Embed Size (px)
Citation preview

This article was downloaded by: [University of Illinois at Urbana-Champaign]On: 07 October 2014, At: 14:01Publisher: Taylor & FrancisInforma Ltd Registered in England and Wales Registered Number: 1072954 Registered office: Mortimer House,37-41 Mortimer Street, London W1T 3JH, UK
Scandinavian Journal of Forest ResearchPublication details, including instructions for authors and subscription information:http://www.tandfonline.com/loi/sfor20
Nutrient dynamics in some decomposing leaf andneedle litter types in a pinus sylvestris forestBjörn Berg a & Jordi Cortina b ca Department of Forest Soils , Swedish University of Agricultural Sciences , P.O. Box 7001,Uppsala, S‐750 07, Swedenb Department Biologia Vegetal, Facultat de Biologia , Universität de Barcelona , Avgda.Diagonal 645, Barcelona, Spainc Dept. Ecologia , Univ. d'Alacant , Ap. 99, Alacant, E‐03080, SpainPublished online: 10 Dec 2008.
To cite this article: Björn Berg & Jordi Cortina (1995) Nutrient dynamics in some decomposing leaf and needle litter types in apinus sylvestris forest, Scandinavian Journal of Forest Research, 10:1-4, 1-11
To link to this article: http://dx.doi.org/10.1080/02827589509382860
PLEASE SCROLL DOWN FOR ARTICLE
Taylor & Francis makes every effort to ensure the accuracy of all the information (the “Content”) containedin the publications on our platform. However, Taylor & Francis, our agents, and our licensors make norepresentations or warranties whatsoever as to the accuracy, completeness, or suitability for any purpose of theContent. Any opinions and views expressed in this publication are the opinions and views of the authors, andare not the views of or endorsed by Taylor & Francis. The accuracy of the Content should not be relied upon andshould be independently verified with primary sources of information. Taylor and Francis shall not be liable forany losses, actions, claims, proceedings, demands, costs, expenses, damages, and other liabilities whatsoeveror howsoever caused arising directly or indirectly in connection with, in relation to or arising out of the use ofthe Content.
This article may be used for research, teaching, and private study purposes. Any substantial or systematicreproduction, redistribution, reselling, loan, sub-licensing, systematic supply, or distribution in anyform to anyone is expressly forbidden. Terms & Conditions of access and use can be found at http://www.tandfonline.com/page/terms-and-conditions

Scand. J. For. Res. 10: 1-11, 1995
Nutrient Dynamics in Some Decomposing Leaf and Needle LitterTypes in a Pinus sylvestris Forest
BJÖRN BERG1 and JORDI CORTINA2
1Department of Forest Soils, P.O. Box 7001, Swedish University of Agricultural Sciences, S-750 07 Uppsala, Sweden,2Department Biologia Vegetal, Facultat de Biologia, Universität de Barcelona, Avgda. Diagonal 645; Barcelona, Spain,Present address: Dept. Ecologia, Univ. d'Alacant, Ap. 99, E-03080, Alacant, Spain
Scandinavian Journalof Forest Research
Berg, B.1 and Cortina, J. (1Department of Forest Soils, P.O. Box 7001, Swedish University ofAgricultural Sciences, S-750 07 Uppsala, Sweden, and 2Departament Biologia Vegetal, Facul-tat de Biologia, Universitat de Barcelona, Avgda. Diagonal 645, Barcelona, Spain). Nutrientdynamics in some decomposing leaf and needle litter types in a Pinus sylvestris forest. RecievedNov. 8, 1993. Accepted Aug. 16, 1994. Scand. J. For. Res. 10: 1-11, 1995.
We investigated the long-term net dynamics of N, P, S, K, Ca, Mg and Mn in decomposinglitter. Green and brown needles of Scots pine (P. sylvestris L.) lodgepole pine (Pinus contorta),as well as green and brown leaves of white birch (Betula pubescens) and leaves of grey alder(Alnus incana) were incubated for up to 4 years in a 130-year-old Scots pine forest. Theconcentrations of N and S increased linearly with accumulated mass loss, also P concentra-tions increased linearly after an initial leaching. Moreover, the rate of increase in N concentra-tions appeared to be proportional to the initial concentration. After the increase, N and Pconcentrations reached a phase in which they remained constant or declined slightly. Concen-trations of Mg and K decreased with accumulated mass loss, especially in litter types with highinitial concentrations of these elements. For Mn the pattern was similar but less pronounced.The relationship between Ca concentrations and accumulated mass loss was largely negative,resembling a negative X2 function. Absolute amounts of nutrients decreased linearly with littermass loss once a net release had started. This applied to all litter types and nutrients with theexception of Mn release from grey alder leaves. Key words: Decomposition, grey alder, leaves,litter, lodgepole pine, needles, nutrients, Scots pine, white birch.
INTRODUCTION
Boreal coniferous forests normally grow on acid,coarse and nutrient-poor soils of low fertility. It haslong been assumed that a certain admixture of birch(Betula spp.) in such habitats has a favourable influ-ence on soil conditions (Tamm 1930, Mork 1942,Nykvist 1963, Gardiner 1968) since the birch compo-nent tends to increase concentrations of, for example,major plant nutrients (Viro 1955, Bonnevie-Svendseh& Gjems 1957, Berg & Wessen 1984) in leaf litter.
In a study of litter decomposition in a nutrient-poor Scots pine forest in central Sweden, Berg &Wessen (1984) found that leaf litter of white birch(Betula pubescens) initially decomposed much fasterthan Scots pine needle litter. After the first year,however, the rate of birch litter decomposition fellbelow that of the Scots pine needles, and over athree-year period the birch Utter had a lower accumu-lated mass loss which was ascribed to slow lignindecomposition (Berg & Wessen 1984).
Another broad-leaved species whose soil-enhancingproperties have been evaluated is grey alder (Alnusincana). This N2-fixing species is fairly common in
coniferous forests, along forest edges and in wetsurroundings. A related species, red alder (Alnus ru-bra), can strongly increase the amount of N stored insoil over a relatively short period (Van Cleve et al.1971). Alnus spp., as well as other N2-fixing species,may promote an increase in C and N content inforest soils (Johnson 1992).
LuUgepole pine (Pinus contorta) generally domi-nates nutrient-poor sites throughout its original geo-graphic distribution (Weetman et al. 1988). Thisspecies has been reported to produce needle litterricher in, P, Mg, Mn and lignin (Berg & Lündmark1987, Berg et al. 1995) compared with that of Scotspine.
In a previous paper, Berg & Ekbohm (1991)showed clear differences in the dynamics of accumu-lated mass loss among seven types of leaves andneedles, and these differences appeared to be relatedto initial N concentrations. The nutrient dynamics ofthe very same litter are described in the present paper.Our aim was to desribe the concentration dynamics.as well as changes in absolute amounts of N, P, S, K,Ca, Mg and Mn in ths incubated brown and green
© 1995 Scandinavian University Press ISSN 0282-7581
Dow
nloa
ded
by [
Uni
vers
ity o
f Il
linoi
s at
Urb
ana-
Cha
mpa
ign]
at 1
4:01
07
Oct
ober
201
4

\
B. Berg and J. Cortina Scand. J. For. Res. 10 (1995)
needles of Scots pine and lodgepole pine, thebrown and green leaves of white birch, and thebrown leaves of grey alder. We wanted the answersto these questions: (i) is there any relation betweeninitial and final concentrations of nutrients in thedecomposing litter, and (ii) how are the releaserates dependent on the concentrations of nutrientsonce the release has started? The relationship betweenchemical properties of the far-decomposed litterand the characteristics of the humus to be formedwill be discussed elsewhere (cf. Laskowski et al.199X).
