Embed Size (px)
Citation preview

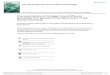
The Snake River basin above Hells CanyonDam comprises approximately 58% of the landsurface area of Idaho (Fig. 1). Since settlementin the 1800s by humans of European descent,aquatic resources in the Snake River basin, as
in the rest of western North America, haveundergone extensive alterations due to hydro -power, agriculture, grazing, mining, logging,and other extraction-based land uses. Despitethese impacts, the only fish species extirpated
Western North American Naturalist 73(1), © 2013, pp. 20–34
NONGAME FISH SPECIES DISTRIBUTION AND HABITAT ASSOCIATIONSIN THE SNAKE RIVER BASIN OF SOUTHERN IDAHO
Kevin A. Meyer1,3, James A. Lamansky Jr.1, Daniel J. Schill1, and Donald W. Zaroban2
ABSTRACT.—The distribution and abundance of game fish populations are commonly monitored closely, whereassampling of nongame species is often neglected. We used a broad-scale salmonid sampling project both to simultane-ously assess the distribution and relative abundance of nongame fish species in small streams (i.e., ≤15 m wetted width)in the Snake River basin of southern Idaho and to relate the distribution and abundance of nongame species to abioticand biotic stream conditions in the study area. Of the 1738 reaches surveyed, 34% were dry or contained too little waterto support fish, and an additional 21% had flowing water but were still absent of fish. At least one species of nongamefish was captured at 30% of the reaches surveyed, and all 18 native nongame fish species believed present in study areastreams were captured. The most widely distributed species was speckled dace Rhinichthys osculus (present in 21% ofsurveyed reaches, excluding dry and nearly dry reaches), followed by bridgelip sucker Catostomus columbianus (19%),Paiute sculpin Cottus beldingi (15%), and redside shiner Richardsonius balteatus (13%). The species least often present(≤1%) were leopard dace Rhinichthys falcatus and peamouth Mylocheilus caurinus. Common carp Cyprinus carpio (cap-tured at 3 locations) and oriental weatherfish Misgurnus anguillicaudatus (2 locations) were the only nonnative nongamespecies encountered. Catostomids and cyprinids generally formed a composite fish assemblage that was associated withhigher stream order (i.e., farther from headwaters), lower elevation, and lower-gradient reaches where streams werewider and deeper. Stream order was the strongest loading factor for canonical correlations constructed for study reachesboth above and below Shoshone Falls, suggesting that the increased abundance of catostomids and cyprinids generallyoccurred on a longitudinal downstream gradient. Biologists sampling game fish populations in streams should alsorecord data on nongame species to more closely monitor their status through time.
RESUMEN.—La distribución y abundancia de las poblaciones de peces de valor deportivo (game fish) comúnmente semonitorean cercanamente, mientras que el muestreo de especies de peces que no son de valor por lo general suele des-cuidarse. Utilizamos un proyecto de muestreos de salmónidos a gran escala para evaluar de manera simultánea la distri-bución y la abundancia relativa de especies de peces que no son de valor en arroyos pequeños (i.e., ≤15 m de ancho), enla cuenca del Río Snake en el sur de Idaho y para relacionar su distribución y abundancia con las condiciones bióticas yabióticas del arroyo en el área de estudio. De los 1738 tramos revisados, el 34% estaban secos o contenían muy pocaagua para mantener a los peces, y un 21% adicional tuvo caudal de agua pero tampoco se encontraron peces. Se capturóal menos una especie de pez que no es de valor en el 30% de los tramos revisados, y se capturaron las 18 especies nati-vas de peces que no son de valor y que se estimaban presentes en los arroyos del área de estudio. Las especies másampliamente distribuidas fueron Rhinichthys osculus (presente en 21% de los tramos revisados, sin incluir los tramossecos y casi secos), seguido por Catostomus columbianus (19%), Cottus beldingi (15%) y Richardsonius balteatus (13%),mientras que las especies menos presentes (≤1%) fueron Rhinichthys falcatus y Mylocheilus caurinus. La carpa comúnCyprinus carpio (capturada en tres sitios) y Misgurnus anguillicaudatus (dos sitios) fueron las únicas especies encon-tradas que son no nativas y que no poseen valor. Los catostómidos y ciprínidos generalmente formaron un ensamblecompuesto de peces que estuvo asociado con arroyos de orden superior (es decir, más lejos de las partes altas del río),menor elevación y tramos de gradiente más bajo donde los arroyos fueron más amplios y profundos. El orden del arroyofue el factor con carga más alta en las correlaciones canónicas construidas para los tramos revisados tanto por encima ypor debajo de las Cataratas Shoshone, lo que sugiere que la creciente abundancia de catostómidos y ciprínidos general-mente ocurrió en un gradiente longitudinal aguas abajo. Los biólogos que toman muestras de las poblaciones de pecesde valor deportivo en arroyos también deben registrar datos de las especies que no poseen valor, para supervisar suestado con mayor detalle a través del tiempo.
1Idaho Department of Fish and Game, 1414 East Locust Lane, Nampa, ID 83686.2Idaho Department of Environmental Quality, 1410 N. Hilton Street, Boise, ID 83706.3E-mail: [email protected]
20

