Embed Size (px)
Citation preview

Cell. Vol. 33, 543-553. June 1983, CopyrIght 0 1983 by MIT
Novel Amplification and Transcriptional Activity of Chorion Genes in Drosophila melanogaster Follicle Cells
0092-8674/83/060543-i 1 $02.00/O
Yvonne Nelson Osheim and 0. L. Miller, Jr. Department of Biology University of Virginia Charlottesville, Virginia 22901
Summary
Single-copy chorion genes coding for egg shell pro- teins are amplified in the follicle cells of Drosophila melanogaster egg chambers. Chromatin spreads of appropriately staged follicle cells reveal complex, multi-forked chromosomal structures in which one chromatin strand branches into two, which them- selves branch out, and so on. In one micrograph, 13 strands originating from a single strand were ob- served. Tliese structures can account for the maxi- mal amplification occurring in the center of the do- main, where the major chorion genes are located, and the decreasing amplification of flanking se- quences to both sides. The amplification, high tran- scriptional rate, stage-specific expression, and cor- relation with known molecular sizes have allowed the putative identification of the single-copy, major chorion genes on the X chromosome and on chro- mosome Ill.
Introduction
In Drosophila melanogaster, the proteins comprising the chorion, or egg shell, are synthesized in the follicle cells surrounding the egg chamber. The 20 or so chorion pro- teins share many structural features and range from 15 to 150 kd. (Waring and Mahowald, 1979; Spradling et al., 1980). Utilized in large quantities over a 5 hr period late in oogenesis, the various chorion proteins are translated in a stage-specific, temporally controlled manner and are de- posited along the inner surface of the follicle cell next to the vitelline membrane, where they form the endochorion and exochorion (Petri et al., 1976; Kafatos et al., 1977; Waring and Mahowald, 1979).
One process that facilitates such high rates of protein production is a preferential amplification in the follicle cells of the single-copy genes coding for chorion proteins (Spra- dling and Mahowald, 1980). The amplification phenome- non begins during stage 8 and continues through stage 12. One domain containing several early-acting chorion genes on the X chromosome is amplified 14 to 16 fold, and a second domain on chromosome III, containing four genes expressed late in choriogenesis, is amplified 60-fold (Spradling and Mahowald, 1980; Spradling, 1981; Griffin- Shea et al., 1982). Each of the two amplified domains extends over 90-100 kb and is maximally amplified in the central region where the major chorion genes are located, with the level of amplification of flanking sequences grad- ually decreasing in both directions (Spradling, 1981).
The mechanism by which amplification occurs has not been determined biochemically. Two extremes that are
compatible with biochemical data are that replicons open up within replicons along the chromatin strand and that the amplified sequences exist as extrachromosomal linear or circular molecules of varying sizes. The existence of a mutant, ocelliless, in which a small chromosome inversion in the X chromosomal domain changes the sequences that are amplified and affects the levels of amplification, suggests that a unique origin for replication may exist in the domain (Spradling and Mahowald, 1981).
A second process that accelerates the production of chorion proteins is the prolific activation of chorion mRNA synthesis in late stage follicle cells (Spradling and Mahow- ald, 1979). mRNA for a particular chorion protein appears in the cytoplasm, coincident with the onset of its transla- tion, and turns over rapidly as production of the protein ceases (Waring and Mahowald, 1979; Spradling and Ma- howald, 1979). For example, mRNA precursors for two abundant chorion proteins, ~36-1 and ~38-1, are tran- scribed from the X chromosome domain mainly during stage 12, while only 2 hr later, at stage 14, transcription predominantly involves the genes for proteins ~15-1 and ~18-1 on chromosome III. (The nomenclature for chorion proteins consists of the prefix “s” for shell, the molecular weight in kilodaltons, and a final digit distinguishing major and minor components at the same molecular weight.) The high levels of mRNA for specific genes that accumu- late over short time periods suggest that the genes must be maximally loaded with RNA polymerase II molecules (Kafatos, 1972).
These two features-the selective amplification of DNA coding for chorion genes and the temporally regulated expression of closely linked genes of the same gene family-provide a useful system for studying the mecha- nism of amplification occurring in the cells and the regula- tion of transcription of linked, single-copy genes. It is usually impractical to study transcriptionally active single- copy genes with the electron microscopy because of the extremely low probability of locating a particular gene in the chromatin of an active nucleus and because, in many cases, no means of clearly identifying particular genes are avallable. The biology of the Drosophila chorion genes suggested that both of these problems could be substan- tially reduced. Amplification should greatly increase the probability of visualizing the genes. Since the genes are present in gene clusters whose topography and times of expression are known, unambiguous recognition of these genes should be possible. Therefore, the electron micro- scope was used to examine ultrastructurally the chromatin from appropriately staged follicle cells, with the objects of elucidating the mechanism of amplification and analyzing the transcriptional morphology of these genes.
