Embed Size (px)
Citation preview

Nitrogen Incorporation Into Decomposing Litter of Spartina alternifloraAuthor(s): David S. White and Brian L. HowesSource: Limnology and Oceanography, Vol. 39, No. 1 (Jan., 1994), pp. 133-140Published by: American Society of Limnology and OceanographyStable URL: http://www.jstor.org/stable/2838369 .
Accessed: 16/06/2014 01:55
Your use of the JSTOR archive indicates your acceptance of the Terms & Conditions of Use, available at .http://www.jstor.org/page/info/about/policies/terms.jsp
.JSTOR is a not-for-profit service that helps scholars, researchers, and students discover, use, and build upon a wide range ofcontent in a trusted digital archive. We use information technology and tools to increase productivity and facilitate new formsof scholarship. For more information about JSTOR, please contact [email protected].
.
American Society of Limnology and Oceanography is collaborating with JSTOR to digitize, preserve andextend access to Limnology and Oceanography.
http://www.jstor.org
This content downloaded from 188.72.126.181 on Mon, 16 Jun 2014 01:55:52 AMAll use subject to JSTOR Terms and Conditions

Notes 133
GARCiA-Novo, F. 1979. The ecology of vegetation of the dunes in Doinana National Park (southwest Spain), p. 571-592. In R. L. Jefferies and A. J. Davy [eds.], Ecological processes in coastal environments. Black- well.
LANGERAAR, W. 1966. Sand waves in the North Sea. Hydrogr. Newslett. 1: 243-246.
LANGHORNE, D. N. 1982. A study of the dynamics of a marine sand wave. Sedimentology 29: 571-594.
MALDONADO, A. 1972. El Delta del Ebro. Estudio se- dimentol6gico y estratigrAfico. Univ. Barcelona Bol. Estratigaf. 1.
PATRIQUIN, D. G. 1973. Estimation of growth rate, pro- duction and age of the marine angiosperm Thalassia testudinum K6nig. Carib. J. Sci. 13: 111-123.
* 1975. Migration of blowouts in seagrass beds at Barbados and Carriacou, West Indies, and its ecolog- ical and geological implications. Aquat. Bot. 1: 163- 189.
SAND-JENSEN, K. 1975. Biomass, net production and growth dynamics in an eelgrass (Zostera marina L.) population in Vellerup Vig, Denmark. Ophelia 14: 185-201.
TWICHELL, D. C. 1983. Sand-wave movement on little Georges Band. U.S. Geol. Surv. Open-File Rep. 83- 746, p. 1-20.
Submitted: 4 March 1992 Accepted: 27 April 1993 Amended: 9 June 1993
Limnol. Oceanogr., 39(1), 1994, 133-140 ? 1994, by the American Society of Limnology and Oceanography, Inc.
Nitrogen incorporation into decomposing litter of Spartina alterniflora
Abstract-Litter N dynamics were followed during decay in aboveground biomass of Spartina alterni- flora (Loisel.) in litter bags and a cohort of intact culms. Initial enrichment with 'IN allowed parti- tioning of the total litter N pool into originally in- corporated N during plant growth vs. N incorporated from external sources during decay. Both the original '5N and total N pools declined through fall but sta- bilized in winter. As temperatures increased in spring, the original IIN pool again declined, but the total N pool remained stable due to significant incorporation of N from external sources. By the midpoint in de- composition, 50-65% of the total detrital N pool was from external sources and at least part of the N in- corporation was biologically mediated. The apparent incorporation of external N based on l sN techniques in this study is greater than previous estimates based on net N accumulation. We conclude that external N incorporation into decaying S. alterniflora is a potentially much more important mechanism of short-term N retention in salt-marsh ecosystems than previously thought.
Only a small fraction of the aboveground primary productivity of Spartina alterniflora
Acknowledgments We thank W. Sylvester and T. Haystead for C, N, and
'5N analyses; Mrs. D. Gifford, Mr. A. Diddel and Salt Pond Sanctuaries for use of their salt marsh property; and J. Teal, T. Millham, D. Schlezinger, and two anonymous reviewers for comments on an earlier version of the manu- script.
This project was supported by NSF grants BSR 85- 07356 and BSR 87-17701 and by a grant from the Island Foundation, Marion, Massachusetts.
Contribution 8272 of the Woods Hole Oceanographic Institution.
(Loisel.) salt marshes along the Atlantic and Gulf coasts is directly consumed by herbivores (Teal 1962). The remainder either decays in place or enters detrital food chains in the marsh or in adjacent coastal waters after export.
The quality of salt-marsh detritus as a food source for consumers depends to a significant degree on the availability of N for assimilation and growth (cf. Tenore 1984). Nitrogen avail- ability in turn depends on the processes which alter the form and amount of N during decom- position. Many types of refractory detritus de- cay slowly, becoming enriched in N as they "age" (Tenore 1984). In subtidal systems mi- crobial N enrichment of fecal pellets and or- ganic-rich sediments improves detrital quality as a food source for consumers (Newell 1965).
