybean (chapter 5) seedling growth under
nutrient deficient conditions. Particularly, affects of Zn priming have
been enormous on shoot growth, in both maize and soybean under
complete absence of Zn supply.. It has been reported that increased P
uptake and its translocation to the shoot is responsible for Zn deficiency
in crop plants grown, both, in solution or soil cultures (Cakmack and
Marschner, 1987; Loneragan et al. 1982, Cakmak and Marschner
1986). Zhu et al. (2001) reported that highly P-efficient wheat
genotypes inadvertently depress Zn uptake and tissue Zn
concentration. Results of the present study suggests that additional Zn
in the seed via Zn priming was readily available for early plant growth
and was not effected by the antagonistic affects of P due to rapid
translocation to the fast growing shoots. This indicates that Zn priming
might be an appropriate strategy to improve plant Zn status on high P
soils and high lvels of P fertilization.
Stimulation of root biomass production in maize (Fig. 3.2b) and soybean
(Fig. 5.4) may be a reaction to Zn limitation, as Zn deficiency-induced
inhibition of P re-translocation to the roots induces P deficiency
responses in the root tissue, such as increased root/shoot ratio,
increased mycorrhization and up-regulation of high affinity root P uptake
systems (Marschner, 2012).
Root dry biomass was also increased under P deficiency in nutrient
solution and also in soil grown maize plants by P seed priming. The
increase in root-shoot ratio could be attributed to some extent by net re-
translocation of P from shoot to roots in P deficient plants (Smith et al.,
1990). Marschner et al. (1996) have suggested that under P deficient
conditions, a higher proportion of uptaken mineral nutrients from the
substrate is retained in the roots, but also the cycling from shoot to roots
provides additional P to the roots. Even though shoot growth is strongly
reduced under P deficiency but the plant struggles at least to maintain

Chapter(6((((((((((((((((((((((((((((((((((((((((((((((((((((((((((((((((((((((((((((((((((((((((((((((((((((((((((((((((((((((((((((((((((((General(Discussion(
( 127(
chlorophyll concentrations and the photosynthetic activity (Fredeen et
al., 1989; Cakmak, 1994), to ensure the constant supply of
photosynthates to the root as a dominant sink in deficient plants
(Marschner et al., 1996). It is shown that shoot dry biomass markedly
reduced under P deficiency, mainly due to severely inhibited growth of
shoot apex in beans plants (Cakmak, 1994a). However, in both P
deficient and sufficient plants, sucrose concentration in primary leaves
(source) and its export in phloem exudate is maintained, therefore, root
growth is similar in the P deficient and sufficient plants (Cakmak et al.,
1994b). In the present study, maize shoot P concentration (Fig. 3.3) in
P-primed plants showed initial translocation of P to the growing shoot,
thereafter, a continuous decrease in shoot P concentration and increase
in root growth indicates that shoot P was translocated to the growing
roots. Therefore, shoot dry biomass of 4 weeks old maize plants was
severely affected.
In soil culture, maize and soybean plants suffered from Zn deficiency in
all treatments. In calcareous soils, the plant faces various stress factors
such as P, Fe and Zn limitation, high pH and higher levels of
bicarbonates with the potential to induce the accumulation of
carboxylates and phenolics in the root tissue (Neumann and Römheld,
2007). The chelating properties of these compounds may promote Zn
sequestration in root tissues and limit their translocation to the shoot.
Therefore, the marginal surplus of Zn supply to the shoot tissue by
Zn+Mn priming in soil-grown plants also induced only a moderate
stimulation of shoot biomass production (+ 18%) as compared to plants
grown in hydroponics (+ 50%) (Fig. 3.5). An intensive stimulation of root
growth as observed in hydroponics was absent in soil culture. This may
be also attributed to additional stress factors in calcareous soils (e.g.
high pH, high bicarbonate levels (Neumann and Römheld, 2007).
In the present study, maize B priming has not produced the expected
positive results but in a separate study on wheat growth under salt

Chapter(6((((((((((((((((((((((((((((((((((((((((((((((((((((((((((((((((((((((((((((((((((((((((((((((((((((((((((((((((((((((((((((((((((((General(Discussion(
( 128(
stress (8 dSm-1), B seed priming has shown alleviating effects of salt
stress on young wheat seedlings by improving shoot and root growth
(Fig. 6.1, 6.2).
Fig. 6.1: Effect of B seed priming on 2 weeks old wheat seedlings grown under salt
stress (8dSm-1).
Fig. 6.2: Effect of B seed priming on root growth of 4 weeks old wheat seedlings
grown under salt stress (8dSm-1).
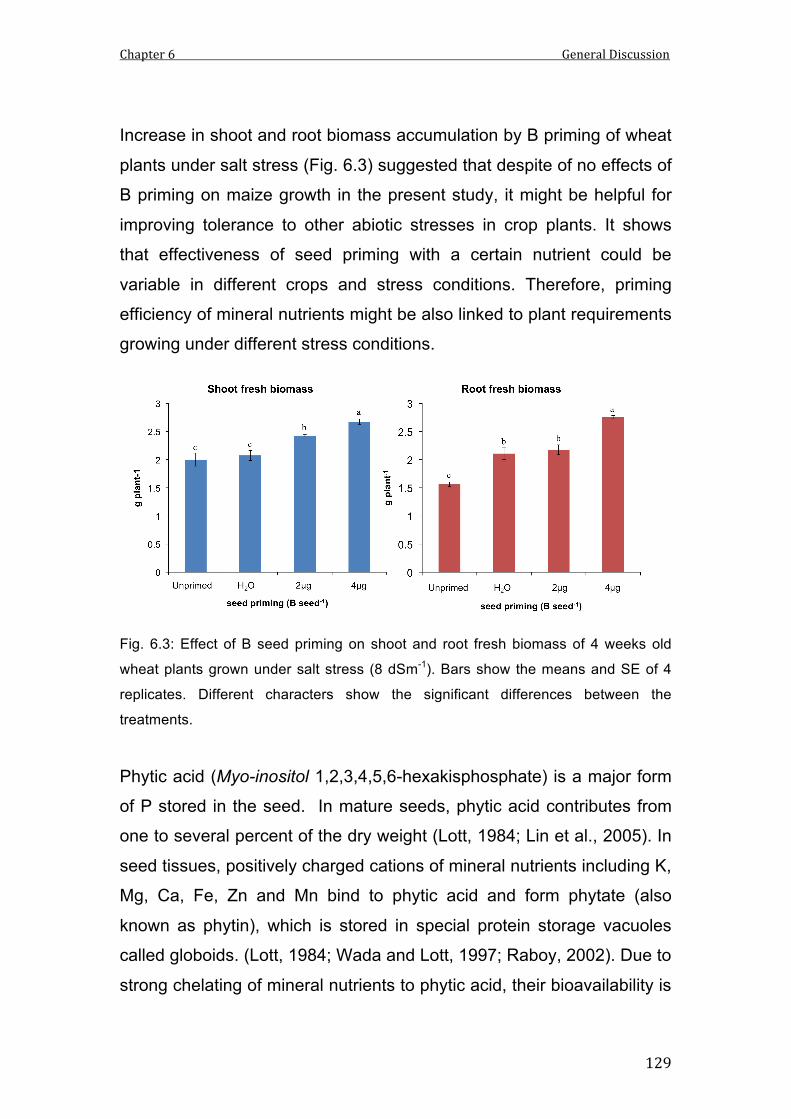
Chapter(6((((((((((((((((((((((((((((((((((((((((((((((((((((((((((((((((((((((((((((((((((((((((((((((((((((((((((((((((((((((((((((((((((((General(Discussion(
( 129(
Increase in shoot and root biomass accumulation by B priming of wheat
plants under salt stress (Fig. 6.3) suggested that despite of no effects of
B priming on maize growth in the present study, it might be helpful for
improving tolerance to other abiotic stresses in crop plants. It shows
that effectiveness of seed priming with a certain nutrient could be
variable in different crops and stress conditions. Therefore, priming
efficiency of mineral nutrients might be also linked to plant requirements
growing under different stress conditions.
Fig. 6.3: Effect of B seed priming on shoot and root fresh biomass of 4 weeks old
wheat plants grown under salt stress (8 dSm-1). Bars show the means and SE of 4
replicates. Different characters show the significant differences between the
treatments.
Phytic acid (Myo-inositol 1,2,3,4,5,6-hexakisphosphate) is a major form
of P stored in the seed. In mature seeds, phytic acid contributes from
one to several percent of the dry weight (Lott, 1984; Lin et al., 2005). In
seed tissues, positively charged cations of mineral nutrients including K,
Mg, Ca, Fe, Zn and Mn bind to phytic acid and form phytate (also
known as phytin), which is stored in special protein storage vacuoles
called globoids. (Lott, 1984; Wada and Lott, 1997; Raboy, 2002). Due to
strong chelating of mineral nutrients to phytic acid, their bioavailability is

Chapter(6((((((((((((((((((((((((((((((((((((((((((((((((((((((((((((((((((((((((((((((((((((((((((((((((((((((((((((((((((((((((((((((((((((General(Discussion(
( 130(
reduced or inhibited in the digestive track of monogastric animals (Zhou
and Erdman, 1995; Lönnerdal, 2003). Development of maize and
soybean mutants for low phytic acid in the seeds have been shown to
improve the bioavailability of mineral nutrient to human and other
monogastric animals (Raboy, 2002; Lönnerdal, 2003).
Recently, Doria et al. (2009) have shown that maize low phytic acid
mutant (lpa1-241) defective in the synthesis of phytic acid during seed
maturation have low germination capacity because of increased
oxidative damages to DNA and other seed proteins. Accordingly, due to
low phytic acid synthesis, alteration in the storage form of mineral
nutrients especially Zn and Mn in the seed would have reduced their
availability during the germination process. This could result in limited
activities of Mn and Zn superoxide dismutases. In this thesis, significant
translocation and utilization of additional Zn and Mn in the seed via
nutrient priming both, in maize and soybean seedlings, suggests their
readily availability for plant growth. Therefore, Zn and Mn seed priming
may be an appropriate strategy to improve germination rate of low
phytate mutants.
6.4 Improving nutritional status of maize and soybean plants grwn
under deficient and limited nutrient available condition by nutrient
seed priming.
Seed dressing or coating of mineral nutrient in soils with low nutrient
availability have been shown an economical and easy method
compared to soil or foliar application (Ajouri et al. 2004; Adhikari and
Rattan, 2000; Slaton et al., 2001; Savithri et al.; 1999; Scott et al.,
1989). Zinc coating of rice seeds or immersing the roots of seedlings in
Zn solution is more efficient than surface application of Zn in calcareous
soils (Giordano and Mordvedt, 1973). Application of nutrients by these
methods require in time availability of the nutrients, may need specific
products, techniques, machinery for foliar application and seed coating;

Chapter(6((((((((((((((((((((((((((((((((((((((((((((((((((((((((((((((((((((((((((((((((((((((((((((((((((((((((((((((((((((((((((((((((((((General(Discussion(
( 131(
furthermore, availability of additional labor might be also required for
handling and sowing. In comparable, nutrient seed priming can be
applied at any time prior to sowing and does not involve extra inputs
other than the nutrients (Ajouri et al., 2004).
In the present study, nutrient seed priming improved the plant nutrient
status of both maize and soybean. Results of shoot mineral contents of
maize grown in nutrient deficient solution culture revealed that more
than 50% of the primed Zn and almost 80% of the Mn reserves were
utilized for shoot growth (Fig. 3.6) until final harvest, that can be
attributed to high mobility of these nutrients in plants.
In contrast to maize plants grown in nutrient solution, in soil grown
plants recovery of Zn seed reserves in shoot of Zn and Zn+Mn primed
plants was low and this level was even lower in water-primed and
unprimed plants. These results can be attributed to low availability of Zn
in the soil culture system (see Materials and methods, Chapter
3),which may also affect Zn distribution within the plants leading to
much lower Zn supply to the shoots as compared with the hydroponic
culture system. In calcareous soils, plants are facing various stress
factors such as P and Fe limitation, high pH and bicarbonate levels with
the potential to induce the accumulation of carboxylates and phenolics
in the root tissue (Neumann and Römheld, 2007.
Although translocation of primed nutrients was reduced in soil culture in
the present study but still it had positive effects on plant growth and
nutrient acquisition. However, there is a need for further investigations
to improve priming efficiency under various soil conditions.
6.5 Improving plant growth and nutrient acquisition at low root
zone temperature stress by nutrient seed priming.
In low root zone temperature experiments on maize, a strong
stimulation of root length (Table 4.3) demonstrated promising effects of
nutrient seed priming on formation of longer and finer roots, particularly

Chapter(6((((((((((((((((((((((((((((((((((((((((((((((((((((((((((((((((((((((((((((((((((((((((((((((((((((((((((((((((((((((((((((((((((((General(Discussion(
( 132(
stimulation of lateral root formation (Fig. 4.2). At low temperature,
reduced uptake and translocation of micronutrients (Zn, Mn, Fe and Cu)
from root to shoot inhibits plant growth (Engels and Marschner, 1996a)
and at the same time limits the translocation of assimilates to the roots
(Engels and Marschner, 1996b). This may cause retarded leaf growth
and limitation of photosynthesis, which strongly depends on the supply
with micronutrients, such as Fe, Mn, Zn. and Cu (Marschner, 1995) with
growth inhibition as a final consequence. Accordingly, Strauss et al.
(2007) reported a functional limitation of photosystem II (PSII) in plants
exposed to low RZT, while Nagasuga et al. (2011) stated that reduced
growth and biomass production of rice plants, grown under low RZT
was mainly attributed to the reduction of shoot growth and leaf area.
Accordingly, the results of the solution culture experiment (Chapter 4)
showed a significant increase in plant leaf area, stomatal conductance
(Fig. 6.4) and net photosynthesis rate per leaf but not per unit leaf area
(Fig. 6.5) in maize plants grown at low root zone temperature.
Results of Zn, Mn and Fe contents in the shoots of maize plants grown
in solution culture (Table 4.6) revealed that improved nutrient
acquisition via nutrient seed priming lead to enhanced root and shoot
growth under low root zone temperature.
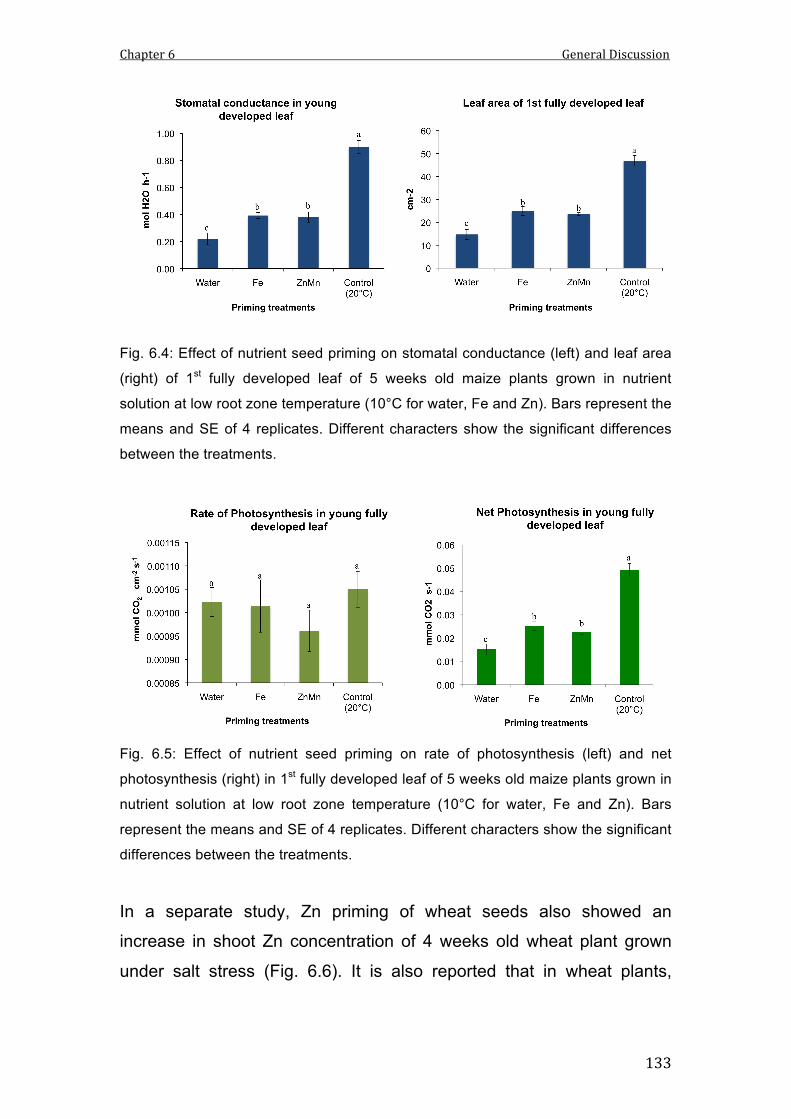
Chapter(6((((((((((((((((((((((((((((((((((((((((((((((((((((((((((((((((((((((((((((((((((((((((((((((((((((((((((((((((((((((((((((((((((((General(Discussion(
( 133(
Fig. 6.4: Effect of nutrient seed priming on stomatal conductance (left) and leaf area
(right) of 1st fully developed leaf of 5 weeks old maize plants grown in nutrient
solution at low root zone temperature (10°C for water, Fe and Zn). Bars represent the
means and SE of 4 replicates. Different characters show the significant differences
between the treatments.
Fig. 6.5: Effect of nutrient seed priming on rate of photosynthesis (left) and net
photosynthesis (right) in 1st fully developed leaf of 5 weeks old maize plants grown in
nutrient solution at low root zone temperature (10°C for water, Fe and Zn). Bars
represent the means and SE of 4 replicates. Different characters show the significant
differences between the treatments.
In a separate study, Zn priming of wheat seeds also showed an
increase in shoot Zn concentration of 4 weeks old wheat plant grown
under salt stress (Fig. 6.6). It is also reported that in wheat plants,
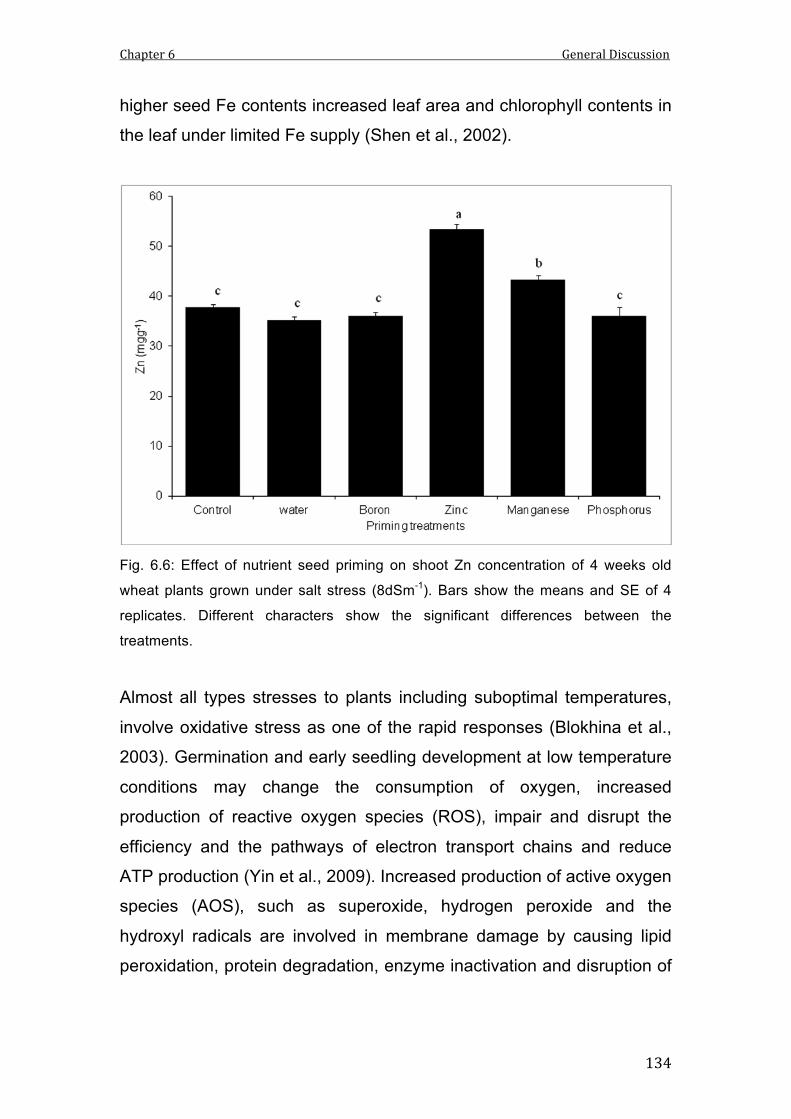
Chapter(6((((((((((((((((((((((((((((((((((((((((((((((((((((((((((((((((((((((((((((((((((((((((((((((((((((((((((((((((((((((((((((((((((((General(Discussion(
( 134(
higher seed Fe contents increased leaf area and chlorophyll contents in
the leaf under limited Fe supply (Shen et al., 2002).
Fig. 6.6: Effect of nutrient seed priming on shoot Zn concentration of 4 weeks old
wheat plants grown under salt stress (8dSm-1). Bars show the means and SE of 4
replicates. Different characters show the significant differences between the
treatments.
Almost all types stresses to plants including suboptimal temperatures,
involve oxidative stress as one of the rapid responses (Blokhina et al.,
2003). Germination and early seedling development at low temperature
conditions may change the consumption of oxygen, increased
production of reactive oxygen species (ROS), impair and disrupt the
efficiency and the pathways of electron transport chains and reduce
ATP production (Yin et al., 2009). Increased production of active oxygen
species (AOS), such as superoxide, hydrogen peroxide and the
hydroxyl radicals are involved in membrane damage by causing lipid
peroxidation, protein degradation, enzyme inactivation and disruption of

