Embed Size (px)
Citation preview

, . 184: 71–76 (1998)
NON-RADIOACTIVE MEASUREMENT OFTELOMERASE ACTIVITY IN HUMAN BLADDERCANCER, BLADDER WASHINGS, AND IN URINE
1, 1*, ̈2, ̈ 2, 2 1
1Institute for Pathology, Universitätsklinikum Benjamin Franklin, Free University Berlin, 12200 Berlin, Germany2Department for Urology, Universitätsklinikum Benjamin Franklin, Free University Berlin, 12200 Berlin, Germany
SUMMARY
Early diagnosis is still the most important prerequisite for successful cancer treatment and this holds true for bladder cancer. Urinecytology is commonly used as a non-invasive screening procedure for the detection of bladder carcinoma, but this method islabour-intensive and often generates false-negative results. The ribonucleoprotein telomerase appears to be a promising new cancermarker, since its activity has been reported to correlate with indefinite growth. The aim of this study was to investigate whethertelomerase activity can be detected in bladder cancer and in corresponding bladder washings. For this purpose, a sensitivenon-radioactive TRAP (telomeric repeat amplification protocol) detection system was developed. With this technique, telomeraseactivity was found in 95 per cent of the carcinomas (n=20), in 70 per cent of the corresponding bladder washings, but in none of the urinesamples obtained from patients with bladder carcinoma. No telomerase activity was detectable in normal urothelium or in samples fromdysplastic urothelium. The data obtained from bladder washings show that superficial carcinoma cells released into the bladder stillharbour telomerase activity. The absence of telomerase activity in voided urine is thus most likely due to degradation or inactivationunder these conditions. The high rate of telomerase activity in bladder carcinoma indicates that the activation of telomerase is a commonstep in the tumourigenesis of bladder cancer. ? 1998 John Wiley & Sons, Ltd.
J. Pathol. 184: 71–76, 1998.
KEY WORDS—telomeres; telomerase activity; TRAP assay; GeneScan; bladder carcinoma; bladder washing; urine cytology
INTRODUCTION
Screening for the presence of bladder carcinoma bymeans of non-invasive procedures such as imaging tech-niques or cytology is not reliable and additional diag-nostic confirmation by invasive methods is required.This is especially true for well-differentiated bladdercarcinoma (G1), as only half of these lesions are detect-able by cytology. The introduction of flow cytometry orimmunocytochemistry has not significantly improvedthe diagnostic efficiency of voided urine cytology1–5 andthere is therefore a need for non-invasive tools enablingan early diagnosis of bladder carcinoma. Telomeraseactivity has been described as being constantly associ-ated with indefinite growth, a feature of malignant cells.The question therefore arose as to whether telomeraseactivity could serve as a suitable marker for malignancyin urine samples.Telomeres are specialized structures at the end of
eukaryotic chromosomes that appear to function inchromosomal protection, positioning, and replication.In humans, they consist mainly of up to a thousand ormore repeats of the 5*-TTAGGG-3* sequence.6–9 Witheach cell division, the telomeres lose about 5–35 repeats,due to the inability of DNA polymerase to replicatecompletely the linear DNA of the telomeres.10–15 This
shortening of the telomeres has been proposed to be themitotic clock by which cells count their divisions,16which implies that if telomeres become critically short,the cells undergo senescence.17 In contrast, malignantcells are able to perpetuate the length of the telomericsequences, which has led to the concept that mainten-ance of the telomeres is required for indefinite prolifer-ation and that indefinitely growing cells must possess anenzyme that enables the de novo synthesis of telomericDNA.This enzyme has been identified and designated as
telomerase. This ribonucleoprotein adds telomericrepeats onto the ends of chromosomes utilizing its RNAcomponent as a template.18–22 A radioactive method formeasuring telomerase activity was developed (telomericrepeat amplification protocol, TRAP) which enabled thedetection of telomerase activity in a wide variety oftumour-derived immortal cell lines and human malig-nancies.23 In contrast, this activity was not detectable innormal cells in vitro and normal somatic tissues in vivo,with only a few exceptions.24–28These findings prompted us to analyse cell pellets of
voided urine samples for telomerase activity frompatients with bladder carcinoma, in order to clarifywhether the activity of this enzyme could serve as areliable marker of malignancy in these samples. Unfor-tunately, our own and other pilot investigations did notreveal telomerase activity in all or nearly all voided urinesamples.29 To clarify the possible reasons for thesenegative measurements we investigated for telomerase
*Correspondence to: Michael Hummel, PhD, Institute of Pathol-ogy, Klinikum Benjamin Franklin, Free University Berlin, 12200Berlin, Germany.
