Embed Size (px)
Citation preview

NUEVO GÉNERO DE CROCODYLIA DEL EOCENO MEDIO DE LA PENÍNSULA IBÉRICA (ZAMORA, ESPAÑA): DUEROSUCHUS PISCATOR NOV. GEN., NOV. SP.
[New genus of Crocodylia from the Middle Eocene of the Iberian Peninsula (Zamora, Spain): Duerosuchus piscator nov. gen., nov. sp.]
Luis Alonso sAntiAgo (*)
Luis Alonso Andrés (*)
(*): Avda. de Portugal, 75 (5.º B). 37005 Salamanca. Correo: [email protected]
(FechA de recepción: 2009-06-10) (FechA de Admisión: 2009-07-20)BIBLID [0211-8327 (2009) 45 (2); 149-173]
RESUMEN: Se determina en el presente estudio un nuevo género y especie de Crocodylia, que denominamos Duerosuchus piscator, partiendo de la descripción del ejemplar holotípico hallado en sedimentos del Eoceno medio de la localidad de Corrales (Zamora, España) y de un análisis comparativo de los osteodermos pertenecientes a la misma localización.
Palabras clave: Crocodylia, Eusuchia, Duerosuchus piscator, Eoceno medio, Zamora, España.
ABSTRACT: In the present study it is described a new genus and species of Crocodylia name Duerosuchus piscator, based on the holotipic ejemplar found in middle Eocene outcrops of Corrales (Zamora, Spain), and the comparative analysis of the osteoderms belonging to the same place.
Key words: Crocodylia, Eusuchia, Duerosuchus piscator, Middle Eocene, Zamora, Spain.
l. Alonso sAntiAgo & l. Alonso Andrés
Nuevo género de Crocodylia del Eoceno medio de la Península Ibérica (Zamora, España):Duerosuchus piscator nov. gen., nov. sp.
© Ediciones Universidad de Salamanca Stud. Geol. Salmant., 45 (2), 2009: pp. 149-173
ISSN: 0211-8327 Studia Geologica Salmanticensia, 45 (2): pp. 149-173

INTRODUCCIÓN
La localidad zamorana de Corrales del Vino y los municipios ubicados en su entorno han dado lugar desde finales del siglo XIX a numerosas publicaciones científicas referidas tanto al estudio geológico de sus sedimentos, como a los hallazgos de restos fósiles de vertebrados eocénicos.
La distribución cronoestratigráfica de los crocodylianos de la cuenca del Duero abarca desde el Cretácico Inferior hasta el Eoceno medio, si bien los afloramientos se concentran en el lapso Eoceno medio y superior. Los yacimientos de Corrales, Casaseca y Cerro del Viso han sido datados en el MP 13-14 (Jiménez, 1992a; cuestA, 1999), si bien el análisis del registro fósil permitió delimitar informalmente en Corrales dos sectores asincrónicos, Corrales B y Corrales A, considerándose más antiguos los sedimentos de este último (Alonso & Alonso, 2005; Alonso et al., 2008).
Hasta el momento sólo se habían hallado tres géneros de Crocodylia en el sector Corrales B: Diplocynodon, Asiatosuchus e Iberosuchus.
SISTEMÁTICA
Neosuchia Clark in Benton & Clark, 1988
Eusuchia Huxley, 1875
Crocodylia Gmelin, 1789
Duerosuchus nov. gen
Especie-tipo: El espécimen n.º STUS 14.133 constituye el holotipo y especie-tipo. Las piezas se hallaron sin conexión anatómica separadas muy escasos centímetros, en un estrato areno-arcilloso.
El cráneo se presenta incompleto y en muy buen estado de fosilización; su longitud aproximada es de 20 cm, por lo que calculamos de manera proporcional, atendiendo a los parámetros de las especies actuales, que la longitud del individuo sería de unos 160 cm. Carece de algunos huesos que se presentan en la figura 1 reconstruidos por aproximación. El pterigoides se conserva desplazado, debido a que la inusual dureza de la matriz areno-arcillosa no aconsejó una intervención más agresiva sobre el fósil. El yugal apareció desconectado. La mandíbula preserva ambos dentarios y un angular. La ornamentación dorsal consiste en alvéolos redondeados, de distinto tamaño y profundidad, bastante discontinuos entre sí, y especialmente concentrados en algunas zonas del cráneo. Hemos incluido en el estudio varias vértebras (figura 17), que consideramos pertenecientes al mismo ejemplar, por la cercanía de su localización (unos 35 cm), dada la circunstancia de que el ejemplar apareció aislado en un estrato que contenía muy escasos restos fosilíferos.
Se incluyen en la descripción varios osteodermos de diversos individuos hallados en el mismo sector Corrales B, atribuidos al nuevo género por la
l. Alonso sAntiAgo & l. Alonso Andrés
Nuevo género de Crocodylia del Eoceno medio de la Península Ibérica (Zamora, España):Duerosuchus piscator nov. gen., nov. sp.
© Ediciones Universidad de Salamanca Stud. Geol. Salmant., 45 (2), 2009: pp. 149-173
150

similitud de su ornamentación, una vez descartada su pertenencia a las especies conocidas (figura 15).
Las referencias numéricas efectuadas a lo largo del presente estudio son las descritas como “características usadas en análisis filogenético” en el Apéndice de sAlisbury et al., 2006.
Distribución temporal: Eoceno medio, parte media; MP 13-14.
Derivatio nominis: El nuevo género se denomina Duerosuchus, o cocodrilo del Duero, por haber sido hallado en la actual cuenca del río Duero.
Diagnosis: El género Duerosuchus presentaría externamente la morfología de los cocodrilos piscívoros de pequeña talla, semejante a los actuales Crocodylus novaeguineae o Crocodylus mindorensis. La longitud total de los ejemplares adultos de Duerosuchus se hallaría en torno a los 160 cm, por unos 20 cm de longitud del cráneo. Destaca especialmente el perfil lanceolado de los siete primeros dientes del maxilar (figura 6), en contraste con los cinco dientes premaxilares, de base cilíndrica y comparativamente más estilizados que los dientes premaxilares de Diplocynodon. La ornamentación de su dentición es tenue, y consiste en finas aristas subparalelas dirigidas hacia el ápice. Coexiste una muesca interna en el premaxilar superior, con una escotadura en la zona dorsal del premaxilar que permitiría la recepción externa del cuarto diente inferior. A su vez se aprecia la oclusión de la hilera inferior del dentario por la dentición del maxilar.
Diferencias con otros géneros. Presencia proporcional: Los ejemplares de Asiatosuchus sp. de la cuenca del Duero se consideran próximos a Asiatosuchus germanicus, presentando peculiaridades que deberán apreciarse
l. Alonso sAntiAgo & l. Alonso Andrés
Nuevo género de Crocodylia del Eoceno medio de la Península Ibérica (Zamora, España):Duerosuchus piscator nov. gen., nov. sp.
© Ediciones Universidad de Salamanca Stud. Geol. Salmant., 45 (2), 2009: pp. 149-173
151
Figura 1. Duerosuchus piscator nov. gen., nov. sp. (STUS 14.133). Eoceno medio. Corrales (Zamora). Vista dorsal. (CO: Cóndilo occipital; FDT: Foramen del primer diente; F: Frontal;
FA: Foramen aereum; FI: Foramen incisivum; FIN: Fenestra infratemporal; FST: Fosa supratemporal; J: Yugal; LA: Lacrimal; MX: Maxilar; NA: Nasal; NE: External naris;
OR: Órbita ocular; PA: Parietal; PMX: Premaxilar; PO: Postorbital; POR: Pilar postorbital; PRF: Prefrontal; Q: Cuadrado; QJ: Cuadradoyugal; SQ: Escamoso).

