Embed Size (px)
Citation preview

www.elsevier.com/locate/cogbrainres
Cognitive Brain Research
Research Report
Now you see it, now you don’t: Variable hemineglect in a
commissurotomized man
Michael C. Corballis a,*, Paul M. Corballis b, Mara Fabri c, Aldo Paggi d, Tullio Manzoni c
aResearch Centre for Cognitive Neuroscience, University of Auckland, Auckland, New ZealandbDepartment of Psychology, Georgia Institute of Technology, Atlanta, GA 30332, USA
cDepartment of Neuroscience, Section of Physiology, Universita Politecnica delle Marche, Ancona, ItalydCentro Epilessia, Ospedale Regionale ‘‘Umberto I’’, Ancona, Italy
Accepted 9 August 2005
Available online 9 September 2005
Abstract
We describe the case of a callosotomized man, D.D.V., who shows unusual neglect of stimuli in the left visual field (LVF). This is manifest
in simple reaction time (RT) to stimuli flashed in the LVF and in judging whether pairs of filled circles in the LVF are of the same or different
color. It may reflect strong left-hemispheric control and consequent attention restricted to the right side of space. It is not evident in simple RT
when there are continuous markers in the visual fields to indicate the locations of the stimuli. In this condition, his RTs are actually faster to
LVF than to right visual field (RVF) stimuli, suggesting a switch to right-hemispheric control that eliminates the hemineglect. Neglect is also
not evident when D.D.V. responds by pointing to or touching the locations of the stimuli, perhaps because these responses are controlled by the
dorsal rather than the ventral visual system. Despite his atypical manifestations of hemineglect, D.D.V. showed evidence of functional
disconnection typical of split-brained subjects, including prolonged crossed–uncrossed different in simple reaction time, inability to match
colors between visual fields, and enhanced redundancy gain in simple RT to bilateral stimuli even when the stimulus in the LVF was neglected.
D 2005 Elsevier B.V. All rights reserved.
Theme: Sensory systems
Topic: Subcortical visual pathways
Keywords: Corpus callosum; Hemineglect; Redundancy gain; Split brain
1. Introduction
Patients with right-sided brain injury commonly show
hemineglect of space on the left side, which is generally
taken to indicate that the left hemisphere directs attention
only to the right side of space. Right-sided hemineglect
occurs only rarely, or transiently, following left-sided injury,
indicating that the right hemisphere directs attention to both
sides of space (e.g., [19]). Rather surprisingly, there is little
evidence for hemineglect in patients with complete section
0926-6410/$ - see front matter D 2005 Elsevier B.V. All rights reserved.
doi:10.1016/j.cogbrainres.2005.08.002
Abbreviations: LVF, left visual field; RVF, right visual field; RT,
reaction time
* Corresponding author. Fax: +64 9 373 7450.
E-mail address: [email protected] (M.C. Corballis).
of the corpus callosum, even though one might expect
attention to be controlled by a single hemisphere. For
example, Joynt [24] observed that split-brained patients
showed no evidence of hemineglect in their everyday lives,
and Plourde and Sperry [29] found no evidence of left
hemineglect in three split-brained patients when they used
their right hands, implying that the left hemisphere mediated
awareness of both sides of space. One of these patients,
L.B., did show evidence of left hemineglect when asked
whether briefly flashed horizontal lines extended further to
the right or to the left, but no such bias when the lines were
available for unlimited viewing [9].
Split-brained patients do show some weakness in
response to stimuli in the left visual field (LVF). For
example, in a simple reaction time (RT) study, two
25 (2005) 521 – 530

M.C. Corballis et al. / Cognitive Brain Research 25 (2005) 521–530522
callosotomized men, M.E. and J.W., responded more slowly
to LVF than to RVF stimuli [3,33], and J.W.’s event-related
potentials (ERPs) also show a reduced P300 component to
LVF stimuli [31]. Both J.W. and M.E. also show extinction
to LVF stimuli when presented simultaneously with RVF
stimuli. When asked to indicate what they saw, J.W.
reported ‘‘both’’ on only 47.5% of bilateral trials [33],
while M.E. responded ‘‘both’’ on only 22.5% of bilateral
trials [4]. On nearly all remaining bilateral trials, they
reported only the stimulus in the RVF.
The most striking, and indeed unusual, example of left-
sided neglect in a callosotomized patient comes from a patient
known as D.D.V. who shows a strong tendency in simple RT
to ignore visual stimuli flashed in the LVF [14]. Out of 60
LVF trials with each hand, he responded only once with his
right hand and 13 times with his left hand. Previous research
indicates that subjects with hemineglect show prolonged RTs
to unilateral LVF stimuli and sometimes fail to respond to
them [28], but D.D.V.’s near total failure to respond might be
taken as indicative of a visual field defect rather than
hemineglect. This is unlikely, however, for several reasons.
First, he shows a strong rightward bias in line bisection [18],
indicative of left hemineglect rather than of a visual field
defect. Second, he showed no evidence of failing to see LVF
stimuli when asked to point to the location of stimuli flashed
in either LVF or RVF [14]. Third, Savazzi and Marzi [34]
report no evidence of hemineglect in D.D.V. in a study very
similar to that of Corballis et al. [14], the main difference
being that guide boxes were continuously present in LVF and
RVF to indicate where the stimuli would be located. While
these last two findings seem to rule out a visual field defect,
they also show that D.D.V.’s hemineglect is intermittent, and
the main aim of the present study was to determine the factors
influencing it.
Although D.D.V.’s hemineglect is unusual, his RTs are
similar to those of other split-brained patients in that he
shows marked redundancy gain when presented with stimuli
in both visual fields, with shorter RTs to bilateral pairs of
stimuli than to single stimuli presented unilaterally. More-
over, RTs to bilateral stimuli in split-brained subjects are
faster than predicted from a race model, in which it is simply
assumed that responses to bilateral stimuli are the outcome
of a race between processes initiated by the two unilateral
stimuli [27,32], whereas in normal subjects, RTs to bilateral
stimuli typically do not violate the race model [10,13,22,
26,33,34]. In the case of D.D.V., violation of the race model
occurred both when there was neglect of LVF stimuli [14]
and when there was no such neglect [36]. Redundancy gain
in excess of that predicted by the race model implies
interhemispheric neural summation.