MATERIALS AND METHODS
Site description
The 130-year-old Scots pine (P. sylvestris L.) stand,where the incubations were made (in 1982), is locatedat the Ivantjärnsheden research site, used previouslyin connection with the Swedish Coniferous ForestProject. The site is situated in central Sweden(60°49' N; 16°30' E), at an altitude of 185 m on a flatarea of deep glacifluvial sand sediments. The meanannual precipitation at a nearby village is 609 mm,and the mean annual temperature is +3.8°C (1931-1960). The length of the growing season (days witha daily temperature higher than 6°C) is about 160days. The tree layer is exclusively composed of 17to 19 m-high Scots pine and the stand density is 393trees ha"1. Common heather (Calluna vulgaris) andcowberry (Vaccinium vitis-idaea) form a well-devel-oped understorey. The bottom layer, completelycovering the ground, is composed mainly of themosses Pleurozium schreberi and Dicranum rugosum,together with Cladonia spp. lichens. The most recentsilvicultural practice was a slight thinning in 1960.A more complete site description is given by Axelsson& Bråkenhielm (1980). The soil profile is an orthichumoferric podzol (iron podzol) with a weakly devel-oped Ae horizon (bleached horizon, 2-7 cm). Thehumus form is a typical mor. A very loose L horizon,interwoven with living mosses and lichens, coversan F/H horizon of 5-10 cm thickness. The F andH horizons (A0 , -A0 2) are almost indistinguishablefrom each other. The pH ranges from 3.9 to 4.2 in theF/H horizon and 4.6 to 4.8 in the upper mineral soil.The parent material, as well as the whole soil, isconsidered to be very poor in essential nutrients.The C-to-N ratio of the F/H horizon (Ao,-Ao2) is42.3.
Decomposition experiment
Scots pine needle litter was sampled at Ivantjärnshe-den in autumn 1981 from branches of trees about 15years old. Brown needles from the falling needlegeneration were taken at abscission from trees grow-ing in an area of about 20 m x 50 m, and stored at—20°C prior to sample preparation. Green need.eswere sampled in the same area, from the 2nd and 3rd(C + 2 and C + 3) needle generations. Brown leavesof white birch (Betula pubescens Ehrh.) and those ofgrey alder (Alnus incana) were sampled at the samesite as Scots pine needles and at the same time. Greenleaves of white birch were sampled at the same site inearly August. Brown needle litter and green needlesfrom lodgepole pine (Pinus contorta) were sampled ina similar way from a research site investigated earlier(cf. Berg & Lundmark 1987).
Before weighing and inclusion in litterbags, needlesand leaves were air-dried at room temperature toabout 5-8% moisture. Exact dry mass was deter-mined at 85°C, and the largest difference in moisturecontent was less than +0.5% units of the average{n = 28). The moisture content was: alder leaves7.1%; green leaves of birch 6.66%; brown leaves ofbirch 8.88%; green needles of lodgepole pine 6.62%;brown needles of lodgepole pine 8.61%; green needlesof Scots pine 5.48%; and for brown needles of Scotspine 6.44%.
The litterbags, made of terylene net with a meshsize of about 1 mm, measured 8 cm x 8 cm. Anamount of 0.6-1.0 g of needle and leaf litter wasenclosed in each bag. The bags were placed on thelitter (L) layer in each measurement plot (1 m x 1 m),according to a randomized block design comprising25 blocks. The whole Scots pine stand measured 2 ha.The litterbags were fastened to the ground with 10-15 cm-long metal pegs. The incubations star+ed inOctober 1982, and samplings were made three timesannually. On each sampling occasion, one samplefrom each of the 25 plots was collected. Five sampleswere used for determination of fungal biömass (Berg1991) and 20 for determination of mass loss, ligninand solubles (Berg & Ekbohm 1991), as well asnutrients. In this type of ecosystem, in which almostall decomposition is microbial, the litterbag clearlyallowed the decomposing organisms free access.
After being collected, the 20 samples of each typewere transported directly to the laboratory and storedin a deep freeze prior to handling. Plant remains,such as mosses, lichens and cowberry, were removed,and the loss of dry mass was determined by drying
2
Dow
nloa
ded
by [
Uni
vers
ity o
f Il
linoi
s at
Urb
ana-
Cha
mpa
ign]
at 1
4:01
07
Oct
ober
201
4

Scand. J. For. Res. 10 (1995) Nutrients in decomposing litter
the samples to a constant mass at 85°C. Mean valuesof mass loss were calculated for each sampling.Thereafter, the samples were pooled into a bulksample for chemical analyses. Thus one analysis wasmade on each bulk sample.
The composite samples were ground in a labora-tory mill equipped with a filter allowing particles lessthan 1 mm diameter to pass through. Nitrogen wasdetermined by a micro-Kjeldahl method using flowinjection analysis (Bifok FAI 05, Tecator, Höganäs,Sweden) with gas diffusion, and phenol red as anindicator (Svensson & Anfält 1982). For analysis ofP, S, Mg, K, Ca and Mn samples were digested for 2days in a 2.5 : 1 (v/v) mixture of nitric and per-chloricacid. The analyses were performed by plasma atomicemission spectrometry, ICP-AES (InstrumentationLaboratory IL P-200, Andover, Massachusetts,USA). Ash content was determined by heating sub-samples to 600°C for 2 h. All basic data are presentedin a report by Berg et al. (1991).
RESULTS AND DISCUSSION
The variation in initial nutrient composition wasfairly large. Nitrogen concentrations ranged from30.7 mgg"1 in alder leaves, to about 3.9 mgg"1 inbrown lodgepole pine needles. Brown leaves of whitebirch were richer in both N (7.7 mgg"1) and P(1.05 mgg"1) than brown needles of both pine spe-cies (4-5 mg g"' and 0.3 mg g~', respectively). Alderleaves had comparatively high concentrations of P(1.4mgg"1), and were also comparatively rich in K(15.6 mgg"1), Ca (12.3 mgg-1) and S (6.12 mgg"1).Brown leaves of white birch also had high concentra-tions of Ca, whereas the other litter types had levelsbelow 6 mg g"1. Relative differences in Mg concentra-tions between litter types were similar to those of Ca.Initial concentrations of Mn were highest in greenneedles of lodgepole pine and in brown leaves ofwhite birch. Alder leaves and Scots pine needles, onthe other hand, had fairly low concentrations of thiselement. Ash contents ranged between 10.5 and13.6 mg g"1 in the pine needles. For brown and greenbirch leaves the values were 22.4 and 23.6 mgg-1,respectively, and for grey alder leaves they were42.4mgg-1. Nutrient concentrations were in thesame range as those reported in other studies forlodgepole pine (Yavitt & Fahey 1986, Berg & Lund-mark 1987, Stump 1991), white birch and Scots pine(Berg & Staaf 1987, Bogatyrev et al. 1983).
