Embed Size (px)
Citation preview

(CANCER RESEARCH 58. 4269-4273. October I. 1998]
Advances in Brief
NUP98-HOXD13 Gene Fusion in Therapy-related Acute Myelogenous Leukemia1
Samina Z. Raza-Egilmez, Sheila N. Jani-Sait, Mauro Grossi, Michael J. Higgins, Thomas B. Shows, andPeter D. Aplan2
Cancer Genetics /S.Z.R-E., M.J.H., T. B. S., P. D.A.I. Pediatrics ¡M.G., P.D.AJ. Immunology [S. Z. R-E.¡, and Clinical Cylogenelics ¡S.N.J-S.], Roswell Park CancerInstitute, Buffalo. NY 14263; anil Department af PediatrìeHematnlogy-Oncnlngy. Children's Hospital of Buffalo. NY 14222 ¡M.G.. P. D. A.¡
Abstract
A novel chromosomal translocation, t(2;ll)(q31;pl5), was identified ina patient with therapy-related acute myelogenous leukemia (I-AMI.).
Fluorescence in \iin hybridization experiments mapped the breakpointnear NUP98; Southern blot analysis demonstrated that the nucleoporingene NUP98 was disrupted by this translocation. We used rapid amplification of cDNA ends to identify a chimi-rie iiiKN A. An in-frame, chimeric
mRNA that fused NVP98 sequences to the homeobox gene HOXD13 wascloned; the predicted fusion protein contains both the GLFG repeats fromNUP98 as well as the homeodomain from HOXD13. The NUP98-HOXD13fusion is structurally similar to the NVP98-HOXA9 fusion previouslyidentified in patients with AML, leading to the speculation that NUP98-
homeobox gene fusions may be oncogenic. Moreover, this report, alongwith a recent study that demonstrated NUP98-DDX10 fusions in patientswith I-AMI,, raises the possibility that NVP98 may be a previously unsuspected target for chromosomal translocations in patients with t-AMI ..
Introduction
Nonrandom, recurrent, chromosomal aberrations such as translocations and inversions are frequently associated with a wide spectrum ofhematological malignancies and are generally thought to be causalevents in the process of leukemic transformation ( 1). In general, thesetranslocations or inversions are thought to be oncogenic either throughthe activation of a latent proto-oncogene [such as activation of c-mvc
by the t(8:14) translocation] or through the generation of a chimericoncoprotein, such as the bcr-abl protein produced by the 1(9:22)
translocation (1). The identification of the genes involved in thesechromosomal aberrations, as well as characterization of the fusionproteins produced, has proven to be a valuable starling point forunderstanding the process of leukemic transformation. NUP98 is a Mr98,000 component of the NPC.3 NUP98 is localized at the nucleo-
plasmic side of the NPC and is involved in the transport of RNA andprotein between the cytoplasm and nucleus (2). Recently, NUP98 hasbeen reported to be involved in a rare but recurrent chromosomaltranslocation, t(7;l I)(pl5pl5.5) (3, 4), as well as a chromosomalinversion, inv(l I)(pl5.5;q31) (5). The t(7;ll) fuses NUP98 in-frame
to a homeobox gene (HOXA9). whereas the inv(l I)(pl5.5:q31) fusesNUP98 to DDXÃŒO.a putative RNA helicase gene. Both of thesechromosomal aberrations involving NUP98 were recognized in patients with AML. Moreover, the inv(l I)(pl5.5;q31) has been associ-
Received 6/22/98: accepted 8/17/98.The costs of publication of this article were defrayed in part hy the payment of page
charges. This article must therefore be hereby marked advertisement in accordance with18 U.S.C. Section 1734 solely to indicate this fact.
1Supported in part by the Roswell Park Alliance Foundation and NIH Grants
CAI6056, CA73773. and CA63333. P. D. A. is a scholar of the Leukemia Society ofAmerica.