en el marco de una reconsideración global del género (buscAlioni, 1986). Sus osteodermos son especialmente gruesos, de contorno rectangular, con barra medial y alvéolos groseros e irregulares distribuidos por casi la totalidad de la superficie de la pieza (figura 16a); este último dato facilita su distinción con los osteodermos atribuidos a Duerosuchus, cuyos alvéolos, con tendencia a la forma circular, aparecen bastante separados y son escasamente pronunciados. Se destaca en ambos casos la coincidencia entre la ornamentación craneal y la de la cubierta ósea.
Los osteodermos del nuevo género tienden a la forma rectangular (apareciendo un segundo morfotipo de contorno ovalado) (figura 15) y suelen presentar una cresta medial, que recorre longitudinalmente parte de la superficie dorsal, carácter también observado en Asiatosuchus.
No existen dudas sobre las diferencias entre la respectiva dentición, que obedece en Asiatosuchus a parámetros próximos a los observados en modernos cocodrilos generalistas de gran talla, adaptándose la parte posterior de la hilera dentaria a una función moledora, previsiblemente destinada a triturar los caparazones de las dos especies de Allaeochelys. Toda la dentición de Duerosuchus se considera idónea para la captura de pequeñas presas, como peces o ejemplares de Allaeochelys en su primer estadio de desarrollo. (Acerca de la dentición de cocodrilos quelonófagos véase KArl & tichy, 2004).
Tampoco se plantean problemas sobre la distinción con las piezas del género Diplocynodon, el cual ha dado lugar a tres determinaciones específicas efectuadas sobre ejemplares de la cuenca del Duero (buscAlioni et al., 1992), considerándose actualmente que el género se restringe a los ejemplares originales franceses y los hallados en el Eoceno de la cuenca del Duero (ortegA & buscAlioni, 1992). Los osteodermos de Diplocynodon son comparativamente más finos, rectangulares, con estilizada barra medial y alvéolos de distribución homogénea (figura 16b), coincidiendo plenamente su ornamentación con la superficie dorsal de los huesos craneales. Únicamente se aprecia una semejanza general que pueda inducir a confusión en caso de hallazgos sueltos, entre los dientes del premaxilar y dentario de Duerosuchus y los de Diplocynodon. Comparativamente estos últimos presentan una ornamentación semejante, en crestas dirigidas hacia el ápice, pero especialmente marcada, con una base de tendencia nítidamente circular, y menor estilización longitudinal que la observada en Duerosuchus.
El género Iberosuchus (Antunes, 1975) se describe sobre material del Eoceno portugués, de Salamanca y de Zamora, y dientes aislados pertenecientes a localizaciones españolas. Se asignan a esta especie los restos de zifodontos hallados en el prepirineo francés y en la cuenca del Duero (mArtín et al., 1987; ortegA et al., 1996). Se observa que los osteodermos de Iberosuchus, de superficie rugosa y no alveolada, poseen un contorno irregular, recorridos longitudinalmente por una gruesa barra medial, que se ve cruzada a nivel variable por una carena transversal, por lo que no se suscitan dudas acerca de su adscripción (figura 16c).
l. Alonso sAntiAgo & l. Alonso Andrés
Nuevo género de Crocodylia del Eoceno medio de la Península Ibérica (Zamora, España):Duerosuchus piscator nov. gen., nov. sp.
© Ediciones Universidad de Salamanca Stud. Geol. Salmant., 45 (2), 2009: pp. 149-173
152

Resulta difícil establecer el porcentaje de presencia relativa de Duerosuchus, por cuanto entendemos que la estadística sobre los osteodermos ofrece resultados escasamente fiables, al resultar desconocida la superficie osificada y su número de placas óseas. A título meramente aproximativo, consideramos que la proporción entre individuos del género Diplocynodon y de Duerosuchus en el Eoceno medio de Corrales B sería de 7 a 1, o incluso superior. En Corrales A, Duerosuchus puede hallarse extremadamente rarificado, o bien ser inexistente (sobre los ecosistemas fluviales del Paleógeno de Castilla y León: Jiménez, 1992a y b; Jiménez et al., 1990, 1991; Jiménez & Alonso, 1993).
La proporción entre Duerosuchus y los restantes géneros de Corrales B resulta difícil de precisar. A su vez no puede afirmarse la presencia en Corrales B de otros géneros, con lo que se cumple por el momento la regla general de distribución usual de crocodylimorfos por depósito singular, normalmente establecida en cinco o inferior (bucKley et al., 2000; brochu, 2001a y b), incluyéndose en los depósitos del nivel superior de Corrales un morfotipo generalista de gran talla, otro de hábitos terrestres y dentición zifodonta que podría superar los cuatro metros, y dos géneros de tamaño inferior a los dos metros, incluyendo aquí a Duerosuchus.
Duerosuchus piscator nov. sp.
Espécimen tipo (holotipo): N.º 14.133 STUS. Incluye el cráneo, mandíbula y dos vértebras del mismo ejemplar.
Localidad tipo: Corrales del Vino, Zamora, España. Consideramos que el género Duerosuchus aparece en el denominado informalmente sector Corrales B (Alonso & Alonso, 2005). No podemos afirmar su presencia en el sector Corrales A, considerado más antiguo. Tampoco se ha podido acreditar su existencia en el resto del área Casaseca, que engloba al sector Corrales B, si bien aquí la presuponemos, por la similitud del registro fósil.
Horizonte: Eoceno medio, parte media; MP 13-14.
Diagnosis: La del género.
Derivatio nominis: La especie se denomina piscator, en referencia a sus presumibles aptitudes para la captura de peces, cuyos restos aparecen abundantemente en los sedimentos del entorno, y a la aparente idoneidad de la configuración lanceolada del sector intermedio de la dentición superior para la retención de tales presas.
Material referido: El cráneo y mandíbula, pertenecientes al mismo individuo, se conservan incompletos (STUS 14.133). Se incluyen en la descripción: dentarios, angular, premaxilar y maxilar, frontal, postorbitales, escamoso, parietal, cuadrados, basioccipital, yugal, supraoccipital, pterigoides y exoccipital. Se describen también dos vértebras atribuidas al mismo espécimen (figura 17), y varios osteodermos de diferentes individuos (STUS 14.134 a 14.139) (figura 15).
l. Alonso sAntiAgo & l. Alonso Andrés
Nuevo género de Crocodylia del Eoceno medio de la Península Ibérica (Zamora, España):Duerosuchus piscator nov. gen., nov. sp.
© Ediciones Universidad de Salamanca Stud. Geol. Salmant., 45 (2), 2009: pp. 149-173
153

DESCRIPCIÓN MORfOLÓGICA
MANDÍBULA INFERIOR
Dentarios: La mandíbula está representada por los dos dentarios, uno de ellos incompleto, y un angular (figuras 2 y 3).
El dental derecho se halla en mejor estado de conservación, preservándose 14 alvéolos. Las siguientes medidas corresponden a éste. La sínfisis se prolonga unos 38 mm hasta cerca del borde posterior del sexto alvéolo. Se observan en la zona lingual pequeñas inserciones alineadas atribuidas a nutrientes y forámenes nerviosos.
La presencia de muescas de recepción en la superficie interna del maxilar para el alojamiento de la dentición inferior, y la distancia entre los dentarios, permiten desechar la hipótesis de la interrelación entre los dientes inferiores y la hilera del maxilar (78-0). La serie superior ocluiría lateralmente a la dentición inferior en todos los tramos del dentario.
Se aprecia una gran separación entre los dientes 1.º-2.º, y entre los 2.º-3.º. En todos los alvéolos el eje medio lateral es de tamaño inferior al eje mesio-distal, configurando una apariencia elíptica.
Las medidas mesio-distales de los alvéolos y sus espacios intraalveolares se reflejan en la tabla 1.
Las distancias del eje mesio-distal de los alvéolos n.º 12 a 15 son estimativas. Desde el borde posterior del alvéolo 11 al borde posterior del alvéolo 17 la distancia es de 37,0 mm.
Se aprecia que el hueso dentario es lineal entre las posiciones 4 y 10 (68-2). A su vez el alvéolo de mayor tamaño es el n.º 4 (52-1), conservándose en el dentario derecho el cuarto diente, que sobresale unos 12 mm. No se conserva el esplenial, si bien la sutura permite advertir su presencia.
Dentición inferior: La dentición mandibular puede ser calificada como pseudoheterodonta. No existen grandes diferencias morfológicas entre los dientes distales y proximales. No existen dientes moledores. Se conservan los dientes 1.º a 4.º, 7.º a 10.º, y 13.º? Todos ellos son alargados y redondeados basalmente en forma elíptica, si bien escasamente comprimidos. La ornamentación del esmalte es muy débil, consiste en unas finas crestas subparalelas orientadas hacia el ápice de la corona. La zona basal carece de ornamentación. Presentan a su vez dos carenas o filos laterales, mesial y distal, totalmente lisas. En los dientes n.º 1 y n.º 4, se observa de forma más nítida una superficie cóncava en el área lingual, y convexa en la vista bucal. Tal característica se percibe a su vez en los demás dientes conservados, si bien de forma menos acusada.
Angular: Sólo se conserva un angular, fosilizado sin conexión anatómica. Formaría la posición caudoventral de la mandíbula externa, sin que se pueda
l. Alonso sAntiAgo & l. Alonso Andrés
Nuevo género de Crocodylia del Eoceno medio de la Península Ibérica (Zamora, España):Duerosuchus piscator nov. gen., nov. sp.
© Ediciones Universidad de Salamanca Stud. Geol. Salmant., 45 (2), 2009: pp. 149-173
154

