Upload
others
View
3
Download
0
Embed Size (px)
Citation preview
Manuscript submitted to: Volume 1, Issue 1, 96-119.
AIMS Neuroscience DOI: 10.3934/Neuroscience.2014.1.96
Received date 7 May 2014, Accepted date 30 May 2014, Published date 20 June 2014
Review article
The Cerebellum in Emotional Processing: Evidence from Human and
Non-Human Animals
Wanda M. Snow 1, 2,*, Brenda M. Stoesz 3 and Judy E. Anderson 4
1 Division of Neurodegenerative Disorders, St. Boniface Hospital Research, R4050-351 Tache
Avenue, Winnipeg, Manitoba, Canada R2H 2A6 2 Department of Pharmacology & Therapeutics, A203 Chown Bldg., 753 McDermot Avenue,
University of Manitoba, Winnipeg, Manitoba, Canada R3E 0T6 3 Department of Psychology, P404 Duff Roblin Bldg, 190 Dysart Rd, University of Manitoba,
Winnipeg, Manitoba, Canada R3T 2N2 4 Department of Biological Sciences, Biological Sciences Building, Faculty of Science, 50 Sifton
Rd., University of Manitoba, Winnipeg, Manitoba, Canada R3T 2N2
* Correspondence: Email: [email protected]; Tel: +1-204-235-3615.
Abstract: The notion that the cerebellum is a central regulator of motor function is undisputed. There exists, however, considerable literature to document a similarly vital role for the cerebellum in the regulation of various non-motor domains, including emotion. Research from numerous avenues of investigation (i.e., neurophysiological, behavioural, electrophysiological, imagining, lesion, and clinical studies) have documented the importance of the cerebellum, in particular, the vermis, in affective processing that appears preserved across species. The cerebellum possesses a distinct laminar arrangement and highly organized neuronal circuitry. Moreover, the cerebellum forms reciprocal connections with several brain regions implicated in diverse functional domains, including motor, sensory, and emotional processing. It has been argued that these unique neuroanatomical features afford the cerebellum with the capacity to integrate information about an organism, its environment, and its place within the environment such that it can respond in an appropriate, coordinated fashion, with such theories extending to the regulation of emotion. This review puts our current understanding of the cerebellum and its role in behaviour in historical perspective, presents an overview of the neuroanatomical and functional organization of the cerebellum, and reviews the literature describing the involvement of the cerebellum in emotional regulation in both humans and non-human animals. In summary, this review discusses the importance of the functional connectivity of the cerebellum with various brain regions in the ability of the cerebellum to effectively regulate emotional behaviour.
97
AIMS Neuroscience Volume 1, Issue 1, 96-119.
Keywords: cerebellum; emotion; non-motor; vermis; fear conditioning
1. Introduction
Our ability to navigate and adapt within our environment relies heavily on the ability to execute the correct movement at the correct time and in the correct sequence in response to sensory and proprioceptive information. A neural substrate important for these functions is the cerebellum. Latin for “little brain”, the cerebellum accounts for only a small portion of the brain’s total volume yet encases more neurons than are housed in the entire remaining human brain [1]. Its putative role in motor behaviours was first noted over 200 years ago. Advances in technical abilities related to studying neurons within the cerebellum, their circuitry, pathways, and electrophysiological properties have vastly improved our understanding of this substrate and its contribution to motor behaviour. Decades of research have confirmed the importance of the cerebellum and its principal neurons, Purkinje neurons (PN), in the refinement of motor behaviours and precision of motor execution. The accumulation of considerable data, however, has prompted a re-evaluation and expansion of this role such that the cerebellum may be viewed as an information processor capable of coordinating and optimizing domains outside of motor function, including emotion.
The present report will begin by offering a brief historical perspective on the scientific inquiry regarding the function of the cerebellum in behaviour that led to the view of the cerebellum as being an integral component of motor functioning, including the acquisition, expression, and learning of motor behaviors. As the unique neuroanatomical organization of the cerebellum was and continues to be instrumental in offering clues to its contribution to behavior, its functional organization and neuronal circuitry will be highlighted. Our current understanding of the distinct contributions of the cerebellum in emotional regulation in both humans and non-human animals will be summarized. Lastly, the view that it is the functional connectivity of the cerebellum with other brain areas that confers its ability to modulate a broad range of capabilities will be also discussed.
2. Historical Perspective of Cerebellar Involvement in Motor Behaviour
The first documented experimental evidence for a putative role of the cerebellum in motor functioning came from Rolando in 1809, who noted that cerebellar lesions resulted in motor deficits but preservation of sensory processing abilities [2]. In 1824, Fluorens astutely observed that animals with cerebellar lesions were not paralyzed and contended that coordination rather than production of movement was influenced by the cerebellum [2].
Advances in microscopy during the middle of the 1800s enabled investigators to visualize cellular features of tissue in a way that was not previously possible. Such advances led to the morphological description of PN in unstained tissue by Purkinje in 1837, for whom these cells were named [2]. Purkinje described them as “corpuscles” and noted their architecturally orderly arrangement throughout the cerebellum. The extensive dendritic arborizations of PNs were not evident, however, as they are not readily obvious in unstained tissue.
It was another 50 years before the unique morphological structure of PNs could be appreciated in more detail. One of the most important experimental techniques in neuroscience, still used routinely today, is the silver impregnation method developed by Golgi in 1873 [2]. The exquisite utility of the method lies in the fact that only a small percentage of neurons is stained in their entirety,
98
AIMS Neuroscience Volume 1, Issue 1, 96-119.
including soma and processes [3]. Thus, the stained neurons are typically observed as single cells, and their processes can be observed in detail without overlapping by other stained cellular processes. Development of this staining procedure (initially from an artifact of histochemical staining), coupled with further advancements in light microscopy, allowed for individual neurons to be visualized with a degree of precision and intensity previously unparalleled. Cajal applied the Golgi method extensively in the late 1880s; his anatomical investigations of multiple brain regions display detailed illustrations. Development of the Golgi technique and its application by Cajal allowed for the characterization of the extensive dendritic arborisation of cerebellar PNs for the first time.
In 1891, Luciani applied fine-surgical techniques to investigate cerebellar functioning in vivo, and elaborated on the earlier work of Fluorens who had used animal experimentation [2]. Luciani described three features of cerebellar damage associated with various aspects of muscle control: 1) asthenia, or muscle weakness; 2) atonia, or a lack of typical muscle tone; 3) and astasia, or the discontinuity of movement, including tremors. This triad of deficits after cerebellar damage was confirmed in humans by Holmes in 1917 in his clinical investigation of World War I victims of gunshot injuries to the brain [4].
Progress in describing the cerebellar circuitry continued in the 20th century. Refinement of electrophysiological techniques allowed a group of collaborators in the 1960s to detail the elaborate cerebellar neuronal circuitry. Based on their findings, the group published a seminal work on the cerebellum, viewing it as a neuronal processing machine based on its intricate circuitry [5]. This monograph provided a jumping-off point for the development of numerous other concepts over the next several decades, in attempts to explain how the unique neuronal circuitry in the cerebellum produces and influences motor behaviour [6–8]. Seminal papers by Leiner and colleagues [9], early studies in clinical populations with cerebellar pathology [8,10], and early neuroimaging studies demonstrating cerebellar activation in tasks with no motor component [11], however, prompted a re-evaluation of the cerebellum as strictly delegated to motor control.
3. Cerebellar Neuroanatomy & Neurocircuitry
The cerebellum houses nearly half of the neurons within the brain [1] and receives input from
multiple regions of the central nervous system. The sources of those inputs subserve a broad array of functions and, in turn, the cerebellar neurons project to nearly all portions of the motor system [8] as well as non-motor areas of the cerebral cortex [12]. The cerebellum consists of a multineuronal cortical layer, white matter projecting from this layer, and the deep cerebellar nuclei (DCN), which are clusters of neurons onto which Purkinje axons synapse. Anatomically, the cerebellum is divided into the flocculonodular lobe, vermis, and lateral cerebellar hemispheres, and functional divisions have been identified based, in part, on these anatomical distinctions [13]. The flocculonodular lobe lies on the ventral side of the cerebellum and is the only cerebellar area to bypass the DCN in its projections to other areas. It receives information on balance and equilibrium via projections from the nuclei of the vestibular nerve in the brainstem and visual information conveyed from the visual cortex via projections from other nuclei. The projections of the flocculonodular lobe terminate back onto vestibular nuclei within the brainstem, forming a processing loop [13]. This region, the phylogenetically oldest region of the cerebellum, is referred to as the “vestibulocerebellum” due to its pivotal role in processing vestibular information [13,14].
The vermal cerebellum lies along the midline of the cerebellum and receives multimodal sensory input from the trunk and proximal muscles via spinocerebellar tracts. Projections from the vermis exit the cerebellum via the fastigial DCN en route to the brainstem and cortical areas involved
99
AIMS Neuroscience Volume 1, Issue 1, 96-119.
in controlling proximal regions of the body, including medial descending pathways [13]. Lateral to the vermal region on either side are the intermediate zones. Although not distinct on gross inspection of the cerebellar surface, the intermediate zone receives distinct somatosensory input about distal muscles from spinocerebellar tracts and from the pontine nuclei. In turn, this zone projects to the interposed DCN, the projections of which enter the descending lateral corticospinal and rubrospinal tracts that coordinate function of the body musculature [13]. Collectively, vermal and intermediate zones are coined “spinocerebellum”, as they constitute the only cerebellar regions to receive spinal cord input [13,14]. With regard to emotional regulation in humans, because of research linking specifically the vermis (discussed below), this area has been referred to as the “limbic cerebellum” [15].