Chapter(6((((((((((((((((((((((((((((((((((((((((((((((((((((((((((((((((((((((((((((((((((((((((((((((((((((((((((((((((((((((((((((((((((((General(Discussion(
( 135(
DNA strands (Stewart and Guinn, 1969; Cohn et al., 1979; Allen 1995;
Prassad, 1996).
Phenolic compounds play an important role in suppressing lipid
peroxidation by scavenging free radicals or destroying peroxides (Rice-
Evans et al., 1997). In plants, proline and other low molecular weight
soluble sugars are produced and accumulated in response to stress
injuries (Hu et al., 2006). Due to their strong hydrophilic character, these
molecules are involved in repairing cell damages by adjusting the water
potential and maintaining the biological activities of other
macromolecules (Rathinasabapathi, 2000; Watanabe et al., 2000).
Recently, Guan et al. (2009) have reported the accumulation of proline
in maize plants in response to chilling stress tolerance.
Micronutrients, such as Zn and Mn are important co-factors of various
enzymes involved in the detoxification of AOS, such as superoxide
dismutases (SODs, Cakmak and Marschner, 1988 a, b, 1992; Cakmak,
2000). Deficiency of Zn, Mn and Cu alters the activities of SOD forms
and these changes depend on the type and the duration of the
deficiency stress (Yu and Rengel, 1999). Moreover, Mn is a cofactor of
various enzymes involved in the biosynthesis of phenolic compounds
with anti-oxidative properties (Thompson and Huber, 2007). Zinc has
functions in direct membrane stabilization, in biosynthesis of auxins
(Salami and Kenefick, 1970) and gibberellins (Sekimoto et al., 1997)
involved in plant growth regulation and in protein synthesis in general.
Therefore, limited supply of these micronutrients under conditions of low
RZT (Marschner and Engels, 1994) may increase oxidative damage to
the root cells and induce disturbances in plant growth. In the present
study, increased concentrations of total phenolics but decreased
concentrations of the stress metabolite proline (Table 6.1) in roots of
Zn/Mn-primed maize plants grown in soil culture at low root zone
temperature, may indicate alleviated root damage by stimulating
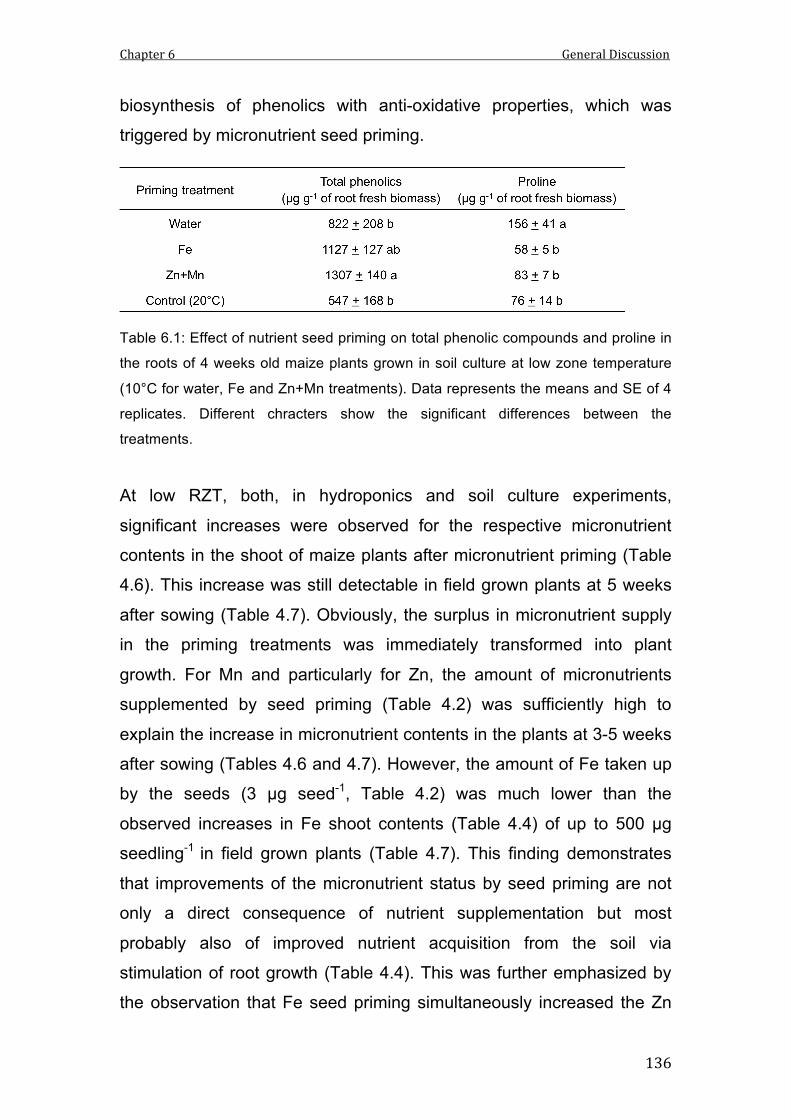
Chapter(6((((((((((((((((((((((((((((((((((((((((((((((((((((((((((((((((((((((((((((((((((((((((((((((((((((((((((((((((((((((((((((((((((((General(Discussion(
( 136(
biosynthesis of phenolics with anti-oxidative properties, which was
triggered by micronutrient seed priming.
Table 6.1: Effect of nutrient seed priming on total phenolic compounds and proline in
the roots of 4 weeks old maize plants grown in soil culture at low zone temperature
(10°C for water, Fe and Zn+Mn treatments). Data represents the means and SE of 4
replicates. Different chracters show the significant differences between the
treatments.
At low RZT, both, in hydroponics and soil culture experiments,
significant increases were observed for the respective micronutrient
contents in the shoot of maize plants after micronutrient priming (Table
4.6). This increase was still detectable in field grown plants at 5 weeks
after sowing (Table 4.7). Obviously, the surplus in micronutrient supply
in the priming treatments was immediately transformed into plant
growth. For Mn and particularly for Zn, the amount of micronutrients
supplemented by seed priming (Table 4.2) was sufficiently high to
explain the increase in micronutrient contents in the plants at 3-5 weeks
after sowing (Tables 4.6 and 4.7). However, the amount of Fe taken up
by the seeds (3 µg seed-1, Table 4.2) was much lower than the
observed increases in Fe shoot contents (Table 4.4) of up to 500 µg
seedling-1 in field grown plants (Table 4.7). This finding demonstrates
that improvements of the micronutrient status by seed priming are not
only a direct consequence of nutrient supplementation but most
probably also of improved nutrient acquisition from the soil via
stimulation of root growth (Table 4.4). This was further emphasized by
the observation that Fe seed priming simultaneously increased the Zn

Chapter(6((((((((((((((((((((((((((((((((((((((((((((((((((((((((((((((((((((((((((((((((((((((((((((((((((((((((((((((((((((((((((((((((((((General(Discussion(
( 137(
and Mn status of the plants and vice versa (Table 4.6) and that also P
content, as an important sparingly soluble macronutrient, was increased
by the priming treatments.
Shen et al., (2002) reported that wheat plants derived from seeds with
high Fe contents showed higher release of phytosiderophores in Fe-
deficient growth medium that leads to increased acquisition of Fe.
Therefore, it should be further elucidated whether Fe priming increases
the release of phytosiderophores in maize plants under low RZT.
Micronutrient seed priming treatments produced a 10-15 % increase in
final grain yield in two independent field experiments conducted in 2010
and 2011 (Table 4.8). The persistence of the priming effects may be
particularly attributed to the enormous stimulation of root growth
observed in model experiments (chapter 3 and 4). This may act as an
advantage not only during stress treatments but may further promote
plant growth during and even after the recovery phase. Moreover, other
studies have demonstrated long-lasting positive effect of micronutrient
seed priming also on alleviating other stress factors. Priming of barley
seeds with Zn significantly improved seed germination, plant growth and
water use efficiency by 44% in 4 weeks-old drought stressed barley
plants (Ajouri et al., 2004). Harris et al., (2007) have reported almost
40% increase in grain yield in maize grown on a Zn deficient soil in
Pakistan.
6.6 Commercial application
Different seed and fertilizer distribution companies are formulating and
selling various products for seed treatment (seed dressing and seed
coating) to improve nutrient uptake in crop plants. ‘Nutrient seed
priming’ is a relatively new approach to increase seed micronutrient
contents (not just soaking into fertilizer solution) as described by Al
Mudaris and Jutzi (1999) and Asgedom and Becker (2001). Some

Chapter(6((((((((((((((((((((((((((((((((((((((((((((((((((((((((((((((((((((((((((((((((((((((((((((((((((((((((((((((((((((((((((((((((((((General(Discussion(
( 138(
fertilizer companies are already selling different solution mixtures of
macro and micronutrients for nutrient seed priming. In Australia, ‘RLF®
The trace element company’ is selling BNS Super Strike® containing P
and micronutrients for seed priming.
In the present study, varying affects of priming methods and different
priming nutrients on germination, plant growth and nutrient acquisition
revealed that the selection of appropriate methods and priming duration
is very important to attain potential benefits of seed priming. In addition,
the selection of particular nutrients and their concentrations in the
priming solution also play a critical role in achieving desired goals of
nutrient seed priming. Priming seeds with low or very high nutrient
concentrations may induce negative effects on seed quality resulting in
decreased germination rate (Ajouri et al., 2004).
In this thesis, Zn priming alone has shown enormous results on plant
growth, but effectiveness of primed Zn was reduced when used
together with Mn, both in maize and soybean. Similarly, as discussed
earlier, B priming improved wheat growth under salt stress but had no
stimulating effect on maize growth. Accordingly, priming solution
containing different nutrients suitable for a crop species might be toxic
for other crops. Figure 6.7 shows soybean growth after seed priming
with a mixture of Zn, Mn, B and P nutrient solution suitable for maize
seed priming (chapter 3). Soybean seed priming with Zn, Mn, B and P
(simultaneously applied in the priming solution) caused leaf shape
deformation. The physiological reasons of these effects need further
elucidation.
For seed producing companies and farmers with larger farm areas, it
might not be easy to manage and handle larger quantities of seeds to
prime at the same time. Hence, persistence of the priming effects during
storage and seed treatment with pesticides is also important for
industrial application of nutrient seed priming. The effect of water
priming on germination in barley seeds was maintained until 9 weeks of

Chapter(6((((((((((((((((((((((((((((((((((((((((((((((((((((((((((((((((((((((((((((((((((((((((((((((((((((((((((((((((((((((((((((((((((((General(Discussion(
( 139(
storage (Ajouri et al., 2004). The persistence of positive affects of
nutrient seed priming should be also investigated for different crop
species.
Fig. 6.7: Effect of P and micronutrients (Zn, Mn and B) seed priming on shoot growth
of 4 weeks old soybean plants.
Therefore, to address various problems and limitations in commercial
application of nutrient seed priming, seed and fertilizer industry should
be actively involved for optimizing nutrient priming for commercial
applications.

Chapter(6((((((((((((((((((((((((((((((((((((((((((((((((((((((((((((((((((((((((((((((((((((((((((((((((((((((((((((((((((((((((((((((((((((General(Discussion(
( 140(
6.7 Future outlook
This thesis demonstrated positive effects of nutrient seed priming in
maize and soybean during early seedling establishment and nutrient
uptake under different plant growth conditions. The physiological role of
primed nutrients needs to be investigated more in detail. The following
section summarises some ideas for further research:
1. Study of the physiological and biochemical changes occurring in
the seed during nutrient seed priming and germination to
elucidate the role of primed nutrients in germination process
under optimal and various stress conditions.
2. Synchrotron X-ray fluorescence analysis of seeds to determine
the localization and forms of nutrients in the seeds.
3. Differential expression of genes responsible for nutrient transport
and synthesis of secondary metabolites in plants and seeds
should be analysed under different growth conditions.
4. Investigating the expression of nutrient seed priming effects
under different environmental conditions e.g. soil type, soil-water
and redox status, fertilisation management, variation in priming
treatments and genotypes etc.