2 To whom requests for reprints should be addressed, at Department of Pediatrics.
Roswell Park Cancer Institute, Buffalo, NY 14263. Phone: (716)845-5773; Fax:(716)845-4502; E-mail: [email protected].
3 The abbreviations used are: NPC. nuclear pore complex; AML. acute myelogenous
leukemia: t-AML. therapy-related AML; t-MDS. therapy-related myclodysplastic syndrome: FISH, fluorescence in situ hybridization: RACE, rapid amplification of cDNAends; AP. adapter primer.
ated with t-AML and t-MDS (5). t-AML and t-MDS arc malignancies
that occur following treatment of a primary malignancy with cytotoxicchemotherapcutic agents or ionizing radiation. As a first approximation, there seem to be two forms of t-AML, which can be distin
guished by clinical and cytogenetic features. The first form is typifiedby the therapeutic use of alkylating agents; a long (5-10-year) latency
period, often associated with a preceding myelodysplastic phase; andthe loss of chromosome 5 or 7 material. The second form is associatedwith the use of topoisomerase II inhibitor, a short (<24-month)
latency period, and balanced translocations involving either the MLLor AMLl gene (6). The speculation that these t-AMLs are caused bychromosomal aberrations induced by cytotoxic, DNA-damaging
chemotherapy is strengthened by the observation that both Eloposide(a topoisomerase II inhibitor) and Melphalan (an alkylating agent) caninduce frequent gross chromosomal abnormalities, such as translocations, inversions, and deletions, in peripheral blood leukocytes (7, 8).Here, we describe the characterization of a novel chromosomal trans-location, a t(2;l I)(q31:pl5.5), which occurred in a patient with t-MDSthat progressed to t-AML.
Materials and Methods
FISH. Cytogenetic analysis of bone marrow cells was performed in accordance with standard techniques, as described previously (9). Fixed cellsstored at —¿�20°Cwere used for FISH studies with previously mapped PAC
clones from 1Ipl5.5 (Fig. IA), as described previously (9, 10).Nucleic Acid Isolation. In accordance with institute guidelines, informed
consent to participate in research studies was obtained from the patient's
parents. Leukemic blasts were isolated from the bone marrow of patient 229 byFicoll-Hypaque (Sigma Chemical Co.) density centrifugaron. Genomic DNAwas isolated using a salting-out technique, as described previously (11). Total
RNA was isolated using Trizol reagent (Life Technologies, Inc.) and themanufacturer's recommended protocol. Plasmid DNA was isolated using Qia-
gen reagents and protocols.Southern Blots and Probes. Ten ^tg of genomic DNA were digested with
the indicated restriction en/.yme (Life Technologies. Inc.). si/e-fractionated on
0.8% agarose gels containing I /xg/ml ethidium bromide, photographed, denatured, neutralized, and transferred to nitrocellulose membranes (Schleicher& Schuell) by the Southern technique (12). Southern blots were hybridized to12P-labeled probes labeled by the random priming technique using Prime-It II
(Stratagene) reagents and protocols. Oligonucleotides were labeled using aterminal deoxynucleotidyl transf'erase end-labeling technique, and hybridiza
tion was performed as described previously (13). The probes used were a1.4-kb tfindIII-£r»RV NUP98 cDNA fragment (nucleotides 1249-2628 of
GenBank accession no. U41815; a kind gift of Dr. Julian Borrow, Center forCancer Research. MIT. Cambridge. MA), a 0.3-kb Mlul-BamHl ETV6 cDNAprobe (nucleotides 813-1142 of GenBank accession no. Ul 1732), and selected
synthetic oligonucleotides.3'-RACE. 3' RACE was performed using reagents from Lite Technolo
gies, Inc., and a modified protocol. Briefly, first strand cDNA was synthesizedfrom 1 /¿gof total RNA using either an oligo(dT) AP (.V-GGCCACGCGTC-GACTAGTAcrrrrrrrrrrrrrrriT-3') or a randomAP IS'-GGC-CACGCGTCGACTAGTACNNNNNN-3'). An aliquot of the first strandcDNA was then amplified using a /VÕ/TOS-specific forward primer (5'-TG-GAGGGCCTCTTGGTACAGG-3': nucleotides 1461-1481 of GenBank ac-