In contrast, microbial biomass makes a rel- atively small contribution to litter N enrich- ment in decaying S. alterniflora. Many field and laboratory studies have shown increases in both concentration (%) and absolute amounts of N over time in decaying Spartina litter (Marinucci 1982; Valiela et al. 1985; Newell et al. 1989). However, estimates, using a variety of methods, suggest that a maximum of only 20-25% of this N increase is due to microbial biomass (Lee et al. 1980; Hicks et al. 1991). Alternatively, microbial exudates which remain physically attached to detrital material after cell death may make a significant contribution to litter N enrichment and may
This content downloaded from 188.72.126.181 on Mon, 16 Jun 2014 01:55:52 AMAll use subject to JSTOR Terms and Conditions

134 Notes
be an important source of available N to con- sumers (Hobbie and Lee 1980).
In addition to direct enrichment from mi- crobial biomass or exudates, a significant por- tion of the increase in the absolute amount of detrital N during decomposition may come from external sources which become chemi- cally incorporated, via condensation reactions, into refractory portions (lignins and bound phenolics) of the detritus as a result of micro- bial attack and subsequent breakdown of the more labile plant components (Rice 1982; Wil- son et al. 1986). Odum et al. (1979) estimated that up to 30% of the total N in aged detritus may be due to such refractory materials and suggested that some of these materials may be of fungal origin.
Although the dynamics of nitrogen loss and accumulation in decaying Spartina litter have been extensively studied and both microbial and nonmicrobial N fractions have been es- timated, little direct evidence exists regarding the amount and timing of loss of original in- ternal plant N vs. incorporation of external N to the total detrital N pool during decompo- sition. Relative (%) increase in N during de- trital aging is by itself not sufficient to prove external N incorporation because N concen- tration can increase without a simultaneous increase in absolute amount (Tenore 1984), for instance by the preferential loss of carbon as CO2 in microbial respiration. In addition, since loss of original plant N may continue even as new external N is being incorporated, calcu- lated amounts of N incorporation based on temporal changes in the total detrital N pool serve only as minimal estimates.
The present study focused directly on the basic process of nitrogen incorporation into decaying Spartina litter using the stable iso- tope 15N to partition the total detrital N pool into original plant N incorporated during growth vs. external N incorporated during de- cay. 5N was used in two different approaches for studying the N dynamics of Spartina de- composition. First, 15N, total N, and C pools were monitored in '5N-enriched litter incu- bated in litter bags in the field for an 18-month period. Second, the same pools were moni- tored in a cohort of intact l 5N-enriched above- ground litter followed over a period of 1 yr under field conditions after 1 5NH4 was injected into the marsh sediments. Problems associated
with litter-bag studies are well documented (Valiela et al. 1985), and recent work (Newell and Fallon 1989) has shown important differ- ences in decomposition dynamics of S. alter- niflora in litter bags compared to that left intact in the field. Both litter bags and intact plants were used in the present study to better gen- eralize the results.
All work was conducted in Great Sippewis- sett Salt Marsh (Cape Cod, Massachusetts) from 1986-1989. The sites used for the cohort and litter-bag studies were colonized by nearly pure stands of short S. alterniflora (20-30 cm high). Litter incubations began in late August and mid-October.
'5N-enriched litter for the litter-bag incu- bations was produced by growing S. alterni- flora in the laboratory in tanks maintained at field water-table conditions. Plants were grown on a 16: 8 diel cycle, watered daily, and sprayed regularly to maintain a saturated substrate and to wash off accumulated salts. Each tank was injected with 20 mmol of 99 atoms%'5N (as ammonium sulfate, Cambridge Isotope Labs), the equivalent of 5.37 g N m-2. The above- ground plant N pool was enriched with 15N to 26.23 atoms%. When plants started to senesce, the aboveground biomass was harvested and air-dried at 20?C (Lee et al. 1980). Subsamples were oven-dried to make air-dried: oven-dried weight conversions of all material before being placed in the field.
Randomly selected subsamples of -7 g air dry wt of intact culms were placed in litter bags made of nylon mesh (2 x 4-mm opening) (Va- liela et al. 1985). Bags were placed in the field in mid-October (time 0) and anchored to the marsh surface. Duplicate bags were destruc- tively sampled at 1, 5, 9, and 18 months after placement.
'5N-enriched field plants were produced by injecting ammonium sulfate (99 atoms% 15N) in four installments over a 1-week period in May into marsh sediments surrounded by thin- walled (3 mm) fiber-glass tubes (27-cm i.d. x 45 cm long) driven directly into the marsh until they were flush with the surface. Water- table movement and geochemical parameters are not changed significantly by the introduc- tion of this size of lysimeter into the sediment (Dacey and Howes 1984). Peak biomass, mea- sured by harvest in August of the current year's biomass, was not affected by either the pres-
This content downloaded from 188.72.126.181 on Mon, 16 Jun 2014 01:55:52 AMAll use subject to JSTOR Terms and Conditions

Notes 135
ence of the lysimeters or injection of the tracer. There were no significant differences between peak biomass estimates in the tubing (380 ? 66 g m-2) vs. control areas (370?33 g m-2). The total amount of 15N injected was 3 mmol N per lysimeter, equivalent to 0.8 g N m-2. A single year's aboveground growth (cohort) was followed for 1 yr beginning with senescence. Duplicate sets of the same cohort of above- ground litter (0.06 m2 area) were harvested in August, December, April, and the following August (3, 7, 1 1, and 15 months respectively after injections were made). For purposes of data reporting, time 0 is late August, the date of the first harvest of senescent aboveground material. The cohort at time 0 was found to be enriched in '5N to 3.63 atoms%. All above- ground biomass was clipped and carefully sort- ed into live, standing dead, and the previous year's dead (litter which was still attached but clearly decomposed and lying on the marsh surface). The time 0 cohort (August) included only the current year's standing culms (dead and alive). The following August, this same cohort became the previous year's dead ma- terial.