Two kinds of explanation for this paradoxical summation
in the split brain have been proposed. According to one, the
effect arises, not because RT is decreased to bilateral
stimulation, but because RTs to unilateral stimuli are slowed
relative to those in the normal brain [33,34]. Others have
argued that it occurs because RTs to bilateral stimuli are
decreased in the split brain either because of a release from
interhemispheric inhibition [10,13,36] or because of
increased neural summation [22]. Whatever the explanation,
it is clear from D.D.V.’s results, as well as those from other
studies [35,37], that redundancy gain can occur even when
unilateral stimuli in one visual field are below the threshold
of detection. It has also been found that redundancy gain
in both normal and split-brained subjects is reduced or
abolished under conditions in which the stimuli are equilu-
minant with the background [10] or when the stimuli are in
monochromatic purple [36]—conditions expected to mini-
mize input to the superior colliculus. These results suggest that
the superior colliculus may be the site of neural summation.
In the present study, we report further data from D.D.V.,
with a view to isolating more explicitly the variables
influencing whether or not he shows neglect for stimuli in
the LVF. In Experiment 1, we examine whether D.D.V.
continues to show hemineglect when asked to make
different keyboard responses to LVF and RVF stimuli. In
Experiment 2, we examine D.D.V.’s performance both in
keyboard responding and in pointing to, and touching, the
stimulus when there are guides to the location of the stimuli,
as in the study by Savazzi and Marzi [36]. In Experiment 3,
we examine whether he shows neglect in a more complex
task involving same–different judgments about stimuli
presented in the LVF, RVF, or bilaterally.
2. Experiment 1
In our earlier study, D.D.V. was able to respond
accurately to stimuli in either visual field by pointing with
his left hand to stimuli in the LVF or with his right hand to
stimuli in the RVF but responded only rarely when
responding with a single hand on the keyboard [14]. One
question was whether his ability to respond by pointing was
due to the use of spatially separated responses (left-sided vs.
right-sided) or whether it was due to pointing per se. In this
experiment, D.D.V. was asked to respond to LVF stimuli by
pressing a key on the left of the keyboard to stimuli in the
LVF, a key on the right of the keyboard to stimuli in the
RVF, and either key if the stimuli appeared in both visual
fields. For comparative purposes, another callosotomized
subject, J.W., also performed this task.
2.1. Method
2.1.1. Subjects
D.D.V. underwent complete callosotomy in two stages,
the final stage in 1994. He was tested in two sessions, one
when he was 38 years old and one when he was 39. J.W.
underwent two-stage callosotomy in 1979 and was 46 when
tested. Both are right-handed. Further details of the neuro-
logical status of D.D.V. are provided by Fabri et al. [15] and
of J.W. by Gazzaniga et al. [16]. There is no evidence of a
visual field defect in D.D.V., and other than his failure to

Table 1
Mean RTs and number of responses under each condition for each subject
in Experiment 1
J.W. D.D.V.
LVF RVF Bilateral LVF RVF Bilateral
Mean RT 398 327 339 391 330 297
Left-hand responses 30 0 6 3 0 0
Right-hand responses 0 30 24 0 28 29
No response 0 0 0 27 2 1
M.C. Corballis et al. / Cognitive Brain Research 25 (2005) 521–530 523
respond to LVF stimuli in simple RT, the only previous
evidence of hemineglect is his marked rightward bias in line
bisection [18]. Both subjects gave informed consent, and the
research was approved by the University of Auckland
Human Participants Ethics Committee.
2.1.2. Stimuli
The stimuli were the same as those used in the experi-
ment by Corballis et al. [14]. They were filled circular disks,
white against a dark gray background, 0.86- in visual angle,
and centered 5- either to the left or right of a central fixation
cross. They were presented on a fast-fade videographics
adapter screen for 100 ms and were viewed from a distance
of 57 cm.
2.1.3. Procedure
There were 99 experimental trials, 30 in which the disks
were presented in the left visual field (LVF), 30 in which they
were in the right visual field (RVF), 30 in which stimuli were
presented simultaneously in both fields (bilateral), and 9
‘‘catch’’ trials in which no stimulus was presented. At the
beginning of each block, a small fixation cross appeared in
themiddle of the screen and remained there for the duration of
the experiment. The subject sat with the forefinger of the left
hand lightly placed on the X key and the forefinger of the
right hand lightly placed on the M key. He was asked to press
the M key if a stimulus appeared in the RVF, the X key if a
stimulus appeared in the LVF, and either key, or both keys, if
stimuli appeared in both fields. RTs were recorded from
stimuli onset, and, in the case of bilateral stimuli, only the first
response was recorded if the subject pressed both keys.
On catch trials, the subject was required to withhold
response for 1700 ms following the stimulus. Following the
response, or a ‘‘time-out’’ period if no response occurred,
there was a variable interval of 1300, 1400, 1500, 1600, or
1700 ms before the stimulus appeared. Each of the five
variable intervals was paired six times with each stimulus
configuration to make up the 90 trials on which stimuli
appeared, and these were randomly intermixed with the nine
catch trials.
Prior to the experimental trials, the subjects were given
10 practice trials in which the stimulus conditions were
randomly selected.
2.2. Results
Table 1 shows the means RTs and number of responses to
the stimuli for each subject. It is clear that, unlike J.W.,
D.D.V showed strong neglect of LVF stimuli. Furthermore,
he always responded with his right hand to bilateral stimuli,
whereas J.W. responded with his right hand on 24 trials and
his left hand on 6. Analysis of variance of J.W.’s RTs
revealed a significant difference between the three con-
ditions (F(2, 87) = 9.51, P < 0.001), and Scheffe tests
further showed that his RTs to LVF stimuli were signifi-
cantly (P < 0.005) longer than those to RVF or bilateral
stimuli, which did not differ significantly from each other
(P = 0.804). Hence, there was no evidence for faster
responding to bilateral stimuli. For D.D.V., the 3 RTs to LVF
stimuli were omitted from an analysis of variance which
showed that RTs were significantly faster to bilateral than to
RVF stimuli (F(1, 55) = 7.06, P = 0.01). Hence, D.D.V,
unlike J.W., did show more rapid responses to bilateral than
to RVF stimuli, even though he only rarely responded to
LVF stimuli when they were presented unilaterally.