CCC 0022–3417/98/010071–06 $17.50 Received 4 March 1997? 1998 John Wiley & Sons, Ltd. Accepted 26 August 1997

activity tissue samples and bladder washings frompatients with transitional carcinomas of different gradesand stages of malignancy, as well as samples frompatients with normal, inflamed, and dysplastic urothe-lium. The results obtained confirm that transitionalcarcinomas consistently upregulate telomerase activityand that superficial carcinoma cells maintain thisactivity. The non-detectability of telomerase activity inthe voided urine samples may therefore be due toenzyme inactivation under the conditions present in theurine.
MATERIALS AND METHODS
Cell lines, tissues and washings
All 40 cell lines were cultured in RPMI medium with10 per cent FCS. As a positive control, K562, amyeloma-derived cell line, was used. Tissue sampleswere removed by transurethral electrical biopsy, or inthe case of normal tissue samples (n=4) in the course ofoperation for other reasons. The tumours were classifiedaccording to WHO criteria for the histological typing ofbladder tumours.30 Bladder washings (n=26) wereobtained transurethrally by washing the bladder withphysiological saline directly before tumour resection.For the investigation of voided urine, 50 ml of urine wascollected by the patients after the morning-urine hadbeen passed.The suspensions were centrifuged at 10 000 g for 10
min and washed twice with phosphate-buffered saline.All samples were immediately processed (in less than30 min) to minimize RNA degradation or telomerasedenaturation.To investigate tissue samples for the presence of
telomerase activity, 15 ìm thick tissue sections wereproduced from fresh frozen tissue blocks and con-trolled by histological examination of 4–7 ìm sectionsdirectly above and beneath the extracted tissue portion.To evaluate bladder washings and urine samples,cell suspensions were counted, cytospun, and used forcytological examination.
ExtractionFifteen-micrometre thick tissue sections and cell pel-
lets from bladder washings were subjected to extraction.Depending on their size, the sections were homogenizedin 200–2000 ìl of lysis buffer [10 m Tris–HCl (pH 7·5),1 m MgCl2, 1 m EGTA, 0·1 m phenylmethylsulpho-nyl fluoride, 5 m â-mercaptoethanol, 0·5 per centCHAPS (Pierce, Oud-Beijerland, The Netherlands),10 per cent glycerol], incubated at 4)C for 30 min, andcentrifuged at 100 000 g for 30 min at 4)C. The cellpellets obtained from bladder washings and from nativeurine were resuspended in 300 ìl of lysis buffer andprocessed according to the above-mentioned protocol.The supernatants were removed, snap-frozen in liquidnitrogen, and stored at "80)C until used. Alkalinephosphatase was measured as an index of enzymeactivity in the lysate.
Telomeric repeat amplification protocol (TRAP)
The protein concentration of the supernatants wasdetermined according to Biuret using a commercialreagent (Sigma). An aliquot of 10 ìg of total protein wasincubated with 20 m Tris–HCl (pH 8·3), 1·5 mMgCl2, 63 m KCl, 0·0005 per cent Tween-20, 1 mEGTA, 50 ì deoxynucleoside triphosphates, 0·1 ìgof TS oligonucleotide (5*-AATCCGTCGAGCAGAGTT-3*), 1 ìg of T4g32 protein (Boehringer-Mannheim,Germany), and 0·1 mg/ml bovine serum albumin atroom temperature for 30 min for the generation oftelomeric repeats. To amplify the de novo synthesizedtelomeric repeats, 2·5 U of Taq DNA polymerase(Perkin-Elmer, Weiterstadt, Germany) and 0·1 ìg of flu-orescence (FAM)-labelled Cx oligonucleotide (3*-AATCCCATTCCCATTCCCATTCCC-5*; Perkin-Elmer,Weiterstadt, Germany) were added to a total reactionvolume of 50 ìl and subjected to 33 PCR cycles (94)C for30 s, 50)C for 30 s, and 72)C for 90 s) in a thermal cycler(TC9600; Perkin Elmer, Weiterstadt, Germany). Todetect Taq DNA polymerase inhibitors, 5 pmol of aninternal standard containing a base proportion of theCAT gene (GenBank Accession number M77788, nt2620–2779) flanked by the Ts and Cx primer sequences(total length 200 bp) was added to each sample.31
Analysis of the TRAP products
An aliquot (2 ìl) of the fluorescence-labelled amplifi-cation products was denatured by boiling for 2 min informamide and then separated on a sequencing gel anddetected on-line in an automated DNA sequencer(373A, ABI/Perkin Elmer, Weiterstadt, Germany). Thesize and intensity of the TRAP products were calculatedafter electrophoresis by application of GeneScansoftware (Perkin Elmer, Weiterstadt, Germany). Onlysamples with four or more amplificates in the appropri-ate size range were scored positive. All negative sampleswere positive for the internal standard. Telomeraseactivity-negative samples (10 ìg) were tested a secondtime by serial dilution between 50 ìg and 0·1 ìg protein.