Nitrogen, phosphorus and sulfur
The dynamics of single nutrients varied stronglyamong litter types. Some patterns were distinguish-able, however. Thus, concentrations of N, P and Sincreased linearly with accumulated mass loss (Table1, Fig. 1) once an initial leaching had taken place insome litter types, viz. in those litter types that werericher in P and S, (e.g. for P in grey alder leaves,green Scots pine needles, and green and brown leavesof white birch; S in grey alder leaves). The quickdecrease in concentration due to leaching was fol-lowed by a more stabilized phase, in which there wasa linear increase in concentration and in which also anet uptake took place in a few cases, e.g. for P inbirch leaves (Fig. 2) and for S in grey alder leaves. Itappears that for P the increases were more linear forthe litter types with lower initial concentrations (Fig.1, Table 1). The heavy initial loss can probably beascribed to leaching (cf. Bogatyrev et al. 1983).
The individual regressions of N concentrationsover mass loss showed very different slopes. Thus, thepositive slopes tended to be steeper the higher theinitial N concentration. Consequently, the relation-ship between rates of increase in N concentration,and the initial N concentrations including all littertypes, was highly significant (r = 0.975, n — 1,p < 0.001) (Table 2). This effect may be due to thefact that there was no real release of N from the litterin this early stage and that the uptake was relativelysmall (cf. Fig. 2), and that the N thus was concen-trated.
We may note that for litter types that were richerin N, P and S, the increase in concentration wasinterrupted and a plateau was reached, in a few casesfollowed by a decrease. This was seen at about40-50% accumulated mass loss. Although such ob-servations appear to be uncommon in the literature,R. Boone (pers. comm.) reported that in needles ofred pine (Pinus resinosa) in late stages of decomposi-tion a clear decrease in N concentration occurredafter the usual increase. In those cases where finalconcentrations of N could be distinguished, theseclearly differed (Fig. 1), depending on litter type, andranged from about 19mgg-1 in brown leaves ofwhite birch to 50.7 mgg"1 in grey alder leaf litter.For P, the final values ranged from about 1.0 mg g"1
(lodgepole pine brown needles) to 1.84 mgg"1 ingreen leaves of white birch. Final measured concen-trations of S varied from 1.05 mg g"1 (brown needlesof lodgepole pine) to 3.35 mg g"1 (grey alder leaves).Initial and finally measured concentrations of N, P
3
Dow
nloa
ded
by [
Uni
vers
ity o
f Il
linoi
s at
Urb
ana-
Cha
mpa
ign]
at 1
4:01
07
Oct
ober
201
4
B. Berg and J. Cortina Scand. J. For. Res. 10 (1995)
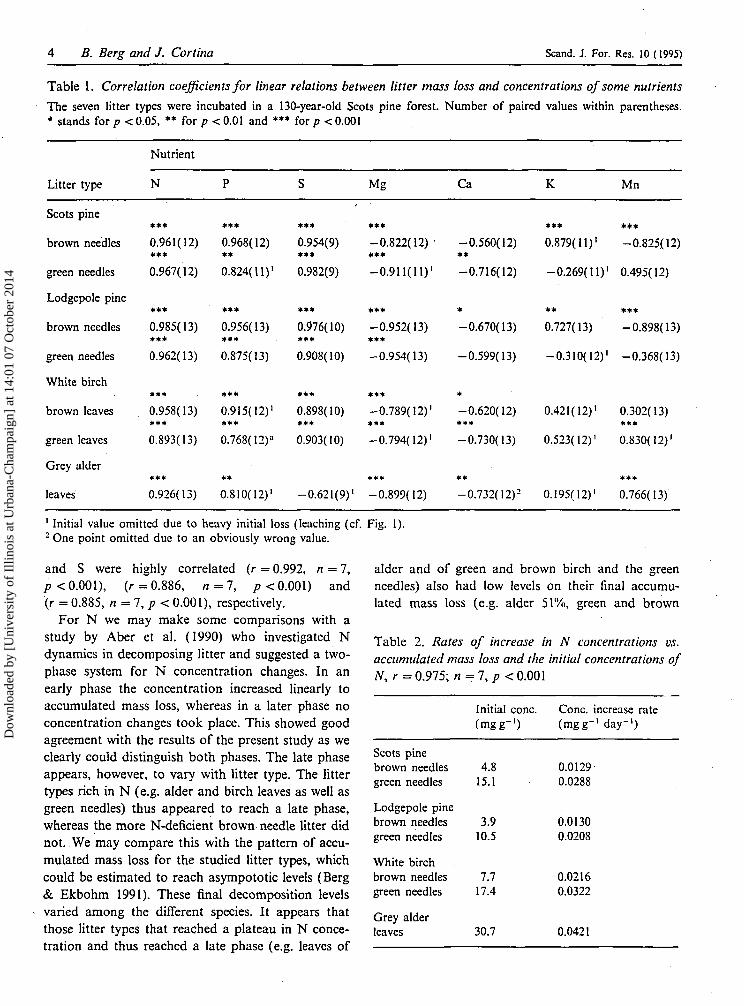
Table 1. Correlation coefficients for linear relations between litter mass loss and concentrations of some nutrients
The seven litter types were incubated in a 130-year-old Scots pine forest. Number of paired values within parentheses.* stands for p < 0.05, ** for p < 0.01 and *** for p < 0.001
Nutrient
Litter type N Mg Ca K Mn
Scots pine
brown needles
green needles
Lodgepole pine
brown needles
green needles
White birch
brown leaves
green leaves
Grey alder
leaves
• * *
0.961(12)***0.967(12)
* * •
0.985(13)• * *
0.962(13)
***
0.958(13)* • •
0.893(13)
***
0.926(13)
***
0.968(12)**0.824(11)'
***
0.956(13)* • •
0.875(13)
• **
0.915(12)'* * •
0.768(12)a
• *
0.810(12)'
• » *
0.954(9)* * •
0.982(9)
* • *
0.976(10)• • *
0.908(10)
***
0.898(10)***0.903(10)
-0.621(9)'
* * •
-0.822(12)***-0.911(11)'
***
-0.952(13)• * *
-0.954(13)
***
-0.789(12)'***-0.794(12)'
***
-0.899(12)
-0.560(12) 0.879(11)' -0.825(12)**-0.716(12) -0.269(11)' 0.495(12)
* ** ***-0.670(13) 0.727(13) -0.898(13)
-0.599(13) -0.310(12)' -0.368(13)
-0.620(12) 0.421(12)' 0.302(13)
-0.730(13) 0.523(12)' 0.830(12)'
-0.732(12)2 0.195(12)' 0.766(13)
' Initial value omitted due to heavy initial loss (leaching (cf. Fig. 1).2 One point omitted due to an obviously wrong value.
and S were highly correlated (r = 0.992, n = 7,p< 0.001), (r = 0.886, n = 7, p< 0.001) and(r = 0.885, n = 7,p< 0.001), respectively.
