Embed Size (px)
Citation preview

Cell, Vol. 6, 395-405, November 1975, Copyright 0 1975 by MIT
Nucleotide Sequence of Salmon Testes and Salmon Liver Cytoplasmic Initiator tRNA
A. M. GillumQ, N. UrquharV, M. Smith*, and U. L. RajBhandary’: Department of Biology” Massachusetts Institute of Technology Cambridge, Massachusetts 02139 and Department of Biochemistry* Faculty of Medicine University of British Columbia Vancouver, B.C., Canada
Summary
Initiator tRNAs from the cytoplasm of salmon testes and salmon liver have been purified. The nucleo- tide sequence of these initiator tRNAs has been determined and found identical to that of initiator tRNA from mammalian cytoplasm. The only dif- ference is the extent of modification of the nucleo- side located between the dihydrouridine and the anticodon stems. In the salmon tRNAs, this modi- fied nucleoside is predominantly N*Nz-dimethyl guanosine, whereas in the mammalian initiator tRNA it is Nz-methyl guanosine.
Introduction
The nucleotide sequence of the first eucaryotic ini- tiator tRNA, that from yeast cytoplasm, was reported recently (Simsek and RajBhandary, 1972). One of the unusual structural features of this tRNA (which distinguishes it from all other tRNAs) is that it lacks the sequence T-#-C-G(A) found so far in all tRNAs active in protein biosynthesis, including the procaryotic initiator tRNAs; instead, it contains the sequence A-U-C-G-. Subsequent studies which led to the total sequence elucidation of three mammalian (Piper and Clark, 1974; Simsek et al., 1974) and the partial sequence of a plant (Ghosh et al., 1974) cytoplasmic initiator tRNA have indicat- ed that the above unique feature of yeast initiator tRNA may be common to all eucaryotic cytoplasmic initiator tRNAs. Besides suggesting an important role of this feature in the function of eucaryotic ini- tiator tRNAs, another interesting outcome of this work is that the total nucleotide sequence of initia- tor tRNAs from three mammalian sources-mouse myeloma, rabbit liver, and sheep mammary gland, which have diverged significantly during evolution- are identical (Piper and Clark, 1974; Simsek et al., 1974).
This finding has prompted us to examine whether the sequence of the mammalian initiator tRNAs has been conserved since an earlier stage in evolution, and whether the mammalian initiator tRNA se- quence is preserved also in primates. This paper describes the results of our work on the purification
and sequence of initiator tRNAs from salmon testes and salmon liver. We show that the sequence of salmon testes and salmon liver initiator tRNA is identical to that of the corresponding mammalian tRNA, except in the extent of modification of a single modified nucleoside. It would thus appear that the structural gene for the nucleotide sequence of cytoplasmic initiator tRNA has remained invariant since the beginning of evolution of the vertebrates.
An accompanying paper describes the purifica- tion and sequence analysis of the initiator tRNA from human placenta (Gillum et al., 1975).
Results
Purification of the Initiator tRNA from Salmon Liver and Salmon Testes For the liver tRNA, this required three steps of coi- umn chromatography, first on BD-cellulose, then on DEAE-Sephadex A-50, and finally on RPC-5. In the case of tRNA from salmon testes, the first two steps of chromatography proved sufficient to obtain the initiator tRNA in a form homogeneous for sequence analysis.
Figure 1 shows the profile obtained upon chro- matography of salmon liver tRNA on BD-cellulose. Assay with the homologous aminoacyl-tRNA syn- thetase yielded three peaks of methionine acceptor activity; only the first of these three peaks could be aminoacylated with methionine using crude E. coli aminoacyl-tRNA synthetase. Since the tRNA in this peak could also be subsequently formylated with the E. coli enzyme, the material in this peak represents the initiator species of methionine tRNA (tRNArMe’) (Lucas-Lenard and Lipmann, 1971). The tRNAiMe’ was then partly separated from the bulk of the nucleotidic material by chromatographing this material on DEAE-Sephadex A-50. This was pooled and used for the final step of chromatog- raphy on RPC-5. The sharp peak of methionine acceptor activity then coincided with the major peak of ultraviolet absorbance, and the patterns of methionine acceptor activity obtained using either
....-.. E coli enzyme --.--Salmon enzyme
Fraction number
Figure 1. Chromatography of Crude Salmon Liver tRNA (1000 AZbo units) on BD-cellulose

Cdl 396
the homologous or the E. coli enzyme could be su- perimposed. The pooled fractions from the RPC-5 column yielded a total of 4.1 AZbO units of pure tRNA+Me+.
Nucleotide Sequences of Initiator tRNAs from Salmon Testes and Salmon Liver General Comments Because the available amounts of these initiator tRNAs were limited, sequence analysis required the use of highly sensitive methods applicable to small amounts of tRNAs. The procedure that we have used involves fhe labeling of 5’ ends of oligonucleo- tides present in T+-RNAase or pancreatic RNAase digests of RNAs with 3*P prior to their sequence analysis (Simsek et al., 1973b, 1974). The 5’ 3*P- labeled oligonucleotides are then separated by two- dimensional electrophoresis (Sanger, Brownlee, and Barrell, 1965). Partial digestion of a 5’ 3*P-labeled oligonucleotide with snake venom phos- phodiesterase yields a series of radioactive prod- ucts ranging in size from the original oligonu- cleotide down to the 5’ terminal mononucleotide. These can be separated by electrophoresis on DEAE-cellulose paper, and the sequence of the 5’- labeled oligonucleotide in question can be derived from the characteristic mobility shifts (M values) re- suiting from the successive removal of nucleotides from the 3’ end (Barrell, 1971). Additionally, in our work the two-dimensional homochromatography system originally described by Sanger et al. (1973) for DNA sequencing proved particularly useful and was in fact essential for establishing the sequence of the larger oligonucleotides. While the methodol- ogy developed for the work will be published in a more detailed paper (M. Silberklang, A. M. Gillum, M. Simsek, and U. L. RajBhandary, manuscript in preparation), the total sequence work was facili- tated to some extent by the fortuitous homology of the sequence to that of a known tRNA, and this combined with the in vitro labeling method allowed us essentially to derive the total sequence of salmon testes and liver initiator tRNAs using a total of lOO- 250 pg of the RNAs. Summary of Evidence for the Sequence The following lines of evidence suggest strongly that the sequence of initiator tRNA from salmon liver and testes is identical to that of mammalian initiator tRNA, except for the extent of modification of a specific G residue.
First, fingerprints of complete pancreatic and T,-RNAase digests of the salmon liver initiator tRNA are identical to the corresponding fingerprints ob- tained from salmon testes initiator tRNA. Since this is a very strong indication that the initiator tRNAs from the two salmon tissues possess identical se-
quences, the detailed results and discussion that follow include only the data from salmon liver.
Second, fingerprints of complete pancreatic RNAase digests of the salmon and rabbit liver initia- tor tRNAs are identical, except that salmon tRNA contains a strong spot due to 3*P-m$G-C and a weak spot of 32P-m2G-C, unlike the tRNA from rab- bit liver which contains 32P-m*G-C. This indicates that a Pyr-m*G-C sequence in rabbit liver is re- placed for the most part by Pyr-m;G-C in salmon liver initiator tRNA.
