Embed Size (px)
Citation preview

Nonlinear Dynamics & Vision
Hugh R. WilsonBiology & Centre for Vision Research
York University

Outline
• Whirlwind tour of nonlinear dynamics• Overview of higher form vision areas• Marroquin illusion• Detailed analysis of competitive
networks in rivalry

Nonlinear Dynamics: Equilibria & Linearization
• Coupled, first order equations• Solve for steady states or
equilibrium points• Compute Jacobian matrix• Evaluate at each steady state• Determine eigenvalues

Linearized Stability Analysis
€
dxdt
= F(x,y)
dydt
=G(x,y)
€
J =
∂F∂x
∂F∂y
∂G∂x
∂G∂y
x,y=Equilibrium
• All real(eig) < 0, asymptotic stability• Any real(eig) > 0, unstable• Pure imaginary eig: theorem does not
apply

Nonlinear Oscillations• Limit cycles: cannot exist in linear systems• Only one general theorem: Hopf Bifurcation
Theorem• Conservative oscillations (analogous to
linear systems) can exist, but not in neural systems
• Chaos can occur in > 2 dimensions• All neural oscillations are limit cycles!

Hopf Bifurcation Theorem
• Equilibrium is asymptotically stable for b < a
• Pair of pure imaginary eig for b = a• For all other eig, real(eig) < 0• Equilibrium point unstable for b > a• Asymptotically stable limit cycle for
b > a; or unstable for b < a
€
dr X
dt=
r F
r X ,β( )

Hopf Addendum
• Limit cycle emerges with infinitesimal amplitude
• Frequency = Im(eig)/2pi• Hodgkin Huxley Euations exhibit
Hopf bifurcation
€
dr X
dt=
r F
r X ,β( )

Nonlinear Oscillations Caveats• Not all limit cycles emerge via Hopf
bifurcations• Example: Mammalian cortical neurons• Conservative oscillations (analogous to
linear systems) can exist, but not in neural systems
• Chaos can occur in > 2 dimensions• All neural oscillations are limit cycles!

Conduction & Spiking Dynamics
• Two-compartment model (Rinzel et al)• Excitatory neurons: slow AHP currents• Simple but accurate cubic model (Wilson, 1999)
dVdt = – a V2 – b V V – ENa – R V – EK + Iinput
τ Rd Rdt = – R + cV2
(conductance)x(potential)

Phase Plane & Spike Generation
-0.2 0 0.2 0.4 0.6 0.8-0.2
0
0.2
0.4
0.6
0.8
1
V
R
-0.2 0 0.2 0.4 0.6 0.8-0.2
0
0.2
0.4
0.6
0.8
1
V
R
0 10 20 30 40 50 60 70 80 90 100-0.1
0
0.1
0.2
0.3
0.4
0.5
0.6
0.7
0.8
0.9
V (N
orm
alize
d Un
its)
Time (arbitrary units)

Fit to Human Action Potentials
-80
-60
-40
-20
0
20
40
0 1 2 3
HumanEqn (9.10)
Pote
ntia
l (m
V)
Time (msec)
A
-80
-60
-40
-20
0
20
40
0 1 2 3
HumanEqn (9.10)
Pote
ntia
l (m
V)
Time (msec)
A
Foehring et al, 1991

Spike Rate Adaptation• Human excitatory cortical neurons:
slow hyperpolarizing current• Causes spike rate adaptation
dVdt = – a V2 – b V V – E Na – R V – EK – H V – EK +I
τ Rd Rdt = – R + cV2
τ Hd Hdt = – H + gV2 Very slow K+ current
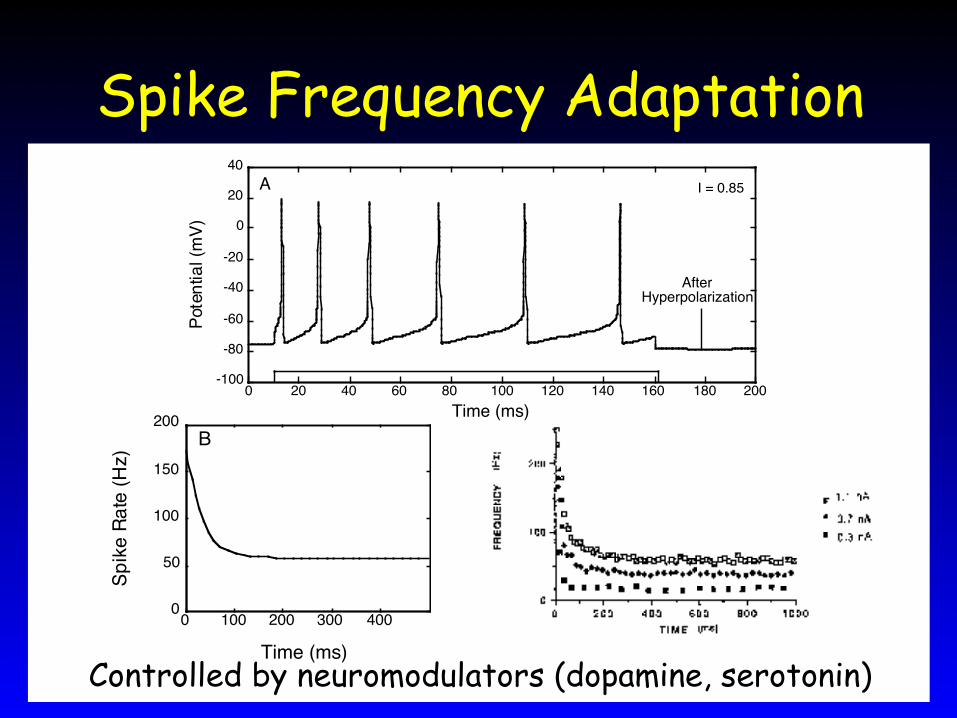
Spike Frequency Adaptation
0 100 200 300 4000
50
100
150
200
Time (ms)
B
Spik
e Ra
te (H
z)
0 20 40 60 80 100 120 140 160 180 200-100
-80
-60
-40
-20
0
20
40I = 0.85
Time (ms)
Pote
ntia
l (m
V)
AfterHyperpolarization
A
Controlled by neuromodulators (dopamine, serotonin)

