Embed Size (px)
Citation preview

Niche Diversity and Niche Size Patterns in Marine FossilsAuthor(s): James W. ValentineSource: Journal of Paleontology, Vol. 43, No. 4 (Jul., 1969), pp. 905-915Published by: SEPM Society for Sedimentary GeologyStable URL: http://www.jstor.org/stable/1302555 .
Accessed: 20/09/2013 08:43
Your use of the JSTOR archive indicates your acceptance of the Terms & Conditions of Use, available at .http://www.jstor.org/page/info/about/policies/terms.jsp
.JSTOR is a not-for-profit service that helps scholars, researchers, and students discover, use, and build upon a wide range ofcontent in a trusted digital archive. We use information technology and tools to increase productivity and facilitate new formsof scholarship. For more information about JSTOR, please contact [email protected].
.
SEPM Society for Sedimentary Geology is collaborating with JSTOR to digitize, preserve and extend access toJournal of Paleontology.
http://www.jstor.org
This content downloaded from 128.233.210.97 on Fri, 20 Sep 2013 08:43:23 AMAll use subject to JSTOR Terms and Conditions

JOURNAL OF PALEONTOLOGY A publication of THE SOCIETY OF ECONOMIC PALEONTOLOGISTS AND MINERALOGISTS and THE PALEONTOLOGICAL SOCIETY with the generous support and cooperation of THE AMERICAN
ASSOCIATION OF PETROLEUM GEOLOGISTS
VOLUME 43 JULY 1969 NUMBER 4
NICHE DIVERSITY AND NICHE SIZE PATTERNS IN MARINE FOSSILS
JAMES W. VALENTINE University of California, Davis
ABSTRACT-The ranges of environment-related functions that can be employed by a species at any time represent the prospective niche size, and the ranges of factors that are actually employed, the realized niche size, of a species. For marine poikilotherms, prospective niches tend to be largest in primitive ecosystems of low species diversity wherein the environment is variable, and smallest in advanced ecosystems of high species diversity wherein the en- vironment is chronologically stable.
Patterns of niche size-frequency distributions of marine poikilotherms have varied as the biosphere has evolved. In shallow tropical waters, modal niche size has decreased as diversification has proceeded in that relatively stable environment, which has been enriched with species through the operation of a latitudinal, climatically-driven "diversity pump." In shallow waters of high latitudes, modal niche size has fluctuated with fluctuations in environ- mental changeability; at present, modal niche sizes are large.
Taxa that produce species of characteristic niche size ranges tend to be most diverse when favored by the niche-size structure; articulate brachiopods, for example, seem to have a rather broad modal niche size and are most diverse in monotonous primitive or in variable advanced ecosystems; most "advanced" bivalves have small niches and are best represented in monotonous advanced ecosystems. There is no reason to believe that diversity maxima have been reached; future ecosystems should be richer in species than those of the present, if the activities of man do not intervene.
INTRODUCTION
THE species niche is one of the more important concepts in paleobiology. As employed here,
the unqualified term niche subsumes the potential interactions of a species with all the factors of the environment, physical and biological. Since evolution is essentially an ecological process operating with genetic machinery, a good case can be made that the niche is the most funda- mental single unit of evolution. It may be reward- ing, therefore, to examine the characteristics of niches, especially the patterns of these character- istics in space and time. The present discussion is focused upon a single aspect of the niche: "size" or "breadth."
To define niche size it is convenient to refer to the concept presented by Hutchinson (1957, 1967). If each separate parameter of the envi- ronment is visualized or represented by a geo- metric dimension, the total environment may be represented as a multidimensional space or hy- perspace containing as many dimensions as
there are environmental parameters. The maxi- mum possible dimensions of all possible envi- ronmental parameters form a prospective envi- ronmental hyperspace. However in any real en- vironment only a small portion of the possible range of these parameters actually exists, and this forms the realized environmental hyper- space (see Simpson, 1944). Each point within the environmental hyperspace represents a unique combination of environmental parame- ters.
The collection of all the points representing the range of conditions under which a species may possibly exist has been called the prospec- tive niche (Valentine, 1968) of that species. A niche thus represents the functional properties of a species and summarizes its environmental requirements, tolerances and preferences. In na- ture the boundaries of the species niche change constantly. Usually the change is quite small compared to the magnitude of the dimensions involved. Nevertheless as gene proportions
905
This content downloaded from 128.233.210.97 on Fri, 20 Sep 2013 08:43:23 AMAll use subject to JSTOR Terms and Conditions

JAMES W. VALENTINE
change, as alleles are added or lost, or even as new gene recombinations occur within gene pools, the frequency distribution of individuals with any given tolerance or requirement usually changes also. All of these events occur fre- quently, some nearly constantly within most natural populations; the niche evolves nearly constantly. Any particular niche is not a con- stant feature of a species, but is more or less in flux. The realized niche is the range of those parameters that actually exists as an aspect of a real species at a given time-in other words, the parts of the prospective functions of a popula- tion niche that overlap with the realized envi- ronment (see Simpson, 1944). The realized niche fluctuates frequently in most species. It is evident that the niche is an extension of the properties of the species, and cannot exist inde- pendently of the species; there are no "vacant niches", although there may be unoccupied envi- ronmental hyperspace (see Connell & Orias, 1964). Text-figure 1 depicts some aspects of the niche of a species at an instant in time.
The niche of any species could presumably be specified at any instant, if we know enough about the species' properties. The size and qual- ity of the niche is underpinned by a gene pool, and thus each species' prospective niche is unique. Most of the niche boundaries would not be represented by a single point within the hy- pervolume that we are using as a conceptual aid, but rather the boundaries have the form of frequency distributions, as the individuals form- ing the species at any time would display varia- tion in any given tolerance or requirement. Fur- thermore there are no doubt many ways in which the niche hypervolume is occupied by in- dividuals. For example, the individuals compos- ing the species population at any time may each have a range of environmental interactions nearly as broad as the realized niche, and thus broadly overlap functionally. The individuals may occupy only small segments of one or sev- eral dimensions of the realized niche, and thus in a manner of speaking each have their own subniche; or there may be numbers of partially isolated populations, each with different niche properties. There may be subpopulations with special subniches inhabiting specialized, atypical environments that interfinger with or are con- tained within the typical species' biotope, with or without broad overlap of individual func- tional ranges. There are surely many additional niche occupation patterns. These patterns may be called niche structure. Functional or genetic aspects of niche structure have been considered by Dempster (1955), Levins (1962), Ludwig (1950), Van Valen (1965) and others; Mayr
(1963, especially chapters 9 and 13) has dis- cussed aspects of niche structure for animals and Stebbins (1950) for plants. Niche structure may be considered as a separate attribute from niche size, although these attributes may often be related.