Samples from both the litter-bag and cohort studies were gently rinsed in seawater over a 0.5-mm-mesh screen, oven-dried at 60?C, and milled through a 40-mesh screen for analysis. Analyses for %C, %N, and atoms% 15N on all litter were performed on a Carlo-Erba model NA1 500 elemental analyzer interfaced with a Europa Scientific tracer mass stable isotope an- alyzer with a precision of 0.001 atoms% 15N (T. Haystead pers. comm.). Results from all analyses were corrected to percent original dry weight to trace changes in mass over time. Means and standard deviations of each dupli- cate set were determined.
Decay rate constants (k) for decomposing litter were determined by a simple first-order decay function X, = Xoexp(-kt), where X, is the amount of material remaining at time t, and X0 is the initial amount. The linear trans- formation of the equation, ln X, = - kt + ln XO, allows for a least-squares analysis of k (Marinucci and Bartha 1982).
Mean monthly air temperatures were cal- culated from daily records at a meteorological station at Long Pond (Falmouth, Massachu- setts).
In general, the patterns of decay and nitro-
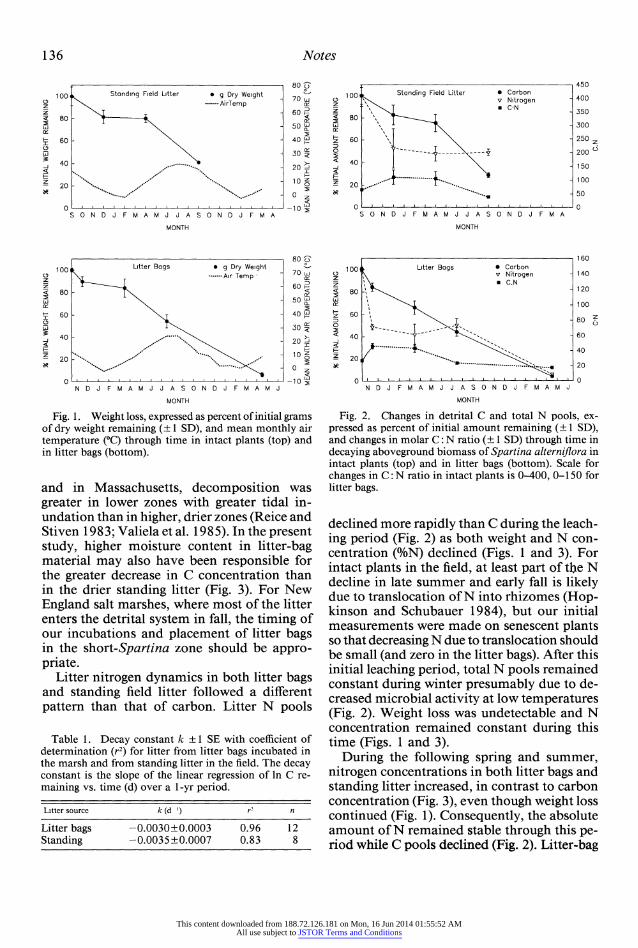
gen dynamics in the present study are consis- tent with other studies of north temperate salt marshes (Lee et al. 1980; Marinucci 1982; Val- iela et al. 1985). There was a rapid weight loss of 10-20% during the initial interval (30-100 d) in fall (September-December) presumably due primarily to leaching (Fig. 1). Loss of or- ganic matter (as total C) followed a similar pattern (Fig. 2).
After leaching, the primary alterations to the litter mass are through microbial respiration, making changes in mass strongly dependent on temperature during the decomposer phase (Valiela et al. 1985). During the next 120 d (December-March) loss of organic matter de- creased in both standing litter and litter-bag material (Fig. 2).
Loss of organic matter increased again over the next 120-140 d during the following spring and summer (March-August) from active de- composition of the remaining material at warmer temperatures (Fig. 2). The study of standing field litter ended at 1 yr because we could no longer confidently follow the cohort of aboveground material. By 540 d (April), litter-bag material had lost >95% of its orig- inal organic matter content (Fig. 2).