4269
Research. on December 25, 2020. © 1998 American Association for Cancercancerres.aacrjournals.org Downloaded from

NUPW-HOXDÕ3 IN l-AML
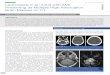
telnmere
II 111
Fig. I. A, map of chromosome Ilpl5.5 in the region of thetranslocation (modified from Ref. 10). Ordered markers and the location of PAC clones used as probes (/I73KI and 47C3) are indicated.Arrow, region of the breakpoint. B. FISH mapping with the l I73K1probe. Metaphase spreads from cells with the t(2;l 1) were hybridizedto the 1173K1 probe. Small arrowhead, the normal chromosome 11;larxe iirriwheatl, the derivative 11. C, FISH mapping with the 47G3probe. Metaphase spreads from cells with the 1(2;11) were hybridizedto the 47G3 probe. Small arrowhead, the normal chromosome 11:large arrowhead, the derivative 2.
cession no. U41815) and an abridged universal AP (5'-GGCCACGCGTC-GACTAGTAC-.V) as the reverse primer. A "hot start" was used by adding the
Taq polymerase after a 3-min incubation at 80°C;35 PCR cycles of 94°Cfor
1 min, 60°Cfor 45 s, and 72°Cfor 2 min were used, followed by a terminal
10-min extension at 72°C.The PCR products were analyzed by agarose gel
electrophoresis and subcloned into either the pGEMR-T or pGEMR-T Easy
vector (Promega).Nucleotide Sequence Analysis. Nucleotide sequencing was performed on
a Perkin-Elmer ABI PRISM Model 373A Stretch Automated Sequencer at the
Roswell Park Cancer Institute Biopolymer facility. Oligonucleotides weresynthesized on an Applied Biosystems oligonucleotide synthesizer at the above
facility.
Results
Case Report. The patient developed B-cell precursor acute lym-
phoblastic leukemia at 4 years of age. Cytogenetic analysis at thattime revealed a normal 46, XY karyotype. He did not have centralnervous system involvement at initial diagnosis, and he receivedstandard induction therapy consisting of vincristine, prednisone, L-asparaginase, and "triple" intrathecal therapy (hydrocortisone, me-
thotrexate, and 1-ß-D-arabinofuranosylcytosine). Consolidation andmaintenance therapies lasted 2.5 years and consisted of intermediate-dose methotrexate and methotrexate with 6-mercaptopurine, respec
tively. The patient was in continuous complete remission for 3 years,
4270
Research. on December 25, 2020. © 1998 American Association for Cancercancerres.aacrjournals.org Downloaded from

NUPW-HOXÛ13 IN t-AML
until he developed an isolated epidural relapse 6 months after cessation of therapy. At that time, he was again treated with vincristine,prednisone, and L-asparaginase, as well as 24 Gy of craniospinalirradiation. He received continuation therapy, consisting of high-dose1-ß-D-arabinofuranosylcytosine,etoposide, vincristine, L-asparaginase, cyclophosphamide, and doxorubicin. Because of religious beliefs that precluded the transfusion of blood products, he was treatedwith recombinant erythropoietin and granulocyte colony-stimulatingfactor following each course of chemotherapy. He completed a 2-yearcourse of therapy and again did well until isolated thrombocytopeniawas noted 9 months after he went off therapy; this progressed, and thepatient was noted to be pancytopenic 13 months after completion oftherapy. A bone marrow aspiration and biopsy at that time wasconsistent with a diagnosis of refractory anemia with excess blasts.Cytogenetic analysis at that time revealed a balanced t(2;l I)(q31;pl5)translocation. The myelodysplastic syndrome progressed to AML (M6variant) within 3 months, and a successful remission was induced with1-ß-D-arabinofuranosylcytosine,etoposide, and mitoxantrone, followed by consolidation with an allogeneic bone marrow transplant.