Niche size may be expressed as a function of the range of tolerances and requirements of in- dividuals composing a species; the range of each parameter may be visualized as measured along the appropriate dimension in the niche hyperspace. It is clear enough which of two niches, A and B, is the larger when niche A in- cludes broader ranges along every parameter of the niche hyperspace than niche B. The hyper- volume of niche A is simply larger than that of niche B. However in a more general case, one niche will be more extensive along some dimen- sions and more restricted along others than will another niche. As the scales of most of the var- ious dimensions have no necessary relation to each other, it cannot be generally specified whether, for example, a certain extension of a tolerance along one dimension is equivalent to,
TEXT-FIG. 1-Diagrammatic representation of realized and prospective aspects of the species niche. 1. Total range of prospective functions of a given gene pool at an instant in time. THE PROSPECTIVE NICHE. 1A, portion of prospective niche not real- ized because it is neither represented in realized phenotypes nor overlaps with realized environment. IB, portion of prospective niche not represented in realized phenotypes although it overlaps with real- ized environment. 2. Prospective functions of the realized population. 2A, portion of prospective func- tion of realized population that does not overlap with realized environment. 2B, realized portion of prospective function of realized population. THE REALIZED NICHE. 3. The realized environment.
906
This content downloaded from 128.233.210.97 on Fri, 20 Sep 2013 08:43:23 AMAll use subject to JSTOR Terms and Conditions

NICHE DIVERSITY AND NICHE SIZE PATTERNS
larger than, or smaller than a certain restriction of another tolerance along another dimension. Furthermore extension a given way along a given dimension for one species may involve much physiological adjustment and require large numbers of gene changes with many sec- ondary changes required to maintain genetic coadaptation. For another species, the same change may involve only a single or at least very few gene substitutions. It is obvious that if these complications are pursued they will lead into a morass of difficulties. These difficulties are compounded when the complexities of niche structure are also considered. Although it ap- pears that these difficulties may be overcome, it is better not to get bogged down here, and therefore I shall define a first approximation of niche size and employ this concept in subsequent discussion, for which it is generally adequate. It is not intended, however, to gloss over the diffi- culties of defining niche size. The definition adopted relies on contrasting two niches; each must be instantaneous but they need not be con- temporaneous. The one with greater extensions along the most parameters is the larger. Any parameter present in one niche but absent in the other represents a greater extension.
The sizes of prospective niches are clearly based on the range and variety of potential en- vironmental interactions with which species are endowed by their gene pools at any time. The sizes of realized niches, however, involve more factors. They are controlled partly by the range of environmental conditions, physical and bio- logical, that happen to be realized in the living region of the species, and they fluctuate as the environment fluctuates. Realized niches also de- pend partly upon the phenotypes that actually compose the population at any time. That is, different phenotypes have different environmen- tal interactions, and as the phenotype mixtures change due to birth and death the realized niche also changes. The nature of the phenotypes is related in turn to the gene combinations that happen to be realized in genotypes and to the environmental history during the ontogenetic history of each individual. All these factors, then, will affect the quality and size of the real- ized niche. Some of these relations are indicated in Text-figure 1.
Niche size has been defined in a variety of ways, and care is required in contrasting the conclusions of different authors, who may be talking about different things. The concept of niche is sometimes employed descriptively to mean habitat, or is functional but restricted only to physical environmental parameters. By niche size, Klopfer and MacArthur (1960)
mean the measurable range of conditions used by a species. This is close to but different from the "realized niche" of the present paper, the difference depending upon the interpretation given to the word "measurable". Van Valen (1965), on the other hand, defines niche size ("width", actually) as ". . . the proportion of the total multidimensional space of limiting re- sources used by a species or segment of a commu- nity." This concept may be close or identical to that of the "realized niche" as employed here, depending partly upon the interpretation of the words "limiting" and "resources."
In this paper it is convenient to distinguish two classes of environmental factors: 1) those for which in any given situation organisms make no significant difference in quality or quantity; these will be called pervasive factors; 2) those which in any given situation are sig- nificantly altered in quality or quantity by orga- nisms, such as living space or food commonly are; these will be called consumable factors. A factor may be consumable in one situation and not in another.
FACTORS INFLUENCING NICHE DIVERSITY
Niche diversity is equivalent to species diver- sity, and many processes of species diversifica- tion are well-known. Mayr and others, from Darwin on, have emphasized that the isolation of populations is a critical factor in speciation, and that isolation is chiefly a geographical phe- nomenon (see for example Mayr, 1942, 1963). Most isolating mechanisms appear to operate fairly randomly with respect to the physical ge- ography of the oceans although exceedingly het- erogeneous regions such as archipelagoes might be sites of higher than average opportunities for isolation. Certainly heterogeneous regions can support a more diverse biota than homoge- neous regions, since more habitats are available. A major factor in niche diversity in any given region, then, is simply habitat diversity.
The major pattern of marine species diversity at present is in the form of an irregular gra- dient, with high species diversity in low lati- tudes grading to low diversity in high latitudes. There has been much discussion on the causes and significance of this gradient, which is found in the terrestrial as well as marine realm; sev- eral major discussions and reviews have ap- peared (Fisher, 1960; MacArthur, 1965; and Pianka, 1966, who includes a classification of hypotheses to account for latitudinal diversity gradients and a bibliography). Recently I have proposed a model of diversification that is re- lated to climatic fluctuations and may account for progressive enrichment in species of the
907
This content downloaded from 128.233.210.97 on Fri, 20 Sep 2013 08:43:23 AMAll use subject to JSTOR Terms and Conditions

JAMES W. VALENTINE
low-latitude biota (Valentine, 1967, 1968). The mechanism of this model is called a "diversity pump." These diversity pumps operate as fol- lows.
In shallow water the latitudinal range of poi- kilotherms is chiefly a function of the latitudinal temperature gradient. The gradient was proba- bly low during most of the Phanerozoic, for polar climates seem to have usually been rela- tively mild, with only a narrow range between polar and equatorial temperature means. Sea- sonal fluctuations are assumed to have been more marked in progressively higher latitudes throughout the Phanerozoic just as they are today, so that some small temperature gradient was always present. In high latitudes the sea- sonal climatic changes, especially in temperature and light intensity, would cause changes in a multitude of factors such as the metabolic rates of poikilotherms, the solubilities of a whole con- stellation of nutrients and other materials, the primary productivity and therefore the energy supply at the base of the trophic pyramid, and so on. Thus for an average year it is necessary for high latitude organisms to have wider real- ized niches than those in low latitudes, for they must inhabit a broader environmental hyper- space. The term niche density will be used for the ratio between the mean realized niche hy- pervolume and the hypervolume of realized en- vironmental hyperspace.