precisar la extensión ventral de la fosa dentroangular, ni su posición relativa frente a los últimos alvéolos. La longitud de la pieza recuperada es de 117 mm.
No se conservó el supraangular, resultando a su vez desconocida la forma del articular, por lo que hasta que pueda ser hallado un ejemplar más completo no podrán establecerse comparaciones con otros géneros.
CRÁNEO. PARTE DELANTERA
Premaxilar y maxilar: (figuras 1, 6, 7, 8 y 9) Presentan cierta ornamentación externa poco acusada. La longitud del maxilar derecho es de 86 mm. El izquierdo se ha preservado aun más incompleto. Se conserva la lámina palatal (figuras 6 y 7). En el espacio intraalveolar, en la zona lingual, se presentan depresiones laterales muy marcadas. En vista lateral el contacto
l. Alonso sAntiAgo & l. Alonso Andrés
Nuevo género de Crocodylia del Eoceno medio de la Península Ibérica (Zamora, España):Duerosuchus piscator nov. gen., nov. sp.
© Ediciones Universidad de Salamanca Stud. Geol. Salmant., 45 (2), 2009: pp. 149-173
155
Figura 2. Duerosuchus piscator nov. gen., nov. sp. (STUS 14.133). Eoceno medio. Corrales (Zamora). Dentario.
Alvéolon.º
Longitudmm
Distancia intraalveolarmm
1 4,00 4,802 3,95 5,503 3,25 1,104 6,20 2,155 4,95 3,006 4,95 1,307 5,15 2,008 3,60 1,009 4,00 2,00
10 4,60 0,9011 5,00 ?12 5,10 ?13 5,00 ?14 4,50 ?15 4,50 ?16 4,00 ?17 3,90 ?
Tabla 1
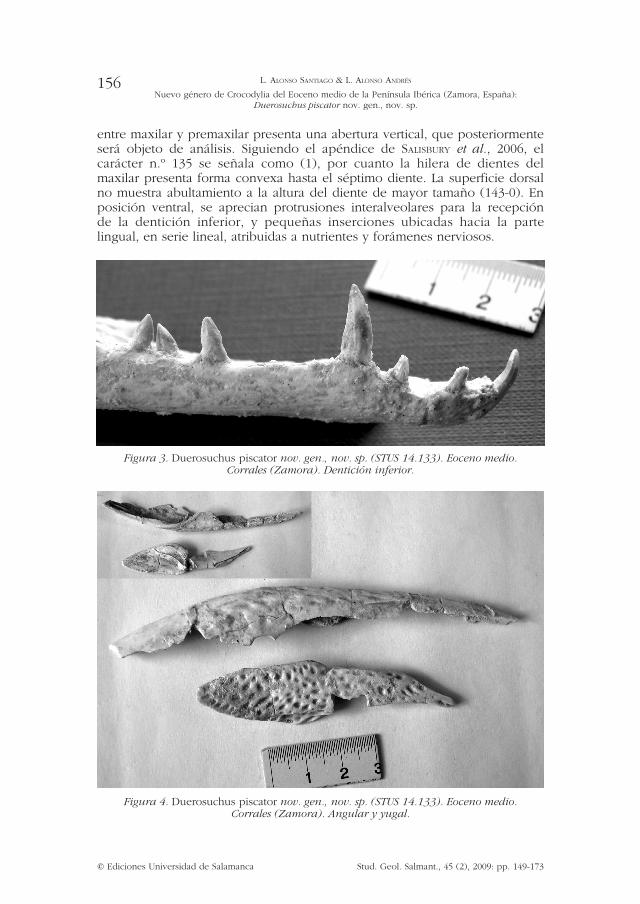
entre maxilar y premaxilar presenta una abertura vertical, que posteriormente será objeto de análisis. Siguiendo el apéndice de sAlisbury et al., 2006, el carácter n.º 135 se señala como (1), por cuanto la hilera de dientes del maxilar presenta forma convexa hasta el séptimo diente. La superficie dorsal no muestra abultamiento a la altura del diente de mayor tamaño (143-0). En posición ventral, se aprecian protrusiones interalveolares para la recepción de la dentición inferior, y pequeñas inserciones ubicadas hacia la parte lingual, en serie lineal, atribuidas a nutrientes y forámenes nerviosos.
l. Alonso sAntiAgo & l. Alonso Andrés
Nuevo género de Crocodylia del Eoceno medio de la Península Ibérica (Zamora, España):Duerosuchus piscator nov. gen., nov. sp.
© Ediciones Universidad de Salamanca Stud. Geol. Salmant., 45 (2), 2009: pp. 149-173
156
Figura 4. Duerosuchus piscator nov. gen., nov. sp. (STUS 14.133). Eoceno medio. Corrales (Zamora). Angular y yugal.
Figura 3. Duerosuchus piscator nov. gen., nov. sp. (STUS 14.133). Eoceno medio. Corrales (Zamora). Dentición inferior.
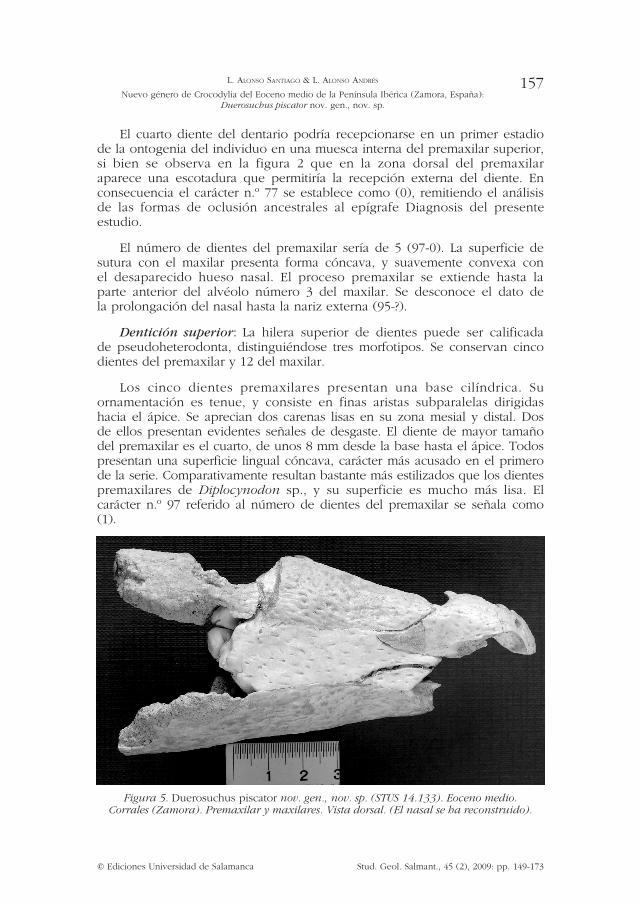
El cuarto diente del dentario podría recepcionarse en un primer estadio de la ontogenia del individuo en una muesca interna del premaxilar superior, si bien se observa en la figura 2 que en la zona dorsal del premaxilar aparece una escotadura que permitiría la recepción externa del diente. En consecuencia el carácter n.º 77 se establece como (0), remitiendo el análisis de las formas de oclusión ancestrales al epígrafe Diagnosis del presente estudio.
El número de dientes del premaxilar sería de 5 (97-0). La superficie de sutura con el maxilar presenta forma cóncava, y suavemente convexa con el desaparecido hueso nasal. El proceso premaxilar se extiende hasta la parte anterior del alvéolo número 3 del maxilar. Se desconoce el dato de la prolongación del nasal hasta la nariz externa (95-?).
Dentición superior: La hilera superior de dientes puede ser calificada de pseudoheterodonta, distinguiéndose tres morfotipos. Se conservan cinco dientes del premaxilar y 12 del maxilar.
Los cinco dientes premaxilares presentan una base cilíndrica. Su ornamentación es tenue, y consiste en finas aristas subparalelas dirigidas hacia el ápice. Se aprecian dos carenas lisas en su zona mesial y distal. Dos de ellos presentan evidentes señales de desgaste. El diente de mayor tamaño del premaxilar es el cuarto, de unos 8 mm desde la base hasta el ápice. Todos presentan una superficie lingual cóncava, carácter más acusado en el primero de la serie. Comparativamente resultan bastante más estilizados que los dientes premaxilares de Diplocynodon sp., y su superficie es mucho más lisa. El carácter n.º 97 referido al número de dientes del premaxilar se señala como (1).
l. Alonso sAntiAgo & l. Alonso Andrés
Nuevo género de Crocodylia del Eoceno medio de la Península Ibérica (Zamora, España):Duerosuchus piscator nov. gen., nov. sp.
© Ediciones Universidad de Salamanca Stud. Geol. Salmant., 45 (2), 2009: pp. 149-173
157
Figura 5. Duerosuchus piscator nov. gen., nov. sp. (STUS 14.133). Eoceno medio. Corrales (Zamora). Premaxilar y maxilares. Vista dorsal. (El nasal se ha reconstruido).
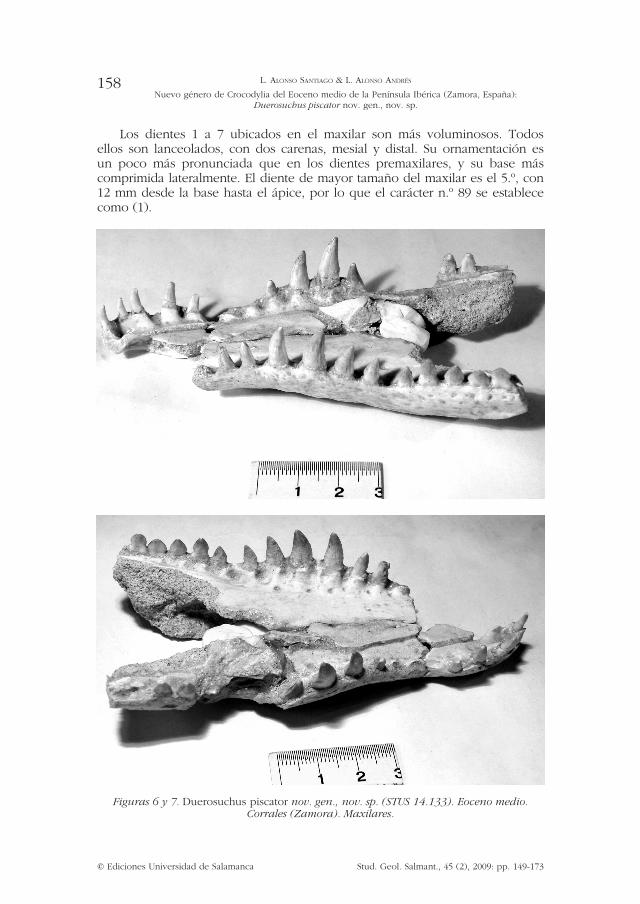
Los dientes 1 a 7 ubicados en el maxilar son más voluminosos. Todos ellos son lanceolados, con dos carenas, mesial y distal. Su ornamentación es un poco más pronunciada que en los dientes premaxilares, y su base más comprimida lateralmente. El diente de mayor tamaño del maxilar es el 5.º, con 12 mm desde la base hasta el ápice, por lo que el carácter n.º 89 se establece como (1).
l. Alonso sAntiAgo & l. Alonso Andrés
Nuevo género de Crocodylia del Eoceno medio de la Península Ibérica (Zamora, España):Duerosuchus piscator nov. gen., nov. sp.
© Ediciones Universidad de Salamanca Stud. Geol. Salmant., 45 (2), 2009: pp. 149-173
158
Figuras 6 y 7. Duerosuchus piscator nov. gen., nov. sp. (STUS 14.133). Eoceno medio. Corrales (Zamora). Maxilares.
l. Alonso sAntiAgo & l. Alonso Andrés
Nuevo género de Crocodylia del Eoceno medio de la Península Ibérica (Zamora, España):Duerosuchus piscator nov. gen., nov. sp.
© Ediciones Universidad de Salamanca Stud. Geol. Salmant., 45 (2), 2009: pp. 149-173
159
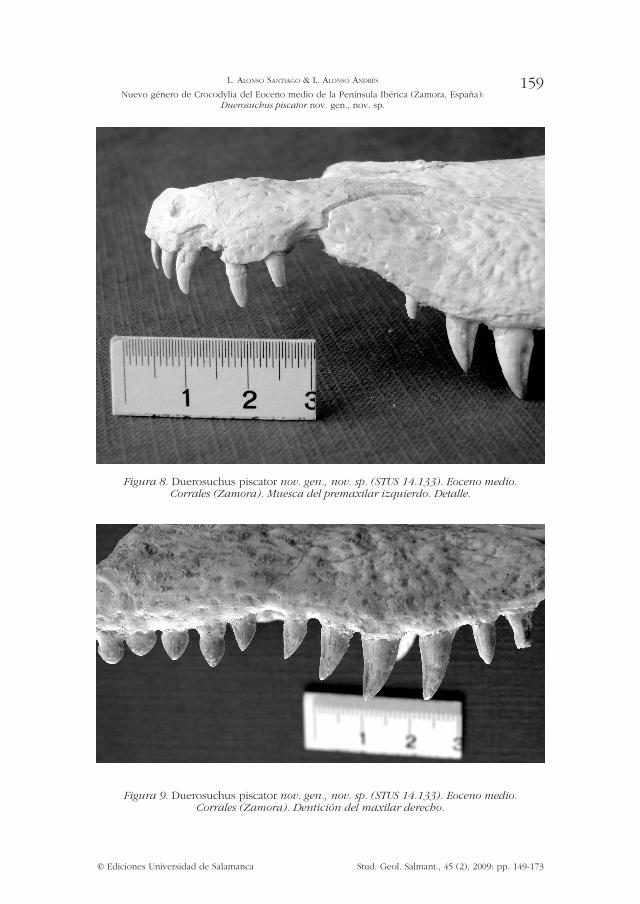
Figura 9. Duerosuchus piscator nov. gen., nov. sp. (STUS 14.133). Eoceno medio. Corrales (Zamora). Dentición del maxilar derecho.
Figura 8. Duerosuchus piscator nov. gen., nov. sp. (STUS 14.133). Eoceno medio. Corrales (Zamora). Muesca del premaxilar izquierdo. Detalle.