For N we may make some comparisons with astudy by Aber et al. (1990) who investigated Ndynamics in decomposing litter and suggested a two-phase system for N concentration changes. In anearly phase the concentration increased linearly toaccumulated mass loss, whereas in a later phase noconcentration changes took place. This showed goodagreement with the results of the present study as weclearly could distinguish both phases. The late phaseappears, however, to vary with litter type. The littertypes rich in N (e.g. alder and birch leaves as well asgreen needles) thus appeared to reach a late phase,whereas the more N-deficient brown needle litter didnot. We may compare this with the pattern of accu-mulated mass loss for the studied litter types, whichcould be estimated to reach asympototic levels (Berg& Ekbohm 1991). These final decomposition levelsvaried among the different species. It appears thatthose litter types that reached a plateau in N conce-tration and thus reached a late phase (e.g. leaves of
alder and of green and brown birch and the greenneedles) also had low levels on their final accumu-lated mass loss (e.g. alder 51%, green and brown
Table 2. Rates of increase in N concentrations vs.accumulated mass loss and the initial concentrations ofN, r = 0.975; n=7,p< 0.001
Scots pinebrown needlesgreen needles
Lodgepole pinebrown needlesgreen needles
White birchbrown needlesgreen needles
Grey alderleaves
Initial cone,(mgg- ' )
4.815.1
3.910.5
7.717.4
30.7
Cone, increase rate(mgg-1 day"1)
0.01290.0288
0.01300.0208
0.02160.0322
0.0421
4
Dow
nloa
ded
by [
Uni
vers
ity o
f Il
linoi
s at
Urb
ana-
Cha
mpa
ign]
at 1
4:01
07
Oct
ober
201
4
Scand. J. For. Res. 10(1995) Nutrients in decomposing litter
10 30 50 70Maas loss (r.)
10 30 SO 70Mass loss (%)
10 30 SO 70Mass loss (•%)
10 30 SO 70Mass loss (•/.)
10 30 50 70Mass loss (•'.)
10 30 SOMass loss (%)
30
£uc
8O10,s :
W\V\
Mass loss {•/.)
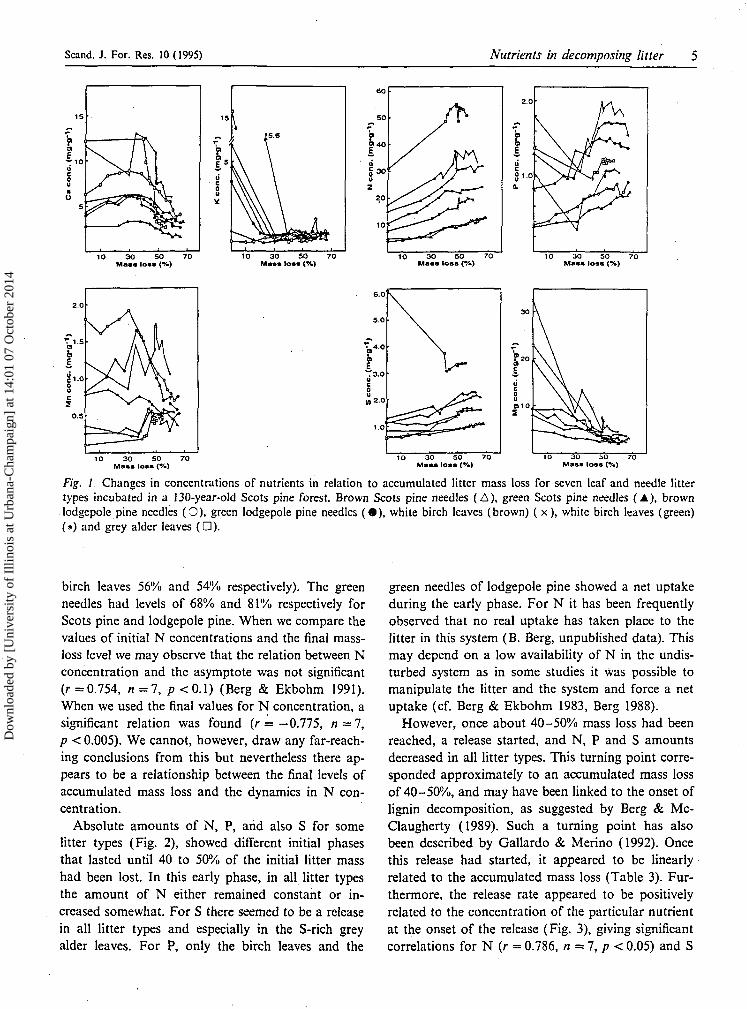
Fig. 1. Changes in concentrations of nutrients in relation to accumulated litter mass loss for seven leaf and needle littertypes incubated in a 130-year-old Scots pine forest. Brown Scots pine needles (A), green Scots pine needles (A), brownlodgepole pine needles (0) , green lodgepole pine needles ( • ) , white birch leaves (brown) (x) , white birch leaves (green)(*) and grey alder leaves (D).
birch leaves 56% and 54% respectively). The greenneedles had levels of 68% and 81% respectively forScots pine and lodgepole pine. When we compare thevalues of initial N concentrations and the final mass-loss level we may observe that the relation between Nconcentration and the asymptote was not significant(r =0.754, n = 7, p <0.1) (Berg & Ekbohm 1991).When we used the final values for N concentration, asignificant relation was found (r = - 0 . 7 7 5 , n = 7,p < 0.005). We cannot, however, draw any far-reach-ing conclusions from this but nevertheless there ap-pears to be a relationship between the final levels ofaccumulated mass loss and the dynamics in N con-centration.
Absolute amounts of N, P, and also S for somelitter types (Fig. 2), showed different initial phasesthat lasted until 40 to 50% of the initial litter masshad been lost. In this early phase, in all litter typesthe amount of N either remained constant or in-creased somewhat. For S there seemed to be a releasein all litter types and especially in the S-rich greyalder leaves. For P, only the birch leaves and the
green needles of lodgepole pine showed a net uptakeduring the early phase. For N it has been frequentlyobserved that no real uptake has taken place to thelitter in this system (B. Berg, unpublished data). Thismay depend on a low availability of N in the undis-turbed system as in some studies it was possible tomanipulate the litter and the system and force a netuptake (cf. Berg & Ekbohm 1983, Berg 1988).
However, once about 40-50% mass loss had beenreached, a release started, and N, P and S amountsdecreased in all litter types. This turning point corre-sponded approximately to an accumulated mass lossof 40-50%, and may have been linked to the onset oflignin decomposition, as suggested by Berg & Mc-Claugherty (1989). Such a turning point has alsobeen described by Gallardo & Merino (1992). Oncethis release had started, it appeared to be linearlyrelated to the accumulated mass loss (Table 3). Fur-thermore, the release rate appeared to be positivelyrelated to the concentration of the particular nutrientat the onset of the release (Fig. 3), giving significantcorrelations for N (r = 0.786, n=7,p < 0.05) and S
5
Dow
nloa
ded
by [
Uni
vers
ity o
f Il
linoi
s at
Urb
ana-
Cha
mpa
ign]
at 1
4:01
07
Oct
ober
201
4
6 B. Berg and J. Cortina Scand. J. For. Res. 10(1995)
O 10 30 SO 70Mass loss (y.)
50 70Mass loss (T.)
