Embed Size (px)
Citation preview

B R A I N R E S E A R C H 1 3 7 2 ( 2 0 1 1 ) 2 2 – 2 8
ava i l ab l e a t www.sc i enced i r ec t . com
www.e l sev i e r . com/ loca te /b ra i n res
Research Report
Novaeguinoside II inhibits cell proliferation and inducesapoptosis of human brain glioblastoma U87MG cells throughthe mitochondrial pathway
Jie Zhoua,1, Gang Chenga,1, Guang Chenga, Hai-feng Tangb, Xiang Zhanga,⁎aInstitute of Neurosurgery, Xijing Hospital, Fourth Military Medical University, Changlexi Road15#, Xi'an 710032, PR ChinabDepartment of Pharmacy, Xijing Hospital, Fourth Military Medical University, Changlexi Road15#, Xi'an 710032, PR China
A R T I C L E I N F O
⁎ Corresponding author.E-mail address: [email protected] (XAbbreviations: DMSO, dimethyl sulfoxid
5-diphenyltetrazolium bromide; PI, propidiumApaf-1, apoptosis protease activating factor-1 Contributed equally to the work.
0006-8993/$ – see front matter © 2010 Elsevidoi:10.1016/j.brainres.2010.11.093
A B S T R A C T
Article history:Accepted 25 November 2010Available online 11 December 2010
Novaeguinoside II is one of the newly found asterosaponins, which are a kind of steroidalglycosides extracted from starfish with an anti-tumor effect. In this study, we investigatedthe bioactivity of novaeguinoside II on human U87MG glioblastoma cells. The resultsshowed that novaeguinoside II significantly suppresses U87MG cell proliferation in a time-and concentration-dependent manner. Flow cytometric analysis of DNA in U87MG cellsshowed that novaeguinoside II induces the prominent appearance of an S phase peak in thecell cycle that is suggestive of apoptosis that is identical to the result of an annexin V/PIassay. Fluorescence and electronmicroscopy revealed apoptotic change of U87MG cells. Theelectrophoresis of DNA showed a typical “ladder” that is consistent with apoptotic DNAfragmentation. Cytofluorometry showed a decreased mitochondrial transmembranepotential in novaeguinoside II-treated U87MG cells. Western blot showed thatnovaeguinoside II increased the expression of cytochrome-c and caspase-3 protein. Thenovel findings suggest that novaeguinoside II can induce apoptosis of U87MG cells by amitochondrial apoptotic pathway.
© 2010 Elsevier B.V. All rights reserved.
Keywords:Novaeguinoside IIApoptosisU87MG cellMitochondriaCytochrome-cCaspase-3
1. Introduction
Glioma is the most malignant of primary brain tumors andcarries the worst clinical prognosis in both adults and children.Until now, the mainstay of treatment has been surgicaldebulking and radiotherapy, although the overall survival rateremains poor. Proapoptotic drugs can induce apoptosis incancer cells without causing excessive damage to normal cells;
. Zhang).e; DMEM, Dulbecco's m
iodide; ΔΨm, mitochon1; MPTP, membrane perm
er B.V. All rights reserved
so, developing new proapoptotic drugs continue to be themainchemotherapeutic strategy inmedical oncology (Shi et al., 2010).In recent years, marine active compounds have become a newfield in the development of new anti-cancer drugs.
Asterosaponin, thepredominantmetaboliteof starfish that isextracted and isolated from bread starfish (Culcita novaeguineae)is a kind of steroidal glycoside that is extracted from starfishtypically having a structure of Δ9(11) -3β, 6α-dioxyl and a
odified Eagle's medium; MTT, 3-(4,5-dimethylthiazol-2-yl)-2,drial transmembrane potential; IC50, 50% inhibitory concentration;eability transition pore
.

