Embed Size (px)
Citation preview

Proc. Natl. Acad. Sci. USAVol. 93, pp. 7059-7062, July 1996Cell Biology
Nuclear protein import: Ran-GTP dissociates the karyopherin axf8heterodimer by displacing a from an overlapping binding site on f8
(liquid phase binding assay/digitonin-permeabilized cells/karyopherin 13 mutant/synthetic peptides)
JUNONA MOROIANU, GUNTER BLOBEL*, AND AURELIAN RADULaboratory of Cell Biology, Howard Hughes Medical Institute, The Rockefeller University, New York, NY 10021
Contributed by Gunter Blobel, April 3, 1996
ABSTRACT The a subunit of the karyopherin het-erodimer functions in recognition of the protein importsubstrate and the 13 subunit serves to dock the trimericcomplex to one of many sites on nuclear pore complex fibers.The small GTPase Ran and the Ran interactive protein, p1O,function in the release of the docked complex. Repeated cyclesof docking and release are thought to concentrate the trans-port substrate for subsequent diffusion into the nucleus.Ran-GTP dissociates the karyopherin heterodimer and formsa stoichiometric complex with Ran-GTP. Here we report themapping of karyopherin 3's binding sites both for Ran-GTPand for karyopherin a. We discovered that karyopherin .3'sbinding site for Ran-GTP shows a striking sequence similar-ity to the cytoplasmic Ran-GTP binding protein, RanBP1.Moreover, we found that Ran-GTP and karyopherin a bind tooverlapping sites on karyopherin 18. Having a higher affinityto the overlapping site, Ran-GTP displaces karyopherin caand binds to karyopherin 13. Competition for overlappingbinding sites may be the mechanism by which GTP boundforms of other small GTPases function in correspondingdissociation-association reactions. We also mapped Ran'sbinding site for karyopherin 13 to a cluster of basic residuesanalogous to those previously shown to constitute karyo-pherin ca's binding site to karyopherin 1.
Four transport factors have been isolated and shown to berequired for nuclear localization sequence (NLS)-mediatedimport of proteins into nuclei (1-14). The a subunit of thekaryopherin a/13 heterodimer binds to the NLS protein (7, 8,15), whereas the 13 subunit mediates docking to those nucleo-porins that share repetitive tetrapeptide motifs (8, 16-18). Thesmall GTPase Ran and the Ran-interactive protein, plO,function in the subsequent translocation step (1-3, 14) bydissociating the docked karyopherin heterodimer (19). Thisdissociation reaction is coordinated by plO (20), which binds topeptide repeat containing nucleoporins, to karyopherin 13, andto Ran-GDP (but not to Ran-GTP) (20). The addition to thiscomplex of GTP causes in situ formation of Ran-GTP, whichin turn dissociates karyopherin a, leaving karyopherin 13 boundto nucleoporin (20). This allows karyopherin a (8, 21) and theNLS protein to diffuse into the nucleus, across the central tube[also termed "central plug" or "transporter" (22)] of thenuclear pore complex. The repeat containing nucleoporins arelocated specifically to cytoplasmic or nucleoplasmic fibersextending from the nuclear pore complex (for review, see ref.23). Repeated cycles of docking and release along the multipledocking sites of these tentacles would serve to concentrateNLS-proteins for diffusion across the central tube, down achemical concentration gradient (19, 20, 24).The Ran-GTP-mediated dissociation of the karyopherin
heterodimer occurs through the formation of a stoichiometriccomplex of Ran-GTP with karyopherin 13 (19, 25). To deter-
mine the mechanism for this association-dissociation reaction,we have undertaken to map the binding sites on karyopherin,B for both Ran-GTP and for karyopherin a. We found thatthese two binding sites overlap and that Ran-GTP, havingmore than a 10-fold higher affinity than karyopherin a for theoverlapping binding site on karyopherin 13, dissociates thekaryopherin heterodimer by competing with karyopherin a forbinding to karyopherin 13. Interestingly, karyopherin ,B's bind-ing site for Ran-GTP shows striking sequence similarity withthe cytoplasmic Ran-GTP binding protein, RanBP1 (26) andwith the four RanBP1-homologous domains of the nucleo-porin Nup358 (27, 28). We also mapped Ran's binding site forkaryopherin 13 and located it to a cluster of primarily basicresidues, analogous to clusters of basic residues of karyopherina that bind to karyopherin 13 (29).