! !!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!References!
! 141
References
Abadía J, Morales F, Abadía A. (1999). Photosystem II efficiency in low chlorophyll, iron deficient leaves. Plant and Soil. 215:183–192. Abbott AJ. (1967). Physiological effects of micronutrient deficiencies in isolated roots of Lycopersicon esculentum. New Phytol. 66:419–437. Adhikari T, Rattan RK. (2000). Modelling Zn uptake by rice crop using a Baber-cushman approach. Plant and soil. 227:235-242. Ajouri H, Asgedom H, Becker M. (2004). Seed priming enhances germination and seedling growth of barley under conditions of P and Zn deficiency. J Plant Nutr Soil Sci. 167:630-636. Allen DJ, Ort DR. (2001) Impact of chilling temperatures on photosynthesis in warm climate plants. Trends Plant Sci. 6:36–42. Alvarez-Uria P, Korner C. (2007). Low temperature limits of root growth in deciduous and evergreen temperate tree species. Funct. Ecol. 21:211–218. An P, Inanaga S, Kafkafi U, Lux A, Sugimoto Y. (2001). Different effect of humidity on growth and salt tolerance of two soybean cultivars. Biol. Plant. 3:405-410.
Arif M, Jan MT, Khan NU, Khan A, Khan MJ, Munir I. (2010). Effect of seed priming on growth parameters of soybean. Pak. J. Bot., 42: 2803-2812.
Arif M, Ali S, Shah A, Javid N, Rashid A. (2005). Seed priming maize for improving emergence and seedling growth. Sarhad J. Agric. 21:539–543.
Arif M, Jan MT, Marwat KB, Khan MA. (2007). Seed priming improves emergence and yield of soybean. Pak. J. Bot., 40: 1169-1177.
Arisoy Z, Erdem H, Yazici A, Gokmen O, Ozturk L, Horst W J. (2010). Biofortification and localization of zinc in wheat grain. J. Agric. Food Chem. 58: 9092–9102. Arshad MA, Azooz RH. (1996). Tillage effects on soil thermal properties in a semiarid cold region. Soil Sci. Soc. Am. J. 60:561–567. Arulanathan AR, Rao IM, Terry N. (1990). Limiting factors in photosynthesis. VI. Regeneration of ribulose 1,5 bisphophate limits photosynthesis at low photochemical capacity. Plant Physiol. 93:1466–1475. Asgedom H, Becker M. (2001). Effect of seed priming with nutrient solutions on germination, seedling growth and weed competitiveness of cereals in Eritrea. In: Proceedings of the Deutscher Tropentag. Univ. of Bonn & ATSAF, Margraf Pub. Press, Weickersheim, p. 282.

! !!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!References!
! 142
Asher CJ. (1987). Crop nutrition during the establishment phase: role of seed reserves. In I.M. Wood, WH Hazard and F From, eds., Crop establishment Problems in Queensland: Recognition, Research and Resolution. Aust. Inst. Agric. Sci. Occasional Publication No. 20. Atkin RK, Barton GE, Robinson DK. (1973). Effect of root-growing temperature on growth substances in xylem exudate of Zea mays. J Exp Bot. 24: 475–48 Atkins CA, Patterson BD, Graham D. (1972). Plant carbonic anhydrases. I. Distribution of type among species. Plant Physiol. 50: 214-217. Auld D S, Bergman T. (2008). The role of zinc for alcohol dehydrogenase structure and function. Cell. Mol. Life Sci. 65, 3961–3970. Barber SA. (1984) Soil Nutrient Bioavailability: A Mechanistic approach. John Wiley and Sons New York Barber J, Ford RC, Mitchell RAC, Millner PA. (1984). Chloroplast thylakoid membrane fluidity and its sensitivity to temperature. Planta. 161: 375-380. Barber S A. (1995). Soil Nutrient Bioavailability: A Mechanistic Approach, 2nd ed., pp. 414. John Wiley, New York. Bradford KJ. (1986). Manipulation of seed water relations via osmotic priming to improve germination under stress conditions. Horti. Sci. 21, 1105-1112. Barr R, Böttger M, Crane FL. (1993). The effect of boron on plasma membrane electron transport and associated proton secretion by cultured carrot cells. Biochem. Mol. Biol. Int. 31: 31-39. Bassil E, Hu HN, Brown PH. (2004). Use of phenylboronic acids to investigate boron function in plants. Possible role of boron in transvacuolar cytoplasmic strands and cell-to-wall adhesion. Plant Physiol. 136, 3383–3395. Bationo, Mokwunye AU. (1991). Role of manures and crop residue in alleviating soil fertility constraints to crop production: With special reference to the Sahelian and Sudanian zones of West Africa. 29:117-125 Beauchamp EG, Lathwell DJ. (1966). Effect of root zone temperature on corn leaf morphology. Can. J. Plant Sci. 46, 593–601. Becraft PW. (2007). Aleurone Cell Development. In: Olsen O-A (ed) Endosperm: Developmental and Molecular Biology. Springer, pp 45–56. Bell RW, McLay L, Plaskett D, Dell B, Loneragan JF. (1989). Germination and vigour of black gram (Vigna mungo (L.) Hepper) seed from plants grown with and without boron. Aust. J. Agric. Res. 40, 272–279.

! !!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!References!
! 143
Bettger WJ, O’Dell BL. (1981). A critical physiological role of zinc in the structure and function of biomembrames. Life Sci. 28: 1425-1438.
Bewley JD. (1997). Seed germination and dormancy. Plant Cell. 9:1055-1066. 1997.
Bewley JD, Black M. (1982). Physiology and biochemistry of seeds. Vol. 2, Viability, dormancy and environmental control. Springer-Verlag, Berlin. Bill Wiebold. MU Agronomy Extension Web Site . http://www.psu.missouri.edu/soyx Blacklow WM. (1972). Influence of temperature on germination and elongation of the radicle and shoot of corn (Zea mays L.). Crop Sci. 12, 647–650. Blevins DG, Lukaszewski KM. (1998) Boron in plant structure and function. Annu. Rev. Plant Physiol. Plant Mol. Biol. 49: 481-500. Bliss RD, Platt-Aloia KA, Thomson WW. (1986). Osmotic sensitivity in relation to salt sensitivity in germinating barley seeds. Plant Cell Environ. 9:721–725. Blokhina O, Violainen E, Fagerstedt KV. (2003) Antioxidants, oxidative damage and oxygen deprivation stress: a review. Ann. Bot. 91:179–194. Bloom AJ, Zwieniecki MA, Passioura JB, Randall LB, Holbrook NM, St Clair DA. (2004). Water relations under root chilling in a sensitive and tolerant tomato species. Plant, Cell and Environment. 27: 971–979. Bohnsack CW. Albert LS. (1977). Early effects of boron deficiency on indoleacetic acid oxidase levels of squash root tips. Plant Physiol. 59:1047–1050. Bolland MDA, Baker MJ. (1988) High phosphorus concen- trations in seed of wheat and annual medic are related to higher rates of dry matter production of seedlings and plants. Aust J Exp Agric. 28:765–770. Bougne S, Job C, Job D. (2000). Sugarbeet seed priming: solublization of the baic subunit of 11– S globulin in individual seeds, Seed Sci. Res. 10: 153 -161. Bouzayen M, Felix G, Latché A, Pech JC, Boller T. (1991). Iron: an essential cofactor for the conversion of 1-aminocyclopro-pane-1-carboxylic acid to ethylene. Planta. 184:244–247. Bowen GD, Rovira AD. (1991). The rhizosphere, the hidden half of the hidden half. In The Plant Roots, the Hidden Half (Y. Waisel, A. Eshel, U. Kafkafi, eds.), pp. 641–669. Marcel Dekker, Inc. New York.

! !!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!References!
! 144
Bradford KJ, Bewley JD. (2002). Seeds; Biology, Technology and Role in Agriculture. Chapter 9, pp. 210-239. In: Plnats, Genes and Biotechnology ( eds. ChrispeelsMJ, Sadava DE), Jones and Bartlet, Boston Bradford KJ. (1995). Water relations in seed germination. p. 351–396. In J. Kigel and G. Galili (ed.) Seed development and germination. Marcel Dekker, New York. Bramlage WJ, Leopold AC, Parrish PJ. (1978). Chilling stress to soybeans during imbibition. Plant Physiol. 61:525-529. Bravo FP, Uribe EG. (1981). Temperature dependence of the con- centration kinetics of absorption of phosphate and potassium in corn roots. Plant Physiol. 67:815–819.
Brown PH, Cakmak I, Zhang Q. (1993). Form and function of zinc in plants. In Zinc in Soils and Plants. Ed. A D Robson. pp 93–106, Kluwer Academic Publishers, Dordrecht.
Brown PH, Bellaloui N, Hu H and Dandekar A. (1999). Transgenically enchanced sorbitol synthesis facilitates phloem boron transport and increases tolerance of tobacco to boron deficiency. Plant Physiol. 119:17-20. Brown PH, Bellaloui N, Wimmer MA, Bassil ES, Ruiz J, Hu H, Pfeffer H, Dannel. (2004). IFA International Symposium on micronutrients. 23-25. February 2004, New Delhi, India Brown PH, Schreier H, Shah PB, Lavkulich LM. (1999). Modelling of soil nutrient budgets: an assessment of agricultural sustainability in Nepal Soil Use and Management. 15: 101-106. Brown TL, Lemay E. (1981). Chemistry: The Central Science. (2nd eds) Prentic-Hall Inc Eaglewood Cliffs, New Jersey. Brown PH. (2008). Micronutrient use in agriculture in the United States of America: current practices, trends and constraints. In Micronutrient Deficiencies in Global Crop Production (B. J. Alloway, ed.). Springer. Brown PH, Cakmak I, Zhang Q. (1993). Form and function of zinc plants. In Zinc in Soils and Plants. Ed. A.D. Robson. pp. 94-106. Kluwer Academic Publishers, Dordrecht. Burnell JN. (1990) Immunological study of carbonic anhydrase in C, and C, plants using antibodies to maize cytosolic and spinach chloroplastic carbonic anhydrase. Plant Cell Physiol 31: 423-427. Burnell J N. (1988). The biochemistry of manganese in plants. In Manganese in Soils and Plants (R. D. Graham, R. J. Hannam and N. C. Uren, eds.), pp. 125–137. Kluwer Academic Publ. Dordrecht.

! !!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!References!
! 145
Cabrera ER, Lansakara H. Open Storage of Soybean Seed in Mississippi. Office of Agricultural Communications, Division of Agriculture, Forestry, and Veterinary Medicine, Mississippi State University. Caffarri S, Frigerio S, Olivieri E, Righetti PG, Bassi R. (2005). Differential accumulation of Lhcb gene products in thylakoid membranes of Zea mays plants grown under contrasting light and temperature conditions. Proteomics. 5: 758-768. Cakmak I. (2000) Possible role of zinc in protecting plant cells from damage by reactive oxygen species. New Phytologist .146: 185–205. Cakmak I, Römheld V. (1997). Boron deficiency-induced impairments of cellular functions in plants. Plant and Soil. 193: 71-83. Cakmak I, Marschner H. (1988a). Increase in membrane permeability and exudation in roots of zinc deficient plants. J Plant Physiol. 132:356-361. Cakmak I, Marschner H. (1988b). Zinc-dependent changes in ESR signals NADPH oxidase and plasma membrane permeability in cotton roots. Physiol Plant. 73:182-186. Cakmak I, Marschner H. (1992). Magnesium deficiency and high light intensity enhance activities of superoxide dismutase, ascor-bate peroxidase, and glutathione reductase in bean leaves. Plant Physiol. 98: 1222-1227. Cakmak I. (2004). Identification and correction of widespread zinc deficiency in Turkey – a success story (a NATO-Science for Stability Project). Proceedings of the International Fertiliser Society 552: 1–26. Cakmak I. (2002). Plant nutrition research: Priorities to meet human needs for food in sustainable ways. Plant and Soil. 247: 3–24. Cakmak I, Marschner H. (1993). Effect of zinc nutritional status on activities of superoxide radical and hydrogen peroxide scavenging enzymes in bean eaves. In Plant Nutrition. From Genetic Engineering to Field Practice. Proceedings of the 12th International Plant Nutrition Colloquium. Ed. N.J. Barrow. pp. 133-137. Kluwer Academic Publishers, Dordrecht. Cakmak I, Marschner H. (1998c) Enhanced superoxide radical production in roots of zinc deficient plants. Journal of Experimental Botany. 39:1449 - 1460. Cakmak I, Marschner H, Bangerth F. (1989). Effect of zinc nutri- tional status on growth, protein metabolism and levels of indole-3- acetic acid and other phytohormones in bean (Phaseolus vulgaris L.). J. Exp.Bot. 40:405–412. Cakmak I, Pfeiffer WH, McClafferty B. (2010b). Biofortification of durum wheat with zinc and iron. Cereal Chem. 87:10–20.

! !!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!References!
! 146
Cakmak I, Sari N, Marschner H, Kalaycı M, Yılmaz A, Eker S, Gulut KY. (1996b). Dry matter production and distribution of zinc in bread and durum wheat genotypes differing in zinc efficiency. Plant and Soil. 180:173–181. Cakmak I, Yilmaz A, Ekiz H, Torun B, Erenoglu B, Braun H J. (1996a). Zinc deficiency as a critical nutritional problem in wheat production in Central Anatolia. Plant and Soil: 180:165–172. Campbell LC, Nable RO. (1988). Physiological functions of manganese in plants. In Manganese in Soils and Plants (R. D. Graham, R. J. Hannan and N. C. Uren, eds.), pp. 139–154. Kluwer Academic Publ., Dordrecht. Castillo AG. (1992). A study of production factors affecting seed vigour in garden peas (Pisum sativum L.) and the relationships between vigour tests and seed lot field and storage performance. Unpublished PhD thesis, Massey University, Palmerston North, New Zealand. Cava F, Zafra O, da Costa MS, Berenguer J. (2008). The role of the nitrate respiration element of Thermus thermophilus in the control and activity of the denitrification apparatus. Environ Microbiol. 10:522–533. Chassot A, Richner W. (2002). Root characteristics and phosphorus uptake of maize seedlings in a bilayred soil. Agronomy Journal. 94:118 – 127. Chassot A. (2000). Early growth or roots and shoots of maize as affected by tillage-induced changes in soil physical properties. ETH Zurich, Diss. ETH No. 13907, Zu- rich, Switzerland. Chouliaras V, Therios I, Molassiotis A, Patakas A, Diamantidis G. (2004) Effect of iron deficiency on gas exchange and catalase and peroxidase activity in citrus. J. Plant Nutr. 27:2085–2099. Chung HJ, Ferl RJ. (1999). Arabidopsis alcohol dehydrogenase expression in both shoots and roots is conditioned by root growth environment. Plant Physiology. 121: 429-436. Clarkson DT, Earnshaw MJ, White PJ, Cooper HD. (1988). Temperature dependent factors influencing nutrient uptake: an analysis of responses at different levels of organization. In Plants and Temperature (S. P. Long and F. I. Woodward, eds.). Symposium of the Society for Experimental Biology. 42:281–309. Company of Biologists, Cambridge. Clarkson DT, Jones LHP, Purves JV. (1992). Absorption of nitrate and ammonium ions by Lolium perenne from flowing solution cultures at low root temperatures. Plant, Cell Environ. 15:99–106.

! !!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!References!
! 147
Codignola A, Verotta L, Panu P, Maffei M, Scannerini S. Bonfante-Fasolo P. (1989). Cell wall bound-phenols in roots of vesicular-arbuscular mycorrhizal plants. New Phytol. 112:221–228.
Cohn MA. (1996). Operational and philosophical decisions in seed dormancyr esearch. Seed Sci. Res. 6:147-153.
Coleman JE. (1992). Zinc proteins: enzymes, storage proteins, tran- scription factors, and replication proteins. Annu. Rev. Biochem. 61:897–946. Conroy JP, Virgona JM, Smillie RM, Barlow EW. (1988). Influence of drought acclimation and CO2 enrichment on osmotic adjustment and chlorophyll a flouresence of sunflower during drought. Plant Physil. 86:1108-1115. Cooper AJ. (1973) Root temperature and plant growth – a review. Res Rev no 4, Commonwealth Bureau of Horticulture and Plantation Crops, Commonwealth Agricultural Bureau, Farnham Royal, England Cooper CS, MacDonald PW. (1970). Energetics of early seedling growth in corn (Zea mays L). Crop Science. 10:136–138. Cordovilla MP, Ocana A, Ligero A, Lluch C. (1995). Salinity effects on growth analysis and nutrient composition in four grain legumes Rhizobium symbiosis. J. Plant Nutr. 18:1595–1609. Corpas FJ, Barroso JB, Del Río LA. (2001). Peroxisomes as a source of reactive oxygen species and nitric oxide signal molecules in plant cells. Trends in Plant Science. 6:145–150. Covey-Crump EM, Attwood RG, Atkin OK. (2002). Regulation of root respiration in two species of Plantago that differ in relative growth rate: the effect of short- and long-term changes in temperature. Plant Cell and Environment. 25: 1501–1513. Crosbie J, Longnecker NE, Robson AD. (1994). Seed Manganese Affects the Early Growth of Lupins in Manganese deficient Conditions. Aust. J. Agric. Res. 45: 1469-1482. Cumbus IP. (1985). Development of wheat roots under zinc deficiency. Plant and Soil. 83: 313–316. Cutforth HW, Shaykewich CF, Cho CM. (1986). Effect of soil water and temperature on corn (Zea mays L) root growth during emergence. Canadian Journal of Soil Science. 66:51- 58 Constance NW, William M, Proebsting PH, Carol JR. (2000). Gibberellins and Seed Development in Maize. I. Evidence That Gibberellin/Abscisic Acid Balance Governs Germination versus Maturation Pathways1. Plant Physiology.122: 1081–1088.