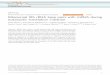
Figures 2A and 2B compare the fingerprints ob- tained respectively from complete pancreatic RNAase digests of the salmon liver and rabbit liver initiator tRNAs. The corresponding spots from each of these fingerprints were analyzed further by 5’ end group analysis and by partial digestion with snake venom phosphodiesterase (Simsek et al., 197313, 1974; Walker and RajBhandary, 1975). The patterns of radioactive spots obtained after partial snake venom phosphodiesterase treatment followed ei- ther by electrophoresis on DEAE-cellulose paper or by two-dimensional homochromatography were identical for each of the corresponding spots. Fig- ure 3 shows a typical example of the use of two- dimensional homochromatography for the se- quence analysis of 3*P-G-G-A-A-G-C, one of the largest oligonucleotides present in the pancreatic RNAase digest, and Table 1 lists the oligonucleo- tides present in the fingerprints of Figures 2A and 2B.
The only difference in the fingerprints of Figures 2A and 28 is the presence of spot 2a in the salmon liver tRNA digest, compared to 2b in the case of rabbit liver tRNA. The results summarized below es- tablish that 2a is 3*P-m;G-C and 2b is 3*P-m*G-C. Thus digestion of spots 2a and 2b with T2-RNAase followed by electrophoresis on Whatman 540 paper at pH 3.5 shows that the radioactive 32P-Np ob- tained from spot 2a had electrophoretic mobility which was faster than that of 3*P-m*Gp present in a similar digest of spot 2b, and identical to a marker of 3*P-m;Gp obtained by T2-RNAase digestion of a known sample of 32P-m2,G-C. Second, 5’ end group analysis of spots 2a and 2b by complete di- gestion with snake venom phosphodiesterase fol- lowed by thin-layer chromatography in solvents 1 and 2 showed that the radioactive QP-N from spots 2a and 2b were clearly separated from each other and that the mobility of the 32P-N from spot 2a was consistent with that expected for pm;G. Third, par- tial snake venom phosphodiesterase digests of spots 2a and 2b followed by DEAE-cellulose paper electrophoresis yielded two radioactive spots in each case, the starting dinucleotide and the 5’ ter- minal mononucleotide (Figure 4). The 3*P-N ob-
Sequence of Salmon Cytoplasmic Initiator tRNA 397
Figure 2. Autoradiographs of 5 ’ SzP-Labeled Oligonucleotides
Oiigonucleotides were obtained from pancreatic RNAase digests of (A) salmon liver tRNA?T (B) rabbit liver tRNAyer; (C) and (0) 5’ and 3’ terminal fragments, respectively, of salmon liver tRNA, Mef after cleavage at m7G (Simsek et al., 1973a). Electrophoresis in the first dimension was on cellulose acetate, and in the second dimension on DEAE-cellulose paper (Sanger et al., 1965). (B) surrounded by dots, blue dye marker. (X) unknown material sometimes seen in fingerprints of pancreatic RNAase digests.

Cell 398
Figure 3. Autoradiograph of Partial Snake Venom Phosphodies- terase Digest on “2P-GGAAGC
Separation was by two-dimensional homochromatography (Sanger et al., 1973). (6) surrounded by dots, blue dye marker. (B) and (P) indicate the location of blue dye and the pink dye in the cellulose acetate electrophoresis in the first dimension.
Figure 4. Autoradiograph of Partial Snake Venom Phosphodies- terase Digests
(A) 32P-mfG-C isolated from yeast tRNAre+; (B) spot 2a of Figure 2A; and (C) spot 2b of Figure 28. The location of ultraviolet ab- sorbing markers of pA and pG are shown. Electrophoresis was on DEAE-cellulose paper at pH 3.5 (Simsek et al., 1974). (B) sur- rounded by dots, blue dye markers. The numbers at the origin indicate the times of incubation in minutes with snake venom phos- phodiesterase.
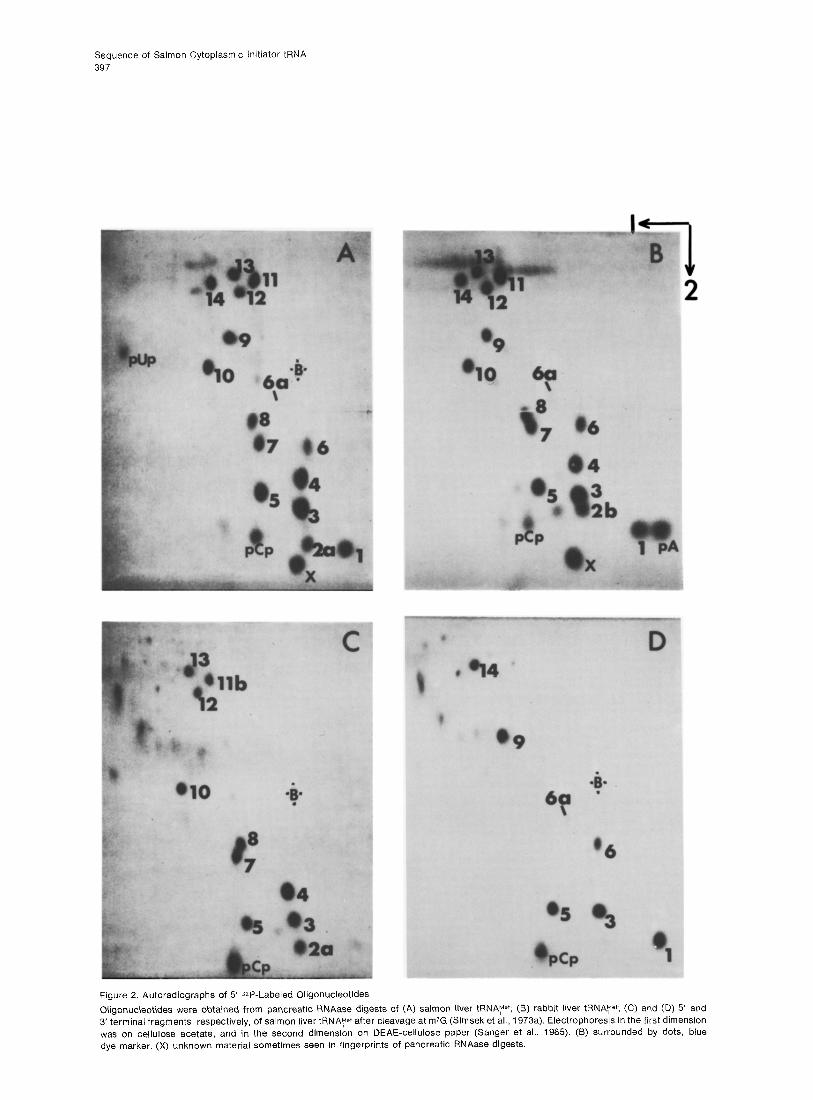
Table 1. Sequence and Molar Yield of Oligonucleotides Present in Fingerprints (Figure 2) of Pancreatic RNAase Digest of Salmon tRNAye+
Fingerprint Sequence of Molar Ratio
spot Oligonucleotide Found0 (Expected)b
1 PAC 0.9 (1)
2a pm;GC 0.66 (1)
2b pm*GC O.llC (0)
3 PGC 2.5 (2) 4 pAGC 2.0 (2) 5 PAU 2.1 (2) 6 pGmrAAAC 0.6
$
(1) 6a pGmbAAACd 0.2
7 pm’GmZGC 1 .o (1)
8 ptbAAC 1 .o (1)
9 pGAU 1.2 (1)
10 PGU 1.2 (1)
Ila pAGAGm7GD
Ilb pGGAAGC I
1.4 VP I
12 pGGGC 0.55 (1)
13 pAGAGU 1 .o (1)
14 pGGAU 1 .o (1)
aBased on Cerenkov counting of fingerprint spots from three separate in vitro labeling reactions. bCalculated from the sequence reported for rabbit liver tRNAyer by Simsek et al. (1974). CRelated to spot 2a. Result of incomplete enzymatic modification of G to m/G in the tRNA (details in text). dProduced by partial conversion of mrA to m&A during digestion of the tRNA and subsequent in vitro labeling of 5’ ends. *These fragments were later separated by one-dimensional chromatography on PEI-cellulose, using 2 M pyridinium formate (pH 3.4) (for details see Gillum et al., 1975).