Take a case when the poles are mild, environ- mental variability is slight, and all species range from equator to poles so that niche diversity is everywhere the same, and niche density is also everywhere the same. Let the poles cool; the realized environmental hyperspace becomes broader poleward and thermophiles and species with niches too narrow in critical dimensions to tolerate polar conditions will have their ranges restricted towards the equator. Cooling need probably be only a degree or so to be effective. As the range of a species A is restricted equa- torward there is an opportunity for another spe- cies B to occupy parts of environmental hyper- space that become vacant poleward. Species B may originate from marginal populations of species A that are finally isolated by growing thermal barriers, or from populations of A that have already been isolated by any of the "ran- dom" mechanisms, or from an entirely different lineage. The point of interest here is that an in- creasing gradient in temperature and variability acts as a diversification mechanism, increasing the number of species. As yet there need be no diversity gradient, however, in the model we are developing. Neither need there be any gradient in niche density. The diversification is due only
to the increased temperature gradient, and is on a provincial level. Poleward communities have fewer species than before, and equatorial com- munities have the same numbers of species as before, but now the poleward and equatorward biotas contain different species, so that the total biota has diversified.
An enrichment of communities in species oc- curs whenever the climatic gradient decreases owing to polar warming. This allows lower-lati- tude species to expand their ranges poleward. Higher-latitude species are either eliminated or find a place in the immigrating ecosystems. Many must become extinct, but some could be accommodated through processes of niche parti- tioning, such as discussed by Brown & Wilson (1956), Klopfer (1962), MacArthur & Levins (1964, 1967), Miller (1967), and others. Thus two or more species niches would be found in the environmental hyperspace formerly occu- pied by a single niche. The sort of species en- richment that may result from niche partition- ing (not necessarily due to a diversity pump) is well shown by the studies of Kohn (1966) on feeding in Conus. In this case feeding special- ization is inversely related to species diversity. Off southern California there is a single small species of Conus (C. californicus) that feeds upon a wide variety of marine animals, at least 28 species in six classes of four phyla, chiefly mollusks and polychaetes. In Hawaii, by con- trast, there is a large number of species of Conus, and six or more may be associated in the same general habitat. The feeding habits of these species are narrower than those of C. cali- fornicus. Eight of the most abundant Hawaiian cones are vermivorous, and each specializes on a different prey species or on a different single prey taxon higher than species.
Another mode of accommodation of addi- tional species in ecosystems during warming trends would be for species to colonize unten- anted portions of environmental hyperspace. Probably most diversifications will involve some of both niche partitioning and colonization of environmental hyperspace, but for many forms one or the other process will be dominant. The species that colonize new environmental hyper- space may sometimes be the forerunners of higher taxa. In any case the enrichment of eco- systems would be accompanied by a decrease in modal niche size, and niche density increases. The next cooling trend, however, forces many species to abandon portions of their ranges that lie in high latitudes and new forms are again evolved poleward. Now there is a niche diver- sity and density difference between the pole- ward regions and low latitudes. Continued fluc-
908
This content downloaded from 128.233.210.97 on Fri, 20 Sep 2013 08:43:23 AMAll use subject to JSTOR Terms and Conditions
NICHE DIVERSITY AND NICHE SIZE PATTERNS
tuations pump more and more species equator- wards, where they accumulate.
Thus climatic fluctuations form the basis of this diversity pump. The magnitude of the fluc- tuations need only be such as to cause a 1?C. or 2?C: change in average annual temperatures in high latitudes. The biosphere is enriched in spe- cies during a cooling trend, but they are provin- cially segregated; the biosphere is somewhat impoverished in species during a warming trend, but communities are enriched. A second cooling trend leads to a new provincial enrich- ment and carries total diversity to new highs, and so the pump continues to operate. The pump works across all latitudes except the low- est, but the communities that are most enriched are equatorward in the regions of relatively mo- notonous physical environment (Valentine, 1967, 1968a), where niches become smaller and smaller as environment space is packed with species. These events establish a diversity gra- dient.
A complication occurs whenever the poles reach freezing in winter. Further cooling now decreases the temperature range by depressing the summer temperature, until temperatures may be near freezing throughout the year. High latitudes are thus becoming thermally monoto- nous at these times. However seasonality is still greatest there, and the appearance of ice intro- duces new sorts of environmental variability, such as salinity changes related to freezing and thawing and narrowing of the period of high primary productivity through restriction of light penetration. The realized environmental hyperspace remains broader in higher latitudes.
A diversity pump should be created whenever a biota is anchored at one end by relatively sta- ble environments and fluctuates between rela- tive environmental monotony and changeability on the other end. This causes major gradients in diversity and in niche size. Increases in di- versity and decreases in niche size are also oc- curring as a result of speciation from "random" processes of genetic isolation not related to the origin of the gradients. There will therefore be an overall tendency towards high diversity that is at first unbiased with respect to depth or lati- tude. However, to the extent that these radia- tions and other "random" diversity processes operate most frequently where diversity is high, they will serve to accentuate the major gra- dients established by diversity pumps.
In summary, species diversification increases during trends of climatic deterioration and polar cooling. During trends of climatic ame- lioration, extinction exceeds speciation, al- though communities become enriched in species
C,)
a.
0
gmall ^I.ae NICHE SIZE
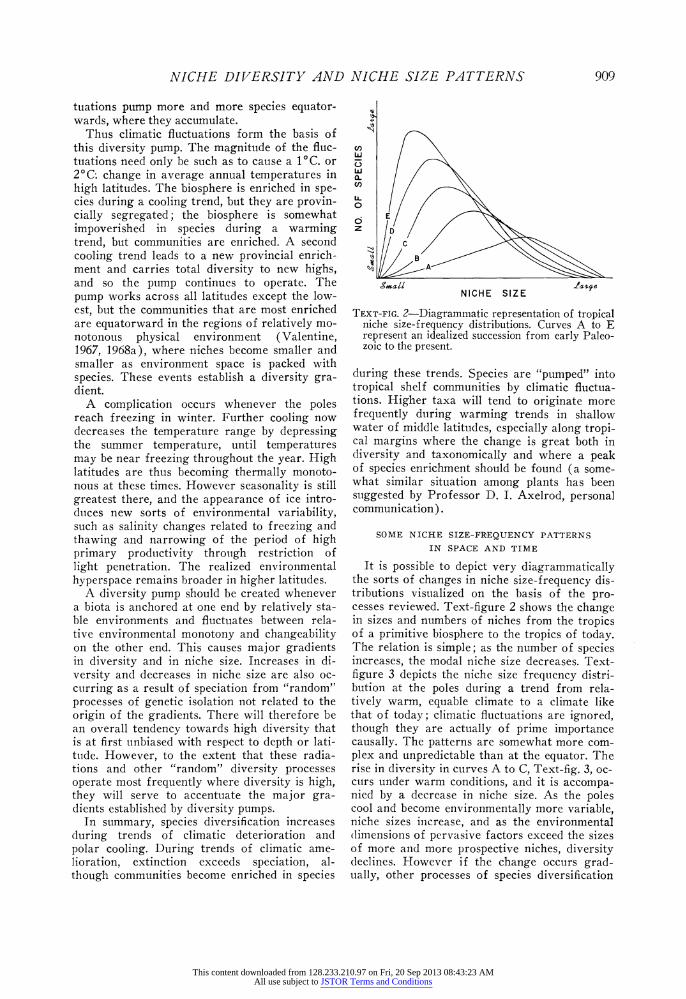
TEXT-FIG. 2-Diagrammatic representation of tropical niche size-frequency distributions. Curves A to E represent an idealized succession from early Paleo- zoic to the present.