! !!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!References!
! 148
Doria E, Luciano G, Lucia C, Calogero P, Roberto P, Elena C, Nielsen E. (2009). Phytic acid prevents oxidative stress in seeds: evidence from a maize (Zea mays L.) low phytic acid mutant. J. Exp Bot. 60: 967–978 Dannel F, Pfeffer H, Römheld V. (2002). Update on boron in higher plants uptake, primary translocation and compartmentation. Plant Biology. 4: 193–204. Dannel F, Pfeffer H, Römheld V. (2000). Characterization of root boron pools, boron uptake and boron translocation in sunflower using the stable isotopes B10and B11. Aust. J. Plant Physiol. 27: 397–405. Delouche JC. (1980). Environmental effects on seed development and quality. Hort. Sci. 15:775-780. DeMarco DG. (1990). Effect of seed weight, and seed phosphorus and nitrogen concentrations on the early growth of wheat seedlings. Aust J Exp Agric. 30:545-549. Derrick JW, Ryan MH. (1998). Influence of seed phosphorus content on seedling growth in wheat: Implications for organic and conventional farm management in South East Australia. Biol. Agric. Horticul. 16:223–237. Dibb DW, Fixen PE, Murphy SE. (1990). Balanced fertilization with particular reference to phosphates: Interaction of phosphorus with other inputs and management practices. Nutrient Cycling in Agroecosystems. 26:29-52. Douglas JL, Grabowski JM, Keith BCA. (2000). Comparison of seed cleaning techniques for improving quality of eastern gamagrass seed. Seed Science and Technology. 28: 163-167. Droillard MJ, Paulin A. (1990). Isoenzymes of superoxide dismutase in mitochondria and peroxisomes isolated from petals of car- nation (Dianthus caryophyllus) during senescence. Plant Physiol. 94:1187–1192. Duffus CM, Rosie R. (1976). Changes in Trace Element Composition of Developing Barley Grain. J Agric Sci. 87:75–79. Duncan WG, Hesketh JD. (1968). Net photosynthetic rates, relative leaf growth rates and leaf numbers of 22 races of maize grown at eight temperatures. Crop Sci. 8:670–674. Dwivedi R, Snehi Takkar PN. (1974). Ribonuclease activity as an index of hidden hunger of zinc in crops. Plant and Soil. 40: 173-181. Egli DB, TeKrony DM, Heitholt JJ, Rupe J. (2005). Air Temperature During Seed Filling and Soybean Seed Germination and Vigor. Crop Sci. 45:1329-1335. Ehdaie B, Merhaut DJ, Ahmadian S, Hoops AC, Khuong T, Layne AP, Waines JG.

! !!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!References!
! 149
(2010) Root System Size Influences Water-Nutrient Uptake and Nitrate Leaching Potential in Wheat. Journal of Agronomy and Crop Science 196: 455–466. Ellis RJ, Davis JF, Thurlow DL. (1964). Zinc avilability in calcareous Michigan soils as influenced by phosphorus level and temperature. Soil Sci Soc Am Proc. 28:83-86. Ellis RJ. (1979). The most abundant protein in the world. Trends Biochem. Sci. 4: 241–244. Elstner EF. (1982). Oxygen activation and oxygen toxicity. Annu. Rev. Plant Physiol. 33: 73–96. Engels C. (1994). Nutrient acquisition by plants and its limitations by low temperatures in maize. In, K Dörffling et al., (eds) Crop adaptation to cool climates. COST 814 workshop ECSP-EEC-EAEC, Brussels Engels C, Marschner H. (1990). Effects of sub-optimal root zone temperatures at varied nutrient supply and shoot meristem temperature on growth and nutrient concentrations in maize seedlings (Zea mays L.). Plant and Soil. 116: 215–225. Engels C, Marschner H. (1992). Root to shoot translocation of macronutrients in relation to shoot demand in maize (Zea mays L.) grown at different root zone temperatures. J plant Nutr Soil Sci. 155:121–128. Engels C, Marschner H. (1996a). Effects of suboptimal root zone temperatures and shoot demand on net translocation of micronutrients from the roots to the shoot of maize. Plant and Soil. 186:311-320. Engels C, Marschner H. (1996b). Effect of root zone temperature and shoot demand on N translocation from the roots to the shoot in maize supplied with nitrate or ammonium. Plant Physiol Biochem. 34:735–742. Engels C. (1993). Differences between maize and wheat in growth- related nutrient demand and uptake of potassium and phosphorus at suboptimal root-zone temperatures. Plant and Soil. 150, 129–138. Equiza MA, Mirave JP, Tognetti JA. (1997). Diჼ�erential root versus shoot growth inhibition and its relationship with carbohydrate accumulation at low temperature in diჼ�erent wheat cultivars. Annals of Botany 80: 657-663. Erbach DC, Cruse RM, Crosbie TM, Timmons DR, Kaspar TC, Potter KN. (1986). Maize response to tillage-induced soil conditions. Trans. ASAE. 29: 690–695. Brown PH, Bellaloui N, Wimmer MA, Bassil ES, Ruiz J, Hu H, Pfeffer H, Dannel F, Römheld V. (2002). Boron in plant biology. Plant Biol. 4:205–223.

! !!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!References!
! 150
Hou FF, Thseng FS. (1991). Studies on the flooding tolerance of soybean seed: varietal differences. Euphytica. Volume 57, Number 2 / September, Farooq M, Wahid A, Kadambot HM. (2012). Micronutrient application through ed treatments. A review. Journal of Soil Sci Plant Nutri.12: 125-142. Fausey NR, McDonald MB Jr. (1985). Emergence of inbred and hybrid corn following flooding. Agron. J. 77:51–56. Ficco DBM, Riefolo C, Nicastro G, De Simeone V, Di Gesu AM, Beleggioa R, Platani C, Cattivelli L, De Vita P. (2009). Phytate and mineral elements concentration in a collection of Italian durum wheat cultivars. Field Crops Research. 111:235–242. Fiedler RJ, Sander DH, Petersen GA. (1983). Relative performance of seed versus broadcast fertilizer on the yield of winter wheat. Agron Abstr . Am Soc Agron
Fincher GB. (1989). Molecular and cellular biology associated with endosperm mobilization in germinating cereal grains. Annu Rev Plant Physiol Plant Mol Biol. 40:305–346
Fleischer A, Titel C, Ehwald R. (1998). The boron requirement and cell wall properties of growing and stationary suspension-cultured Chenopodium album L. cells. Plant Physiol. 117: 1401-1410. Fleischer A, O’Neill MA, Ehwald R. (1999). The pore size of non-graminaceaous plant cell walls is rapidly decreased by borate ester cross-linking of the pectic polysaccharide rhamnogalacturonan II. Plant Physiol. 121:829–838. Flowers TJ. (2004). Improving crop salt tolerance. Journal of Experimental Botany. 55:307-319. Fortin MC, Pierce FJ. (1991). Timing and nature of mulch retardation of corn vegetative development. Agron J. 83:258–263. Foyer CH, Hélène VLD, Gomez JH. (2002). Review: Regulation of photosynthesis and antioxidant metabolism in maize leaves at optimal and chilling temperatures. Plant Physiology and Biochemistry. 40:659-568. Foyer CH, Halliwell B. (1976). The presence of glutathione and glutathione reductase in chloroplasts: A proposed role in ascorbic acid metabolism. Planta. 133: 21-25. Foyer CH, Halliwell B. (1977). Purification and properties of dehyroascorbate reductase from spinach leaves. Phytochemistry. 16: 1347-135. Fracheboud Y. (1999). Cold adaptation of the photosynthetic apparatus of maize by growth at suboptimal temperature. In: Sánchez-Díaz, M., Irigoyen, J.J., Aguirreolea, J., Pi- than, K. (eds) Crop Development for Cool and Wet Cli- mate of Europe, pp. 88-98. Office for Official Publications of the European Communities, Luxembourg.

! !!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!References!
! 151
Fracheboud Y, Jompuk C, Ribaut JM, Stamp P, Leipner J. (2004). Genetic analysis of cold tolerance of photosynthesis in maize. Plant Molecular Biology. 56: 241-253. Fridovich I. (1983). Superoxide radical: an endogenous toxicant. Annu. Rev. Pharmacol. Toxicol. 23: 239–257. Fukamatsu Y, Yabe N, Hasunuma K. (2003). Arabidopsis NDK1 is a component of ROS signaling by interacting with three catalases. Plant & Cell Physiology. 44: 982–989. Fukuyama K. (2004). Structure and function of plant ype ferredoxins. Photosynthesis Research. 81: 289–301. Gallardo K, Job C, Groot SPC, Puype M, Demol H, Vande- kerckove J, Job D. (2001). Proteomic analysis of Arabidopsis seed germination and priming. Plant Physiol.126: 835-848. Gomez-Cadenas A, Zentella R, Walker-Simmons MK, Ho TH. (2001). Gibberellin/abscisic acid antagonism in barley aleurone cells: Site of action of the protein kinase PKABA1 in relation to gibberellin signaling molecules. Plant Cell. 13: 667–679. Jacobsen JV, Chandler PM. (1987). Gibberellin and abscisic acid in germinating cereals. In PJ Davies, ed, Plant Hormones and Their Role in Plant Growth and Development. Martinus Nijhoff, Boston, pp 164–193. Genc Y, McDonald GK, Graham RD. (2000). Effect of seed zinc content on early growth of barley (Hordeum vulgare L.) under low and adequate soil zinc supply. Aust J Agric Res. 51:37–46. Gericke S, Kurmis B. (1952). Die kolorimetrische Phophorsäurebestimmung mit Ammonium-Vanadat-Molybdat und ihre Anwendung in der Pflanzenanalyse. Z Pflanzenernaehr Bodenkd. 59:235–247.
Ghassemi-Goleani K, Farshbaf-Jafari S, Shafagh-Kolvanagh J. (2011). Seed Priming and Field Performance of Soybean (Glycine max L.) in Response to Water Limitation. Not. Bot. Horti. Agrobo. 39(2): 186-189. Giauffre G, Bonhomme R, Derieux M. (1995). Genotypic differences for temperature response of leaf appearance rate and leaf elongation rate in field grown maize. Agronomie (Paris). 15: 123–137.
Giordano PM, Mortvedt JJ. (1978). Response of corn to Zn in ortho- and pyrophosphate fertilizers as affected by soil temperature and moisture. Agron J. 70:531-534. Gladish DK, Rost TL. (1993). The effects of temperature on primary root growth dynamics and lateral root distribution in garden pea (Pisum sativum L., cv. ‘Alaska’). Environ. Experim. Bot. 33: 243–258.

! !!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!References!
! 152
Goldbach HE. (1997). A critical review on current hypothesis concerning the role of boron in higher plants: suggestions for further research and methodological requirements. J. Trace & Microprobe Techniques .15: 51-91. Goldbach HE, Hartmann D, Rötzer T. (1990). Boron is required for the stimulation of the ferricyanide-induced proton release by auxins in suspension-cultured cells of Daucus carota and Lycopersicon esculentum. Physiol. Plant. 80: 114-118. Goldbach HE, Wimmer MA. (2007). Boron in plants and animals: is there a role beyond cell-wall structure? J. Plant Nutr. Soil Sci. 170:39–48. Gong X, Wang Y, Liu C, Wang S, Zhao X, Zhou M, Li N, Lu Y, Hong F. (2010). Effects of manganese deficiency on spectral characteristics and oxygen evolution in maize chloroplasts. Biol. Trace Elem. Res. 136: 372–382.
Graham RD, Ascher JS, Hynes SC. (1992). Selecting zinc-efficient genotypes for soils of low zinc status. Plant and Soil. 146: 241–250. Graham RD, Rengel Z. (1993). Genotypic variation in zinc uptake and utilization by plants, in: Robson A.D. (Ed.), Zinc in Soils and Plants, Kluwer Academic Publishers, Dordrecht, pp. 107–118.
Graham D, Patterson BD. (1982). Responses of plantsto low, non-freezing temperatures: Proteins, metabolism and acclimation. Annu. Rev. Plant Physiol. 33:347-372. Graham MY, Graham TL. (1991). Rapid accumulation of anionic peroxidase and phenolic polymers in soybean cotyledon tissues following treatment with Phytophthora megasperma f. sp. Glycinea woll glucan. Plant Physiol. 97:1445–1455. Granier C, Tardied F. (1998). Is thermal time adequate for expressing the effects of temperature on sunflower leaf development? Plant, Cell & Environ. 21: 695-703. Greaves JA. (1996). Improving suboptimal temperature tolerance in maize - the search for variation. J. Exper. Bot. 47: 307-323. Greenway H, Munns R. (1980). Mechanisms of salt tolerance in nonhalophytes. Annu. Rev. Plant Physiol. 31: 149–190. Greiner R. (2002). Purification and characterization of three phytate degrading enzymes from germinated lupin seeds (Lupinus albus var. Amiga). J Agric Food Chem. 50: 6858–6864. Greiner R, Egli I. (2003). Determination of the activity of acidic phytate-degrading enzymes in cereal seeds. J Agric Food Chem. 51:847–850. Greiner R, Muzquiz M, Burbano C, Cuadrado C, Pedrosa MM. (2001). Purification and characterization of a phytate degrading enzyme from germinated faba beans (Vicia faba

! !!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!References!
! 153
var. Alameda). J Agric Food Chem 49: 2234–2240. Grewal HS, Zhonggu L, Graham RD. (1997). Influence of subsoil zinc on dry matter production, seed yield and distribution of zinc in oilseed rape genotypes differing in zinc efficiency. Plant and Soil. 192: 181 – 189.
Grewal HS, Graham RD. (1994). Genetic variation in micronutrient uptake of wheat. In: Proceedings of the seventh Assembly Wheat Breeding Society of Australia. Eds. J Paull, IS Dundas, KJ Shepard and GJ Hollaniby. pp 23–26. Wheat Breeding Society of Australia, Adelaide.
Grundon NJ. (1987). Hungry Crops: A Guide to Nutrient Deficiencies in Field Crops. Queensland Department of Primary Industries, Brisbane. 242p. Guan Y, HU X, Wangi C, SHAO. (2009). Seed priming with chitosan improves maize germination and seedling growth in relation to physiological changes under low temperature stress. J Zhejiang Univ Sci B. 10:427-433. Gupta UC. (1993). Boron and its role in crop production. CRC Press, Inc. Boca Raton, FL., USA. 237p. Guy CL. (1990). Cold acclimation and freezing stress tolerance: Role of protein metabolism. Ann. Rev. Plant. Physiol. Mol. Biol. 41: 187-223. Magen M. (2004). Potential impact of crop diversification and biotechnological inventions on the use of micronutrients. IFA International Symposium on Micronutrients. 23-25 February 2004, New Delhi, India. Hacisalihoglu G, Kochian LV. (2003). How do some plants tolerate low levels of soil zinc? Mechansims of zinc efficiency in crop plants. New Phytol 159:341–350 Haldimann P. (1998). Low growth temperature induced changes to pigment composition and photosynthesis in Zea mays genotypes differing in chilling sensitivity. Plant, Cell and Environment 21: 200-208. Haldimann P, Fracheboud Y, Stamp P. (1995). Carotenoid composition in Zea mays developed at sub-optimal tem- perature and different light intensities. Physiologia Plantarum. 95: 409-414. Haldimann P, Fracheboud Y, Stamp P. (1996). Photosynthetic performance and resistance to photoinhibition of Zea mays L. leaves grown at suboptimal temperature. Plant, Cell and Environment. 19: 85-92. Halliwell B. (2009). The wanderings of a free radical. Free Radical Biol. Med. 46: 531–542. Halliwell B, Gutteridge JMC. (1986). Iron and free radical reactions: two aspects of

! !!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!References!
! 154
antioxidant protection. TIBS. 11: 372–375. Harris D, Joshi A, Khan PA, Gothkar P, Sodhi PS. (1999). On-farm seed priming in semi-arid agriculture: development and evaluation in maize rice and chickpea in India using participatory methods. Exp Agric. 35:15-29 Harris D, Rashid A, Miraj G, Arif M, Shah H. (2007). ‘On-farm’ seed priming with zinc sulphate solution—A cost-effective way to increase the maize yields of resource-poor farmers. Field Crops Research. 102:119–127. Harris D, Tripathi RS, Joshi A. (2000). On-farm' seed priming to improve crop establishment and yield in direct-seeded rice in IRRI: International Workshop on Dry-seeded Rice Technology. Bangkok 25-28 January, 2000. The International Rice Research Institute. Manila, The Philippines. Harris D, Raghuwanshi BS, Gangwar JS, Singh SC, Joshi KD, Rashid A. Hollington PA. (2001). Participatory evaluation by farmers of on-farm seed priming in Wheat in India, Nepal and Pakistan. Experimental Agriculture. 37: 403-415. Harris D. (1996). The effects of manure, genotype, seed priming, depth and date of sowing on the emergence and early growth of Sorghum bicolor (L.) Moench in semi-arid Botswana. Soil and Tillage Research. 40:73-88.
Harris D, Jones M. (1997). On-farm seed priming to accelerate germination in rainfed, dry seeded rice. International Rice Research Notes 22, 30
Hasegawa PM, Bressan RA, Zhu JK, Bohnert HJ. 2000. Plant cellular and molecular responses to high salinity. Annu Rev. Plant Physiol. Plant Mol. Biol. 51:463–499. Hedley MJ, Nye PH, White RE. (1982a). Plant-induced changes in the rhizosphere of rape (Brassica napus var. Emerald) seedlings. II. Origin of the pH change. New Phytol. 91:31–44. Hedley MJ, Stewart JWB, Chauhan BS. (1982b). Changes in inorganic and organic soil phosphorus fractions induced by cultivation practices and by laboratory incubations. Soil Sci. Soc. Am. J. 46:970–976. Hedrich R, Kurkdjian A. (1988). Characterization of an anion- permeable channel from sugar beet vacuoles: effect of inhibitors. EMBO Journal. 7: 3661–3666. Hendricks T, van Loon L. C. (1990). Petunia peroxidase is local- ized in the epidermis of aerial plant organs. J. Plant Physiol. 136:519–525. Hether NH, Olsen RA, Jackson LL. (1984). Chemical identification of iron reductants exuded by plant roots. J. Plant Nutr. 7:667–676. Hewitt EJ. (1966). Sand and Water Culture Methods Used in the Study of Plant Nutrition. 2nd edition. Technical Communication No. 22. Commonwealth Agricultural

! !!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!References!
! 155
Bureaux, Farnham Royal, England. pp. 547. Hil R, Scarisbrick R. (1951). The haematin compounds of leaves. New Phytologist. 50: 98. Hil R. (1954). The cytochrome b component of chloroplasts. Nature. 174: 501. Hildebrand DF. (1989). Lipoxygenases. Physiol. Plant. 76: 249–253. Hillel D. (1998). Environmental soil physics. Academic Press, San Diego Hillel M, Imas P. (2004). Potential Impact of Crop Diversification and Biotechnological Inventions on the Use of Micronutrients. IFA international symposium on micronutrients. 23-25 February 2004, New Delhi, India. Ogle HJ, DE Byth, McLean R. (1979). Effect of rust (Phakopsora pachyrhizi) on soybean yield and quality in south-eastern Queensland Australian Journal of Agricultural Research. 30: 883-889.
Hobbs PR, Obendorf RL. (1972). Interaction of initial seed moisture and inhibitional temperature on germination and productivity of soybean. Crop Sci 12:664–667.
Hodson MJ, Di Nola L, Mayer AM. (1987). The effect of changing temperatures during imbibition on ultrastructure in germinating pea embryonic radicles. J. exp. Bot. 38: 525–534. Hölzl G, Dörmann P. (2007). Structure and function of glycoglycerolipids in plants and bacteria. Prog. Lipid Res. 46: 225–243. Hope AB, Stevens PG. (1952). Electrical potential differences in bean roots on their relation to salt uptake. Aust. J. Sci. Res., Ser. B. 5: 335–343. Hopkins WG. (1995). Introduction to plant physiology. Wiley, New York. Hoppler M, Zeder C, Walczyk T. (2009). Quantification of ferritin-bound iron in plant samples by isotope tagging and species-specific isotope dilution mass spectrometry. Anal. Chem. 81: 7368–7372. Horvatic M, Balint L. (1996). Relationship among the phytic acid and protein content during maize grain maturation. J Agron Crop Sci. 176:73–77.
Hou FF, Thseng FS. (1991). Studies on the flooding tolerance of soybean seed: Varietal differences. Euphytica. 57:169–173.
Howarth CJ, Ougham HJ. (1993). “Gene expression under temperature stres”, New Phytol. 125:1-26.