MATERIALS AND METHODSPreparation of Recombinant Transport Factors. Recombi-
nant, full-length, human karyopherin al (referred herein as a),rat karyopherin 1 (lacking 12 N-terminal residues), and humanplO were expressed as His6 tagged proteins and purified fromEscherichia coli BL21(DE3) lysates on a Ni-nitrilotriacetic acid(NTA) column (7, 8). Recombinant human Ran was preparedas described (26).
Preparation of Karyopherin 13 Mutant. A karyopherin 13mutant, containing a deletion of residues 329-342 and termed1A329-342, was prepared by using PCR with primers con-taining restriction sites for insertion in the pET21b vector;expression in E. coli BL21(DE3) was induced by 1 mMisopropyl 1-D-thiogalactoside (IPTG). The recombinant His6tagged protein was purified as described (7) except that cellswere lysed in a French pressure cell. The purified protein wasdialyzed against transport buffer (20 mM HepesKOH, pH7.3/110 mM potassium acetate/2 mM magnesium acetate/imM EGTA/2 mM dithiothreitol), aliquoted, and stored at-70°C until use.
Synthetic Peptides. The following peptides were synthe-sized: ATAEEETEEE (residues 51-60 of karyopherin a);LTKQDENDDDDDWN (residues 329-342 of karyopherin13); NPDWRYRD (residues 375-382 of karyopherin 13);KDRKVKAKSLVFHRKK (residues 127-142 of Ran); andCYTPPKKKRKV, the NLS of simian virus 40 T antigen withan N-terminal Cys added for coupling.
Solution Binding Assay. Binding assays were performedwith one of the proteins immobilized on AffiGel 10 (2 ,ug ofprotein per 10 Al of beads) (29). The 50% bead slurry inbinding buffer [20mM Hepes-KOH, pH 7.3/110 mM KOAc/2mM Mg(OAc)2/1 mM EGTA/2 mM dithiothreitol/0.1%Tween-20] (20 ,ul) was incubated in a final volume of 40 ,ul withvarious components for 30 min at 20°C. Beads were collected
The publication costs of this article were defrayed in part by page chargepayment. This article must therefore be hereby marked "advertisement" inaccordance with 18 U.S.C. §1734 solely to indicate this fact.
7059
Abbreviations: NLS, nuclear localization sequence; HSA, humanserum albumin.*To whom reprint requests should be addressed. e-mail:[email protected].

7060 Cell Biology: Moroianu et al.
by centrifugation at 2000 x g for 30 sec. The collectedsupernatant (29.5,l) represented the unbound fraction. Beadswere washed 2x in 0.5 ml binding buffer and proteins thatremained associated with the beads represented the boundfraction. Bound and unbound fractions were analyzed bySDS/PAGE and Coomassie blue staining as described (19).
Nuclear Import Assays. Import assays were done usingBuffalo rat liver cells permeabilized with digitonin as described(7, 8).
RESULTSAn Acidic Region of Karyopherin (3 Binds to Karyopherin
a. As clusters of primarily basic residues in the N-terminalregion of karyopherin a were previously found to constitutekaryopherin a's determinants for binding to karyopherin (3(29), we reasoned that a corresponding cluster of acidicresidues may comprise the cognate binding region in karyo-pherin (3, particularly as binding is sensitive to high salt (9). Thelongest stretch of acidic residues of karyopherin (3, and a highlyconserved one among ( homologs of other species, isD333ENDDDDD340. Indeed, a synthetic peptide, (329-342, thatcontained this acidic region inhibited binding of karyopherina to immobilized karyopherin ( (Fig. lA, compare lane 1 withlane 2). Moreover, this peptide also inhibited import ofNLS-HSA (nuclear localization sequence of simian virus 40 Tantigen coupled to human serum albumin) into nuclei ofdigitonin-permeabilized cells (Fig. 1Db) in a concentration-dependent manner (Fig. 1B). This inhibition was specific asanother highly acidic peptide, A51TAEEETEEE6O of karyo-pherin a, had no effect either on binding of karyopherin a toimmobilized karyopherin 3 or on import of NLS-HSA intonuclei of digitonin-permeabilized cells (data not shown).These data indicated that P329-342 is a determinant of karyo-pherin ('s binding site for karyopherin a.