Sequence of Salmon Cytoplasmic Initiator tRNA 399
tained from spot 2a migrates faster than ‘ZP-m*G present in the digest of spot 2b, and has mobility identical to that of a marker of 3*P-m$G.
Third, the fingerprint of complete T1-RNAase di- gest of the salmon liver tRNA is identical to the cor- responding digest of rabbit liver tRNA, except that the salmon tRNA contains mostly J*P-U-m$G and a weak spot due to 3*P-U-m2G, unlike the case of rabbit liver tRNA which contains 3*P-U-m*G pre- dominantly. This combined with the evidence from the pancreatic RNAase digests suggests that a G-U-m*G-C-sequence in the rabbit liver tRNA is replaced by G-U-m;G-C- in salmon liver tRNA.
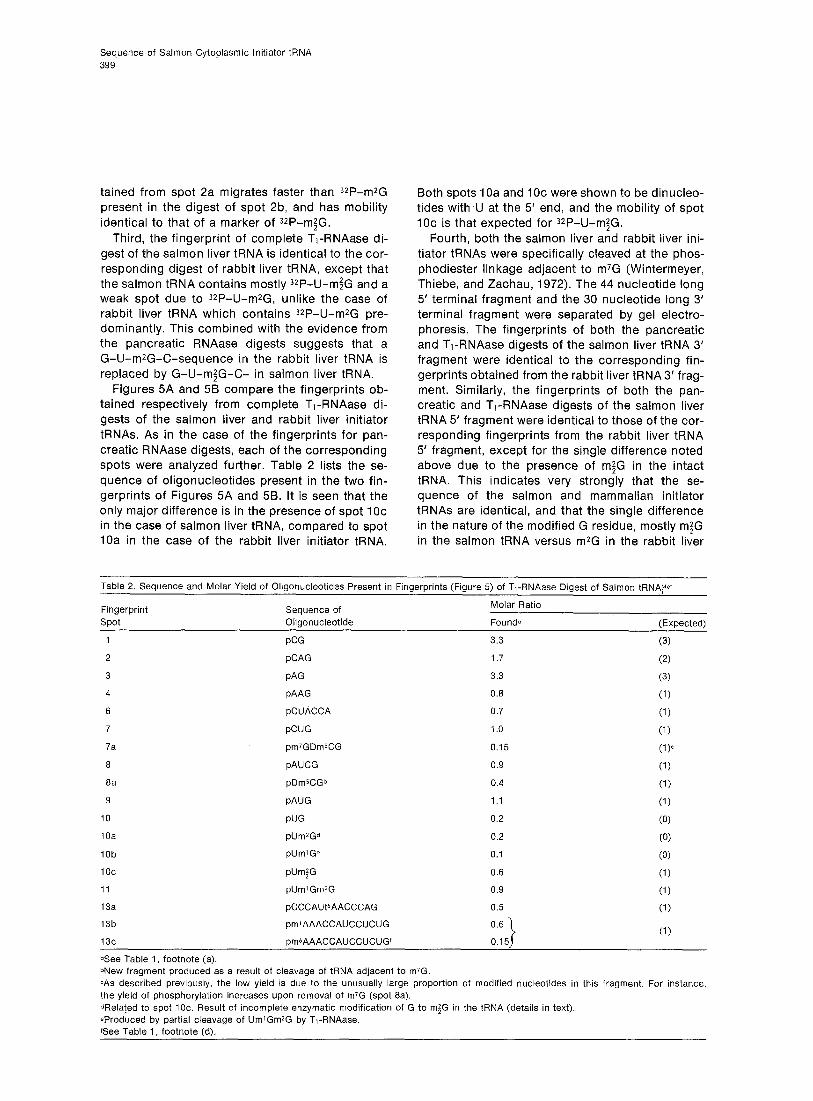
Figures 5A and 58 compare the fingerprints ob- tained respectively from complete T,-RNAase di- gests of the salmon liver and rabbit liver initiator tRNAs. As in the case of the fingerprints for pan- creatic RNAase digests, each of the corresponding spots were analyzed further. Table 2 lists the se- quence of oligonucleotides present in the two fin- gerprints of Figures 5A and 5B. It is seen that the only major difference is in the presence of spot IOc in the case of salmon liver tRNA, compared to spot 10a in the case of the rabbit liver initiator tRNA.
Both spots 1 Oa and 1 Oc were shown to be dinucleo- tides with ,lJ at the 5’ end, and the mobility of spot IOc is that expected for 3*P-U-m;G.