Fig. 1 plots cumulative distributions of responses over
time for the two subjects. If it is assumed that each
hemisphere independently races for control of response,
the following relation should hold:
pB ¼ pL þ pR � pL I pRð Þ;
where pB is the probability of a response having occurred to
stimuli in both fields, pL is the probability of a response
having occurred to the stimulus in the LVF alone, and p is
the probability of a response having occurred to the stimulus
in the RVF alone [32]. In Fig. 1, responses were collected
cumulatively in bins of 5, with bin 1 representing the
baseline, bin 2 the fastest 5 responses, bin 3 the fastest 10
responses, and so on. Responses in each bin were then
allocated to the different visual field conditions (LVF, RVF,
and bilateral). The frequencies in each bin were then divided
by 30 to produce cumulative probabilities for each visual
field. D.D.V. responded only 3 times to stimuli in the LVF.
It is clear that, for D.D.V., the responses to bilateral
stimuli cannot be explained in terms of this model since
pL = 0 and pB > pR over nearly all of the distribution. In a
more stringent version of this so-called race model, in which
the races in two hemispheres are not independent, the
following inequality should still hold [27]:
pB V pL þ pR;
D.D.V.’s responses also fail to conform to this inequality.
These results suggest that D.D.V.’s RTs to bilateral stimuli
result from neural summation, rather than probability
summation, between the hemispheres.
2.3. Discussion
In this experiment, the response arrangement was some-
what analogous to that in which the subject points to the
stimuli in that the subject responded to different locations
depending on the location of the stimuli. Nevertheless, D.D.V.

Fig. 1. Cumulative distributions of RT for J.W. and D.D.V. in Experiment 1.
M.C. Corballis et al. / Cognitive Brain Research 25 (2005) 521–530524
continued to show the strong hemineglect of LVF stimuli
reported earlier [14], suggesting that the absence of neglect
when he pointed to the stimuli themselves was not due to
differential response mapping. Rather, it suggests that point-
ing may depend on different pathways. D.D.V. made only
three (very late) responses with his left hand to LVF stimuli,
and none to bilateral stimuli. This may have been due simply
to his perceptual neglect of the LVF stimuli. As in previous
studies [13,33], J.W. also showed a bias in favor of the RVF
and the right hand, even when the stimuli were bilateral (see
Table 1), implying dominant left-hemispheric control.
It is also clear that, despite his neglect of LVF stimuli,
D.D.V. responded more quickly to bilateral stimuli than to
RVF stimuli. Since this exceeded the race model, it implies
neural summation. J.W., in contrast, did not show any
advantage in responding to bilateral stimuli, despite always
responding to unilateral LVF stimuli. It seems likely that
J.W.’s responses to bilateral stimuli were slowed somewhat
by uncertainty as to which hand to respond with, counter-
acting any summation effect. D.D.V., on the other hand,
appears to have been unaware of LVF stimuli even under the
bilateral condition and responded effectively as though there
were stimuli only in the RVF. Yet, the strong advantage to
bilateral stimuli shows that LVF stimuli did register under
the bilateral condition, speeding the response. This is further
evidence that neural summation can occur even though one
of the stimuli, when presented alone, is below detection
threshold [14,35,37].
3. Experiment 2
Savazzi and Marzi [36] reported that D.D.V. showed no
neglect of LVF stimuli in an experiment in which the stimuli
appeared in ‘‘guide boxes’’ to the left or right of fixation.
Each box was a 3- by 3- square comprised of 9 smaller
squares that changed randomly in luminance from 0.98 to
2.23 cd/m2 every 67 ms, against a background of 0.01 cd/
m2. After a variable interval, the stimulus appeared in the
center of this arrangement. Savazzi and Marzi suggest that
the presence of the guide boxes may have eliminated the
neglect of LVF stimuli observed by Corballis et al. [14].
In the present experiment, circles were located in the left
and right visual fields, and the stimuli appeared inside these
circles. The question was whether these markers would have
the same effect as the guide boxes in eliminating neglect of
the LVF stimuli. D.D.V. was given trials in which he
responded by touching the circles in which a stimulus had
appeared, or by touching the central fixation cross, or by
pressing the space bar. The touch responses were essentially
the same as the pointing responses in the experiment by
Corballis et al. [14], but D.D.V. was required to actually
touch the screen so that his RTs could be recorded.
3.1. Method
3.1.1. Subject
D.D.V. was the only subject tested in this experiment.
3.1.2. Stimuli
The stimuli were white disks, 0.86- cm in diameter,
centered 5- left or right of a fixation cross. They appeared
for 100 ms within white unfilled circles of diameter 1- thatwere continually present, and that served as markers for
touching responses. The screen was a touch screen that
allowed touches on the screen to be recorded as responses,
and RTs were measured from the onset of the stimulus.
3.1.3. Procedure
Stimulus presentations were arranged in randomly
ordered blocks of 90 trials, 30 in the LVF, 30 in the RVF,
and 30 in both fields. Following response to a stimulus, or
an interval of 2 s if there was no response, there was an
interval of 1500, 1700, 1900, 2100, or 2300 ms before the
next stimulus appeared. There were no catch trials. D.D.V.
was asked to respond to stimulus presentations in the
following ways:
1. Touching the stimulus locations. If he saw a disk appear
within either circle, he was to raise his hand and touch
that circle with his forefinger. In one block, he used the
forefinger of his left hand for LVF stimuli, his right hand
for RVF stimuli, and either (or both) hands for bilateral
stimuli. In a second block, he used only the left hand and
in a third block only the right hand.
2. Touching the fixation cross. Whenever he saw a stimulus,
he was to touch the fixation cross, with his right hand on
one block and his left hand on another block.