A IMMOBILIZED 1 B ioa + + a 0329-3421329-342 - + 80
.|-a BB: 132-4
R ". -4 -a -
0 25 50 75 10012 MOR zXCESSOFPEPTE
D
CWMOBilIZEDPA329-342
m
To further substantiate that 1329-342 constitutes a bindingdeterminant for karyopherin a, we prepared a recombinantkaryopherin 3 mutant that lacked residues 329-342, termed13A329-342. Indeed, immobilized l3A329-342 was unable tobind karyopherin a (Fig. 1C). Moreover, (3A329-342 wasunable to substitute for karyopherin B in import of NLS-HSAinto nuclei of digitonin-permeabilized cells (Fig. lDc). Incontrast, 3PA329-342 retained its ability to bind to peptide-repeat-containing nucleoporins (data not shown), indicatingthat another binding domain of karyopherin 13 remainedfunctional. Hence, (3A329-342's inability to bind to karyo-pherin a is likely to have resulted from a specific loss of abinding determinant rather than from general misfolding.
Overlapping Binding Sites for a and for Ran-GTP on f.Yeast karyopherin , has been shown to form a stoichiometriccomplex with yeast Ran-GTP (19, 25). As expected, immobi-lized mammalian karyopherin , also bound mammalian Ran-GTP (Fig. 2A, lane 1). We then tested whether syntheticpeptides representing regions of karyopherin 3would competefor this binding. Surprisingly, we found that a peptide, (3329-342,that inhibited binding of karyopherin a to immobilized karyo-pherin (3 (Fig. 1A) also inhibited binding of Ran-GTP toimmobilized (3 (Fig. 2A, compare lane 1 with lane 2). Thisinhibition was specific as another highly acidic peptide,A51TAEEETEEE6o of karyopherin a, had no effect on bindingof Ran-GTP to immobilized karyopherin (3 (data not shown).Likewise, the karyopherin (3 mutant lacking residues 329-342that failed to bind karyopherin a (Fig. 1C) also showed areduced ability to bind Ran-GTP (Fig. 2B). These data suggestthat karyopherin a and Ran-GTP share a binding site deter-minant on karyopherin (3.The mapping in karyopherin 3 of a determinant for Ran-
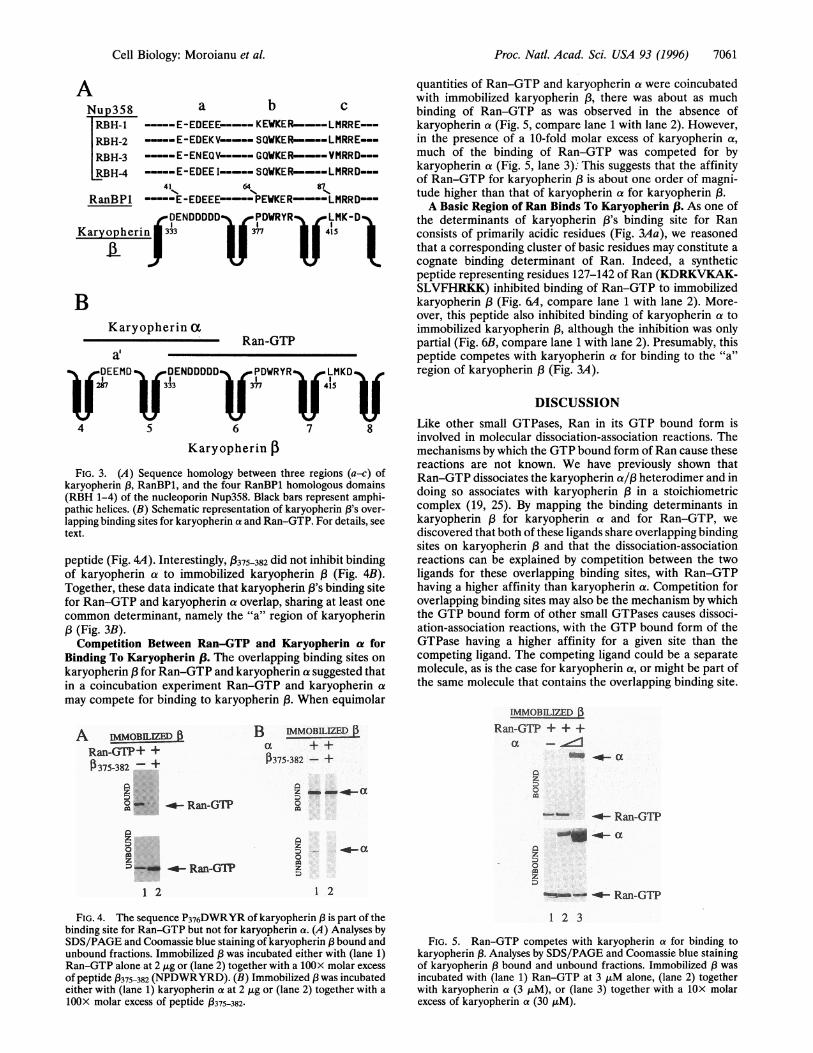