! !!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!References!
! 156
Hu H, Penn SG, Lebrilla CB, Brown PH. (1997). Isolation and characterization of soluble boron complexes in higher plants: the mechanism of phloem mobility of boron. Plant Physiol. 113: 649-655. Hu YQ, Liu S, Yuan HM, Li J, Yan DW, Zhang JF, Lu YT. (2010). Functional comparison of catalase genes in the elimination of photorespiratory H2O2 using promoter- and 3′-untranslated region exchange experiments in the Arabidopsis cat2 photorespiratory mutant. Plant, Cell & Environment. 33: 1656–1670. Huang B, Taylor HM, McMichael BL. (1991). Effects of temperature on the development of metaxylem in primary wheat roots and its hydraulic consequences. Ann Bot 67: 63–166 Huang J, Reddman RE. (1995). Salt tolerance of Hordeum and Brassica species during germination and early seedling growth. Can. J. Plant Sci. 75:815–819. Huang, C., Barker, S. J., Langridge, P., Smith, F. W. and Graham, R. D. (2000). Zinc deficiency up-regulates expression of high-affinity phosphate transporter genes in both phosphate-sufficient and -defi- cient barley roots. Plant Physiol. 124, 415–422 Huber DM, Wilhelm NS. (1988). The role of manganese in resistance of plant diseases. In: Graham, R.D, R.J. Hannam, and N.C. Uren (eds). Manganese in Soils and Plants. Kluwer Academic Publishers, Dordrecht, The Netherlands. pp 155-173. Hughes NP, Williams RJP. (1988). An introduction to manganese biological chemistry In: Graham, R.D, R.J. Hannam, and N.C. Uren (eds). Manganese in Soils and Plants. Kluwer Academic Publishers, Dordrecht, The Netherlands. pp 155-173. Hund A, Richner W, Soldati A, Fracheboud Y, Stamp P. (2007). Root morphology and photosynthetic performance of maize inbred lines at low temperature, Eur J Agron. 27:52-61. Hund A. (2003). Genetic analysis of the response of maize (Zea mays L.) seedlings to long-term mild chilling stress: a morpho-physiological approach. ETH Zurich, Diss. ETH No. 15181, Zurich, Switzerland. Husted S, Laursen KH, Hebbern CA, Schmidt SB, Pedas P, Haldrup A, Jensen PE. (2010). Manganese deficiency leads to genotype-specific changes in fluorescence induction kinetics and state transitions. Plant Physiol. 150: 825–833. Ignaeva OS. (1969). Effect of presowing treatment of seeds with micronutrients on some physiological characteristics and yield of cereals grown in Buryat. Physiology and productivity of plants Zabaikal, e Ulan-Ude, 84-92. Imran M, V Römheld, G Neumann. 2013. Impact of nutrient seed priming on germination, seedling development, nutritional status and grain yield of maize. (In press: Journal of Plant Nutrition).

! !!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!References!
! 157
Imran M, A Mehmood, V Römheld, G Neumann. 2013. Nutrient seed priming improves seedling development and increases grain yield of maize exposed to low root zone temperatures during early growth. European Journal of Agronomy. 49: 141-148.
Imran M, V Römheld, G Neumann. 2013. Accumulation and distribution of Zn and Mn in soybean seeds after nutrient seed priming and its contribution to plant growth under Zn and Mn deficient conditions (Submitted: Journal of Plant Nutrition). Ishii T, Matsunaga T. (2001). Pectic polysaccharide rhamnogalacturonan II is covalently linked to homogalacturonan. Phytochemistry. 57: 969-974.
James AB. (1998). Effects of environment during growth on the sensitivity of leaf conductance to changes in humidity Global Change Biology Volume 4 Page 269 - March 1998
Janowiak F, Markowski A. (1994). Changes in leaf water relations and injuries in maize seedlings induced by different chilling conditions. Journal of Agronomy and Crop Science. 172: 19–28. Jean FC, Leopold AC. (1985). Ultra structural Aspects of Chilling Injury in the Soybean Seed Radicle. American Journal of Botany. 72:1120-1126. Jiang L, Phillips TE, Hamm CA, Drozdowicz YM, Rea PA, Maeshima M, Rogers SW, Rogers JC. (2001). The protein storage vacuole: a unique compound organelle. Journal of Cell Biology. 155:991-1002. Jin CW, You GY, He YF, Tang C, Wu P. Zheng SJ. (2007). Iron deficiency-induced secretion of phenolics facilitates the reutilization of root apoplastic iron in red clover. Plant Physiol. 144:278–285.
Job C, Rajjou L, Yoann L, Maya B, Dominique J. (2005). Patterns of Protein Oxidation in Arabidopsis Seeds and during Germination Plant Physiol. 138: 790–802.
Job C, Kersulec A, Ravasio L, Chareyre S, Pepin R, Job D. (1997). The solubilization of the basic subunits of sugarbeet seed 11-S globulin during priming and early germination. Seed Sci. Res. 7:225-243. John BC, Lersten NR. (1987). Reproductive morphology. In J.R.Wilcox(ed.)Soybean: Improvement, Production and Uses Madison. American Society of Agronomy. Wisconsin.U.S.A. Johnson IR, Thornley JHM. (1985). Temperature dependence of plant and crop processes. Ann Bot. 55:1–24. Johnson JR. Cobb BG, Drew MC. (1994). Hypoxic induction of anoxia tolerance in roots of Adh1 null Zea mays. Plant Physiology. 105: 61-67 Johnson AD, Simons JG. (1979). Diagnostic indices of zinc deficiency in tropical

! !!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!References!
! 158
legumes. J. Plant Nutr. 1:123–149. Johnson JG, Lauren JG, Welch RM, Duxbury JM. (2005). A comparison of the effects of micronutrient seed priming and soil fertilization on the mineral nutrition of chickpea (Cicer arietinum), Lentil (Lens culinaris), Rice (Oryza sativa) and wheat (Triticum aestivum) in Nepal. Expl Agric. 41:427–448. Jungk A. (1991). Dynamics of nutrient movement at the soil–root interface. In Plant Roots, The Hidden Half (J.Waisel, A. Eshel and U. Kafkafi, eds.), pp. 455–481. Marcel Dekker Inc., New York. Kanai R, Edwards GE. (1999). The biochemistry of C4 photosynthesis. In R Sage, R Monson, eds, C4 Plant Biology. Academic Press, San Diego, pp 49–87. Kanauchi M, Milet J, Bamforth CW. (2009). Oxalate and oxalate in malt. J. Inst. Brewing 115: 232–237. Kasper TC, Bland WI. (1992). Soil temperature and root growth. Soil science 154:290-299. Kaya Cengiz, David Higgs and Halil Kirnak. (2001). The effect of high salinity (NaCl) and supplementary phosphorus and potassium on physiology and nutrtion development of spinach. Bulg. J. Plant Physiol. 27: 47-59. Kedrov-Zikhman OK, Deeva VP. (1958). The effect of trace elements on the yield of sugarbeet and flax on weekly acid sub pudzolic soils. udobr. Urozk. 11: 36-39. Kendall AC, Keys AJ, Turner JC, Lea PJ, Miflin BJ. (1983). The isolation and characterisation of a catalase-deficient mutant of barley (Hordeum vulgare L.). Planta. 159:505–511. Khalid BY. Malik NSA. (1982). Presowing soaking of wheat seeds in copper and manganese solutions. Communications in Soil Science and Plant Analysis. 13:981–986. Kim TH, Bohmer M, Hu H, Nishimura N, Schroeder JI. (2010). Guard cell signal transduction network: advances in understanding abscisic acid, CO2, and Ca2 signaling. Annu Rev Plant Biol. 61: 561–591. Kingston-Smith AH, Harbinson J, Foyer CH. (1999). Acclimation of photosynthesis, H2O2 content and antioxidants in maize (Zea mays) grown at sub-optimal temperatures. Plant, Cell and Environment. 22: 1071-1083. Kitagishi K, Obata H. (1986). Effects of zinc deficiency on the nitrogen metabolism of meristematic tissues of rice plants with reference to protein synthesis. Soil Sci. Plant Nutr. 32: 397–405. Kitagishi K, Obata H, Kondo T. (1987). Effect of zinc deficiency on 80S ribosome content of meristematic tissues of rice plant. Soil Sci. Plant Nutr. 33: 423–430.

! !!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!References!
! 159
Klepper B. (1991). Root-shoot relationships. In Plant Roots: The Hidden Half (Y. Waisel, A. Eshel and U. Kafkafi, eds.), pp. 265–286. Marcel Dekker Inc., New York. Kobayashi M, Nakagawa H, Asaka T, Matoh T. (1999). Borate rhamnogalacturonan II bonding reinforced by Ca2+ retains pectic polysaccharides in higher plant cell walls. Plant Physiol. 119: 199-203. Kobayashi M, Matoh T, Azuma J. (1996). Two chain of rhamnogalacturonan II are cross-linked by boratediol ester bonds in higher plant cell walls. Plant Physiol. 110: 1017-1020. Kramer PJ, Boyer JS. (1995). Water relations of plants and soils. Academic Press, San Diego Kriedemann PE, Graham RD, Wiskich JT. (1985). Photosynthetic disfunction and in vivo changes in chlorophyll a fluorescence from manganese deficient wheat leaves. Aust. Agric. Res. 36: 157–169. Kropff MJ, van Laar HH. (1993). Modeling Crop-Weed Interactions. CAB International, UK. Kutík J, Holá D, Koová M, Rothová O, Haisel D, Wilhelmová N, Tichá I. (2004). Ultrastructure and dimen- sions of chloroplasts in leaves of three maize (Zea mays L.) inbred lines and their F1 hybrids grown under moderate chilling stress. Photosynthetica. 42: 447-455. Kwiatowsky J, Safianowska A, Kaniuga, Z. (1985). Isolation and characterization of an iron-containing superoxide dismutase from tomato leaves, Lycopersicon exculentum. Eur. J. Biochem. 146:459–466. Kolotelo D, Steenie EV, Peterson M, Bennett R, Trotter D, Dennis J. (2001). Seeds Handling Guidebook. - British Columbia. III. British Columbia. Tree Improvement Branch. Lin L, Ockenden I, Lott JNA. (2005). The concentrations and distribution of phytic acid-phosphorus and other mineral nutrients in wild-type and low phytic acid1-1 (lpa1-1) corn (Zea mays L.) grains and grain parts. Canadian Journal of Botany. 83: 131–141. Liu YQ, Bergervoet JHW, deVos CHR, Hilhorst HWM, Kraak HL, Karssen CM, Bino RJ. (1994). Nuclear replication activities during imbibition of abscisic acid and gibberellin-deficient tomato (Lycopersicon esculentum Mill.) seeds. Planta. 194: 368–373. Lönnerdal B. (2003). Genetically modified plants improved trace element nutrition. Journal of Nutrition.133: 1490–1493.

! !!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!References!
! 160
Laboure AM, Gagnon J, Lescure AM. (1993). Purification and characterization of a phytase (myo-Inositol-hexaki- sphosphate phosphohydrolase) accumulated in maize (Zea mays) seedlings during germination. Biochem J. 295:413–419. Ladror U, Dyck RL, Silbernagel MJ. (1986). Effects of oxygen and temperature during imbibition of seeds of two bean lines at two moisture levels. J. Am. Soc. Hortic. Sci. 111: 572–577. Lane BG. (1994) Oxalate, germin, and the extra cellular matrix of higher plants. FASEB J. 8: 294-301. Langan TD, Pendleton JW, Oplinger ES. (1986). Peroxide coated seed emergence in water-saturated soil. Agron. J. 78:769–772. Larbi A, Abadía A, Abadía J, Morales F. (2006). Down coregulation of light absorption, photochemistry, and carboxylation in Fe-deficient plants growing in different environments. Photosynth. Res. 89: 113–126. Larbi A, Abadía A, Morales F, Abadía J. (2004). Fe resupply to Fe-deficient sugar beet plants leads to rapid changes in the violaxanthin cycle and other photosynthetic characteristics without significant de novo chlorophyll synthesis. Photosynth. Res. 79:59–69. Larcher W, Bauer H. (1981). Ecological significance of resistance to low temperature. In: Lange OL, Nobel PS, Osmond CB, Ziegler H (eds.) Physiological plant ecology I. Responses to the physical environment. (Encyclopedia of plant physiology. new ser. Vol. 12A). pp. 403-428. Springer-Verlag, Berlin-Heidelberg. Lauchli A. (1984). Salt exclusion: an adaptation of legumes for crops and pastures under saline conditions. p. 171–187. In R.C. Staples and G.H. Toenniessen (ed.) Salinity tolerance in plants: strategies for crop improvement. John Wiley and Sons, New York. Lauchli A, Wieneke J. (1979). Studies on growth and distribution of Na1, K1 and Cl2 in soybean varieties differing in salt tolerance. Z. Pflanzenernahr Bodenked. 142:3–13. Lee EA, Staebler MA, Tollenaar M, (2002). Genetic variation in physiological discriminators for cold tolerance - early autotrophic phase of maize development. Crop Science 42: 1919–1929. Leipner J, Stamp P. (2009). Chilling stress in maize seedlings. In: Bennetzen, J.L., Hake, S.C. (eds) Handbook of Maize: Its Biology, pp. 291-310. Springer, Heidelberg. Lemanceau P, Bauer P, Kraemer S, Briat J F. (2009). Iron dynamics in the rhizosphere as a case study for analyzing interactions between soils, plants and microbes. Plant and Soil 321: 513–535. Levitt J. (1980). Responses of Plants to Environmental Stresses, 2nd. ed., Vol. 2. Academic Press, New York.

! !!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!References!
! 161
Lin CH, Stocking CR. (1978). Influence of leaf age, light, dark and iron deficiency on polyribosome levels in maize leaves. Plant Cell Physiol. 19: 461–470. Lindsay WL, Norvell WA. (1978). Development of a DTPA soil test for zinc, iron, manganese, and copper. Soil Sci. Soc. Amr. J. 42: 421–428. Loıc R, Manuel D, Karine G, Julie C, Julia B, Claudette J, Dominique J. (2012). Seed germination and vigor. Annu. Rev. Plant Biol. 63:507–33 Huang L, Ye L, Bell RW, and Dell B. (2005). Boron Nutrition and Chilling Tolerance of Warm Climate Crop Species Annals of Botany. 96: 755–767. Longnecker NE, Marcar NE, Graham RD. (1991). Increased manganese content of barley seeds can increase grain yield in manganese deficient conditions. Aust. J. Agric. Res. 42:1065-1074. Lönnerdal B. (2000). Dietary factors influencing zinc absorption. J. Nutr. 130:1378–1383. Lönnerdal B. (2002). Phytic acid trace element (Zn, Cu, Mn) interactions. J. Food Sci. Technol. 37: 749–758. Loomis WD, Durst RW. (1992). Review: chemistry and biology of boron. Biofactors. 3: 229-239. Lott, J. N. A. and Buttrose, M. S. (1978). Globoids in protein bodies of legume seed cotyledons. Aust. J. Plant Physiol. 5: 89–111. Lott, J. N. A., Bojarski, M., Kolasa, J., Batten, G. D. and Campbell, L. C. (2009). A review of the phosphorus content of dry cereal and legume crops of the world. Intern. J. Agric Res. Govern. Ecol. 8: 351–370. Lott JNA, Greenwood JS, Batten GD. (1995). Mechanisms and regulation of mineral nutrient storage during seed development. In. Seed Development and Germination (J. Kigel and G. Galili, eds.), pp. 215–235. Marcel Dekker, New York. Lowell WW, Kar LJT. (1981). Prevention of imbibitional injury in low vigor soybean embryonic axes by osmotic control of water uptake. Physiologia Plantarum. Vol.51 p 133. Lütke-Entrup N, Oehmichen J. (2000). Lehrbuch des Pflanzenbaus. Grundlagen Verlag Thomas Mann, Gelsenkirchen, Germany. Lynch, J. P. (2007). Roots of the second green revolution. Aust. J. Bot. 55: 493–512 Ascher-Ellis JS, Graham RD, Hollamby GJ, Paull J, Davies P, Huang C, Pallotta MA, Howes N, Khabaz-Saberi H, Jeffries SP, Moussavi-Nik M. (2001). Chapter 19.