from the Snake River basin in southern Idahoare anadromous species (salmon and lamprey)whose passage has been blocked by dams lack -ing fish ladders. Due to their sportfishing andaesthetic value, native game fish populationsin the basin have been closely monitored (e.g.,Meyer et al. 2006, 2009, High et al. 2008, Schill2009), in part so that management practicessuch as harvest regulations can be adjusted if
fish population metrics indicate changes areneeded. In general, the economic or intrinsicvalues of native nongame fish species arerarely altered by the recreational community,thus management regulations for native non -game species have rarely been necessary, andpopulation monitoring has therefore rarelybeen conducted. This lack of information is animpediment to understanding the conservation
2013] NONGAME FISH STATUS IN SOUTHERN IDAHO 21
##
##
#
#
#
#
#
#
#
##
#
#
#
#
# #
#
#
##
##
##
#
!(
!(!(
!(
!(
!( !(!(
!(!(!(!(!(
!( ##
#
#
##
#
#
#
#
!(!(!(
!(
!(
!( !(!(
!(
!(
!(!(!(
!(!(
!(
!(
#
#
#
#
##
#
###
#!(
!(!(!(
!( !(!(
!(!(!(
!(!(!(
!(
!(!(!(!(!(!(!( !(
!(
!(!(
!(!(!(
!(!(!(!(
!(!(!(!(!(
!(
!(
!(
!(
!(
!(
!(
!(!(
#
!(!(
!(
!(
!(!(!(!(!(!(!(!(
!( !(!(
!(!(
!(
!(
!(!(
!(
!(
!(
!(!(
!(!(
!(!(!(!(!(
!(
!(
!(!(!(
!(
!(!(
!(
!(
!(
!(
!(!(
#
##
#
#
#
##
#
#####
####
#
#
#
##
##
#
###
#
##
# #
!(
!(
!(!(!(
!(
!(!(!(
!(
!(!(
!(
!(!(
!(
!(
!(
!(
!(
#
##
#!(
!(!(!(
!(
!(
#
!(
!(!(
!(
!(!(!(
!(
!(
!(
!(!(!(
!(
!(
!(
!(
!(!(!(!(
!(
!(!(
!(!(
##!(!(
##!(
#########
##
####
!(!(!(!(!(!(!(!(!(!(
!(
!(!(!(!(#
#
#
!(!(!(!(
!(!(
!(!(
!(!(
!(!(
!(
!(
!(!(
!(
####
#
##
#
###
##
#
#
###
#
##
#
#
#
#
#
#
#
#
#
#
##
#
####
###
####
#
#
##
##
###
##
###
#
# ###
# ###
###
##
#
##
#
#
####
#
#
#
#
##
#
###
#
#
#
# ###
#
#
###
# #
#
### ##
###
#
#
#
##
!(!(!(
!(!(
!(
!(
!(
!(!(
!(!(
!(
!(
!(
!(
!(
!(
!(!(
!(!(
!(!(
!(
!(
!(
!(
!(
##
#
#
#
#
#
##
#
#
#
!(
!(
!(
!(!(
!(
!(
!(
!(!(
!(
!(
!(!(
!(
!(
!(
!(!(!(!( !(
!(
!(
!(!(!(
!(
!(
!(
!(!(
!(!(
!(
!(
!(
!(
!(
!(!(
!(
!(
!(
!(
!(
!(#
#####
#
##
##!(
!(
!(!(!(
!(!(
!(
!(
!(
!(!(!(
!(!( !(
!(
!(
!(
!(
!(
!(
!(
!(!(!(
!(
!(!(!(!(!(!(!(!(
!(!(
!(!(!(
!(
!(
!(
!(
!( !(
!(!(
!(!(!(!(
!(!(!(
!(
!(
!(
!(
!(!(
!(!(
!(
!(!(
!(
!(!(
!(
!(
!(!(
!(
!(
!(
!(!(
!(
!(
#
#
!(
!(
!(
!(
!(
!(
!(
!(
!(
!( !(
!(
!(!(
!(
!(
!(
!(
!(
#
##
##
#
#
#!( !(!(
!(
#
##!(!(
!(
!(
##
#
#
#### #
#
#
##
###
##
####
##
#
#
#
#
##!( !(!(
!(
!(
!(
!(
!(
#
#
!(
!(
!(!(
!(
!(
!(
!(
!(!(!(
!(!(
!(
!(
!( !(
!(
!(!(
!(!(
!(
!(!(!(
!(
!(
#
## # #####
#
###
###
#####
###
##
###
###
#####
#####
###
#
#
##
##
#
#
#
##
!(
!(
!(!(!(
!(
!(
!(!(!(
!(
!(!(
!(
!(!(!(!(!(
!(!(
!(
!(
!(!(!(
!(!(!(!(
!(!(
!(
!(!( !(
!(!(
!(
!(!(!(
#
##
##
##
!(
!(!(
!(!(!(!(
!(!(
!(
!(
!(
!(
!(
!(!(
!(!(
!(!(
!(!(
!(
!(!(!(!(
!(
!(!(
!(
!(!(
!(!(
!(!(
!(
!( !(!(
!(
!(
!(
!(
!(
!(
!(
!(
!(
##
#
#
#
###
#
!(!(
!(
!(
!(
!(
!(
!(
!(!(!(!(!(!(!(
!(!(!(!(!(
!(!(!(
!(
!(
!(
!(!(!(!(
##
# !(!(!(
!(
!(
!(
!(
!(
!(!(
!(
!(
!(!(
!(
!(
!(
##!(
!(!(!(
#
#######
##
#!(!(
!(!(!(!( !(!(!(
!(
#
#####
#
##
#
!(!(
!(
!(!(
!(!(
!(
!(!(
!(
!(
!(!(!(
!(
!(
!(
!(!(
!(
!(
!(!(
!(!(
!(!(
!(
#
#
##
!(
!(
!(
!(
!(
!(!(
!(!(!(!(!(!(
!(!(!(!(!(!(!(
!(!(
!(
!(!(
!(!(!(!(!(
!(
!(!(
##
#
####
!(
!(!(!(
!(!(
!(!(!(
!(!(
!(
!(
!(
!(####
#
!(
!(
!(!(
##
#
#
#
## #
###
#
#
!(!(!( !(
!(
!( !(
!(!(
!(!( !(!(
!(
!(!(!(!(
!(!(
!(!(
!(!(!(
!(!(!(!(!( !(!(
!(
!(!(!(
!(!(
!(!(
!(!(#
#
#
!(!(!(
!(!(!(!(
!(
!(
!(
!(
!( !(!(!(
!(
!(
!(!(
!(!(!(
!(
!( !(!(
!(!(!(
##!(!(
!(!(!(!(!(!(
#
##
#
#
!(!(
!(!(
!( !(
!(!(
!(!(!(
!(!(
!(
!(
!(!(!(
!(
!(
!(
!(
!(!(!(!(!(!(
!(
!(!(!(!(
!(!(!(!(
!(!(
!(!(
!(
!(##
##
#
!(!(
!(
!(
!(
!(!(!(!(!(!( !(!(!(!(!(
!(!(!(
!(
!(
!(
!(
!(
!(!(
!(
!(
!(
!(
##
##
# ###
######
# ####
#
#!(
!(
!(!(
!(!(
!( !(
!(!(
!(
!(!(!(
!(
!(!(!(
!(
!(!(
!(
!(
!(!(!(!(
!(!(!(!(!(!(!(!(!(
!(
#
##
###
#
#!(
!(
!(
!(
!(
!(!(
!(
!(!(!(!(
!(!(
!(
!(!(!(
!(
!(
!(!(!(!(!(
#########
####
!(
#!(
#
#
#
##
#!(
!(
!(
!(
!(!(!(
!(
!(
!(!(
!(!(!(!(!(
!(!(!(!(
!(
!(!( !(
!(
!(
!(!(!(!(!(
!(
!(
!(
!(
!(!(
!(
!(
!(
!(!(!(
!(
!(!(
!(!(
!(
!(
!(
!(
!(!(
!(
!(
!(!(
!(#####
###
####
#####
####
!(
!(
!(!(
!(
!(!(
!(
!(!(!(!(!(!(!(!(
!(
!(!(
!(!(!(
!(!(
!(!( !(!(!(!(#####
#
!(!(!(!(
!(!(
!(!(!(
!(!(!(!(
!(!(!(
!(
!(!(
!(!(
!(
!(!(###
##
# ######
!(!(!(!( !(!(!(
!(!(!(!( !(!(!(!(
!(!(!(!(!( !(
!(!(!(!(!(!(!(
!(!(
!(!(!(!(
!(!(!( !(!(!(
!(!(!(!(
####
#
###!(!( !(!(
!(!(
!(!(!(
!(!( !(
####
#
##
#
##
##
### ###
##
!(!(
!(!(!(!(!(!(!(!(!(!(!(!(
!(
!(
!(!(
!(
!(!(
!(!(!(
!(
!(!(!(
!(
!(!(
!(!( !(
!(!(!(!( !(
!(
!(!(!(
!(
!(!(
!(
!(
!(
#
#
# # !(
!(!(!(!(!(
!(!(
!(!(!(
!(
!(!(!(!(
!(
######
#
####
##
!(
!(!(!(!(
!(!(!(!(
!(
##
#
###
###
#!(!(
!(!(!( !(!(!(
!(!(
!(!(!(!(!(!( !(!( !(!(!(
!(
!(!(!(
%2
#0
Fort Hall
Duck Valley
Thousand Springs
²0 10050
Kilometers
Streamflow & Non-Game Presence# Dry!( Non-game not present!( Non-game present
Barriers
%2 Hells Canyon Dam
#0 Shoshone Falls
MT
AL
OR
NV
BC
UT
WY
WA
Fig. 1. Locations where the distribution and relative abundance of nongame fish species were examined in the SnakeRiver basin of southern Idaho.

status of many native nongame fish species inIdaho (IDFG 2007).
The Snake River basin above Hells CanyonDam historically supported 26 native fish spe -cies (Simpson and Wallace 1982), 20 of whichare considered nongame species. Fish distrib-ution patterns in the Snake River basin havebeen influenced by numerous climatic andgeologic events. Smith (1978) argued that thepredominant factors affecting fish fauna pat-terns in the Intermountain Region of the west-ern United States are the degree of isolationbetween basins and the late Cenozoic historyof fluctuating wet and dry periods, which alsoincluded glaciation. In addition, late Pleisto -cene volcanic activity in the region repeatedlyaltered drainage patterns and may have exter-minated some or all fishes from the upperSnake River basin (Hubbs and Miller 1948,Johnson 2002). Shoshone Falls, a 65-m highwaterfall on the Snake River at river kilometer992 (Fig. 1), precluded the invasion of manyColumbia River fish species (such as redbandtrout Oncorhynchus mykiss gairdneri andnorthern pikeminnow Ptychocheilus oregonen-sis) into the upper drainages of the Snake Riverbasin. Taken collectively, these processes forma multifaceted history of hydrologic isolation,diversion, and reconnection and have resultedin a complex pattern of fish distribution in theSnake River basin.
Overlaying this complex hydrologic historyis the fact that fish distribution patterns arealso affected by combinations of local bioticand abiotic factors, and associated regionalfac tors, that determine how suitable streamreaches are for particular species (Angermeierand Winston 1998, Hughes et al. 2006). Forexample, stream size, elevation, channel gradi-ent, water temperature, and substrate compo-sition have commonly been shown to influ-ence the distribution and relative abundanceof nongame fish species across North America(e.g., Grossman et al. 1998, Quist et al. 2004c,2004a, Rashleigh et al. 2005, Torgersen et al.2006). Understanding fish distribution patternsacross the landscape and mechanisms that in -fluence fish assemblages can contribute im -portant knowledge for management and con-servation purposes. For instance, a commonpurpose of developing relationships betweenfish assemblages and both abiotic and bioticstream conditions is to develop indices of bio-logical integrity to assess environmental quality
in flowing waters (e.g., Mebane et al. 2003) orto facilitate preservation and rehabilitation ofriverine ecosystems (e.g., Scott and Hall 1997,Hughes et al. 1998).
Information regarding species assemblagesand stream habitat conditions across the land-scape is especially lacking in the Intermoun-tain Region of the western United States (Quistet al. 2004c), where fish assemblages are rela-tively depauperate compared to streams ineastern North America (Lee et al. 1980). Es -tablishing relationships between stream-dwell -ing fish and their environment is often prob-lematic because these types of studies areoften focused at small scales (Fausch et al.1988). Although they may adequately describefish distribution or abundance patterns withina particular study area, small-scale studies mayhave limited ability to explain patterns acrossthe landscape if inadequate sample sizes failedto fully characterize fish–habitat associations,or if the limited study area was not represen-tative of other areas of a species’ range.
The primary objective of this paper was tofill the aforementioned data gap regarding thedistribution and abundance of nongame fish(native and nonnative) species in the SnakeRiver basin above Hells Canyon Dam. To ac -complish this objective, we used field crewsdeployed to collect data on stream-dwellingsalmonids across the landscape to simultane-ously gather quantitative information on non -game species occupying lotic environments inthe study area. Because little work has beendone to describe fish–habitat associations fornongame fish species in the Snake River basinin southern Idaho, a second objective was toassess which abiotic and biotic stream condi-tions were associated with the distribution andrelative abundance of these species. Our large,spatially balanced sample size (over 1700 streamsurveys) over a broad geographic scale helpedcircumvent the aforementioned limitations ofstudies of fish–habitat associations.
METHODS
Study Area
The Snake River in Idaho flows approxi-mately 1000 km from east to west across south -ern Idaho, then flows north to the confluencewith the Columbia River. The study area in -cluded most of the Snake River basin (approxi-mately 84,000 km2) above Hells Canyon Dam
22 WESTERN NORTH AMERICAN NATURALIST [Volume 73