GTP binding to residues 329-342, prompted us to search forsequence similarities between this determinant and the se-quence of the cytosolic Ran-GTP-binding protein, Ran BP1(26). Indeed, the region comprising residues 329-342 wasamong three discontinuous regions (Fig. 3Aa-c) in karyo-pherin ( that showed a striking sequence similarity to threediscontinuous regions in RanBP1 and in the four Ran BP1-homologous domains of the nucleoporin, Nup358 (27,28). Thethree regions a-c of karyopherin 3 may be positioned as loopson the surface of karyopherin 3 by intervening nonidenticalrepeats, each of about 40 residues and each forming a pair ofantiparallel amphipathic helices (Fig. 3).To test whether the "b" region of karyopherin (3 (Fig. 3) is
involved in Ran-GTP binding, we synthesized the peptide,(375-382, containing this segment. Indeed, binding of Ran-GTPto immobilized karyopherin (3 was competed for by this
Control 1(329-342)
I IPA329-342
FIG. 1. An acidic region of karyopherin , binds to karyopherin a.(A) SDS/PAGE and Coomassie blue staining analyses of karyopherin,B bound and unbound fractions. Immobilized 13 was incubated witheither (lane 1) karyopherin a alone at 0.5 jig or (lane 2) together witha 10OX molar excess of peptide /3329-342. (B) Quantitative analysis ofnuclear import of NLS-HSA into nuclei of digitonin-permeabilizedcells in the presence of increasing molar excess of peptide /3329-342 withrespect to karyopherin /B. (C) Analysis by SDS/PAGE of karyopherinf3A329-342 bound and unbound fractions. Immobilized 3lA329-342was incubated with karyopherin a at 0.5 jig. (D) Inhibition of nuclearimport of NLS-HSA into nuclei of digitonin-permeabilized cells bypeptide (329-342 or by mutant /A329-342. (Da) Import of NLS-HSA(200 nM) in the presence of karyopherin a and /3 (200 nM), Ran (3,iM), and plO (6 jiM). (Db) Import reaction contained in addition a10OX molar excess of peptide (329-342. (Dc) Karyopherin / wassubstituted by the mutant /3A329-342. (Bar = 10 ,um.)
A IMMOBnI 13Ran-GTP+ +0329-342 - +
-- <- Ran-GTP
9
9 -_- 0 Ran-GT?
BIMMOBILIZEDPA329-342
--w- Ran-GTP
- - Ran-GTP
1 2
FIG. 2. Karyopherin /3 contains overlapping binding sites forkaryopherin a and Ran-GTP. (A) Analysis by SDS/PAGE andCoomassie blue staining of karyopherin /3 bound and unboundfractions. Immobilized / was incubated with either (lane 1) Ran-GTPalone at 1 jig or (lane 2) together with a 10OX molar excess of peptide/329-342. (B) Analyses by SDS/PAGE and Coomassie blue staining ofmutant karyopherin B3A329-342 bound and unbound fractions. Im-mobilized /3A329-342 was incubated with 1 jig of Ran-GTP.