Los dientes 8 a 12 del maxilar son cortos y simétricos, presentándose comprimidos lateralmente. No se aprecia curvatura en su superficie lingual. Se observan igualmente dos carenas, mesial y distal, y una ornamentación de finas aristas ascendentes hacia el ápice.
CRÁNEO. APERTURAS CRANEALES
Narinas externas: Posiblemente, las narinas externas no se hallasen rodeadas enteramente por el premaxilar, si bien al no conservarse el hueso nasal, que aparece reconstruido en la figura 10 con masilla blanca, se desconoce si su extremo distal irrumpe en aquélla (95-?).
En la reconstrucción del cráneo representamos al nasal conectado con las narinas externas.
Se proyecta dorsalmente (79-1), y su forma es elipsoidal, con el eje mayor longitudinal de 21 mm, y el eje menor, transversal, de 16 mm (161-1).
Órbitas y fenestra infratemporal: Se desconoce la forma exacta de las órbitas y de la fenestra infratemporal, reconstruidas en la figura 1 por aproximación. No parece que el borde de las órbitas formado por la superficie del cráneo se eleve considerablemente (103-0). Se representan en la figura 1 de forma meramente aproximativa.
l. Alonso sAntiAgo & l. Alonso Andrés
Nuevo género de Crocodylia del Eoceno medio de la Península Ibérica (Zamora, España):Duerosuchus piscator nov. gen., nov. sp.
© Ediciones Universidad de Salamanca Stud. Geol. Salmant., 45 (2), 2009: pp. 149-173
160
Figura 10. Duerosuchus piscator nov. gen., nov. sp. (STUS 14.133). Eoceno medio. Corrales (Zamora). Caja craneal. Vista dorsal.
l. Alonso sAntiAgo & l. Alonso Andrés
Nuevo género de Crocodylia del Eoceno medio de la Península Ibérica (Zamora, España):Duerosuchus piscator nov. gen., nov. sp.
© Ediciones Universidad de Salamanca Stud. Geol. Salmant., 45 (2), 2009: pp. 149-173
161
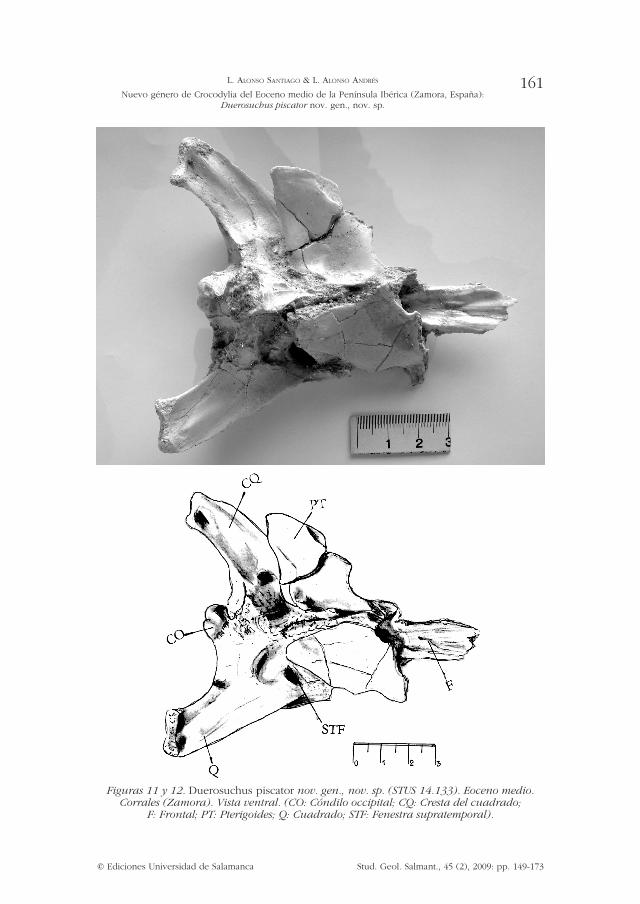
Figuras 11 y 12. Duerosuchus piscator nov. gen., nov. sp. (STUS 14.133). Eoceno medio. Corrales (Zamora). Vista ventral. (CO: Cóndilo occipital; CQ: Cresta del cuadrado;
F: Frontal; PT: Pterigoides; Q: Cuadrado; STF: Fenestra supratemporal).