! !!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!References!
! 162
Micronutrients. In Application of Physiology to Wheat Breeding. Reynolds, M.P., Ortiz-Monasterio, J.I. and McNab, A. (eds). CIMMYT, Mexico DF. pp. 219-240. Maas EV, Ogata G, Garber MJ. (1972). Influence of salinity on Fe, Mn and Zn uptake by plants. Agron. J. 64:793–795. Mahan JR, McMichael BL, Wanjura DF. (1995). Methods for reducing the adverse effects of temperature stress on plants: a review. Environ. Exp. Bot. 35: 251-258. Marcar NE, Graham RD. (1986). Effect of seed manganese content on the growth of wheat (Triticum aestivum) under manganese deficiency. Plant and Soil 96:165-173. Marentes E, Grusak MA. (1998). Iron transport and storage within the seed coat and embryo of developing seeds of pea (Pisum sativum L.) Seed Sci. Res. 8:367–375. Marschner H. (1995). Mineral nutrition of higher plants. London UK, Academic Press Marschner H. (2012). Mineral nutrition of higher plants. London UK, Academic Press Marschner H, Römheld V, Kissel M. (1987). Localization of phytosiderophore release and of iron uptake along intact barley roots. Physiol Plant. 71:157-162. Marschner H. (1993). Zinc uptake from soils. In Zinc in Soils and Plants (A. D. Robson, ed.), pp. 59–77. Kluwer Academic Publishers, Dordrecht, The Netherlands. Marsh HV, Evans HJ, Matrone G. (1965). Investigations of the Role of Iron in Chlorophyll Metabolism I. Effect of Iron Deficiency on Chlorophyll and Heme Content and on the Activities of Certain Enzymes in Leaves. Plant Physiol. 38:632–638. Martin WE, McLean JG, Quick J. (1965). Effect of temperature on the occurrence of phosphorus-induced zinc deficiency. Soil Sci Soc Amer Proc. 29: 411-413. Mathewes NIA, Boyer JS. (1984). Acclimation of photosynthesis to low leaf water potentials. Plant Physiol. 74:161-166. Matoh T, Kobayashi M. (1998). Boron and calcium, essential inorganic constituents of pectic polysaccharides in higher plant cell walls. J. Plant Res. 111: 179-190. Matoh T. (1997). Boron in plant cell walls. Plant and Soil. 193: 59–70. Mayer AM, Poljakoff MA. (1975). The germination of seeds. 3rd ed. Pergamon, New York Mazzolini AP, Pallaghy CK, Legge GJF. (1985). Quantitative micro analysis of Mn, Zn and other elements in mature wheat seed. New Phytol. 100: 483–509. McMichael BL, Burke JJ. (1996). Temperature effects on root growth. In Plant Roots: The Hidden Half. 2 nd. Edition. Eds. Y. Waisel, A. Eshel and U, Kafkafi. pp. 383-396. Marcel Dekker, New York.

! !!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!References!
! 163
McMichael BL, Burke JJ. (1998). Soil temperature and root growth. HortScience. 33: 947-951. Melkonian J, Yu LX, Setter TL. (2004). Chilling responses of maize (Zea mays L.) seedlings: root hydraulic conductance, abscisic acid, and stomatal conductance. Journal of Experimental Botany. 55: 1751-1760. Mengel K, Kirkby EA, (1987). Principles of Plant Nutrition. 4th (eds), International Potash Institute. Bern, Switzerland Miedema P, (1982). The effects of low temperature on Zea mays. Advances in Agronomy. 35:93-128 Miedema P, Post J, Groot PJ, (1987). The effects of low temperature on seedling growth of maize genotypes. Pudoc Wageningen, NL Miller GA, Youngs VL, Oplinger ES. (1980). Effect of available soil-phosphorus and environment on the phytic acid concentration in oats. Cereal Chem. 57:192–194. Milligan S, Dale J. (1988). The effects of root treatments on growth of the primary leaves of Phaseolus vulgaris L: general features. New Phytol. 108: 27–35. Miraj G. (2005). Effect of phosphorus and zinc priming on germination, seedling growth and yield of maize. M.Sc. (Hons.) Thesis. Dept. Agric. Chem., NWFP Agricultural University, Peshawar, Pakistan. Moiseev KA, Aleksandrova MI. (1960). The effect on silage and vegetable plants of pre-sowing treatment of their seeds with micronutrients. Trudy Komi Fil. Ahad. Nauk No. 9: 23-32. Monasterio JI, McNab A. (eds). CIMMYT, Mexico DF. pp. 219-240. Moore PA Jr., Patrick WH Jr. (1988). Effect of zinc deficiency on alcohol dehydrogenase activity and nutrient uptake in rice. Agron. J. 80: 882–885. Moraghan JT. (1985). Manganese deficiency in soybeans as affected by FeEDDHA and low soil temperature. Soil Sci. Soc. Am. J. 49: 1584-1586. Moraghan JT. (1987). Effects of phosphorus and iron fertilizers on the growth of two soybean varieties at two soil temperatures. Plant and Soil. 104: 121-127. Moraghan JT, Freemand TP, White D. (1986). Influence of FeEDDHA and soil temperature on the growth of two soybean varieties. Plant and Soil. 95: 574-579. Moraghan JT, Mascagni HJ. (1991). Environmental and soil factors affecting micronutrients deficiency and toxicity. In, JJ Mortvedt JJ, Cox FR, Shuman LM, Welch RM 2nd (eds) Micronutrients in Agriculture. Soil Sci Soc Amer Book Series No 4,

! !!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!References!
! 164
Madison Wisconsin, USA Morales F, Abadia A, Abadia J. (1991). Chlorophyll fluorescence and photon yield of oxygen evolution in iron–deficient sugar beet (Beta vulgaris L.) leaves. Plant Physiol. 97: 886–893. Morgan PW, Taylor DM, Joham HE. (1976). Manipulation of IAA-oxidase activity and auxine deficiency symptoms in intact cotton plants with manganese nutrition. Plant Physiol. 37: 149–156. Morgounov A, Gomez-Becerra HF, Abugalieva A, Dzhunusova M, Yessimbekova M, Muminjanov H, Zelenskiy Y, Ozturk L, Cakmak I. (2007). Iron and zinc grain density in common wheat grown in Central Asia. Euphytica. 155: 193–203. Mozafar AP, Schreiber, Oertli JJ. (1993). Photoperiod and root- zone temperature: Interacting effects on growth and mineral nutrients of maize. Plant and Soil. 153:71-78. Mühling KH, Wimmer M, Goldbach HE. (1998). Apoplastic and membrane-associated Ca2+ in leaves and roots as affected by boron deficiency. Physiol. Plant. 102: 179-184. Muldoon DK, Wheeler JL, Pearson CJ. (1984). Growth mineral composition and digestibility of maize sorghum and barnyard millets at different temperatures. Aus J Agric Res. 35:367-378. Munns R. Termaat A. (1986) Whole plant responses to salinity. Aus. J. Plant Physiol. 13, 143–160. Murungu FS, Chiduza C, Nyamugafata P, Clark LJ, Whalley WR, Finchsavage WR, Finchsavage WE. (2004). Effects of 'on-farm seed priming' on consecutive daily sowing occasions on the emergence and growth of maize in semi-arid Zimbabwe. Field crops res. 89: 49-57. Musakhandov AN. (1984). Effect of trace elements and methods of their application on yield of horse beans. Dokl. Ahad. SKh. Nauk No. 3: 26-88.
Dashti N, Zhang F, Hynes R, Smith DL. (1998). Plant growth promoting rhizobacteria accelerate nodulation and increase nitrogen fixation activity by field grown soybean (Glycine max (L.) Merr) under short season conditions, Plant and Soil. 200: 205-213.
Nable RO, Bar-Akiva A, Loneragan JF. (1984). Functional manganese requirement and its use as a critical value for diagnosis of manganese deficiency in subterranean clover (Trifolium subterraneum L. cv. Seaton Park). Ann. Bot. 54: 39–49. Nagasuga K, Murai-Hatano M, Kuwagata T. (2011). Effects of low root temperature on dry matter production and root water uptake in rice plants. Plant Production Science. 14:22-29 Neumann G, Römheld V. (2007). The release of root exudates as affected by the plant

! !!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!References!
! 165
physiological status. In: Pinton R, Varanini Z, Nannipieri Z (eds) The rhizosphere: biochem- istry and organic substances at the soil-plant interface, 2nd edn. CRC, Boca Raton, Florida, USA, pp 23–72 Neumann KH, Steward FC. (1968). Investigations on the growth and metabolism of cultured explants of Daucus carota. I. Effects of iron, molybdenum and manganese on growth. Planta. 81: 333–350. Nie GY, Baker NR. (1991). Modifications to thylakoid composition during development of maize leaves at low growth temperature. Plant Physiology. 95: 184-191. Nie GY, Long SP, Baker NR. (1992). The effects of development at sub-optimal growth temperatures on photosynthetic capacity and susceptibility to chilling-dependent photoinhibition in Zea mays. Physiologia Plantarum. 85: 554-560. Nie GY, Robertson EJ, Freyer MJ, Leech RM, Baker NR. (1995). Response of the photosynthetic appa- ratus in maize leaves grown at low temperature on trans- fer to normal growth temperature. Plant, Cell and Envi- ronment. 18: 1-12. Dogar, M. A. and van Hai, T. (1980). Effect of P, N and HCO32 levels in the nutrient solution on rate of Zn absorption by rice roots and Zn content in plants. Z. Pflanzenphysiol. 98, 203–212. Forno, D. A., Yoshida, S. and Asher, C. J. (1975). Zinc deficiency in rice. I. Soil factors associated with the deficiency. Plant Soil 42, 537–550. Nikos A, Gavalas, Clark HE. (1971). On the Role of Manganese in Photosynthesis. Plant Physiol. 47, 139-143. Nishio JN, Abadía J, Terry N. (1985). Chlorophyll-proteins and electron transport during iron nutrition mediated chloroplast development. Plant Physiol. 78: 296–299. Niu X, Bressan RA, Hasegawa PM, Pardo JM. (1995). Ion homeostasis in NaCl stress environments. Plant Physiol. 109: 735–742. O’’Neill MA, Warrenfeltz D, Kates K, Pellerin P, Doco T, Darvill AG, Albersheim P. (1996). Rhamnogalacturonan-II, a pectic polysaccharide in the walls of growing plant cell, forms a dimer that is covalently cross-linked by a borate ester: In vitro conditions for the formation and hydrolysis of the dimer. J. Biol. Chem. 271: 22923-22930. Obata H, Umebayashi M. (1988). Effect of zinc deficiency on protein synthesis in cultured tobacco plant cells. Soil Sci. Plant Nutr. 34: 351–357.
Ogle HJ, Byth DE, Mclean RJ. (1979). Effect of rust (Phakopsora pachyrhizi) on soybean yield and quality in South-eastern Queensland. Australian Journal of Agricultural Research. 30: 883-893.
Önnerud H, Zhang L, Gellerstedt G, Henriksson G. (2002). Polymerization of monolignols by redox shuttle-mediated enzymatic oxidation. Plant Cell. 14: 1953–1962.

! !!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!References!
! 166
Osborne S, Riedell W. (2006). Soybean Growth Response to Low Rates of Nitrogen Applied at Planting in the Northern Great Plains Journal of Plant Nutrition. 29: 985-1002.
Pahlavanian AM, SilkWK. (1988). Effect of temperature on spatial and temporal aspects of growth in the primary maize root. Plant Physiol87: 529-532. Parera CA, Cantliffe DJ. (1994). Presowing seed priming. Horticultural Reviews. 16:109-141. Park SH, Sung JK, Lee SY, Park JH, Lee JY, Jang BC, Lee BH, Kim TW. (2006). Early growth, carbohydrate, and phytic acid contents of germinating rice seeds under NaCl stress. Korean J Crop Sci. 51:137–141. Parr AJ, Loughman BC. (1983). Boron and membrane functions in plants. In Metals and Micronutrients: Uptake and Utilization by Plants (Ann. Proc. Phytochem. Soc. Eur. No. 21; D. A. Robb and W. S. Pierpoint, eds.), pp. 87–107. Academic Press, London. Paul MJ, Lawlor DW, Driscoll SP. (1990). The effect of temperature on photosynthesis and carbon fluxes in sunflower and rape. J. Exp. Bot. 41: 547-555. Pavel EW, Fereres E. (1998). Low soil temperatures induce water deficits in olive (Olea europaea) trees. Physiol. Plant. 104: 525-532. Peeran SN, Natanasabapathy S. (1980). Potassium chloride pretreatment on rice seeds. International Rice Research Newsletter 5:19. Penfield S. (2008). Temperature perception and signal transduction in plants. New Phytol. 179, 615–628. Perur NG, Smith RL, Wiebe HH. (1961). Effect of iron chlorosis on protein fraction on corn leaf tissue. Plant Physiol. 36: 736–739. Peterson CJ, Johnson VA, Mattern PJ. (1986). Influence of cultivar and environment on mineral and protein concentrations of wheat flour, bran and grain. Cereal Chem. 63: 118–186. Petruzzelli L, Melillo MT, Bleve ZT, Taranto T. (1992). Physiological and Ultrastructural Changes in Isolated Wheat Embryos During Salt and Osmotic Shock. Annals of Botany 69: 25-31. Pfeffer H, Dannel F, Römheld V. (1998). Are there connections between phenol metabolism, ascorbate metabolism and membrane integrity in leaves of boron-deficient sunflower plants? Physiol. Plant. 104: 479-485. Pietrini F, Iannelli MA, Battistelli A, Moscatello S, Loreto F, Massacci A. (1999). Effects on photosynthesis, carbohydrate accumulation and regrowth induced by temperature increase in maize genotypes with different sensitivity to low temperature.

! !!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!References!
! 167
Australian Journal of Plant Physiology 26: 367-373. Pietrzak LN, Fregeau-Reid J, Chatson B, Blackwell B. (2002). Observations on Water Distribution in Soybean Seed During Hydration Processes Using Nuclear Magnetic Resonance Imaging. Can. J. Plant Science. 82: 513-519 Pinton R, Cakmak I, Marschner H. (1994). Zinc deficiency enchanced NAD(P) H dependent superoxide radical production in plasma membrane vesicles isolated from roots of bean plants. J. Exp. Bot. 45: 45-50. Pollard AS, Parr AJ, Loughman BC. (1977). Boron in relation to membrane function in higher plants. J. Exp. Bot. 28: 831–841. Pollock BM, Vivian KT. (1996). Imbibition Period as the Critical Temperature Sensitive Stage in Germination of Lima Bean Seeds. Plant Physiol. 41: 221–229. Pollock BM. (1972). Effects of environment after sowing on viability. p. 150–171. In E.H. Roberts (ed.) Viability of seeds. Syracuse University Press, Syracuse, NY. Pollock CJ, Eagles CF. (1988). Low tem perature and the growth of plants. In Plants and Temperature: Proceedings of the Society for Experimental Biology 42, eds S. P. Long and F. I. Woodward, pp. 157-180. Company of Biologists, Cambridge.
Powell AA, Matthews S. (1978). The damaging effect of water on dry pea embryos during imbibition. Journal of Experimental Botany. 29: 1215–1229.
Pregitzer KS, King JS. (2005). Effects of soil temperature on nutrient uptake. In, BassiriRad H (eds) Nutrient acquisition by plants: an ecological perspective. Springer-Verlag, Berlin Heidelberg. Pregitzer KS, King JS, Burton AJ, Brown SE. (2000). Responses of tree fine roots to temperature. New Phytol. 147:105–115. Pritchard J, Barlow PW, Adam JS, Tomos AD. (1990). Biophysics of the inhibition of the growth of maize roots by lowered temperature. Plant Physiol. 93: 222–230. Pulschen L. Application of micronutrients: pros and cons of the different application strategies. Ifa international symposium on micronutrients. 23-25 february 2004, new delhi, India. Pushnik JC, Miller GW. (1989). Iron regulation of chloroplast photosynthetic function: mediation of PS I development. J. Plant Nutr. 12: 407–421. Pushnik JC, Miller GW, Manwaring JH. (1984). The role of iron in higher plant chlorophyll biosynthesis, maintenance and chloroplast biogenesis. J. Plant Nutr. 7: 733–758. Raboy V, Below FE, Dickinson DB. (1989). Alteration of maize kernel phytic acid

! !!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!References!
! 168
levels by recurrent selection for protein and oil. J Hered. 80:311–315. Raboy V, Gerbasi PF, Young KA, Stoneberg SD, Pickett SG, Bauman AT, Murthy PPN, Sheridan WF, Ertl DS. (2000). Origin and seed phenotype of maize low phytic acid 1-1 and low phytic acid 2-1. Plant Physiol. 124:355–368. Raboy V, Noaman MM, Taylor GA, Pickett SG. (1991). Grain phytic acid and protein are highly correlated in winter wheat. Crop Sci. 31:631–635. Raboy V, Young KA, Dorsch JA, Cook A. (2001). Genetics and breeding of seed phosphorus and phytic acid. J Plant Physiol. 158: 489–497. Ragus L.N. (1987). Role of water absorbing capacity in soybean germination and seedling vigour. Seed sci. technol. 15: 285-296 Ramón A M, Carpena-Ruiz R O and Gárate A. (1990). The effects of short-term deficiency of boron on potassium, calcium and magnesium distribution in leaves and roots of tomato (Lycopersicon esculentum) plants. In Plant Nutrition Physiology and Applications. Ed. M L van Beusichem. pp. 287–290. Kluwer Academic Publishers, Dordrecht. Ranieri A, Castagna A, Baldan B, Soldatini GF. (2001). Iron defiency differently affects peroxidase isoforms in sunflower. J. Exp. Bot. 52: 25–35. Rapacz M. (2002). Cold-deacclimation of oilseed rape (Brassica napus var. oleifera) in response to fluctuating temperatures and photoperiod. Ann. Bot. (Lond.) 89: 543-549. Rashid A, Harris D, Hollington PA and Khattak RA. (2002). On-farm seed priming: a key technology for improving the livelihoods of resource-poor farmers on saline lands. Pp 423-431 in: R. Ahmad and K.A. Malik (eds.) Prospects for Saline Agriculture_. Kluwer Academic Publishers. The Netherlands. Raven JA, Evans MCW, Korb RE. (1999). The role of trace metals in photosynthetic electron transport in O2-evolving organisms. Photosynth. Res. 60:111–149. Ravindran V, Ravindran G, Sivalogan S. (1994). Total and phytate phosphorus contents of various foods and feed stuffs of plant origin. Food Chem. 56:335–343. Rengel Z. (2001). Genotypic differences in micronutrient use efficiency in crops. Commun. Soil Sci. Plant Anal. 32: 1163–1186. Rengel Z, Graham RD. (1995). Importance of seed Zn content for wheat growth on Zn- deficient soil. II. Grain yield. Plant and Soil. 173:267-274. Rerkasem B, Bell RW, Lordkaew S, Loneragan JF. (1997). Relationship of seed boron concentration to germination and growth of soybean (Glycine max L. Merr.). Nutr. Cycl. Agroecosyst. 48: 217–223.