A third functional subdivision in the cerebellum lies adjacent to the intermediate zones in the most lateral portion of the cerebellar hemispheres. This area, found bilaterally, receives input solely from the cerebral cortex [13], including motor, premotor, prefrontal, and parietal areas (as reviewed in [12]). These cortical inputs travel through the pontine nuclei en route to the cerebellum. Information processed within the lateral cerebellum projects back to motor, premotor, prefrontal, and parietal areas via the dentate DCN to thalamic and red nuclei, thus forming a cerebrocerebellar loop [12,16]. As the lateral cerebellar hemispheres receive input exclusively from the cerebral cortex, they are referred to as the “cerebrocerebellum”, which, phylogenetically, is the newest part of the cerebellum [13,14].
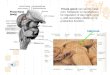
At the cellular level, the cerebellar cortex (Figure 1) consists of three cellular layers comprised of five neuronal types (as reviewed in [17]). The granule cell layer is the deepest cortical layer and is comprised of vast numbers of granule cells, Golgi interneurons, and axons from various inputs, depending on the functional subdivision, collectively referred to as mossy fibres (MF). A second neuronal layer consists of a monolayer of PN bodies (somata). The outermost layer, the molecular layer, is composed of dendrites of PNs, basket and stellate cells, and granule cell axons. The latter are termed parallel fibres due to their arrangement parallel to the parasagittal plane of PN dendrites.
This cortical neuronal arrangement is preserved across the subdivisions of the cerebellum (i.e. flocculonodular lobe, vermal and lateral regions); the only difference among the regions is the source of the MF input as well as the DCN through which the processed signal ultimately leaves the cerebellum [18]. Excitatory influences onto PNs arise indirectly from the MFs from three sources (i.e. from vestibular nuclei in the vestibulocerebellum or flocculonodular lobe; from the spinal cord via spinocerebellar tracts and reticular formation in the spinocerebellum or vermal and intermediate regions; and from cerebral cortex via the pontine nuclei in the cerebrocerebellum or lateral region). MFs form excitatory synaptic connections with granule cell dendrites, the axons of which extend into the molecular layer in the form of parallel fibres that form excitatory synapses with PNs. The excitatory influence of MFs is modified by Golgi interneurons in the granule layer. Axons of Golgi cells form inhibitory synaptic contacts with MF axon terminals and granule cell dendrites in synaptic clusters called cerebellar glomeruli [17]. The interconnections suppress the excitatory effects of parallel fibres onto PNs. The only other source of excitatory projections to PNs arises from inferior olivary nuclei in the medulla. The long axons projecting from the inferior olive, termed climbing fibres (CF), wind around PN somata and their dendrites, and synapse with multiple PNs [19]. Basket and stellate cells within the molecular layer exert an inhibitory influence onto PNs. Thus, PNs are subject to inhibitory influences from cerebellar cortical circuits that collectively modulate the excitatory influences from projections originating outside the cerebellum [17].
100
AIMS Neuroscience Volume 1, Issue 1, 96-119.
Figure 1. A schematic representation of the internal neuronal circuitry of the
cerebellum. + = excitatory synapse; – = inhibitory synapse; PF = parallel fibre; BC
= basket cell; PN = Purkinje neuron; GC = granule cell; GO = Golgi cell; SC =
stellate cell; CF = climbing fibre; DCN = deep cerebellar nuclei; MF = mossy fibre.
After integrating multiple inhibitory and excitatory synapses, PNs send GABA-ergic inhibitory projections to the DCN [19] (i.e., the fastigial nucleus in the vermal region, interposed in the intermediate portion, and the dentate nucleus in the lateral hemispheres). Fibres from the DCN project to many regions of the brain, including vestibular nuclei, medial and lateral descending motor pathways, and cerebral cortical association areas [13]. In addition to the inhibitory signals from cerebellar PNs, neurons in the DCN also receive direct excitatory projections from the two main inputs to the cerebellum, MFs and CFs, via axon collaterals. This direct excitation of the DCN is mediated and dampened by the inhibitory input from PNs, the central processors that receive a barrage of both excitatory incoming signals from outside the cerebellum and inhibitory signals from interneurons within the cerebellar cortex. Therefore, PNs play a key role in modulating and inhibiting the excitatory influence of the DCN and their broad distribution of terminations in multiple brain regions.
4. Evidence of the Cerebellum and Non-motor Domains in Humans-Relevance to Error
Correction and Timing
Although the importance of the cerebellum to motor function is uncontested, precisely how the cerebellum achieves this has been the source of considerable debate. Studies using experimental paradigms with clinical populations have elucidated roles for the cerebellum in both the correction of errors during motor learning and in ensuring accurate timing of both motor and perceptual tasks. In a
101
AIMS Neuroscience Volume 1, Issue 1, 96-119.
now classic experiment, Thach et al. [8] examined response patterns to throwing a dart at a target before, during, and after visual displacement of the target with prism glasses among individuals with a degenerative disease affecting the inferior olive, a source of excitatory input to cerebellar PNs. An individual’s fixation on a target provides a point of aim for the arm throw. When the subject wears prism glasses that bend the optical path to the right, individuals typically throw the dart to the left of the target; this is in response to the person adjusting his/her gaze to the left to compensate for the bending of the optic path, as the arm throw follows the direction of gaze. With repeated trials, throws become closer to the target. After removal of the glasses, adjustments made to compensate for the optical displacement persist, despite the gaze now being in-line; as a result, after removing the prism from the optical path, the throws are to the right of the target. Again after practice [this time without the prism glasses], errors in aim are reduced, and throws are on target. Individuals with disease affecting the excitatory input to cerebellar PNs fail to adjust their aim in response to the initial optical displacement with glasses on, and the target is repeatedly to the left, with little improvement in reducing the distance from the target with practice. Upon removal of the glasses, throws from those with cerebellar damage are not far to the right of the target, as is the case for typical individuals, suggesting that the initial adjustments for the prism were never made, and explaining the failure to correct for the visual displacement.
This reduction in error adaptation implicates the cerebellum in providing error correction during the execution of movement. Anatomically, its massive inputs from huge numbers of granule cells are capable of conveying vast amounts of information, and this provides the cerebellum with the neuronal machinery to create a fine representation of incoming information (and adjust to it in near-real time), a feature required for moment-to-moment fine-tuning of motor output [6–8].
The cerebellum has also been implicated in patterning the timing of both motor and perceptual tasks. Keele and Ivry [10] examined the motor and perceptual timing abilities of patients with neurological conditions, including cerebellar pathology, cortical pathology, and Parkinson’s disease compared to typical controls. In the motor component, subjects tapped their fingers in time to an auditory tone. After the pacing tone was turned off, subjects continued tapping for a specified time, and data were collected regarding the inter-tap variability across trials. In this test, patients with both cerebral and cerebellar pathologies had increased variability in the duration of the inter-tap interval compared to typical controls. During the perceptual component, subjects were presented with two pairs of tones and asked to indicate which one of the two pairs had a longer interval between them. The first pair of tones presented was of a constant duration, and the interval between the second pair of tones was either longer or shorter in duration. The patients with cerebellar and cortical pathology were impaired on tests of motor timing compared to both typical control subjects and those with Parkinson’s disease, ruling out the effect of the diseased basal ganglia to decrease timing abilities. Only patients with cerebellar damage were impaired on the perceptual timing task and the motor timing task. Although the increased variability of motor taps could be due to motor impairments in both cortical and cerebellar patient groups, only cerebellar patients were impaired in both timing assessments. To rule out decreased auditory processing abilities as an explanation for the increased variability in the perceptual component, a third task was included in which subjects were presented with pairs of tones as before but were asked to judge which tones were louder. Only cortical patients were impaired on this additional task, suggesting that the altered perceptual timing seen in cerebellar patients was not related to a deficit in general auditory processing or hearing impairments. Based on such findings, the authors hypothesized that the cerebellum attends to and provides temporal information regarding not only the execution of movement but also sensory perception.
102
AIMS Neuroscience Volume 1, Issue 1, 96-119.
5.1. The Cerebellum and Emotional Processing in Humans
The aforementioned studies by Thach et al. [8] and Keele and Ivry [10] were among the first to scientifically evaluate the presence of deficits in non-motor functions after cerebellar damage in humans. In addition to studies demonstrating the importance of the cerebellum in perceptual timing, recent advances in functional brain imaging have provided further evidence for the putative role of the cerebellum in non-motor functions. These data are changing the classic view of the cerebellum as a substrate exclusively and intricately involved in motor behaviour, including motor coordination and motor learning; it is now considered to have a broad role in human behaviour including mediation of cognitive processing [20].
Proposed by Leiner, Leiner, and Dow in the 1980’s [9], the cerebellum’s putative role in higher order cognitive functions has since garnered a great deal of interest. Experimental evidence supports cerebellar involvement in mental activity, including the generation of appropriate verbs when presented with nouns [11], silent counting [21], mental imagery [22] and completion of a pegboard puzzle, with cerebellar activation significantly higher than with simple peg movements [23].
Simultaneous to research identifying cerebellar activation associated with cognitive processing, its role in emotional processing was beginning to be appreciated. In humans, data supporting the notion of the cerebellum as a mediator of emotion stems primarily from multiple lines of research, including: 1) studies of patients with cerebellar lesions and, in some cases, resulting clinical syndromes; 2) experimental paradigms to assess associative fear learning; 3) neuroimaging and neurostimulation studies; 4) the emotional processing of human faces, organically rich in emotional context; and 5) studies in neurodevelopmental and psychiatric disorders.
5.2. Lesion Studies & Clinical Syndromes
A pioneering investigation of cerebellar regulation of emotional behaviour was conducted by Schmahmann and Sherman [24]. These authors conducted exhaustive neuropsychological testing in conjunction with bedside mental state examinations and anatomical imaging in adult patients with pathology localized to the cerebellum (i.e., from stroke, post-infectious cerebellitis, cerebellar cortical atrophy, and midline tumor resection). Based on clinical and neuropsychological data, they described a cluster of disturbances spanning numerous aspects of functioning, characterized by deficits in executive function (i.e., working memory, planning, set-shifting), visual-spatial domains (i.e., processing and memory), language production (i.e., anomia, agrammatism), and behavioural-emotional alterations (i.e., behavioural disinhibition, blunted affect). They coined this clinical syndrome “cerebellar cognitive affective disorder” (CCAS) and argued that these deficits could not be a result of cerebellar-associated motor deficits alone. Further, they highlighted the fact that symptoms typically associated with cortical damage, including aphasia and agnosia, were devoid in these patients. Although the heterogeneity of pathological region and small sample size precluded definitive association of cerebellar region to symptomology, these authors noted more severe emotional dysregulation with lesions specifically affecting the vermis [24].