23B R A I N R E S E A R C H 1 3 7 2 ( 2 0 1 1 ) 2 2 – 2 8
sulfate group at C3 and the penta- or hexaglycosides at C62
(Tang et al., 2005a). Tang et al. (2005b) isolated and identified15 asterosaponins from C. novaeguineae. Experiments haveshown that asterosaponins have various biological activities,including anti-tumor activity (Cheng et al., 2006; Tang et al.,2005c, 2006). Novaeguinoside II (Fig. 1) is one of the newlyfound asterosaponin compounds. It has been established as(20R,24S)-6α-O-[3-O-methyl-β-D-quinovopyranosyl-(1→2)-β-D-xylopyranosyl-(1→3)-β-D-glucopyranosyl]-5α-cholest-9(11)-en-3β,24-disulfate disodium salt with a molecular formulaof C45H74O22S2Na2 (molecular weight: 1076) (Tang et al., 2005a).Until now, there has been no systematic study of its bioactivityon tumors. In this study, we have investigated the effect ofnovaeguinoside II on human U87MG glioblastoma cells. Wehave discovered that novaeguinoside II can induce apoptosis ofU87MG cells by mitochondrial apoptotic pathways.
Fig. 2 – Effect of novaeguinoside II on the cell viability of thehuman glioblastoma U87MG cells by MTT assay. *P<0.05;**P<0.01 vs. control group.
2. Results
2.1. Cytotoxicity assay
Exposure of the astrocytes to different concentrations ofnovaeguinoside II (2.32–37.12 μM) for 12–72 h result in thedecline of cell viability with increased concentrations andtreating periods, but the viability remained over 90% (n=10)(Fig. 2A). Novaeguinoside II inhibited the growth of U87MGcells in a concentration- and time-dependentmanner (Fig. 2B).Judging from the growth curve, the concentrations at whichnovaeguinoside II inhibited cell growth by 50% (IC50) at 12, 24,48 and 72 h were 9.20, 4.93, 3.53 and 2.42 μM, respectively.Therefore, novaeguinoside II reduced the viability of U87MGcells but was only slightly toxic to the primary cultured humanastrocytes.
2.2. Cell cycle change in U87MG cells
U87MG cells were treatedwith 2.32 and 4.64 μMnovaeguinosideII for 12, 24 and 48 h. The cellular DNA content was analyzed byflow cytometry to detect changes in the cell cycle distribution(Fig. 3A). The sub-G1 phase (0.43%) appeared in cells that wereincubated with 2.32 μM novaeguinoside II for 12 h, whichincreased to 17.97% in the group incubated with 4.64 μM for48 h. The presence of the sub-G1 peak in cell cycle analysisindicates cell fragmentation, which is generally interpreted asevidence of apoptosis (Table 1) (Schmid et al., 1994). To quantifythe apoptotic effects, the cells were stained with annexin V andPI following novaeguinoside II exposure (Fig. 3B). When
Fig. 1 – The chemical structure of novaeguinoside II.
compared to the control group, there was a statisticallysignificant difference (P<0.05) (Table 2).
2.3. Cellular and nuclear morphology
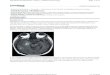
After treatment with novaeguinoside II, the U87MG cellsshrank, aggregated and detached from the surface of theculture flask. For Hoechst 33342 staining, the cells weretreated by 9.28 μM novaeguinoside II for 24 h. Observedunder an epifluorescence microscope, the cells' nuclei under-went alterations that represent apoptosis. The typical changeswere irregularly shaped marginal chromatin condensation.For observation by electron microscope, the cells were treatedby 3.53 μM novaeguinoside II for 24 and 48 h. The electronmicroscope showed marked morphological changes of theU87MG cells, including chromatin condensation, cell shrink-age, nuclear fragmentation and formation of apoptotic bodies.In many cells, there was a developed cytoplasmic blabbingand large vacuoles as a result of cytoplasmic loss. Moreover,the chromatin was condensed and marginalized. Typicalapoptotic bodies appeared in the nuclei. The mitochondrialmorphology changed markedly throughout the apoptosisprocess, including swelling, vacuolization and mitochondrialcristae rupturing (Fig. 4).