FISH Analysis. We used a set of previously characterizedchromosome 1Ipl5.5 probes (9, 10) to localize the t(2;l 1) breakpoint. Fig.1 demonstrates that the 1173K1 probe hybridized to the derivative 11chromosome and the normal 11, whereas the 47G3 probe hybridizedto the derivative 2 and the normal 11 (Fig. 1). Additionally, two moretelomeric 1Ipl5.5 probes (74K15 and 915F1; Ref. 10) also mapped tothe derivative 11 (data not shown). These findings placed the breakpoint between 1173K1 and 47G3. Because the 1173K1 mapped quiteclose to NUP98 and NUP98 was known to be translocated in otherAML cases, NUP98 was an obvious candidate gene for this translocation.
NUP98 Rearrangement. To investigate whether NUP98 was disrupted by this translocation, we searched for genomic DNA rearrangements within the NUP98 locus by Southern blot hybridization to the1.4-kb Hindlll-EcoRV NUP98 cDNA probe. As seen in Fig. 2A,non-germ-line-sized fragments are present in genomic DNA digestedwith multiple different restriction enzymes, suggesting a genomicDNA rearrangement had occurred within the NUP98 gene, most likelydue to a chromosomal translocation.
To rule out the possibility that the non-germ-line-sized bands weredue to a polymorphism, as well as to investigate whether the NUP98rearrangement was present in the initial leukemia, we searched forNUP98 rearrangements in genomic DNA from the original B-cellprecursor leukemia, genomic DNA from the t-AML, and genomicDNA from a remission sample obtained after treatment of the t-AML.In addition, because the initial leukemia had an ETV6-AMLI translocation, we were able to determine whether the initial leukemia hadevolved into an AML, through a lineage shift of the initial leukemicclone. Fig. IB shows that the ETV6 rearrangement was present in theinitial leukemia but not the t-AML or the remission sample. Similarly,the NUP98 rearrangement was present in the t-AML but not the initialleukemia nor the remission sample, demonstrating that the NUP98rearrangement was not due to a polymorphism or to a lineage shift ofthe initial leukemia.
Detection of a Fusion Transcript in the t-AML Sample. Weused 3' RACE to search for a NUP98 fusion transcript in RNA
from the t-AML sample. Because the previously reported NUP98fusion transcripts (3-5) join NUP98 exons to HOXA9 or DDX10exons at either nucleotide 1552 or 1864 of the NUP98 cDNA(GenBank accession no. U41815) in a head-to-tail fashion, wereasoned that the t(2;ll) may produce a NUP98 fusion transcriptwith a similar fusion point. We used two complementary 3' RACE
approaches to clone the NUP98 fusion transcript. For one set ofexperiments, we used an oligo(dT) tailed AP and a JVWPS-specific
4271
IECUU)co^M^CI^ooLU
evi Oo>eviCM Ü
O)evievi oO)evievi Ü
23-
9.6-
6.4-
4.4-
2.3_2.0-
BBamHI BamHI
4.4 —¿�
2.3 —¿�2.0 —¿�
Probe: ETV6 Probe: NUP98
Fig. 2. A, Soulhern blot hybridized lo Ihe 1.4-kh NUPW Hind\ll-Eti>RV cDNA probe.The restriction enzymes used are indicated above the lanes. 229. patient DNA; C. controlDNA from a lymphoblastoid cell line. Size standards are in kb. Rearranged fragments areseen in all four lanes containing patient DNA. B, comparison of primary acute lympho-blastic leukemia and t-AML. Lanes 1-4. Southern blot of fiiimHI-digcsled DNA fromindicated patient samples or control lymphoblastoid DNA (O hybridi/ed to the ETV6Mlu\-BanM\ cDNA probe. Lanes 5-8. the identical Southern blot was stripped andrehybridized to the NUP98 H/ndlll-EcoRV cDNA probe. Size standards are in kb.Non-germ-line sized fragments are evident in Lane\ 2 and 7.