Other studies have examined the consequences of pathology as a function of cerebellar region. In a study of those with vascular lesions affecting the cerebellum, lesions within the posterior region due to infarction of the posterior inferior cerebellar artery were associated with deficits in executive function, memory, and emotional withdrawal, whereas infarctions in the region of the superior cerebellar artery region were not [25].
Further support of cerebellar involvement in emotional regulation comes from studies
103
AIMS Neuroscience Volume 1, Issue 1, 96-119.
describing a collection of symptoms that are associated with pathology affecting the posterior fossa. Similar to CCAS, posterior fossa syndrome (PFS) is a clinical syndrome typically affecting children and adolescents upon resection of cerebellar tumors [26], with symptoms generally not appearing until hours or days post-operatively [26]. The syndrome is characterized by symptoms associated with diverse domains, including language and speech production (i.e., cerebellar mutism, dysarthria) as well as behavioural-emotional control. Transient mutism is generally considered a cardinal feature of the syndrome, and, hence, it is often referred to as cerebellar mutism in the literature [27]. There are reports, however, of behavioural-emotional disturbances in the absence of mutism after tumor resection of the posterior fossa [28]. Given the temporary nature of the mutism, PFS has been considered a transient entity [26]. Many studies, nonetheless, have noted long-term neurocognitive and neurobehavioural sequelae, including specific deficits in executive function, memory, attention, spatial processing, as well as more general diminished intellectual functioning and academic achievement [29–31].
With respect to the affective symptoms of PFS, emotional lability has been described after resection of posterior fossa tumors in children [32]. Irritability, depressed affect, lack of initiation, apathy, disinterest, and withdrawal are also reported [29,32–34] . Severe mood changes were noted in over half of a group of children that underwent tumor resection involving the vermis [24]. Concurring with work in adults by this group [24], these findings provide confirmatory evidence of the role of the cerebellar vermis in emotional regulation in humans, as no such changes were reported in resections that did not involve the vermis in children with PFS [24]. In addition to its occurrence after cerebellar surgical resection, symptoms associated with PFS have also been documented as a consequence of infections [35,36], trauma [37,38], or vascular alterations [39] affecting the cerebellum.
Although more typical in children, where reported incidence rates range from 8% to 31% after tumor resection (as reviewed in [26]), PFS has been documented in adults after tumor resection. The syndrome is rare in adults, however, occurring in 1% of postoperative adults [40]. In a review of the literature, Marien et al. [26] found only 21 cases of the syndrome after cerebellar surgery in adults (reported from 1969 to 2011). In addition to classical symptom of mutism, these patients displayed behavioral-emotional dysregulation, including depressed mood [41], emotional lability [41,42], lethargy [43], add emotional aspontaneity [44]. PFS has also been confirmed in a small number of patients that underwent posterior fossa surgery to repair vascular pathology [42,45–47]. Recently, Marien et al. [48] reported PFS of purely vascular etiology (i.e., with no surgical intervention) in an adult after cerebellar stroke, with the patient displaying apathy and blunting of affect post-stroke.
5.3. Fear Conditioning and Emotional Learning
Fear is a strong emotion, and the capacity to form fear-related memories serves an evolutionary advantage in that associating fear to a relevant stimulus and encoding a memory trace of this relationship can aid an organism in survival upon future exposures to that stimulus. The regulation of fear and formation of fear memories is multifaceted, consisting of aspects of motor/autonomic (i.e., changes in blood pressure and heart rate, dilation of pupils), endocrine, and behavioural aspects (i.e. freezing behaviour, potentiated startle reflex) (as reviewed in [49]), in addition to the affective component. Not surprisingly, given its vast connections with multiple brain regions that subserve diverse domains, the cerebellum has been implicated in fear conditioning, whereby an organism learns to associate a neutral stimulus (referred to as the conditioned stimulus; CS) to a painful stimulus (referred to as the unconditioned stimulus; US) [50]. Maschke et al. [51] examined fear
104
AIMS Neuroscience Volume 1, Issue 1, 96-119.
conditioning in individuals with vermal cerebellar lesions after tumor resection using classical conditioning paradigms designed to elicit the conditioned bradycardic response, a fear-related slowing of the heart rate. Here, the presentation of an auditory tone (CS) was paired with the presentation of a painful stimulus, an electrical shock to the finger (US). After repeated pairings, the previously neutral CS, the tone, elicits fear-associated behaviours, including bradycardia, in healthy individuals. In those with cerebellar vermal lesions, however, heart rate did not decrease after the conditioning protocol [51].
In another study, Maschke et al. [52] also examined fear-related conditioning associated with potentiation of the startle response. In this experimental paradigm, a neutral stimulus (CS) is paired with an aversive stimuli (i.e., shock; US) such that the neutral stimulus alone elicits the fear response, as described above. A second phase occurs, whereby the now fear-eliciting CS is paired with another neutral stimulus (i.e., auditory tone). The potentiated amplitude of the response to the second neutral stimulus is considered demonstrative of the degree of conditioning that has occurred. Compared to healthy controls, those with cerebellar lesions restricted to the vermis showed decreased potentiation of the blink reflex [52]. In those with lesions to the lateral hemispheres, however, potentiation was within the normal range, although only two subjects with lateral hemisphere lesions were included [52].
Interestingly, the studies by Maschke demonstrated intact autonomic responses to the fear-inducing stimuli. Further, Turner et al. [53] noted that in patients with cerebellar stroke, both the evaluation of an image as emotional/fearful and the autonomic response to fear-inducing stimuli were intact. Thus, the ability to form an association between sensory and aversive stimuli in fear conditioning may rely on the cerebellar vermis. The brain, however, does seem to make compensatory changes, in some cases, in efforts to preserve the ability to form such critical fear associations via recruiting other neural circuits to a greater degree after cerebellar damage, as evidenced by increase activation of several brain regions after the presentation of fearful stimuli in cerebellar stroke patients relative to healthy controls [53].
5.4. Neuroimaging & Neurostimulation Studies
In addition to studies investigating symptomology of affect and cerebellar lesions, neuroimaging studies provide considerable evidence for the integral role of the cerebellum in emotional processing. In humans, electrical stimulation of the cerebellum can induce feelings of fear and anxiety [54]. Moreover, activation of the vermis was reported when individuals were asked to recall an emotional past memory versus a neutral one [55]. Studies also document localization of aspects of fear and fear memories within the vermis; pain-inducing heat results in activation of the anterior portion of the vermis, whereas a sensory anticipatory cue of the painful stimulus activates the posterior vermis [56,57]. Further, Singer et al. [58] also noted distinct activation patterns within the vermis in a functional magnetic resonance imaging (fMRI) study investigating the neural correlates of feelings of pain and of the empathy of pain. Here, the authors noted anterior activation with receipt of a painful stimulus, whereas the posterior vermis was activated upon witnessing pain in others. Unlike the majority of studies examining cerebellar regulation of emotion, which report contributions specifically by the vermis, Singer et al. [58] found that both conditions activated the lateral cerebellum.
Others also report lateral cerebellar activation in emotional processing. In a positron emission tomography (PET) evaluation of fear conditioning, activation was heightened in the left lateral cerebellum after pairing of shocks to videotape cues [59]. Frings et al. [60] reported similar findings,
105
AIMS Neuroscience Volume 1, Issue 1, 96-119.
with increased regional cerebellar blood flow in the left lateral cerebellum in a fear conditioning paradigm. Using a fear-conditioned potentiation protocol, however, elicited an increase in the regional cerebellar blood flow in the vermis [60]. These data point to separable roles for regions of the cerebellum in the associative learning that precedes the formation of a fear memory from its autonomic aspects as well as in the experience of pain vs. the empathy of pain in others. This topographical activation noted with various components of conditioned fear has led some to argue that the vermis is involved in the somatic and autonomic aspects of fear conditioning, whereas the lateral cerebellum contributes to the emotional aspects of the association [61].
In addition to the investigation of the neural substrates central to the processing of fear, studies have been devised to uncover the biological underpinnings of other basic emotions. Using fMRI, Baumann and Mattingley [62] reported emotion-specific topographical cerebellar activation in association with feelings of happiness, sadness, anger, fear, and disgust. To investigate the neural connectivity associated with localized cerebellar activation seen with various primary emotions, Schienle and Scharmuller [63] examined not only cerebellar activation patterns but also connectivity patterns involved in the regulation of happiness and disgust. Only female subjects were recruited, given this group’s prior research demonstrating increased elicitation of disgust in women relative to men [64]. In accordance with the previous study by Baumann and Mattingley [62], Schienle and Scharmuller [63] found localized activation as a function of the emotion elicited. In contrast to images evoking happiness, presentation of images evoking disgust, as rated by the participants, activated both the vermal and lateral cerebellum. Images evoking happiness activated crus II of the posterior lateral cerebellar hemispheres in contrast to disgust-evoking stimuli. Elicitation of either emotion enhanced connectivity with the canonical emotional centres of the brain that comprise the limbic system, including the amygdala. Feelings of disgust have been previously shown to activate the amygdala [64]. Stoodly and Schmahmann [65] recently summarized the results of nine neuroimaging studies reporting cerebellar activation involved in emotion processing. The results of this meta-analysis confirmed activation of left lobule VIIAt, lobules VI and Crus I of the left cerebellar hemisphere as well as right lobule VI in response to various emotional stimuli (e.g., emotion in a speaker’s voice, emotional pictures). These data further support the concept of regional localization of emotional processing within the cerebellum and extend this to emotion specificity.