RESULTS
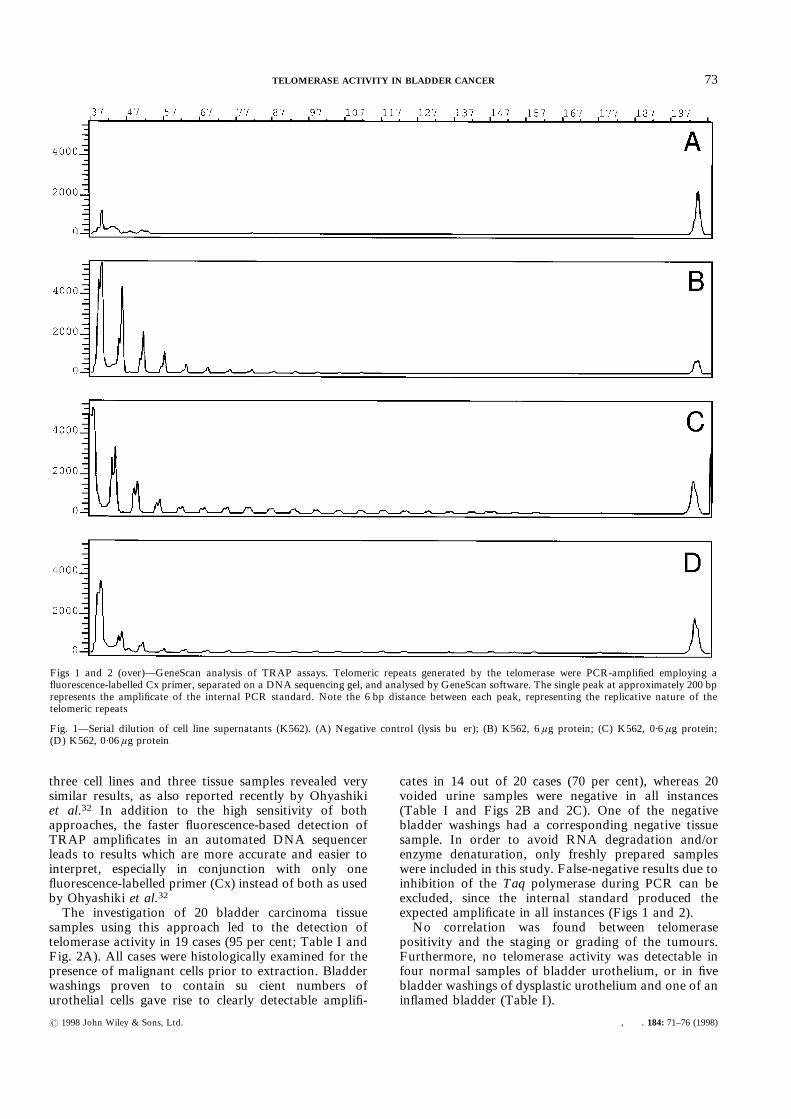
Application of the TRAP detection system to 39human and one monkey cell lines displayed telomeraseactivity in 95 per cent (38/40) of the cases, including fourcell lines derived from human transitional cell carcino-mas. This activity was illustrated as a chromatogramwith successive peaks of diminishing size representingthe amplificates produced with the TRAP assay (Fig. 1).However, two of the 40 cell lines investigated (MoT andMT-2, both HTLV-positive) did not show this pattern,indicating complete absence of telomerase activity. Non-malignant tissue samples were devoid of any specificfluorescence-labelled amplificates (Fig. 2D), as well asRNase-treated supernatants, clearly demonstrating thevalidity of our approach.The comparison of our fluorescence-based method