Supratemporal: La ventana supratemporal se halla formada por el postorbital, parietal y escamoso, sin participación del frontal (87-0). Los márgenes laterales son claramente más largos que los mediales. Su forma externa se asemeja a la figura de un trapecio, cuya base mayor se orientaría hacia el postorbital y escamoso, y la base menor hacia el parietal, con las siguientes dimensiones; base mayor 17 mm, base menor 11,5 mm, altura 13 mm. Se aprecia una fosa de 1 mm de diámetro, levemente insinuada en la esquina anteromedial de la fenestra supratemporal, cercana a la superficie del parietal. Existe a su vez en la pared posterior de la fenestra supratemporal una fosa elipsoidal de 5 por 2 mm, situada entre la sutura del parietal y escamoso, a 3 mm de la superficie dorsal del cráneo.
Foramen incisivum: No se conserva completo, si bien se aprecia uno de sus bordes que permite precisar su localización relativa en el premaxilar, coincidiendo su borde anterior con el segundo alvéolo (153-0). Por la orientación del borde conservado consideramos que sería de reducido tamaño (124-0).
El foramen del primer diente es de forma elipsoidal y escaso tamaño Su eje mayor es de 5 mm, por 3,5 mm de su eje menor. Su distancia con la base del segundo diente es de 6 mm, y con el borde de la nariz externa es de 1 mm.
Narinas internas (choana): Se hallan rodeadas por los pterigoides (71-1), si bien éstos se encuentran desplazados de su posición original. Sus medidas aproximadas serían de 7,5 mm en su eje longitudinal, por 6,5 mm transversal. Se localiza al extremo opuesto de la sutura con el palatino (72 -0). Las narinas internas presentan forma de lágrima, con la parte ancha orientada posteriormente y con un nítido reborde anterior. Su margen anterior y lateral se halla circundado por el pterigoides. El margen posterior, cercano al cóndilo occipital, se aprecia con dificultad debido al desplazamiento de la pieza.
CRÁNEO. HUESOS
Frontal: En posición dorsal es liso y escasamente ornamentado salvo en su parte posterior. No existen crestas en la superficie frontal inter-orbital; también es plano el sector latero-posterior de las órbitas. La distancia longitudinal de la pieza entre la sutura con el nasal y el parietal es de 46,5 mm, por 27 mm de la distancia transversal entre las suturas con los postorbitales. La sutura con el parietal es de tendencia cóncava, si bien se aprecia un vértice intermedio que se adentra en la pieza parietal (86-1). No existe superficie de contacto con la fenestra supratemporal (81-2) por lo que el triple cruce frontal-parietal-postorbital se halla en la caja craneal.
Postorbitales: Relativamente delgados (70-1) y con el borde antero-lateral redondeado. La línea de sutura se aprecia difícilmente. Aparentemente conectaría
l. Alonso sAntiAgo & l. Alonso Andrés
Nuevo género de Crocodylia del Eoceno medio de la Península Ibérica (Zamora, España):Duerosuchus piscator nov. gen., nov. sp.
© Ediciones Universidad de Salamanca Stud. Geol. Salmant., 45 (2), 2009: pp. 149-173
162
con el margen lateral del yugal. La sutura con el escamoso se orienta ventralmente sobre el margen lateral de la caja craneal.
Escamoso: El contorno occipital de la caja craneal presenta forma cóncava, formada por los lóbulos posteriores de los escamosos. El escamoso aparece bastante ornamentado y con superficie plana, salvo en su parte posterior que presenta una zona deprimida, desde la que arranca la proyección posterolateral del escamoso. El borde del lóbulo escamoso se extiende ventrolateralmente hacia la extensión lateral del proceso paraoccipital.
l. Alonso sAntiAgo & l. Alonso Andrés
Nuevo género de Crocodylia del Eoceno medio de la Península Ibérica (Zamora, España):Duerosuchus piscator nov. gen., nov. sp.
© Ediciones Universidad de Salamanca Stud. Geol. Salmant., 45 (2), 2009: pp. 149-173
163
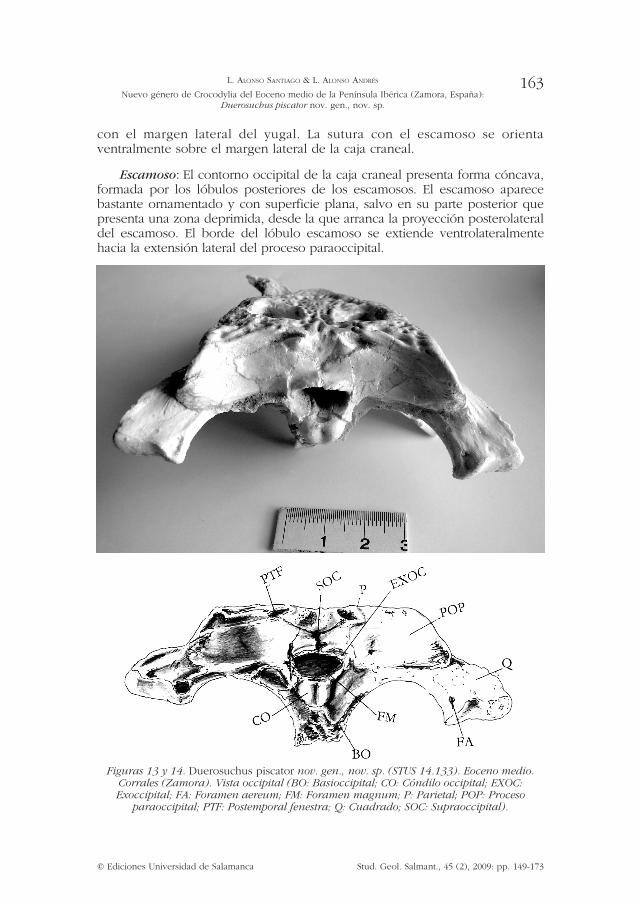
Figuras 13 y 14. Duerosuchus piscator nov. gen., nov. sp. (STUS 14.133). Eoceno medio. Corrales (Zamora). Vista occipital (BO: Basioccipital; CO: Cóndilo occipital; EXOC: Exoccipital; FA: Foramen aereum; FM: Foramen magnum; P: Parietal; POP: Proceso
paraoccipital; PTF: Postemporal fenestra; Q: Cuadrado; SOC: Supraoccipital).

La sutura escamoso-cuadrado se extiende dorsalmente a lo largo del margen posterior del meatus auditivo externo.
El escamoso forma el techo del oído externo, resultando visibles lateralmente las suturas con el exoccipital y el cuadrado. El contacto con el cuadrado tiene lugar en la parte anterior del escamoso y su relación con el exoccipital y cuadrado es visible lateralmente. La sutura con la parte superior del exoccipital discurre hasta el borde lateral del escamoso.
La sutura con el postorbital es rectilínea y se aprecia con dificultad. La participación del escamoso en la parte occipital del cráneo es restringida en comparación con la superficie exoccipital, presentándose en plano inferior al proceso paraoccipital, con suturas muy marcadas, siendo franqueada por las fenestras postemporales.
Parietal: Constituye el anterior, medial y parte del posterior margen de cada abertura supratemporal, apreciándose una superficie plana y ornamentada en el mismo nivel que el resto de la caja craneal. Se proyecta ventralmente formando en su margen anterolateral la pared medial de la fenestra supratemporal.
La línea de sutura con el frontal es convexa, con una inserción central en forma de v. La sutura con los escamosos es rectilínea. Presenta sendas proyecciones anterolaterales que contactan con los postorbitales. La longitud total del hueso parietal es de 27,5 mm; la distancia entre las suturas con los escamosos es de 16 mm. La superficie de contacto con los bordes de la fenestra supratemporal presenta un relieve plano.
Cuadrados: En vista posterior contacta con la región ventrolateral del exoccipital y dorsalmente con la expansión del proceso paraoccipital. No se aprecia el foramen craneocuadrado, que se ubicaría entre el borde interno del cuadrado y el extremo distal del proceso paraoccipital.
La sutura con la base del escamoso se extiende dorsalmente a lo largo del margen posterior del oído externo, compuesto por tres fosas, de 3, 5 y 6 mm de diámetro respectivamente, según la orientación anteroposterior. No se conserva la pieza cuadrato-yugal; la línea de sutura con esta pieza sería suavemente convexa.
Se observa el foramen aereum hacia el borde posterolateral de la superficie dorsal articular. En vista ventral el cuadrado presenta una cresta longitudinal, no especialmente robusta, que lo recorre hacia su extremo distal, dividiéndolo en dos facetas, la lateral de menor tamaño que la medial. Desde su nacimiento posteromedial hasta el fin de su extensión la cresta va decreciendo paulatinamente hasta desaparecer, por lo que la restante extensión ventral del cuadrado presenta una superficie lisa, con suave tendencia cóncava.
l. Alonso sAntiAgo & l. Alonso Andrés
Nuevo género de Crocodylia del Eoceno medio de la Península Ibérica (Zamora, España):Duerosuchus piscator nov. gen., nov. sp.
© Ediciones Universidad de Salamanca Stud. Geol. Salmant., 45 (2), 2009: pp. 149-173
164