5.5. Processing Emotional Expression in Faces
The idea that the cerebellum is involved in processing the emotional expressions in other people’s faces has only recently been acknowledged. This is an important discovery, as it may partially explain why some patients with degenerative cerebellar disorders or discrete ischemic cerebellar lesions show impairments in identifying the intentions of others correctly, and/or show difficulty responding appropriately in social situations [66].
Some research suggests that the cerebellum is involved in processing facial expression in general. For example, Fusar-Poli et al. [67] reported that the processing of several different emotional facial expressions (i.e., happy, angry, and fearful) were associated with increased activation in the cerebellum (declive), as well as subcortical areas (putamen), limbic areas (amygdala and parahippocampal gyrus, posterior cingulate cortex), temporoparietal areas (parietal lobule, middle temporal gyrus, insula), prefrontal areas (medial frontal gyrus), and a number of visual areas (fusiform gyrus, inferior and middle occipital gyri, lingual gyrus). These results confirm that the processing of emotion from facial expressions draws on diverse psychological processes implemented in a large array of neural structures (see also [68]). Moreover, no differential brain
106
AIMS Neuroscience Volume 1, Issue 1, 96-119.
activation in the cerebellum occurred across emotions, suggesting that the cerebellum plays a general role in processing expressions in faces.
Interestingly, other research suggests that the cerebellum is involved in the processing of particular expressions. Which facial expression is preferentially processed by the cerebellum, however, varies across studies. For example, one study reported that cerebellar transcranial magnetic stimulation (TMS), compared with occipital cortex and sham TMS, resulted in enhanced implicit processing of happy facial expressions with changes in self-reported mood [69]. Other studies report that cerebellar structures are more involved in recognizing and forwarding information about facial expression conveying negative emotions, such as fear or anger [53,70]. Adamaszek [71] reported that compared to controls, patients with discrete ischemic cerebellar lesions showed impaired selection and matching of emotional facial expressions, which was more pronounced for fear. Moreover, the larger the volume of lesion, the greater the deficit on the expression processing tasks. Together, these results confirm a significant role of the cerebellum in processing emotional recognition from faces, a component of social cognition.
It is important to note, however, that the differing results described above come from studies using static face stimuli. Emotional expressions in static face stimuli imply motion, but static stimuli do not contain the additional information, provided in the non-rigidly moving faces, that viewers process in the real world. Indeed, face processing regions in the occipital and parietal areas activate more for dynamic compared to static face stimuli [72]; thus, it is logical that the cerebellum would show different levels of activation during dynamic, as opposed to static, facial expression processing. Indeed, Kilts et al. [73] found that compared to judgments of spatial orientation of dynamic neutral expressions, judgment of anger intensity in dynamic expressions was associated with prominent right hemisphere-lateralized superior, medial, middle, and inferior frontal cortex and cerebellar activations. Further, the direct contrast of images acquired during anger recognition in dynamic versus static expressions was associated with activations in the right occipital–temporal junction corresponding to area V5 and in the periamygdaloid cortex, parahippocampal gyrus, STS, and cerebellum. Relative to happiness, anger perception was also associated with differential activation of the right amygdala and gyrus rectus, left fusiform gyrus, and cerebellum. Thus, there is evidence to suggest that the cerebellum plays an important role in processing naturally unfolding facial expressions of fear.
5.6. Neurodevelopmental & Psychiatric Disorders
Another line of evidence in support of a pivotal cerebellar role in emotional processing comes from studies of symptomology of psychiatric and neurodevelopmental disorders with emotional components, including depression, anxiety, obsessive-compulsive disorder, bipolar disorder, schizophrenia, fetal alcohol syndrome, and autism spectrum disorders [see [74] for review]. For example, in an MRI study, the volume of the cerebellar vermis, not the lateral hemispheres, was reduced in males and females with schizophrenia relative to psychiatrically well controls [75]. Others have confirmed the finding of reduced volume in the vermis in both adult populations [76] and in childhood-onset schizophrenia [77]. A neuroimaging study in males with schizophrenia, however, reported increased volume of the vermis, suggesting possible gender-mediated differences [78]. As chronic administration of antipsychotic medication used to treat the disorder may influence brain morphology, Ichimiya et al. [79] examined cerebellar volume in neuroleptic-naïve schizophrenics and also noted decreased vermal volume, ruling out the possible effects of antipsychotic medication as a confounder. A small number of studies, however, failed to detect differences in cerebellar volume in schizophrenia [80,81]; thus, the literature documenting volume reductions in the vermis in
107
AIMS Neuroscience Volume 1, Issue 1, 96-119.
schizophrenia is not unanimous. As early as the 1970s, chronic stimulation of the vermal cortex was investigated as a putative
treatment to alleviate emotional dysregulation in a heterogeneous group (n = 11) of patients with intractable psychiatric illness [81]. Prior to implantation, patients exhibited a range of symptoms, from severe aggression, suicidal and homicidal behaviour, to depression and auditory hallucinations. The results were striking: ten out of eleven patients showed improvements in symptomology and behaviour such that medications could be terminated and/or patients discharged. Interestingly, Heath [82] noted changes in affect as among the first reported after pacemaker implantation. Based on such early work, Demirtas-Tatlidede et al. [83] more recently applied non-invasive theta burst stimulation to the cerebellar vermis via MRI-guided TMS in patients with treatment-refractory schizophrenia. Although the follow-up period was minimal (one week), results were promising, as TMS was well-tolerated and was associated with significant elevation in mood and improvements in cognition, including working memory. Such results are particularly promising, given that symptoms associated with schizophrenia are often resistant to treatment, and current medications can yield significant aversive side effects (for example, see [84]).
In accordance with data regarding the cerebellum and schizophrenia, reduced cerebellar volume is also reported in depression [85]. Moreover, a meta-analysis demonstrated reduced activity in a network of brain regions, including frontal and temporal cortex, insula, and the cerebellum, with increased activation after antidepressant use [86]. In a study of psychiatric morbidity in patients with adult-onset progressive disorders that induce cerebellar degeneration (i.e., spinocerebellar ataxia or multisystem atrophy), the rate of psychiatric disorders was approximately two-fold higher in those with cerebellar disease relative to neurologically healthy controls and was accompanied by a significantly later onset than seen in controls [87]. Of the psychiatric conditions diagnosed, major depression occurred more frequently in this patient group in comparison to controls. In geriatric depression, Alalade et al. [88] reported altered connectivity between the posterior vermis and cingulate cortex that was positively correlated with depression severity. Such data strongly implicate the cerebellum in the pathophysiology of psychiatric disorders, including depression, across the lifespan.
The neurodevelopment disorders under the autism spectrum represent an etiologically diverse range of disorders with clinical heterogeneity. Autism spectrum disorder (ASD) is typically accompanied by impairments, to varying degrees, associated with social interaction and social reciprocity, nonverbal and verbal communication, and behavioural patterns, interests, and activities that are repetitive, restrictive, and stereotyped. In the first descriptions of the disorder, Kanner [89] acknowledged autism as having a significant emotional component. Depression, as well as increased fear and anxiety, are reported in ASD [90–92]. Further, in some cases, decreased motivation to engage emotionally with parents during the first year of life in those later diagnosed with ASD has been reported [93].
Although clinically heterogeneous, neuroanatomical abnormalities specifically of the cerebellum are a common, well-documented finding in ASD [94–101,99], having been found in ninety-five percent of autopsies of individuals with ASD [102]. Cerebellar abnormality in ASD patients includes cases of both hyperplasia, resulting in greater cell packing density, as well as hypoplasia [99]. The latter, however, is the dominant finding and is characterized by a reduction of Purkinje cells of the posterior vermis [99]. Cerebellar cortical atrophy, with a reduction in both Purkinje and granule cells, has been described [95].
Despite the consistency of findings regarding cerebellar morphology in ASD, contradiction exists regarding the direction of abnormality, as both hypoplasia and hyperplasia have been found
108
AIMS Neuroscience Volume 1, Issue 1, 96-119.
[99]. A reduction in the number of neurons within a given structure can be assumed to decrease the communicative capability of that structure and cause dysfunction. Greater neuron density and packing within a structure, however, may also contribute to impairment [103]. In line with this, Snow et al. [104] documented increased dendritic complexity of pyramidal neurons in the motor cortex of rats exposed to valproic acid in utero, a recognized animal model of the disorder.
The evidence for cerebellar pathology in ASD suggests that dysfunction of this structure is a critical feature of the disorder. A common behavioural symptom in ASD is a restriction in the range of interests, behaviours, or movements, and exploratory behaviour is consequently often absent or limited [100]. To assess the role of the cerebellum in exploratory behaviour, Pierce and Courchesne used MRI to document the degree of cerebellar hypoplasia of the vermis in ASD [100]. The same children were then rated in terms of time exploring a novel environment as measured by duration of exploration, number of containers explored, and motor activity. Children with ASD spent substantially less time in exploratory behaviour than control children, and this temporal difference was negatively correlated with the magnitude of hypoplasia. This differentiation was independent of overall motor activity, which did not significantly differ between the two groups.
Allen and Courchesne [102] used fMRI to observe cerebellar activation during a selective attention task and during a motor task and found that the cerebellar cortex of individuals with ASD showed decreased activation during the attention task but increased activation during the motor task when compared to healthy controls. These researchers previously demonstrated that regions of the cerebellum differentially respond to aspects of attention versus motor involvement [105], further confirming the topographical nature of the cerebellum. Moreover, the diversity of symptoms and their severity against the backdrop of consistent findings of cerebellar involvement in the pathology and functional activation patterns seen in ASD provide further evidence of significant motor, cognitive, and emotional-behaviour regulation that may be attributed to intact cerebellar function.