(Fig. 1). We excluded the Malheur River, PowderRiver, Burnt River, and Pine Creek drainagesbecause they reside entirely in Oregon, andwe also excluded the Snake River drainage inWyoming (upstream of Palisades Reservoir), ex -cept for those tributaries that were containedat least in part within Idaho. Similarly, Nevada,Utah, and Oregon basins were in cluded if theywere contained at least partly within Idaho.Within Idaho, we did not sample the Lost Riverdrainages (Birch Creek, Little Lost River, BigLost River) or streams located within the bound-aries of the Duck Valley and Fort Hall Indianreservations (Fig. 1). We also excluded the copi-ous springs directly adjacent to the Snake Riverin south central Idaho, known as the ThousandSprings, for 2 reasons. First, these springs rep-resent very unique habitat in the study area, andmany of these springs are actually more likelentic habi tats than streams. Also, most of thesesprings are immediately adjacent or closelyproximal to the Snake River, and species pres-ence is often more strongly influenced by thefish as semblage of the Snake River than thehabitat characteristics of the spring.
Within this study area, native nongame fishspecies include 5 catostomids (bluehead suckerCatostomus discobolus, bridgelip sucker Cato -stomus columbianus, largescale sucker Catosto -mus macrocheilus, mountain sucker Catostomusplatyrhynchus, and Utah sucker Catostomus ar -dens); 9 cyprinids (chiselmouth Acrocheilusalutaceus, leopard dace Rhinichthys falcatus,longnose dace Rhinichthys cataractae, northernleatherside chub Lepidomeda copei, northernpikeminnow, peamouth Mylocheilus caurinus,redside shiner Richardsonius balteatus, speck-led dace Rhinichthys osculus, and Utah chubGila atraria); and 4 cottids (mottled sculpinCottus bairdi, Paiute sculpin Cottus beldingi,shorthead sculpin Cottus confusus, and WoodRiver sculpin Cottus leiopomus). Shoshone scul -pin Cottus greenei is also native to the studyarea but only occurs in the Thousands Springsarea of south central Idaho, not in any particu-lar river drainages. Pacific lamprey Entosphenustridentata were native below Shoshone Fallsbut were extirpated from the study area longago by a series of ladderless dams built on theSnake River and many tributaries.
Fish Sampling
Between 1999 and 2005, we surveyed streamreaches throughout the Snake River basin in
southern Idaho from mid-June to mid-October,after high spring flows had receded. Studyreaches were distributed randomly across a1:100,000-scale stream network in first- throughsixth-order streams (Strahler 1964) based pro-portionally on the length of stream withineach stream-order stratum. However, streamsthat were greater than 15 m wide (mean wet-ted width) or 1 m deep (average depth) wereexcluded so that backpack electrofishing couldbe used to collect fish. These exclusions elimi-nated 3% of the randomly drawn samples butwere necessary because the wide stream widthand deeper water reduced our ability to effi-ciently collect small nongame fish, making fishcapture data unreliable at these locations.
We attempted to sample 100 linear metersof stream at each survey location, but due tostream conditions, riparian vegetation, and ourability to set lower and upper blocknets, studyreaches ranged from 25 to 337 m in length (x– =98 m). Blocknets (7-mm mesh size) were in -stalled to prevent fish movement out of thereach during sampling.
We collected fish with Smith-Root Model15D backpack electrofishers at settings of 1–3ms pulses, 30–60 Hz, and 200–700 V. One to 4electrofishing passes were conducted using1–3 electrofishers (depending on stream size,water volume, and stream gradient) to depletesalmonids for population estimation. At thesame time, nongame fish species were nettedand identified to species, but they were notdepleted. Rather, while electrofishing, we ap -proximated an index of abundance by visuallyestimating the numerical abundance of eachnongame fish species. This approximationoften was accomplished by netting each non -game fish observed, but, at reaches wheresome species were highly abundant, estimatesof abundance for some species were approxi-mated by counting fish of that species as theywere stunned, without netting every fish. Afterall the passes were completed, we used a scalesimilar to Grossman et al. (1998) to categorizeeach species (at a scale of 100 linear meters ofstream, which was about our average reachlength) as absent (0), sparse (1–10), common(10–50), or abundant (50–100 or more).
We vouchered a total of 981 nongame fishspecimens from 107 study reaches throughoutthe study area to corroborate our identifica-tions of fish species. Vouchered specimens werepreserved in 10% formalin and were later
2013] NONGAME FISH STATUS IN SOUTHERN IDAHO 23

identified by zoological staff at the Orma J.Smith Museum of Natural History. Field crewsidentified species correctly 86% of the time,and most errors involved incorrectly identify-ing Paiute or shorthead sculpin as mottledsculpin. Because the nonsculpin error rate waslow (6%), we assumed our occupancy (i.e.,present or absent) and relative abundance datawere correct for all species except sculpin. We corrected errors for these 3 sculpin spe -cies at reaches where vouchers were col-lected. In addition, we collected an additional409 sculpin specimens from 41 additional studyreaches to verify sculpin species distributionin the study area. Finally, the database of vouch -ered sculpin specimens at the Orma J. SmithMuseum (n = 1361 records for the study area)was used to corroborate any further question-able identifications.
Abiotic and Biotic Stream Conditions
We assessed several abiotic and biotic streamconditions that we felt could potentially influ-ence nongame fish occurrence or abundanceat a study reach. Elevation (m) was determinedfrom USGS 1:24,000-scale maps at the down-stream end of each reach, based on coordi-nates obtained from GPS units. Stream gradi-ent (%) was estimated using All Topo MapsVersion 2.1 for Windows software (iGage Map-ping Corporation, Salt Lake City, UT) by trac-ing (at 1:24,000 scale) the length of streambetween 2 or 3 contour lines incorporating thereach (approximately 1 km) and dividing thestream length by the elevation change betweenthe contour lines. Specific conductivity (mS ⋅cm–1) was measured with a calibrated, hand-held conductivity meter accurate to +–2%.
Ten equally spaced transects were estab-lished throughout each sample reach and wereused for the remaining measurements. Aver-age stream wetted width (m) was calculatedfrom the 10 transect readings. Across transects,mean water depth was estimated by using astadia rod to measure depth at 1/4, 1/2, and3/4 distance across the channel, and the sum ofthe measurements was divided by 4 to ac countfor zero depths at the stream margins for trape -zoidal-shaped channels (Arend 1999). Fromthese measurements, we calculated the width :depth ratio. Substrate composition was visu-ally estimated as the percentage of stream bot-tom within 1 m of each transect that was com-prised of silt (<0.06 mm, diameter of b axis),
sand (0.06–1.99 mm), gravel (2–63 mm), cobble(64–249 mm), boulder (250–3999 mm), or bed -rock (>4000 mm). To reduce the number ofindependent variables considered in our analy -ses, however, silt, sand, and bedrock were dis-carded because they were heavily skewed to -ward zeros or low values. Cobble and bouldersubstrate likely affect many fishes in a similarmanner (Allan 1995) and were therefore com-bined into one percentage value. Percent un -stable banks and stream shading were alsovisually estimated within 1 m of each transect.All ocular estimates were averaged across alltransects, yielding an overall mean for eachstudy reach.
One additional stream condition that wasconsidered to potentially affect occupancy andrelative abundance of nongame fish specieswas trout density in the stream reach. In ourstudy area, we encountered rainbow trout On -corhynchus mykiss, cutthroat trout Oncorhyn -chus clarkii, brook trout Salvelinus fontinalis,bull trout Salvelinus confluentus, and browntrout Salmo trutta, all of which prey on non -game fish species in some stream settings(Scott and Crossman 1973, Wydoski and Whit-ney 2003). Consequently, we pooled the abun-dance of all trout as an additional stream con-dition that may have influenced the distributionand abundance of nongame fish species in ourstudy area.
Data Analyses
For the 9 species that occupied at least 30study reaches, we used logistic regression analy -sis to assess the relationship between the abioticand biotic stream conditions and the relativeabundance of nongame fish species. A benefit ofusing logistic regression is that the techniquedoes not require that variables be normally dis-tributed or of equal variance (Ta bachnick andFidell 1989). We assessed multicollinearity inthe independent variables by calculating vari-ance inflation factors (VIFs), all of which were<5.0, indicating that multicol linearity was lowin our data set (Menard 1995). Moreover, cor-relation coefficients (r) between independentvariables were >0.40 for only 2 comparisons(stream width vs. stream depth, r = 0.53; streamwidth vs. width : depth ratio, r = 0.58).
Separate logistic regression models wereconstructed for each species. For the responsevariable, we assigned index values to the relativeabundance data (0 = absent, 1 = sparse, 2 =
24 WESTERN NORTH AMERICAN NATURALIST [Volume 73