El cóndilo del cuadrado presenta un proceso articular ligeramente más saliente que la faceta medial; su tercio interior es cóncavo y liso. La longitud entre los bordes del cóndilo de articulación cráneo-mandibular es de 16 mm, por 7 de alto, que se reducen a 5,8 mm en su tercio medial.
Basioccipital: Formaría el cuerpo del cóndilo occipital y el borde ventral del foramen magnum. El cóndilo presenta en vista posterior forma circular, su forma superior es bilobulada y su cuello corto, proyectándose perpendicular al plano occipital, del que sobresale en total unos 6 mm.
Yugal: La pieza se halló aislada, sin conexión anatómica. La parte posterior del yugal es subcilíndrica y alargada, constituyendo la base de la fenestra infratemporal. Su longitud es de 74 mm, presentando una superficie alveolada. El proceso dorsal del yugal formaría la porción ventral de la barra postorbital, no conservada.
Supraoccipital: Largo y de forma trapezoidal, más ancho que alto, formando en vista posterior una superficie ligeramente convexa con sus bordes dorsales salientes y suturas muy nítidas. Se extiende unos 2 mm más allá del borde dorsal del foramen magnum. Tiene sendas pequeñas crestas verticales acompañadas por grandes forámenes postemporales a cada lado. Contacta con el parietal en sutura rectilínea apenas apreciable. No contacta con el borde superior del foramen magnum, enmarcado por la unión de los exoccipitales.
Pterigoides: Se presenta desplazado de su posición original. No parece que se proyecte de manera destacada sobre la horizontal. Sus dimensiones son de unos 35 mm de distancia ventrolateral por 31 mm en la medida anteroposterior. No se conserva el ectopterygoide. La choana aparece en su superficie ventral localizada al extremo opuesto de la sutura con el palatino, hacia la zona occipital. En el espacio inmediatamente anterior a la choana se aprecia una superficie deprimida que linda con aquélla; esta cavidad está presente en otros crocodyloideos como Prodiplocynodon, Osteolamus o Leydosuchus, si bien su ubicación exacta no es coincidente, así en este último se posiciona lateralmente (véase Wu et al., 2001).
Exoccipitales: El exoccipital izquierdo presenta un ligero desplazamiento por compresión producida en el proceso de fosilización. De superficie lisa. Integran la mayor parte de la superficie occipital del cráneo, contactando a nivel dorsal con el supraoccipital, y laterodorsalmente con escamoso. Enlazan entre sí enmarcando la parte superior del foramen magnum. Se aprecian en distinto plano la superficie del sector superior, que forma el proceso paraoccipital, y la expansión ventrolateral. El extremo distal del proceso paraoccipital presenta una cresta cercana a la ubicación del inapreciable foramen cráneo-cuadrado.
l. Alonso sAntiAgo & l. Alonso Andrés
Nuevo género de Crocodylia del Eoceno medio de la Península Ibérica (Zamora, España):Duerosuchus piscator nov. gen., nov. sp.
© Ediciones Universidad de Salamanca Stud. Geol. Salmant., 45 (2), 2009: pp. 149-173
165

l. Alonso sAntiAgo & l. Alonso Andrés
Nuevo género de Crocodylia del Eoceno medio de la Península Ibérica (Zamora, España):Duerosuchus piscator nov. gen., nov. sp.
© Ediciones Universidad de Salamanca Stud. Geol. Salmant., 45 (2), 2009: pp. 149-173
166
Figura 15. Duerosuchus piscator nov. gen., nov. sp. (STUS 14.133). Eoceno medio. Corrales (Zamora). Osteodermos.
Figura 16. Osteodermos de: a. Asiatosuchus. b. Diplocynodon. c. Iberosuchus.Todos del Eoceno medio de Corrales del Vino (Zamora).
OSTEODERMOS Y VÉRTEBRAS
Atribuimos a Duerosuchus, fundándonos en la comparación con la ornamentación craneal, unos osteodermos de alvéolos separados irregularmente y escasamente pronunciados, que presentan dos morfotipos básicos, unos de tendencia rectangular, y otros de forma oval, apareciendo generalmente una cresta o barra medial que recorre longitudinalmente parte de su superficie dorsal. En los primeros se percibe en su extremo una superficie dorsal estrecha carente de alvéolos (figura 15).
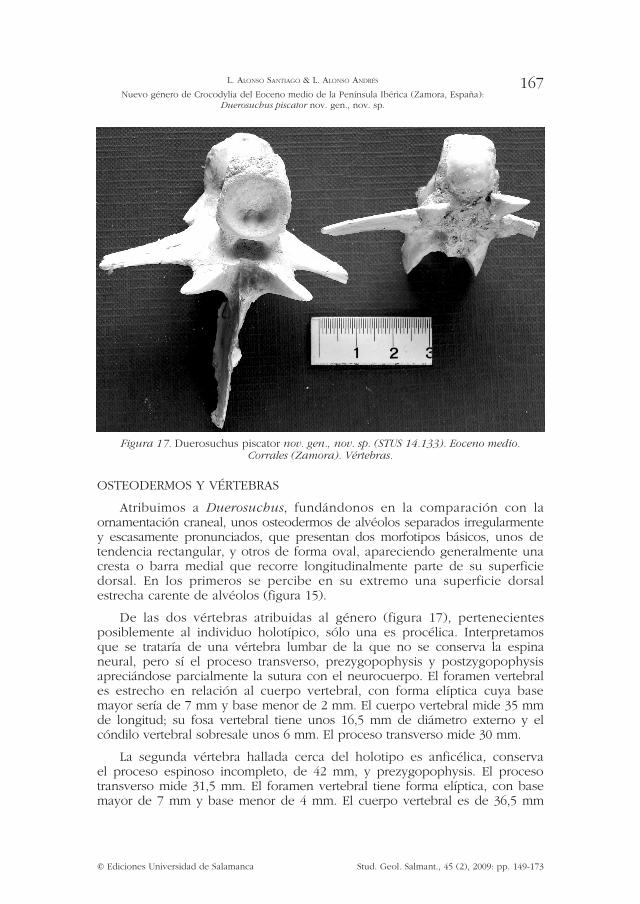
De las dos vértebras atribuidas al género (figura 17), pertenecientes posiblemente al individuo holotípico, sólo una es procélica. Interpretamos que se trataría de una vértebra lumbar de la que no se conserva la espina neural, pero sí el proceso transverso, prezygopophysis y postzygopophysis apreciándose parcialmente la sutura con el neurocuerpo. El foramen vertebral es estrecho en relación al cuerpo vertebral, con forma elíptica cuya base mayor sería de 7 mm y base menor de 2 mm. El cuerpo vertebral mide 35 mm de longitud; su fosa vertebral tiene unos 16,5 mm de diámetro externo y el cóndilo vertebral sobresale unos 6 mm. El proceso transverso mide 30 mm.
La segunda vértebra hallada cerca del holotipo es anficélica, conserva el proceso espinoso incompleto, de 42 mm, y prezygopophysis. El proceso transverso mide 31,5 mm. El foramen vertebral tiene forma elíptica, con base mayor de 7 mm y base menor de 4 mm. El cuerpo vertebral es de 36,5 mm
l. Alonso sAntiAgo & l. Alonso Andrés
Nuevo género de Crocodylia del Eoceno medio de la Península Ibérica (Zamora, España):Duerosuchus piscator nov. gen., nov. sp.
© Ediciones Universidad de Salamanca Stud. Geol. Salmant., 45 (2), 2009: pp. 149-173
167
Figura 17. Duerosuchus piscator nov. gen., nov. sp. (STUS 14.133). Eoceno medio. Corrales (Zamora). Vértebras.