Research. on December 25, 2020. © 1998 American Association for Cancercancerres.aacrjournals.org Downloaded from

NUP9K-HOXDI.ÃŒIN I-AML
primer (5'-TGGAGGGCCTCTTGGTACAGG-3') complementary
to NUP98 nucleotides 1461-1481. However, if the fusion transcript encoded a very large RNA, we realized that this approachmay not be successful if the reverse transcriptase had to reversetranscribe a large segment prior to reaching the NUP98 fusionpoint. Therefore, a second set of 3' RACE experiments used a
random hexamer AP and the same yVW9£-specificprimer. Bothapproaches were performed with templates from patient RNA, aswell as three control cell lines (HEL, K562, and Molt4) that did nothave a t(2;l 1) translocation: both approaches yielded PCR products that hybridized to NUP98 probes.
PCR products from the oligo(dT) primed experiment were sub-cloned into the pGEMR-T Easy vector. Twenty-five clones that hybridized to a nested, internal oligonucleotide 5' of the predictedbreakpoint (5'-GCCACTTTGGGCTTTGGAGC-3'; complementary
to NUP98 nucleotides 1516-1525) were evaluated. None of theseclones hybridized to a NUP98 probe 3' of the predicted breakpoint(5'-AGAAGTTGGTTTrGAAGAACC-3'), suggesting that none of
these clones represented germ-line NUP98 clones. We analyzed sixindependent subclones; these fell into two classes, with either a 0.5- or1.6-kb insert. We sequenced one clone from each class; both clonesdiverged from NUP98 germ-line sequences at nucleotide 1552 ofGenBank accession no. U41815, precisely where the NUP98-HOXA9fusion occurs (Fig. 3A). The sequence 3' of the fusion in each case
was a perfect match for the human HOXD13, which maps to humanchromosome 2q31: we did not detect a NUP98-HOXD13 fusionamong the PCR products from three control cell lines (HEL, K562,and Molt4). Therefore, the t(2;l I)(q31;pl5) generated a specific fusion mRNA between NUP98 and HOXD13. The 1.6-kb clones had a
Table 1 Translocations involving nucleoporin genes
Translocation1(2:1
1)q3l:pl5.5>t(7;ll)(pl5:pl5)inv(ll(pl5:q22)t(6;9)(p23;q34)t(4;ll(q21:pl5.5p)NPC
geneNUP9S
NUP98NUP98NUP214NUP98Partner
geneHOXDI3
HOXA9DDX10DEKUnknownDiseaset-AML
AMLDe novo and t-AML/MDS
AMLAMLRefs.3
and 45
2122
consensus polyadenylation signal (AATAAA) located 23 nucleotidesupstream of the poly(A) tail; the 0.5-kb clones did not have a polyadenylation signal and were generated by fortuitous annealing of theAP to a complementary sequence within the HOXD13 3'-untranslated
region. We were unable to amplify a potential reciprocal HOXDI3-NUP98 fusion transcript; this result is not surprising in light of theobservation that the HOXD13 promoter is not normally active inhematopoietic cells (14).
Discussion
This report demonstrates that NUP98 is fused to HOXDI3 in anovel t(2;11)(q31;p15) seen in a patient with t-AML. Because this wasthe sole cytogenetic abnormality seen in this t-AML sample, wesuspect that the NUP98-HOXD13 fusion was a causal event in thisdisease. The fusion transcript is composed of the 5' region of theNUP98 gene fused in frame to exon 2 and the 3' untranslated region
of the HOXD13 gene. The NUP98-HOXD13 fusion transcript is fusedat the same NUP98 exon as NUP98-HOXA9 and NUP98-DDX10fusions, suggesting the existence of a breakpoint cluster region withinthe NUP98 gene (14-16).