with the previously published radioactive procedure in
72 B. HEINE ET AL.
? 1998 John Wiley & Sons, Ltd. , . 184: 71–76 (1998)
three cell lines and three tissue samples revealed verysimilar results, as also reported recently by Ohyashikiet al.32 In addition to the high sensitivity of bothapproaches, the faster fluorescence-based detection ofTRAP amplificates in an automated DNA sequencerleads to results which are more accurate and easier tointerpret, especially in conjunction with only onefluorescence-labelled primer (Cx) instead of both as usedby Ohyashiki et al.32The investigation of 20 bladder carcinoma tissue
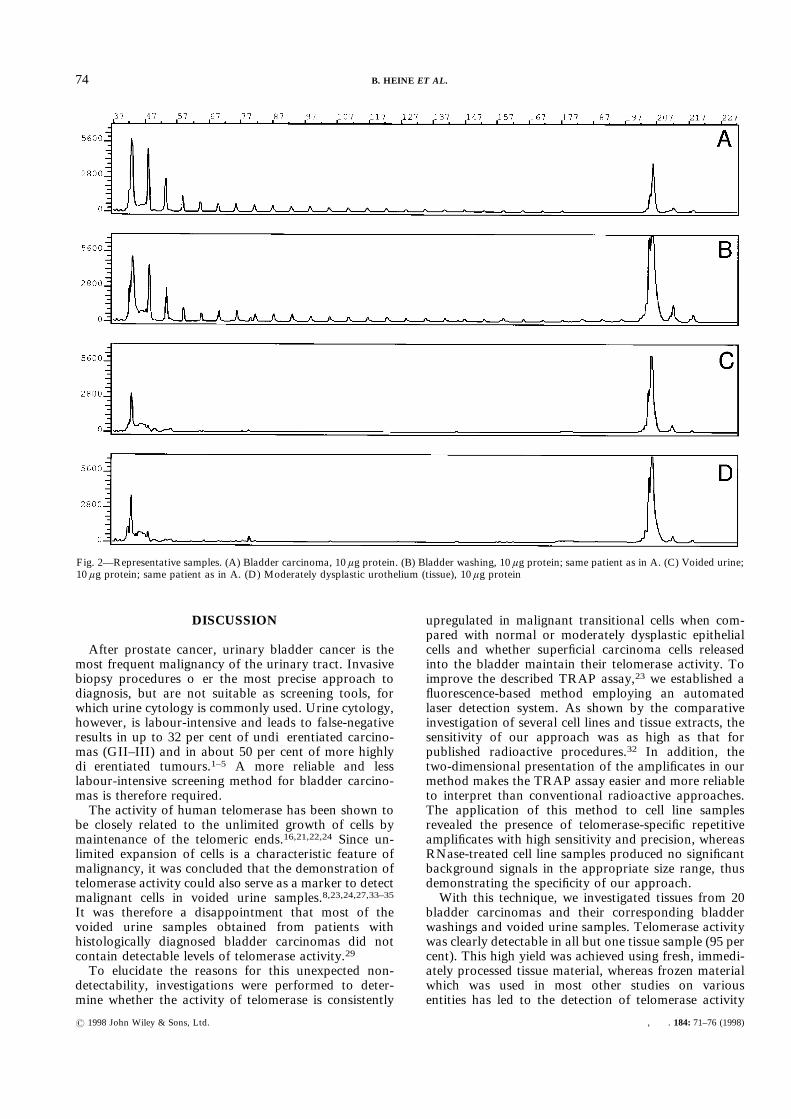
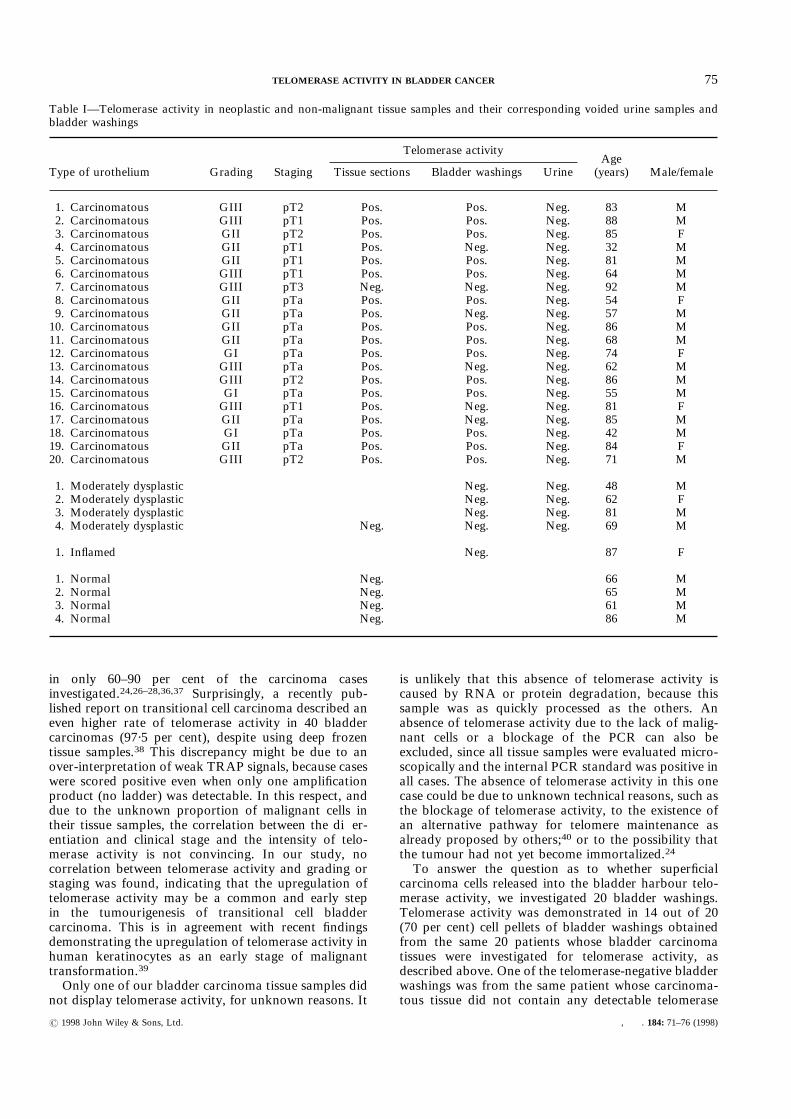
samples using this approach led to the detection oftelomerase activity in 19 cases (95 per cent; Table I andFig. 2A). All cases were histologically examined for thepresence of malignant cells prior to extraction. Bladderwashings proven to contain sufficient numbers ofurothelial cells gave rise to clearly detectable amplifi-
cates in 14 out of 20 cases (70 per cent), whereas 20voided urine samples were negative in all instances(Table I and Figs 2B and 2C). One of the negativebladder washings had a corresponding negative tissuesample. In order to avoid RNA degradation and/orenzyme denaturation, only freshly prepared sampleswere included in this study. False-negative results due toinhibition of the Taq polymerase during PCR can beexcluded, since the internal standard produced theexpected amplificate in all instances (Figs 1 and 2).No correlation was found between telomerase
positivity and the staging or grading of the tumours.Furthermore, no telomerase activity was detectable infour normal samples of bladder urothelium, or in fivebladder washings of dysplastic urothelium and one of aninflamed bladder (Table I).
Figs 1 and 2 (over)—GeneScan analysis of TRAP assays. Telomeric repeats generated by the telomerase were PCR-amplified employing afluorescence-labelled Cx primer, separated on a DNA sequencing gel, and analysed by GeneScan software. The single peak at approximately 200 bprepresents the amplificate of the internal PCR standard. Note the 6 bp distance between each peak, representing the replicative nature of thetelomeric repeats
Fig. 1—Serial dilution of cell line supernatants (K562). (A) Negative control (lysis buffer); (B) K562, 6 ìg protein; (C) K562, 0·6 ìg protein;(D) K562, 0·06 ìg protein
73TELOMERASE ACTIVITY IN BLADDER CANCER
? 1998 John Wiley & Sons, Ltd. , . 184: 71–76 (1998)
DISCUSSION