Results and Discussion
Identification of Major Chorion Genes on the X Chromosome To examine amplification and transcription of chorion gene sequences, chromatin was spread from the homogeneous

Cell 544
population of follicle cells surrounding the oocyte (Mahow- ald and Kambysellis, 1980). In spreads from stage 11 and stage 12 follicle cells, prominent clusters of short, closely linked transcription units are frequently observed. The two genes are densely packed with RNA polymerase mole- cules and consist of distinct matrix units of nascent RNA fibrils of progressively greater length (Figure 1).
We have carefully analyzed 20 of these pairs of short transcription units observed on chromatin derived from stage 11 and stage 12 follicle cells. In every case, the adjacent genes were found to be transcribed in the same direction. The upstream, shorter gene averages 0.36 pm in length (n = 20, 0.06 SD) with an average of 13 polym- erase molecules per gene, The longer, downstream gene covers 0.56 pm (n = 20, 0.07 SD), and is loaded with an average of 19 polymerase molecules. A nontranscribed chromatin segment, averaging 0.35 pm in length, sepa- rates the genes. The morphology of the nascent RNP fibrils on the two genes is characterized by a granular, electron dense knob, averaging 350 A in diameter, at the distal tip of each fibril. A short distance in from the distal knob, a second, smaller particle is often present.
The clusters of gene pairs tend not to disperse easily. The tandem set of transcription units is most frequently observed along the periphery of a partially spread nucleus, where scores of electron-dense RNA polymerase mole- cules and knob-like RNA fibrils overlay each other and inactive chromatin strands. In an attempt to improve chro- matin dispersal, we partially digested samples with the restriction enzyme Hind Ill before spreading. The enzyme cuts the chromatin both upstream and downstream from the gene pairs. Figure 2 shows a cluster of gene pairs cut away from the bulk of the chromatin. Because the salt levels required to get a reasonable number of restriction cuts cause nucleoprotein precipitation, the nascent RNP fibrils appear as dark, beaded molecules condensed along the chromatin axis. This was also observed when Xenopus laevis ribosomal RNA transcription units were cut with restriction enzymes (Reeder et al., 1976). Whereas the salt precipitation presents a drawback to ultrastructural reso- lution, the effects of restriction digestion have facilitated several quantitations. Two hundred and four actively tran- scribing gene pairs were counted in a gene cluster, char- acteristic of stage 11 and stage 12 chromatin. Figure 2 shows a portion of this cluster. Since by stage 11, follicle cells have completed their last round of DNA synthesis and reached a ploidy of 45C (Mahowald et al., 1979) the existence of 200+ gene pairs in one partially dispersed cluster indicates that we are seeing more transcription than can be accounted for by one transcribed gene pair per homolog. Additionally, these gene pairs are never ob- served in spreads of follicle cells from stage 10 or earlier, indicating that their activation is under stage-specific con- trol.
We identify the two transcription units as active copies of ~36-1 and ~38-1 chorion protein genes, based on their congruence with the molecular anatomy of the genes. s36-
1 and ~38-1 are positioned at 7Fl-2 on the X chromosome and code for highly abundant chorion proteins (Waring and Mahowald, 1979; Spradling et al., 1979; Spradling and Mahowald, 1979). mRNA molecules from the two genes first appear during stage 11 and disappear by the end of stage 13 when production of ~36-1 and ~38-1 proteins ceases (Waring and Mahowald, 1979; Spradling and Ma- howald, 1979). The two gene products accumulate at approximately the same rate and show the same temporal developmental expression (Waring and Mahowald, 1979). Molecular analysis of genomic DNA indicates that ~36-1 and ~38-1 are transcribed from the same strand and that ~36-1 lies upstream from ~38-1 (Spradling, 1981). Figure 3 diagrams the arrangement and spacing of the two genes, as well as their position within the amplified domain on the X chromosome. Several other chorion genes coding for less abundant proteins are present in the X chromosomal domain, but their lengths and relative positions are un- known (Spradling, 1981).
The shorter, upstream transcription unit that we observe in chromatin spreads corresponds to ~36-1, and the longer, downstream unit corresponds to ~38-1. The transcription units occupy the same relative positions as ~36-1 and s38- 1, with the shorter gene located upstream from the longer gene. The spacer between the two genes extends for 0.35 pm, very near the 0.31 pm length measured by Spradling (1981). In addition, both genes are highly transcribed from the same DNA strand and share a common RNP mor- phology. The tandem transcription units appear in large clusters that are only observed during the stages when ~36-1 and ~38-1 are expressed. Moreover, the fact that they occur in greater numbers than the chromosomal ploidy suggests that they are amplified. The genes ob- viously are not rRNA genes, in that they are not arranged as multiple tandem repeats with varying spacer lengths (Glover and Hogness, 1977; White and Hogness, 1977; Wellauer and Dawid, 1977; and Pelligrini et al., 1977) and do not approach the 2.55 pm length reported for active rRNA genes in Drosophila melanogaster embryos (Mc- Knight and Miller, 1976; Long and Dawid, 1979).