CCCCAGGCCCCAGTAGCTTTTGACAGATCCPQAPVACDRS
NUP98
CCCCAQGCCCCAGTAGGGGATGTGGCTCTAPQAPVGDVAL
NUP98/HOXD13
Fig. 3. A NUP98-HOXDIJ fusion cDNA. Thegerm-line NUP9X nucleotide sequence is shown (lupiin holdfiict' rvpt1. The germ-line HOXDI3 sequence
(hotlum) and the fusion cDNA (middle) are also shown.The fusion between NUP9X and HOXDI3 occurs between two Gs (nntlrrlineil). The amino acids encodedare shown below the nucleotide sequence. B, schematicdiagram of wild-type proteins and deduced chimericprotein. Arrm-s. fusion point of NUP98 and HOXD1J.
Regions that correspond to structural and functionaldomains of the proteins are also shown: GLFG, GLFGrepeats: RNA hind. RNA-binding domain (^): HD.
homeodomain (B); and HOXDIÃŒexon 1 (H).
CTTTTGTTTGTATCAGGGGATGTGGCTCTALLFVSGDVAL
HOXD13
B
NUP98
GLFG GLFG RNA bind
NUP98/HOXD13
HOXD13
HD
4272
Research. on December 25, 2020. © 1998 American Association for Cancercancerres.aacrjournals.org Downloaded from

NUPW-HOXD13 IN I-AML
BOX genes encode a large family of transcription factors containing a conserved DNA-binding homeobox domain (14, 15). HOX
genes are important for a wide spectrum of developmental events,including axial patterning and hematopoiesis (14, 16). Most of themammalian HOX genes are found in four conserved clusters (HOX A,B, C, and D); a minority of mammalian HOX genes (such as HOX 11)are "orphans" that are located outside of one of the principal clusters
(14). Members of the HOX A, B, and C clusters are normally expressed during distinct stages of hematopoietic differentiation. Incontrast, with the exception of HOXD3 expression in the erythroleu-
kemia cell line HEL (17), HOX D cluster genes are not normallyexpressed during hematopoietic development (16). Several homeoboxgenes, including HOXA9, HOX11, and PBXl, are known to be involved in recurrent, nonrandom chromosomal translocations associated with lymphoid and myeloid malignancies (3, 4, 18, 19).
The predicted NUP98-HOXD13 fusion protein (illustrated in Fig.
3ß)retains the conserved GLFG repeats OÕNUP98,which are thoughtto function as docking sites for karyopherin b at the nuclear pore (20).The HOXD13 portion of the predicted fusion protein retains thehomeobox domain, which is a helix-turn-helix DNA-binding domain.
Therefore, the putative fusion protein retains the ability to bind bothDNA as well as any transcription factors that bind to the homeodo-
main.It is not clear how a NUP98-HOXDÌ3fusion protein might con
tribute to leukemogenesis. The NUP98 portion of the fusion proteindoes not retain the RNA-binding domain OÕNUP98,which is replaced
by HOXDI3. Therefore, aberrant RNA transport could be responsiblefor oncogenesis. Alternatively, the localization of HOXD13 as part ofa NUP98-HOXDÌ3fusion protein at the nuclear pore might act as atrap for homeobox-binding transcription factors that normally bind
HOXDI3, resulting in their sequestration at the nuclear membrane. Ofnote, the erythroleukemia cell line HEL (17) demonstrates overex-
pression of HOXD3, raising the possibility that dysregulation ofHOXD family members may be generally associated with a subset oferythroleukemias.