After prostate cancer, urinary bladder cancer is themost frequent malignancy of the urinary tract. Invasivebiopsy procedures offer the most precise approach todiagnosis, but are not suitable as screening tools, forwhich urine cytology is commonly used. Urine cytology,however, is labour-intensive and leads to false-negativeresults in up to 32 per cent of undifferentiated carcino-mas (GII–III) and in about 50 per cent of more highlydifferentiated tumours.1–5 A more reliable and lesslabour-intensive screening method for bladder carcino-mas is therefore required.The activity of human telomerase has been shown to
be closely related to the unlimited growth of cells bymaintenance of the telomeric ends.16,21,22,24 Since un-limited expansion of cells is a characteristic feature ofmalignancy, it was concluded that the demonstration oftelomerase activity could also serve as a marker to detectmalignant cells in voided urine samples.8,23,24,27,33–35It was therefore a disappointment that most of thevoided urine samples obtained from patients withhistologically diagnosed bladder carcinomas did notcontain detectable levels of telomerase activity.29To elucidate the reasons for this unexpected non-
detectability, investigations were performed to deter-mine whether the activity of telomerase is consistently
upregulated in malignant transitional cells when com-pared with normal or moderately dysplastic epithelialcells and whether superficial carcinoma cells releasedinto the bladder maintain their telomerase activity. Toimprove the described TRAP assay,23 we established afluorescence-based method employing an automatedlaser detection system. As shown by the comparativeinvestigation of several cell lines and tissue extracts, thesensitivity of our approach was as high as that forpublished radioactive procedures.32 In addition, thetwo-dimensional presentation of the amplificates in ourmethod makes the TRAP assay easier and more reliableto interpret than conventional radioactive approaches.The application of this method to cell line samplesrevealed the presence of telomerase-specific repetitiveamplificates with high sensitivity and precision, whereasRNase-treated cell line samples produced no significantbackground signals in the appropriate size range, thusdemonstrating the specificity of our approach.With this technique, we investigated tissues from 20
bladder carcinomas and their corresponding bladderwashings and voided urine samples. Telomerase activitywas clearly detectable in all but one tissue sample (95 percent). This high yield was achieved using fresh, immedi-ately processed tissue material, whereas frozen materialwhich was used in most other studies on variousentities has led to the detection of telomerase activity
Fig. 2—Representative samples. (A) Bladder carcinoma, 10 ìg protein. (B) Bladder washing, 10 ìg protein; same patient as in A. (C) Voided urine;10 ìg protein; same patient as in A. (D) Moderately dysplastic urothelium (tissue), 10 ìg protein