2.4. DNA ladder
U87MG cells were treated for 12 and 24 h with 9.28 μMnovaeguinoside II. Isolated DNA was analyzed in agarose gelelectrophoresis. The ladder pattern did not appear in the DNAthat was isolated from the control cells. In the novaeguinoside

Fig. 3 – Cell cycle change in U87MG cells analyzed by flow cytometry and annexin V/PI staining. A. Flow cytometry of U87MGcells after they have been treated by different concentrations of novaeguinoside II for the indicated time periods. B. Apoptosisrate of U87MG cells incubated by novaeguinoside II for different time periods by annexin V/PI staining. A1: U87MG cells culturedfor 12 h in novaeguinoside II-free culturemedia. A2: U87MG cells incubated by 3.53 μMof novaeguinoside II for 12 h. A3: U87MGcells incubated by 3.53 μMof novaeguinoside II for 24 h. A4: U87MG cells incubated by 3.53 μMof novaeguinoside II for 48 h. B1:U87MG cells cultured for 24 h in novaeguinoside II-free culturemedia. B2: U87MG cells incubated by 2.32 μMof novaeguinosideII for 24 h. B3: U87MG cells incubated by 4.64 μM of novaeguinoside II for 24 h. B4: U87MG cells incubated by 9.28 μM ofnovaeguinoside II for 24 h.
24 B R A I N R E S E A R C H 1 3 7 2 ( 2 0 1 1 ) 2 2 – 2 8
II-treated groups, electrophoresis of cellular DNA revealed adistinctive ladder pattern of DNA cleavage in the apoptoticU87MG cells that represent discrete fragments of 180–200 basepairs or multiples (Fig. 5). This showed that novaeguinoside IIcould induceapoptosis by specific DNAcleavage inU87MGcells.
2.5. ΔΨm
The dissipation of ΔΨm is a key event during apoptosis. In thestudy, we found that novaeguinoside II could decrease ΔΨm in
a concentration-dependentmanner. The fluorescent intensityin the novaeguinoside II (3.53, 4.64, 9.28 μM)-treated andcontrol groups were 21501±449, 14810±407, 5105±328 and27753±552, respectively (Fig. 6). There was a statisticallysignificant difference of ΔΨm in different groups (P<0.01).
2.6. Western blot
To further explore themechanismof apoptosis in novaeguino-side II-treated U87MG cells, we undertook Western blot

Table 1 – Cell cycle distribution of novaeguinoside II-treated U87MG cells (%).
Cell cyclestatus (%)
Control 2.32 μM 4.64 μM
12 h 24 h 48 h 12 h 24 h 48 h
Sub-G1 0 0.43±0.03 2.71±0.19 10.15±0.68 9.18±0.23 16.43±1.21 17.97±1.51G0/G1 62.91±3.61 49.88±2.51 49.35±2.65 46.31±3.41 49.27±5.30 42.13±3.55 45.09±2.88S 19.62±2.72 33.13±1.25 33.21±2.03 40.29±3.69 34.52±3.66 30.63±2.58 37.70±4.37G2/M 17.47±1.25 16.99±1.01 17.44±1.48 13.40±1.33 16.20±2.40 27.25±1.98 17.21±2.54
25B R A I N R E S E A R C H 1 3 7 2 ( 2 0 1 1 ) 2 2 – 2 8
analysis to determine whether the expression of cytochrome-cand caspase-3, two important proteins in the mitochondrialapoptotic pathway, changed. The cells were treated by 3.53, 4.64and 9.28 μM novaeguinoside II for 12, 24 and 48 h, respectively.Western blot showed that the expression of cytochrome-c andcaspase-3 increased in a concentration- and time-dependentmanner in the novaeguinoside II-treated group. Furthermore,cleavage of caspase-3 from full-length to the 17 kDa cleavagefragment was present and increased with elongation ofincubation time (Fig. 7). Compared to the control group, all testgroups displayed statistically significant differences (P<0.01).
3. Discussion
There is little knowledge of the bioactivity of newly foundasterosaponins. In this study,we first investigated the bioactivityof novaeguinoside II on U87MG cells. Our investigation showedthat novaeguinoside II could inhibit the growth of U87MG cells ina concentration- and time-dependent manner that was con-firmed by an MTT assay. Next, cell cycle analyses showed thatnovaeguinoside II arrested the cell cycle mainly at the S phase,which meant that DNA synthesis was inhibited by novaeguino-side II. At the same time, the sub-G1 peak increased withincreasing novaeguinoside II concentration and elongation ofincubation time.This suggested that chromosomefragmentationoccurred in the presence of novaeguinoside II. The results provedthat novaeguinoside II could arrest the cell cycle of U87MG cells.