30 50Mass loss (
0 10 3O 50 7OMass loss (%)
30 SO 70Mass loss (•/.)
0 10 30 50 70Mass loss (%)
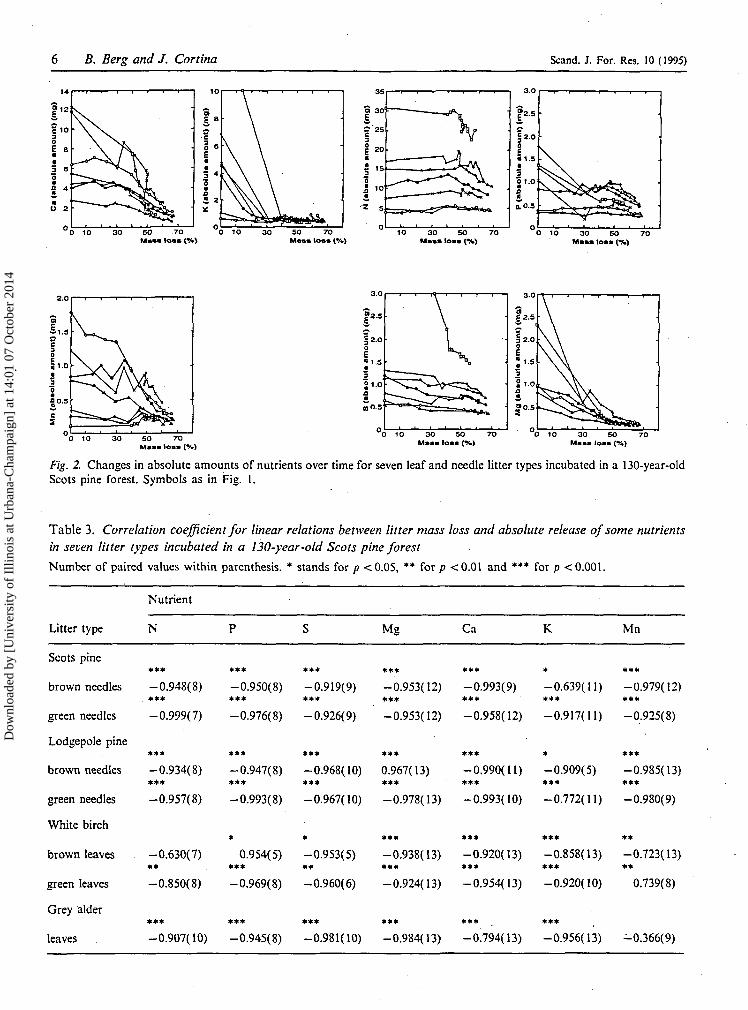
Fig. 2. Changes in absolute amounts of nutrients over time for seven leaf and needle litter types incubated in a 130-year-oldScots pine forest. Symbols as in Fig. 1.
Table 3. Correlation coefficient for linear relations between litter mass loss and absolute release of some nutrientsin seven litter types incubated in a 130-year-old Scots pine forest
Number of paired values within parenthesis. * stands for p < 0.05, ** for/7 < 0.01 and *** for p <0.00I.
Nutrient
Litter type N Mg Ca K Mn
-0.950(8) -0.919(9) -0.953(12) -0.993(9) -0.639(11) -0.979(12)• * * * * • * * • * * * * * * * * *
-0.976(8) -0.926(9) -0.953(12) -0.958(12) -0.917(11) -0.925(8)
-0.947(8) -0.968(10) 0.967(13) -0.990(11) -0.909(5) -0.985(13)* * * »** • * * * * * * * * * * *
-0.993(8) -0.967(10) -0.978(13) -0.993(10) -0.772(11) -0.980(9)
0.954(5) -0.953(5) -0.938(13) -0.920(13) -0.858(13) -0.723(13)
-0.969(8) -0.960(6) -0.924(13) -0.954(13) -0.920(10) 0.739(8)
* * * • * * * * * * * * * • •
-0.945(8) -0.981(10) -0.984(13) -0.794(13) -0.956(13) -0.366(9)
Scots pine
brown needles
green needles
Lodgepole pine
brown needles
green needles
White birch
brown leaves
green leaves
Grey alder
leaves
**•
-0.948(8)
-0.999(7)
***
-0.934(8)*+•
-0.957(8)
-0.630(7)**-0.850(8)
* * •
-0.907(10)
Dow
nloa
ded
by [
Uni
vers
ity o
f Il
linoi
s at
Urb
ana-
Cha
mpa
ign]
at 1
4:01
07
Oct
ober
201
4
Scand. J. For. Res. 10(1995) Nutrients in decomposing litter
o<0CB
a
rel
c»0.3
Itro
c0o«5E 0.1
-
X
-
/
/
i
x7
//
//
//
/
//
X
I 1 1
A
r2 = 0.62n =7p <0.05
i i
10 30 50
Nitrogen cone, (mg -g"1)
70
•da
y')
»0.02
> re
leas
e (
1»0.01
Rat
e of
phc
X
-
/
X
/
/
X
B
r2 = 0.62n =7p <0.05
1 2
Phosphorus eonc. (mg • g"1)
0.03
0.02
0.01
X
x/
x/
1
/
/
X
1
/
/
r2
nP
i
> c
= 0.69= 7<0.05
i
1 • 2 3 4
Sulfur conc. (mg • g*1)
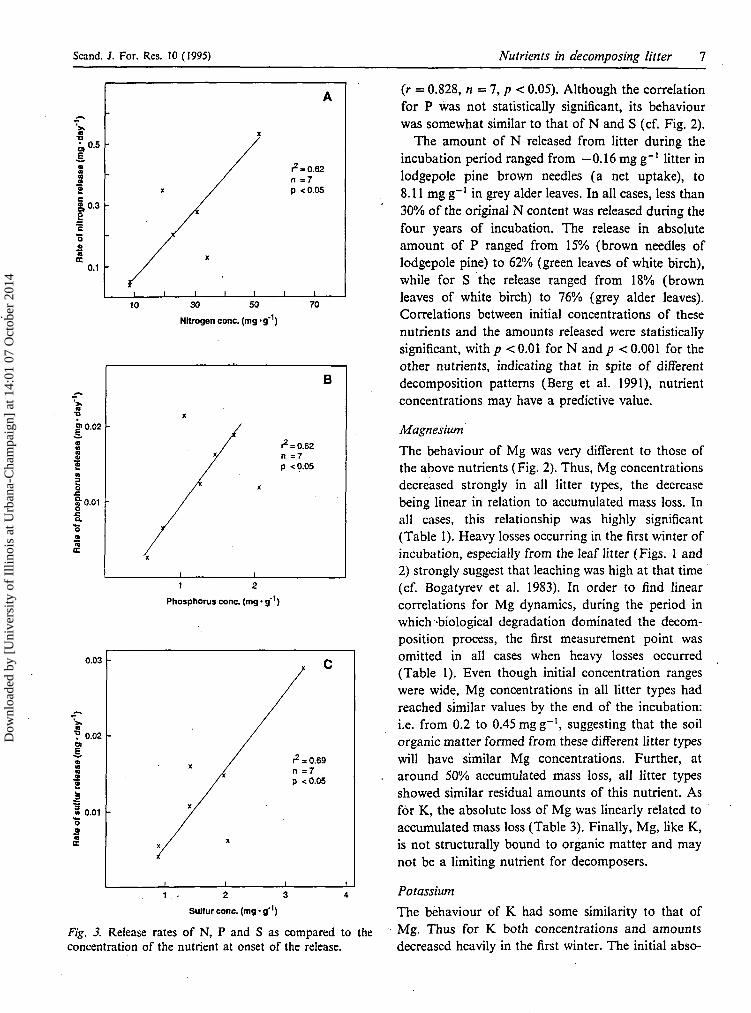
Fig. 3. Release rates of N, P and S as compared to theconcentration of the nutrient at onset of the release.