during these trends. Species are "pumped" into tropical shelf communities by climatic fluctua- tions. Higher taxa will tend to originate more frequently during warming trends in shallow water of middle latitudes, especially along tropi- cal margins where the change is great both in diversity and taxonomically and where a peak of species enrichment should be found (a some- what similar situation among plants has been suggested by Professor D. I. Axelrod, personal communication).
SOME NICHE SIZE-FREQUENCY PATTERNS
IN SPACE AND TIME
It is possible to depict very diagrammatically the sorts of changes in niche size-frequency dis- tributions visualized on the basis of the pro- cesses reviewed. Text-figure 2 shows the change in sizes and numbers of niches from the tropics of a primitive biosphere to the tropics of today. The relation is simple; as the number of species increases, the modal niche size decreases. Text- figure 3 depicts the niche size frequency distri- bution at the poles during a trend from rela- tively warm, equable climate to a climate like that of today; climatic fluctuations are ignored, though they are actually of prime importance causally. The patterns are somewhat more com- plex and unpredictable than at the equator. The rise in diversity in curves A to C, Text-fig. 3, oc- curs under warm conditions, and it is accompa- nied by a decrease in niche size. As the poles cool and become environmentally more variable, niche sizes increase, and as the environmental (limensions of pervasive factors exceed the sizes of more and more prospective niches, diversity declines. However if the change occurs grad- ually, other processes of species diversification
909
This content downloaded from 128.233.210.97 on Fri, 20 Sep 2013 08:43:23 AMAll use subject to JSTOR Terms and Conditions
JAMES W. VALENTINE
Cn, w o w a, cn
LL 0
o z
S~all NICHE SIZE
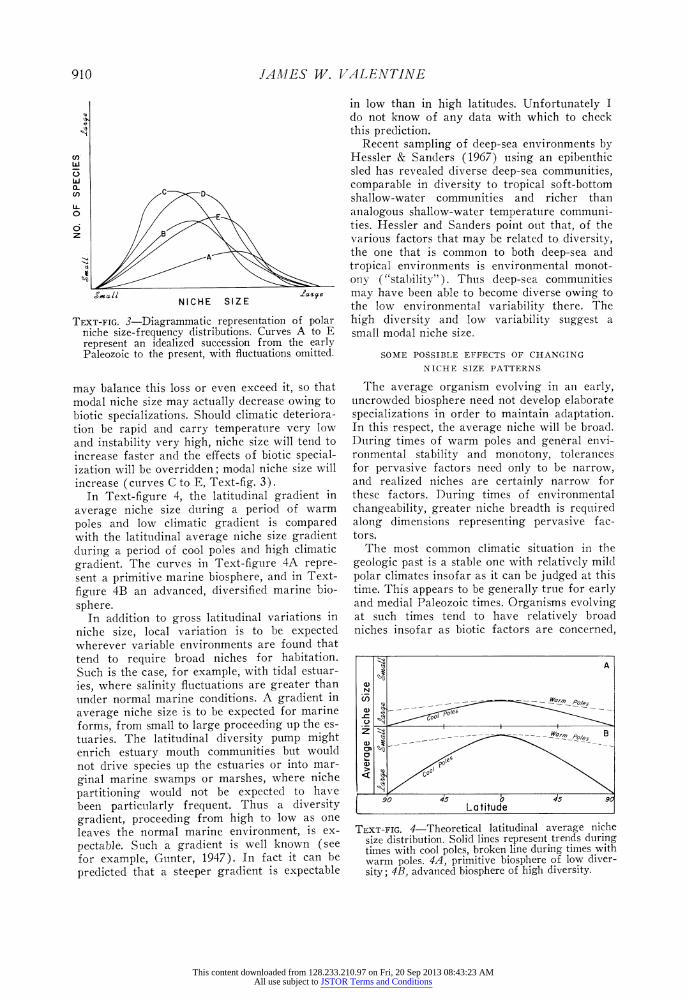
TEXT-FIG. 3-Diagrammatic representation o niche size-frequency distributions. Curves : represent an idealized succession from th Paleozoic to the present, with fluctuations c
may balance this loss or even exceed it, modal niche size may actually decrease ov biotic specializations. Should climatic det tion be rapid and carry temperature ve and instability very high, niche size will I increase faster and the effects of biotic s ization will be overridden; modal niche si increase (curves C to E, Text-fig. 3).
In Text-figure 4, the latitudinal grad average niche size during a period of
poles and low climatic gradient is coi with the latitudinal average niche size g during a period of cool poles and high c
gradient. The curves in Text-figure 4A sent a primitive marine biosphere, and ir
figure 4B an advanced, diversified mari sphere.
In addition to gross latitudinal variat niche size, local variation is to be e: wherever variable environments are fou tend to require broad niches for hab Such is the case, for example, with tidal ies, where salinity fluctuations are great under normal marine conditions. A grac average niche size is to be expected for forms, from small to large proceeding up tuaries. The latitudinal diversity pump enrich estuary mouth communities but not drive species up the estuaries or ini
ginal marine swamps or marshes, wher
partitioning would not be expected t been particularly frequent. Thus a d gradient, proceeding from high to low leaves the normal marine environment, pectable. Such a gradient is well knov for example, Gunter, 1947). In fact it
predicted that a steeper gradient is ex]
in low than in high latitudes. Unfortunately I do not know of any data with which to check this prediction.