Then, we used Hoechst 33342 stain and an electronmicroscope to observe morphological changes of U87MG cellsand found marked morphological changes and apoptotic bodyformation. This revealed that novaeguinoside II could inducethe apoptosis of U87MG cells. One of the hallmarks of apoptosisis the cleavage of chromosomal DNA into nucleosomal units.Thiswas confirmed by DNA electrophoresis in the study, whichshowed that novaeguinoside II induced apoptosis specific DNAcleavage in U87MG cells.
Table 2 – Apoptosis of novaeguinoside II-treated U87MG cells (%
Group Normal Early apoptosis
control 97.9±5.6 0.9±0.13.53 μM, 12 h 96.6±6.1 1.6±0.1 ⁎
3.53 μM, 24 h 81.8±5.3 ⁎ 5.7±0.3 ⁎
3.53 μM, 48 h 67.1±5.4 ⁎ 20.1±0.8 ⁎
2.32 μM, 24 h 93.4±6.2 3.6±0.2 ⁎
4.64 μM, 24 h 72.8±4.9 ⁎ 17.7±1.2 ⁎
9.28 μM, 24 h 64.8±5.7 ⁎ 24.8±1.8 ⁎
⁎ P<0.05 vs. control group.
Next, we probed the exact mechanism of apoptosis ofU87MG cells induced by novaeguinoside II. Apoptosis istriggered by a variety of stimuli that induce through the cellsurface death receptors, such as Fas, mitochondrial andendoplasmic reticulum stress (Sinha et al., 2005). Cytochrome-c and caspase-3 are two proteins that normally reside in theintermembrane space of mitochondria. In response to a varietyof apoptotic stimuli, they are released to the cytosol and/or tothe nucleus. The release of cytochrome-c will form apoptoticcomplex with Apaf-1 and caspase-9 and activate caspase-3, anexecutioner caspase for apoptosis. In the study, we measuredthe expression of cytochrome-c and caspase-3. The resultsshowed that the expression of the two kinds of proteinincreased in novaeguinoside II-treated groups in a concentra-tion- and time-dependent manner. Furthermore, there wasproteolytic cleavage of caspase-3. This suggested that therelease of cytochrome-c from mitochondria and the activationof caspase-3 involved in novaeguinoside II induced apoptosis.
Normal ΔΨm is essential for various mitochondrial func-tions. Depolarization of ΔΨm is an important indicator in theearly stage of apoptosis (Krysko et al., 2001). In the stimulationof apoptosis-inducing factors, the release of cytochrome-c canpotentially halt the electron transfer, leading to a failure inmaintaining the ΔΨm and ATP synthesis. Energy deprivationwill lead to increased mitochondrial outer membrane per-meabilization. Furthermore, an altered ΔΨ may lead tocytochrome-c release and activation of caspases, or converse-ly, cytochrome-c release may alter mitochondrial function,which—in the absence of caspase activity—may lead to thedeath of the cell (Waterhouse et al., 2001). This indicates thatnovaeguinoside II could decrease ΔΨm in a concentration-dependent manner, perhaps by increasing the cytochrome-crelease. When U87MG cells were treated by different concen-trations of novaeguinoside II in the study, we found decreasedexpression of cytochrome-c. It proved that novaeguinoside IIcould induce apoptosis by increasing cytochrome-c releasefrom mitochondria and depolarization of ΔΨm.
).