(r = 0.828, n = 7, />< 0.05). Although the correlationfor P was not statistically significant, its behaviourwas somewhat similar to that of N and S (cf. Fig. 2).
The amount of N released from litter during theincubation period ranged from —0.16 mg g~' litter inlodgepole pine brown needles (a net uptake), to8.11 mg g"1 in grey alder leaves. In all cases, less than30% of the original N content was released during thefour years of incubation. The release in absoluteamount of P ranged from 15% (brown needles oflodgepole pine) to 62% (green leaves of white birch),while for S the release ranged from 18% (brownleaves of white birch) to 76% (grey alder leaves).Correlations between initial concentrations of thesenutrients and the amounts released were statisticallysignificant, with p < 0.01 for N and p < 0.001 for theother nutrients, indicating that in spite of differentdecomposition patterns (Berg et al. 1991), nutrientconcentrations may have a predictive value.
Magnesium
The behaviour of Mg was very different to those ofthe above nutrients (Fig. 2). Thus, Mg concentrationsdecreased strongly in all litter types, the decreasebeing linear in relation to accumulated mass loss. Inall cases, this relationship was highly significant(Table 1). Heavy losses occurring in the first winter ofincubation, especially from the leaf litter (Figs. 1 and2) strongly suggest that leaching was high at that time(cf. Bogatyrev et al. 1983). In order to find linearcorrelations for Mg dynamics, during the period inwhich ••biological degradation dominated the decom-position process, the first measurement point wasomitted in all cases when heavy losses occurred(Table 1). Even though initial concentration rangeswere wide, Mg concentrations in all litter types hadreached similar values by the end of the incubation:i.e. from 0.2 to 0.45 mgg"1 , suggesting that the soilorganic matter formed from these different litter typeswill have similar Mg concentrations. Further, ataround 50% accumulated mass loss, all litter typesshowed similar residual amounts of this nutrient. Asfor K, the absolute loss of Mg was linearly related toaccumulated mass loss (Table 3). Finally, Mg, like K,is not structurally bound to organic matter and maynot be a limiting nutrient for decomposers.
Potassium
The behaviour of K had some similarity to that ofMg. Thus for K both concentrations and amountsdecreased heavily in the first winter. The initial abso-
7
Dow
nloa
ded
by [
Uni
vers
ity o
f Il
linoi
s at
Urb
ana-
Cha
mpa
ign]
at 1
4:01
07
Oct
ober
201
4
8 B. Berg and J. Cortina Scand. J. For. Res. 10 (1995)
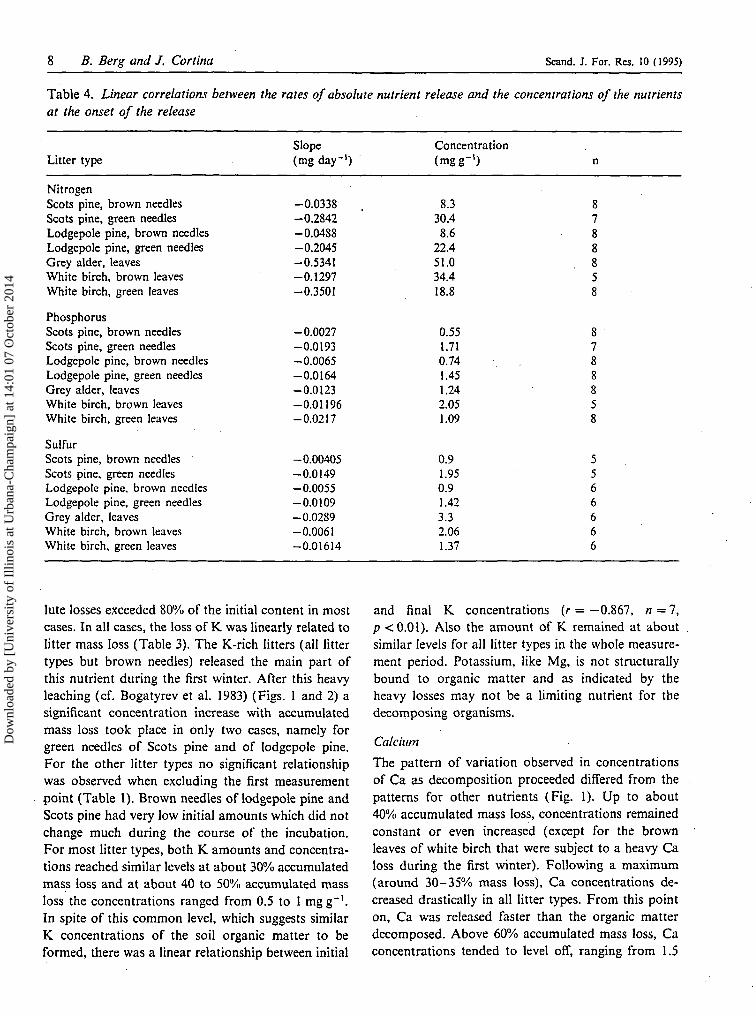
Table 4. Linear correlations between the rates of absolute nutrient release and the concentrations of the nutrientsat the onset of the release
Litter type
NitrogenScots pine, brown needlesScots pine, green needlesLodgepole pine, brown needlesLodgepole pine, green needlesGrey alder, leavesWhite birch, brown leavesWhite birch, green leaves
PhosphorusScots pine, brown needlesScots pine, green needlesLodgepole pine, brown needlesLodgepole pine, green needlesGrey alder, leavesWhite birch, brown leavesWhite birch, green leaves
SulfurScots pine, brown needlesScots pine, green needlesLodgepole pine, brown needlesLodgepole pine, green needlesGrey alder, leavesWhite birch, brown leavesWhite birch, green leaves
Slope(mg day"1)
-0.0338-0.2842-0.0488-0.2045-0.5341-0.1297-0.3501
-0.0027-0.0193-0.0065-0.0164-0.0123-0.01196-0.0217
-0.00405-0.0149-0.0055-0.0109-0.0289-0.0061-0.01614
Concentration(mgg"1)
8.330.4
8.622.451.034.418.8
0.551.710.741.451.242.051.09
0.91.950.91.423.32.061.37
n
8788858
8788858
5566666
lute losses exceeded 80% of the initial content in mostcases. In all cases, the loss of K was linearly related tolitter mass loss (Table 3). The K-rich litters (all littertypes but brown needles) released the main part ofthis nutrient during the first winter. After this heavyleaching (cf. Bogatyrev et al. 1983) (Figs. 1 and 2) asignificant concentration increase with accumulatedmass loss took place in only two cases, namely forgreen needles of Scots pine and of lodgepole pine.For the other litter types no significant relationshipwas observed when excluding the first measurementpoint (Table 1). Brown needles of lodgepole pine andScots pine had very low initial amounts which did notchange much during the course of the incubation.For most litter types, both K amounts and concentra-tions reached similar levels at about 30% accumulatedmass loss and at about 40 to 50% accumulated massloss the concentrations ranged from 0.5 to 1 mgg"1.In spite of this common level, which suggests similarK. concentrations of the soil organic matter to beformed, there was a linear relationship between initial
and final K concentrations (/• =—0.867, n = 7,p < 0.01). Also the amount of K remained at aboutsimilar levels for all litter types in the whole measure-ment period. Potassium, like Mg, is not structurallybound to organic matter and as indicated by theheavy losses may not be a limiting nutrient for thedecomposing organisms.