common, 3 = abundant) and to the occupancydata (0 = absent, 1 = present), and we pro-duced models for each, but because the re sultswere nearly identical, we present only the oc -cupancy models. Stepwise methods were usedfor including independent variables in the mod -els, and adjusted R2 for discrete models (here-after termed R̃2; Nagelkerke 1991) was used toassess the amount of variation explained bythe models. Only first-order in teractions weretested for significance, and they were rarelysignificant for the best models. To reduce type-I error rate inflation caused by the inclusion ofmultiple independent variables in multiple in -dividual logistic regressions, we set a = 0.01for stepwise inclusion. For each species, weincluded in our analyses only those studyreaches within the native distribution for thatparticular species. We used the Hosmer andLemeshow goodness-of-fit test to ensure thedata adequately fit logistic regression modelsfor each species, and this test failed to rejectthe fit of any of our final models. We used theestimate of the area under the receiver operat-ing characteristic (ROC) curve to assess thepredictive ability of the final mod els (Hanleyand McNeil 1982). We did not in clude anyquadratic terms because species oc cupancydata was not parabolic in shape when plottedagainst any abiotic or biotic stream conditions.
In addition to the above logistic regressionanalyses, we used canonical correlation analy-ses to relate the suite of abiotic and bioticstream conditions to the relative abundance of
all non game fish species simultaneously. Canoni -cal correlation analysis is a multivariate statis-tical model that facilitates the study of relation-ships among a set of multiple dependent vari-ables and multiple independent variables. Thisanaly sis reduces the concern of committing typeI errors due to multiple statistical tests on thesame independent variables (thus we relaxed ato 0.05). Canonical correlation analysis is there-fore an ideal analytical tool for elucidating rela-tionships between biological assemblages ofspecies and their environment (ter Braak andVerdonschot 1995).
Since sampling efficiency of using backpackelectrofishers is generally low for most stream-dwelling nongame fish species (Reynolds et al.2003, Burns 2007, Reid et al. 2008), we as -sumed that, when indexing fish abundance, weobserved or captured only a fraction of thetotal number of fish of each species in anygiven study reach. Accordingly, we roundedeach abundance index value to the upper endof the category (e.g., all “sparse” delineationswere given an abundance value of 10). Thesedata were square-root transformed prior tomodel construction to increase linearity (i.e.,moderate skewness) in the fish–habitat rela-tionships. As recommended by Hair et al. (1998),we interpreted relationships between fish andhabitat parameters by using canoni cal cross-loadings, and we used loading values of 0.3and greater to indicate which parameters wererobust contributors for each interpretable ca -nonical function (Tabachnick and Fidell 1989).Because the native distribution of several spe -cies was interrupted by Sho shone Falls, we con -ducted separate canonical correlation analy sesfor data above and below the falls. All statisti-cal analyses were performed with SAS statisticalsoftware (SAS Institute, Inc. 2009).
Within the study area, 4 species of sculpinoccur, one of which (Wood River sculpin) isendemic only to the Big Wood River drainage.Fish–habitat relationships have already beendeveloped for this species (Meyer et al. 2008)based on data collected as part of this study;therefore, we did not include this species inour analyses. Although the remaining 3 spe -cies of sculpin (mottled, Paiute, and short-head) sometimes have disparate habitat pref-erences and distribution patterns (Peden et al.1989, Quist et al. 2004b), in the Snake Riverbasin of southern Idaho they appear to fill asimilar habitat niche across the landscape,
2013] NONGAME FISH STATUS IN SOUTHERN IDAHO 25
TABLE 1. Descriptive statistics for various abiotic andbiotic stream parameters at study reaches (n = 1152) inthe Snake River basin of southern Idaho. Dry reaches (n= 586) were not included in these calculations.
Stream habitatparameter Mean SD Min. Max.
Stream order 2.2 1.0 1.0 6.0Elevation (m) 1747 328 696 2964Stream gradient (%) 3.6 3.6 0.1 32.5Specific conductivity 200 170 4 1585
(mS ⋅ cm–1)Stream width (m) 3.3 2.4 0.5 14.2Average depth (m) 0.13 0.15 <0.01 4.30Width : depth ratio 26.9 13.1 1.7 127.0Gravel substrate (%) 29 11 0 80Cobble-boulder 34 19 0 100
substrate (%)Stream shading (%) 21 21 0 88Unstable banks (%) 8 15 0 88Trout density 0.16 0.32 0.00 3.91
(number ⋅ m–2)

with allopatric-sympatric distribution patternsthat appear somewhat indiscriminate withinand among drainages. Thus, although we mod -eled each species separately, we also com-bined all sculpin species into one metric toassess whether this combination better de -scribed the distribution and abundance of scul -pin in the study area, compared to what wasascertained for individual species.
RESULTS
A total of 1738 reaches were surveyed insmall streams in the Snake River basin aboveHells Canyon Dam. Stream characteristicsvaried widely at the study reaches we samp -led (Table 1), ranging from 696 to 2964 m inele vation, 0.1% to 32.5% in gradient, and 4 to
1585 mS ⋅ cm–1 in specific conductivity. Of thereaches surveyed, 30% were in first-orderstreams, 39% were second-order, 22% werethird-order, 7% were fourth-order, and <3%were fifth- and sixth-order.
Of the 1738 reaches surveyed, 34% were dryor nearly dry (i.e., contained too little water tosupport any fish species; Table 2). An additional21% did not contain any fish, even though flow -ing water was present during sampling. At leastone species of nongame fish was captured at30% of the stream reaches surveyed. All 18native nongame fish species still present in thestudy area were captured during our surveys.
The most widely distributed species wasspeckled dace (occupying 21% of surveyedreaches where they were potentially native,
26 WESTERN NORTH AMERICAN NATURALIST [Volume 73
TABLE 2. Number of reaches surveyed and percentage of reaches in which native nongame fish species were collected known or probable native range for each species. Reaches unoccupied by a species are left blank to improve visualiza-
Number of reaches surveyed 156 208 61 115 105 91 90 19 67 7Percentage of dry or nearly 41.0 63.9 19.7 11.3 11.4 42.9 54.4 57.9 47.8 57.1
dry reachesPercentage of reaches with 50.0 33.7 65.6 62.6 76.2 54.9 32.2 42.1 47.8 42.9
fish (trout included)Percentage of reaches with 32.7 22.1 29.5 16.5 44.8 36.3 27.8 21.1 40.3 14.3
nongame speciesPercentage of reaches with 38.5 21.6 57.4 55.7 69.5 46.2 23.3 36.8 34.3 42.9
troutPercentage of reaches withc
Bluehead suckerBridgelip sucker 29.3 22.7 6.1 4.9 6.5 46.2 14.6 12.5 45.7Largescale sucker 5.3 2.9 1.9 2.9Mountain sucker 2.7 1.9 5.7Utah suckerChiselmouth 3.3 2.7 2.2 7.7 8.6Leopard dace 12.5Longnose dace 7.6 4.0 10.2 2.9 3.2 30.8 7.3 12.5 2.9Speckled dace 46.7 54.7 12.2 11.8 2.2 48.1 14.6 50.0 65.7Redside dace 21.7 30.7 8.2 5.9 6.5 51.9 7.3 12.5 48.6Northern leatherside chub 6.3a
Utah chubPeamouth 1.1Northern pikeminnow 4.3 13.3 2.0 3.9 4.3 11.5 8.6Mottled sculpin 4.0 10.2 8.8 5.4 3.8 12.5 33.3Paiute sculpin 13.3 48.6Shorthead sculpin 1.3 20.4 3.9 44.1 5.8Wood River sculpin 48.8
aMay have been misidentifiedbWe did not include Big Lost River, Little Lost River, or Birch Creek in the Sinks drainagescEstimated from the surveryed reaches where each species was potentially native, excluding dry and nearly dry reaches.
Snak
etr
ibut
arie
sbe
-lo
wSh
osho
neFa
lls
Ow
yhee
Riv
er
Wei
ser
Riv
er
Paye
tte
Riv
er
Boi
seR
iver
Bru
neau
Riv
er
Big
Woo
dR
iver
Roc
kC
reek
(Tw
in)
Salm
onFa
llsC
reek
Dry
Cre
ek