Fourth, both the salmon liver and rabbit liver ini- tiator tRNAs were specifically cleaved at the phos- phodiester linkage adjacent to m7G (Wintermeyer, Thiebe, and Zachau, 1972). The 44 nucleotide long 5’ terminal fragment and the 30 nucleotide long 3’ terminal fragment were separated by gel electro- phoresis. The fingerprints of both the pancreatic and T,-RNAase digests of the salmon liver tRNA 3’ fragment were identical to the corresponding fin- gerprints obtained from the rabbit liver tRNA 3’ frag- ment. Similarly, the fingerprints of both the pan- creatic and T,-RNAase digests of the salmon liver tRNA 5’ fragment were identical to those of the cor- responding fingerprints from the rabbit liver tRNA 5’ fragment, except for the single difference noted above due to the presence of m;G in the intact tRNA. This indicates very strongly that the se- quence of the salmon and mammalian initiator tRNAs are identical, and that the single difference in the nature of the modified G residue, mostly m$G in the salmon tRNA versus m*G in the rabbit liver
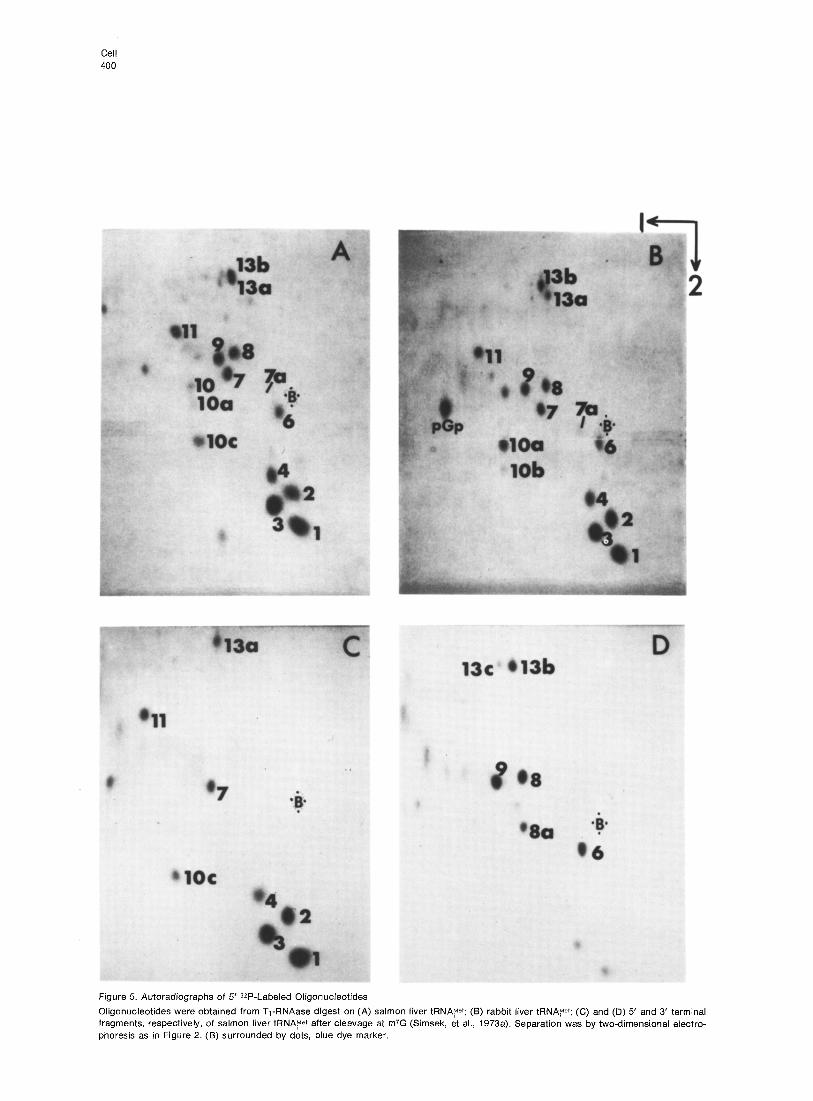
Table 2. Sequence and Molar Yield of Oligonucleotides Present in Fingerprints (Figure 6) of T,-RNAase Digest of Salmon tRNA?et
Fingerprint Sequence of Molar Ratio
spot Oligonucleotide Founda (Expected)
1 PCG 3.3 (3)
2 pCAG 1.7 (2) 3 PAG 3.3 (3)
4 pAAG 0.8 (1)
6 pCUACCA 0.7 (1)
7 pCUG 1 .o (1)
7a pm7GDmSCG 0.15 VP
8 pAUCG 0.9 (1)
8a pDmSCGb 0.4 (1)
9 pAUG 1.1 (1)
10 PUG 0.2 (0)
10a pUm2Gd 0.2 (0)
10b pUmrGe 0.1 (0)
ioc pUm;G 0.6 (1)
11 pUm’Gm2G 0.9 (1)
13a pCCCAUt6AACCCAG 0.5 (1)
13b pmrAAACCAUCCUCUG 0.6 (1)
13c pm6AAACCAUCCUCUGf 1 0.15
aSee Table 1, footnote (a). bNew fragment produced as a result of cleavage of tRNA adjacent to m/G. cAs described previously, the low yield is due to the unusually large proportion of modified nucleotides in this fragment. For instance, the yield of phosphorylation increases upon removal of m7G (spot 8a). dRela!ed to spot 10~. Result of incomplete enzymatic modification of G to m$G in the tRNA (details in text). eProduced by partial cleavage of UmlGmZG by T,-RNAase. See Table 1, footnote (d).
Cell 400
Figure 5. Autoradiographs of 5 ’ 32P-Labeled Oligonucleotides
Oligonucleotides were obtained from T,-RNAase digest on (A) salmon liver tRNAy+; (6) rabbit liver tFlNA~r; (C) and (D) 5’ and 3’ terminal fragments, respectively, of salmon liver tRNA, Mer after cleavage at m’G (Simsek, et al., 1973a). Separation was by two-dimensional electro- phoresis as in Figure 2. (6) surrounded by dots, blue dye marker.

Sequence of Salmon Cytoplasmic Initiator tRNA 401
tRNA in the sequence G-U-m*-G-C-, is located on the 5’ fragment of the tRNA.
Figures 2C and 2D show the fingerprints of pan- creatic RNAase digests of the 5’ and 3’ terminal fragments, respectively. Except for spots 3 and 5, which are present in more than 1 mole per mole of the intact tRNA, the remainder of the spots are uniquely present in the digest of either the 5’ (Fig- ure 2C) or of the 3’ terminal fragment (Figure 2D), but not in both. The oligonucleotides present in Fig- ures 2C and 2D together account for all of the oligonucleotides originally present in pancreatic RNAase digest of the intact tRNA (Figure 2A), ex- cept for A-G-A-G-m7G-D (spot 1 la) which con- tains within it the m7G cleavage site. Although the oligonucleotide generated by this cleavage was not located in these studies, it has been previously ana- lyzed in similar studies of rabbit liver initiator tRNA (Simsek, 1974).
Figures 5C and 5D show the fingerprint of T,- RNAase digests on the 5’ and 3’ terminal fragments, respectively. The oligonucleotides present in these autoradiographs together account for all of the oli- gonucleotides present in a similar digest of the orig- inal tRNA (Figure 5A), except that spot 7a due to QP-m7G-D-m%-G is totally absent and is now re- placed by a new spot 8a due to 32P-D-m5C-G. This finding has been reported previously during studies on the rabbit liver initiator tRNA (Simsek, Petrissant, and RajBhandary, 1973a), and is expected from the removal of m7G residue and cleavage of the phos- phodiester bond adjacent to it in the tRNA (Winter- meyer et al., 1972).
Discussion
The finding that the sequence of cytoplasmic initia- tor tRNA from three diverse mammalian sources was identical (Piper and Clark, 1974; Simsek et al., 1974) prompted us to examine whether this se- quence has been conserved since an earlier stage in evolution. Our choice of salmon tissue for such studies was dictated by the considerations that salmon as a member of the bony fish family repre- sents one of the earlier vertebrates, that salmon liver and testes tissue are readily available, and that preliminary studies on arginine tRNAs of salmon testes were already underway (Urquhart, 1973). Our results demonstrate that the sequence of salmon testes and liver initiator tRNA is also identical to that of the corresponding mammalian tRNA, except in the extent of methylation of a specific guanosine residue (Figure 6). It is therefore possible that the nucleotide sequence of the gene for cytoplasmic initiator tRNAs has remained invariant since the be- ginning of evolution of the vertebrates, some 500 million years ago.