Lyapunov Functions & Memory
• Positive definite function U(t) around an equilibrium
• dU/dt < 0 along trajectories in a region surrounding equilibrium
• Then equilibrium is asymptotically stable• Lyapunov fcns. always exist, but not unique
€
dUdt
=∂U∂xii
∑ dxidt

Lyapunov Functions & Memory• Apply where linearization fails• Permit estimate of domain of attraction
€
dxdt
= −y − x 3
dydt
= x − y 3
€
U(x,y) =x 2 + y 2
2dUdt
= −x 4 − y 4
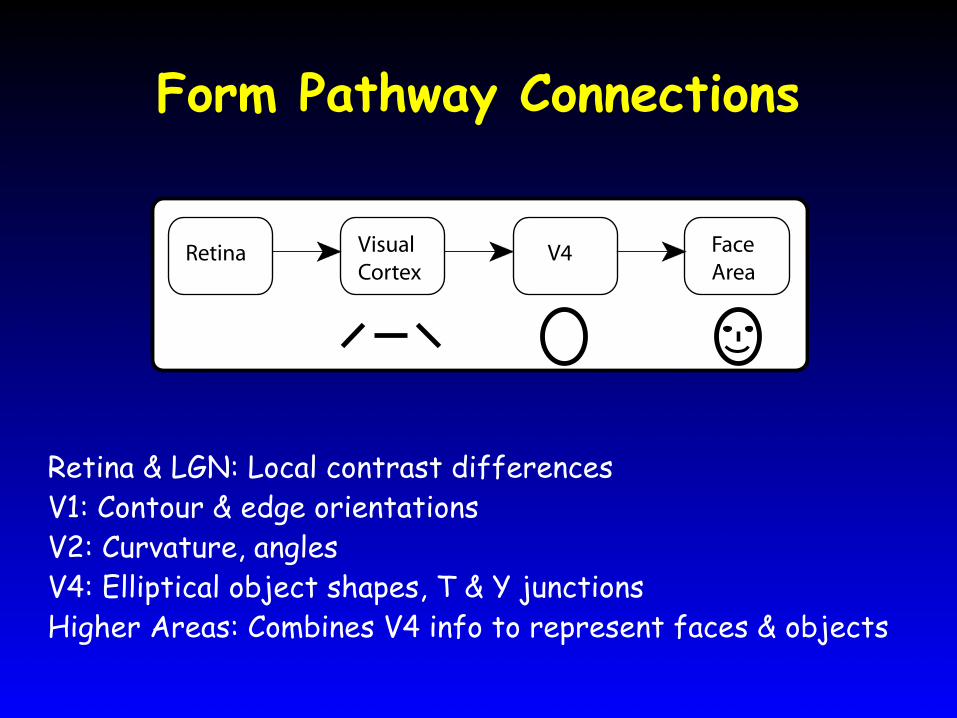
Form Pathway Connections
Retina & LGN: Local contrast differencesV1: Contour & edge orientationsV2: Curvature, anglesV4: Elliptical object shapes, T & Y junctionsHigher Areas: Combines V4 info to represent faces & objects
Retina VisualCortex
FaceArea
V4

Form Pathway Connections
Area to area feedback connectionsSkipping connectionsFeedback local but patchy
V1 V4 TEO TEV2

fMRI of V4
Wilkinson et al, Current Biology (2000)