Recent sampling of deep-sea environments by Hessler & Sanders (1967) using an epibenthic sled has revealed diverse deep-sea communities, comparable in diversity to tropical soft-bottom shallow-water communities and richer than analogous shallow-water temperature communi- ties. Hessler and Sanders point out that, of the various factors that may be related to diversity, the one that is common to both deep-sea and tropical environments is environmental monot- ony ("stability"). Thus deep-sea communities
2a4e, may have been able to become diverse owing to the low environmental variability there. The
f polar high diversity and low variability suggest a A to E small modal niche size. e early )mitted. SOME POSSIBLE EFFECTS OF CHANGING
NICHE SIZE PATTERNS
so that The average organism evolving in an early, ving to uncrowded biosphere need not develop elaborate eriora- specializations in order to maintain adaptation. ry low In this respect, the average niche will be broad. tend to During times of warm poles and general envi-
special- ronmental stability and monotony, tolerances ize will for pervasive factors need only to be narrow,
and realized niches are certainly narrow for ient in these factors. During times of environmental
warm changeability, greater niche breadth is required mpared along dimensions representing pervasive fac- radient tors. :limatic The most common climatic situation in the
repre- geologic past is a stable one with relatively mild n Text- polar climates insofar as it can be judged at this ne bio- time. This appears to be generally true for early
and medial Paleozoic times. Organisms evolving :ions in at such times tend to have relatively broad xpected niches insofar as biotic factors are concerned, nd that fitation. estuar- er than dient in marine
)the es- ) might
would to mar- e niche o have iversity as one
, is ex- vn (see can be
pectable
TEXT-FIG. 4-Theoretical latitudinal average niche size distribution. Solid lines represent trends during times with cool poles, broken line during times with warm poles. 4A, primitive biosphere of low diver- sity; 4B, advanced biosphere of high diversity.
910
This content downloaded from 128.233.210.97 on Fri, 20 Sep 2013 08:43:23 AMAll use subject to JSTOR Terms and Conditions
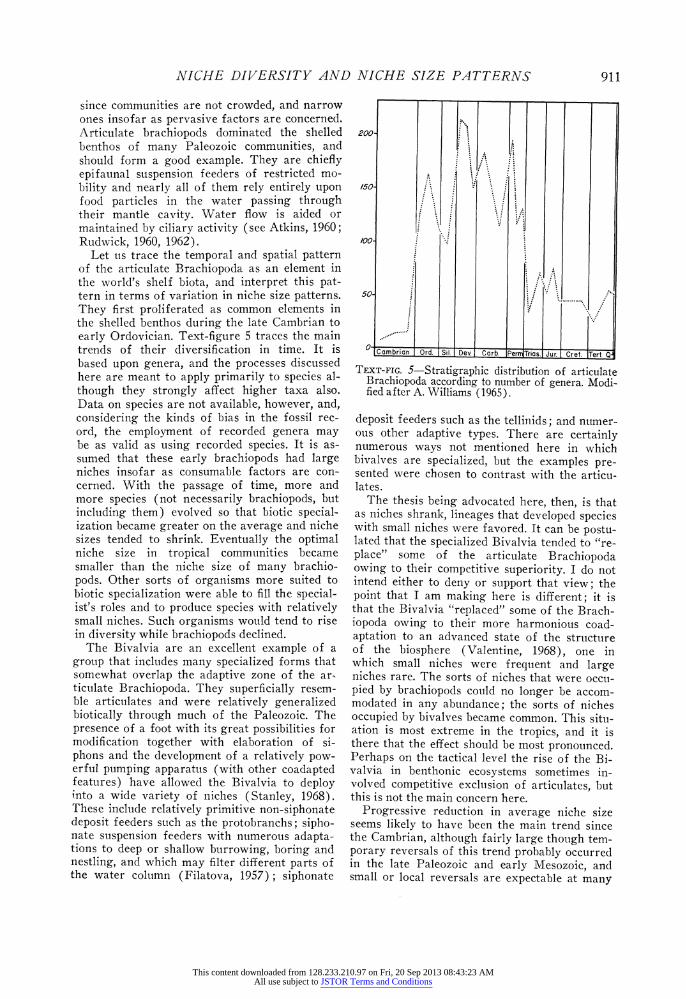
NICHE DIVERSITY AND NICHE SIZE PATTERNS
since communities are not crowded, and narrow ones insofar as pervasive factors are concerned. Articulate brachiopods dominated the shelled benthos of many Paleozoic communities, and should form a good example. They are chiefly epifaunal suspension feeders of restricted mo- bility and nearly all of them rely entirely upon food particles in the water passing through their mantle cavity. Water flow is aided or maintained by ciliary activity (see Atkins, 1960; Rudwick, 1960, 1962).
Let us trace the temporal and spatial pattern of the articulate Brachiopoda as an element in the world's shelf biota, and interpret this pat- tern in terms of variation in niche size patterns. They first proliferated as common elements in the shelled benthos during the late Cambrian to early Ordovician. Text-figure 5 traces the main trends of their diversification in time. It is based upon genera, and the processes discussed here are meant to apply primarily to species al- though they strongly affect higher taxa also. Data on species are not available, however, and, considering the kinds of bias in the fossil rec- ord, the employment of recorded genera may be as valid as using recorded species. It is as- sumed that these early brachiopods had large niches insofar as consumable factors are con- cerned. With the passage of time, more and more species (not necessarily brachiopods, but including them) evolved so that biotic special- ization became greater on the average and niche sizes tended to shrink. Eventually the optimal niche size in tropical communities became smaller than the niche size of many brachio- pods. Other sorts of organisms more suited to biotic specialization were able to fill the special- ist's roles and to produce species with relatively small niches. Such organisms would tend to rise in diversity while brachiopods declined.
The Bivalvia are an excellent example of a group that includes many specialized forms that somewhat overlap the adaptive zone of the ar- ticulate Brachiopoda. They superficially resem- ble articulates and were relatively generalized biotically through much of the Paleozoic. The presence of a foot with its great possibilities for modification together with elaboration of si- phons and the development of a relatively pow- erful pumping apparatus (with other coadapted features) have allowed the Bivalvia to deploy into a wide variety of niches (Stanley, 1968). These include relatively primitive non-siphonate deposit feeders such as the protobranchs; sipho- nate suspension feeders with numerous adapta- tions to deep or shallow burrowing, boring and nestling, and which may filter different parts of the water column (Filatova, 1957); siphonate
200-
/50-
/00-
50-
0
\ . .
-: :' :
Cambrian Ord. . Dev. Carb. PerTrias. Jur. Cret. ertQ
TEXT-FIG. 5-Stratigraphic distribution of articulate Brachiopoda according to number of genera. Modi- fied after A. Williams (1965).
deposit feeders such as the tellinids; and numer- ous other adaptive types. There are certainly numerous ways not mentioned here in which bivalves are specialized, but the examples pre- sented were chosen to contrast with the articu- lates.