! !!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!References!
! 169
Rerkasem B, Bell RW, Loneragan JF. (1990). Effects of seed and soil boron on early seedling growth of black and green gram (Vigna mungo and V. radiata). In: van Beusichem ML (ed.) Plant nutrition - physiology and applications. pp. 281-285. Kluwer Acad. Publ., Dordrecht. Reuter DJ, Alston AM, McFarlane JD. (1988). occurance and correction of manganese deficiency in plants. In, ‘Manganese in soils and plants. Graham RD, Hannan RJ, Uren NC (eds) Kluwer Academic, Dordrecht, pp205-224. Richner WC, Kiel, Stamp P. (1997). Is seedling root morphology predictive of seasonal accumulation of shoot dry matter in maize? Crop Sci. 37:1237-1241. Riekels JW, Lingle JC. (1966). Iron uptake and translocation by tomato plants as influenced by root temperature and manganese nutrition. Plant Physiol. 41: 1095-1101. Ritchie SW, Hanway JJ, Benson GO. (1986). How a corn plant develops? Iowa State Univ, Coop Exten Ser Spec Rep 48: 1-21. Robert LT, Sherlie HW. (1985). Reversal of the Effects of Aging in Soybean Seeds' Plant Physiol.77: 584-586. Roberth MD, Francis HW. (1983). Plant Physiology. 4th Edition. Willard Grany Press, Boston p. 577. Roberts WO. (1948). Preservation of mineral deficiency by soaking in nutrient solution. J. Agric. Sci. 38: 458-468. Roberts WO. (1973). Prevention of mineral deficiency by soaking seed in nutrient solution. J Agric Sci. 38:458-468. Robertson EJ, Baker NR, Leech RM. (1993). Chlorophyll thylakoid protein changes induced by low growth tem- perature in maize revealed by immunocytology. Plant, Cell and Environment. 16: 809-818. Robertson GA, Loughman BC. (1974). Response to boron defi- ciency: a comparison with responses produced by chemical methods of retarding root elongation. New Phytol. 73: 821–832. Römheld V, Marschner H. (1981). Rhythmic iron stress reactions in sunflower at suboptimal iron supply. Physiol. Plant. 53: 347–353. Römheld V, Marschner H. (1991). Functions of micronutrients in plants. In Micronutrients in Agriculture, 2nd ed. (J. J. Mordvedt, F. R. Cox, L. M. Shuman and R. M. Welch, eds.), pp. 297–328. SSSA Book Series, No. 4, Madison, WI, USA. Ros C, Bell RW, White PF. (1997). Effect of seed phosphorus and soil phosphorus applications on early growth of rice (Oryza sativa L.) cv IR66. Soil Sci Plant Nutr. 43: 499-509.

! !!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!References!
! 170
Ros CR, Belland W, White PF. (2000). Phosphorous seed coating and soaking for improving seedling growth of Oryza sativa (rice) cv IR66. Seed Sci Tech. 28:391-401. Rowse HR. (1995): Drum priming: A non-osmotic method of priming seeds. Seed Sci. Techn. 24. 281-294. Rymen B, Fiorani F, Kartal F, Vandepoele K, Inzé D, Beemster GTS. (2007). Cold nights impair leaf growth and cell cycle progression in maize through transcriptional changes of cell cycle genes. Plant Physiology. 143: 1429- 1438. Raboy V. (2002). Progress in breeding low phytate crops. Journal of Nutrition. 132: 503–505. Steber CM, Cooney SE, McCourt P. (1998). Isolation of the GA- response mutant sly1 as a suppressor of ABI1–1 in Arabidopsis thaliana. Genetics. 149: 509–521. Sadana US, Kusum L, Claassen N. (2002). Manganese efficiency of wheat cultivars as related to root growth and internal manganese requirement. J. Plant Nutr. 25, 2677–2688. Sadana US, Sharma P, Castañeda ON, Samal D, Claassen N. (2005). Manganese uptake and Mn efficiency of wheat cultivars are related to Mn-uptake kinetics and root growth. J. Plant Nutr. Soil Sci. 168: 581–589. Salami AU, Kenefick DG. (1970). Stimulation of growth in zinc- deficient corn seedlings by the addition of tryptophan. Crop Sci. 10:291-294. Salisbury FB, Ross CW. (1992). Plant physiology. Wadsworth Publishing Company Inc, Belmont Sandmann G, Böger P. (1983). The enzymatological function of heavy metals and their role in electron transfer processes of plants. In Encyclopedia of Plant Physiology, New Series (A. Läuchli and R. L. Bieleski, eds) Vol. 15A, pp 563–593. Springer-Verlag, Berlin and New York. Sarkar AN, Wynjones RJ. (1982). Effect of rhizosphere pH on the availability and uptake of Fe, Mn and Zn Plant and Soil. 66: 361-372. Sattelmacher B, Marschner H. Kuehne R. (1990). Effects of the temperature of the rooting zone on the growth and development of roots of potato (Solanum tuberosum). Ann. Bot. 65: 27–36. Savithri PR, Perumal R, Nagarajan R. (1999). Soil and crop management technologis fro enhancing rice production under micronutrient constraits. Nutrient cycling in agroecosytem. 53:121-136.

! !!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!References!
! 171
Scaife MA, Smith R. (1973). The phosphorus requirement of lettuce. II. A dynamic model of phosphorus uptake and growth. J Agric Sci. 80:353–361
Schlemmer U, Frølich W, Prieto RM, Grases F. (2009). Phytate in foods and significance for humans: Food sources, intake, process- ing, bioavailability, protective role and analysis. Mol. Nutr. Food Res. 53: 330–375. Schon MK, Novacky A, Blevins DG. (1990). Boron induces hyper- polarisation of sunflower root cell membranes and increases membrane permeability to K+. Plant Physiology 93: 566–577. Schwartz SM, Welch RM, Grunes DL, Cary EE, Norvell WA, Gilbert MD, Meredith MP, Sanchirico CA. (1987). Effect of zinc, phosphorus and root-zone temperature on nutrient uptake by barley. Soil Sci Soc Am J. 51:371-375 Seckbach J. (1982). Ferreting out the secrets of plant ferritin – a review. J. Plant Nutr. 5: 369–394. Sekimoto H, Hoshi M, Nomura T, Yokota T. (1997). Zinc deficiency affects the levels of endogenous gibberellins in Zea mays L. Plant Cell Physiol. 38:1087–1090. Sevilla F, del Rio L A, Hellin E. (1984). Superoxide dismutases from a citrus plant: Presence of two iron-containing isoenzymes in leaves of lemon trees (Citrus limonum L.). J. Plant Physiol. 116: 381–387. Shannon LM. (1948). Plant isozymes.Annu.Rev.Plant.Physiol. 19:187-210. Sharma KC, Krantz BA, Brown AL, Quick J. (1968). Interactions of Zn and P with soil temperatures in rice. Agron J. 60:652-655.
Sharma CP, Sharma PN, Bisht SS, Nautiyal BD. (1982). Zinc deficiency induced changes in cabbage (Brassica oleracea var. capitata). In Plant Nutrition 1982. Proceedings of the Ninth International Plant Nutrition Colloquium. Ed. A. Scaife. pp. 601-606. Warwick, England.
Sharratt BS. (1991). Shoot growth, root length density, and water use of barley grown at different soil temperatures. Agron. J. 83: 237-239. Shenker M, Plessner OE, Tel OE. (2004). Manganese nutrition effects on tomato growth, chlorophyll concentration, and superoxide dismutase activity. J. Plant Physiol. 161: 197–202. Sherrell CG. (1984). Effect of molybdenum concentration in the seed on the response of pasture legumes to molybdenum. New Zealand Journal of Agricultural Research. 27:417–423. Shive JB Jr. Barnett NM. (1973). Boron deficiency effects on peroxidase, hydroxyproline, and boron in cell walls and cytoplasm of Helianthus annuus L. hypocotyls. Plant Cell Physiol. 14: 573-583.

! !!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!References!
! 172
Sijmons PC, Kolattukudy PE, Bienfait HF. (1985). Iron- deficiency decreases suberization in bean roots through a decrease in suberin-specific peroxidase activity. Plant Physiol. 78: 115–120. Silberbush M, Ben-Asher J. (2001). Simulation study of nutrient uptake by plants from soilless cultures as affected by salinity buildup and transpiration. Plant and Soil. 233: 59–69. Sillanpaa M. (1982). Micronutrients and the nutrient status of soils: a global study. FAO Soils Bulletin 48. Food and Agriculture Organization of the United Nations, Rome. Simpson DJ, Robinson SR. (1984). Freeze-fracture ultrastructure of thylakoid membranes in chloroplasts from manganese deficient plants. Plant Physiol. 74: 735–741. Singh BP. (1998). Soil environment and root growth: introduction to the colloquium. HortSci. 33: 946-7. Singh DK, Bharti S. (1985). Seed manganese content and its relationship with the growth characteristics of wheat cultivars. New Phytologist. 101:387-391. Skok J, McIlrath WJ. (1958). Distribution of boron in cells of dicotyledonous plants in relation to growth. Plant Physiol. 33: 428-431. Smalik M. (1959). The effect of trace elements on the yield and quality of flax and potatos in Northern Slovaki Pol noskospodarstov. 6: 29-44. Smith P, Dale J. (1988). The effect of root cooling and excision treatments on the growth of primary leaves of Phaseolus vulgaris L. Rapid and reversible increases in abscisic acid content. New Phytol. 110: 293–300. Snowball K, Robson AD. (1986). Symptoms of nutrient deficiencies: lupins. Nedlands: The University of Western Australia Press. Søren B, Henrik BP, Birgitte T, Preben BH. (2009). Iron transport, deposition and bioavailability in the wheat and barley grain. Plant and Soil. 325:15–24. Sousa SF, Lopes AB, Fernandes PA, Ramos MJ. (2009). The zinc proteome: a tale of stability and functionality. Dalton Transactions. 7946–7956. Sowinski P, Maleszewski S. (1989). Chilling-sensitivity in maize seedlings. I. Growth and functioning of shoots and roots. Acta Physiologiae Plantarum. 11: 165-171. Sowinski, P, Rudzinska LA, Adamczyk J, Kubica I, Fronk J. (2005). Recovery of maize seedling growth, development and photosynthetic efficiency after initial growth at low temperature. Journal of Plant Physi- ology. 162: 67-80. Sowinski P, Rudzinska LA, Kobus P. (2003). Changes in plasmodesmata frequency in

! !!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!References!
! 173
vascular bundles of maize seedling leaf induced by growth at sub-optimal temperatures in relation to photosynthesis and assimilate export. Environ. Exp. Bot. 50: 183- 196. Spiller SC, Kaufman LS, Thompson WF, Briggs WR. (1987). Specific mRNA and rRNA level in greening pea leaves during recov- ery from iron stress. Plant Physiol. 84: 409–414. Staehelin LA, Newcomb EH. (2000). Membrane structure and membranous organelles. In Biochemistry & Molecular Biology of Plants (B. B. Buchanan, W. Gruissem and R. L. Jones, eds.), pp. 2–50. American Society of Plant Physiologists, Rockville, MD. Stamp P. (1986). Chilling stress in maize. In, Dolstra O, Miedema P (eds) Breeding of silage maize. Eucarpia Proceeding, Wageningen, pp 43–50 Stamp P. (1984). Chilling tolerance of young plants demonstrated on the example of maize (Zea mays L.). Adv. Agron. Crop Sci. 7. Paul Parey, Berlin. Stanely HD, Genichi K, Tamara MH. (1983). Differential Leakage of Intracellular Substances from Imbibing Soybean Seeds. Plant Physiol. 72: 919-924.
Stewart J, Mc. Guinn. (1969). Chilling injury and changes in adenosine triphosphate of cotton seedlings. Plant Physiol. 44:605-608
Stone PJ, Sorensen IB, Jamieson PD. (1999). Effect of soil temperature on phenology, canopy development, biomass and yield of maize in a cool temperature climate. Field Crops Research. 63:169-178.
Strauss AJ, Krüger GHJ, Strasser RJ, Van Heerden PDR. (2007). The role of low soil temperature in the inhibition of growth and PSII function during dark chilling in soybean genotypes of contrasting tolerance. Physiol. Plantarum. 131: 89–105.
Sung JM. (1995). The effect of sub-optimal O2 on seedling emergence of soybean seeds of different seed size. Seed Sci. Technol. 23:807–814.
Fujita TK, Mae T, Matsumoto H, Mori S, Sekiya J. (eds.) Plant nutrition - for sustainable food production and environment. pp. 153-156. Kluwer Acad. Publ., Dordrecht. Tachibana, S. (1987). Effect of root temperature on the rate of water and nutrient absorption in cucumber cultivars and fig leaf gourd. J. Japan. Soc. Hort. Sci. 55: 461–467. Taishi U, Katsuyoshi S, Morio K, Takao U. (2000). Enhancement of salt tolerance in soybean with NaCl pretreatment. Physiologia Plantarum. 110: 59-66. Takkar PN, Walker CD. (1993). The distribution and correction of zinc deficiency, in: Robson A.D. (Ed.), Zinc in Soils and Plants, Kluwer Academic Publishers, Dordrecht, The Netherlands, pp. 151–165.

! !!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!References!
! 174
Tanada T. (1983). Localization of boron in membranes. J. Plant Nutr. 6: 743-749. Tanaka Y, Hibino T, Hayashi Y, Tanaka A, Kishitani S, Takabe T, Yokota S, Takabe T. (1999). Salt tolerance of transgenic rice overexpressing yeast mitochondrial Mn-SOD in chloroplasts. Plant and Science. 148:131–138. Teasdale RD, Richards DK. (19909. Boron deficiency in cultured pine cells. Quantitative studies of the interaction with Ca andMg. Plant Physiol. 93: 2071–2078. Tehrani MM, Malakouti MJ, Savaghebi GR, Lotfollahi M, Zieyan A, Balali MR, Bybordi A, Majadi A. (2003). Can the zinc fortified seed increase wheat grain yield in zinc deficient soils of Iran? In: 2nd International Congress of Plant Physiology, eds. M. C. Ghildiyal. New Delhi, India. Teixeira MG, Guerra JGM, Lopes de Almeida D, Araujo AP, Franco AA. (1999). Effect of seed phosphorus concentration on nodulation and growth of three common bean cultivars. J. Plant Nutr. 22: 1599–1611. Terry N. (1980). Limiting factors in photosynthesis. I. Use of iron stress to control photochemical capacity in vivo. Plant Physiol. 65: 114–120. Terry N, Abadia J. (1986). Function of iron in chloroplasts. J. Plant Nutr. 9: 609–646. Tester M, Davenport R. (2003). Na+ Tolerance and Na+ transport in higher plants. Ann. Bot. 91: 503–527. Tewari RK, Kumar P, Neetu Sharma PN. (2005). Signs of oxidative stress in the chlorotic leaves of iron starved plants. Plant and Science. 169: 1037–1045. Thompson IA, Huber DM. (2007). Manganese and plant disease. In: Datnoff, L.E., Elmer, W.H., Huber, D.M. (Eds.), Mineral Nutrition and Plant Disease. APS Press, St. Paul, MN Thomson CJ, Bolger TP. (1993). Effects of seed phosphorus concentration on the emergence and growth of subterranean clover (Trifolium subterraneum L.). Plant and Soil 156: 285-288. Timperio AM, D’Amici GM, Barta C, Loreto F, Zolla L. (2007). Proteomics pigment composition, and organization of thylakoid membranes in iron-deficiency spinach leaves. J. Exp. Bot. 13: 3695–3710. Tinker BT, Nye PH. (2000). Solute Movement in the Rhizosphere. Oxford University Press, Oxford, UK. Tollner EW, Hargrove WL, Langdale GW. (1984). Influence of conventional and no-till practices on soil physical properties in the southern Piedmont. J. Soil Water Conserv. 39: 73–76.