Evidence available to date indicates that the strong conservation of sequence observed above is unique to the initiator tRNAs and that it is not a general feature of all small RNAs. The partial or total sequence of several eucaryotic 5s and 5.8s RNAs are now known. While it appears that all mam- malian 5s RNAs examined have the same sequence (Walker et al., 1975) the 5s RNA sequences of chicken (Pace et al., 1974) and Xenopus (Brownlee et al., 1972) are different from each other and from that of the mammalian 5s RNAs. Although the total sequence information on 5.8s RNAs is limited (Rubin, 1973) these RNAs may also be much less conserved. It has been found that within a single family of flowering plants (Woledge, Corry, and Payne, 1974), there are several differences in the sequence of 5.8s RNAs. Whether the sequence of non-initiator tRNAs is also very strongly conserved among the vertebrates is not known at the moment, since except for avian tRNATrp, the only non-initiator tRNAs from the vertebrates whose sequences are known are from mammalian sources. It is interest- ing to note, however, that the sequence of avian tRNArrp (Harada, Sawyer and Dahlberg, 1975) is identical to that of beef liver tRNATI-p (G. Dirheimer, personal communication). This combined with re- cent findings of identical sequence between corre- sponding aminoacid acceptor RNAs from diverse mammalian sources (Keith, Ebel, and Dirheimer,
A OH C
cGA d: m2 G
G CGCG
GAA G C G U,
G +G flC
cu G
CG C
U AA
‘CUACC $l
GAUGGA cG #CD U
A c”G
ii C C C A ?A
C AU
Figure 6. The Nucleotide Sequence of Salmon Liver, Salmon Testes, and Mammalian Cytoplasmic tFiNA”et
The salmon tRNA contains predominantly m$G in the interstem region between the D and the anticodon stems as indicated by the arrow.

Cell 402
1974; B. A. Roe, M. P. J. S. Anandraj, L. S. Y. Chia, E. Randerath, R. C. Gupta, and K. Randerath, in press) would suggest that even the sequence of non-initiator tRNAs may be more conserved than previously assumed on the basis of data available from tRNAs of yeast and E. coli (Holmquist, Jukes, and Pangburn, 1973).
That initiator tRNAs in general have been more strongly conserved throughout can be concluded from a comparison of the known initiator tRNA se- quences of procaryotes and eucaryotes. These now include initiator tRNAs from E. coli (Dube et al., 1968),blue-greenalgae(R.Cedergren,personalcom- munication), Thermus thermophilus (S. Nishimura, personal communication), B. subtilis (Yamada and Ishikura, 1975), yeast(Simsek and RajBhandary, 1972), salmon, rabbit liver (Simsek et al., 1974), sheep mammary gland (Simsek et al., 1974), mouse myeloma (Piper and Clark, 1974), and human pla- centa (Gillum et al., 1975). In a pairwise comparison of the sequences of initiator tRNAs covering this wide evolutionary span of some 3 x 109 years, the maximum percentage nucleotide difference be- tween any two initiator tRNAs is only of the order of 28%. In contrast, the initiator and non-initiator species of methionine tRNA isolated from the same organism (either in the case of E. coli or of rabbit liver) or the non-initiator methionine tRNAs from dif- ferent organisms (Barrel1 and Clark, 1974) differ in about 44% of the total nucleotides present. This strong conservation in the sequence of initiator tRNAs compared to non-initiator tRNAs is most probably a reflection of the highly specialized func- tion of the initiator species of tRNAs.
In view of the possibility that the cytoplasmic ini- tiator tRNA sequence may have remained fixed since the beginning of evolution of vertebrates, it would be desirable now to retrace the evolution of this tRNA still further and to examine whether the sequence may have been conserved even earlier. The methodology that has been developed in this and previous work is expected to be particularly useful for this purpose. First, it has been possible to purify initiator tRNAs on a scale much smaller than used previously in tRNA purifications. Hitherto, a limiting factor in the undertaking of sequence analysis of any tRNA from a potentially interesting source has been the availability of large amounts of crude tRNA necessary for the purification step. It is of considerable technical importance in the pu- rification of initiator tRNAs that all initiator tRNAs accept methionine, and that 35S-methionine of very high specific activity is commercially available. Sec- ond, the in vitro labeling procedure (Simsek et al., 1973b) we have used is particularly useful for com- paring the extent of homology of nucleotide sequence between two tRNAs (Figures 2A and 28,
5A and 56) and can be carried out on as little as 0.5 pg of an RNA (M. Silberklang, unpublished results).
The only difference between the sequence of salmon testes and salmon liver initiator tRNA on one hand and mammalian initiator tRNA on the other is that the salmon tRNA contains predomi- nantly m;G at a site occupied mostly by m*G in rab- bit liver, sheep mammary gland, or mouse myeloma initiator tRNA. Initially, our main reason for investi- gating the salmon liver tRNA was based on prelimi- nary indications from the fingerprints of salmon testes tRNA that this tRNA may contain m;G in place of m2G. Since both liver and testes tissues contain m$G, the presence of m$G in place of m2G in eucaryotic cytoplasmic initiator tRNAs is proba- bly not a reflection of their tissue of origin.
Finally, besides their homology of sequence with mammalian initiator tRNAs, salmon testes and liver initiator tRNAs provide yet another example of the unusual sequence in loop IV common to several eu- caryotic cytoplasmic initiator tRNAs. Thus yeast (Simsek and RajBhandary, 1972), Neurospora (A. M. Gillum, L. Hecker, W. E. Barnett, and U. L. RajBhan- dary, unpublished results), wheat germ (Ghosh et al., 1974), salmon testes and liver, rabbit liver (Sim- sek et al., 1974), sheep mammary gland (Simsek et al., 1974), mouse myeloma (Piper and Clark, 1974) and human placenta (Gillum et al., 1975), ini- tiator tRNAs all have been shown to lack the se- quence T-J/-C-G- and to contain the sequence A-U (or U’s)-C-G instead. This strong conservation of a feature unique only to eucaryotic cytoplasmic initiator tRNAs implies an essential role for this in their function. Studies to be published elsewhere (J. Heckman, G. Temple, N. Gupta, C. Woodley, and U. L. RajBhandary, manuscript in preparation) indi- cate that one important role of this sequence may be in the initiation step of eucaryotic protein synthe- sis Whether the presence of this sequence addi- tionally prevents initiator tRNAs from binding to the A site of the ribosome and hence from inserting methionine internally is another possibility, and re- mains to be examined further.
Experimental Procedures
General BD-cellulose was purchased from Boehringer; DEAE-Sephadex A-50 from Pharmacia; cellulose acetate strips (2.5 x 55 cm) and glass backed PEI-cellulose plates (20 x 20 cm) from Schleicher and Schuell; glass-backed cellulose thin-layer plates without fluo- rescent indicator from Brinkman; and Whatman DEAE-cellulose paper sheets or rolls were from Reeve Angel and Co. DEAE thin- layer plates for homochromatography were either prepared as de- scribed by Agarwal et al. (1975) or obtained commercially from Analtech. RPC-5 resin was made according to Kelmers and Heatherly (1971), and provided by Joyce Heckman.
Triethylammonium bicarbonate was prepared by bubbling CO? into triethylamine which had been distilled previously. Markers of

Sequence of Salmon Cytoplasmic Initiator tRNA 403
NJ-methyl adenosine 5’ phosphate and N7-methyl guanosine 5’ phosphate were obtained from Terra Marine Bioresearch; pGp was a gift of Dr. Hans J. Gross; the source of pAp, pCp, and pup was as described previously (Simsek et al., 1973b). Crude yeast RNA used for the preparation of homochromatography solutions was from Sigma, and ultra-pure urea used for gel electrophoresis was from Schwartz-Mann. For the detection of ultraviolet absorbing bands on slab gels by direct visualization under an ultraviolet lamp, the acrylamide used for gel electrophoresis was recrystallized from chloroform.