excluding dry and nearly dry reaches; Table 2),followed by bridgelip sucker (19%), Paiute scul -pin (16%), redside shiner (14%), shortheadsculpin (12%), and mottled sculpin (12%). Al -though Wood River sculpin occupied 49% ofthe surveyed reaches within their native range,excluding dry and nearly dry reaches, theywere native to only one river drainage. Spe -cies found in ≤1% of the reaches where theywere potentially native included leopard daceand peamouth, whereas northern leathersidechub, largescale sucker, Utah chub, and blue-head sucker were found in ≤2% of the reacheswhere they were potentially native.
Common carp Cyprinus carpio and orien -tal weatherfish Misgurnus anguillicaudatuswere the only nonnative nongame fish speciesencountered during our surveys. Common
carp were captured at one location each in theRock Creek (Twin Falls), Portneuf River, andBoise River drainages. Oriental weatherfishwere caught at 2 locations in the Boise Riverdrainage, including one reach that also con-tained common carp. These 2 species werecategorized as ‘sparse’ at each location wherethey were captured, and they were too rare toinclude in our statistical analyses.
When present, redside shiners were mostoften categorized as abundant (59% of thereaches they occupied; Table 3), as were speck -led dace (55%) and all species of sculpin (onaverage, 50%). Commonly occurring spe ciesthat were rarely categorized as abundant in -cluded bluehead sucker (10%), mountain sucker(15%), chiselmouth (21%), and northern pike -minnow (22%).
2013] NONGAME FISH STATUS IN SOUTHERN IDAHO 27
with backpack electrofishers in small streams in the Snake River basin of southern Idaho. Shaded gray areas indicate the tion of the data.
10 81 92 16 6 82 76 80 64 58 89 87 78 173820.0 37.0 31.5 87.5 33.3 12.2 18.4 33.8 9.4 39.7 27.0 18.4 25.6 33.7
80.0 51.9 58.7 6.3 50.0 63.4 75.0 60.0 78.1 51.7 60.7 62.1 56.4 55.2
30.0 27.2 27.2 6.3 33.3 36.6 43.4 28.8 51.6 43.1 18.0 33.3 15.4 30.2
80.0 45.7 58.7 0.0 50.0 58.5 56.6 57.5 75.0 39.7 60.7 58.6 55.1 47.8
13.7 3.8 1.7 2.8 2.019.21.6
3.9 6.3 8.3 16.1 3.8 12.1 20.0 2.8 4.13.9 1.6 2.8 3.2 3.4 17.1 1.7 2.6
2.80.2
13.7 9.5 4.8 19.0 8.6 7.7 5.6 7.031.4 9.5 5.6 27.4 7.5 19.0 25.7 9.2 14.1 1.7 21.417.6 11.1 1.4 6.5 1.9 10.3 31.4 1.5 9.9 1.7 13.511.8 1.7 1.2
3.9 1.6 1.4 11.3 1.80.26.3
37.5 29.4 36.5 11.1 17.7 9.4 22.4 42.9 7.7 12.7 12.011.8 4.8 50.0 50.0 31.9 22.6 30.2 58.6 22.9 23.1 19.7 15.5
19.0 12.448.8
Mar
shC
reek
Goo
seC
reek
Raf
tRiv
er
Roc
kC
reek
Ban
nock
Cre
ek
Port
neuf
Riv
er
Bla
ckfo
otR
iver
Sout
hFo
rkSn
ake
(Ban
nock
)
Riv
er
Salt
Riv
er
Will
owC
reek
Teto
nR
iver
Hen
rys
Fork
Snak
eR
iver
Sink
sdr
aina
gesb
TO
TAL

The canonical correlation model constructedfor species occurring below Shoshone Fallsyielded 12 canonical functions, which collec-tively produced a statistically significant model(Wilks’ l = 0.27, F144, 4307 = 4.91, P < 0.0001)that explained a combined 73% of the varianceshared between the variable sets across allfunctions. However, dimension reduction analy -sis indicated that only the first 4 functions(with squared canonical correlations of 0.49,0.18, 0.16, and 0.11) explained a significant (a= 0.05) amount of shared variance betweenthe variable sets. Moreover, only the first func-tion contained variables with cross-loadings of0.30 and greater, which were thereby consid-ered interpretable. This function explained49% of the shared variance between the vari-able sets and indicated that the relative abun-dance of several species, including redsideshiner, speckled dace, bridgelip sucker, north-ern pikeminnow, and chiselmouth, were allpositively related to an increase in stream order(i.e., farther from headwaters), stream width,mean depth, and conductivity (Table 4). Thesespecies were all negatively related to streamgradient, stream shading, and elevation.
For species occurring above Shoshone Falls,the canonical correlation model was strikinglysimilar, yielding 12 canonical functions whichcollectively produced a statistically significantmodel (Wilks’ l = 0.33, F132, 4290 = 4.91, P<0.0001) that explained a combined 67% ofthe variance shared between the variable sets
across all functions. Dimension reduction analy -sis indicated that the first 4 functions (withsquared canonical correlations of 0.47, 0.15,0.09, and 0.09) explained a significant amountof shared variance between the variable sets,but, once again, only the first function con-tained variables with cross-loadings 0.30 andgreater. This function explained 47% of theshared variance between the variable sets andindicated that the relative abundance of speck -led dace, mottled sculpin, longnose dace, red-side shiner, mountain sucker, and Paiute scul -pin were all positively related to an increase instream order, mean depth, and stream wettedwidth (Table 4). These species were all nega-tively related to stream gradient and elevation.
Similar relationships were observed withlogistic regression analyses (Table 5), and re -sults were largely consistent for families offish. For example, bridgelip and mountainsucker presence was more likely in reacheswith lower gradient and less shading. Moun-tain sucker were also more likely to occupyreaches with less cobble-boulder substrate,whereas bridgelip sucker were more likely tooccupy higher-order, lower-elevation streamreaches. Cyprinids (i.e., longnose dace, speck-led dace, redside shiner, and northern pike -minnow) were also more likely to occupylower gradient, lower elevation, higher-orderreaches. In addition, longnose dace were morelikely where cobble-boulder substrate wasmore abundant, whereas speckled dace and
28 WESTERN NORTH AMERICAN NATURALIST [Volume 73
TABLE 3. Percentage of study reaches (excluding dry and nearly dry reaches) in small streams in the Snake River basinof southern Idaho where 18 fish species were absent, sparse, common, and abundant. Calculations for a species includeddata only from the drainages in which it was native.
Species Absent Sparse Common Abundant
Bluehead sucker 98.0 0.8 1.0 0.2Bridgelip sucker 80.8 4.0 5.5 9.7Largescale sucker 98.4 0.9 0.0 0.7Mountain sucker 95.9 1.7 1.8 0.5Utah sucker 97.4 0.3 1.6 0.7Chiselmouth 97.2 1.2 1.0 0.6Leopard dace 99.8 0.2 0.0 0.0Longnose dace 93.0 1.6 2.8 2.6Speckled dace 78.6 3.5 6.1 11.8Redside shiner 86.5 1.8 3.7 7.9Northern leatherside chub 98.8 1.2 0.0 0.0Utah chub 98.2 0.5 0.7 0.7Peamouth 99.8 0.2 0.0 0.0Northern pikeminnow 93.7 2.6 2.4 1.4Mottled sculpin 88.0 2.8 4.6 4.6Paiute sculpin 84.5 3.6 5.5 6.4Shorthead sculpin 87.6 1.8 4.3 6.4Wood River sculpin 51.2 4.9 9.8 34.1

redside shiner were more likely where troutdensity was lower.
There was less consistency in the logisticregression models, and the models were gen-erally weaker for cottids. Paiute and shortheadsculpin were both positively associated withhigher stream order and more cobble-bouldersubstrate. Paiute sculpin were also positivelyassociated with higher elevation but were neg-atively associated with gradient, conductivity,and stream width. Mottled sculpin were posi-tively associated with conductivity and nega-tively associated with stream gradient. Whenall cottid species were combined into a con-glomerate ‘occupancy’ metric, sculpin specieswere positively associated with stream order,elevation, gradient, stream width, and cobble-boulder substrate, but negatively associated
with conductivity. Across all species, individ-ual logistic regression models on average ex -plained 34% of the variation (range 12%– 53%)in the presence/absence of native non gamefish species (Table 5). Predictive power washigh for nearly all models, with area under theROC curve averaging 0.84 across all modelsand ranging from 0.73 to 0.92 (Table 5).
DISCUSSION
The results of our study suggest that thedistribution and abundance of many nativenongame fish species in the Snake River basinof southern Idaho were influenced by abioticand biotic stream conditions across the land-scape. In general, catostomids and cyprinidsformed a composite fish assemblage associatedwith higher-order, lower-elevation, lower-gra-dient reaches where conductivity was higherand streams were wider and deeper. Previousstudies have demonstrated that similar cato -stomid-cyprinid fish assemblages (of generallythe same species) were associated with similarstream conditions in western Wyoming (Quistet al. 2004b), eastern Oregon (Torgersen et al.2006), and southern British Columbia (Porteret al. 2000). Surprisingly, nearly the exact sameabiotic and biotic stream conditions scoredcanonical cross-loadings of 0.30 or greater formodels applied to river drainages above andbelow Shoshone Falls (Table 4), despite thefact that 6 of the 14 species included in theanalyses occurred only above or only belowthe falls. The similarity between these studies,and between our models applied to drainagesboth above and below Shoshone Falls, high-lights the consistency in the influence thatsome reach-scale abiotic and biotic streamconditions may have on the abundance of non -game fish species, regardless of species com-position or the additional influence of larger,regional-scale environmental conditions suchas climate or geology.
Stream order was the strongest loading factor for canonical correlations in modelsapplied to drainages above and below Sho -shone Falls, suggesting that the increase incatostomids-cyprinids was occurring along alongitudinal downstream gradient. Longitudi-nal changes in fish assemblage structure aregenerally thought to be due either to continualspecies additions to downstream reaches (Shel -don 1968) or to biotic zonation resulting from
2013] NONGAME FISH STATUS IN SOUTHERN IDAHO 29
TABLE 4. Canonical cross-loadings for the first canonicalfunction relating abiotic and biotic stream conditions tothe relative abundance of select nongame fish species inthe Snake River basin of southern Idaho. Separate modelswere run for sections downstream and upstream fromShoshone Falls. Only variables with loadings 0.30 andgreater (in bold) should be considered interpretable.Missing values indicate where the species was not native.
From Shoshone Falls_______________________Variable Downstream Upstream
Stream habitat parameterStream order 0.63 0.56Elevation –0.35 –0.30Gradient –0.47 –0.42Conductivity 0.36 0.25Stream width 0.39 0.44Average depth 0.30 0.49Width : depth ratio 0.12 0.01Percent gravel substrate 0.10 –0.17Percent cobble- –0.20 –0.06
boulder substrateStream shading –0.38 –0.21Unstable banks 0.12 0.19Trout density –0.22 –0.10
Species relative abundanceBluehead sucker — 0.17Bridgelip sucker 0.54 —Largescale sucker 0.13 —Mountain sucker 0.16 0.37Utah sucker — 0.29Chiselmouth 0.31 —Longnose dace 0.28 0.39Speckled dace 0.56 0.42Redside shiner 0.60 0.38Northern pikeminnow 0.40 —Utah chub — 0.14Mottled sculpin 0.07 0.39Paiute scuplin 0.10 0.37Shorthead sculpin –0.02 0.07Sculpin combined 0.09 0.54