Proc. Natl. Acad. Sci. USA 93 (1996)
Proc. Natl. Acad. Sci. USA 93 (1996) 7061
ANup358 a b cRBH-1 E-EDEEE----- KEWKER----LMRRE--RBH-2 -----E-EDEKV-----SQWKER----LMRRE--RBH-3 -----E-ENEQV-----GQWKER----VMRRD---RBH-4 E-EDEE I-----SQWKER----LMRRD---
41 64 87
RanBP -----E-EDEEE-----PEWKER----LMRRD--DENDDDDDiJ PDWRYR LMK-D
Karyopherin 33 15
BKaryopherin a
a'Ran-GTP
DEEMD DENDDDDD PDWRYR LMKD
3~~33 415
4 5 6 7 8
Karyopherin
FIG. 3. (A) Sequence homology between three regions (a-c) ofkaryopherin (3, RanBP1, and the four RanBP1 homologous domains(RBH 1-4) of the nucleoporin Nup358. Black bars represent amphi-pathic helices. (B) Schematic representation of karyopherin (3's over-lapping binding sites for karyopherin a and Ran-GTP. For details, seetext.
peptide (Fig. 4A). Interestingly, 3375382 did not inhibit bindingof karyopherin a to immobilized karyopherin (Fig. 4B).Together, these data indicate that karyopherin 3's binding sitefor Ran-GTP and karyopherin a overlap, sharing at least onecommon determinant, namely the "a" region of karyopherin(3(Fig. 3B).Competition Between Ran-GTP and Karyopherin at for
Binding To Karyopherin 13. The overlapping binding sites onkaryopherin for Ran-GTP and karyopherin a suggested thatin a coincubation experiment Ran-GTP and karyopherin a
may compete for binding to karyopherin (3. When equimolar
A IMMOBILZED B3Ran-GTP+ +0375-382 - +
...
.......
0 ; Ran-GTP
D'-- 4- Ran-GTP
1 2
B IMMOBILIZED
a ++13375-382 - +
az - a.4-E1
0
z0 1z
1 2
FIG. 4. The sequence P376DWRYR of karyopherin , is part of thebinding site for Ran-GTP but not for karyopherin a. (A) Analyses bySDS/PAGE and Coomassie blue staining of karyopherin bound andunbound fractions. Immobilized , was incubated either with (lane 1)Ran-GTP alone at 2 ,ig or (lane 2) together with a 10OX molar excessof peptide (3375-382 (NPDWRYRD). (B) Immobilized 3was incubatedeither with (lane 1) karyopherin a at 2 jig or (lane 2) together with a
10OX molar excess of peptide ,B375-382.
quantities of Ran-GTP and karyopherin a were coincubatedwith immobilized karyopherin (3, there was about as muchbinding of Ran-GTP as was observed in the absence ofkaryopherin a (Fig. 5, compare lane 1 with lane 2). However,in the presence of a 10-fold molar excess of karyopherin a,much of the binding of Ran-GTP was competed for bykaryopherin a (Fig. 5, lane 3). This suggests that the affinityof Ran-GTP for karyopherin ,B is about one order of magni-tude higher than that of karyopherin a for karyopherin P3.A Basic Region of Ran Binds To Karyopherin ,B. As one of
the determinants of karyopherin (3's binding site for Ranconsists of primarily acidic residues (Fig. 3Aa), we reasonedthat a corresponding cluster of basic residues may constitute acognate binding determinant of Ran. Indeed, a syntheticpeptide representing residues 127-142 of Ran (KDRKVKAK-SLVFHRKK) inhibited binding of Ran-GTP to immobilizedkaryopherin 3 (Fig. 6A, compare lane 1 with lane 2). More-over, this peptide also inhibited binding of karyopherin a toimmobilized karyopherin (3, although the inhibition was onlypartial (Fig. 6B, compare lane 1 with lane 2). Presumably, thispeptide competes with karyopherin a for binding to the "a"region of karyopherin ( (Fig. 3A).