Gene lengths are given in micrometers and have not been corrected for compaction ratios based on the chro- mosome condensation characteristic of this EM prepara- tive technique. An earlier study by McKnight et al. (1978) suggests that the degree of DNA condensation in a tran- scription unit is inversely proportional to the number of RNA polymerase molecules attached to the chromatin template. Minimally transcribed transcription units exhibit- ing beaded elements between RNA polymerase molecules were estimated to contain 4.8 kb/pm, whereas maximally transcribed chromatin associated with 30 RNA polymerase molecules/km is extended to nearly B-form, or 3.3 kb/pm. Since presumptive ~36-1 and ~38-1 average 36 and 33 polymerases per pm, respectively, the chromatin template may be more extended than previously observed. Calcu- lating the number of nucleotides covered by an RNA polymerase II molecule gives 80 to 90 nucleotides per

r$rion Gene Amplification and Transcription
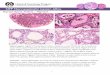
Figure 1. Transcription of Chorion Genes on the X Chromosome
Each electron micrograph shows a cluster of hrghly transcribed chorion genes from a stage 12 follrcle cell. Occasionally, one can trace a chromatrn strand and see two genes, transcribed in the same directron (arrows). Both genes are usually fully loaded with RNA polymerase molecules and share an RNP structure charactenzed by electron-dense terminal knobs. Bars = 0.5 Mm.

Cdl 546
Figure 2. Large Cluster of Short, Actiie Genes
A cluster of chorion genes from a stage 11 follicle cell treated with Hind Ill has, for the most part, been cut away from the bulk of the chromatin. The dark, particulate regions along the chromatin strands are actively transcribed genes whose nascent transcripts have been somewhat precipitated by the relatively high salt concentration of the digestion medium. Arrows point to gene pairs tentatively identifed as ~36-1 (shorter gene) and ~38-1 (longer gene). Twenty-four measurements of chromatin lengths from the free, cut ends to the active genes suggest that many of the restriction cuts are nonspecific, although some specific restriction sites (arrowhead) also appear to have been cleaved. Precise determinations of specific and nonspecific cuts are difficult, since the salt levels required greatly affect the degree of chromatin compaction. Depending on the compaction ratio used (see text for discussion), varying results can be obtained. Ear = 1 Mm.
polymerase for ~38-1 and ~36-1, compared to 110 nucleo- tides if the 3.3 kb/pm compaction ratio is used.
Identification of Chorion Genes on Chromosome Ill Chromatin spreads from stage 13 follicle cells also show large clusters of active genes (Figure 4a), which differ in length and spacing from those observed in stages 11 and 12. The transcription units are shorter, averaging 0.35 pm in length (n = 43; 0.06 SD), and are separated by spacers averaging 0.26 pm (n = 15; 0.05 SD). We most commonly see two transcription units transcribed in the same direc- tion, but the occasional presence of a single nascent transcript downstream suggests that there are at least three genes at the locus (Figure 4~). Most of these genes are completely covered with nascent RNP fibrils, and the distal ends of the fibrils are organized into granular particles averaging 270 A in diameter. Typically, there is a second, smaller particle averaging 230 A in diameter immediately adjacent to the distal particle. Gene clusters with genes of these sizes are not observed in chromatin spreads from follicle cells of stage 12 and earlier.
Based on the presence of large clusters of these short
transcription units, their similar RNP morphology, and their stage-specific expression, we conclude that they corre- spond to chorion genes at the 66 Dl l-15 locus of chro- mosome III (Figure 5). Three genes known to be tran- scribed from this locus share the same polarity and extend 0.29 pm, 0.26 pm, and 0.27 pm; the genes are separated from each other by spacers of 0.24 pm (Griffin-Shea et al., 1982; Spradling, 1981). The fourth gene at the locus covers 0.24 pm and is separated from the other three genes by 0.41 pm. The transcriptional polarity of this fourth gene is not known (Griffin-Shea et al., 1982). All four genes are expressed during stage 13, but gene products accumulate to differing degrees during stages 13 and 14 (Griffin-Shea et al., 1982; Spradling and Mahowald, 1979; Waring and Mahowald, 1979). Thus because of the similarities in size, spacing, and time of expression of the genes, we cannot unambiguously designate which specific genes have been observed. A further complication is that all nuclei and/or loci within a cluster may not express the same subset of genes. Like the major chorion genes on the X chromo- some, the major chorion genes on chromosome Ill reside in the center of the amplified domain (Spradling, 1981).