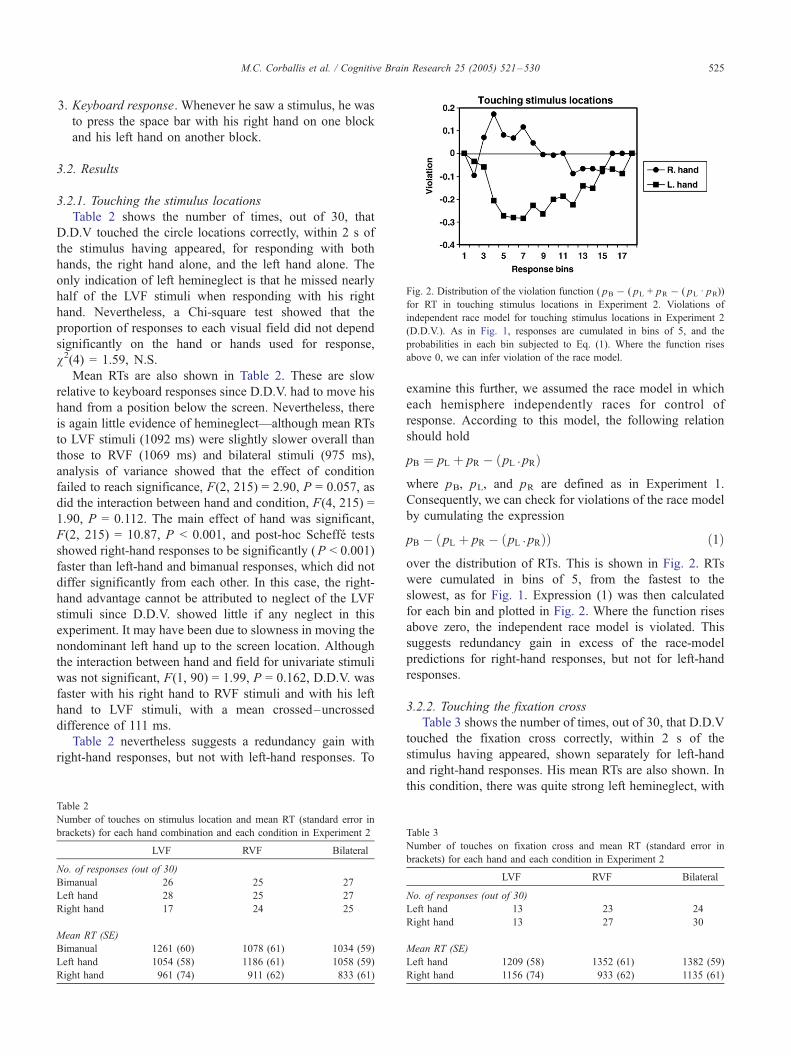
Fig. 2. Distribution of the violation function ( pB � ( pL + pR � ( pL I pR))for RT in touching stimulus locations in Experiment 2. Violations of
independent race model for touching stimulus locations in Experiment 2
(D.D.V.). As in Fig. 1, responses are cumulated in bins of 5, and the
probabilities in each bin subjected to Eq. (1). Where the function rises
above 0, we can infer violation of the race model.
M.C. Corballis et al. / Cognitive Brain Research 25 (2005) 521–530 525
3. Keyboard response. Whenever he saw a stimulus, he was
to press the space bar with his right hand on one block
and his left hand on another block.
3.2. Results
3.2.1. Touching the stimulus locations
Table 2 shows the number of times, out of 30, that
D.D.V touched the circle locations correctly, within 2 s of
the stimulus having appeared, for responding with both
hands, the right hand alone, and the left hand alone. The
only indication of left hemineglect is that he missed nearly
half of the LVF stimuli when responding with his right
hand. Nevertheless, a Chi-square test showed that the
proportion of responses to each visual field did not depend
significantly on the hand or hands used for response,
v2(4) = 1.59, N.S.
Mean RTs are also shown in Table 2. These are slow
relative to keyboard responses since D.D.V. had to move his
hand from a position below the screen. Nevertheless, there
is again little evidence of hemineglect—although mean RTs
to LVF stimuli (1092 ms) were slightly slower overall than
those to RVF (1069 ms) and bilateral stimuli (975 ms),
analysis of variance showed that the effect of condition
failed to reach significance, F(2, 215) = 2.90, P = 0.057, as
did the interaction between hand and condition, F(4, 215) =
1.90, P = 0.112. The main effect of hand was significant,
F(2, 215) = 10.87, P < 0.001, and post-hoc Scheffe tests
showed right-hand responses to be significantly (P < 0.001)
faster than left-hand and bimanual responses, which did not
differ significantly from each other. In this case, the right-
hand advantage cannot be attributed to neglect of the LVF
stimuli since D.D.V. showed little if any neglect in this
experiment. It may have been due to slowness in moving the
nondominant left hand up to the screen location. Although
the interaction between hand and field for univariate stimuli
was not significant, F(1, 90) = 1.99, P = 0.162, D.D.V. was
faster with his right hand to RVF stimuli and with his left
hand to LVF stimuli, with a mean crossed–uncrossed
difference of 111 ms.
Table 2 nevertheless suggests a redundancy gain with
right-hand responses, but not with left-hand responses. To
Table 2
Number of touches on stimulus location and mean RT (standard error in
brackets) for each hand combination and each condition in Experiment 2
LVF RVF Bilateral
No. of responses (out of 30)
Bimanual 26 25 27
Left hand 28 25 27
Right hand 17 24 25
Mean RT (SE)
Bimanual 1261 (60) 1078 (61) 1034 (59)
Left hand 1054 (58) 1186 (61) 1058 (59)
Right hand 961 (74) 911 (62) 833 (61)
examine this further, we assumed the race model in which
each hemisphere independently races for control of
response. According to this model, the following relation
should hold
pB ¼ pL þ pR � pL I pRð Þ
where pB, pL, and pR are defined as in Experiment 1.
Consequently, we can check for violations of the race model
by cumulating the expression
pB � pL þ pR � pL I pRð Þð Þ ð1Þ
over the distribution of RTs. This is shown in Fig. 2. RTs
were cumulated in bins of 5, from the fastest to the
slowest, as for Fig. 1. Expression (1) was then calculated
for each bin and plotted in Fig. 2. Where the function rises
above zero, the independent race model is violated. This
suggests redundancy gain in excess of the race-model
predictions for right-hand responses, but not for left-hand
responses.