A number of themes regarding NUP98 translocations are underscored by this report. First, this is now the second form of NUP98translocation associated with t-MDS or t-AML, suggesting that the
NUP98 locus, like MLL and AMLI, may be a target for nonrandomchromosomal translocations or inversions occurring as a result ofcytotoxic chemotherapy. Second, there are now at least five examples(Table 1) of translocations involving NPC genes in association withacute leukemia. Last, this is the second example of a translocationfusing NUP98 to a HOX gene, suggesting that both NUP98 and thehomeobox domains are important for malignant transformation.
Acknowledgments
We thank Drs. Carolyn Felix, Harish Ahuja. and Alex Dobrovic for helpfuldiscussions and Elaina Greco for artwork.
References
1. Rabbilt, T. H. Chromosomal translocations in human cancer. Nature (Lond.), 372:143-149. 1994.
2. Radu. A.. Moore. M. S., and Blobel, G. The peptide repeat domain of nucleoporinNUP98 functions as a docking sile in transport across the nuclear pore complex. Cell.81: 215-222. 1995.
3. Borrow. J., Shearman, A. M.. Stanton. V. P.. Jr.. Becher. R.. Collins. T.. Williams,A. J.. Dube, I.. Katz. F.. Kwong. Y. L.. Morris. C.. Ohyashiki, K.. Toyama, K.,Rowley. J.. and Houseman. D. E. The 1(7:11)(pl5;pl 5) translocation in acute myeloidleukemia fuses the genes for nucleoporin NUP98 and class 1 homeoprotein HOXA9.Nat. Genet.. 12: 159-167, 1996.
4. Nakamura. T.. Largaespada. D. A.. Lee, M. P.. Johnson. L. A.. Ohyashiki. K..Toyama. K.. Chen. S. J.. Willman. C. L.. Chen. 1. M.. Feinberg. A. P.. Jenkins, N. A.,Copeland. N. G.. and Shaughnessy. J. D. Fusion of the nucieoporin gene NUP9K toHOXA9 by the chromosomal translocation 1(7:11Hpl5:pl5l in human myeloid leukemia. Nat. Genet.. 12: 154-158, 1996.
5. Yasuhito. A.. Hosoda, F., Kohayashi. H.. Kyoko. A.. Hayashi. Y.. Nanao. K.. Kaneko.Y.. and Ohki. M. The inv(l I)(pl5.5:q31 )(pl5.5:q31 Xpl5;cj22). chromosome trans-location of de novo and therapy-related myeloid malignancies results in fusion of the
nucleoporin gene. NUP98. with the putative RNA helicase. DDX10. Blood, 89:3936-3944. 1997.
6. Smith. M. A.. Rubinstein. L., and Ungerleider, R. S. Therapy-related acute myeloid
leukemia following treatment with epipodophyllotoxins: estimating the risks. Med.Pediatr. Oncol., 23: 86-98. 1994.
7. Maraschin. J., Dutrillaux, B., and Aurias. A. Chromosome aberrations induced byetoposide are not random. Int. J. Cancer. 46: 808-812. 1990.
8. Mamuris. Z.. Prieur, M.. Dutrillaux. B.. and Aurias. A. The chemotherapcutic drugMelphalan induces breakage of chromosome regions rearranged in secondary leukemia. Cancer Genet. Cytogenet., 37: 65-77, 1989.
9. Sait, S. N. J., Nowak, N. J.. Singh-Kahlon. P.. Weksberg. R.. Squire. J.. Shows. T. B..and Higgins. M. J. Localization of Beckwith-Wiedemann and rhabdoid tumor chro
mosome rearrangements to a defined interval in chromosome band 1Ipl5.5. GenesChromosomes Cancer, //: 97-105, 1994.
10. Reid, L. H., Davies. C.. Cooper. P. R.. Cridermiller. S. J.. Sait. S. N. J.. Nowak, N. J.Evans. G., Stanbridge. E. J.. DeJong. P.. Shows. T. B.. Weissman. B. E.. and Higgins,M. J. A I-Mb physical map and pac contig of the imprinted domain in I Ipl5.5 thatcontains tapal and the bwscrl/wt2 region. Genomics. 43: 366-375. 1997.