Calcium
The pattern of variation observed in concentrationsof Ca as decomposition proceeded differed from thepatterns for other nutrients (Fig. 1). Up to about40% accumulated mass loss, concentrations remainedconstant or even increased (except for the brownleaves of white birch that were subject to a heavy Caloss during the first winter). Following a maximum(around 30-35% mass loss), Ca concentrations de-creased drastically in all litter types. From this pointon, Ca was released faster than the organic matterdecomposed. Above 60% accumulated mass loss, Caconcentrations tended to level off, ranging from 1.5
Dow
nloa
ded
by [
Uni
vers
ity o
f Il
linoi
s at
Urb
ana-
Cha
mpa
ign]
at 1
4:01
07
Oct
ober
201
4

Scand. J. For. Res. 10 (1995) Nutrients in decomposing litter
to 5.0 mg g '. The range of Ca concentrations, there-fore, substantially narrowed during the incubationperiod. Even though the linear regressions were sig-nificant in most cases, the negative X2 functionspossibly describe the change in Ca concentrationmore accurately (cf. Fig. 1). There was no significantrelationship between initial and final concentrationsof this nutrient.
The changes in absolute amounts of Ca were simi-lar to those of N and P (Fig. 2). Initially, theamounts of Ca in the lodgepole pine needles re-mained constant, maybe combined with some smallimmobilization in some litter types, irrespective of theinitial concentration. The other litter types showed aninitial absolute release of Ca. Once the litter reached40-50% accumulated mass loss (all litter types), therate of release of Ca changed. The release of Ca atthis final stage was linearly related to accumulatedmass loss (Table 3). Cromack et al. (1975) andSollins et al. (1981) demonstrated that Ca can beaccumulated by fungi, probably as oxalate salts. It ispossible that a withdrawal of the active fungalbiomass (which could help to explain the decrease inN concentration in late stages of the decomposition)caused the precipitation of such salts, resulting in adecline in the Ca content. In this case, the rate of Carelease at this stage could be faster than the rate ofmass loss.
Manganese
Concentrations of Mn varied with accumulated massloss in a relatively complicated way. In Mn-poor litter(green leaves of white birch and leaves of grey alder)the concentration increased linearly with mass loss,levelling off at around 0.4-0.5 mg g~' (Fig. 1). Littertypes richer in Mn (green needles of Scots pine andbrown ones of lodgepole pine) showed a strong de-crease in concentration. For these litters the concen-tration range narrowed from 0.2 to 1.8 mgg"1
initially, to between 0.4 and 0.7mgg"1 at 60% massloss.
Changes in absolute amount of Mn showed avariety of complex patterns in the various litter types(Fig. 2). The range of absolute amounts the differentlitter types narrowed as decomposition proceeded,mainly owing to a release in the litters with highinitial Mn contents combined with an increase inabsolute amounts in those with initially lower con-centrations. Green needles of lodgepole pine, forinstance, showed an absolute uptake of Mn inspite of their relatively high initial concentration
(0.82 mg g"')- As previously described for Mg, K andCa, once 40-50% accumulated loss had been reached,all litter types showed a release of Mn, which was inlinear proportion to accumulated mass loss (Table 3).One exception was grey alder leaves with no loss,which was probably due to low initial levels.
CONCLUDING REMARKS
Even if such an expression as "soil improvement"normally is reflected by an increased growth rate oftrees, or assumed to be seen from soil classification,we may make some comparisons among actual chem-ical factors in the decomposing litter and attempt aninterpretation of the present data. There are somedifferent approaches to such an interpretation, suchas: (i) a comparison of the initial chemical composi-tion which reflects the incoming material to the sys-tem and (ii) a comparison of the final levels ofnutrients in the decomposing litter which may reflectthe nutrient status of the humus formed from theparticular litter.
The initial concentrations of the falling litter willreflect the input concentration. An important factorhere would have been basic knowledge of the amountof litter formed in these systems, reflecting the actualamounts of the nutrients reaching the systems. It is ashame to say, though, that we lack enough good dataon birch and alder litter fall, particularly, to developthis approach. However, assuming that nutrient con-centrations and availability are important factorswithin the concept of "soil improvement", we can see(above) that the initial concentrations of nutrients inalder and birch leaf litter were considerably higherthan those of the brown pine litter. For N, P, S, Mg,Ca and K. the concentrations were between 3 and 10times as high as in the brown pine needles, whereasfor Mn the picture was less clear. We may concludethat the litter of alder and birch clearly add litter tothe soil with considerably higher nutrient concentra-tions as compared to the pine species, and especiallyas compared to Scots pine, and if this is the impor-tant factor for soil improvement the difference is largeenough to justify such a difference. However, theinput rate of green needle litter may affect this con-clusion, as the green litter of both pine species werevery rich in the same nutrients.
When regarding concentrations in the late stagesfor the major nutrients, N, S and P appear to formone group with seemingly steady levels in the latedecomposition stages which could be expected to
9
Dow
nloa
ded
by [
Uni
vers
ity o
f Il
linoi
s at
Urb
ana-
Cha
mpa
ign]
at 1
4:01
07
Oct
ober
201
4

10 B. Berg and J. Cortina Scand. J. For. Res. 10(1995)
produce soil organic matter with different levels ofthese nutrients. Comparing brown needles withbrown leaves of birch and those of alder, we thuswould expect an alder soil to contain levels of Nabout 4 times as high, and a soil from brown birchleaves levels about twice as high. For P and S thealder soil could be 1.5 to 2 times as rich and the birchsoil possibly somewhat richer. For the nutrients Mgand K the concentrations would not be different andthe tendencies for Mn and Ca are not clear enough toallow any conclusion in this respect. We cannot ex-clude that the primary effect would be caused by highlevels of the major nutrients such as N, P and S, sincethe litter then also would have high levels of the othernutrients (cf. Berg & Staaf 1982).
ACKNOWLEDGEMENTS
We are most indebted to the very skilful technicalassistance of Mrs Annette Ewertsson. Mrs SirkkaKoivuoja is gratefully acknowledged for preparingthe litterbags. We also want to thank two anonymousreferees for their thorough work on this manuscript.
REFERENCES
Aber, J. D., Melillo, J. M. & McClaugherty, C. A. 1990.Predicting long-term patterns of mass loss, nitrogendynamics, and soil organic matter formation from initialfine litter chemistry in temperate forest ecosystems. Can.J. Bot. 68: 2201-2208.
Axelsson, B. & Bråkenhielm, S. 1980. Investigation sites ofthe Swedish Coniferous Forest Project. Biological andPhysiographical Features. Ecol. Bull. (Stockholm) 32:25-64.