! !!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!References!
! 175
Torchia RA, Hirsh AM. (1982). Analysis of membrane fractions from boron-deficient and control sunflower root tips by (n, a) nuclear reaction. Plant Physiol. 69: 44-51. Tully RE, Musgrave ME, Leopold AC. (1981). The seed coatas a control of imbibitional chilling injury. Crop Sci. 21:312–317. Turbe-Omemaetxe I, Moran JF, Arrese-Igor C, Gogorcena Y, Klucas RV, Becana M. (1995). Activated oxygen and antioxidant defences in iron-deficient pea plants. Plant, Cell Environ. 18: 421–429. Urbano G, Lopez-Jurado M, Aranda P, Vidai-Valverde C, Tenorio E, Porres J. (2000). The role of phytic acid in legumes: antinutrient or beneficial function? Journal of Physiology and Biochemistry. 56:283-294. Vandenabeele S, Vanderauwera S, Vuylsteke M, Rombauts S, Langebartels C, Seidlitz HK, Zabeau M, Van Montagu M, Inzé D, Van Breusegem F. (2004). Catalase deficiency drastically affects gene expression induced by high light in Arabidopsis thaliana. The Plant Journal .39: 45–58. Verheul MJ, Picatto C, Stamp P. (1996). Growth and development of maize seedlings under chilling conditions in the field. European Journal of Agronomy. 5:31-43 Verslues PE, Batelli G, Grillo S, Agius F, Kim YS, Zhu J, Agarwal M, Katiyar-Agarwal S, Zhu JK. (2007). Interaction of SOS2 with nucleoside diphosphate kinase 2 and catalases reveals a point of connection between salt stress and H2O2 signaling in Arabidopsis thaliana. Molecular and Cellular Biology. 27: 7771–7780. Wall MJ, Quinn AJ, D’Cunha GB. (2008). Manganese (Mn2)- dependent storage stabilization of Rhodotorula glutinis phenylalanine ammonia-lyase activity. J. Agric Food. Chem. 56: 894–902. Wallis JG, Browse J. (2002). Mutants of Arabidopsis reveal many roles for membrane lipids. Prog. Lipid Res. 41: 254–278. Wan X, Zwiazek JJ, Lieffers VJ, Landhausser M. (2001). Hydraulic conductance in aspen (Populus tremuloides) seedlings exposed to low root temperatures. Tree Physiol. 21:691–696. Wang FZ, Wang QB, Kwon SY, Kwak SS, Su WA. (2005a). Enhanced drought tolerance of transgenic rice plants expressing a pea manganese superoxide dismutase. J. Plant Physiol. 162: 465–472. Wang MY, Siddiqi MY, Ruth TJ, Glass ADM. (1993). Ammonium uptake by rice roots. 2. Kinetics of NH4 N13 influx across the plasmalemma. Plant Physiol. 103: 1259–1267. Weast RC. (1978). CRC. handbook of chemistry and physics, 58th edn. CRC Press,

! !!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!References!
! 176
West Palm Beach. Welbaum GE, Tissaoui T, Bradford KJ. (1990). Water relations of seed development and germination in muskmelon (Cucumis melo L.). III. Sensitivity of germination to water potential and abscisic acid during development. Plant Physiol. 92:1029–1037. Welch RM. (1986). Effects of nutrient deficiencies on seed production and quality. In. Advances in Plant Nutrition (B. Tinker and A. Läuchli, eds.), pp. 205–247. Praeger Scientific, New York. Welch RM, Webb MJ, Loneragan JF. (1982). Zinc in mem- brane function and its role in phosphorus toxicity. In Proceedings of the Ninth Plant Nutrition Colloquium, Warwick, England (A. Scaife, ed.), pp. 710–715. Commonw. Agric. Bur., Farnham Royal, Bucks. Welch RM. (1993). Zinc concentrations and forms in plants for humans and animals. p. 183!195. In A.D. Robson (ed.) Zinc in Soils and Plants. Kluwer Academic Publishers, Dordrecht, Boston, London. Welch RM. (1995). Micronutrient nutrition of plants. Critical Reviews in Plant Science. 14: 49-82. Welti R, Li W, Li M, Sang Y, Biesiada H, Zhou HE, Rajashekar CB, Williams TD, Wang X. (2002). Profiling membrane lipids in plant stress responses. Role of phospholipase Da in freezing induced lipid changes in Arabidopsis. J. Biol. Chem. 277: 31994–32002. Westerman L, Roddick JG. (1981). Annual variation in sterol levels in leaves of Taraxacum officinale Weber. Plant Physiol. 68: 872–875.
Whalley RDB, Mckell CM, Green LR. (1966). Seedling vigor and the early nonphotosynthetic stage of seedling growth in grasses. Crop Sci. 6:147–150. White JG, Zasoski RJ. (1999) Mapping soil micronutrients. Field Crops Res. 60, 11- 26.
White PJ, Broadley MR, (2009). Biofortification of crops with seven mineral elements often lacking in human diets – iron, zinc, copper, calcium, magnesium, selenium and iodine. New Phytol. 182: 49–84. White PJ, Hammond JP. (2008). Phosphorus nutrition of terrestrial plants. In: The Ecophysiology of Plant-Phosphorus Interactions (P. J. White and J. P. Hammond, eds.), pp. 51–81. Springer, Dordrecht. Wilhelm NS, Graham RD, Rovira AD. (1988). Application of different sources of manganese sulphate decreases take-all (Gaeumannomyces graminis var. tritici) of wheat grown in a manganese deficienct soil. Australian Journal of Agricultural Research. 39:1–10.

! !!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!References!
! 177
Willekens H, Langebartels C, Tire C, Van Montagu M, Inzé D, Van Camp W. (1994). Differential expression of catalase genes in Nicotiana plumbaginifolia (L.). Proceedings of the National Academy of Sciences of the United States of America 91, 10450–10454. William J, Bramlage A, Leopold C, David JP. (1978). Chilling Stress to Soybeans during Imhibition. Plant Physiol. 61: 525–529. Woodstock LW, Taylorson RB. (1981).Soaking injury and its reversal with polyethylene glycol in relation to respiratory metabolism in high and low vigor soybean seeds. Physiol. Plant. 53:263–268. Wuebker EF, Mullen RE, Koehler K. (2001). Flooding and Temperature Effects on Soybean Germination. Crop Science. 41: 1857-1861. !Xinhua Y, Tony JV. (2003). Potassium Placement Effects on Yield and Seed Composition of No-Till Soybean Seeded in Alternate Row Widths. Agronomy Journal. 95:126-132. Yilmaz AH, Ekiz I, Gültekin B, Torun H, Barut K, Cakmak I. (1998). Effect of seed zinc content on grain yield and zinc concentration of wheat grown in zinc-deficient calcareous soils. J Plant Nutr. 21:2257-2264 Yoshida S, Uemura M. (1986). Lipid composition of plasma mem- branes and tonoplasts isolated from etiolated seedlings of mung bean (Vigna radiata L.). Plant Physiol. 82: 807–812. Yu Q, Osborne LD, Rengel Z. (1998). Micronutrient deficiency changes activities of superoxide dismutase and ascorbate- peroxidase in tobacco plants. J Plant Nutr. 21:1427–1437 Yu Q, Rengel Z. (1999). Micronutrient deficiency influences plant growth and activities of superoxide dismutases in narrow-leafed lupins. Ann. Bot. 83: 175–182. Yu Q, Osborne LD, Rengel Z. (1999a). Increased tolerance to Mn deficiency in transgenic tobacco over producing superoxide dismutase. Ann. Bot. 84, 543–547. Wada T, Lott JNA. (1997). Light and electron microscopy and energy dispersive X-ray microanalysis studies of globoids in protein bodies of embryo tissues and the aleurone layer of rice (Oryza sativa L.) grains. Canadian Journal of Botany. 75: 1137–1147. Zhou JR, Erdman JW. (1995). Phytic acid in health and disease. Critical Reviews in Food Science and Nutrition. 35: 495–508. Zhu YG, Smith SE, Smith FA. (2001). Zinc (Zn)-phosphorus (P) Interactions in Two Cultivars of Spring Wheat (Triticum aestivum L.) Diჼ�ering in P Uptake Efficiency. Annals of Botany 88: 941-945.

! !!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!References!
! 178
Zhang M, Nyborg M, Mcgill WB. (1990). Phosphorus concen- tration in barley (Hordeum vulgare L.) seed: Influence on seedling growth and dry matter production. Plant and Soil 122:79–83. Zhang F, Römheld V, Marschner H. (1991a). Release of zinc mobilizing root exudates in different plant species as affected by zinc nutritional status. J. Plant Nutr. 14: 675–686. Zhang F, Römheld V, Marschner V. (1991b). Diurnal rhythm of release of phytosiderophores and uptake rate of zinc in iron-deficient wheat. Soil Sci. Plant Nutr. 37: 671–678.
Zhang M, Nyborg M, McGill WB. (1998). Phosphorus imbibed by barley seeds: location within the seed and assimilation by seedlings. Seed Sci. Techn. 26: 325-332.
Zhao FJ, Su YH, Dunham SJ, Rakszegi M, Bedo Z, McGrath SP, Shewry PR. (2009). Variation in mineral micronutrient con- centrations in grain of wheat lines of diverse origin. J. Cereal Sci. 49: 290–295. Zhu YG, Smith SE. (2001). Seed phosphorus (P) content affects growth, and P uptake of wheat plants and their association with arbuscular mycorrhizal (AM) fungi. Plant and Soil. 231: 105–112.

179
Curriculum Vitae Name: Muhammad Imran Date of birth: 12th July, 1978. Residential Address: Muhammad Imran Snedkervej 5, 1th 4200, Slagelse, Danmark Nationality: Pakistani Phone No.: 0045 7153 6241 E mail : [email protected] Accademics:
Degree Board/university Year of passing Division
PhD Uni. of Hohenheim, Stuttgart, Germany 2013
M.Sc. (Tropen. Master)
Uni. of Hohenheim, Stuttgart, Germany 2006 1st
M.Sc.(Hons.) Soil Science
University of Agriculture, Faisalabad 2004 1st
B.Sc.(Hons.) Soil Science
University of Agriculture, Faisalabad 2001 1st
F.Sc. (Pre-medical) Govt. College Sargodha 1997 1st
Matriculation (Science group)
G.H.S. Chak No. 104 NB, Sargodha 1994 1st

180
Phd Thesis Title: Nutrient seed priming improves early seedling establishment, plant growth and
nutritional status in Zea mays L. and Glycine max L. under abiotic stress
conditions
M.Sc. Thesis Title: Nutrient Seed Priming Improves Germination of Low Quality Soybean Seeds
and Induces Tolerance to Low Temperature Submergence and Salt Stress.
(2006, Hohenheim, Germany)
List of Publications:
Imran M, V Römheld, G Neumann. 2013. Impact of nutrient seed priming on germination, seedling development, nutritional status and grain yield of maize. (In press: Journal of Plant Nutrition).
Imran M, A Mehmood, V Römheld, G Neumann. 2013. Nutrient seed priming improves seedling development and increases grain yield of maize exposed to low root zone temperatures during early growth. European Journal of Agronomy. 49: 141-148.
Imran M, V Römheld, G Neumann. 2013. Accumulation and distribution of Zn and Mn in soybean seeds after nutrient seed priming and its contribution to plant growth under Zn and Mn deficient conditions (Submitted: Journal of Plant Nutrition).
Imran, Rahmatullah, M. A. Gill and Tariq Aziz.2004. Differential phosphorus mobilization by two wheat genotypes on a calcareous soil fertilized with ammonium sulphate, potassium nitrate, ammonium nitrate and urea. Pak. J. Soil Sci. Vol. 22(4) 64-67.
Hafiza salma kousar, M. A. Gill, Rahmatullah, Tariq Aziz, M. S. Akhtar and M. Imran. Solublization of tri-calcium phosphate by different wheat genotype. 2002. Pak. J. Agric. Sci. 39(4); 273-277.

181
Tahir, M. A., Rahmatullah, M. A. Gill, Tariq Aziz and M. Imran. 2003. Response of wheat and oat crops to potassium application and artificial irrigation with canal water. Pak. J. Agric. Sci. 40(3-4): 114-118.
Oral presentations
M. Imran. Nutrient seed priming improves low temperature stress tolerance in maize. International Plant Nutrition Conference. 2013. Istanbul. Turkey. M. Imran. Seed fertification: an approach to improve low temperature stress tolerance in maize. DanSeed symposium 2013. Denmark.
M. Imran. Seed priming with Fe and micronutrients improves seedling development and increases grain yield of maize exposed to low root zone temperatures during early growth. 16th International Symposium on Iron Nutrition and Interactions in Plants, June 17-20. Massachusetts. USA
Poster presentations:
Imran M., Asim M., Römheld V., Neumann G. Seed priming with micronutrients improves seedling development and increases grain yield of maize exposed to low root zone temperatures during early growth. ISRR 2012. “Roots to the Future”. 26-29 June. Dundee. Scotland.
Imran M., T. Müller, V. Römheld and G. Neumann. Nutrient seed priming enhances soil productivity under abiotic stress conditions. International Conference on Soil Fertility and Productivity.17-20 March 2010. Berlin, Germany.
Imran M., G. Neumann and V. Römheld. Nutrient seed priming improves germination rate and seedling growth under submergence stress at low temperature. Tropentag 2008. October 7-9. Uni. Hohenheim. Stuttgart.
Nora B., D. Sebastian, M. Imran, M. Weinmann, G. Neumann, T. Muller and V. Römheld. Effectiveness of commercial bio-fertilizers for improves phosphorus acquisition: Use of rapid screening test. Tropentag 2008. October 7-9. Uni. Hohenheim. Stuttgart.
Imran M., Rahmatullah, M.A. Gill and T. Aziz. 2004. Ammonium, nitrate and urea-N fertilizers influence P mobilization by wheat genotypes on a

182
calcareous soil. (Abst. 75-76). 10th Ints. Cong. Soil Sci., Soc. Pak., March 16-18, 2004, Sindh Agriculture University Tandojam.
Akhtar, M. S., M. A. Gill, T. Aziz, Rahmatullah, and M. Imran. 2002. Phosphorus nutrition of Brassica species /genotypes supplied with deficient and adequate P levels. (Abst. 40, Plant Sciences) 33rd All Pakistan Science Conference, 26-28 December, Uni. of Agric, Faisalabad, Pakistan.
Research projects:
1. Supervised master thesis of 3 M.Sc. students during my PhD studies in University of Hohenheim.
2. Work as a Research Assistant for two years in a project for the measurement of NO3 leaching in the ground water as a consequence of high input of N fertilizers in Germany.
3. Effect of different bio-fertilizers to control Take-all disease of wheat. 4. Effect of Boron seed priming on germination and growth of wheat under
salinity stress. 5. Effect of different biofertilizers on maize growth at low root zone
temperature. Academic supervisors: PD. Dr. Gunter Neumann Prof. Dr. Volker Römheld University of Hohenheim University of Hohenheim Stuttgart, Germany. Stuttgart, Germany. [email protected] [email protected] Prof. Dr. Torsten Müller University of Hohenheim Stuttgart, Germany. [email protected]

! 183!
Acknowledgements.. "In the name of God, most Gracious, most Compassionate"
I would like to express my sincere gratitude to my supervisor Prof.. Dr.. Günter.
Neumann. for his continues support, guidance and also for his vast knowledge that
helped me to reach this milestone. I am also very grateful to Prof.. Dr.. Volker.
Römheld.for his worthy ideas, guidance, motivation and enthusiasm that helped me a
lot to accomplish the objectives of my Ph.D research work. I have no hesitation to say
that both of them are wonderful persons.
I would like to extend my sincere regards to my thesis committee members Prof..Dr..
Michael.Kruse.(Institute of Seed Science Technology) and Prof..Dr..Uwe.Ludewig.
(Institute of Crop Science) for their kind guidance and for giving valuable advice.
My sincere thanks also go to the technical staff in the institute Maria.Ruckwied,.
Helene. Ochott,. Heide. Zimmermann,. Charlotte. Haake,. Hinrich. Bremer. for
their help and guidance, and especially to Hans.Buchar,.who always stood with me
in the fields. I will not forget Mrs. Christa. Shöllhammer. and Mrs. Ursla.
Berghammer.for their help to solve administrative problems in Hohenheim.
Words do not count easy to pay heartiest thanks to my uncle Muhammad.Amin.
Salim. and his family, who always encouraged and support me during my stay in
Germany. I am grateful to my brothers Adnan,.Rizwan,.Irfan.and.Naveed.and my
friends Rai Mukkaram Ali Tahir, Shoaib.Nasir.and Muhammad.Ajmal.for their
love, affection, prayers for my success and consistent encouragement. I am also
thankful to all my Pakistani friends in Pakistan, Stuttgart and to all my fellows in the
lab, office and coffee room who always wished me for success. The sweet memories
of my stay at Hohenheim will always be with me.
My wife, Fariha.Tanwir, whose love and companionship has boosted me at every
step during my PhD. Her endless love and indispensable help have made all this
possible. I must say thanks to Baasil Imran, my son, whose smile is always
energizing me to continue-
.
.
Muhammad.Imran.
!





