6 Cerebellar-Mediated Emotional Processing in Non-human Animals
In other species, the localization of circuitry within the cerebellum that subserves non-motor versus motor aspects of behaviour provide evidence that the cerebellum is associated with
higher-order information processing outside the parameters of pure motor behaviour. Localization of mental, non-motor activity separable from its motor components have been found within the cerebellum of non-human animals. For example, lesions to the lateral cerebellum in rats result in deficits in spatial learning that do not affect overt motor abilities [106–108] or impair the acquisition of new memories by observation [109]. Lesions to the vermal region, however, lead to impairments in visual guidance to the target platform [108].
In animals, much of what we have learned about the contribution of the cerebellum to emotional processing has been gained via examination of fear. Fear is a trans-species emotional state central to survival that is well-defined in many species, with corresponding autonomic alterations that can be monitored and measured. Early studies by Dow [110] provided evidence for involvement of the vermal region in emotional processing in animals, as stimulation of this region disrupted autonomic responses associated with fear, including prevention of increases in blood pressure and breathing, with no such changes with stimulation of the lateral hemispheres.
In addition to stimulation, lesions of the vermis can alter an organism’s affect and responses to fearful stimuli in several species. In cats, potentiated pleasure reactions have been reported after lesion to the vermal cerebellar cortex [111,112] including instances of purring and kneading, relative to sham-operated controls [113]. Further, blunting of responses to sudden stimuli and spontaneous
109
AIMS Neuroscience Volume 1, Issue 1, 96-119.
movement accompanied the lesions [113]. A similar taming effect and lack of spontaneous behaviour after similar lesions has also been reported in untamed squirrel monkeys [113]. In rhesus monkeys, vermal lesions result in docile behaviour and reduced aggression, whereas no such emotional-behavioural consequences were found after lesions to the lateral cerebellum [114]. Freezing behaviour, a defensive mechanism in rats in response to fearful or painful stimuli, diminished in the presence of a cat in rats with vermal lesions [115]. In addition to freezing behaviour, rats are known to exhibit specific behavioural responses in the presence of fear-inducing stimuli, such as thigmotaxis (wall hugging), considered to be an adaptive response in attempts to reduce possible predator attacks [116]. Vermal-lesioned rats also demonstrate reduced freezing behaviour and thigmotaxis in an open field test [115]. Such comparative data suggest an evolutionarily conserved role for the vermal cerebellum in regulating emotional behaviour.
As in human research, fear-conditioning paradigms have also been used extensively in non-human animals to investigate the cerebellum and emotional processing. Corroborating the significance of the vermis in this context, several studies report impaired acquisition of elements of the conditioned fear response after vermal lesions, including diminished bradycardia in rats [115,117] and rabbits [118]. Of note is the finding that lesioning did not affect baseline heart rate or heart rate in response to the tone alone [117]. These results support the role specifically of the vermis in associative fear learning.
Fear conditioning has also been examined in mice in which cerebellar dysfunction comes not from lesioning but from mutations affecting cerebellar synaptic integrity. In hotfoot mice, transmission at the PF-PN synapse is impaired due to an absence of the glutamate receptor delta2 subunit (GluRδ2) [119], which resides exclusively in spines of PN dendrites that receive PF innervation. In a comprehensive series of experiments, Sacchetti et al. [120] sought to investigate potential underlying physiological mechanisms by which the cerebellum contributes to emotional behaviour, specifically fear conditioning, using hotfoot mutants. GluR δ2-deficient hotfoot mice were not impaired in the acquisition of the learned fear association after conditioning, as freezing behaviour was similar in mutant and control mice. When tested 10-minute and 24-hours post-training, however, hotfoot mice demonstrated reduced freezing behaviour in response to the tone presentation relative to controls, indicative of impaired fear memory consolidation. This could not be explained by an overall increased state of anxiety-related arousal or spontaneous fear level, as no differences were reported between hotfoot mice and controls in the open field or light-dark test, both well-characterized measures of anxiety in rodent. Moreover, these authors noted a heightened long-term potentiation (LTP) effect in PNs after stimulation of PFs in fear-conditioned rats relative to either naïve untrained rats or unpaired controls (received tone and shock in unpaired manner) [120]. Such data confirm the importance of the learned association underlying fear memory formation in the physiological response, as no such potentiation was noted in animals that received equivalent exposure to the aversive stimuli. The LTP response was only observed in lobules V and VI, known sites of convergence for both acoustic and nociceptive inputs [121], and were notably absent in lobules IX and X, a region previous reported [118] as uninvolved in the regulation of aversive behavior. Given this region-specific convergence, these authors theorize that LTP response seen after associative fear learning may represent concomitant activation of the separate PF inputs from the CS and US. Further, this learning-induced potentiation was restricted to PF inputs, as no changes in synaptic transmission were noted at CF-PN synapse after fear conditioning Sacchetti et al. [120]. Therefore, transmission at the PF-PN synapse appears to play a central role in the consolidation of emotional memory formation regulated by the cerebellum, with plasticity at this site constituting a putative physiological mechanism by which such regulation may occur.
110
AIMS Neuroscience Volume 1, Issue 1, 96-119.
Fear conditioning under cases of reversible inactivation of the cerebellum has also been investigated as a means of uncovering how the cerebellum contributes to fear memories. The experimental setup includes administration of drugs with reversible effects (i.e., tetrodotoxin, a Na+-channel blocker) after acquisition of a fear-conditioned response. Retention of the fear memory is then examined at a point when the effects of the drug have passed. This allows one to dissociate the effects of the drug on sensory and motor functions that could be attributed to the cerebellum from those related to memory consolidation [122,123]. Using this technique, Sacchetti et al. [122] found amnesic effects with injection of tetrodotoxin into the cerebellar vermis 72 and 96 hours after the acquisition of a fear memory, with decreased freezing behaviour in rats up to 192 hours after TTX administration in response to presentation of the conditioned tone. The consolidation of contextual fear conditioning, in which the animal forms an association to the experimental environment in which the pairing takes place, was impaired up to 96 hours, as evidence by decreased freezing behaviour.
In addition to work in mammals, research investigating the contribution of the cerebellum to aspects of emotional regulation in non-mammal vertebrates has been conducted. Such studies have relevance to a general understanding of the impact imparted by the cerebellum in affective processing, as although there exists considerable heterogeneity in terms of relative size, all vertebrates studied share comparable and consistent cytoarchitecture and neuronal circuitry [124]. In goldfish, cerebellar lesions prevent the classical conditioning of an eye-retraction response in response to light after repeated pairings of the visual stimulus with a shock [125], akin to eyeblink conditioning paradigms used in mammals, demonstrating a preserved role for the cerebellum in classical conditioning across diverse vertebrate species. Further, this lab confirmed a similarly conserved role for the cerebellum in emotional learning [125]. As has been reported in mammals, cerebellar lesions in goldfish disrupted the conditioned bradycardia response that accompanies CS-US pairings relative to sham-operated goldfish [125]. Importantly, alterations in cardiac activity were exclusive to the conditioned response in the lesioned fish, as baseline heart rate and heart rate to the CS alone were unaffected [125], again as the case found in mammalian studies [117,118].
To further elucidate putative mechanisms involved in cerebellar-mediated fear conditioning, Yoshida and Kondo [126] recorded activity of PNs in vivo from the goldfish corpus cerebelli, homologous to the mammalian vermis, throughout the acquisition phase of fear conditioning. In the rabbit, responses of vermal PNs to a neutral stimulus in untrained rabbits have been shown to differ from PN responses to the same stimulus after fear conditioning in trained rabbits [127]. Pre- and post-conditioning responses within the same animal, however, were not examined. In goldfish, PN firing rates were altered in response to the CS after pairing with the US in comparison to firing rates to the unpaired CS alone, demonstrating fear conditioned-induced changes in vivo in the same neurons [126]. Thus, in addition to analogous cytoarchitecture and microcircuitry, the cerebella of various species seem to share consistent neurophysiological mechanisms and a similar functional relevance to associative emotional learning.
7. Cerebellar connectivity is crucial to its ability to integrative emotion and other functions
Physiological, behavioural, clinical, and imaging data investigating the cerebellum and its contribution to various functional domains support the view that local circuits within the cerebellum are distinct both anatomically and functionally and that such circuits are not confined to motor domains. The intrinsic neural connectivity and anatomical connectedness of the cerebellum can provide insights into just how the cerebellum contributes to such a diverse range of functions (i.e.,
111
AIMS Neuroscience Volume 1, Issue 1, 96-119.
motor, emotional-behavioural, cognitive). For example, neuroanatomical studies have confirmed the presence of reciprocal connections between the cerebellum and numerous brain regions, including the frontal and prefrontal cortex [128–130] and the hypothalamus [131,132]. The cerebellum also possesses reciprocal connections with brainstem regions that are intricately associated with limbic regions [133]. Furthermore, vermal stimulation alters firing properties of neurons in limbic regions, including the hippocampus, amygdala, and septum [134,135], confirming functional connectivity between the cerebellum and brain regions central to emotional processing.
This reciprocity provides the cerebellum with the physiological capacity to integrate information from various brain regions, compute/process the information, then send the processed information back to the regions of interest for fine-tuning. This is consistent with a role for the cerebellum in optimizing an organism’s internal and external response to its environment, akin to a master regulatory structure for the integration of motor, emotional, and sensory information [136]. It stands to reason, then, that it is the connectivity with a specific area of the cerebellum that dictates its role in processing [137]; such theories are in line with relatively consistent neuronal circuitry and cytoarchitecture within the cerebellum. It must be noted, however, that distinctions do arise in synaptic transmission, electrophysiological, biochemical, and morphological properties of cerebellar neurons as a function of cerebellar region. For example, information processed through the vestibulocerebellum (i.e., flocconodular lobe) bypasses the DCN, unlike transmission in other cerebellar regions [4]. There exist regional neurochemical (i.e., [138]) and electrophysiological distinctions (i.e., intrinsic firing properties; [139]) between cerebellar neurons as well. Generally speaking, however, the microcircuitry and cytoarchitecture appear fairly preserved across cerebellar functional domains.