The thesis being advocated here, then, is that as niches shrank, lineages that developed species with small niches were favored. It can be postu- lated that the specialized Bivalvia tended to "re- place" some of the articulate Brachiopoda owing to their competitive superiority. I do not intend either to deny or support that view; the point that I am making here is different; it is that the Bivalvia "replaced" some of the Brach- iopoda owing to their more harmonious coad- aptation to an advanced state of the structure of the biosphere (Valentine, 1968), one in which small niches were frequent and large niches rare. The sorts of niches that were occu- pied by brachiopods could no longer be accom- modated in any abundance; the sorts of niches occupied by bivalves became common. This situ- ation is most extreme in the tropics, and it is there that the effect should be most pronounced. Perhaps on the tactical level the rise of the Bi- valvia in benthonic ecosystems sometimes in- volved competitive exclusion of articulates, but this is not the main concern here.
Progressive reduction in average niche size seems likely to have been the main trend since the Cambrian, although fairly large though tem- porary reversals of this trend probably occurred in the late Paleozoic and early Mesozoic, and small or local reversals are expectable at many
911
This content downloaded from 128.233.210.97 on Fri, 20 Sep 2013 08:43:23 AMAll use subject to JSTOR Terms and Conditions
JAMES W. VALENTINE
60-
50-
40-
30-
20-
10-
7-80 160-70 150-60 40-50130-40 120-301 0-20 10 -/ I 0-0 I /0-20 1 20 -30 1 30-40 1 40-50
N 50-60 60-70 170-80 |
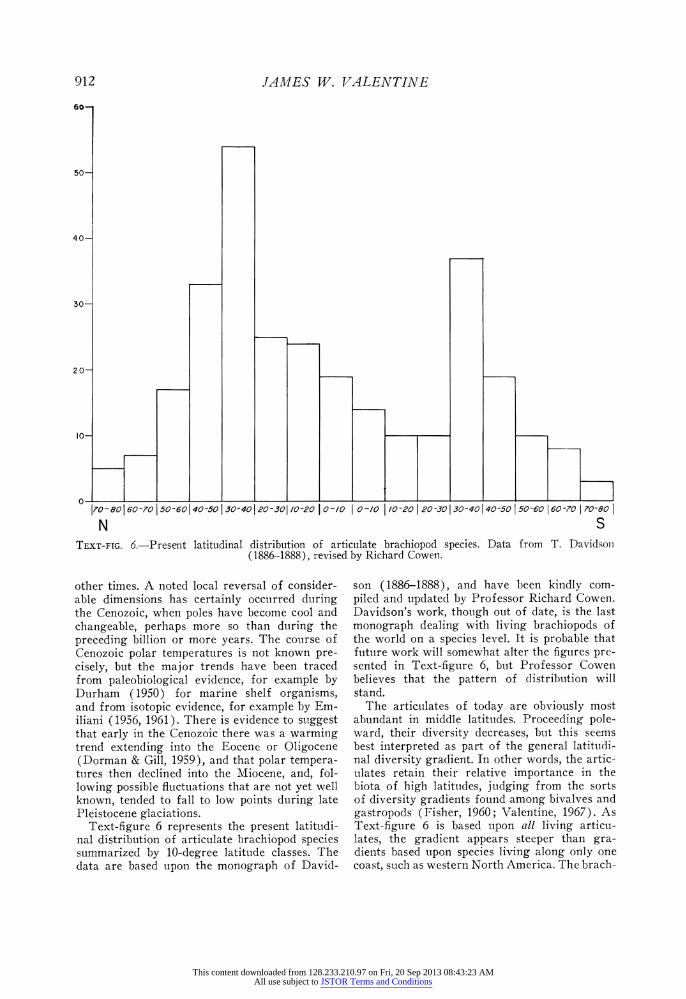
S TEXT-FIG. 6.-Present latitudinal distribution of articulate brachiopod species. Data from T. Davidson
(1886-1888), revised by Richard Cowen.
other times. A noted local reversal of consider- able dimensions has certainly occurred during the Cenozoic, when poles have become cool and changeable, perhaps more so than during the preceding billion or more years. The course of Cenozoic polar temperatures is not known pre- cisely, but the major trends have been traced from paleobiological evidence, for example by Durham (1950) for marine shelf organisms, and from isotopic evidence, for example by Em- iliani (1956, 1961). There is evidence to suggest that early in the Cenozoic there was a warming trend extending into the Eocene or Oligocene (Dorman & Gill, 1959), and that polar tempera- tures then declined into the Miocene, and, fol- lowing possible fluctuations that are not yet well known, tended to fall to low points during late Pleistocene glaciations.
Text-figure 6 represents the present latitudi- nal distribution of articulate brachiopod species summarized by 10-degree latitude classes. The data are based upon the monograph of David-
son (1886-1888), and have been kindly com- piled and updated by Professor Richard Cowen. Davidson's work, though out of date, is the last monograph dealing with living brachiopods of the world on a species level. It is probable that future work will somewhat alter the figures pre- sented in Text-figure 6, but Professor Cowen believes that the pattern of distribution will stand.
The articulates of today are obviously most abundant in middle latitudes. Proceeding pole- ward, their diversity decreases, but this seems best interpreted as part of the general latitudi- nal diversity gradient. In other words, the artic- ulates retain their relative importance in the biota of high latitudes, judging from the sorts of diversity gradients found among bivalves and gastropods (Fisher, 1960; Valentine, 1967). As Text-figure 6 is based upon all living articu- lates, the gradient appears steeper than gra- dients based upon species living along only one coast, such as western North America. The brach-
01
912
I I
This content downloaded from 128.233.210.97 on Fri, 20 Sep 2013 08:43:23 AMAll use subject to JSTOR Terms and Conditions

NICHE DIVERSITY AND NICHE SIZE PATTERNS
iopods therefore partake in the normal diver- sity trend from middle to high latitudes, and can be considered as chiefly temperate to arctic (or antarctic) forms today. Although few in number, some high-latitude species are repre- sented by large populations. From middle to low latitudes, the brachiopod diversity change op- poses the normal trend. In fact, some of the tropical species are found only in deep water. In the Paleozoic, however, articulates flourished in tropical and subtropical climates in shallow wa- ters according to present interpretations.
Bivalves are proportionately most diverse, relative to brachiopods, in the tropics. However they are far more diverse and abundant than articulates even in temperate and cold marine shelf communities.