The decay constants, based on the loss of total organic C (Fig. 2), were similar for litter- bag material and for standing litter (Table 1), although monitoring of decomposition in standing litter began during the warmer tem- peratures of late August and the litter-bag study began in mid-October. We expected a higher rate of organic matter loss in the standing litter than in litter bags due to differences in initial incubation temperatures. In a New Jersey study, litter bags containing S. alterniflora placed in the low marsh in summer exhibited very rapid decay with >90% of initial litter weight lost after 120 d (Marinucci 1982). Since, in the present study, both litter-bag material and standing litter decayed at similar rates, factors other than temperature-possibly dif- ferences in moisture content-may have had a significant effect on decomposition rates. Halupa (1993) found that rates of litter decom- position in a New Jersey salt marsh were di- rectly related to moisture content and that standing field litter was drier than litter incu- bated in bags on the marsh surface. Similarly, in studies where litter was placed in zones with different flooding regimes in North Carolina
This content downloaded from 188.72.126.181 on Mon, 16 Jun 2014 01:55:52 AMAll use subject to JSTOR Terms and Conditions
136 Notes
80 i3 100 Standing Field Litter * g Dry Weight 70
. 8 0... AirTemp 70
i Z 60 M ~ 80
50
60 40 uJ
H 60 < 30 < 40 ..
- 20 ' ***.j..? z0
0 -10. SON D J F M A M J J A S O N D J F M A
MONTH
80 3 in ol Litter Begs o g Dry Weight
greater in lower zones with.. Airrempe tia n
Z D z 60H
80~~~~~~~~~~~~~~5 40 u9
Si ~~~~~~~~~~~~~~~30 20~~~~~~~~~~~~0
10 z
Stve .93 .ail et a. 195) In th rsn
NOD J F M A M J J A SON 0 J F M A MJ
MONTH
Fig. 1. Weight loss, expressed as percent of initial grams of dry weight remaining (? 1 SD), and mean monthly air temperature (r through time in intact plants (top) and in litter bags (bottom).
and in Massachusetts, decomposition was greater in lower zones with greater tidal in- undation than in higher, drier zones (Reice and Stiven 1 83; Valiela et al. 1985). In the present study, higher moisture content in litter-bag material may also have been responsible for the greater decrease in C concentration than in the drier standing litter (Fig. 3). For New England salt marshes, where most of the litter enters the detrital system in fall, the timing of our incubations and placement of litter bags in the short-Spartina zone should be appro- priate.
Litter nitrogen dynamics in both litter bags and standing field litter followed a different pattern than that of carbon. Litter N pools
Table 1. Decay constant k ?1I SE with coefficient of determination (r2) for litter from litter bags incubated in the marsh and from standing litter in the field. The decay constant is the slope of the linear regression of In C re- maining vs. time (d) over a 1 -yr period.
Litter source k (d I) r n
Litter bags -0.0030?0.0003 0.96 12 Standing -0.0035?0.0007 0.83 8
450 Standing Field Litter v Carbon
lZ o v Nitrogen 400
z O 350 80 300
z 60 250 z O 2000
<40 j _ T 150
.......... 100
lr ~~~~~~~~~~~~50 0 . . . . . . . .0
S O N D J F M A M J J A S O N D J F M A
MONTH
160
100i Litter Bags * Carbon ID X V Nitrogen - 140
z *C.N
ZE. - 120
< 80
NJ D J F MA M J A S O D J FM A M 100
z 60 D IT 8
< 40 60 -j ? .........4 z 20,
20 0 0
ND J FMA M J JA SON DJ FM AM J
MONTH
Fig. 2. Changes in detrital C and total N pools, ex- pressed as percent of initial amount remaining (? 1 SD), and changes in molar C: N ratio (? 1 SD) through time in decaying aboveground biomass of Spartina alterniflora in intact plants (top) and in litter bags (bottom). Scale for changes in C: N ratio in intact plants is 0-400, 0-150 for litter bags.
declined more rapidly than C during the leach- ing period (Fig. 2) as both weight and N con- centration (%N) declined (Figs. 1 and 3). For intact plants in the field, at least part of the N decline in late summer and early fall is likely due to translocation of N into rhizomes (Hop- kinson and Schubauer 1984), but our initial measurements were made on senescent plants so that decreasing N due to translocation should be small (and zero in the litter bags). After this initial leaching period, total N pools remained constant during winter presumably due to de- creased microbial activity at low temperatures (Fig. 2). Weight loss was undetectable and N concentration remained constant during this time (Figs. 1 and 3).