Chorion Gene Amplification and Transcription 547
X Chromosome
S36-l 838-l
Figure 3. Map of the Amplified Chorion Domain on the X Chromosome
A schematic representation shows the locus containing the X chromosome genes coding for two major chorion proteins and located at 7Fl-2. Two genes at this locus, ~36-1 and ~36.1, code for very abundant egg shell proteins. These genes are maximally expressed during stages 11 and 12 of egg chamber development and are transcribed from the same strand. Upstream and downstream from the genes are other regions codrng for poly(A)+ chorion mRNAs that are far less abundant and which have not been characterized (see dark boxes). The upstream region may contain 4 to 8 genes (Parks and Spradling. 1961). Lengths of genes and spacers are given in micrometers. The chromatrn containing the chorion genes is preferentrally amplrfied 14 to 16 fold above background DNA levels. Amplr- fication IS maximum at the center of the locus, and then decreases to background levels some 45 kb rn erther direction away from the genes (Spradling, 1961).
Novel Mechanism of Amplification of Chorion Genes The last rounds of preferential chorion domain amplification occur during stage 11 and early in stage 12. Concomi- tantly, the chorion genes ~36-1 and ~38-1 on the X chro- mosome become activated. We have therefore concen- trated on spreading follicle cell chromatin from these two stages in order to see recognizable gene pairs in the process of amplification.
In spreads from stage 11 follicle cells, replication forks in the vicinity of ~36-1 and ~38-1 are occasionally observed. Figure 6 exhibits at least two replication forks downstream from ~38-1. The two strands arising from one of the forks traverse 1.26 pm and 1.27 Mm before encountering the 3’ ends of ~38-1. Regions of a gene cluster partially digested with Hind Ill provide additional examples of forks near genes. Figures 7a, 7b, and 7c exhibit replication forks 0.27 pm, 0.51 pm, and 0.94 pm, respectively, from the 5’ ends of ~36-1. Because we have observed replication forks that were moving away from the genes, both upstream from the transcription unit encoding ~36-1 and downstream from the transcription unit encoding ~38-1, a rough approxima- tion can be made of where the origin of replication is located, if one fixed origin exists. The origin must be positioned near the genes, that is, between 0.27 pm 5’ to 236-l and 1.26 pm 3’ to ~38-1, a distance extending 2.80
pm, based on the positions of forks in Figure 6 and Figure 7a. All electron micrographs exhibiting forks in the vicinity of the major chorion genes are consistent with the exist- ence of a fixed origin of replication very near ~36-1 and ~38-1.
When follicle cell chromatin from stage 11 is well dis- persed, complex, multi-forked structures are occasionally observed. The periphery of the structure reveals a single chromatin strand that branches into two strands, with each of the resulting strands branching again, and so on. Figure 8a shows a structure in which 13 chromatin strands arise from one strand. We assume that each branch represents either an active or a quiescent replication fork. The repli- cation forks do not occur at the same positions along the chromatin axis, suggesting that the process of amplifica- tion is not synchronous within a replicating structure. It is noteworthy that a transcript is apparent on one of the amplified strands (arrowhead), but not on homologous strands. A schematic drawing of the amplifying structure is shown in Figure 8b.
Our measurements of lengths of amplified chromatin strands are minimal, since the strands become increasingly entangled as replication forks multiply and the chromatin strands become impossible to follow. Tracing from the first replication fork along the various branch points (until the strands become too entangled to discern) gives distances extending 8.09 to 10.86 pm. The maximum observed strand length of 10.86 pm corresponds to 31.9 kb of B- form DNA, or 60.6 kb if the DNA is compacted 1.9-fold into nucleosomal chromatin, as has been reported for nontranscribed Drosophila embryonic chromatin (Mc- Knight and Miller, 1977). Because a restriction enzyme was added to these preparations, the salt level was higher than is normally used in chromatin spreading. It is impos- sible, therefore, to know how many proteins were removed or how this has affected nucleosome unraveling and, hence, the compaction ratio. Chromatin stretching during grid preparation can also influence the compaction ratio. The multi-forked structure in Figure 8a shows great varia- tion in nucleosome periodicity along the DNP strands- some short regions appear nucleosome-free, while other regions exhibit typical nucleosome arrays. Two independ- ent determinations of nucleosomes per pm have been performed on each of two homologous strands, One strand is complexed with 15.8 nucleosomes/pm, giving a compaction ratio of 1.0. The second strand contains 29 nucleosomes/Km, with a compaction ratio of 1.8 (see Figure legend 8a for compaction ratio formula). If we assume that most strands have a compaction ratio some- where between the two extremes and assign an average compaction ratio of 1.4, the amount of chromatin in the 10.86 pm multi-forked structure is 45 kb. According to Spradling (1981) the amplified domains extend 45 and 50 kb in each direction, which suggests that we have visual- ized about one half of an amplified domain.
The distances between replication forks from six elec- tron micrographs exhibiting multi-forked structures have

Cell 548
d
Figure 4. Transcription of Chonon Genes on Chromosome III
Short, active, tandem genes (a and c) are present in these chromatfn spreads from stage 13 foflicle cells. The genes are transcribed from the same strand and are characterized by terminal knobs on nascent transcripts. The line drawing in (b) points out three gene pairs; however, additional genes may lie uostream or downstream from the cairs and be obscured bv otfrer chromatin strands. A lone, nascent transcript is apparent downstream from one of the gene pairs in (c) (see arrow and the’line drawrng d). Sars = 6.2 pm.
been tabulated and are presented in Table 1. The dis- tances from the first to second replication forks average 2.9 Km. The distances between forks then decrease (ex- cluding the distance between the fifth and sixth replication forks where n = 2). However, individual amplified structures exhibit a great degree of variability in the lengths of chro- matin between forks.