74 B. HEINE ET AL.
? 1998 John Wiley & Sons, Ltd. , . 184: 71–76 (1998)
in only 60–90 per cent of the carcinoma casesinvestigated.24,26–28,36,37 Surprisingly, a recently pub-lished report on transitional cell carcinoma described aneven higher rate of telomerase activity in 40 bladdercarcinomas (97·5 per cent), despite using deep frozentissue samples.38 This discrepancy might be due to anover-interpretation of weak TRAP signals, because caseswere scored positive even when only one amplificationproduct (no ladder) was detectable. In this respect, anddue to the unknown proportion of malignant cells intheir tissue samples, the correlation between the differ-entiation and clinical stage and the intensity of telo-merase activity is not convincing. In our study, nocorrelation between telomerase activity and grading orstaging was found, indicating that the upregulation oftelomerase activity may be a common and early stepin the tumourigenesis of transitional cell bladdercarcinoma. This is in agreement with recent findingsdemonstrating the upregulation of telomerase activity inhuman keratinocytes as an early stage of malignanttransformation.39Only one of our bladder carcinoma tissue samples did
not display telomerase activity, for unknown reasons. It
is unlikely that this absence of telomerase activity iscaused by RNA or protein degradation, because thissample was as quickly processed as the others. Anabsence of telomerase activity due to the lack of malig-nant cells or a blockage of the PCR can also beexcluded, since all tissue samples were evaluated micro-scopically and the internal PCR standard was positive inall cases. The absence of telomerase activity in this onecase could be due to unknown technical reasons, such asthe blockage of telomerase activity, to the existence ofan alternative pathway for telomere maintenance asalready proposed by others;40 or to the possibility thatthe tumour had not yet become immortalized.24To answer the question as to whether superficial
carcinoma cells released into the bladder harbour telo-merase activity, we investigated 20 bladder washings.Telomerase activity was demonstrated in 14 out of 20(70 per cent) cell pellets of bladder washings obtainedfrom the same 20 patients whose bladder carcinomatissues were investigated for telomerase activity, asdescribed above. One of the telomerase-negative bladderwashings was from the same patient whose carcinoma-tous tissue did not contain any detectable telomerase
Table I—Telomerase activity in neoplastic and non-malignant tissue samples and their corresponding voided urine samples andbladder washings
Type of urothelium Grading Staging
Telomerase activityAge(years) Male/femaleTissue sections Bladder washings Urine
1. Carcinomatous GIII pT2 Pos. Pos. Neg. 83 M2. Carcinomatous GIII pT1 Pos. Pos. Neg. 88 M3. Carcinomatous GII pT2 Pos. Pos. Neg. 85 F4. Carcinomatous GII pT1 Pos. Neg. Neg. 32 M5. Carcinomatous GII pT1 Pos. Pos. Neg. 81 M6. Carcinomatous GIII pT1 Pos. Pos. Neg. 64 M7. Carcinomatous GIII pT3 Neg. Neg. Neg. 92 M8. Carcinomatous GII pTa Pos. Pos. Neg. 54 F9. Carcinomatous GII pTa Pos. Neg. Neg. 57 M10. Carcinomatous GII pTa Pos. Pos. Neg. 86 M11. Carcinomatous GII pTa Pos. Pos. Neg. 68 M12. Carcinomatous GI pTa Pos. Pos. Neg. 74 F13. Carcinomatous GIII pTa Pos. Neg. Neg. 62 M14. Carcinomatous GIII pT2 Pos. Pos. Neg. 86 M15. Carcinomatous GI pTa Pos. Pos. Neg. 55 M16. Carcinomatous GIII pT1 Pos. Neg. Neg. 81 F17. Carcinomatous GII pTa Pos. Neg. Neg. 85 M18. Carcinomatous GI pTa Pos. Pos. Neg. 42 M19. Carcinomatous GII pTa Pos. Pos. Neg. 84 F20. Carcinomatous GIII pT2 Pos. Pos. Neg. 71 M
1. Moderately dysplastic Neg. Neg. 48 M2. Moderately dysplastic Neg. Neg. 62 F3. Moderately dysplastic Neg. Neg. 81 M4. Moderately dysplastic Neg. Neg. Neg. 69 M
1. Inflamed Neg. 87 F
1. Normal Neg. 66 M2. Normal Neg. 65 M3. Normal Neg. 61 M4. Normal Neg. 86 M
75TELOMERASE ACTIVITY IN BLADDER CANCER
? 1998 John Wiley & Sons, Ltd. , . 184: 71–76 (1998)