During the following spring and summer, nitrogen concentrations in both litter bags and standing litter increased, in contrast to carbon concentration (Fig. 3), even though weight loss continued (Fig. 1). Consequently, the absolute amount of N remained stable through this pe- riod while C pools declined (Fig. 2). Litter-bag
This content downloaded from 188.72.126.181 on Mon, 16 Jun 2014 01:55:52 AMAll use subject to JSTOR Terms and Conditions
Notes 137
50 2 0 Standing Field Litter * % Carbon
, , + A % Nitrogen 1 8 40' 16
14 z ~~~~~~~~~~~~z
0o 30 - 12 m ~~~~~~~~~~~~~0 cr_ ~~~~~~~~10cr
S OO8 AMJJASO
520 0
06
10 * - 0.4 - 0.2
0 . . . . . . . . . . . , , 0
C 00N D J F M A M J J A S O N D J F M A
MONTH
Litter Bags 1 8
1.4
%C is 0-0an Nis02
20 declined again after 270 d (July) dueto8 Z
0 6
crobial % Carbon d 0.4
a % Nitrogen - 0.2
MONTH
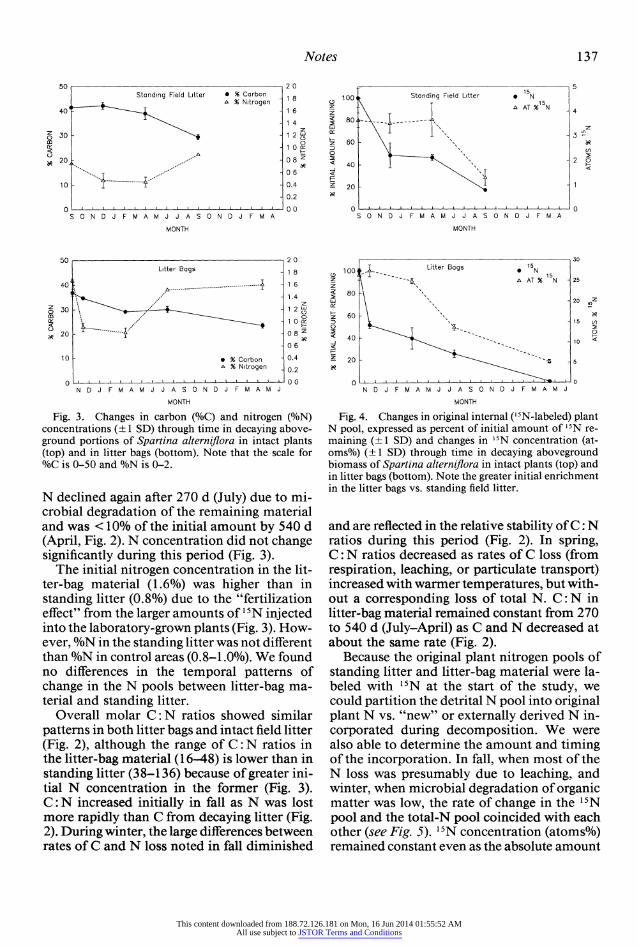
Fig. 3. Changes in carbon (%C) and nitrogen (%N) concentrations (?1 SD) through time in decaying above- ground portions of Spartina alterniflora in intact plants (top) and in litter bags (bottom). Note that the scale for %C is 0-50 and %N is 0-2.
N declined again after 270 d (July) due to mi- crobial degradation of the remaining material and was <0 % of the initial amount by 540 d (April, Fig. 2). N concentration did not change significantly during this period (Fig. 3).
The initial nitrogen concentration in the lit- ter-bag material (1.6%) was higher than in standing litter (0.8%) due to the "fertilization effect" from the larger amounts of 15 N injected into the laboratory-grown plants (Fig. 3). How- ever, %N in the standing litter was not different than %N in control areas (0.8-1.0%). We found no differences in the temporal patterns of change in the N pools between litter-bag ma- terial and standing litter.
Overall molar C: N ratios showed similar patterns in both litter bags and intact field litter (Fig. 2), although the range of C: N ratios in the litter-bag material (1 6-4 8) is lower than in standing litter (38-136) because of greater ini- tial N concentration in the former (Fig. 3). C: N increased initially in fall as N was lost more rapidly than C from decaying litter (Fig. 2). During winter, the large differences between rates of C and N loss noted in fall diminished
5
100 Standing Field Litter A N
0 \ _
z .at~ ~ - AT % N -2
<- 80 \ " 20 < 40
_ "I~~~~~~~~~~~~~ z 2011
C 0 SON D J F M A M J J A S O N D J F M A
MONTH
30
15
100 l itter Bags (bto) Not 1h5rae ntilercmn
Z AT % N -i25
< 80 Lu ~~~~~~~~~~~~~~~~~~~-20Z
z 60 1
0
Fig.er4.aChangesrinogal rmintedcnsal t ( frombed plan
mi i40 (? 1uSD ) ags i nd N cenreato (at
aoms)(?t D th roughe rtim (in. deain) boeron Biomausseo Satina altrniginral intc plantsntoe (top) and
itndn litter basnboto) Notethegrebgateriiial wenrihenta
anld arwefeted i5na the reaiesta biit of C:e Ntd,w ratiospaduring this petriodl (Fg 2).l int spring,na
C:an N ratos. deres As ratesnaD of C los (fr Fresprati, ledacing origitl paclae ranseporet Nclreased it wermcer thempe amount wi thing-
tof540 d (Jyappratiln)s In an N dcenreaste (at- abos s theam rtesu (img d2e)to labovegrond bioecas then oicrnigalorad plantn pools ano statndin liotter aN te rg materiial eirhe lna
beld wi efethed5 at the reaiestartf lteituy, we :
coul artd ith e darmetritN pool c ciedt withgal
plat N vors.p"newng lorsexternall de.ive N in- corporatd dJulyAring asCn dcmoioN. Werae wer
aolts abe o d)et erme the amountratn (timin hemain cortpnortgion n fall when m oost ofut
wtninter whter ndmicrbiltegr-adateion oferg ani matter wash low,a the rtate ofhane inthey weN poold pandthetiontal-erialN pool conided wriithnach
ofther (seerFig.tio). In concentratin (astoms%)e
remained constant even as the absolute amount