3.2.2. Touching the fixation cross
Table 3 shows the number of times, out of 30, that D.D.V
touched the fixation cross correctly, within 2 s of the
stimulus having appeared, shown separately for left-hand
and right-hand responses. His mean RTs are also shown. In
this condition, there was quite strong left hemineglect, with
Table 3
Number of touches on fixation cross and mean RT (standard error in
brackets) for each hand and each condition in Experiment 2
LVF RVF Bilateral
No. of responses (out of 30)
Left hand 13 23 24
Right hand 13 27 30
Mean RT (SE)
Left hand 1209 (58) 1352 (61) 1382 (59)
Right hand 1156 (74) 933 (62) 1135 (61)

Fig. 3. Distribution of the violation function ( pB � ( pL + pR � ( pL I pR))for RT in keyboard responses in Experiment 2. Violations of independent
race model for keyboard responses in Experiment 2 (D.D.V.).
M.C. Corballis et al. / Cognitive Brain Research 25 (2005) 521–530526
D.D.V. responding to LVF stimuli on only 13 out of 30 trials
with either hand. Mean RTs show a compatibility effect,
with faster responses to LVF than to RVF stimuli with the
left hand, and the opposite for the right hand, resulting in a
crossed–uncrossed difference of 183 ms.
There was no evidence of redundancy gain. We have no
explanation for this, but note that the number of responses
was very low compared to that in previous studies showing
redundancy gain in split-brained subjects.
3.2.3. Keyboard responses
The numbers of responses and mean RTs are shown for
each hand and visual field condition in Table 4. There was
no evidence of left hemineglect. Instead, there was some
weakness in response to stimuli in the RVF. Analysis of
variance of RTs revealed a significant main effect of field
condition, F(2, 159) = 11.58, P < 0.001, and a significant
interaction between field condition and hand, F(2, 159) =
3.79, P = 0.025. Overall, RTs were considerably longer to
RVF (362 ms) than to either LVF (265 ms) or bilateral (234
ms) stimuli. When the bilateral presentations were excluded,
the effect of field remained significant, F(1, 103) = 8.65, P =
0.004, indicating faster responses to LVF stimuli than to
RVF stimuli, and the interaction between hand and field was
also significant, F(1, 103) = 4.88, P = 0.029, indicating
longer RTs to crossed hand-field combinations than to
uncrossed ones. The computed crossed–uncrossed differ-
ence was 73 ms.
For both hands, RTs to bilateral presentations were
shorter to bilateral than to LVF or RVF presentations,
suggesting redundancy gain. However, when Expression
(1), the violation from the independent race model, was
plotted over responses bins, it exceeded zero only for left-
hand responses, as shown in Fig. 3. That is, interhemi-
spheric neural summation is implied only for left-hand
responses. This is in interesting contrast to what occurred
when the response was to touch the stimulus locations,
where neural summation was implied for right-hand
responses only.
3.3. Discussion
The results of this experiment largely confirm the
suggestion of Savazzi and Marzi [35] that D.D.V.’s left
Table 4
Number of keyboard responses and mean RT (standard error in brackets)
for each hand and each condition in Experiment 2
LVF RVF Bilateral
No. of responses (out of 30)
Left hand 29 28 30
Right hand 26 24 28
Mean RT (SE)
Left hand 236 (27) 406 (27) 219 (26)
Right hand 295 (28) 319 (29) 249 (27)
hemineglect is essentially eliminated when continuous
markers indicating the locations of the stimuli are provided.
The only exception occurred when D.D.V. was instructed to
respond by touching the fixation cross. Under this con-
dition, he responded to just under half of the LVF stimuli.
It is especially striking that the left hemineglect was no
longer present when D.D.V. responded by pressing the
spacebar since this condition essentially duplicates that in
which he previously showed LVF neglect [14]—except for
the presence of the markers. Furthermore, he was especially
slow in responding to RVF stimuli. This suggests that the
presence of the markers shifted control to the right hemi-
sphere, which controls attention to both sides of space,
whereas the left hemisphere controls attention largely if not
exclusively to the right side [19].
With respect to redundancy gain, the pattern of results is
somewhat inconsistent perhaps because the number of
responses under each condition was relatively low. Never-
theless, it is of interest that keyboard responses showed
redundancy gain only for the left hand, whereas touching
the stimulus locations showed a redundancy gain only for
the right hand. It is unclear what might explain this switch,
but it is perhaps noteworthy that in each case it occurred for
the hand that responded more quickly.
4. Experiment 3
In this experiment, we compare visual fields in a more
complex task to determine whether hemineglect might be
alleviated in a task requiring more cognitive resources. The
task was to decide whether pairs of filled circles flashed
either in the LVF, RVF, or on either side of fixation were of
the same or different color. This experiment is essentially a
continuation of that reported by Corballis and Corballis
[12], in which three split-brained subjects (other than
D.D.V.) performed at chance when the two circles were
flashed to opposite visual fields but were generally above
chance when they were flashed within either the left or the
right visual field.
Table 5
Number of ‘‘same–different’’ responses and number of correct responses,
both out of 96, for each subject under each condition in Experiment 3
J.W. D.D.V.
LVF RVF Bilateral LVF RVF Bilateral
No. of responses
(out of 96)
96 96 96 6 94 91
No. correct 88 86 54 5 94 45
M.C. Corballis et al. / Cognitive Brain Research 25 (2005) 521–530 527
4.1. Method
4.1.1. Subjects
D.D.V. was the principle subject of interest. For
comparative purposes, data from J.W. are also presented.
J.W.’s data are also available in the earlier publication by
Corballis and Corballis [12].
4.1.2. Stimuli
The stimuli (as also described in [12]) were filled red,
green, and blue circles of diameter 7.15 mm, presented
against a background of 5 cd/m2. The red and green circles
were matched for apparent luminance using flicker photo-
metry, and their luminances set at 23.36 and 28.28 cd/m2,
respectively. It proved impossible to match the luminance of
the blue to the other two colors using flicker photometry,
and its luminance was set at 13.47 cd/m2; although this was
close to its maximum, the blue appeared vivid but was
physically and subjectively darker than the other two colors.