Berg, B. 1988. Dynamics of nitrogen (15N) in decomposingScots pine (Pinus sylvestris L.) needle litter. Long-termdecomposition in a Scots pine forest VI. Canadian Jour-nal of Botany 66: 1539-1546.
Berg, B. 1991. Live fungal mycelium and lignin concentra-tions in some needle and leaf litter types. Scand. J. For.Res. 6: 451-462.
Berg, B. & Ekbohm, G. 1983. Nitrogen immobilization todecomposing needle litter at variable carbon-nitrogenratios. Ecology 64: 63-67.
Berg, B. & Ekbohm, G. 1991. Litter mass loss and decom-position patterns in some needle and leaf litter types.Long-term decomposition in a Scots pine forest VII.Can. J. Bot. 69: 1449-1456.
Berg, B. & Lundmark, J.-E. 1987. Decomposition of needlelitter in lodgepole pine and Scots pine monoculturalsystems - a comparison. Scand. J. For. Res. 2: 3-12.
Berg, B. & McClaugherty, C. 1989. Nitrogen and phospho-rus release from decomposing forest litter in relationto the disappearance of lignin. Can. J. Bot. 67: 1148—1156.
Berg, B. & Staaf, H. 1980. Decomposition rate and chemi-cal changes of Scots pine needle litter. II. Influence ofchemical composition. In: Persson, T. (ed). Structureand Function of Northern Coniferous Forests - AnEcosystem Study. Ecological Bulletins (Stockholm) 32:373-390.
Berg, B. & Staaf, H. 1987. Release of nutrients fromdecomposing white birch leaves and Scots pine needlelitter. Pedobiologia 30: 55-63.
Berg, B. & Wessen, B. 1984. Changes in organic-chemicalcomponents and ingrowth of fungal mycelium in decom-posing birch leaf litter as compared to pine needles.Pedobiologia 26: 285-298.
Berg, B., Booltink, H. G. W., Breymeyer, A., Ewertsson,A., Gallardo, A., Holm, B., Johansson, M.-B.,Koivuoja, S., Meentemeyer, V.; Nyman, P., Olofsson, J.,Pettersson, A.-S., Staaf, H., Staaf, I. & Uba, L. 1991.Data on needle litter decomposition and soil climate aswell as site characteristics for some coniferous forestsites. 2nd Edition, Section 2. Data on needle litterdecomposition. Swedish University of AgriculturalSciences. Department of Ecology and EnvironmentalResearch. Report No 42. ISSN 0348-422X.
Berg, B., Calvo de Anta, R., Ekbohm, G., Escudero, A.,Johansson, M.-B., Laskowski, R., Madeira, M., Mc-Claugherty, C, Meentemeyer, V., Reurslag, A. & VirzoDe Santo, A. 1995. The chemical composition of newlyshed needle litter of different pine species and Scots pinein a climatic transect. Long-term decomposition in aScots pine forest X. Can. J. Bot. (in press).
Bogatyrev, L., Berg, B. & Staaf, H. 1983. Leaching ofnutrients from leaf and needle litter. Swedish ConiferousForest Project. Swedish University of AgriculturalSciences, Uppsala. Technical Report 33, 25 pp. ISSN0346-7708.
Bonnevie-Svendsen, C. & Gjems, O. 1957. Amount andchemical composition of the litter from larch, beech,Norway spruce and Scots pine stands and its effects onthe soil. Meddelelser fra det Norske Skogsfor-söksvesende 48: 111 -174.
Cromack, K. Jr., Todd, R. L. & Monk, C. D. 1975.Patterns of basidiomycete nutrient accumulation inconiferous and deciduous forest litter. Soil Biol.Biochem. 7: 265-268.
Gallardo, A. & Merino, J. 1992. Nitrogen immobilizationin leaf litter at two Mediterranean ecosystems of SWSpain. Biogeochemistry 15: 13-228.
Gardiner, A. S. 1968. The reputation of birch for soilimprovement. Forestry Commission, London. Researchand Development Paper No. 67. 9 pp.
Johnson, D. W. 1992. Effects of forest management on soilcarbon storage. NCASI Technical Bulletin No. 628.New York, USA. 66 pp.
Laskowski, R., Berg, B., Johansson, M. & McClaugherty,C. Release pattern potassium from decomposing forestleaf litter. Long-term decomposition in a Scots pineforest IX. (unpubl.)
Mork, E. 1942. Om ströfallet i våre skoger. Meddelelser fradet Norske Skogsforsöksvesende 8: 297-365. (In Nor-wegian, with German summary.)
Dow
nloa
ded
by [
Uni
vers
ity o
f Il
linoi
s at
Urb
ana-
Cha
mpa
ign]
at 1
4:01
07
Oct
ober
201
4

Scand. J. For. Res. 10(1995) Nutrients in decomposing litter 11
Ny kvist, N. 1963. Leaching and decomposition of water-soluble substances from different types of leaf andneedle litter. Stud. Forest. Suec. 3: 1-31.
Sollins, P., Cromack, K., Fogel, R. & Li, C. Y. 1981. Roleof low molecular-weight organic acids in the inorganicnutrition of fungi and higher plants. In Wicklow, D. &Carroll, G. (eds). The Fungal Community, pp. 607-619.M. Decker, New York.
Stump, L. 1991. Relationships between litter quality and 'nitrogen availability in Rocky Mountain forests. M.Sc.Thesis, Colorado State University, Fort Collins CO.150 pp.
Svensson, G. & Anfält, T. 1982. Rapid determination ofammonia in whole blood and plasma using flow injec-tion analysis. Clinica Chimica Acta 119: 7-14.
Tamm, O. 1930. Om brunjorden i Sverige. SverigesSkogsvadsförbunds Tidskrift 28: 1-41. (In Swedish,with German summary.)
Van Cleve, K., Vierech, L. A. & Schlentner, R. L. 1971.Accumulation of nitrogen in alder (Alnus) ecosystems nearFairbanks, Alaska. Arctic and Alpine Res. 3: 101-114.
Viro, P. J. 1995. Investigations of forest litter. Comm. Inst.For. Fenn. 45(6): 1-65.
Weetman, G. F., Fournier, R. M. & Schnorbus, E. 1988.Lodgepole pine fertilization screening trials: four-yeargrowth response following initial prodictions. Soil Sei.Soc. Am. J. 52: 833-839.
Yavitt, J. B. & Fahey, T. J. 1986. Litter decay and leachingfrom the forest floor in Pinus contorta (lodgepole pine)ecosystems. J. Ecol. 74: 525-545.
Dow
nloa
ded
by [
Uni
vers
ity o
f Il
linoi
s at
Urb
ana-
Cha
mpa
ign]
at 1
4:01
07
Oct
ober
201
4











![Pine - Wikipedia · Pine - Wikipedia 6/20/2017 2:11:22 PM] Pinus sylvestris prepared for transport, Hungary Pine cone Tongue and groove solid pine](https://img.pdfslide.us/doc/110x75/5d62849d88c99309088b4588/pine-pine-wikipedia-6202017-21122-pm-pinus-sylvestris-prepared-for.jpg)

