DISCUSSIONLike other small GTPases, Ran in its GTP bound form isinvolved in molecular dissociation-association reactions. Themechanisms by which the GTP bound form of Ran cause thesereactions are not known. We have previously shown thatRan-GTP dissociates the karyopherin a/,3 heterodimer and indoing so associates with karyopherin 3 in a stoichiometriccomplex (19, 25). By mapping the binding determinants inkaryopherin (3 for karyopherin a and for Ran-GTP, wediscovered that both of these ligands share overlapping bindingsites on karyopherin (3 and that the dissociation-associationreactions can be explained by competition between the twoligands for these overlapping binding sites, with Ran-GTPhaving a higher affinity than karyopherin a. Competition foroverlapping binding sites may also be the mechanism by whichthe GTP bound form of other small GTPases causes dissoci-ation-association reactions, with the GTP bound form of theGTPase having a higher affinity for a given site than thecompeting ligand. The competing ligand could be a separatemolecule, as is the case for karyopherin a, or might be part ofthe same molecule that contains the overlapping binding site.
IMMOBELIZEDRan-GTP + + +
a_ a
z001
mz
_-~ -'4-Ran-GTP
1 2 3
FIG. 5. Ran-GTP competes with karyopherin a for binding tokaryopherin ,B. Analyses by SDS/PAGE and Coomassie blue stainingof karyopherin ,B bound and unbound fractions. Immobilized f3 wasincubated with (lane 1) Ran-GTP at 3 ,uM alone, (lane 2) togetherwith karyopherin a (3 J,M), or (lane 3) together with a 1OX molarexcess of karyopherin a (30 ,IM).
Cell Biology: Moroianu et al.

7062 Cell Biology: Moroianu et al.
A IMMOBILIZED f B IMMOBILIZEDRan-GTP++ a ++Ran 127-142 - + Ran 127-142 - +
o 0- _- Ran-GTP
z zZ 3) ___ ao 0m mzD- - 4- Ran-GTP
1 2 1 2
FIG. 6. The Ran binding site for karyopherin , contains clusters ofbasic amino acids. (A) Analyses by SDS/PAGE and Coomassie bluestaining of karyopherin ,B bound and unbound fractions. Immobilized( was incubated either with (lane 1) Ran-GTP alone at 2 ,Lg or (lane2) together with a 10OX molar excess of the synthetic peptideRanl27m142 (KDRKVKAKSLVFHRKK). (B) Immobilized (3 was in-cubated either with (lane 1) karyopherin a alone at 2 ,ug or (lane 2)together with a 10OX molar excess of the synthetic peptide Ranl27_142.
The ligand-binding site on karyopherin A that is shared byboth Ran-GTP and karyopherin a is a stretch of acidicresidues (see Fig. 3Aa). The Ran-GTP binding site containsadditional downstream determinants (Figs. 3 and 4) and thekaryopherin a binding site is likely to comprise additionalupstream determinants (see Fig. 3B). Interestingly, the bindingdeterminants of karyopherin , for Ran-GTP showed a strik-ing sequence similarity to Ran BP1, a cytoplasmic Ran-GTPbinding protein (26) and to four RanBP1 homologous domainsof the nucleoporin Nup358 (27, 28) (see Fig. 3A). Hence,cytoplasmic RanBP1 or the RanBP1 homologous domains ofthe nucleoporin Nup358 might serve as competing ligands aswell and therefore might function in the displacement ofRan-GTP from karyopherin (B. We have previously shown thatthe cytoplasmic Ran-GAP (GTPase activating protein) isunable to hydrolyze GTP when Ran-GTP is complexed withkaryopherin (3 (25). Hence, displacement of Ran-GTP fromkaryopherin (3 may be a requisite for GTP hydrolysis byRan-GAP and may serve to recycle karyopherin (3 as Ran-GDP has only a low affinity for karyopherin (3 (25). Yetanother potential means for competitively displacing Ran-GTP from karyopherin ,B may be high local concentrations ofkaryopherin a.
Karyopherin 3 has been proposed to contain 11 "HEAT"repeats (30). These repeats consist of about 40 residues thatappear to form two antiparallel a helices with the loopsbetween the repeats serving as ligand binding sites (ref. 31; seeFig. 3).We have also mapped a determinant of Ran that binds to
karyopherin (. It was found to consist of a cluster of basicresidues (Fig. 6), not unlike the karyopherin a binding deter-minants for karyopherin (3 (29).
We thank Robert O'Neill and Peter Palese for karyopherin al/NPI-1-expressing vector pSE420; Philip Bernstein for recombinanthuman Ran; Makoto Hijikata for recombinant human plO; andmembers of the Protein/DNA Technology Center of the RockefellerUniversity for peptide synthesis.