changes in geomorphology or thermal charac-teristics (Huet 1959). In our study, speciessuch as sculpin, which are more closely associ-ated with headwater streams, were still pre-sent at many of the lower elevation reaches,suggesting that biotic zonation perhaps hadnot yet materialized in our study for at leastsome species. However, as previously men-tioned, we could not quantitatively samplestreams larger than 15 m wetted width, thuszonation may have been unlikely to have oc -curred (or perhaps could not be detected) atthe edge of our sampling framework.
Two cyprinids, speckled dace and redsideshiner, were especially widespread and abun-dant. They were captured in all major riverdrainages (i.e., those greater than 1000 km2)and most of the smaller drainages (Table 2),and were more often categorized as abundantthan were most other nongame species (Table3). Both species are habitat generalists (Pear-sons et al. 1992, Wydoski and Whitney 2003)and thus would be expected in a variety ofstream conditions. Nevertheless, both showedan affinity for higher-order, lower-gradient,lower-elevation reaches. Longnose dace wasthe only species of the catostomid-cyprinidfish assemblage that was directly associatedwith cobble-boulder substrate. A positive as -sociation for longnose dace with cobble-boul-der substrate has been demonstrated previ-ously (Mullen and Burton 1995, Thompson etal. 2001) and probably results from longnosedace using substrates as shelter from the cur-rent (Culp 1989) or as a foraging mechanismto feed on macroinvertebrates on the surfaceof larger substrates (Thompson et al. 2001).None of the remaining cyprinids or catosto-mids were positively associated with cobble-boulder substrate, but 2 of the 3 sculpin spe -cies (and the conglomerate sculpin metric)were. Direct associations with rocky substratehave been repeatedly demonstrated for manystream-dwelling sculpin (e.g., Brown 1991,Hesthagen and Heggenes 2003, Meyer et al.2008) and are likely the result of shelter, forag-ing, and spawning needs (Wydoski and Whit-ney 2003).
At least one of the 3 main Cottus species in the Snake River basin of southern Idaho(Paiute, mottled, and shorthead sculpin) waspresent in 317 reaches, and 2 species weresympatric in 49 reaches; no reaches containedall 3 species. The prevalence of Cottus species
30 WESTERN NORTH AMERICAN NATURALIST [Volume 73TA
BL
E5.
Log
istic
reg
ress
ion
mod
el r
esul
ts r
elat
ing
fish
occu
panc
y (i.
e., p
rese
nt o
r ab
sent
) at
stud
y re
ache
s in
the
Sna
ke R
iver
bas
in t
o ab
iotic
and
bio
tic s
trea
m c
ondi
tions
with
inth
e re
ach.
Str
eam
con
ditio
ns t
hat
wer
e si
gnifi
cant
ly (a
= 0
.01)
rel
ated
to
a sp
ecie
s’ o
ccup
ancy
are
ind
icat
ed b
y pl
us (
dire
ct r
elat
ions
hip)
or
min
us (
inve
rse
rela
tions
hip)
sym
bols
.A
naly
ses
incl
uded
onl
y th
ose
spec
ies
that
occ
upie
d at
leas
t 30
reac
hes.
RO
C =
rec
eive
r op
erat
ing
char
acte
rist
ic.
Stre
am h
abita
t par
amet
erM
odel
str
engt
h__
____
____
____
____
____
____
____
____
____
____
____
____
____
____
____
____
____
____
____
____
____
____
____
____
____
___
____
____
____
____
__W
idth
:Pe
rcen
tPe
rcen
tPe
rcen
tPe
rcen
tE
stim
ated
Stre
amE
leva
-G
radi
-C
ondu
c-M
ean
Mea
nde
pth
grav
elco
bble
-bou
lder
stre
amun
stab
leTr
out
area
und
erSp
ecie
sor
der
tion
ent
tivity
wid
thde
pth
ratio
subs
trat
esu
bstr
ate
shad
ing
band
sde
nsity
R̃2
RO
C c
urve
Bri
dgel
ip s
ucke
r+
––
–0.
530.
90M
ount
ain
suck
er–
––
0.22
0.85
Lon
gnos
e da
ce+
–+
0.35
0.88
Spec
kled
dac
e+
––
+–
0.48
0.89
Red
side
shi
ner
+–
––
0.52
0.92
Nor
ther
n pi
kem
inno
w+
––
0.47
0.92
Mot
tled
scul
pin
–+
0.15
0.74
Paiu
te s
culp
ina
++
––
–+
0.26
0.81
Shor
thea
d sc
ulpi
n+
++
0.12
0.73
Scul
pin
com
bine
db+
++
–+
+0.
320.
80a A
lso
sign
ifica
nt w
ere
the
inte
ract
ion
term
s co
nduc
tivity
* w
idth
(+) a
nd c
ondu
ctiv
ity *
gra
dien
t (+
)b A
lso
sign
ifica
nt w
ere
the
inte
ract
ion
term
s el
evat
ion
* gr
adie
nt (–
), el
evat
ion
* co
nduc
tivity
(+),
and
cond
uctiv
ity *
wid
th (+
)

was not surprising since they are known toinhabit small mountain streams more so thanmost other nongame fish species in the Inter-mountain West. However, fish–habitat associa-tions for sculpin were generally weaker andless consistent than for other nongame speciesin our study. Also, and logistic regression modelresults differed, and the models were weakerfor individual sculpin species compared to theconglomerate sculpin metric. As we originallyhypothesized, perhaps these sculpin speciesfill a somewhat similar habitat niche across theSnake River basin in southern Idaho. If so, theweakness of individual sculpin models couldstem in part from a somewhat haphazard dis-tribution influenced more by geomorphic pro -cesses (Smith 1978) and stream capture events(Burridge et al. 2007) than by abiotic and bio -tic stream conditions. At smaller scales withinthe Snake River basin, habitat features maymore prominently influence distribution pat-terns for sculpin populations. For instance, inone of the drainages included in our study(i.e., the Salt River), allopatric and sympatricdistributions of Paiute and mottled sculpinwere associated with elevation and thermalcharacteristics (Quist et al. 2004b). Alterna-tively, others have argued that, for sculpin popu -lations, microhabitat and macrohabitat con di -tions are of little importance; rather, they areregulated most strongly by density-dependentprocesses such as competition for food andspace (Petty and Grossman 1996, Grossman etal. 2006).
Surprisingly, nonnative nongame fish spe -cies were almost entirely absent from smallstreams in the Snake River basin in southernIdaho. While this is an encouraging finding, itshould be noted that nonnative nongame spe -cies are prevalent in many larger rivers andlentic habitats in southern Idaho (IDFG 2007).In fact, 13 nonnative nongame fish speciesexist in the Snake River basin in southernIdaho, 9 of which were likely established fromthe release of aquarium pet fish, a problemthat will probably worsen in the coming dec -ades (Gertzen et al. 2008, Strecker et al. 2011).Of the remaining 4 species, only common carpand grass carp Ctenopharyngodon idella arewidely distributed in southern Idaho. Idaho islargely devoid of commonly used bait fishsuch as fathead minnow Pimephales promelas,golden shiner Notemigonus crysoleucas, andred shiner Cyprinella lutrensis, in part because
the use of live bait fish has been prohibited inIdaho since 1960, with other regulations dat-ing back to 1941 that restricted most use oflive bait for Idaho anglers. At present, exoticnongame fish have apparently not yet invadedsmall streams in southern Idaho, although cli-mate change may increase the likelihood ofsuccessful invasions as warmer water tempera-tures alter the types of fish that can establishself-sustaining populations (Rahel and Olden2008).
In our study, much of the variation in theoccupancy and relative abundance of nativenongame fish species in the Snake River basinwas unexplained by our fish–habitat models(both canonical correlation and logistic regres-sion), suggesting that nongame fish speciespopulation metrics were additionally influ-enced by other environmental factors that wedid not measure. One such factor may bestream flow, which has been shown to influ-ence nongame fish species assemblages (Poffand Allan 1995, Grossman et al. 1998), usuallyvia effects on mortality and subsequent re -cruitment. Water temperature has also beenshown to affect fish species assemblages (Porteret al. 2000, Quist et al. 2004b), although ourmetric of elevation might be considered a sur-rogate for water temperature (Bozek and Hu -bert 1992). Moreover, as our models consis-tently indicated, most of the species found inour study (except sculpin species) occurred atlower-elevation streams; thus, their occupancyand abundance were likely more closely asso-ciated with stream conditions in larger streamsat lower elevations, rather than the smallstreams in our study. Unfortunately, quantita-tive sampling techniques for nongame fishspecies in larger streams are difficult to imple-ment, and the abundance of many of thesespecies (e.g., dace and shiners) can be over-whelming both in small streams and largerivers, due mainly to their schooling nature,making it difficult to quantitatively samplethem in an unbiased manner. Fish–habitat rela -tionships are often weak and not applicable toareas outside the study area where the rela-tionships were developed (Fausch et al. 1988),in part because complex interactions betweenmultiple habitat factors simultaneously affect-ing organisms are not all measured and ac -counted for in statistical models (Cade and Noon2003), nor are they necessarily consistent spa-tially or temporally.
2013] NONGAME FISH STATUS IN SOUTHERN IDAHO 31