Late apoptosis Total apoptosis Necrosis
0.8±0.1 1.7±0.2 0.3±0.01.4±0.1 ⁎ 3.0±0.1 ⁎ 0.4±0.19.7±0.2 ⁎ 15.4±1.2 ⁎ 2.8±0.3 ⁎
10.0±0.2 ⁎ 30.1±2.0 ⁎ 2.8±0.4 ⁎
2.8±0.1 ⁎ 6.4±0.6 ⁎ 0.2±0.07.3±0.2 ⁎ 25.0±2.6 ⁎ 2.2±0.4 ⁎
7.3±0.2 ⁎ 32.1±2.8 ⁎ 3.1±0.6 ⁎

Fig. 4 – Electron micrographs of U87MG cells in different groups. A: normal cells (×20,000); B: cells treated by novaeguinoside II(3.53 μM, 24 h) showed apoptotic body in nuclei (arrowhead) (×20,000); C: cells treated by novaeguinoside II (3.53 μM, 48 h)showed a typical apoptotic body (arrowhead) (×20,000); D: normalmitochondria (×100,000); E: cells treated by novaeguinoside II(3.53 μM, 24 h) showed a swelling mitochondria with large vacuoles in it (arrowhead) (×200,000); F: cells treated bynovaeguinoside II (3.53 μM, 48 h) showed a swelling mitochondria with large vacuoles in it (arrowhead) (×50,000).
26 B R A I N R E S E A R C H 1 3 7 2 ( 2 0 1 1 ) 2 2 – 2 8
In the study, we investigated the mechanism of U87MGapoptosis induced by novaeguinoside II. We found thatnovaeguinoside II could induce apoptosis of U87MG cellsthrough mitochondrial apoptotic pathway by increasing therelease of cytochrome-c and caspase 3. Such an effect is related
Fig. 5 –DNA ladder of U87MG cells treated for 12 and 24 hwith9.28 μM novaeguinoside II. Lane 1: DL2000 marker. Lane 2:normal control. Lane 3: 9.28 μM novaeguinoside II for 12 h.Lane 4: 9.28 μM novaeguinoside II for 24 h.
to theconcentrationand incubationperiodofnovaeguinoside II.However, there is a need for further investigation to explore theexact mechanism of novaeguinoside II on mitochondria andΔΨm. As a newly found compound, there is still much studyrequired to further clarify the bioactivity of novaeguinoside II.
Fig. 6 – Changes of U87MG cells mitochondrialtransmembrane potential (ΔΨm) after incubated withnovaeguinoside II. A: control group, B: novaeguinosideII-treated group (3.53 μM, 24 h), C: novaeguinoside II-treatedgroup (4.64 μM, 24 h), D: novaeguinoside II-treated group(9.28 μM, 24 h).

Fig. 7 – Effect of treatment of novaeguinoside II on caspase-3 and cytochrome-c level in U87MG cells by Western blotting.Control: no novaeguinoside II; Lane 1: 3.53 μM novaeguinoside II for 48 h. Lane 2: 4.64 μM novaeguinoside II for 48 h. Lane 3:9.28 μM novaeguinoside II for 48 h. Lane 4: 9.28 μM novaeguinoside II for 24 h. Lane 5: 9.28 μM novaeguinoside II for 12 h.
27B R A I N R E S E A R C H 1 3 7 2 ( 2 0 1 1 ) 2 2 – 2 8
For example, can it cross the blood–brain barrier? Does it havean anti-tumor effect in vivo? The answers to these and otherquestions require further research.
4. Experimental procedures
4.1. Cell culture and reagents
Novaeguinoside II (>98% pure), provided by Prof. Tang (Depart-ment of Pharmacy, Xijing Hospital, Xi'an, PR China), wasprepared inDMSO and stored at −20 °C. Prior to the experiment,the stock solutionwas thawed and diluted inDMEM (Gibco-BRL,USA) as needed. In no experiment did the final DMSOconcentration exceed 1% (vol./vol.). The human glioblastomaU87MG cells (originally obtained from American Tissue CultureCollection and stored in our laboratory) that were used in thisstudy were grown as a monolayer culture in DMEM supple-mented with 10% fetal bovine serum (Gibco-BRL, USA) andantibiotics in a humid atmosphere containing 5% CO2. The cellswere treated by novaeguinoside II (2.32–37.12 μM) for differentincubation periods (12, 24, 48 and 72 h) before further experi-ments were carried out.