1. Moore, M. S. & Blobel, G. (1993) Nature (London) 365,661-663.2. Melchior, F. B., Paschal, J.; Evans, J. & Gerace, L. (1993) J. Cell
Biol. 123, 1649-1659.3. Moore, M. S. & Blobel, G. (1994) Proc. Natl. Acad. Sci. USA 91,
10212-10216.4. Adam, E. J. H. & Adam, S. A. (1994) J. Cell Bio. 125, 547-555.5. Gorlich, D., Prehn, S., Laskey, R. & Hartmann, E. (1994) Cell 79,
767-778.6. Radu, A., Blobel, G. & Moore, M. S. (1995) Proc. Natl. Acad. Sci.
USA 92, 1769-1773.7. Moroianu, J., Blobel, G. & Radu, A. (1995) Proc. Natl. Acad. Sci.
USA 92, 2008-2011.8. Moroianu, J., Hijikata, M., Blobel, G. & Radu, A. (1995) Proc.
Natl. Acad. Sci. USA 92, 6532-6536.9. Gorlich, D., Kosika, S., Kraft, R., Dingwall, C., Laskey, R.,
Hartmann, E. & Prehn, S. (1995) Curr. Biol. 5, 383-392.10. Chi, N., Adam, E. J. H. & Adam, S. A. (1995) J. Cell Biol. 130,
265-274.11. Imamoto, N., Tachibana, T., Matsubae, M. & Yoneda, Y. (1995)
J. Biol. Chem. 270, 8559-8565.12. Imamoto, N., Shimamoto, T., Takao, T., Tachibana, T., Kose, S.,
Matsubae, M., Sekimoto, T., Shimonishi, Y. & Yoneda, Y. (1995)EMBO J. 14, 3617-3626.
13. Enenkel, C., Blobel, G. & Rexach, M. (1995) J. Biol. Chem. 270,16499-16502.
14. Paschal, B. M. & Gerace, L. (1995) J. Cell Biol. 129, 925-937.15. Weis, K., Mattaj, I. & Lamond, A. (1995) Science 268, 1049-1051.16. Radu, A., Moore, M. S. & Blobel, G. (1995) Cell 81, 215-222.17. Kraemer, D., Strambio-de-Castillia, M. C., Blobel, G. & Rout,
M. P. (1995) J. Biol. Chem. 270, 19017-19021.18. Iovine, M. K., Watkins, J. L. & Wente, S. R. (1995) J. Cell Biol.
131, 1699-1713.19. Rexach, M. & Blobel, G. (1995) Cell 83, 683-692.20. Nehrbass, U. & Blobel, G. (1996) Science 272, 120-122.21. Gorlich, D., Vogel, F., Mills, A., Hartmann, E. & Laskey, R.
(1995) Nature (London) 377, 246-248.22. Akey, C. W. & Radermacher, M. (1993) J. Cell Biol. 122, 1-19.23. Rout, M. P. & Wente, S. R. (1994) Trends Biochem. Sci. 4,
357-365.24. Paine, L. P. (1993) Trends Cell Biol. 3, 325-329.25. Floer, M. & Blobel, G. (1996) J. Biol. Chem. 271, 5313-5316.26. Coutavas, E., Ren, M., Oppenheim, J., D'Eustachio, P. & Rush,
M. (1993) Nature (London) 366, 585-587.27. Wu, J., Matunis, M. J., Kraemer, D., Blobel, G. & Coutavas, E.
(1995) J. Biol. Chem. 270, 14209-14213.28. Yokoyama, N., Hayashi, N., Seki, T., Pante, N., Ohba, T., Nishii,
K., Kuma, K., Hayashida, T., Miyata, T., Aebi, U., Fukui, M. &Nishimoto, T. (1995) Nature (London) 376, 184-186.
29. Moroianu, J., Blobel, G. & Radu, A. (1996) Proc. Natl. Acad. Sci.USA, in press.
30. Andrade, M. A. & Bork, P. {1995) Nat. Genet. 11, 115-116.31. Ruediger, R., Hentz, M., Fait, J., Mumby, M. & Walter, G. (1994)
J. Virol. 68, 123-129.
Proc. Natl. Acad. Sci. USA 93 (1996)





