Given neuroimaging and clinical data coupled with the general neuronal homogeneity of the cerebellum, Stoodley and Schmahmann [140] postulated a topographical organization of the cerebellum based on functional domains, each corresponding to specific cerebellar regions; 1) sensorimotor cerebellum (includes anterior lobe and lobule VIII of posterior lobe); 2) cognitive cerebellum (includes lobules VI and VII of the posterior region); 3) limbic cerebellum (includes primarily the posterior vermis); and 4) vestibulocerebellum (include the flocculonodular lobule, lobule X), involved in equilibrium and balance. Although there is sufficient evidence in support of such a topographical representation, there are some discrepancies in the literature, as imaging studies have pointed to a role for both the vermis and the lateral cerebellar hemispheres in emotional processing [58]. In non-human animals, the contribution of the cerebellum to emotional regulation appears to be more restricted to the vermal region, however. Thus, the cerebellar regions devoted to emotional regulation in humans seems to be more expansive than in other species.
Irrespective of topographical region, the cerebellum is assumed to perform a similar computation on the incoming information [18]; this has been deemed the “universal cerebellar transform” [141]. The cerebellum is considered an information-processing “machine” [5,18]. Therefore, the breadth of function in which the cerebellum has been implicated is assumed to arise due to its interconnectedness with a multiplicity of brain regions, of which it forms part of a distributed neural network for a given domain or response [9,142,143]. As speculated approximately 200 years ago by Flourens, the cerebellum appears integral for the coordination of movement rather than the generation of movement [2]. For instance, cerebellar damage is not associated specifically with paralysis, as may occur with damage to primary motor cortex [143,144]. The organism can still execute a motor response but not in a coordinated manner. These findings are similar to research investigating non-motor capabilities of the cerebellum (i.e., cognition and emotion) in cases of disease or damage, whereby cerebellar pathology affects the ability to execute the desired response
112
AIMS Neuroscience Volume 1, Issue 1, 96-119.
(internal or external) in an optimal, coordinated fashion. The contribution of the cerebellum to this optimization with regard to emotional regulation has been considered worthy of deeming the cerebellum an “emotional pacemaker”, modulating an organism’s affective response such that it is appropriate to the context [135].
8. Conclusions
Although our understanding of cerebellar function and the involvement of the cerebellum in a multitude of abilities has expanded immensely over the last 200 years, there remains much to be discovered. Numerous imaging, anatomical, behavioural, electrophysiological, and clinical studies across several species have revealed a broader role for the cerebellum in overall information processing, including affective regulation. At the core of the ability of the cerebellum to regulate distinct and diverse aspects of an organism’s capabilities and internal states appears to be its high degree of interconnectedness with several brain regions, for which its cytoarchitecture is highly suited. At the cellular level, synaptic plasticity at the PF-PN synapse seems integral for appropriate emotional regulation. Although there is a general consensus that the cerebellum, regardless of region, processes information in a homogeneous manner, the precise computation performed by the cerebellum in this regard is still unclear. As such, there is great and growing interest in learning how this “little brain” and its exquisitely elegant circuitry contribute to emotional-behavioural regulation.
Conflict of Interest
The authors report no conflict of interest associated with this review.
References
1. Andersen BB, Korbo L, Pakkenberg B. (1992) A quantitative study of the human cerebellum with unbiased stereological techniques. J Comp Neurol 326(4): 549-60.
2. Glickstein M, Doron K. (2008) Cerebellum: Connections and functions. Cerebellum 7(4):589-94. 3. Gibb R, Kolb B. (1999) A method for vibratome sectioning of golgi-cox stained whole rat brain.
J Neurosci Methods 79(1): 1-4. 4. Ito M. (2002) Historical review of the significance of the cerebellum and the role of purkinje
cells in motor learning. Ann N Y Acad Sci 978: 273-88. 5. Eccles JC, Ito M, Szentágothai J. (1967) The Cerebellum as a Neuronal Machine. New York: NY
Springer-Verlag. 6. Marr D. (1960) A theory of cerebellar cortex. J Physiol 202(2): 437-70. 7. Albus JS. (1971) A theory of cerebellar function. Math Biosci 10: 25-61. 8. Thach WT, Goodkin HP, Keating JG. (1992) The cerebellum and the adaptive coordination of
movement. Ann Rev Neurosci 15: 403-42. 9. Leiner HC, Leiner AL, Dow RS. (1986) Does the cerebellum contribute to mental skills? Behav
Neurosci 100(4): 443-54. 10. Keele SW, Ivry R. (1990) Does the cerebellum provide a common computation for diverse tasks?
A timing hypothesis. Ann NY Acad Sci 608: 207-11. 11. Petersen SE, Fox PT, Posner MI, et al. (1989) Positron emission tomographic studies of the
processing of single words. J Cognitive Neurosci 1: 153-170. 12. Strick PL, Dum RP, Fiez JA. (2009) Cerebellum and nonmotor function. Annu Rev Neurosci 32:
113
AIMS Neuroscience Volume 1, Issue 1, 96-119.
413-34. 13. DOW RS. (1961) Some aspects of cerebellar physiology. J Neurosurg 18: 512-30. 14. Kandel ER, Schwartz JH, Jessell TM. (2000) Principles of neural science. 4Eds, New York:
McGraw-Hill, Health Professions Division, 1414 . 15. Berntson GG, Torello MW. (1982) The paleocerebellum and the integration of behavioural
function. Physiol Psychol 10: 2-12. 16. Middleton FA, Strick PL. (1997) Cerebellar output channels. Int Rev Neurobiol 41: 61-82. 17. Hashimoto M, Hibi M. (2012) Development and evolution of cerebellar neural circuits. Dev
Growth Differ 54(3): 373-89. 18. Ito M. (2006) Cerebellar circuitry as a neuronal machine. Prog Neurobiol 78(3-5): 272-303. 19. Eccles JC. (1967) Circuits in the cerebellar control of movement. Proc Natl Acad Sci USA
58(1): 336-43. 20. Akshoomoff NA, Courchesne E. (1992) A new role for the cerebellum in cognitive operations.
Behav Neurosci 106(5): 731-8. 21. Decety J, Sjoholm H, Ryding E, Stenberg G, Ingvar DH. (1990) The cerebellum participates in
mental activity: Tomographic measurements of regional cerebral blood flow. Brain Res 535(2): 313-7.
22. Ryding E, Decety J, Sjoholm H, et al. (1993) Motor imagery activates the cerebellum regionally. A SPECT rCBF study with 99mTc-HMPAO. Brain Res Cogn Brain Res 1(2): 94-9.
23. Kim SG, Ugurbil K, Strick PL. (1994) Activation of a cerebellar output nucleus during cognitive processing. Science 265(5174): 949-51.
24. Schmahmann JD, Sherman JC. (1998) The cerebellar cognitive affective syndrome. Brain 121: 561-79.
25. Exner C, Weniger G, Irle E. (2004) Cerebellar lesions in the PICA but not SCA territory impair cognition. Neurology 63(11): 2132-5.
26. Marien P, de Smet HJ, Wijgerde E, et al. (2013) Posterior fossa syndrome in adults: A new case and comprehensive survey of the literature. Cortex 49(1): 284-300.
27. Kuper M, Timmann D. (2013) Cerebellar mutism. Brain Lang 127(3): 327-33. 28. Baillieux H, De Smet HJ, Paquier PF, et al. (2008) Cerebellar neurocognition: Insights into the
bottom of the brain. Clin Neurol Neurosurg 110(8): 763-73. 29. Steinlin M, Imfeld S, Zulauf P, et al. (2003) Neuropsychological long-term sequelae after
posterior fossa tumour resection during childhood. Brain 126(Pt 9): 1998-2008. 30. Levisohn L, Cronin-Golomb A, Schmahmann JD. (2000) Neuropsychological consequences of
cerebellar tumour resection in children: Cerebellar cognitive affective syndrome in a paediatric population. Brain 123: 1041-50.
31. de Smet HJ, Baillieux H, Wackenier P, et al. (2009) Long-term cognitive deficits following posterior fossa tumor resection: A neuropsychological and functional neuroimaging follow-up study. Neuropsychology 23(6): 694-704.
32. Pollack IF, Polinko P, Albright AL, et al. (1995) Mutism and pseudobulbar symptoms after resection of posterior fossa tumors in children: Incidence and pathophysiology. Neurosurgery 37(5): 885-93.
33. Ozimek A, Richter S, Hein-Kropp C, et al. (2004) Cerebellar mutism––report of four cases. J Neurol 251(8): 963-72.
34. Daniels SR, Moores LE, DiFazio MP. (2005) Visual disturbance associated with postoperative cerebellar mutism. Pediatr Neurol 32(2): 127-30.