Given that the central region of chromosome III is ampli- fied 60-fold and that the amplification extends 100 kb (Spradling, 1981) one would predict that the distances between replication forks would be less than for the X chromosome, which is amplified only 14 to 16 fold over 90 kb (Spradling, 1981). It should therefore be possible to discriminate between amplified structures on chromosome Ill and on the X chromosome. Unfortunately, observations are too limited at present to test this prediction. It seems most likely, however, that the structure in Figure 8a corre- sponds to one half of the amplified domain on the X chromosome, based on the number of strands observed and the distance covered by the structure. Since the structure was spread from a very early stage 11 follicle cell, it is probable that transcription of the two major chorion genes has not yet been activated. Alternatively, the struc- ture could represent a portion of the amplified domain on
chromosome Ill, where the chorion genes are not ex- pressed until stage 13.
We cannot determine whether or not the replication forks visualized were actually moving at the time of isola- tion, or if some or all were stationary. The first round of chorion gene amplification begins in stage 8, approxi- mately 15 hr earlier than stage 11 from which the electron micrographs in Figures 7 and 8 were obtained (Spradling, 1981; Mahowald and Kambysellis, 1980). If DNA replication in polyploid follicle cells is bidirectional and follows the same kinetics as replication in Drosophila virilis salivary gland cells, fork movement would proceed at 0.1 pm/min at 25“C (Steinemann, 1981). Replicon lengths in D. virilis salivary gland cells average 46.7 pm (Steinemann, 1981) which is only slightly larger than the 30.6 pm amplified region on the X chromosome and the 34 pm amplified region on chromosome Ill (using 2.94 kb B-form DNA/Mm). It would therefore take approximately 2.6 hr to complete the first round of amplification on the X chromosome and only a bit longer (2.8 hr) on chromosome Ill. Each succes- sive round of amplification would require less time, since a shorter distance would be traversed by each replication fork. According to Spradling and Mahowald (1980) ampli- fication continues from stage 8 through stage 13. The

Chorion Gene Amplification and Transcription 549
Chromosome I I I
918-l 915-l 919-l 9164
\ ‘\\
I’ /’ \ \
‘\\ //’
,’ ‘\ /
‘\ /’ \ / \ / \ I
-00 -40 -20 0 m 40 04 kb
Figure 5. Map of the Amplified Chorion Domain on the Thrrd Chromosome
This dragram shows the chorion locus at 6601 i-15 on chromosome III. The locus contains four short, linked genes, which are expressed during stages 13 and 14. Three of the genes are transcribed from the same strand; the drrection of transcription has not been determined for the fourth gene, ~16. 1. Gene and spacer lengths are given in mcrometers. The region containrng the four genes is amplified 60.fold above background (from GriffInShea et al., 1982; Spradlrng, 1981).
central, maximally amplified region encompasses IO-20 kb of DNA, and the flanking regions decrease in DNA content as a smooth continuum (Spradling et al., 1981). On the X chromosome, the entire region could be com- pletely amplified in about 6.4 hr if each successive round of amplification begins shortly after the previous forks stop moving and if the bidirectional forks stop at equidistant positions relative to the putative origin of replication we have observed. Chromosome Ill amplification would require 9.9 hr, given the above conditions, suggesting that the next round of amplification does not begin until the pre- vious round has been complete for some time.
The complex, multi-forked structures observed are con- sistent with a mechanism in which DNA amplification initiates at an origin of replication located very near the major chorion genes, and bidirectional replication then proceeds from that origin. Sometime after the replication forks have stopped moving, a second initiation event occurs at the origin. The corresponding replication forks cover less distance before ceasing replication than in the preceding replicon. Multiple initiation events continue within this structure until the centrally located major chorion genes are maximally amplified (4-6 rounds). Within homologs of one complex structure, replication forks are located at
Figure 6. Replication Forks Downstream from Active ~38-1 Genes
Frgure 6 shows several gene pairs of ~36-1 and ~38-1 from stage 11 follicle cells. A close examination of the lower region reveals at least two replication forks (large arrows). The two strands arising from one of these forks each lead to the 3’ ends of ~38-1. Each pair of opposing small arrows points to homologous genes. Bar = 0.2 pm.