de longitud, su fosa vertebral superior tiene unos 17 mm de diámetro exterior y la fosa inferior es de 18 mm de diámetro. No se aprecia la sutura con el neurocuerpo.
fILOGENIA
Los estudios moleculares de la secuencia evolutiva sitúan estimadamente al ancestro común más reciente del género Crocodylus en una radiación posterior al Mioceno medio, ofreciendo escasas divergencias entre las especies actuales (densmore, 1983; densmore & oWen, 1989; brochu, 2003). Los fósiles más antiguos concuerdan con tales estimaciones, datando su origen sobre el tránsito Mioceno-Plioceno o posterior (brochu, 2000, 2003; delFino et al., 2007; rex, 2006; grigg & gAns, 1993; densmore, 1983; sAlisbury et al., 2006).
El término Crocodylia (Gmelin, 1789) se restringe actualmente al crown group inclusivo del último ancestro común de Gavialoidea y todos los Brevirrostros más sus descendientes (buscAlioni et al., 2001; brochu, 2001b, 2003), si bien algunos autores discrepan en cuanto a la posición de Crocodylia, considerando que Crocodylia + Sphenosuchia representarían la subclase Crocodylomorfa, constituyendo Eusuchia un suborden del orden Crocodylia (mArtin & benton, 2008).
A su vez se considera que el lugar del nodo de Eusuchia referido al grupo inclusivo del último ancestro común de Hylaeochampsa y Crocodylia y todos sus descendientes (brochu, 1999) es inestable, por cuanto aquél depende de la distribución de características en conflicto o pérdidas de los taxones que conforman el grupo Crocodylia.
La transición entre Neosuchia y Eusuchia y la transformación de los miembros de Eusuchia en crocodylianos se considera oscura, por lo incompleto del registro fósil (norell & clArK, 1991; buscAlioni et al., 2001; sAlisbury et al., 2006). En consecuencia, tanto los orígenes de Eusuchia como la subsiguiente aparición de los taxones ancestrales inmediatos de Crocodylia constituyen parcelas insuficientemente conocidas de la evolución (clArK & norell, 1992; sAlisbury et al., 2006).
El hallazgo de Duerosuchus aporta nuevos datos sobre la evolución de los cocodrilianos del Eoceno del viejo continente, aunque el estudio de su posición filogenética presenta el obstáculo de la incompleta conservación del holotipo, que adolece de determinados caracteres considerados esenciales, dificultad a la que se agrega la posibilidad de que algunos parámetros fenotípicos obedezcan a evolución paralela, originada por la respuesta adaptativa a semejantes condiciones ecológicas (iordAnsKi, 1973).
Sin necesidad de reiterar todos los datos ya presentados a lo largo del presente estudio, y basándonos esencialmente en los análisis de brochu (1999, 2003) y de buscAlioni et al. (2001) sobre Eusuchia y su grupo corona
l. Alonso sAntiAgo & l. Alonso Andrés
Nuevo género de Crocodylia del Eoceno medio de la Península Ibérica (Zamora, España):Duerosuchus piscator nov. gen., nov. sp.
© Ediciones Universidad de Salamanca Stud. Geol. Salmant., 45 (2), 2009: pp. 149-173
168

Crocodylia, y el Apéndice al trabajo de sAlisbury et al. (2006), nos centraremos en determinados parámetros fenotípicos. Se destaca que algunos de los osteodermos atribuidos a Duerosuchus presentan una gruesa quilla dorsal, carácter (35-1). Las vértebras atribuidas al género son nítidamente procélicas (18-1). El carácter número 52 referido a los alvéolos dentarios se señala como (1), por cuanto el tercer y cuarto diente se hallan separados, siendo éste de mayor longitud mesio-distal. A su vez se constata la oclusión de la hilera de dientes del dentario por la dentición del maxilar (78-0), y la existencia de una muesca entre maxilar y premaxilar (77-0). A tal respecto, se considera que los modelos de alligator y crocodiliano derivan de una condición ancestral común, y que tanto en los basales alligatoroideos y crocodyloideos, como en los pristichampsinos, el basal Borealosuchus y los outgroups inmediatos a Crocodylia, los dientes inferiores ocluyen lingualmente a la hilera del maxilar, si bien se conserva una muesca entre maxilar y premaxilar para el cuarto diente inferior (brochu, 1999a, 2003). Coexiste en Duerosuchus la hendidura externa del premaxilar, capaz de alojar el cuarto diente del dentario en algún momento del desarrollo del espécimen, con una cavidad interna en la superficie ventral del premaxilar, y un modelo de oclusión de la hilera del dentario semejante a los actuales aligátores.
El carácter n.º 68 se señala como (0), dado que el dentario se presenta lineal entre el 4.º y 10.º alveolos. El diente de mayor tamaño y longitud del maxilar es el quinto, carácter que se asocia a los miembros más avanzados del clado Neosuchia (89-1) (ortegA et al., 2000). El premaxilar tiene 5 dientes (97-0).
El carácter 81 fue señalado como (2) en la anterior descripción, y el número 86, también referido a la sutura frontoparietal, se indica como (1) por presentar forma cóncavo-convexa.
La choana se encuentra rodeada por el pterigoides (71-1), muy desplazada hacia la parte posterior del palatino (72-0). Si bien se admite que la posición de la choana resulta un carácter de conflicto entre los más avanzados neosuchios (buscAlioni et al., 2001). A su vez, se reconoce la gran variabilidad en la morfología de los pterigoides (brAndAlise et al., 2006), aunque la posición y morfología del espécimen estudiado se aparta muy poco de las de los modernos cocodrilianos. Se aprecia una depresión en la superficie del pterigoides, ubicada en la parte inmediata anterior al margen de la choana (73-1).
Se constata a su vez la presencia de una leve fosa de 1 mm en la esquina rostromedial de la fenestra supratemporal, lindando con la superficie del parietal, carácter que actualmente plantea dudas sobre su repercusión en el análisis filogenético. Existe a su vez una fosa elipsoidal de 5 por 2 mm en la pared posterior de la fenestra supratemporal, ubicada entre la sutura del parietal y escamoso, a unos 3 mm de la superficie dorsal de aquéllos.
Se desconoce si el extremo distal del nasal irrumpe en la external naris (95-?), como se representa en la figura 1. Las narinas externas se proyectan
l. Alonso sAntiAgo & l. Alonso Andrés
Nuevo género de Crocodylia del Eoceno medio de la Península Ibérica (Zamora, España):Duerosuchus piscator nov. gen., nov. sp.
© Ediciones Universidad de Salamanca Stud. Geol. Salmant., 45 (2), 2009: pp. 149-173
169

dorsalmente (79-1), y su forma es elipsoidal, con el eje mayor longitudinal de 21 mm, y el eje menor, transversal, de 16 mm (161-1).
El borde anterior del foramen incisivum coincidiría con el segundo alvéolo del premaxilar (153-0). Analizada la forma del borde conservado, se estima que tendría un tamaño reducido (124-0).
Una vez esbozados los caracteres fenotípicos de mayor relevancia en orden a la verificación de su posición filogenética, debe señalarse, a su vez, que la ausencia de indicios del nuevo género en otros depósitos del Eoceno de la Península Ibérica dificulta las conclusiones sobre su distribución cronoestratigráfica. Tampoco encuentra respaldo la hipótesis de que Duerosuchus constituya un endemismo europeo, por cuanto la experiencia demuestra la tendencia a una amplia extensión geográfica de los taxones crocodylianos del inicio del Terciario, no restringidos por las barreras marinas. De hecho, la aparición de endemismos en las faunas crocodylianas se hace especialmente constatable en el Mioceno de Australia y África (brochu, 2003).
A la espera de nuevos hallazgos que permitan un conocimiento más exacto de la morfología del cráneo y aporten claridad tanto sobre las características fenotípicas del nuevo género como acerca del alcance de las posibles variaciones interindividuales, podemos concluir que la fisonomía de Duerosuchus permitiría su consideración como taxón basal de Crocodylia, filogenéticamente cercano a los brevirrostros, anticipando detalles morfológicos que lo aproximan a las especies actuales. Esperamos que el presente descubrimiento contribuya a impulsar futuros estudios en el entorno, que con toda seguridad depararán novedades sobre la fauna eocénica del sector denominado informalmente Corrales B (Alonso & Alonso, 2005).
AGRADECIMIENTOS
Nuestro especial agradecimiento al Dr. Emiliano Jiménez Fuentes, principal referente e impulsor de la paleoqueloniología en España.
A: Carlos Alonso Santiago (abogado), Carlos Revenga Alonso, Alberto Revenga Alonso, Miguel Temprano García (biólogo); Gregorio Panero y Francisco Núñez (vecinos de Corrales); Pedro Gómez y Luis Ángel Díez (Ayuntamiento de Corrales). Asimismo también queremos expresar nuestro agradecimiento por su continua colaboración y las facilidades prestadas a Buddy (DEYME), Paco Mateo (Cerámicas SAZA), Maximiliano San Gregorio y Pablo Criado; a Federico Hernández y al resto de personal de la Cerámica por su constante y amistoso apoyo.
l. Alonso sAntiAgo & l. Alonso Andrés
Nuevo género de Crocodylia del Eoceno medio de la Península Ibérica (Zamora, España):Duerosuchus piscator nov. gen., nov. sp.
© Ediciones Universidad de Salamanca Stud. Geol. Salmant., 45 (2), 2009: pp. 149-173
170