The stimuli were presented in horizontal pairs centered
7.15 cm apart. For unilateral presentations, the near stimulus
was centered 3.575 cm from a central fixation cross. For
bilateral presentations, the two stimuli were centered 3.575
cm on either side of fixation. Each color pairing was
presented equally often, so there were equal numbers of
trials on which the stimuli were the same or different in
color.
4.1.3. Procedure
Stimulus presentations were initiated by the experimenter
pressing the ‘‘1’’ key on the computer keyboard. The
stimulus pair appeared 500 ms later and was on the screen
for 100 ms. The subjects responded by pressing the ‘‘N’’ key
if the colors were judged to be the same and the ‘‘M’’ key if
they were judged to be different, using the forefinger and
middle finger of a single hand. Each pair was presented 8
times in each visual field condition, making a total of 144
trials, which were randomly ordered.
Each subject received 10 practice trials followed by two
blocks of 144 experimental trials. J.W. performed the first
block with his left hand and the second with his right, while
this order was reversed for D.D.V. At least 3 s was allowed
for response.
4.2. Results
Table 5 shows the number of responses each subject
made under each condition, along with the number of
correct responses. Again, D.D.V. shows striking left hemi-
neglect but responded on most trials to bilateral or RVF
presentations. In contrast, J.W. responded on all trials.
Each subject’s responses were subjected to multidimen-
sional Chi-square [38] to determine the effects of stimulus
(same vs. different), location (LVF, bilateral, RVF), and
hand on the response selection (‘‘same’’ vs. ‘‘different’’). As
previously reported [12], J.W.’s responses depended sig-
nificantly on location, v2(1, N = 144) = 49, P < 0.001, and
were significantly above chance in the LVF (91.7% correct),
v2(1, N = 48) = 33.33, P < 0.001, and in the RVF (89.6%
correct), v2(1, N = 48) = 30.56, P < 0.001, but not with
bilateral presentation (56.3%).
D.D.V.’s responses to LVF presentations were too rare
for inclusion in the analysis, but it is noteworthy that, of the
6 responses (2 with the left hand and 4 with the right), 5
were correct, suggesting that when LVF stimuli were
attended to D.D.V. was capable of correct discrimination.
Multidimensional Chi-square computed on his responses to
bilateral and RVF stimuli revealed no significant effects of
hand. His responses were significantly related to whether
the stimuli were the same or different, v2(1, N = 185) =
52.93, P < 0.001, indicating above-chance performance
overall, and also on the visual field, v2(1, N = 185) = 26.91,
P < 0.001, indicating a strong bias to respond ‘‘same’’ to
bilateral stimuli (78 ‘‘same’’ responses vs. 17 ‘‘different’’
responses). More critically, the triple contingency between
field, same–different stimuli, and response was significant,
v2(1, N = 185) = 54.74, P < 0.001, indicating a significant
difference in accuracy between fields. He failed to respond
on 2 RVF trials but was 100% correct on the remaining 94
trials. On bilateral trials, he failed to respond 5 times but he
was correct on only 49.5% of the remaining 91 trials, which
is almost exactly at the chance level of 50%.
4.3. Discussion
In common with other callosotomized subjects [12],
including J.W., D.D.V. was at chance when making same–
different judgments about bilateral pairs, implying that
information about color cannot be matched subcortically. He
was, however, the most accurate of all callosotomized
subjects on stimuli in the RVF, indicating that he had no
difficulty understanding the task. Unlike the other subjects,
he seldom responded at all to LVF pairs, again reflecting
strong left hemineglect. Adding to the cognitive load of the
task therefore did not attenuate the left hemineglect.
5. General discussion
Earlier research showed that a callosotomized patient,
D.D.V., exhibits strong neglect of stimuli flashed in the LVF
in a simple RT task [14]. The present experiments show that
this neglect persists when D.D.V. is required to respond to

M.C. Corballis et al. / Cognitive Brain Research 25 (2005) 521–530528
LVF stimuli on a leftward key with his left hand and to RVF
stimuli on a rightward key with his right hand (Experiment
1) and when he is asked to judge pairs of circles to be the
same or different in color (Experiment 3). His hemineglect
in line bisection is also manifest regardless of which hand he
uses to bisect the lines [18]. It seems likely that performance
on these tasks is strongly dominated by the left hemisphere,
with consequent attention directed almost exclusively to the
right side of space.
The left hemineglect is no longer evident, however, when
there are fixed markers in the LVF and RVF to indicate the
locations of the stimuli. In the experiment by Savazzi and
Marzi [36], these markers consisted of squares containing
components that varied in luminance, in a twinkling fashion,
while in Experiment 2 of the present study the markers were
outline circles within which the filled circles that served as
stimuli would appear. In Experiment 2, not only was there
no evidence of left hemineglect when D.D.V. made key-
board responses, but also RTs were actually shorter to LVF
than to RVF stimuli, regardless of the hand used. This
suggests that the presence of markers may have shifted
control to the right hemisphere, with consequent attention to
both sides of space, albeit with something of a bias toward
the left. (Normal subjects typically show a slight bias to the
left in line bisection—so-called ‘‘pseudoneglect’’—suggest-
ing that attention directed by the right hemisphere is not
distributed entirely evenly [6].) Some support for the idea
that attentional control can shift from one hemisphere to the
other in the split brain is provided by evidence that a split-
brained patient, J.W., was faster to RVF stimuli when
making key-pressing responses but faster to LVF stimuli
when making target-directed saccades [20]. Levy and
Trevarthen [25] also showed that different instructions
could elicit left- or right-hemispheric control in the
interpretation of ambiguous pictures by split-brained sub-
jects, suggesting brainstem metacontrol of attention.
There was also little sign of hemineglect when D.D.V.
was asked to point to the locations of the stimuli [14] or
when he actually touched these locations (Experiment 2). It
is perhaps unlikely that this can be attributed to right-
hemispheric control since there was no indication that RTs
to LVF stimuli were faster than those to RVF stimuli—if
anything, they were slightly slower. An alternative possi-
bility is that pointing depends on the dorsal visual system
and keyboard responses on the ventral system [17]. There is
a cluster of neurons in the posterior parietal lobe of the
macaque specialized for reaching to a visible or remembered
target [8], and a similar system in the human brain may be
involved in pointing and touching.