35. Riva D. (1998) The cerebellar contribution to language and sequential functions: Evidence from a
114
AIMS Neuroscience Volume 1, Issue 1, 96-119.
child with cerebellitis. Cortex 34(2): 279-87. 36. Drost G, Verrips A, Thijssen HO, et al. (2000) Cerebellar involvement as a rare complication of
pneumococcal meningitis. Neuropediatrics 31(2): 97-9. 37. Fujisawa H, Yonaha H, Okumoto K, et al. (2005) Mutism after evacuation of acute subdural
hematoma of the posterior fossa. Childs Nerv Syst 21(3): 234-6. 38. Ersahin Y, Mutluer S, Saydam S, Barcin E. (1997) Cerebellar mutism: Report of two unusual
cases and review of the literature. Clin Neurol Neurosurg 99(2): 130-4. 39. Al-Anazi A, Hassounah M, Sheikh B, et al. (2001) Cerebellar mutism caused by arteriovenous
malformation of the vermis. Br J Neurosurg 15(1): 47-50. 40. Dubey A, Sung WS, Shaya M, et al. (2009) Complications of posterior cranial fossa surgery––an
institutional experience of 500 patients. Surg Neurol 72(4): 369-75. 41. Afshar-Oromieh A, Linhart H, Podlesek D, et al. (2010) Postoperative cerebellar mutism in adult
patients with lhermitte-duclos disease. Neurosurg Rev 33(4): 401-8. 42. Coplin WM, Kim DK, Kliot M, et al. (1997) Mutism in an adult following hypertensive
cerebellar hemorrhage: Nosological discussion and illustrative case. Brain Lang 59(3): 473-93. 43. Moore MT. (1969) Progressive akinetic mutism in cerebellar hemangioblastoma with
"normal-pressure hydrocephalus". Neurology 19(1): 32-6. 44. Caner H, Altinors N, Benli S, et al. (1999) Akinetic mutism after fourth ventricle choroid plexus
papilloma: Treatment with a dopamine agonist. Surg Neurol 51(2): 181-4. 45. Idiaquez J, Fadic R, Mathias CJ. (2001) Transient orthostatic hypertension after partial cerebellar
resection. Clin Auton Res 21(1): 57-9. 46. De Smet HJ, Marien P. (2012) Posterior fossa syndrome in an adult patient following surgical
evacuation of an intracerebellar haematoma. Cerebellum 11(2): 587-92. 47. Dunwoody GW, Alsagoff ZS, Yuan SY. (1997) Cerebellar mutism with subsequent dysarthria in
an adult: Case report. Br J Neurosurg 11(2): 161-3. 48. Marien P, Verslegers L, Moens M, et al. (2013) Posterior fossa syndrome after cerebellar stroke.
Cerebellum 12(5): 686-91. 49. Sacchetti B, Scelfo B, Strata P. (2005) The cerebellum: Synaptic changes and fear conditioning.
Neuroscientist 11(3): 217-27. 50. LeDoux JE. (1994) Emotion, memory and the brain. Sci Am 270(6): 50-7. 51. Maschke M, Schugens M, Kindsvater K, et al. (2002) Fear conditioned changes of heart rate in
patients with medial cerebellar lesions. J Neurol Neurosurg Psychiatry 72(1): 116-8. 52. Maschke M, Drepper J, Kindsvater K, et al. (2000) Fear conditioned potentiation of the acoustic
blink reflex in patients with cerebellar lesions. J Neurol Neurosurg Psychiatry 68(3):358-64. 53. Turner BM, Paradiso S, Marvel CL, et al. (2007) The cerebellum and emotional experience.
Neuropsychologia 45(6): 1331-41. 54. Nashold BS, Wilson WP, Slaughter DG. (1969) Sensations evoked by stimulation in the midbrain
of man. J Neurosurg 30(1): 14-24. 55. Damasio AR, Grabowski TJ, Bechara A, et al. (2000) Subcortical and cortical brain activity
during the feeling of self-generated emotions. Nat Neurosci 3(10): 1049-56. 56. Ploghaus A, Tracey I, Clare S, et al. (2000) Learning about pain: The neural substrate of the
prediction error for aversive events. Proc Natl Acad Sci USA 97(16): 9281-6. 57. Ploghaus A, Tracey I, Gati JS, et al. (1999) Dissociating pain from its anticipation in the human
brain. Science 284(5422): 1979-81. 58. Singer T, Seymour B, O'Doherty J, et al. (2004) Empathy for pain involves the affective but not
sensory components of pain. Science 303(5661): 1157-62.
115
AIMS Neuroscience Volume 1, Issue 1, 96-119.
59. Fischer H, Andersson JL, Furmark T, et al. (2000) Fear conditioning and brain activity: A positron emission tomography study in humans. Behav Neurosci 114(4): 671-80.
60. Frings M, Maschke M, Erichsen M, et al. (2002) Involvement of the human cerebellum in fear-conditioned potentiation of the acoustic startle response: A PET study. Neuroreport 13(10): 1275-8.
61. Timmann D, Drepper J, Frings M, et al. (2010) The human cerebellum contributes to motor, emotional and cognitive associative learning. A review. Cortex 46(7): 845-57.
62. Baumann O, Mattingley JB. (2012) Functional topography of primary emotion processing in the human cerebellum. Neuroimage 61(4): 805-11.
63. Schienle A, Scharmuller W. (2013) Cerebellar activity and connectivity during the experience of disgust and happiness. Neuroscience 246: 375-81.
64. Schienle A, Stark R, Walter B, et al. (2002) The insula is not specifically involved in disgust processing: An fMRI study. Neuroreport 13(16): 2023-6.
65. Stoodley CJ, Schmahmann JD. (2009) Functional topography in the human cerebellum: A meta-analysis of neuroimaging studies. Neuroimage 44(2): 489-501.
66. Heilman KM, Gilmore RL. (1998) Cortical influences in emotion. J Clin Neurophysiol 15(5): 409-23.
67. Fusar-Poli P, Placentino A, Carletti F, et al. (2009) Functional atlas of emotional faces processing: A voxel-based meta-analysis of 105 functional magnetic resonance imaging studies. J Psychiatry Neurosci 34(6): 418-32.
68. Vuilleumier P, Pourtois G. (2007) Distributed and interactive brain mechanisms during emotion face perception: Evidence from functional neuroimaging. Neuropsychologia 45(1): 174-94.
69. Schutter DJ, Enter D, Hoppenbrouwers SS. (2009) High-frequency repetitive transcranial magnetic stimulation to the cerebellum and implicit processing of happy facial expressions. J Psychiatry Neurosci 34(1): 60-5.
70. Ferrucci R, Giannicola G, Rosa M, et al. (2012) Cerebellum and processing of negative facial emotions: Cerebellar transcranial DC stimulation specifically enhances the emotional recognition of facial anger and sadness. Cogn Emot 26(5):786-99.
71. Adamaszek M, D'Agata F, Kirkby KC, et al. (2014) Impairment of emotional facial expression and prosody discrimination due to ischemic cerebellar lesions. Cerebellum 13(3):338-45.
72. Furl N, van Rijsbergen NJ, Kiebel SJ, et al. (2010) Modulation of perception and brain activity by predictable trajectories of facial expressions. Cereb Cortex 20(3): 694-703.
73. Kilts CD, Egan G, Gideon DA, et al. (2003) Dissociable neural pathways are involved in the recognition of emotion in static and dynamic facial expressions. Neuroimage 18(1): 156-68.
74. Villanueva R. (2012) The cerebellum and neuropsychiatric disorders. Psychiatry Res 198(3): 527-32.
75. Loeber RT, Cintron CM, Yurgelun-Todd DA. (2001) Morphometry of individual cerebellar lobules in schizophrenia. Am J Psychiatry 158(6): 952-4.
76. Okugawa G, Sedvall GC, Agartz I. (2003) Smaller cerebellar vermis but not hemisphere volumes in patients with chronic schizophrenia. Am J Psychiatry 160(9): 1614-7.
77. Jacobsen LK, Giedd JN, Berquin PC, et al. (1997) Quantitative morphology of the cerebellum and fourth ventricle in childhood-onset schizophrenia. Am J Psychiatry 154(12): 1663-9.
78. Nasrallah HA, Schwarzkopf SB, Olson SC, et al. (1991) Perinatal brain injury and cerebellar vermal lobules I-X in schizophrenia. Biol Psychiatry 29(6): 567-74.
79. Ichimiya T, Okubo Y, Suhara T, et al. (2001) Reduced volume of the cerebellar vermis in neuroleptic-naive schizophrenia. Biol Psychiatry 49(1): 20-7.
116
AIMS Neuroscience Volume 1, Issue 1, 96-119.
80. Staal WG, Hulshoff Pol HE, Schnack HG, et al. (2000) Structural brain abnormalities in patients with schizophrenia and their healthy siblings. Am J Psychiatry 157(3): 416-21.
81. Aylward EH, Reiss A, Barta PE, et al. (1994) Magnetic resonance imaging measurement of posterior fossa structures in schizophrenia. Am J Psychiatry 151(10): 1448-52.
82. Heath RG. (1977) Modulation of emotion with a brain pacemaker. Treatment for intractable psychiatric illness. J Nerv Ment Dis 165(5): 300-17.
83. Demirtas-Tatlidede A, Freitas C, Cromer JR, et al. (2010) Safety and proof of principle study of cerebellar vermal theta burst stimulation in refractory schizophrenia. Schizophr Res 124(1-3): 91-100.
84. Manu P, Sarpal D, Muir O, et al. (2011) When can patients with potentially life-threatening adverse effects be rechallenged with clozapine? A systematic review of the published literature. Schizophr Res 134(2-3): 180-6.
85. Soares JC, Mann JJ. (1997) The anatomy of mood disorders––review of structural neuroimaging studies. Biol Psychiatry 41(1): 86-106.
86. Fitzgerald PB, Laird AR, Maller J, et al. (2008) A meta-analytic study of changes in brain activation in depression. Hum Brain Mapp 29(6): 683-95.
87. Leroi I, O'Hearn E, Marsh L, Lyketsos CG, et al. (2002) Psychopathology in patients with degenerative cerebellar diseases: A comparison to huntington's disease. Am J Psychiatry 159(8): 1306-14.
88. Alalade E, Denny K, Potter G. (2011) Altered cerebellar-cerebral functional connectivity in geriatric depression. PLoS One 6(5): e20035.
89. Kanner L. (1943) Autistic disturbances of affective contact. Nervous Child 2: 217-50. 90. Senju A, Johnson MH. (2009) Atypical eye contact in autism: Models, mechanisms and
development. Neurosci Biobehav Rev 33(8): 1204-14. 91. Ospina MB, Krebs Seida J, Clark B, et al. (2008) Behavioural and developmental interventions
for autism spectrum disorder: A clinical systematic review. PLoS One 3(11): e3755. 92. Hughes JR. (2009) Update on autism: A review of 1300 reports published in 2008. Epilepsy
Behav 16(4): 569-89. 93. Muratori F, Maestro S. (2007) Autism as a downstream effect of primary difficulties in
intersubjectivity interacting with abnormal development of brain connectivity. Inter J Dialog Science 2007(2): 93-118.