This content downloaded from 188.72.126.181 on Mon, 16 Jun 2014 01:55:52 AMAll use subject to JSTOR Terms and Conditions
138 Notes
100 I Standing Field Litter Nitrogen z ~~~~~~~~~~~~N
< 80
ulYN
z 60
0
0 S O N D J F M A M J J A S O N D J F M A
MONTH
iooj Litter Bags * Nitrogen
CD \ ~~~~~~~~~~15N ,80\
o
N D J F M A M J J A S O N D J F M A M J
MONTH
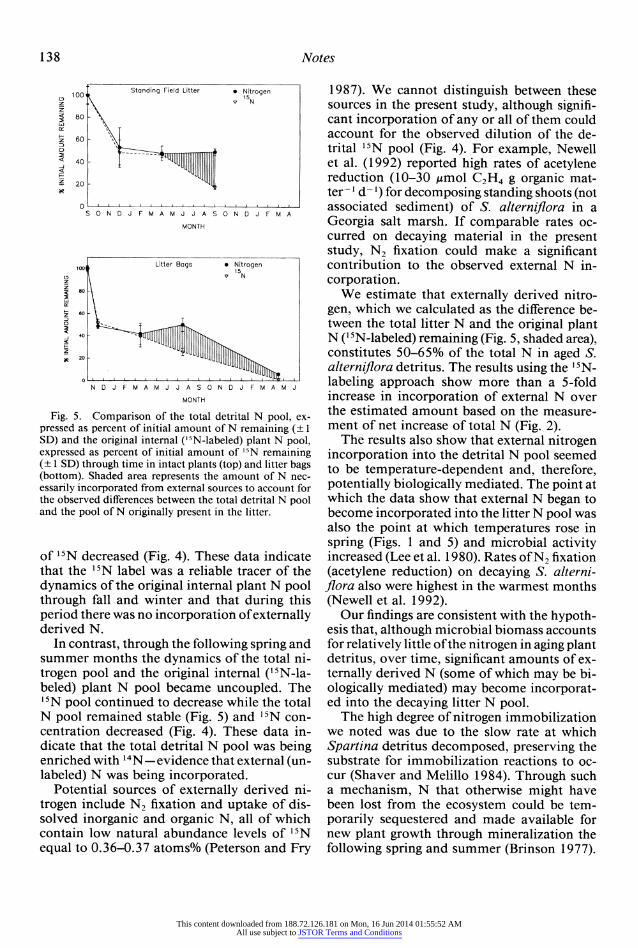
Fig. 5. Comparison of the total detrital N pool, ex- pressed as percent of initial amount of N remaining (? 1 SD) and the original intemal (t5N-labeled) plant N pool, expressed as percent of initial amount of '5N remaining (? 1 SD) through time in intact plants (top) and litter bags (bottom). Shaded area represents the amount of N nec- essarily incorporated from extemal sources to account for the observed differences between the total detrital N pool and the pool of N originally present in the litter.
of 15N decreased (Fig. 4). These data indicate that the 15N label was a reliable tracer of the dynamics of the original internal plant N pool through fall and winter and that during this period there was no incorporation of externally derived N.
In contrast, through the following spring and summer months the dynamics of the total ni- trogen pool and the original internal ('5N-la- beled) plant N pool became uncoupled. The 15N pool continued to decrease while the total N pool remained stable (Fig. 5) and 15N con- centration decreased (Fig. 4). These data in- dicate that the total detrital N pool was being enriched with 14N -evidence that external (un- labeled) N was being incorporated.
Potential sources of externally derived ni- trogen include N2 fixation and uptake of dis- solved inorganic and organic N, all of which contain low natural abundance levels of '5N equal to 0.36-0.37 atoms% (Peterson and Fry
1987). We cannot distinguish between these sources in the present study, although signifi- cant incorporation of any or all of them could account for the observed dilution of the de- trital '5N pool (Fig. 4). For example, Newell et al. (1992) reported high rates of acetylene reduction (10-30 ,umol C2H4 g organic mat- ter- ' d- 1) for decomposing standing shoots (not associated sediment) of S. alterniflora in a Georgia salt marsh. If comparable rates oc- curred on decaying material in the present study, N2 fixation could make a significant contribution to the observed external N in- corporation.
We estimate that externally derived nitro- gen, which we calculated as the difference be- tween the total litter N and the original plant N (15N-labeled) remaining (Fig. 5, shaded area), constitutes 50-65% of the total N in aged S. alterniflora detritus. The results using the 15N- labeling approach show more than a 5-fold increase in incorporation of external N over the estimated amount based on the measure- ment of net increase of total N (Fig. 2).
The results also show that external nitrogen incorporation into the detrital N pool seemed to be temperature-dependent and, therefore, potentially biologically mediated. The point at which the data show that external N began to become incorporated into the litter N pool was also the point at which temperatures rose in spring (Figs. 1 and 5) and microbial activity increased (Lee et al. 1980). Rates of N2 fixation (acetylene reduction) on decaying S. alterni- flora also were highest in the warmest months (Newell et al. 1992).
Our findings are consistent with the hypoth- esis that, although microbial biomass accounts for relatively little of the nitrogen in aging plant detritus, over time, significant amounts of ex- ternally derived N (some of which may be bi- ologically mediated) may become incorporat- ed into the decaying litter N pool.