Some degree of left hemineglect did return, however,
when D.D.V. was asked to touch the fixation cross rather
than the actual stimulus locations when a stimulus appeared
in either visual field, despite the fact that the markers were
present. Since this response was no longer the relatively
hard-wired act of reaching for or touching the location of an
event, control may have been returned to the ventral system,
and attention to the fixation cross for response as well as for
stimulus presentation may have favored left- rather than
right-hemispheric control. It is possible that this task may
have increased the demand on cognitive resources since RTs
were very slow. D.D.V. also showed strong neglect in
making same–different judgments in Experiment 3, a task
that presumably made greater demands on cognitive
resources than any of the other tasks. It is not clear,
however, whether hemineglect in these cases was due to
left-hemispheric control or to increased cognitive demands
per se. The circumstances under which one or other
hemisphere assumes control are not fully understood and
may in fact be fickle. Nevertheless, the results, taken
together, suggest three possible mechanisms of control—
one left-hemispheric, one right-hemispheric, and one via the
dorsal visual system.
Most researchers have interpreted hemineglect in terms
of an attentional imbalance (e.g., [3,4,6,19,24,28]), as have
we. There are nevertheless other possible interpretations.
Some of these interpretations relate to impaired registration
of a stimulus in the damaged hemisphere, whether due to
low arousal [19] or to disturbed mental imagery [5]. In the
case of D.D.V., there is no evidence of right-hemispheric
impairment, and his ability to respond to LVF stimuli when
markers were present, or to point to LVF stimuli, suggests
that his perception of LVF stimuli is intact. His failure to
make keyboard responses to LVF stimuli when no markers
are present therefore seems better attributed to attentional
failure rather than a failure of perception or registration.
Another possibility is that neglect is due to failure to
disengage from a previously attended object and move
attention to the left [30]. Perhaps, then, D.D.V.’s hemi-
neglect, when it occurs, is due to attention to the fixation
cross and subsequent failure to move his attention to the left.
This might be tested by removing the fixation cross.
Aside from his unusual degree of hemineglect, D.D.V.
showed other typical signs of disconnection. Like other
split-brained patients, he was unable to compare colors
between visual fields at better than chance [12,23]. In the
earlier study, the crossed–uncrossed difference in RT, as a
measure of interhemispheric transfer time, could not be
calculated due to the paucity of responses to LVF stimuli.
Experiment 2, however, provided three estimates. When the
response was to touch the locations, the estimate was 111
ms; when it was to touch the fixation cross, it was 183 ms;
when it was to press the spacebar, it was 78 ms. The last of
these is probably the most valid since RTs were relative
short and the responses did not involve movements of the
hand. In any event, as in previous studies of split-brained
patients [1,10,21], these values are clearly well in excess of
those typically obtained from normal subjects, which are
typically in the range of 2–6 ms [2,7,11].
D.D.V. also showed evidence of enhanced redundancy
gain, characteristic of split-brained patients but not of
normal subjects [10,13,22,26,33,34]. In Experiment 1, as
in the earlier study by Corballis et al. [14], this effect was

M.C. Corballis et al. / Cognitive Brain Research 25 (2005) 521–530 529
present despite neglect of the LVF stimuli, adding to
evidence that redundancy gain can occur even when one
of the stimuli is below response threshold [35]. Experiment
2 was more equivocal; D.D.V. showed redundancy gain
when touching the locations of the stimuli with his right
hand, but not when touching with his left hand, and when
pressing the spacebar with his left hand, but not when
touching with his right hand. In both cases, redundancy gain
was associated with the hand showing the faster responses.
These results raise the possibility that redundancy gain may
vary depending on the neural locus of control, but it will
require further study to clarify this.
In summary, the most important finding is that the striking
left hemineglect shown by the callosotomized patient D.D.V.
is sometimes present, sometimes not, depending on the
conditions. It appears to be present when response is under
control of the left cerebral hemisphere, but not when it is
under the control of the right hemisphere. It is also absent
when the subject points to or touches the locations of stimuli
on the screen perhaps because these responses are under the
separate control of the dorsal visual system. It is also clear that
D.D.V.’s hemineglect persisted when he was required to
make same–different judgments, indicating that hemineglect
is not diminished, and is, if anything, increased, by increasing
demands on cognitive resources.
Acknowledgments
We thank Gabriella Venanzi and Gabriele Polonara for
arranging the testing of D.D.V., and we also thank D.D.V.
for his willing cooperation. We also thank Margaret Francis
for writing the programs for Experiment 2. This research
was supported by grants from the Marsden Fund of New
Zealand and MIUR, Cofin 2003 in Italy.
References
[1] S. Aglioti, G. Berlucchi, R. Pallini, G.F. Rossi, G. Tassinari,
Hemispheric control of unilateral and bilateral responses to lateral-
ized light stimuli after callosotomy and in callosal agenesis, Exp.
Brain Res. 95 (1993) 151–165.
[2] G. Berlucchi, W. Heron, R. Hyman, G. Rizzolatti, C. Umilta, Simple
reaction time of ipsilateral and contralateral hand to lateralized stimuli,
Brain 94 (1971) 419–430.
[3] G. Berlucchi, S. Aglioti, G. Tassinari, Rightward attentional bias and
left hemisphere dominance in a cue– target light detection task in a
callosotomy patient, Neuropsychologia 35 (1997) 941–952.
[4] G. Berlucchi, G.R. Mangun, M.S. Gazzaniga, Visuospatial attention
and the split brain, News Physiol. Sci. 12 (1997) 226–231.
[5] E. Bisiach, M. Neppi-Modona, R. Genero, R. Pepi, Anisometry of
space representation in unilateral neglect: empirical test of a former
hypothesis, Conscious Cogn. 8 (1999) 577–584.
[6] D. Bowers, K.M. Heilman, Pseudoneglect: effects of hemispace on a
tactile line bisection task, Neuropsychologia 18 (1980) 491–498.
[7] C.M.J. Braun, Estimates of interhemispheric dynamics from simple
unimanual reaction time to extrafoveal stimuli, Neuropsychol. Rev. 3
(1992) 321–365.