11. Miller. S. A., Dykes, D. O.. and Polesky, H. F. A simple salting out procedure forextracting DNA from human nucleated cells. Nucleic Acids Res.. 16: 1215,1988.
12. Davis, L. G.. Dibner, M. D.. and Battey. J. F. Basic methods in molecular biology.New York: Elsevier. 1986.
13. Ausubel. F. M.. Brent. R.. Kingston, R. E.. Moore. D. D.. Seudman, J. G., Smith.J. A., and Sir uhi. K. Current Protocols in Molecular Biology. Vol. 1. New York: JohnWiley & Sons. 1995.
14. Scott. M. P. Vertebrate homeobox gene nomenclature. Cell. 71: 551-553. 1992.
15. Carroll. S. B. Homeotic genes and the evolution of arthropods and ehordates. Nature(Lond.), 376: 479-485, 1995.
16. Thorsleindottir, U., Sauvageau. G.. and Humphries. R. K. HOX homeobox genes asregulators of normal and leukemic hematopoiesis. Hematol. Oncol. Clin. North Am..//.•1221-1237, 1997.
17. Taniguchi. Y.. Komatsu. N.. and Moriuchi. T. Overexpression of HOX4A (HOXD3)homeobox gene in human erythroleukemia HEL cells results in altered adhesiveproperties. Blood. 85: 2786-2794. 1995.
18. Hatano. M., Roberts. C. W., Minden, M.. Crist. W. M., and Korsmeyer. S. J.Deregulation of a homeobox gene, by the 1(10:14) in T cell leukemia. Science(Washington DC), 253: 79-82, 1991.
19. Kamps. M. P.. Murre. C.. Sun. X. H., and Baltimore. D. A new homeobox gènecontributes the DNA binding domain of the t(l;19) translocation protein in prc-BALL. Cell, 60: 547-555. 1990.
20. Moroiannu. J.. Hijikala. M.. Blobel. G.. and Radu. A. Mammalian karyopherin alband a2b heterodimers: a! or a2 subunil binds nuclear localization signal and b subunitinteracts with peptide repeat-containing nucleoporins. Proc. Nati. Acad. Sci. USA. 92:6332-6536. 1995.
21. von Lindern. M., Poustka. A.. Lerach. H.. and Grosveld. G. The translocation (6:91.associated with a specific subtype of acule myeloid leukemia, results in the fusion ofa chimaeric. leukemia-specific deli-can mRNA. Mol. Cell. Biol.. 12: 1687-1697.
1992.22. Dobrovic. A., Hussey. D.. and Sage. R. E. NUP9H is the gene at the chromosome 11
breakpoint of the translocation t(4; 11)(q2l ;pl5.5) in a patient with acute lymphocylicleukemia. Blood. 90 (Suppl.): 3l8a. 1997.
4273
Research. on December 25, 2020. © 1998 American Association for Cancercancerres.aacrjournals.org Downloaded from

1998;58:4269-4273. Cancer Res Samina Z. Raza-Egilmez, Sheila N. Jani-Sait, Mauro Grossi, et al. Myelogenous Leukemia
Gene Fusion in Therapy-related AcuteNUP98-HOXD13
Updated version
http://cancerres.aacrjournals.org/content/58/19/4269
Access the most recent version of this article at:
E-mail alerts related to this article or journal.Sign up to receive free email-alerts
Subscriptions
Reprints and
To order reprints of this article or to subscribe to the journal, contact the AACR Publications
Permissions
Rightslink site. Click on "Request Permissions" which will take you to the Copyright Clearance Center's (CCC)
.http://cancerres.aacrjournals.org/content/58/19/4269To request permission to re-use all or part of this article, use this link
Research. on December 25, 2020. © 1998 American Association for Cancercancerres.aacrjournals.org Downloaded from