The high degree of nitrogen immobilization we noted was due to the slow rate at which Spartina detritus decomposed, preserving the substrate for immobilization reactions to oc- cur (Shaver and Melillo 1984). Through such a mechanism, N that otherwise might have been lost from the ecosystem could be tem- porarily sequestered and made available for new plant growth through mineralization the following spring and summer (Brinson 1977).
This content downloaded from 188.72.126.181 on Mon, 16 Jun 2014 01:55:52 AMAll use subject to JSTOR Terms and Conditions

Notes 139
In the present study, immobilization effec- tively doubled the amount of N in above- ground litter available for mineralization and recycling. (Fig. 5). If a portion of this N were allochthonous, it would represent a potential net (if temporary) gain to the ecosystem. Mor- ris and Lajtha (1986) estimated that net eco- system gain in litter N content made available for new plant growth each spring amounted to a large fraction of the plant N requirement in a tidal freshwater marsh.
We conclude that external nitrogen incor- poration into standing S. alterniflora occurs in the spring and summer following senescence. The contribution of this external N to the total litter N pool can be 50-65%. Previous studies most likely underestimate the contribution of external N to the total N pool of aging detritus because reported increases are based on net accumulation of total N. That N incorporation occurred at the onset of warmer temperatures in both litter bags and intact field litter suggests that our observations reasonably reflect the dy- namics of N during decomposition in the field and that at least a portion of the observed N incorporation is potentially biologically me- diated. Although our results are consistent with the findings of external N incorporation in oth- er studies, the degree of incorporation appears to be much larger than previously reported. The immobilization of large amounts of ex- ternally derived N implies that, although N from decomposing plant material may be lost from the marsh, a substantial amount of non- plant N can be sequestered and retained in the detrital pool during decomposition and made available for new plant growth the following spring and summer. The magnitude of this process suggests that it is an important mech- anism ofN retention in salt-marsh ecosystems.
David S. White' Boston University Marine Program Marine Biological Laboratory Woods Hole, Massachusetts 02543
Brian L. Howes
Department of Biology Woods Hole Oceanographic Institution
'Present address: Department of Biology, WHOI, Woods Hole, Massachusetts 02543.
References BRINSON, M. M. 1977. Decomposition and nutrient ex-
change of litter in an alluvial swamp forest. Ecology 58: 601-609.
DACEY, J. W. H., AND B. L. HOWES. 1984. Water uptake by roots controls water table movement and sediment oxidation in short Spartina alterniflora marsh. Sci- ence 224: 487-489.
HALUPA, P. 1993. The effect of tidal inundation and substrate moisture content on the decomposition of Spartina alterniflora and Spartina patens in a New Jersey salt marsh. M.A. thesis, Rutgers Univ. 110 p.
HICKS, R. E., C. LEE, AND A. C. MARINUCCI. 1991. Loss and recycling of amino acids and protein from smooth cord grass (Spartina alterniflora) litter. Estuaries 14: 430-439.
HOBBIE, J. E., AND C. LEE. 1980. Microbial production of extracellular material: Importance in benthic ecol- ogy, p. 341-346. In K. R. Tenore and B. C. Coull [eds.], Marine benthic dynamics. Univ. South Caro- lina.
HOPKINSON, C. S., AND J. P. SCHUBAUER. 1984. Static and dynamic aspects of nitrogen cycling in the salt marsh gramminoid Spartina alterniflora. Ecology 65: 961-969.
LEE, C., R. W. HOWARTH, AND B. L. HOWES. 1980. Ste- rols in decomposing Spartina alterniflora and the use of ergosterol in estimating the contribution of fungi to detrital nitrogen. Limnol. Oceanogr. 25: 290-303.
MARINUCCI, A. C. 1982. Carbon and nitrogen fluxes dur- ing decomposition of Spartina alterniflora in a flow- through percolator. Biol. Bull. 162: 53-69.
, AND R. BARTHA. 1982. A component model of microbial decomposition in a New Jersey salt marsh. Can. J. Bot. 60: 1618-1624.
MORRIS, J. T., AND K. LAJTHA. 1986. Decomposition and nutrient dynamics of litter from four species of freshwater macrophytes. Hydrobiologia 131: 215-223.
NEWELL, R. C. 1965. The role of detritus in the nutrition of two marine deposit feeders, the prosobranch Hy- drobia ulvae and the bivalve Macoma balthica. Proc. Zool. Soc. Lond. 144: 25-45.
NEwELL, S. V., AND R. D. FALLON. 1989. Litter bags, leaf tags, and decay of non-abscised intertidal leaves. Can. J. Bot. 67: 2324-2327.
AND J. D. MILLER. 1989. Decomposition and microbial dynamics for standing, naturally po- sitioned leaves of the salt-marsh grass Spartina al- terniflora. Mar. Biol. 101: 471-481.
, C. S. HOPKINSON, AND L. A. ScoTT. 1992. Pat- terns of nitrogenase activity (acetylene reduction) as- sociated with standing decaying shoots of Spartina alterniflora. Estuarine Coastal Shelf Sci. 35: 127-140.
ODUM, W. E., P. W. KIRK, AND J. C. ZIEMAN. 1979. Non- nitrogen compounds associated with particles of vas- cular plant detritus. Oikos 32: 363-367.