Gel Electrophoresis Gel electrophoresis under denaturing conditions was carried out on 15% polyacrylamide gel slabs (17 x 17 x 0.2 cm) containing 0.75% bisacrylamide and 6 M urea. Running buffer was 90 mM Tris-borate (pH 8.3) containing 4 mM EDTA (Simsek et al., 1973a). The sample (usually 15 pl) was heated for 1 min at 1OOOC in the presence of 5-6 M urea and then applied. Electrophoresis was at 350 volts for 14 hr at 4°C (or until xylene cyanol tracking dye had migrated about 9 cm). Analytical slots containing 0.2 AZbO units of tRNA or fragments were visualized by staining with toluidine blue. Preparative slots containing 1.5-2.5 Alba units of tRNA or fragments could be visualized directly under a Mineralight ultravio- let lamp when recrystallized acrylamide was used.
tRNA or fragments of tRNA were extracted from the gel pieces by homogenization in 1-2 ml of 0.3 M NaCI, 30 mM sodium citrate (pH 8.0) in the absence of any added carrier, and were worked up as before (Walker and RajBhandary, 1975).
Radioactive Material Carrier-free QP-phosphoric acid, 3H-met (5-11 Ci/mmole), and 35S-met (50-300 Ci/mmole) were all obtained from New England Nuclear Corporation. y-32P-ATP was prepared according to Glynn and Chappell (1964) up to a specific activity of 350 CVmmole, and purified by chromatography on DEAE-Sephadex A-25.
Chromatographic Solvents The following solvent systems were used for thin-layer chroma- tography: first, isobutyric acid-cont. NH40H-Hz0 (66:1:33 v/v) (pH 3.7); second, 0.1 M sodium phosphate (pH 6.8) (100 ml), (NH&SO4 (60 g), and n-propanol (2 ml); third, n-propyl alcohol- cont. NH40H-Hz0 (55:10:35 v/v); fourth, isobutyric acid-cont. NHdOH-Hz0 (557:38:385 v/v); and fifth, t-butanol-cont. HCI-Hz0 (70:15:15, v/v).
Enzymes The sources of pancreatic RNAase, purified T,-RNAase, T2- RNAase, and snake venom phosphodiesterase have all been de- scribed previously (Simsek et al., 1973b; Walker and RajBhandary, 1975). Snake venom phosphodiesterase was stored at 4°C as a 1 mg/ml solution in 100 mM Tris-HCI and IO mM potassium phos- phate at pH 8.6. E. coli alkaline phosphatase and yeast hexokinase were purchased from Boehringer Mannheim Corp.; the E. coli alka- line phosphatase was stored frozen at -20°C at a concentration of 2.5-10 U/ml, and hexokinase was stored at -20°C in 50% glycerol, 5 mM sodium acetate (pH 5.0) at a concentration of 2.1 U/ml. An assay after 5 weeks showed negligible loss of activity of hexokinase when stored as above. T4-polynucleotide kinase was isolated from E. coli infected with Tq amN82 and purified according to Panet et al. (1973), and stored at -20°C in 50% glycerol at a concentration of 5 U/PI.
Crude E. coli aminoacyl-tRNA synthetases which also contained met-tRNA formylase activity were prepared from E. coli B according to Muench and Berg (1966). Crude salmon liver aminoacyl-tRNA synthetases were prepared by adaptation of procedures of Rosen, Murray, and Novelli (1967) for tissue homogenization and centrifu- gation, and then chromatographed on DEAE-cellulose as described by Muench and Berg (1966). The final enzyme preparation (14 mg/ ml) was stored at -20°C in 50% glycerol, 10 mM potassium phos-
phate (pH 7.4), 1 mM MgClz, and 10 mM mercaptoethanol. The total yield of crude enzyme from 25 g of frozen liver was about 140 mg protein.
Assays for Aminoacylation E. co/i Enzyme Incubation mixture (0.12 ml) contained 50 mM Tris-HCI (pH 7.5), 15 mM MgCIz, 2.5 mM ATP, 0.5-l .5 nmole of 3H- or 35S-methio- nine, tRNA, and 0.02-0.04 mg of crude E. coli aminoacyl-tRNA synthetases. After incubation at 37°C for IO-15 min, an aliquot (100 ,LLI) was withdrawn and used for assay of acid-precipitable radioactivity using the filter paper technique (Hoskinson and Khorana, 1965). Salmon Liver Enzyme The incubation mixture was identical to that described above, ex- cept that MgClz was IO mM, and the incubation mixture contained in addition 10 mM P-mercaptoethanol, 80 mM NH4CI, and 0.07 mg crude salmon liver aminoacyl-tRNA synthetase instead of the E. coli enzyme. Incubation was at 25°C for 20-30 min.
Assay for the Extent of Formylation of Salmon Testes Initiator tRNA This was carried out as before (Walker and RajBhandary, 1975). The purified tRNA was aminoacylated and formylated in the pres- ence of 35S-methionine and Nra-formyltetrahydrofolate (100 pg/ ml) using E. coli aminoacyl-tRNA synthetases. After phenol extrac- tion, 200,000 cpm of the isolated fmet-tRNA was treated with 0.1 N NaOH at 37°C for 15 min. The radioactive products were separat- ed by electrophoresis on Whatman 3 MM paper at either pH 7.5 (30 mM phosphate buffer) or at pH 3.5 (pyridinium acetate buffer). The extent of formylation for salmon testes tRNAMer was 93%.
Isolation of Salmon Testes RNA This was carried out essentially as before (Urquhart, 1973). Frozen salmon tissue (220 g) was pulverized into small chunks and was left at 4°C until the tissue was slightly soft. Phenol extraction was performed in a Waring blender with 150 ml extraction buffer [O.l M sodium acetate (pH 4.7) 0.14 M NaCI, and 5 mM EDTA]. and 150 ml phenol equilibrated with 0.1 M sodium acetate (pH 4.7) per 110 g of salmon testes tissue. The mixture was homogenized in a Waring blender for 60 set at 4°C and then centrifuged. The aqueous and phenol layers were separated, and were each back- extracted once with phenol and extraction buffer, respectively. Use of pH 4.7 during phenol extraction of testes tissue minimizes DNA contamination of crude tRNA, which can otherwise be extensive. Ethanol (2.5 vol) was added to the combined aqueous layers to precipitate RNA, and then left at 4°C. The ethanol precipitate was dissolved in 25 ml water and dialyzed against IO mM Tris (pH 8.0). The 4400 Alao units were then applied on a column (2.2 x 30 cm) of DEAE-cellulose previously equilibrated with IO mM Tris (pH 8.0) at 4°C. The column was eluted with a linear gradient (total volume, 3 I) of NaCl [0 -1.5 M in 10 mM Tris (pH 8.0)]. The tRNA eluted at about 0.5 M NaCI; total yield of crude tRNA was 1360 Azao units.
Isolation of Salmon Liver tRNA This was carried out by minor modifications of the method used for mammalian liver by Rogg, Wehrli, and Staehelin (1969). Frozen salmon liver (200 g) was pulverized and then homogenized in a Waring blender with 150 ml of 0.1 M Tris-HCI (pH 7.5) containing 0.1% bentonite, 45 ml of 1 M sucrose, and 150 ml of phenol equili- brated with 0.1 M Tris-HCI (pH 7.5) and 0.1% in 8-hydroxy quino- line per 100 g of liver tissue. After centrifugation to separate the aqueous and phenol layers, the total nucleic acid was recovered from the aqueous layer by ethanol precipitation, and then partially purified by batchwise absorption to and subsequent elution from DEAE-cellulose. The crude tRNA (2260 AzbO units) was further freed from DNA and rRNA by fractional precipitation from isopropyl alco- hol. Final yield of crude tRNA was 1460 Azao units.