Figure 7. Replicatron Forks Upstream from Active s36-1 Genes
The chromatin was treated with Hind III before spreading and displays cuts in the vicinity of replication forks near active genes, In (a), the replication fork is 0.27 Am from the 5’ ends of sister s361 genes (see arrow). (b) shows a replication fork 0.51 pm from ~36-1 genes. Note that the two sister strands have been cut at the same position (arrowheads). The arrow in (c) indicates a replication fork that is 0.94 pm from the 5’ ends of ~36-1. Note
Table 1. Distances between Replication Forks in Micrometers
Electron Micrographs
Forks
1st 2nd and and 2nd 3rd
3rd 4th 5th and and and 4th 5th 6th
average
n
Standard Deviation
2.9 1.6 1.1 0.8 0.8 = 1.57
16 20 19 16 2 = 73
1.4 1.4 0.8 0.4 0.1
The distances between replication forks in micrometers from six electron micrographs showing multi-forked structures were calculated. When DNP fibers downstream from a replication fork could no longer be measured because of blockage by a grid bar or because they were entangled in chromatin, the partial distance was used. Thus the distances are, in some cases, underestimates of the actual average distances between replication forks.
various positions along the chromatin strands, suggesting that fixed termination sites of DNA replication do not exist. Moreover, if one assumes that the rate of replication is the same for each fork, then it follows that DNA initiation events do not occur with synchrony within an amplifying structure. These observations, obtained from ultrastructural exami- nation of spread follicle ceil chromatin, are remarkably compatible with biochemical evidence for differential am- plification of chorion genes reported by Spradling and Mahowald (1980) and Spradling (1981).
Less complicated structures in which a second round of replication is initiated before the first round is completed have been reported in bacteriophage T7, 8. subtilis, and T4 chromosomes (Dressler et al., 1972; Wake, 1972; and Delius et al., 1971). The existence of multi-forked strands, however, has not previously been documented in higher organisms. Models invoking such structures have been proposed for SV40 and adenovirus excision from cellular chromosomes (Sambrook et al., 1974; Botchan et al., 1978) and to account for the differences in polyteny between euchromatic bands and interbands in Drosophila melanogaster salivary glands (Laird, 1980): Earlier, Laird postulated that either discontinuities exist or multi-forked sites are produced during polyploidization as a result of unequal replication of euchromatic and heterochromatic sequences (Laird, 1973). In agreement with this sugges- tion, we see many free chromatin ends, but whether they are due to random chromosomal breakage during grid preparation or to unstable replication loops (Zannis-Had- jopoulous, 1981) or are, in fact, normal in vivo structures cannot be resolved by the present study. Isolated replica- tion forks that are not contiguous with other forks are also occasionally observed.
The haploid genome of Drosophila melanogaster con-
that the bifurcated strands adjacent to the replication fork are not nucleo somal, but are complexed with fibrous molecules. Whether these lateral extensions represent nascent transcripts of a minor chorion gene or whether they are diverse proteins bound to the DNA in the region containing minor chorion gene sequences cannot be ascertained. Bars = 0.2 Am.

Chorion Gene Amplification and Transcription 551
Figure 8. An Amplrfied Chromatin Strand
A chromatrn strand from an early stage 11 follrcle cell has undergone several bifurcations (arrows), resulting in a complex. multi-forked structure. We have interpreted each bifurcation as a replication fork. An arrowhead pornts to a lone nascent transcript. Regions of the multi-forked structure show homologous strands with varying numbers of particles (nucleosomes) on each. Two strands were selected, and the number of particles per 2 pm stretches were counted. A chromatrn compaction ratio for each strand was calculated based on the following formula:
Compaction ratio
# particles (nucleosomes)/pm x 185 bp/nucleosome zz
2940 bp/pm B-form DNA
Bar= l.Opm (b) The line drawing depicts each replrcatron fork in (a) and the distance to the next repltcation fork rn mrcrometers. The forks do not occur at the same positions along the chromatin strands. Measuring from the first replication fork as far as possible along each strand, we find the longest distance to be 10.86 Grn; the shortest distance IS 8.09 pm.
tains 11 x IO” daltons of DNA, or approximately 50,000 pm (Rasch et al., 1971). Each amplified chorion gene domain extends, at most, 34 pm, which is equivalent to 0.07% of the genome. Coupling this small percentage with a mechanism in which from 8 to 30 replication units open up within each other, we have been fortunate to see multi- forked structures in which the replicons are not completely entangled and collapsed upon each other. This, in turn, has allowed us to visualize ultrastructurally how the fruit fly satisfies the need for huge amounts of protein within a short developmental time period-it amplifies the DNA encoding the chorion genes, and it transcribes these genes prolifically.