Adequate data on the niche size of bivalve species in high latitudes compared with their congeners or other close allies in low latitudes is not available, apart from the obvious observa- tion that the great variability in many environ- mental factors in high latitudes requires broader realized niches. A clue to niche size may be afforded by the great variation in shell morphology exhibited by many high-latitude species-myaceans and astartaceans, for exam- ple. Among gastropods, some families with nu- merous species in high latitudes, such as the Neptuneidae and Buccinidae, exhibit unusually large variation in shell morphology. Whether or not these particular shell variations may corre- late with enlarged functional ranges and there- fore enlarged niche sizes is not certain. Varia- tion in the shells of many forms in marginal marine environments, forms that face large variations in salinity and other factors, is well known for some groups, including crustaceans and a few mollusks, such as thaidid gastropods. In nonmarine groups the phenomenon is also well-known, for example among plants (Steb- bins, 1950, especially p. 176-181), wherein phenotypic plasticity tends to be high among species, chiefly annuals, that inhabit variable en- vironments or are pioneers in fluctuating asso- ciations. Again, the evidence is suggestive but the case remains open. Abundant data on mor- phologic variation between high and low lati- tude forms and correlation of the variation with functions would be most valuable.
ADDITIONAL CONSIDERATIONS
Many of the generalities that have been pro- posed to explain major or recurrent evolution- ary events lend themselves easily to consider- ation as phenomena of niche size change. For example, the observation that groups commonly
produce specialized or bizarre species as they approach extinction has been shown to be due, not to any "law of racial senility," but to the natural but not universal tendency for evolution to result in greater complexity and increasing narrowness of adaptation. That extinction would be likely in the event of unusual environ- mental changes related to the specialization is attributed to the relative inflexibility of nar- rowly specialized forms in adapting to new con- ditions. This explanation may be recast in terms of niche size, as follows: evolution commonly produces species with smaller and smaller niches within evolving groups; environmental change that eliminates parts of the small niches or that places these narrowly adapted forms at any disadvantage relative to other species tends to cause extinction. An additional explanation is suggested by the preceding discussion of niche size distributions: environmental change that broadens the average niche size will eliminate numbers of species with small niches, and these are often forms that have been evolving in- creasingly narrow specializations (see also Simpson, 1944).
In a similar way a correlation of reduction of climatic equability with extinction, such as is persuasively argued by Axelrod for large Qua- ternary mammals (1967) and by Axelrod & Bailey (1968) for dinosaurs, may be considered as a case of extinction due to environmental ex- tensions of niche parameters that exceed the niche dimensions of certain species.
The present distributional pattern displayed by living articulate brachiopods is shared by other marine invertebrates, and it seems likely that similar explanations may apply at least in part to these others.
Major temporal changes in diversity of taxa are exhibited by such patterns as are depicted in Text-figure 5. In any analysis of such patterns it is necessary to carefully separate the effects owing to different levels of organization of the biosphere (Valentine, 1968). For example, the Permo-Triassic decline in articulates is the most dramatic change known in their generic diver- sity, except for their original deployment in the Upper Cambrian to Lower Ordovician. Before the diversity change can be evaluated, however, it is necessary to understand something of the general ecological situation for these times. If provinciality is high, high species diversity counts might mainly be reflecting provincial di- versity, while relatively few species might ac- tually be living within any given province. This provincial effect is certainly one of the causes of high over-all species diversity today among
913
This content downloaded from 128.233.210.97 on Fri, 20 Sep 2013 08:43:23 AMAll use subject to JSTOR Terms and Conditions

JAMES W. VALENTINE
shelled benthonic forms relative to times in the Mesozoic. Similarly, if community diversity is high, species diversity counts within a province may reflect chiefly the environmental heteroge- neity (as distinct from changeability) and rela- tively few species might actually live together in the same community. For example, articu- lates were common in the large reef communi- ties of the Permian (Newell, and others, 1953). In fact the niches of the brachiopods in these reef ecosystems may have been narrower on the average than at any other time or place (Rud- wick & Cowen, 1968). In the Upper Permian of the Caucasus, there are reported to be 281 spe- cies of articulate brachiopods in reef facies, and 91 in the non-reef facies (Ruzhentsev & Sary- cheva, 1965, p. 65). With the disappearance of these reefs, brachiopod diversity must have been markedly lowered. This did not necessarily af- fect brachiopod diversity in other communities, however, even in the same provinces as the reefs; the disappearance of reef-related niches would not relieve biotic pressure on niches in non-reefal communities. These considerations strengthen a point made in another context (Valentine, 1958), that evaluation of evolution- ary pathways of populations, of communities, of provinces, and of the entire planetary biota, should go hand in hand.
In sum, niche size patterns of the marine realm have changed as the ecological structure of the biosphere has evolved. Insofar as differ- ent taxa have different limitations or different potentials for developing species with certain niche-size ranges, the relative importance of these taxa in the biota will be altered as niche size-frequency patterns are altered. Thus some of the patterns of diversity exhibited by many taxa in space and time may be owing to the evolution of this structural aspect of the bio- sphere. There is no special reason to suspect that communities will not continue to be en- riched in species. Biotic diversity should con- tinue to rise in the future, providing that man does not continue to interfere.
ACKNOWLEDGMENTS
Richard Cowen, Department of Geology, Uni- versity of California, Davis, has drawn upon his broad knowledge of the Brachiopoda to provide many of the examples employed herein, and to contribute to the interpretation of their distribu- tional patterns; and he has reviewed the manu- script. The manuscript was additionally re- viewed by D. I. Axelrod, W. M. Hamner, and J. H. Lipps, University of California, Davis; these reviews resulted in much improvement. It is
therefore a pleasure to express my debt and thanks for all this aid. Drafting is by Mrs. Ruth Darden.
REFERENCES
ATKINS, D., 1960, The ciliary feeding mechanism of the Megathyridae (Brachiopoda), and the growth stages of the lophophore: Jour. Marine Biol. Assoc. United Kingdom, v. 39, p. 459-479.
AXELROD, D. I., 1967, Quaternary extinctions of large mammals: Univ. of California Pub. Geol. Science, v. 74, p. 1-42. -, & BAILEY, H. P., 1968, Cretaceous dinosaur ex- tinction: Evolution, v. 22, p. 595-609.
BROWN, W. L., & WILSON, E. O., 1956, Character displacement: Sys. Zoology, v. 5, p. 49-64.
CONNELL, J. H., & ORIAS, EDUARDO, 1964, The eco- logical regulation of species diversity: Am. Natu- ralist, v. 98, p. 339414.
DAVIDSON, THOMAS, 1886-1888, A monograph of Re- cent Brachiopoda: Linnean Soc. London, Trans., ser. 2, v. 4 (Zool.), p. 1-248, Pls. 1-30.
DEMPSTER, E. R., 1955, Maintenance of genetic het- erogeneity: Cold Spring Harbor Symp. Quant. Biology, v. 20, p. 25-32.
DORMAN, F. H., & GILL, E. D., 1959, Oxygen isotope paleotemperature measurements on Australian fos- sils: Roy. Soc. Victoria, Proc., new ser., v. 71, p. 73-98.
DURHAM, J. W., 1950, Cenozoic marine climates of the Pacific Coast: Geol. Soc. America, Bull., v. 61, p. 1243-1264.