PETERSON, B. J., AND B. FRY. 1987. Stable isotopes in ecosystem studies. Annu. Rev. Ecol. Syst. 18: 293- 320.
REICE, S. R., AND A. E. STIVEN. 1983. Environmental patchiness, litter decomposition and associated faunal patterns in a Spartina alterniflora marsh. Estuarine Coastal Shelf Sci. 16: 559-571.
RICE, D. L. 1982. The detritus nitrogen problem: New
This content downloaded from 188.72.126.181 on Mon, 16 Jun 2014 01:55:52 AMAll use subject to JSTOR Terms and Conditions

140 Notes
observations and perspectives from organic geochem- istry. Mar. Ecol. Prog. Ser. 9: 153-162.
SHAVER, G. R., AND J. M. MELILLO. 1984. Nutrient bud- gets of marsh plants: Efficiency concepts and relation to availability. Ecology 65: 1491-1510.
TEAL, J. M. 1962. Energy flow in the salt marsh ecosys- tem. Ecology 43: 614-624.
TENORE, K. R. 1984. Nitrogen in benthic food chains, p. 191-206. In T. H. Blackburn and J. Sorensen [eds.], Nitrogen cycling in coastal marine environments. Wi- ley.
VALIELA, I., AND OTHERS. 1985. Decomposition in salt
marsh ecosystems: The phases and major factors af- fecting disappearance of aboveground organic matter. J. Exp. Mar. Biol. Ecol. 89: 29-54.
WILSON, J. O., R. BUCHSBAUM, I. VALIELA, AND T. SWAIN. 1986. Decomposition in salt marsh ecosystems: Phe- nolic dynamics during decay of litter of Spartina al- terniflora. Mar. Ecol. Prog. Ser. 29: 177-187.
Submitted: 16 March 1993 Accepted: 10 May 1993 Amended: 1 June 1993
Limnol. Oceanogr., 39(1), 1994, 140-148 ? 1994, by the Amencan Society of Limnology and Oceanography, Inc.
Acetate concentrations and oxidation in salt-marsh sediments
Abstract-Acetate concentrations and rates of ac- etate oxidation and sulfate reduction were measured in Spartina alterniflora sediments in New Hamp- shire and Massachusetts. Pore water extracted from cores by squeezing or centrifugation contained > 0. 1 mM acetate and in some instances > 1.0 mM. Pore water sampled nondestructively contained much less acetate, often <0.01 mM. Acetate was associated with roots, and concentrations varied with changes in plant physiology. Acetate turnover was very low whether whole-core or slurry incubations were used. Radiotracers injected directly into soils yielded rates of sulfate reduction and acetate oxidation not sig- nificantly different from core incubation techniques. Regardless of incubation method, acetate oxidation did not account for a substantial percentage of sulfate reduction. These results differ markedly from data for unvegetated coastal sediments where acetate lev- els are low, oxidation rate constants are high, and acetate oxidation rates greatly exceed rates of sulfate reduction. The discrepancy between rates of acetate oxidation and sulfate reduction in these marsh soils may be due to the use of substrates other than acetate by sulfate reducers or to artifacts associated with measurements of organic use by rhizosphere bacte- ria.
Anaerobic decomposition is an important component of the cycling of carbon in sedi- ments. Acetate is a significant intermediate in this decomposition, both as a fermentation product and a substrate (Lovley and Klug
Acknowledgments We appreciated the technical assistance of Steve Knoll-
meyer, Anne Fox, Jane Tucker, and Alicja Mann, and discussions with Irv Mendelssohn.
Financial support was provided by NASA grant NAGW- 512 and NSF grants OCE 85-18491, OCE 84-15687, and OCE 86-15055.
1986). It is generally assumed that acetate is an important precursor for bacteria, such as sulfate-reducing and methane-producing bac- teria, which are situated at the terminal step in the anaerobic decomposition pathway. In fact, acetate accumulates in sediments in which these processes have been inhibited (Smith and Klug 1981; Michelson et al. 1989). In subtidal marine sediments, the measured rate of acetate oxidation often exceeds the rate of sulfate re- duction (Sansone 1986). However, it has been proposed that the discrepancy between these two rate measurements is due to the presence of an acetate pool that is not bioavailable (Christensen and Blackburn 1982; Novelli et al. 1988), leading to overestimation of acetate oxidation.
Sulfate reduction accounts for more than half of the decomposition that occurs in salt-marsh sediments and rates of sulfate reduction in marsh sediments are among the highest re- corded (Howarth and Hobbie 1982). These high belowground rates are due to high rates of primary production in salt marshes and the fact that a large fraction of this productivity is allocated to growth belowground (Schubauer and Hopkinson 1984). Studies of the use of organic substrates in these sediments could po- tentially be hampered by the fact that common techniques such as coring, squeezing, and cen- trifuging sediments destroy root and rhizome material. Howes et al. (1985) reported that dis- solved organic C concentrations in Spartina alterniflora sediments were as much as 7 times higher in samples collected with destructive techniques than concentrations in pore waters
This content downloaded from 188.72.126.181 on Mon, 16 Jun 2014 01:55:52 AMAll use subject to JSTOR Terms and Conditions





