Chromatin Spreading Well fed Drosophila melanogaster Oregon-R flies were transferred to a small capped tube and shaken several times to disorient the flies. Flies
1 were not etherized, in order to avoid rnducrng the heat shock response. A female was caught with forceps and her ovaries dissected out over ice in

Cell 552
distilled H20. adjusted to pH 9.0 with NaDH-borate buffer (all subsequent pHs were obtained by adjusting with this buffer). An egg chamber of the desired stage (Mahowald and Kambysellis, 1980; King, 1970) was teased free of the ovariole, the end containing nurse cells and chorionic appen- dages was squeezed off, and the remaining follicle cells were transferred to 100 i of 0.05% Joy, 0.05% Na-desoxycholate. pH 8.2. The cells were pulled apart, macerated, and then stirred with forceps for 2 min. After vigorous pipetting, the chromatin was allowed to disperse for 20-40 min, at which time 25 X of 0.1 M sucrose, 10% formalin, pH 8.5, was added. The solution was layered over a sucrose-formalin cushion in a Lucite centrifuge chamber (Miller and Bakken, 1972) and spun at 1500 x g for 6 min in a Sorvall GLC tabletop centrifuge onto a carbon-coated electron microscope grid, which had previously been rinsed in 95% EtDH for 1 mm to render it hydrophilic. The grid was dipped in a 0.5% Photo-flo solution, pH 8.5, dried, and then stained with 1% phosphotungstic acid in 70% EtOH, followed by 1% uranyl acetate in 70% EDH. After rotary shadow- casting with platinum, grids were observed using a JEOL 1OOC transmission electron microscope. A Durst enlarger was used for printing and tracing. Measurements were made with a Numonics graphic calculator.
Restriction Enzyme Digestion Dissections were done in ice-cold 8 mM NaCI, 0.14 mM MgCI,, pH 8.0. The remaining follicle cells were transferred to 50 X of room temperature 8 mM NaCI, 0.14 mM MgCI,, pH 8.0, containing 10 U of Hind Ill (Bethesda Research Lab). The follicle cells were then macerated and stirred for 1 min with forceps. Over a 10 min period, follicle cells from three additional egg chambers of the same stage, but from different females, were added, with stirring after each. The enzyme was allowed to digest for 20 more min. when 200 X 0.05% Joy, 0.05% Na-desoxycholate, pH 8.2. was added. The mixture was stirred and pipetted, and allowed to disperse for 26 min, at which time 50 X 0.1 M sucrose, 10% formalin. pH 8.5. was added and the dispersed chromatin was processed as above.
Acknowledgments
We thank Dr. Allan Spradling for helpful discussions that contributed significantly to thus research and for his critical reading of the manuscript. We are also grateful to Drs. Charles Emerson, Ann Beyer, Steven McKnight, and Kathy Martin for their comments on the manuscript. Martha Farrell provided essential technical assistance. This work was funded by National Institutes of Health GM 21020 to 0. L. M.
The costs of publication of this article were defrayed in pad by the payment of page charges. This article must therefore be hereby marked “advertisement” In accordance with 18 U.S.C. Section 1734 solely to indicate this fact.
Received March 1, 1983
References
Botchan, M., Topp, W., and Sambrook, J. (1978). Studies on simian virus 40 excision from cellular chromosomes. Cold Spring Harbor Symp. Quant. Biol. 43, 709-719.
Delius, H., Howe, C., and Kozinski, A. W. (1971). Structure of replicating DNA from bacteriophage T4. Proc. Nat. Acad. Sci. USA 68, 3049-3053.
Dressier, D.. Wolfson, J., and Magazin. M. (1972). Initiation and reinitiation of DNA synthesis during replication of bacteriophage T7. Proc. Nat. Acad. Sci. USA 69, 998-1002.
Glover, D. M., and Hogness, D. S. (1977). A novel arrangement of the 18s and 28s sequences in a repeating unit of Drosophila melanogaster rDNA. Cell 10, 167-176.
Griffin-Shea, Ft., Thireos, G., and Kafatos, F. C. (1982). Organization of a cluster of four chorion genes in Drosophila and its relationship to develop- mental expression and amplification. Dev. Biol. 97, 325-336.
Kafatos, F. C. (1972). The cocoonase zymogen cells of silk moths: a model of teninal cell differentiation for specific protein synthesis, Curr Topics Dev. Biol. 7, 125-191.
Kafatos, F. C.. Regier. J., Mazur. G.. Nadel. M.. Blau. H., Petri, W. H., Wyman, A. R.. Gelinas, R., Moore, P., Paul, M., Efstratiadis, A., Vournakis,
J., Goldsmith, M., Hunsley, J., Baker, B.. and Nardi, J. (1977). The eggshell of insects: differentiation-specific proteins and the control of their synthesis and accumulation during development. In Results and Problems in Cell Differentiation, 8, W. Beermann, ed. (Benin: Springer-Verlag), pp. 45-145.
King, R. C. (1970). Ovarian development in Drosophila melanogaster. (New York: Academic Press).
Laird, C. D. (1973). DNA of Drosophila chromosomes. Ann. Rev. Genet. 7, 177-204.
Laird. C. D. (1980). Structural paradox of polytene chromosomes. Cell 22, 869-874.
Long, E. 0.. and Dawid, I. 8. (1979). Expression of ribosomal DNA insertions in Drosophila melanogaster. Cell 18, 1185-l 196.
Long, E. O., and Dawid, I. B. (1980). Repeated genes in eukaryotes. In Ann. Rev. Biochem. 49, 727-764.