94. Bailey A, Luthert P, Dean A, et al. (1998) A clinicopathological study of autism. Brain 121 ( Pt 5)(Pt 5): 889-905.
95. Bauman M, Kemper TL. (1985) Histoanatomic observations of the brain in early infantile autism. Neurology 35(6): 866-74.
96. Courchesne E, Yeung-Courchesne R, Press GA, et al. (1988) Hypoplasia of cerebellar vermal lobules VI and VII in autism. N Engl J Med 318(21): 1349-54.
97. Courchesne E, Hesselink JR, Jernigan TL, et al. (1987) Abnormal neuroanatomy in a nonretarded person with autism. unusual findings with magnetic resonance imaging. Arch Neurol 44(3): 335-41.
98. Hashimoto T, Tayama M, Murakawa K, et al. (1995) Development of the brainstem and cerebellum in autistic patients. J Autism Dev Disord 25(1):1-18.
99. Ingram JL, Peckham SM, Tisdale B, et al. (2000) Prenatal exposure of rats to valproic acid reproduces the cerebellar anomalies associated with autism. Neurotoxicol Teratol 22(3): 319-24.
100.Pierce K, Courchesne E. (2001) Evidence for a cerebellar role in reduced exploration and stereotyped behavior in autism. Biol Psychiatry 49(8): 655-64.
117
AIMS Neuroscience Volume 1, Issue 1, 96-119.
101.Sparks BF, Friedman SD, Shaw DW, et al. (2002) Brain structural abnormalities in young children with autism spectrum disorder. Neurology 59(2): 184-92.
102.Allen G, Courchesne E. (2003) Differential effects of developmental cerebellar abnormality on cognitive and motor functions in the cerebellum: An fMRI study of autism. Am J Psychiatry 160(2): 262-73.
103.Waterhouse L, Fein D, Modahl C. (1996) Neurofunctional mechanisms in autism. Psychol Rev 103(3): 457-89.
104.Snow WM, Hartle K, Ivanco TL. (2008) Altered morphology of motor cortex neurons in the VPA rat model of autism. Dev Psychobiol 50(7): 633-9.
105.Allen G, Buxton RB, Wong EC, et al. (1997) Attentional activation of the cerebellum independent of motor involvement. Science 275(5308): 1940-3.
106.Lalonde R, Strazielle C. (2003) The effects of cerebellar damage on maze learning in animals. Cerebellum 2(4): 300-9.
107.Joyal CC, Strazielle C, Lalonde R. (2001) Effects of dentate nucleus lesions on spatial and postural sensorimotor learning in rats. Behav Brain Res 122(2): 131-7.
108.Joyal CC, Meyer C, Jacquart G, et al. (1996) Effects of midline and lateral cerebellar lesions on motor coordination and spatial orientation. Brain Res 739(1-2): 1-11.
109.Leggio MG, Molinari M, Neri P, et al. (2000) Representation of actions in rats: The role of cerebellum in learning spatial performances by observation. Proc Natl Acad Sci USA 97(5): 2320-5.
110.Dow RS, Moruzzi G. (1958) The physiology and pathology of the cerebellum. Minneapolis: University of Minnesota Press.
111.Sprague JM, Chambers WW. (1959) An analysis of cerebellar function in the cat, as revealed by its complete and partial destruction, and its interaction with the cerebral cortex. Archs ital Biol 97: 68-88.
112.Chambers WW, Sprague JM. (1955) Functional localization in the cerebellum II. Arch Neurol Psychiat (Chic) 74: 653-680.
113.Peters M, Monjan AA. (1971) Behavior after cerebellar lesions in cats and monkeys. Physiol Behav 6(2): 205-6.
114.Berman AS, Berman D, Prescott JW. (1974) The effect of cerebellar lesions on emotional behavior in the rhesus monkey. In: Cooper IS, Riklan M, Snider RS, editors. Ihe Cerebellum, Epilepsy, and Behavior. New York: Plenum, 1974.
115.Supple WF, Leaton RN, Fanselow MS. (1987) Effects of cerebellar vermal lesions on species-specific fear responses, neophobia, and taste-aversion learning in rats. Physiol Behav 39(5): 579-86.
116.Barnett SA, Barnett SA. (1975) The rat: a study in behavior. Chicago: University of Chicago. Press.
117.Supple WF, Leaton RN. (1990) Cerebellar vermis: Essential for classically conditioned bradycardia in the rat. Brain Res 509(1): 17-23.
118.Supple WF, Kapp BS. (1993) The anterior cerebellar vermis: Essential involvement in classically conditioned bradycardia in the rabbit. J Neurosci 13(9): 3705-11.
119.Yuzaki M. (2003) The delta2 glutamate receptor: 10 years later. Neurosci Res 46(1): 11-22. 120.Sacchetti B, Scelfo B, Tempia F, et al. (2004) Long-term synaptic changes induced in the
cerebellar cortex by fear conditioning. Neuron 42(6): 973-82. 121.Saab CY, Willis WD. (2003) The cerebellum: Organization, functions and its role in nociception.
Brain Res Brain Res Rev 42(1): 85-95.
118
AIMS Neuroscience Volume 1, Issue 1, 96-119.
122.Sacchetti B, Baldi E, Lorenzini CA, et al. (2002) Cerebellar role in fear-conditioning consolidation. Proc Natl Acad Sci USA 99(12): 8406-11.
123.Sacchetti B, Scelfo B, Strata P. (2009) Cerebellum and emotional behavior. Neuroscience 162(3): 756-62.
124.Butler AB, Hodos H. (1996) Comparative vertebrate neuroanatomy. New York: Wiley-Liss. 125.Rodriguez F, Duran E, Gomez A, et al. (2005) Cognitive and emotional functions of the teleost
fish cerebellum. Brain Res Bull 66(4-6): 365-70. 126.Yoshida M, Kondo H. (2012) Fear conditioning-related changes in cerebellar purkinje cell
activities in goldfish. Behav Brain Funct 8: 52. 127.Supple WF, Sebastiani L, Kapp BS. (1993) Purkinje cell responses in the anterior cerebellar
vermis during pavlovian fear conditioning in the rabbit. Neuroreport 4(7): 975-8. 128.Middleton FA, Strick PL. (1994) Anatomical evidence for cerebellar and basal ganglia
involvement in higher cognitive function. Science 266(5184): 458-61. 129.Middleton FA, Strick PL. (2000) Cerebellar projections to the prefrontal cortex of the primate. J
Neurosci 21(2):700-12. 130.Middleton FA, Strick PL. (1998) Cerebellar output: Motor and cognitive channels. Trends Cogn
Sci 2(9): 348-54. 131.Dietrichs E. (1984) Cerebellar autonomic function: Direct hypothalamocerebellar pathway.
Science 223(4636): 591-3. 132.Haines DE, Dietrichs E. (1984) An HRP study of hypothalamo-cerebellar and
cerebello-hypothalamic connections in squirrel monkey (saimiri sciureus). J Comp Neurol 229(4): 559-75.
133.Snider RS, Maiti A. (1976) Cerebellar contributions to the papez circuit. J Neurosci Res 2(2): 133-46.
134.Heath RG. (1972) Physiologic basis of emotional expression: Evoked potential and mirror focus studies in rhesus monkeys. Biol Psychiatry 5(1):15-31.
135.Heath RG, Harper JW. (1974) Ascending projections of the cerebellar fastigial nucleus to the hippocampus, amygdala, and other temporal lobe sites: Evoked potential and histological studies in monkeys and cats. Exp Neurol 45(2): 268-87.
136.Prescott JW. (1971) Early somatosensory deprivation as ontogenic process in the abnormal development of the brain and behavior. In: Moor-Jankowski E, editor. Basel: Karger.
137.D'Angelo E, Casali S. (2013) Seeking a unified framework for cerebellar function and dysfunction: From circuit operations to cognition. Front Neural Circuits 6: 116.
138.Larouche M, Hawkes R. (2006) From clusters to stripes: The developmental origins of adult cerebellar compartmentation. Cerebellum 5(2): 77-88.
139.Snow WM, Anderson JE, Fry M. (2014) Regional and genotypic differences in intrinsic electrophysiological properties of cerebellar purkinje neurons from wild-type and dystrophin-deficient mdx mice. Neurobiol Learn Mem 107: 19-31.
140.Stoodley CJ, Schmahmann JD. (2010) Evidence for topographic organization in the cerebellum of motor control versus cognitive and affective processing. Cortex 46(7): 831-44.
141.Schmahmann JD, Caplan D. (2006) Cognition, emotion and the cerebellum. Brain 129(Pt 2): 290-2.
142.Leiner HC, Leiner AL, Dow RS. (1989) Reappraising the cerebellum: What does the hindbrain contribute to the forebrain? Behav Neurosci 103(5): 998-1008.
143.Schmahmann JD. (2010) The role of the cerebellum in cognition and emotion: Personal reflections since 1982 on the dysmetria of thought hypothesis, and its historical evolution from
119
AIMS Neuroscience Volume 1, Issue 1, 96-119.
theory to therapy. Neuropsychol Rev 20(3): 236-60. 144.Nudo RJ, Plautz EJ, Frost SB. (2001) Role of adaptive plasticity in r

![Altered Cerebellar Functional Connectivity with Intrinsic ... · order functions of the cerebellum [2], including cognitive processing, emotional control, as well as learning and](https://img.pdfslide.us/doc/110x75/5fdc05992d3b4b5b3e4287b2/altered-cerebellar-functional-connectivity-with-intrinsic-order-functions-of.jpg)