Cultured primary astrocytes were obtained from a slightlyimpaired brain tissue fragment in a patient with cerebraltrauma who consented to the procedure. The gray matter wasdissected after being removed and dispersed repeatedly afterwashing in phosphate buffered sodium (PBS). The cell suspen-sion was filtered and cultured in DMEMwith 10% newborn calf
serum for 2 weeks. The remaining cells weremostly astrocytes.Acquisition of the tissue was approved by the local medicalresearch ethics committee at Xijing Hospital, Fourth MilitaryMedical University. Twenty-four hours before the experiments,thecellswere transferred to a serum-freemedium.Novaeguino-side II in different concentrations (2.32–37.12 μM) was added tothe culture medium and left for 12–72 h to determine if theformer has any effect on the viability of normal astrocytes.
4.2. Cytotoxicity assay
The MTT (Sigma-Aldrich, USA) assay was used to determinethe number of viable cells as described previously (Carmichaelet al., 1988). Briefly, the cells were cultured in 96-well plates ata density of 2×104/ml in the presence of novaeguinoside II atthe indicated concentrations. After incubation for 12, 24, 48 or72 h, the MTT dissolved in PBS was added to each well at afinal concentration of 5 mg/ml and then incubated at 37 °C for4 h. The reduction in viability of the novaeguinoside II-treatedU87MG cells was expressed as a percentage of the novaeguino-side II-free control cells.
4.3. Cell cycle analysis
U87MG cells were trypsinized, counted, centrifuged and fixed inethanol at various times after incubation with novaeguinosideII. Then, the cells were washed twice in PBS and centrifuged. Thepelletwas resuspended ina solutionof RNAse andPI (0.02mg/ml,

28 B R A I N R E S E A R C H 1 3 7 2 ( 2 0 1 1 ) 2 2 – 2 8
Sigma-Aldrich, USA) and incubated at 4 °C for 30 min. Thefluorescence of approximately 10,000–20,000 stained cells wasmeasured. The data were collected and analyzed. The resultswere expressed as a plot of fluorescence intensity vs. cell number(Vindelov and Christensen, 1990).
4.4. Cellular and nuclear morphology
After cells were grown in 24-well plates in the presence ofnovaeguinoside II, the cellular and nuclear morphology wereobserved by fluorescence and electron microscope. To observethe fluorescence, the cells were washed in PBS and fixed in 70%ethanol for 2 h at 4 °C. The cell nucleiwere stainedwith 5 μg /mlHoechst 33342 (Sigma-Aldrich, USA). After a final washing inPBS, the changes in nuclear morphology were observed byfluorescence microscopy (Leica Microsystems, German). Forelectron microscope observation, the cells were trypsinizedwith 0.25% trypsin and centrifuged at 1400g for 15 min. Thepelletswere fixed in0.1 MPBS (pH7.4)with 2.5%glutaraldehyde,postfixed in 2% buffered osmium tetroxide for 2 h and finallydehydrated by graded ethanol solutions. After the specimenswere embedded in EPON resin, thin sections were cut on anultramicrotome and double stained with uranyl acetate andlead citrate. Electronmicrographswere taken ona transmissionelectron microscope (JEM-2000EX, JEOL Ltd., Japan).
4.5. Analysis of DNA fragmentation
For DNA fragmentation analysis, the U87MG cells were treatedfor 12 and 24 h with 9.28 μM of novaeguinoside II. The DNAfragmentationwasdeterminedby theDNA-laddering technique.Briefly, cells were lysed in a buffer containing 5mM Tris–HCl(pH 7.5), 20 mM EDTA and 0.2% Triton-X 100 at 4 °C for 30 min.The lysates were centrifuged at 13,000g for 15 min at 4 °C.FragmentedDNAwas extracted from the supernatant inphenol/chloroform and in chloroform/isoamylalcohol (24:1) and subse-quently precipitated in 100%ethanol. Thepelletswerewashed in70% ethanol and resuspended in 25 μl of Tris/EDTA buffer(pH 7.5). Ten microliters of each preparation were subjected to2.0% agarose gel electrophoresis and stained with ethidiumbromide.
4.6. ΔΨm
The cells were harvested and treated with the final concentra-tion of 5 μg/ml Rhodamine123 (Sigma-Aldrich, USA) at 37 °C for30min. After the loading period, the cells were rinsed with PBSand resuspendedat roomtemperature. TheΔΨmwasmeasuredby fluorescence emission in flow cytometry (Beckman Coulter,USA).