Mahowald, A. P., and Kambysellis, M. P. (1980). Oogenesis. In TheGenetIcs and Biology of Drosophila 2d, M. Ashburner and T. R. F. Wright, eds. (New York: Academic Press), pp. 141-225.
Mahowald, A. P., Caulton, J. H., Edwards, M. K., and Floyd, A. P. (1979). Loss of centrioles and polyploidization in follicle cells of Drosophila melan- ogasfer. Exp. Cell Res. 118, 404410.
McKnight, S. L.. and Mifler, 0. L., Jr. (1976). Ultrastructural patterns of RNA synthesis during early embrycgenesis of Drosophila melanogaster. Cell 8, 305-319.
McKnight, S. L., and Miller, 0. L., Jr. (1977). Electron microscopic analysis of chromatin replication in the cellular blastoderm Drosophila melanogaster embryo. Cell 72. 795-804.
McKnight, S. L.. Bustin, M., and Miller, 0. L., Jr. (1978). Electron microscopic analysis of chromosome metabolism in the Drosophila melanogaster em- bryo. Cold Spring Harbor Symp. Quant. Biol. 42, 741-754.
Miller, 0. L., Jr., and Bakken, A. H. (1972). Morphological studies of transcription. Acta Endocrinol. Suppl. 168, 155-177.
Parks, S., and Spradling, A. (1981). The temporal program of chorion gene expression. Carnegie Inst. Wash. Year Book, 80. 188-191,
Pellegrini, M., Manning, J., and Davidson, N. (1977). Sequence arrangement of the rDNA of Drosophila melanogaster. Cell 70. 213-224.
Petri, W. H., Wyman, A. R., and Kafatos, F. C. (1976). Specific protein synthesis in cellular differentiation. Ill. The eggshell proteins of Drosophila melanogaster and their program of synthesis. Dev. Biol. 49, 185-199.
Rasch, E. M., Barr, H. J., and Rasch, R. W. (1971). The DNA content of sperm of Drosophila melanogaster. Chromosoma 33, I-1 8.
Reeder, R. H., Higashinakagawa. T., and Miller, 0. L., Jr. (1976). The 5’.3’ polarity of the Xenopus ribosomal RNA precursor molecule. Cell 8, 449- 454.
Sambrook, J., Botchan, M., Gallimore, P., Ozanne. B., Petterson, U., Williams, J.. and Sharp, P. A. (1974). Viral DNA sequences in cells trans- formed by simian virus 40, adenovirus type 2 and adenovims type 5. Cold Spring Harbor Symp. Quant. Biol. 39, 615-632.
Spradling, A. C. (1981). The organization and amplification of two chro- mosomal domains containing Drosophila chorion genes. Cell 27, 193-201,
Spradling, A. C., and Mahowald, A. P. (1979). Identification and genetic localization of mRNAs from ovarian follicle cells of Drosophila melanogaster. Cell 16, 589-598.
Spradling, A. C., and Mahowald, A. P. (1980). Amplification of genes for chorion proteins during oogenesis in Drosophila melanogasrer. Proc. Nat. Acad. Sci. USA 77, 1096-1100.
Spradling, A. C., and Mahowald. A. P. (1981). A chromosome inversion alters the pattern of specific DNA replication in Drosophila follicle cells. Cell 27,203-209.
Spradling, A. C.. Waring, G. L., and Mahowald, A. P. (1979). Drosophrla bearing the ccelliless mutation underproduce two major chorion proteins both of which map near this gene. Cell 16, 609-616.
Spradling, A. C., Digan, M. E.. Mahowald, A. P., Scott, M., and Craig, E. A. (1980). Two clusters of genes for major chorion proteins of Drosophila melanogaster. Cell 19, 905-914.
Spradling, A. C., Levine, J., and Wakimoto. B. (1981). The mechanism of chorion gene amplification. Carnegie Inst. Wash. Year Book 80, 191-193.

t5$onon Gene Amplificatron and Transcription
Sternemann, M. (1961). Chromosomal replication in Drosophila virilis III. Organization of actrve ongins rn the highly polytene salivary gland cells. Chromosoma (Bert.) 82, 289-307.
Wake, Ft. G. (1972). Visualization of reinitiated chromosomes in Bacillus subtilis. J. Mol. Biol. 68, 501509.
Waring, G. L., and Mahowald, A. P. (1979). Identification and time of syntheses of chorion proteins in Drosophila melanogaster. Cell 76, 599- 607.
Wellauer, P. K., and Dawid, I. 6. (1977). The structural organization of ribosomal DNA in Drosophila melanogaster. Cell 70, 193-212.
Whrte, R. L., and Hogness, D. S. (1977). R loop mapping of the 185 and 28s sequences in the long and short repeating units of Drosophila melan- ogaster rDNA. Cell 70, 177-192.
Zannis-Hadjopoulous, M., Persico, M., and Marttn, R. G. (1961). The remarkable instability of replication loops provides a general method for the isolation of origrns of DNA replication. Cell 27, 155-163.