Our probability of detecting a species at anygiven reach was not equal to one. Thus, at anunknown number of locations, we mistakenlyconcluded that a species was absent when infact it was present at a reach. Not accountingfor imperfect detection may have led to in -accurate conclusions regarding the magnitudeof effect for each variable included in our mod -els (Mac Kenzie 2005). Indeed, imperfect detec -tion probability is an active area of research inapplied ecology (e.g., MacKenzie et al. 2002,He and Gaston 2003, Royle et al. 2005). Falsespecies absences in our study probably oc -curred most often for the rarest species, and wedid not attempt to identify stream conditionsthat influenced their occupancy. For the re -maining species that were more abundant, er -rors still occurred at locations where their abun-dance was very low, but we do not be lieve thatfalse absences in our data introduced substan-tial bias to our conclusions. Meyer and High(2011) concluded that, at measured capture effi-ciencies of 20%–60% for salmonids in the SnakeRiver basin, if abundance was as low as only 2fish in 100 m of stream, the likelihood of catch-ing at least one of these fish with 3 depletionpasses would be about 95%. Although sculpinare more difficult to capture during electrofish-ing than salmonids (because sculpin are smallerand are less buoyant be cause they have no swimbladder), capture probabilities for Wood Riversculpin were only about 35% lower than forsalmonids (K. Meyer unpublished data).
Within the small streams of the Snake Riverbasin above Hells Canyon Dam, we captured all18 native nongame fish species still present inthe study area. This was somewhat surprisingbecause we surveyed <1% of the stream kilo-meters included in our study area, and somespecies such as northern leatherside chub areextremely rare in the Snake River basin, whileothers are more closely associated with largerrivers (e.g., chiselmouth, northern pikeminnow)that we excluded from our study. While it isencouraging that all native nongame fish spe -cies were represented, most species were ab -sent from a high percentage of study reaches,and, even when present, most species wererarely abundant. We believe that, for many spe -cies, this characteristic may be reflective of astronger affinity for larger rivers rather thanan actual decline in distribution or abundance.
Whether the distribution and abundance ofnative nongame fish species in southern Idaho
has declined from historical levels cannot beascertained from our results because (1) thereis little previous work for comparison and (2)our exclusion of large rivers makes our currentassessment incomplete. The ubiquity of stud-ies across North America demonstrating adecline in the distribution and abundance ofnative nongame fish species (e.g., Warren etal. 2000, Haslouer et al. 2005, Hoagstrom etal. 2007, Moyle et al. 2011), as well as thedeclines in native game fish within our studyarea (e.g., Thurow et al. 1997), suggest thatnongame fish species in southern Idaho havelikely experienced some level of decline. How -ever, perhaps more important is monitoringwhether existing populations maintain theircurrent distribution and abundance for theforeseeable future. Accordingly, future studiesthat fill the gaps in our data set or use our dataas a baseline for direct comparison may morecompletely elucidate the current status of na -tive nongame fish species in the Snake Riverbasin of southern Idaho. This study under-scores the need for management agencies tofocus more effort on recording data for non -game fish species (rather than just the tar-geted game species of interest) during stan-dard fish-sampling surveys, both in small,higher-elevation streams, where quantitativesampling is more easily conducted, and inlarger downstream rivers, where quantitativesampling may be more difficult but manynongame species may be more likely to occur.An added benefit is that population metricsand distributions of nongame species mayserve as better ecological indicators of changebecause the population dynamics of non gamespecies are less influenced by managementactions.
ACKNOWLEDGMENTS
We thank the many field personnel whoassisted in data collection, most notably JohnCassinelli. Reviews by Tim Copeland, ChrisSullivan, Erin Larson, Chris Walser, and 2anonymous reviewers greatly improved themanuscript. The states of Oregon, Nevada,Utah, and Wyoming generously permitted usaccess to streams within their jurisdiction.
LITERATURE CITED
ALLAN, J.D. 1995. Stream ecology, structure and functionof running water. Chapman and Hall, New York, NY.
32 WESTERN NORTH AMERICAN NATURALIST [Volume 73

ANGERMEIER, P.L., AND M.R. WINSTON. 1998. Local vs.regional influences on local diversity in stream fishcommunities of Virginia. Ecology 79:911–927.
AREND, K.K. 1999. Macrohabitat identification. Pages 75–93 in M.B. Bain and N.J. Stevenson, editors, Aquatichabitat assessment: common methods. AmericanFisheries Society, Bethesda, MD. 216 pp.
BOZEK, M.A., AND W.A. HUBERT. 1992. Segregation of res-ident trout in streams as predicted by three habi-tat dimensions. Canadian Journal of Zoology 70:886–890.
BROWN, L.R. 1991. Differences in habitat choice and be -havior among three species of sculpin (Cottus) inartificial stream channels. Copeia 1991:810–819.
BURNS, A.D. 2007. Comparison of two electrofishing gears(backpack and parallel wires) and abundances offishes of the upper Greenbrier River drainage. Mas-ter’s thesis, West Virginia University, Morgantown,WV. 72 pp.
BURRIDGE, C.P., D. CRAW, AND J.M. WATERS. 2007. Anempirical test of freshwater vicariance via river cap-ture. Molecular Ecology 16:1883–1895.
CADE, B.S., AND B.R. NOON. 2003. A gentle introductionto quantile regression for ecologists. Frontiers inEcology and the Environment 1:412–420.
CULP, J.M. 1989. Nocturnally constrained foraging of alotic minnow (Rhinichthys cataractae). Canadian Jour -nal of Zoology 67:2008–2012.
FAUSCH, K.D., C.L. HAWKES, AND M.G. PARSONS. 1988.Models that predict standing crop of stream fish fromhabitat variables: 1950–1985. General Technical Re -port PNW-GTR-213, USDA Forest Service, PacificNorthwest Research Station, Portland, OR.
GERTZEN, E., O. FAMILIAR, AND B. LEUNG. 2008. Quanti-fying invasion pathways: fish introductions from theaquarium trade. Canadian Journal of Fisheries andAquatic Sciences 65:1265–1273.
GROSSMAN, G.D., R.E. RATAJCZAK, M. CRAWFORD, AND
M.C. FREEMAN. 1998. Assemblage organization instream fishes: effects of environmental variation andinterspecific interactions. Ecological Monographs 68:396–420.
GROSSMAN, G.D., R.E. RATAJCZAK JR., J.T. PETTY, M.D.HUNTER, J.T. PETERSON, AND G. GRENOUILLET. 2006.Population dynamics of mottled sculpin (Pisces) in a variable environment: information theoretic ap -proaches. Ecological Monographs 76:217–234.
HAIR, J.F., JR., R.E. ANDERSON, R.L. TATHAM, AND W.C.BLACK. 1998. Multivariate data analysis. 5th edition.Prentice Hall, Inc., Upper Saddle River, NJ.
HANLEY, J.A., AND B.J. MCNEIL. 1982. The meaning anduse of the area under a receiver operating character-istic (ROC) curve. Radiology 143:29–36.
HASLOUER, S.G., M.E. EBERLE, D.R. EDDS, K.B. GIDO,C.S. MAMMOLITI, J.R. TRIPLETT, AND J.T. COLLINS.2005. Current status of native fish species in Kansas.Transactions of the Kansas Academy of Science 108:32–46.
HE, F., AND K.J. GASTON. 2003. Occupancy, spatial vari-ance, and the abundance of species. American Natu-ralist 162:332–342.
HESTHAGEN, T., AND J. HEGGENES. 2003. Competitivehabitat displacement of brown trout by Siberian scul -pin: the role of size and density. Journal of Fish Biol-ogy 62:222–236.
HIGH, B., K.A. MEYER, D.J. SCHILL, AND E.R.J. MAMER.2008. Distribution, abundance, and population trends
of bull trout in Idaho. North American Journal ofFisheries Management 28:1687–1701.
HOAGSTROM, C.W., S.S. WALL, J.G. KRAL, B.G. BLACK-WELL, AND C.R. BERRY JR. 2007. Zoogeographic pat-terns and faunal changes of South Dakota fishes.Western North American Naturalist 67:161–184.
HUBBS, C.L., AND R.R. MILLER. 1948. The zoological evi-dence: correlation between fish distribution andhydrographic history in the desert basins of westernUnited States. Pages 17–166 in The Great Basin,with emphasis on glacial and postglacial times. Bul-letin of the University of Utah, Biological Series 10,no. 38.
HUET, M. 1959. Profiles and biology of western Europeanstreams as related to fish management. Transactionsof the American Fisheries Society 88:155–163.
HUGHES, R.M., P.R. KAUFMANN, A.T. HERLIHY, T.M. KIN-CAID, L. REYNOLDS, AND D.P. LARSEN. 1998. A pro -cess for developing and evaluating indices of fishassemblage integrity. Canadian Journal of Fisheriesand Aquatic Sciences 55:1618–1631.
HUGHES, R.M., L.W. WANG, AND P.W. SEELBACH. 2006.Landscape influences on stream habitats and biolog-ical assemblages. American Fisheries Society Sym-posium 48, American Fisheries Society, Bethesda,MD. 697 pp.
[IDFG] IDAHO DEPARTMENT OF FISH AND GAME. 2007.Fisheries Management Plan 2007–2012. Boise, ID.
JOHNSON, J.B. 2002. Evolution after the flood: phylogeog-raphy of the desert fish Utah chub. Evolution 56:948–960.
LEE, D.S., C.R. GILBER, C.H. HOCUTT, R.E. JENKINS,D.E. MCALLISTER, AND J.R. STAUFFER. 1980. Atlas ofNorth American freshwater fishes. North CarolinaMuseum of Natural History, Raleigh, NC.
MACKENZIE, D.I. 2005. What are the issues with pres-ence-absence data for wildlife managers? Journal ofWildlife Management 69:849–860.
MACKENZIE, D.I., J.D. NICHOLS, G.B. LACHMAN, S.DROEGE, J.A. ROYLE, AND C.A. LANGTIMM. 2002.Estimating site occupancy rates when detec-tion probabilities are less than one. Ecology 83:2248–2255.
MEBANE, C.A., T.R. MARET, AND R.M. HUGHES. 2003. Anindex of biological integrity (IBI) for Pacific North-west rivers. Transactions of the American FisheriesSociety 132:239–261.
MENARD, S. 1995. Applied logistic regression analysis.Sage Publications, Thousand Oaks, CA.
MEYER, K.A., F.S. ELLE, AND J.A. LAMANSKY. 2009. Envi-ronmental factors related to the distribution, abun-dance, and life history characteristics of mountainwhitefish in Idaho. North American Journal of Fish-eries Management 29:753–767.
MEYER, K.A., AND B. HIGH. 2011. Accuracy of removalelectrofishing estimates of trout abundance in RockyMountain streams. North American Journal of Fish-eries Management 31:923–933.
MEYER, K.A., D.J. SCHILL, M.R. CAMPBELL, AND C.C.KOZFKAY. 2008. Distribution, abundance, and geneticpopulation structure of Wood River sculpin, Cottusleiopomus. Western North American Naturalist 68:508–520.
MEYER, K.A., D.J. SCHILL, J.A. LAMANSKY, M.R. CAMP-BELL, AND C.C. KOZFKAY. 2006. Status of Yellowstonecutthroat trout in Idaho. Transactions of the Ameri-can Fisheries Society 135:1329–1347.
2013] NONGAME FISH STATUS IN SOUTHERN IDAHO 33