BIBLIOGRAfÍA
Alonso sAntiAgo, l. & Alonso Andrés, l. (2005): Diferencias anatómicas entre dos formas de Allaeochelys del Eoceno de Corrales (Zamora). Nueva especie de tortuga, Allaeochelys jimenezi nov. sp. Studia Geol. Salmant., 41: 103-126.
Alonso sAntiAgo, l.; Alonso Andrés, L. & Jiménez Fuentes, E. (2008): Primer espaldar en conexión anatómica de Allaeochelys casasecai. Cazurra: nuevo yacimiento del Eoceno de Zamora (España). Stud. Geol. Salmant., 44 (2): 177-186.
AndrAde, m. b. de; bertini, r. J. & piAcentini piñeiro, A. e. (2006): Observations on the palate and choanae structures in Mesoeucrocodylia (Archosauria, Crocodylomorpha): Phylogenetic implications. Rev. Bras. Paleont., 9 (3): 323-332.
brochu, C. A. (1999a): Phylogenetics, taxonomy, and historical biogeography of Alligatoroidea. J. Vertebr. Paleontol. Memoir, 6 19: 9-100.
brochu, C. A. (1999b): Taxon sampling and reverse successive weighting. Syst. Biol., 48: 808-13.
brochu, C. A. (2000): Phylogenetic relationships and divergence timing of Crocodylus based on morphology and the fossil record. Copeia, 2000: 657-673.
brochu, C. A. (2001a): Congruence between physiology, phylogenetics, and the fossil record on crocodylian historical biogeography. In: Crocodilian biology and evolution (edits. grigg, g.; seebAcher, F. & FrAnKlin, c. e.). Surrey Beatt & Sons, Sydney, pp. 9-28.
brochu, C. A. (2001b): Crocodylian snouts in space and time: phylogenetic approaches toward adaptive radiation. Am. Zool., 41: 564-585.
brochu, C. A. (2003): Phylogenetic approaches toward crocodylian history. Ann. Rev. Earth Planet. Sci., 31: 357-397.
bucKley, G. A.; brochu, C. A.; KrAuse, d. W. & pol, D. (2000): A bizarre pug-nosed crocodyliform from the Late Cretaceous of Madagascar. Nature, 405: 941-944.
buscAlioni, A. D. (1986): Cocodrilos fósiles del registro español. Tesis Doctoral. Fac. Ci. Univ. Autónoma Madrid. 363 pp. (inédita).
buscAlioni, A. D.; ortegA, F.; WeisAmpel, D. B. & JiAnu, C. M. (2001): A revision of the crocodyliform Allodaposuchus precedens from the Upper Cretaceous of the Hateg Basin, Romania. Its relevance in the Phylogeny of Eusuchia. J. Vertebr. Paleontol., 21: 74-86.
buscAlioni, A. d.; sAnz, J. l. & cAsAnovAs, M. L. (1992): A new species of the eusuchian crocodile Diplocynodon from the Eocene of Spain. N. Jb. Geol. Paläont., Mh. Abh., 187: 1-29.
clArK, J. m. & norell, m. A. (1992): The Early Cretaceous crocodylomorph Hylaeochampsa vectiana from the Wealden of the Isle of Wight. Am. Mus. Novit., 3032: 1-19.
cuestA, M. Á. (1999): Las faunas de mamíferos del Eoceno de la cuenca del Duero (Castilla y León, España). Síntesis bioestratigráfica y biogeográfica. Rev. Española Pal., 14 (2): 203-216.
l. Alonso sAntiAgo & l. Alonso Andrés
Nuevo género de Crocodylia del Eoceno medio de la Península Ibérica (Zamora, España):Duerosuchus piscator nov. gen., nov. sp.
© Ediciones Universidad de Salamanca Stud. Geol. Salmant., 45 (2), 2009: pp. 149-173
171

delFino, M.; bohme, m. & rooK, l. (2007): First European evidence for transcontinental dispersal of Crocodylus (late Neogene of southern Italy). Zool. J. Linnean Soc., 149: 293-307.
densmore, L. D. (1983): Biochemical and immunological systematics of the order Crocodylia. Evol. Biol., 15: 397-465.
densmore, L. D. & oWen, R. D. (1989): Molecular systematics of the order Crocodilia. Am. Zool., 29: 831-841.
grigg, g. & gAns, c. (1993): Morphology and physiology of the Crocodylia. In: Fauna of Australia, vol. 2A: Amphibia and Reptilia. Austr. Govern. Publ. Serv., Canberra, pp. 326-336.
iordAnsKi, N. N. (1973): The skull of the Crocodilians. In: Biology of the Reptilia, vol. 4: Morphology (edits. GAns, c. & pArsons, t. S.). Academic Press, London, pp. 201-262.
Jiménez Fuentes, E. (1992a): Las dataciones del Paleógeno de Castilla y León. In: Vertebrados Fósiles de Castilla-León (coord. Jiménez Fuentes, e.). Museo de Salamanca, pp. 39-41, 2 figs.
Jiménez Fuentes, E. (1992b): Quelonios Fósiles de Castilla y León. In: Vertebrados Fósiles de Castilla-León (coord. Jiménez Fuentes, e.). Museo de Salamanca, pp. 71-100, 26 figs.
Jiménez Fuentes, E. & Alonso Andrés, L. (1993): Nuevos hallazgos de Trionychidae (chelonii) en el Eoceno de Salamanca y Zamora. Consideraciones sobre los ecosistemas fluviales durante el Eoceno en la cuenca del Duero. Stud. Geol. Salmant., 29: 95-113, 6 figs. Salamanca.
Jiménez Fuentes, E.; ortegA, F. J.; gil, s.; Alonso Andrés, l. & Jiménez gArcíA, s. (1991): Excavaciones en Corrales (Zamora) 1990-1991. Notas informativas. Sala de las Tortugas, 15: 4 pp., 3 figs. Salamanca.
Jiménez Fuentes, E.; ortegA, F. J.; gil, s.; mArtín, s. & Alonso Andrés, L. (1990): Excavaciones paleontológicas en la provincia de Zamora realizadas durante 1990. An. Ins. Est. Zamoranos “Florián de Ocampo”, 1990: 17-27, 2 figs. Zamora.
KArl, H.-V. & tichy, G. (2004): The structure of fossil teeth of chelonophagous crocodiles (Diapsida: Crocodylia). Stud. Geol. Salmant., 40: 115-124.
mcAliley, l. r.; Willis, r. e.; rAy, d. A.; White, p. c.; brochu, c. A. & densmore, l. d. (2006): Are crocodiles really monophyletic? Evidence for subdivisions from sequence and morphological data. Molecular Phylogenetics and Evolution, 39: 16-32.
mArtin, J. E. & benton, M. J. (2008): Crown clades in vertebrate nomenclature: Correcting the definition of Crocodylia. Syst. Biol., 57 (1): 173-181.
mArtin, M. A. & clArK, J. M. (1991): A reanalysis of Bernissartia fagesii, with comments on its phylogenetic position and its bearing on the origin and diagnosis of the Eusuchia. Bull. Inst. Nat. Sci. Nat. Belgique, 60: 115-128.
mArtín, S.; Jiménez, e.; FinciAs, b.; prAdo, J. m. del & mulAs, e. (1987): Los Crocodylia del Eoceno y Oligoceno de la cuenca del Duero. Dientes y Osteodermos. Rev. Española Paleont., 2: 95-108, 4 figs., 3 láms.
l. Alonso sAntiAgo & l. Alonso Andrés
Nuevo género de Crocodylia del Eoceno medio de la Península Ibérica (Zamora, España):Duerosuchus piscator nov. gen., nov. sp.
© Ediciones Universidad de Salamanca Stud. Geol. Salmant., 45 (2), 2009: pp. 149-173
172

l. Alonso sAntiAgo & l. Alonso Andrés
Nuevo género de Crocodylia del Eoceno medio de la Península Ibérica (Zamora, España):Duerosuchus piscator nov. gen., nov. sp.
© Ediciones Universidad de Salamanca Stud. Geol. Salmant., 45 (2), 2009: pp. 149-173
173
norell, m. A. & clArK, J. M. (1991): A reanalysis of Bernissartia fagesii, with comments on its phylogenetic position and its bearing on the origin and diagnosis of the Eusuchia. Bull. Inst. R. Sci. Natur. Belgique. Sci. Terre, 60: 115-128.
ortegA, F. J. & buscAlioni, A. D. (1992): Cocodrilos fósiles de Castilla y León. In: Vertebrados Fósiles de Castilla-León (coord. Jiménez Fuentes, e.). Museo de Salamanca, pp. 59-70, 8 figs.
ortegA, F.; buscAlioni, A. D. & gAspArini, Z. (1996): Reinterpretation and new denomination of Atacisaurus crassiproratus (Middle Eocene; Issel, France) as cf. Iberosuchus (Crocodylomorpha, Metasuchia). Geobios, 29: 353-364.
ortegA, F.; gAspArini, Z.; buscAlioni, A. D. & cAlvo, J. O. (2000): A new species of Araripesuchus (Crocodylomorpha, Mesoeucrocodylia) from the lower Cretaceus of Patagonia (Argentina). J. Vert. Paleont., 20 (1): 57-76.
sAlisbury, s. W.; molnAr, r. e.; Frey, e. & Willis, p. m. A. (2006): The origin of the modern crocodylyforms: new evidence from the Cretaceous of Australia. Proc. R. Soc. London, (Ser. B) 273: 2439-2448.
Wu, x.-c.; russell, A. p. & brinKmAn, d. b. (2001): A review of Leydosuchus canadiensis Lambe 1907 (Archosauria: Crocodylia) and an assessment of cranial variation based upon new material. Can. J. Earth Sci., 38: 1665-1687.















![Catálogo de Capacitaciones - · PDF fileIntroducción a IBM i para Usuarios Nuevos 8 Programación Básica en RPG IV 40 ... [As400 / RPG] 40 PMI - Project Management Institute CURSO](https://img.pdfslide.us/doc/110x75/5ab7eb6b7f8b9ac1058c07cd/catlogo-de-capacitaciones-a-ibm-i-para-usuarios-nuevos-8-programacin-bsica-en.jpg)