Cell 404
Purification of Salmon Liver and Salmon Testes Cytoplasmic Initiator tRNAs Small amounts of crude salmon testes and salmon liver tRNAs were incubated at 37’C with 1 M Tris-HCI (pH 8.8) and then as- sayed for methionine acceptor activity. No increase in the total methionine acceptance of the tRNAs was observed. Consequently, the bulk of the crude tRNAs was not subjected to the usual deacyla- tion with Tris-HCI prior to the purification steps.
Chromatography on BD-Cellulose Crude salmon liver tRNA (1000 A 260 units) was applied on a column (0.9 x 90 cm) of BD-cellulose equilibrated with 0.4 M NaCI, IO mM MgC12, and 2% (v/v) dimethylformamide (Urquhart, 1973). The column was then eluted with a linear gradient (0.4 + 1 .I M of NaCI; total volume of gradient, 2.11) containing 10 mM MgC12 and 2% dimethylformamide. Fractions (5 ml) were collected every 5 min. Aliquots (IO pl) were used for the assay of methionine acceptor activity using either salmon liver or E. coli aminoacyl-tRNA synthe- tases. Fractions 50-61 (Figure 1) were pooled to obtain 93 AZbO units of partially purified cytoplasmic initiator tRNA (tRNA?t).
Chromatography on DEAE-Sephadex A-50 Salmon liver tRNAp (93 A*&0 units) was applied to a column (0.8 x 22 cm) of DEAE-Sephadex A-50 previously equilibrated with 0.325 M NaCl in 10 mM MgClz and 20 mM Tris-HCI (pH 7.5), and the column was eluted with a linear gradient (0.325 - 0.41 M; total volume of gradient, 1.1 I) of NaCl in 10 mM MgC12 and 20 mM Tris-HCI (pH 7.5). Fractions (4 ml) were collected every 12 min. Fractions 64-93 were pooled to obtain 18.6 Azao of salmon liver tRNAW.
Chromatography on RPC-5 Salmon liver tRNAye+ isolated from the DEAE-Sephadex A-50 col- umn was applied to a column (0.3 x 60 cm) of RPC-5 equilibrated with 0.4 M NaCl in IO mM MgC12 and IO mM sodium acetate (pH 5.3). The column was eluted at 300 psi with a linear gradient (0.4 - 0.67 M; total volume, 320 ml) of NaCl in IO mM MgC12 and IO mM sodium acetate (pH 5.3). Fractions (1.5 ml) were collected every 5 min. Fractions 78-84 were pooled to yield 4.1 AZbO units of pure salmon liver tRNA+.
Purification of salmon testes initiator tRNA was essentially simi- lar, except that 750 Azao units of crude tRNA were used and 6.1 AzbO units of homogeneous testes tRNAye+ were obtained after the first two steps of column chromatography.
Solutions for Homochromatography These were prepared as described by Jay et al. (1974), except that the pH of the final homomix was adjusted to 4.7 with acetic acid.
5’ End Group Labeling of tRNA Fragments with 3*P The procedure for complete T,- or pancreatic RNAase digestion and subsequent labeling of the oligonucleotide fragments present in such digests was essentially the same as before (Simsek et al., 1973b), with the following modifications: initial digestion of the tRNA was scaled down to a total volume of 20 ~1, and the incubation mixture contained 5 pg tRNA; the scale of the in vitro labeling reaction was reduced to contain 0.5 pg of digested tRNA in a total incubation of 10 ~1; the specific activity of y-3*P-ATP used was about 100 Ci/mmole; and the amount of hexokinase used to de- stroy the excess of ATP was reduced to 0.002-0.003 unit. The labeled fragments were then separated by two-dimension electro- phoresis (Sanger et al., 1965). The first-dimensional electrophore- sis on cellulose acetate was run in a tank prewarmed to 30-35°C rather than a cooled tank, since this procedure was found to give sharp and compact spots.
5’ End Group Analysis of 32P-Labeled Fragments The radioactive spots from DEAE-cellulose paper were eluted with 2 M triethylammonium bicarbonate (pH 9) into Eppendorf polypropylene tubes. The eluate was evaporated to dryness, and evaporation was repeated several times with addition of water to remove all traces of the eluant. For fragments longer than 4 nucleo- tides, crude yeast tRNA (0.5-I .O A zbO unit) was added to the tubes prior to evaporation to avoid losses due to adsorption to the polypropylene tubes.
Two different methods were used for the 5’ end group analyses (Simsek et al., 1973b; Simsek, 1974). The first involves treatment of the 5’ labeled radioactive oligonucleotide with T&NAase and characterization of the S*P-Np thus produced. The second method involves complete digestion of the 5’ end labeled oligonucleotide with snake venom phosphodiesterase and characterization of the 32P 5’ mononucleotide thus produced.
The incubation mixture (5 ~1) for T,-RNAase digestion contained 8000-l 0,000 cpm of the radioactive fragment, 25 mM ammonium acetate (pH 4.6), and 0.5-I .O unit of T2-RNAase. Incubation was at 37°C for 2 hr. The mixture was diluted with 10 ~1 of water and then lyophilized. The residue was dissolved in 2 ~1 of a marker solution containing 0.05 AZ&,, unit per ~1 of pAp, pCp, and pup. An aliquot (1 ~1) was then applied on a thin-layer cellulose plate and chromatographed in solvent 1 for 6-8 hr at 25°C. The nonra- dioactive markers were clearly separated and located under a Min- eralight ultraviolet lamp, and the 32P-labeled products were located by autoradiography for 12-I 5 hr.
The incubation mixture (3 ~1) for complete digestion with snake venom phosphodiesterase contained 6000-10,000 cpm of radioac- tive fragment, 50 mM Tris-HCI and 5 mM potassium phosphate at pH 8.6, and 0.1-0.2 pg of snake venom phosphodiesterase per pmole of the oligonucleotide. Incubation was at 37°C for 1 hr. The mixture was evaporated to dryness and then dissolved in 3 ~1 of a marker solution containing 0.05 AZbO unit per pl of pA, pG, pU, PC, pmlA, and pm7G. Aliquots (1 ~1) were spotted onto two cellulose plates and chromatographed in solvents 1 and 2.
Partial Digestion of 5’ End-Labeled Fragments with Snake Venom Phosphodiesterase The incubation mixture (50 pl) usually contained about 100,000 Cerenkov cpm of the 5’ labeled fragment, 20 mM Tris-HCI and 2 mM potassium phosphate (pH 8.6), 0.1-1.0 AZbO unit of carrier yeast tRNA, and snake venom phosphodiesterase to give an en- zyme to carrier RNA ratio of IO-20 pg enzyme per Anao unit RNA. In a few cases when the presence of some modified nucleotides slowed down the digestion by snake venom phosphodiesterase, it was necessary to increase the enzyme to carrier RNA ratio to 50 pg enzyme per AZbO unit RNA. Incubation was at room tempera- ture or at 37°C. Aliquots (5 ~1) were removed at 0, 1, 2, 5, IO, 20, 40, 80, and 160 min; the enzyme was inactivated by heating at 100°C for 1 min, and the digests were stored at -20°C until use. To test the extent of digestion, 0.5 pl of the various timed aliquots were applied on DEAE-cellulose plates, subjected to ho- mochromatography (Sanger et al., 1973), and the pattern of diges- tion followed by autoradiography. The appropriate timed aliquots were then pooled to yield an even display of all the intermediates produced in the partial digest. The oligonucleotides present in the partial digest were separated and identified by either one or both of the systems described below
One-dimensional electrophoresis on DE-81 paper at pH 3.5 (pyri- dinium acetate buffer) of selected pooled aliquots along with a control sample of undigested fragment has been described pre- viously(Barrell, 1971; Simsek et al., 1973b). Assignment of oligonu- cleotide sequences was based on M values (Barrell, 1971) and/or direct comparison of the pattern obtained from salmon liver frag- ment with that of the corresponding fragment from mammalian initiator tRNA.