EMILIANI, CESARE, 1956, On paleotemperatures of Pacific bottom waters: Science, v. 123, p. 460-461. - , 1961, The temperature decrease of surface sea-water in high latitudes and of abyssal-hadal water in open oceanic basins during the past 75 million years: Deep-Sea Research, v. 8, p. 144-147.
FILATOVA, Z. A., 1957, General review of the bivalve mollusks in the northern seas of the U.S.S.R.: Trans. Internat. Oceanol., v. 20, p. 1-44 [in Rus- sian].
FISHER, A. G., 1960, Latitudinal variations in organic diversity: Evolution, v. 14, p. 64-81.
GUNTER, GORDON, 1947, Paleoecological import of certain relationships of marine animals to salinity: Jour. Paleontology, v. 21, p. 77-79.
HESSLER, R. R., & SANDERS, H. L., 1967, Faunal di- versity in the deep-sea: Deep-Sea Research, v. 14, p. 65-79.
HUTCHINSON, G. E., 1957, Concluding remarks: Cold Spring Harbor Symp. Quant. Biology, v. 22, p. 415-427.
--, 1967, A treatise on limnology, v. 2, Introduction to lake biology and the limnoplankton: New York, John Wiley & Sons, Inc., xi + 1115 p.
KLOPFER, P. H., 1959, Environmental determinants of faunal diversity: Am. Naturalist, v. 93, p. 337-342. -, 1962, Behavioral aspects of ecology: Engle- wood Cliffs, New Jersey, Prentice-Hall, Inc., 161 p.
,& MACARTHUR, R. H., 1961, On the causes of tropical species diversity: niche overlap: Am. Naturalist, v. 95, p. 223-226.
KOHN, A. J., 1966, Food specialization in Conus in Hawaii and California: Ecology, v. 47, p. 1041-1043.
LEVINS, R., 1962, Theory of fitness in a heteroge- neous environment. II. Developmental flexibility and niche selection: Am. Naturalist, v. 97, p. 75-90.
914
This content downloaded from 128.233.210.97 on Fri, 20 Sep 2013 08:43:23 AMAll use subject to JSTOR Terms and Conditions

NICHE DIVERSITY AND NICHE SIZE PATTERNS
LUDWIG, W., 1950, Zur Theorie der Konkurrenz: N. Engeb. Zoologie, Klatt-Festschrift, p. 516-537.
MACARTHUR, R. H., 1965, Patterns of species di- versity: Biol. Reviews, v. 40, p. 510-533.
, & LEVINS, R., 1964, Competition, habitat se- lection and character displacement in a patchy en- vironment: Nat. Acad. Science, Proc., v. 51, p. 1207-1210.
- , & - , 1967, The limiting similarity, conver- gence, and divergence of coexisting species: Am. Naturalist, v. 101, p. 377-385.
MAYR, ERNST, 1942, Systematics and the origin of species: New York, Columbia Univ. Press, xiv + 334 p.
1963, Animal species and evolution: Cambridge, Harvard Univ. Press, iv + 797 p.
MILLER, R. S., 1967, Pattern and process in competi- tion, in CRAGG J. B., editor, Advances in ecological research, v. 4: New York, Academic Press, p. 1-74.
NEWELL, N. D.; RIGBY, J. K.; FISCHER, A. G.; WHITEMAN, A. J.; HICKOX, J. E.; & BRADLEY, J. S., 1953, The Permian reef complex of the Guada- lupe Mountains Region, Texas and New Mexico: San Francisco, W. H. Freeman & Co., xix + 236 p., 32 Pls.
NICOL, DAVID, 1967, Some characteristics of cold-wa- ter marine pelecypods: Jour. Paleontology, v. 41, p. 1330-1340.
PIANKA, E. R., 1966, Latitudinal gradients in species diversity: a review of concepts: Am. Naturalist, v. 100, p. 33-46.
PROSSER, C. L., & BROWN, F. A., 1961, Comparative animal physiology: Philadelphia, W. B. Saunders, ix + 688 p.
RUDWICK, M. J. S., 1960, The feeding mechanisms of spire-bearing fossil brachiopods: Geol. Mag., v. 97, p. 369-383.
-, 1962, Filter-feeding mechanisms in some brach- iopods from New Zealand: Linnean Soc. London, Jour., v. 44, no. 300, p. 592-615.
,- & COWEN, RICHARD, 1968, The functional mor- phology of some Permian brachiopods from the
Sosio Valley, Sicily: Soc. Paleont. Ital., Boll., v. 6, p. 113-176, Pls. 3243.
RUZHENTSEV, V. E., & SARYCHEVA, T. G., editors, 1965, Development and change of marine organisms at the Paleozoic and Mesozoic boundary: Akad. Nauk SSSR, Trudy, v. 108, 431 p., 58 Pls. [in Rus- sian].
SIMPSON, G. G., 1944, Tempo and mode in evolution: New York, Columbia Univ. Press, xviii + 237 p.
- , 1953, The major features of evolution: New York, Columbia Univ. Press, xx + 434 p.
STANLEY, S. M., 1968, Post-Paleozoic adaptive radia- tion of infaunal bivalve molluscs-a consequence of mantle fusion and siphon formation: Jour. Paleon- tology, v. 42, p. 214-229.
STEBBINS, G. L., JR., 1950, Variation and evolution in plants: New York, Columbia Univ. Press, 643 p.
VALENTINE, J. W., 1967, Influence of climatic fluctua- tion on species diversity within the Tethyan pro- vincial system, in ADAMS, C. G., & AGER, D. V., editors, Aspects of Tethyan biogeography: Syste- matics Assoc., Pub. 7, p. 153-166.
--, 1968a, Climatic regulation of species diversifica- tion and extinction: Geol. Soc. America, Bull., v. 79, p. 273-275.
-, 1968b, The evolution of ecological units above the population level: Jour. Paleontology, v. 42, p. 253-267.
VAN VALEN, LEIGH, 1965, Morphological variation and width of ecological niche: Am. Naturalist, v. 99, p. 377-390.
WILLIAMS, ALWYN, 1965, Stratigraphic distribution: Treatise on Invertebrate Paleontology, R. C. MOORE, editor, Part H, Brachiopoda, p. 237-250, Lawrence, Kansas, Univ. Kansas Press and Geol. Soc. America.
WILLIAMS, G. C., 1966, Adaptation and natural selec- tion, a critique of some current evolutionary thought: Princeton, Princeton Univ. Press, x + 307 p.
MANUSCRIPT RECEIVED MARCH 4, 1968.
915
This content downloaded from 128.233.210.97 on Fri, 20 Sep 2013 08:43:23 AMAll use subject to JSTOR Terms and Conditions