activity, whereas the other telomerase-negative washingsstemmed from patients with telomerase-positive carcino-mas. The absence of telomerase activity in these fivebladder washings may reflect the problematic handlingand processing of washings and body fluids. We assumethat in these five samples the telomerase RNA com-ponent was degraded or telomerase was inactivated forunknown reasons, even though they were processedunder optimized conditions.In contrast to the results obtained in samples containingmalignant cells, no telomerase activity was demonstrablein normal or moderately dysplastic tissue samples orwashings. The absence of telomerase activity in normaland dysplastic cells is largely in line with publisheddata.24,26,34 Only a few exceptions to this rule areconvincingly described. Such as germ cells, peripheral Bcells, and germinal centre B cells. The description of onetelomerase-positive dysplastic tissue sample by Lin et al.might be a rare exception or an over-interpretation offaint signals. Our results obtained in bladder carcinomaand bladder washings support the view that the presenceof telomerase activity is restricted to cells which requirethis activity for their unlimited growth.To conclude, bladder carcinomas are regularly associ-ated with high levels of telomerase activity, independentof their grade and stage, whereas normal or dysplasticcells fail to show detectable activity. Superficial malig-nant cells released into the bladder also harbour elevatedlevels of telomerase activity. The detection of telomeraseactivity in bladder washings and its absence in urinesamples thus justify the conclusion that telomerase isinactivated, or that its RNA component is degraded inthe urine by unknown mechanisms. New studies areneeded to clarify these mechanisms, with the aim offinding procedures to prevent this negative outcome.Such studies are currently in progress.
ACKNOWLEDGEMENTS
We thank Dr Thomas Emrich (Fa. BoehringerMannheim, Penzberg, Germany) for providing us withthe internal standard. We are indebted to Hans-HenningMüller and Birgit Schaak for their excellent technicalassistance and to Janet Yates for her critical editorialcontribution. This work was supported by the DeutscheKrebshilfe; grant number 70-1923-St3.
REFERENCES
1. Esposti PI, Moberger G, Zajicek J. The cytologic diagnosis of transitionalcell tumors of the urinary bladder and its histologic basis. Acta Cytol(Baltimore) 1970; 14: 145–155.
2. Koss LG, Deitch D, Ramanathan R, Sherman AB. Diagnostic value ofcytology in voided urine. Acta Cytol (Baltimore) 1985; 29: 810–816.
3. Wawroschek F, Rathert P. Urine cytology. Urologe A 1995; 34: 69–75.
4. Mollhof S, Rathert P. What is the value of urine cytology in after-care ofpatients with urothelial cancers? Urologe A 1992; 31: 203–205.
5. National Bladder Cancer Cooperative Group A. Cytology and histopathol-ogy of bladder cancer cases in a prospective longitudinal study. Cancer Res1977; 37: 2911–2915.
6. Allshire RC, Gosden JR, Cross SH, et al. Telomeric repeat from T.thermophila cross hybridizes with human telomeres. Nature 1988; 332:656–659.
7. Moyzis RK, Buckingham JM, Cram IS, et al. A highly conserved repetitiveDNA sequence, (TTAGGG)n, present at the telomeres of human chromo-somes. Proc Natl Acad Sci USA 1988; 85: 6622–6626.
8. Zakian V. Telomeres. Beginning to understand the end. Science 1995; 270:1601–1607.
9. Chong L, Steensel B, Broccoli D, et al. A human telomeric protein. Science1995; 270: 1663–1667.
10. Harley CB, Futcher AB, Greider CW. Telomeres shorten during aging ofhuman fibroblasts. Nature 1990; 345: 458–460.
11. Lindsey J, McGill NI, Lindsey LA, Green DK, Kooke HJ. In vivo loss oftelomeric repeats with age in humans. Mutat Res 1991; 256: 45–48.
12. Allsopp RC, Vaziri H, Patterson C, et al. Telomere length predictsreplicative capacity of human fibroblasts. Proc Natl Acad Sci USA 1992; 89:10114–10118.
13. Allsopp RC, Chang E, Kashefi-Aazam M, et al. Telomere shortening isassociated with cell division in vitro and in vivo. Exp Cell Res 1995; 220:194–200.
14. Rogan E, Bryan T, Hukku B, et al. Alterations in p53 and p16INK4expression and telomere length during spontaneous immortalization ofLi–Fraumeni syndrome fibroblasts. Mol Cell Biol 1995; 15: 4745–4753.