V4 & FFA Activation
0
0.2
0.4
0.6
0.8
1
1.2
1.4
V1 V4 FFA
ConcentricRadialParallelFaces
Resp
onse
(% s
igna
l cha
nge)
Cortical Area
0
0.2
0.4
0.6
0.8
1
1.2
1.4
V1 V4 FFA
ConcentricRadialParallelFaces
Resp
onse
(% s
igna
l cha
nge)
Cortical Area
0
0.2
0.4
0.6
0.8
1
1.2
1.4
V1 V4 FFA
ConcentricRadialParallelFaces
Resp
onse
(% s
igna
l cha
nge)
Cortical Area
0
0.2
0.4
0.6
0.8
1
1.2
1.4
V1 V4 FFA
ConcentricRadialParallelFaces
Resp
onse
(% s
igna
l cha
nge)
Cortical Area
* *
**

Gallant et al, 2000: V4 damage
Normal V4Damaged V4
But: Orientation Discrimination normal (normal V1)

Receptive Field Size Increases
0.1°
1°
10°
100°Receptive Field Sizes
RF DiameterKobatake & TanakaOp de Beeck et al
RF D
iam
eter
Cortical Area
Diameter = 0.09°·Area3.04
V1 V2 V4 TEO TE
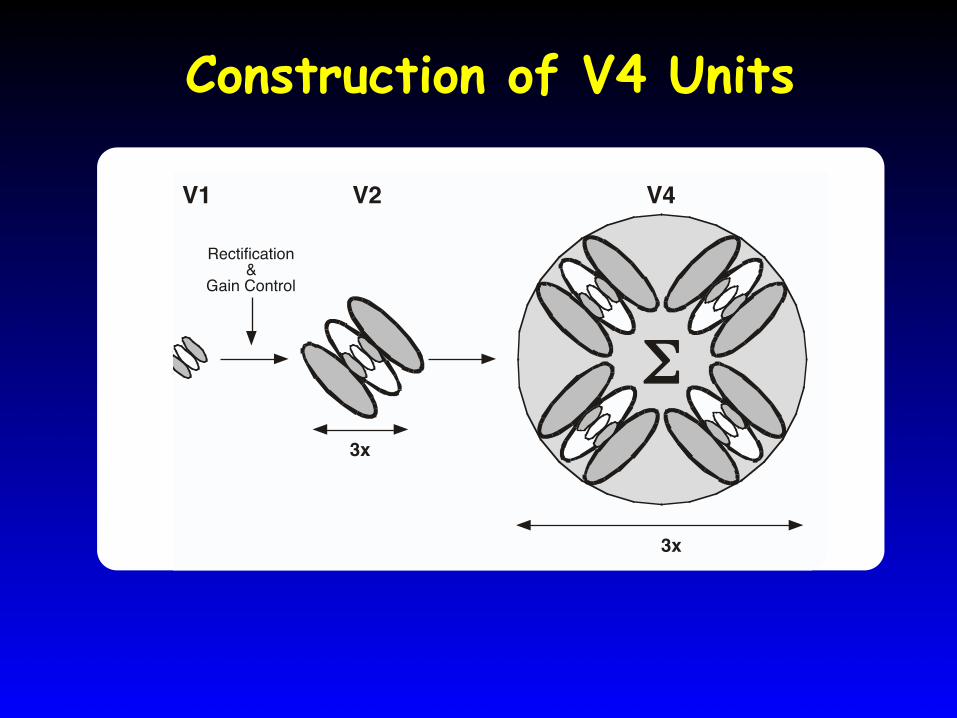
Construction of V4 Units
3x
3x
Σ
Rectification&
Gain Control
V1 V2 V4
3x
3x
Σ
Rectification&
Gain Control


fMRI & Distance from MeanMean Face
1
1.2
1.4
1.6
2 4 6 8 10 12 14 16Face geometry (%)
FFA neurons increase firing with distance from mean faceNature Neuroscience, October, 2005

Marroquin Illusion (1976)

Competitive Marroquin Model
τEdE n
d t= – E n +
100P +2
10+H n2+P +
2
where P = S Marroquin – 0.6 I k exp – R nk5 σ5– R nk5 σ5Σ
n ≠ k
τId I n
d t= – I n + E n
τHdH n
d t= – H n +gE n
Slow adapt (400-900 ms) Ca++ - K+ potential McCormick, Avoli
Spatial Competitive Inhibition
Describe neurons by spike rate equations
Sigmoid Nonlinearity

Competitive Model (demo)
• Spatially Regional Winner Take All• Winner slowly adapts, so new winners
emerge• Model generates gamma distribution

Gamma Distribution
0
5
10
15
20
0 2 4 6 8 10
FWGamma
Inte
rval
Cou
nt
Interval Duration (sec)
N = 103
0
10
20
30
40
50
60
70
0 2 4 6 8 10
ModelGamma
Interval Duration (sec)
N = 478
Too many long intervals for true Gamma!





