Sequence of Salmon Cytoplasmic Initiator tRNA 405
Two-dimensional analysis of partial digest of longer oligonucleo- tides was based on a method previously described by Sanger et al. (1973) for DNA sequencing. The first dimension is electrophore- sis on cellulose acetate strips at pH 3.5. The removal of C, A, U, or G from an oligonucleotide results in characteristic speeding up or slowing down of the oligonucleotide which is now one nucleo- tide shorter. The radioactive oligonucleotides are then transferred onto a DEAE-cellulose thin-layer plate for homochromatography at pH 4.7 in the second dimension. Separation in the second dimen- sion is based mostly on chain lengths (generally, the vertical dis- tance between two oligonucleotides which differ by a purine nu- cleotide is larger than those which differ by a pyrimidine nucleotide). Thus the stepwise removal of nucleotides can be read as angular “jumps” between successively shorter and shorter frag- ments. In the special cases where “unusual” modified nucleotides were expected to grossly alter the type of jump (for example, mrA, m7G, tbA, modified nucleotides which carry a charge on the ring), comparison of the salmon liver tRNA fragment with that generated from the corresponding mammalian initiator tRNA fragment was used to confirm the sequence of the salmon tRNA fragment.
Cleavage of IRNA Adjacent to m’G Residue This was performed as described previously (Simsek et al., 1973a; Simsek, 1974) using 3.5 Azao units of salmon liver tRNA?et. The products were separated by gel electrophoresis and recovered as described above.
Acknowledgments
This work was supported by a grant from the USPHS and grants from the American Cancer Society to U. L. R. and the Medical Research Council of Canada to M. S. A. M. G. was supported by a USPHS training grant.
Received August 21, 1975
Agarwal, K. A., Berlin, Y., Kleid, D., Smirnov, V., and Khorana, H. G. (1975). J. Biol. Chem. 250, 5563-5573.
Barrell, B. G. (1971). In Procedures in Nucleic Acids Research, 2, G. L. Cantoni and D. R. Davies, eds. (New York: Harper and Row), pp. 751-779.
Barrell, B. G. and Clark, B. F. C. (1974). Handbook of Nucleic Acid Sequences (Oxford, England: Joynson-Bruvers Ltd.),
Brownlee, G. G., Cartwright, E., McShane, T., and Williamson, R. (1972). FEBS Letters 25, 8-12.
Dube, S. K., Marcker, K. A., Clark, B. F. C., and Cory, S. (1968). Nature 278, 232-234.
Ghosh, K., Ghosh, H. P., Simsek, M., and RajBhandary, U. L. (1974). J. Biol. Chem. 249, 4720-4729.
Gillum, A. M., Anandaraj, M. P. J. S., Roe, B, A., and RajBhandary, U.L. (1975). Cell 6, 407-413.
Glynn, I. M., and Chappell, J. B. (1964). Biochem. J. 90, 147-149.
Harada, F., Sawyer, R. C., and Dahlberg, J. E. (1975). J. Biol. Chem. 250, 3487-3497.
Holmquist, R., Jukes, T. H., and Pangburn, S. (1973). J. Mol. Biol. 78, 91-116.
Hoskinson, R. M., and Khorana, H. G. (1965). J. Biol. Chem. 240, 2129-2134.
Jay, E., Bambara, R., Padmanabhan, R., and Wu, R. (1974). Nucleic Acids Res. 7, 331-353.
Keith, G., Ebel, J. P., and Dirheimer, G. (1974). FEBS Letters 48, 50-52.
Kelmers, A. D., and Heatherly, D. E. (1971). Anal. Biochem. 44, 486-495.
Lucas-Lenard, J., and Lipmann, F. (1971). Annu. Rev. Biochem. 40, 409-448.
Muench, K., and Berg, P. (1966). In Procedures in Nucleic Acids Research, 1, G. L. Cantoni and D. R. Davies, eds. (New York: Harper and Row), pp. 375-383.
Pace, N. R., Walker, T. A., Pace, B., and Erickson, R. L. (1974). J. Mol. Evol. 3, 151-159.
Panet, A., van de Sande, J. H., Loewen, P. C., Khorana, H. G., Paae, A. J., Lillehaug, J. R., and Kleppe, K. (1973). Biochemistry 72, 5045-5050.
Piper, P. W., and Clark, B. F. C. (1974). Nature 247, 516-518.
Roe, B. A., Anandaraj, M. P. J. S., Chia, L. S. Y., Randerath, E., Gupta, R. C., and Randerath, K. (1975). Biochem. Biophys. Res. Commun., in press.
Rogg, H., Wehrli, W., and Staehelin, M. (1969). Biochim. Biophys. Acta 795, 13-22.
Rosen, L., Murray, E. L., and Novelli, G. D. (1967). Can. J. Biochem. 45, 2005-2014.
Rubin, G. M. (1973). J. Biol. Chem., 248, 3860-3875.
Sanger, F., Brownlee, G. G., and Barrell, B. G. (1965). J. Mol. Biol. 13, 373-389.
Sanger, F., Donelson, J. E., Carlson, A. R., Kossel, H., and Fischer, D. (1973). Proc. Nat. Acad. Sci. USA 70, 1209-1213.
Simsek, M. (1974). Ph.D. Thesis, Massachusetts Institute of Tech- nology, Cambridge, Massachusetts.
Simsek, M., and RajBhandary, U. L. (1972). Biochem. Biophys. Res. Commun. 49, 508-511.
Simsek, M., Petrissant, G., and RajBhandary, U. L. (1973a). Proc. Nat. Acad. Sci. USA 70, 2600-2604.
Simsek, M., Ziegenmeyera J., Heckman, J., and RajBhandary, U. L. (1973b). Proc. Nat. Acad. Sci. USA 70, 1041-1045.
Simsek, M., RajBhandary, U. L., Boisnard, M., and Petrissant, G. (1974). I\lature 247, 518-521.
Urquhart, N. (1973). Ph.D. Thesis, University of British Columbia, Vancouver, British Columbia, Canada.
Walker, R. T., and RajBhandary, U. L. (1975). Nucleic Acids Res. 2, 61-78.
Walker, T. A., Betz, J. L., Olah, J., and Pace, N. R. (1975). FEBS Letters 54, 241-244.
Wintermeyer, W., Thiebe, R., and Zachau, H. G. (1972). Hoppe- Seyler’s Z. Physiol. Chem. 353, 1626-1632.
Woledge, J., Corry, M. J., and Payne, P. I. (1974). Biochim. Biophys. Acta 349, 339-350.
Yamada, Y., and Ishikura, H. (1975). FEBS Letters 54, 155-158.