MOYLE, P.B., J.V.E. KATZ, AND R.M. QUIÑONES. 2011.Rapid decline of California’s native inland fishes: a status assessment. Biological Conservation 144:2414–2423.
MULLEN, D.M., AND T.M. BURTON. 1995. Size-relatedhabitat use of longnose dace (Rhinichthys catarac-tae). American Midland Naturalist 133:177–183.
NAGELKERKE, N.J.D. 1991. A note on a general definitionof the coefficient of determination. Biometrika 78:691–692.
PEARSONS, T.N., H.W. LI, AND G.A. LAMBERTI. 1992. Influ-ence of habitat complexity on resistance and resil -ience of stream fish assemblages to flooding. Trans-actions of the American Fisheries Society 121:427–436.
PEDEN, A.E., G.W. HUGHES, AND W.E. ROBERTS. 1989.Morphologically distinct populations of the short-head sculpin, Cottus confusus, and mottled sculpin,Cottus bairdi, (Pisces, Cottidae), near the westernborder of Canada and the United States. CanadianJournal of Zoology 67:2711–2720.
PETTY, J.T., AND G.D. GROSSMAN. 1996. Patch selection bymottled sculpin (Pisces: Cottidae) in a southern Ap -palachian stream. Freshwater Biology 35:261–276.
POFF, N.L., AND J.D. ALLAN. 1995. Functional organizationof stream fish assemblages in relation to hydrologicalvariability. Ecology 76:606–627.
PORTER, M.S., J. ROSENFELD, AND E.A. PARKINSON. 2000.Predictive models of fish species distribution in theBlackwater drainage, British Columbia. North Ameri -can Journal of Fisheries Management 20:349–359.
QUIST, M.C., W.A. HUBERT, AND D.J. ISAAK. 2004a. Fishassemblage structure and relations with environ-mental conditions in a Rocky Mountain watershed.Canadian Journal of Zoology 82:1554–1565.
QUIST, M.C., W.A. HUBERT, AND D.J. ISAAK. 2004b. Factorsaffecting allopatric and sympatric occurrence of twosculpin species across a Rocky Mountain watershed.Copeia 2004(3):618–624.
QUIST, M.C., W.A. HUBERT, AND F.J. RAHEL. 2004c. Rela-tions among habitat characteristics, exotic species,and turbid-cyprinids in the Missouri River drainageof Wyoming. Transactions of the American FisheriesSociety 133:727–742.
RAHEL, F.J., AND J.D. OLDEN. 2008. Assessing the effectsof climate change on aquatic invasive species. Con-servation Biology 22:521–533.
RASHLEIGH, B., R. PARMAR, J.M. JOHNSTON, AND M.C.BARBER. 2005. Predictive habitat models for the oc -currence of stream fishes in the Mid-Atlantic High-lands. North American Journal of Fisheries Manage-ment 25:1353–1366.
REID, S.M., N.E. JONES, AND G. YUNKER. 2008. Evaluationof single-pass electrofishing and rapid habitat assess-ment for monitoring redside dace. North AmericanJournal of Fisheries Management 28:50–56.
REYNOLDS, L., A.T. HERLIHY, P.R. KAUFMANN, S.V. GRE-GORY, AND R.M. HUGHES. 2003. Electrofishing ef -fort requirements for assessing species richness andbiotic integrity in western Oregon streams. NorthAmerican Journal of Fisheries Management 23:450–461.
ROYLE, J.A., J.D. NICHOLS, AND M. KÉRY. 2005. Modelingoccurrence and abundance of species when detec-tion is imperfect. Oikos 353:353–359.
SAS INSTITUTE, INC. 2009. SAS/STAT® 9.2 user’s guide.2nd edition. SAS Institute, Inc., Cary, NC.
SCHILL, D.J. 2009. Population studies of desert redbandtrout. Doctoral dissertation, University of Idaho,Moscow, ID. 180 pp.
SCOTT, M.C., AND L.W. HALL. 1997. Fish assemblages asindicators of environmental degradation in Marylandcoastal plain streams. Transactions of the AmericanFisheries Society 126:349–360.
SCOTT, W.B., AND E.J. CROSSMAN. 1973. Freshwater fishesof Canada. Fisheries Research Board of Canada Bul-letin 184.
SHELDON, A.L. 1968. Species diversity and longitudinalsuccession in stream fishes. Ecology 49:193–198.
SIMPSON, J.C., AND R.L. WALLACE. 1982. Fishes of Idaho.University of Idaho Press, Moscow, ID. 238 pp.
SMITH, G.R. 1978. Biogeography of Intermountain fishes.Great Basin Naturalist Memoirs 2:17–42.
STRAHLER, A.N. 1964. Quantitative geomorphology of drain -age basins and channel networks. Section 4-2 in V.T.Chow, editor, Handbook of applied hydrology. Mc -Graw-Hill, New York, NY.
STRECKER, A.L., P.M. CAMPBELL, AND J.D. OLDEN. 2011.The aquarium trade as an invasion pathway in thePacific Northwest. Fisheries 36:74–85.
TABACHNICK, B.G., AND L.S. FIDELL. 1989. Using multi-variate statistics. 2nd edition. HarperCollins, NewYork, NY.
TER BRAAK, C.J.F., AND P.F.M. VERDONSCHOT. 1995. Canoni -cal correspondence analysis and related multivariatemethods in aquatic ecology. Aquatic Sciences 57:255–289.
THOMPSON, A.R., J.T. PETTY, AND G.D. GROSSMAN. 2001.Multi-scale effects of resource patchiness on forag-ing behavior and habitat use by longnose dace,Rhinichthys cataractae. Freshwater Biology46:145–160.
THUROW, R.F., D.C. LEE, AND B.E. RIEMAN. 1997. Distrib-ution and status of seven native salmonids in theinterior Columbia River Basin and portions of theKlamath River and Great basins. North AmericanJournal of Fisheries Management 17:1094–1110.
TORGERSEN, C.E., C.V. BAXTER, H.W. LI, AND B.A. MCIN-TOSH. 2006. Landscape influences on longitudinalpatterns of river fishes: spatially continuous analysisof fish–habitat relationships. American FisheriesSociety Symposium 48:473–492.
WARREN, M.L., H.W. ROBISON, S.T. ROSS, W.C. STARNES,B.M. BURR, S.J. WALSH, H.L. BART JR., R.C. CASH-NER, D.A. ETNIER, B.J. FREEMAN, ET AL. 2000. Di -versity, distribution, and conservation status of thenative freshwater fishes of the southern UnitedStates. Fisheries 25:7–31.
WYDOSKI, R.S., AND R.R. WHITNEY. 2003. Inland fishes ofWashington. 2nd edition. University of WashingtonPress, Seattle, WA.
Received 18 May 2012Accepted 16 November 2012
34 WESTERN NORTH AMERICAN NATURALIST [Volume 73