4.7. Western blot analysis
The total protein was harvested from the U87MG cells andquantified by the Lowry assay. For each sample, 50 μg ofprotein were loaded onto 12.5% SDS–polyacrylamide gel,electrophoresed and transferred to a nitrocellulosemembrane(0.22 μm). The membrane was blocked for 1 h at roomtemperature with a blocking buffer. The primary antibodieswere used at a concentration of 1:400 for cytochrome-c (mouse
monoclonal, Neomarkers, USA), 1:600 for caspase-3 (rabbitpolyclonal, Neomarkers, USA) or 1:400 for β-actin (mousemonoclonal, Neomarkers, USA). Then, the membranes wereincubated for 1 hwith HRP-labeled secondary antibodies (anti-rabbit IgG and anti-mouse IgG, both from Cell SignalingTechnology) and then developed by an ECL system accordingto the manufacturer's instructions (Alington Heights, USA).
4.8. Statistical analysis
Statistical analysis was carried out using one-way ANOVAfollowed by Fisher's post hoc test for multiple comparisonsusing SPSS13.0 software. Results with a P value of less than orequal to 0.05 were considered to be statistically significant.Data were expressed as mean±SD (n≥3, where n representsthe number of independent experiments).
Acknowledgment
This work was supported by The Natural Science Foundationof China (No. 30873402).
R E F E R E N C E S
Carmichael, J., Mitchell, J.B., DeGraff, W.G., et al., 1988.Chemosensitivity testing of human lung cancer cell lines usingthe MTT assay. Br. J. Cancer 57, 540–547.
Cheng, G., Zhang, X., Tang, H.F., et al., 2006. Asterosaponin 1, acytostatic compound from the starfish Culcita novaeguineae,functions by inducing apoptosis in human glioblastomaU87MG cells. J. Neurooncol. 79, 235–241.
Krysko, D.V., Roels, F., Leybaert, L., et al., 2001. Mitochondrialtransmembrane potential changes support the concept ofmitochondrial heterogeneity during apoptosis. J. Histochem.Cytochem. 49, 1277–1284.
Schmid, I., Uittenbogaart, C.H., Giorgi, J.V., 1994. Sensitive methodfor measuring apoptosis and cell surface phenotype in humanthymocytes by flow cytometry. Cytometry 15, 12–20.
Shi, L., Chen, J., Yang, J., et al., 2010. MiR-21 protected humanglioblastoma U87MG cells from chemotherapeutic drugtemozolomide induced apoptosis by decreasing Bax/Bcl-2 ratioand caspase-3 activity. Brain Res. 1352C, 255–264.
Sinha, I., Sinha-Hikim, A.P., Hannawa, K.K., et al., 2005.Mitochondrial dependent apoptosis in experimental rodentabdominal aortic aneurysms. Surgery 138, 806–811.
Tang, H.F., Yi, Y.H., Li, L., et al., 2005a. Bioactive asterosaponinsfrom the starfish Culcita novaeguineae. J. Nat. Prod. 68, 337–341.
Tang, H.F., Yi, Y.H., Li, L., et al., 2005b. Three new asterosaponinsfrom the starfish Culcita novaeguineae and their bioactivity.Planta Med. 71, 458–463.
Tang, H.F., Yi, Y.H., Li, L., et al., 2006. Asterosaponins from thestarfish Culcita novaeguineae and their bioactivities. Fitoterapia77, 28–34.
Tang, H.F., Yi, Y.H., Li, L., et al., 2005c. A new asterosaponin fromthe starfish Culcita novaeguineae. Chin. Chem. Lett. 16, 619–622.
Vindelov, L., Christensen, I.J., 1990. An integrated set of methodsfor routine flow cytometric DNA analysis. Methods Cell Biol. 33,127–137.
Waterhouse, N.J., Goldstein, J.C., von Ahsen, O., et al., 2001.Cytochrome c maintains mitochondrial transmembranepotential and ATP generation after outer mitochondrialmembrane permeabilization during the apoptotic process. J.Cell Biol. 153, 319–328.