15. Watson JD. Origin of concatemeric T7 DNA. Nature New Biol 1972; 239:197–201.
16. Harley C. Telomere loss: mitotic clock or genetic time bomb. Mutat Res1991; 256: 271–282.
17. Counter CM, Botelho FM, Wang P, Harley CB, Bacchetti S. Stabilizationof short telomeres and telomerase activity accompany immortalization ofEpstein–Barr virus-transformed human B lymphocytes. J Virol 1994; 68:3410–3414.
18. Morin G. The human telomere terminal transferase enzyme is a ribonucleo-protein that synthesizes TTAGGG repeats. Cell 1989; 59: 521–529.
19. Greider CW, Blackburn EH. Identification of a specific telomere terminaltransferase activity in tetrahymena extracts. Cell 1985; 43: 405–413.
20. Feng J, Funk W, Wang S, et al. The RNA component of human telomerase.Science 1995; 269: 1236–1241.
21. Shay J, Pereira-Smith OM, Wright WE. A role for both RB and p53 in theregulation of human cellular senescence. Exp Cell Res 1991; 196: 33–39.
22. Rhyu MS. Telomeres, telomerase, and immortality. J Natl Cancer Inst 1995;87: 884–894.
23. Kim NW, Piatyszek MA, Prowse KR, et al. Specific association of humantelomerase activity with immortal cells and cancer. Science 1994; 266:2011–2015.
24. Hiyama E, Hiyama K, Yokohama T, Matsuura Y, Piatyszek M, Shay JW.Correlating telomerase activity levels with human neuroblastoma outcomes.Nature Med 1995; 1: 249–255.
25. Counter CM, Gupta J, Harley CB, Leber B, Bacchetti S. Telomeraseactivity in normal leukocytes and in hematologic malignancies. Blood 1995;85: 2315–2320.
26. Tahara H, Nakanishi T, Kitamoto M, et al. Telomerase activity in humanliver tissues: comparison between chronic liver disease and hepatocellularcarcinomas. Cancer Res 1995; 55: 2734–2736.
27. Chadeneau C, Hay K, Hirte H, Gallinger S, Bacchetti S. Telomerase activityassociated with acquisition of malignancy in human colorectal cancer.Cancer Res 1995; 55: 2533–2536.
28. Counter C, Hirte H, Bacchetti S, Harley C. Telomerase activity in humanovarian carcinoma. Proc Natl Acad Sci USA 1994; 91: 2900–2904.
29. Oral presentation by Professor J. Shay at the AACR Meeting ‘NovelStrategies Against Resistant Cancers’, Fort Myers, Florida, November 1995and own unpublished data.
30. Mostofi F, Sobin L, Torloni H. International Histological Classification ofTumours (10): Histological Typing of Urinary Bladder Tumours. Geneva:World Health Organization, 1973.
31. Wright W, Shay J, Piatyszek M. Modifications of a telomeric repeatamplification protocol (TRAP) result in increased reliability, linearity andsensitivity. Nucleic Acids Res 1995; 23: 3794–3795.
32. Ohyashiki J, Ohyashiki K, Sano T, Toyama K. Non-radioisotopic andsemi-quantitative procedure for terminal repeat amplification protocol. JpnJ Cancer Res 1996; 87: 329–331.
33. Nilsson P, Mehle C, Remes K, Roos G. Telomerase activity in vivo inhuman malignant hematopoietic cells. Oncogene 1994; 9: 3043–3048.
34. Norrback KF, Dahlenborg K, Carlsson R, Roos G. Telomerase activationin normal B lymphocytes and non-Hodgkin’s lymphomas. Blood 1996; 88:222–229.
35. Morin G. Is telomerase a universal cancer target? J Natl Cancer Inst 1995;87: 859–861.
36. Hiyama K, Hiyama E, Ishioka S, et al. Telomerase activity in small-cell andnon-small-cell lung cancers. J Natl Cancer Inst 1995; 87: 895–902.
37. Ohyashiki K, Ohyashiki J, Iwama H, Hayashi S, Shay J, Toyama K.Telomerase reactivation in leukemia cells. Int J Oncol 1996; 8: 417–421.
38. Lin Y, Miyamato H, Fujinami K, et al. Telomerase activity in humanbladder cancer. Clin Cancer Res 1996; 2: 929–932.
39. Klingelhutz A, Foster S, McDougall J. Telomerase activation by the E6gene product of human papillomavirus type 16. Nature 1996; 380: 79–82.
40. Bryan T, Englezou A, Gupta J, Bacchetti S, Reddel R. Telomere elongationin immortal human cells without detectable telomerase activity. EMBO J1995; 14: 4240–4248.
76 B. HEINE ET AL.
? 1998 John Wiley & Sons, Ltd. , . 184: 71–76 (1998)