[8] J.L. Calton, A.R. Dickinson, L.H. Snyder, Non-spatial, action-specific
activation in posterior parietal cortex, Nat. Neurosci. 5 (2002) 580–588.
[9] M.C. Corballis, Line bisection in a man with complete forebrain
commissurotomy, Neuropsychology 9 (1995) 147–156.
[10] M.C. Corballis, Interhemispheric neural summation in the absence of
the corpus callosum, Brain 121 (1998) 101–113.
[11] M.C. Corballis, Hemispheric interactions in simple reaction time,
Neuropsychologia 40 (2002) 423–434.
[12] M.C. Corballis, P.M. Corballis, Interhemispheric visual matching in
the split brain, Neuropsychologia 39 (2001) 1395–1400.
[13] M.C. Corballis, J.P. Hamm, K.J. Barnett, P.M. Corballis, Paradoxical
interhemispheric summation in the split brain, J. Cogn. Neurosci. 14
(2002) 1151–1157.
[14] M.C. Corballis, P.M. Corballis, M. Fabri, Redundancy gain in simple
reaction time following partial and complete callosotomy, Neuro-
psychologia 42 (2004) 71–81.
[15] M. Fabri, G. Polonara, A. Quattrini, U. Salvolini, M. Del Pesce, T.
Manzoni, Role of the corpus callosum in the somatosensory activation
of the ipsilateral cerebral cortex: an fMRI study of callosotomized
subjects, Eur. J. Neurosci 11 (1999) 3983–3994.
[16] M.S. Gazzaniga, J.D. Holtzman, M.D.E. Deck, B.C.P. Lee, MRI
assessment of human callosal surgery with neuropsychological
correlates, Neurology 35 (1985) 682–685.
[17] M.A. Goodale, A.D. Milner, Separate visual pathways for perception
and action, Trends Neurosci. 15 (1992) 20–25.
[18] M. Hausmann, M.C. Corballis, M. Fabri, Line bisection in the split
brain, Neuropsychology 17 (2003) 602–609.
[19] K.M. Heilman, R.T. Watson, E. Valenstein, Neglect and related
disorders, in: K.M. Heilman, E. Valenstein (Eds.), Clinical Neuro-
psychology, 3rd edition, Oxford University Press, Oxford, 1993,
pp. 274–336.
[20] H.C. Hughes, P.A. Reuter-Lorenz, R. Fendrich, M.S. Gazzaniga,
Bidirectional control of saccadic eye-movements by the disconnected
cerebral hemispheres, Exp. Brain Res. 91 (1992) 335–339.
[21] M. Iacoboni, E. Zaidel, Channels of the corpus callosum: evidence
from simple reaction time to lateralized flashes in the normal and the
split brain, Brain 123 (1995) 759–769.
[22] M. Iacoboni, A. Ptito, N.Y. Weekes, E. Zaidel, Parallel visuomotor
processing in the split brain: cortico-subcortical interactions, Brain
123 (2000) 759–769.
[23] L.E. Johnson, Bilateral cross-integration by human forebrain commis-
surotomy subjects, Neuropsychologia 22 (1983) 167–175.
[24] R.J. Joynt, Inattention syndromes in split-brained man, Adv. Neurol.
81 (1977) 33–39.
[25] J. Levy, C. Trevarthen, Metacontrol of hemispheric function in human
split-brain patients, J. Exp. Psychol.: Hum. Percept. Perform. 2 (1976)
299–312.
[26] C.A. Marzi, N. Smania, M.C. Martini, G. Gambina, G. Tomelleri, A.
Palamara, et al., Implicit redundant-target effects in visual extinction,
Neuropsychologia 24 (1997) 749–758.
[27] J. Miller, Divided attention: evidence for coactivation with redundant
signals, Cogn. Psychol. 14 (1982) 247–279.
[28] E. Natale, L. Posteraro, M. Prior, C.A. Marzi, What kind of spatial
attention is impaired in neglect? Neuropsychologia 43 (2005)
1072–1085.
[29] G. Plourde, R.W. Sperry, Left hemisphere involvement in left spatial
neglect from right-sided lesions: a commissurotomy study, Brain 107
(1984) 95–106.
[30] M.I. Posner, S.E. Peterson, The attention system of the human brain,
Annu. Rev. Neurosci. 13 (1990) 25–42.
[31] A.M. Proverbio, A. Zani, M.S. Gazzaniga, G.R. Mangun, ERP and RT
signs of a rightward bias for spatial orienting in a split-brain patient,
NeuroReport 5 (1994) 2457–2461.
[32] D. Raab, Statistical facilitation of simple reaction times, Trans. N. Y.
Acad. Sci. 24 (1962) 574–590.
[33] P.A. Reuter-Lorenz, G. Nozawa, M.S. Gazzaniga, H.C. Hughes, Fate
of neglected targets: a chronometric analysis of redundant target

M.C. Corballis et al. / Cognitive Brain Research 25 (2005) 521–530530
effects in the bisected brain, J. Exp. Psychol. Hum. Percept. Perform.
21 (1995) 211–230.
[34] M. Roser, M.C. Corballis, Interhemispheric neural summation in the
split brain with symmetrical and asymmetrical displays, Neuro-
psychologia 40 (2002) 1300–1312.
[35] S. Savazzi, C.A. Marzi, Speeding up reaction time with invisible
stimuli, Curr. Biol. 12 (2002) 403–407.
[36] S. Savazzi, C.A. Marzi, The superior colliculus subserves interhemi-
spheric neural summation in both normals and patients with a total
section or agenesis of the corpus callosum, Neuropsychologia 42
(2004) 1608–1618.
[37] F. Tomaiuolo, M. Ptito, C.A. Marzi, T. Paus, A. Ptito, Blindsight in
hemispherectomized subjects as revealed by spatial summation across
the vertical meridian, Brain 120 (1997) 795–803.
[38] B.J. Winer, D.R. Brown, K.M. Michels, Statistical Principles in
Experimental Design, 3rd edition, McGraw-Hill, New York, 1991.





























