Embed Size (px)
Citation preview

J. CM Set. 33, 185-195 (1978) 185Printed in Great Britain © Compdny of Biologists Limited i<)j8
NEW STAINING METHOD FOR THE
DETECTION OF SISTER-CHROMATID
EXCHANGES IN BrdU-LABELLED
CHROMOSOMES
P. ALVES AND J. JONASSONInstitute of Medical Genetics, University of Uppsala,V. Agatan 24, S-752 20 Uppsala, Sweden
SUMMARY
A direct Giemsa technique is described for the selective staining of BrdU-substitutedchromatin. It can advantageously be used for sister-chromatid exchange analysis and for thedetailed cytological study of the mouse kinetochores. Presumptive inversions within the centro-meric region of chromosomes 4 and 18 in the CS7Black/6J mouse are described.
INTRODUCTION
Methods have recently been developed that allow for the rapid scoring of sister-chromatid exchanges in the metaphase chromosomes of cells that have been grownfor two replication cycles in the presence of 5-bromodeoxyuridine (BrdU) (Latt,1973; Perry & Wolff, 1974; Korenberg & Freedlender, 1974). By means of the alteredHoechst 33258 and/or Giemsa-staining properties of BrdU-substituted chromatinthese methods will positively stain only chromatids containing a parental DNA strandwhich does not contain BrdU.
We report here a simple procedure, based upon the principles of the Giemsa-11technique (Bobrow, Madan & Pearson, 1972), by which it is possible to stain insteadthe BrdU-substituted chromatin. The present direct Giemsa technique produces anintense magenta staining of fully BrdU-substituted chromatin, whereas half-sub-stituted chromatin remains pale. Non-substituted chromatin is refractory to staining.The microscopical resolution and information gained is far greater than what isgenerally achieved with the conventional FPG technique (Perry & Wolff, 1974).
MATERIALS AND METHODS
Cells
Ci7Bl melanoma Clone PG19. This cell line from the mouse has been fully described byJonasson, Povey & Harris (1977).
Cell culture
The cells were maintained in plastic flasks in Eagle's Minimal Essential Medium (Eagle et al.1956) plus 10 % foetal calf serum. 5-bromodeoxyuridine was added to a final concentration of15 /ig/ml growth medium o, 12, 24 or 36 h prior to harvesting. Thereafter the cultures were

18 6 P. A Ives and J. Jonasson
kept in the complete dark. Details of culture techniques can be found in Klein, Bregula, Wiener& Harris (1971) and subsequent papers in that series.
Preparation of cells for chromosome analysis
Cells in tissue culture were treated with colchicine at a concentration of o-i /tg/ml for aperiod of 60 min. They were then exposed to a hypotonic solution of 0-56 % KC1 for 8 minand fixed (with 2 changes of fixative) in 1 part acetic acid and 3 parts methanol for at least30 min. Spreads were made on dry slides and air-dried.
Microscopical analysis
The metaphase spreads were stained for 10 min at room temperature in a 2 % solution ofGiemsa R66 (Gurr) in 0-3 M Na2HPO4 adjusted to pH 10-4 with 1 N NaOH. The preparationswere rinsed in distilled water, dried and examined under a 100 x oil-immersion planapochromaticlens in a Zeiss photomicroscope. Kodak SO 410 35-mm film was used for photography. As withall chromosome-banding techniques slides may respond differently to the staining procedure.However, by altering the pH between 102 and 106 and the phosphate concentration between0-2 and 04 M one can generally achieve satisfactory results. The study of the mouse kineto-chores is often made easier if the slides are preincubated in the buffer at 42 °C for 10 minbefore staining.
RESULTS AND DISCUSSION
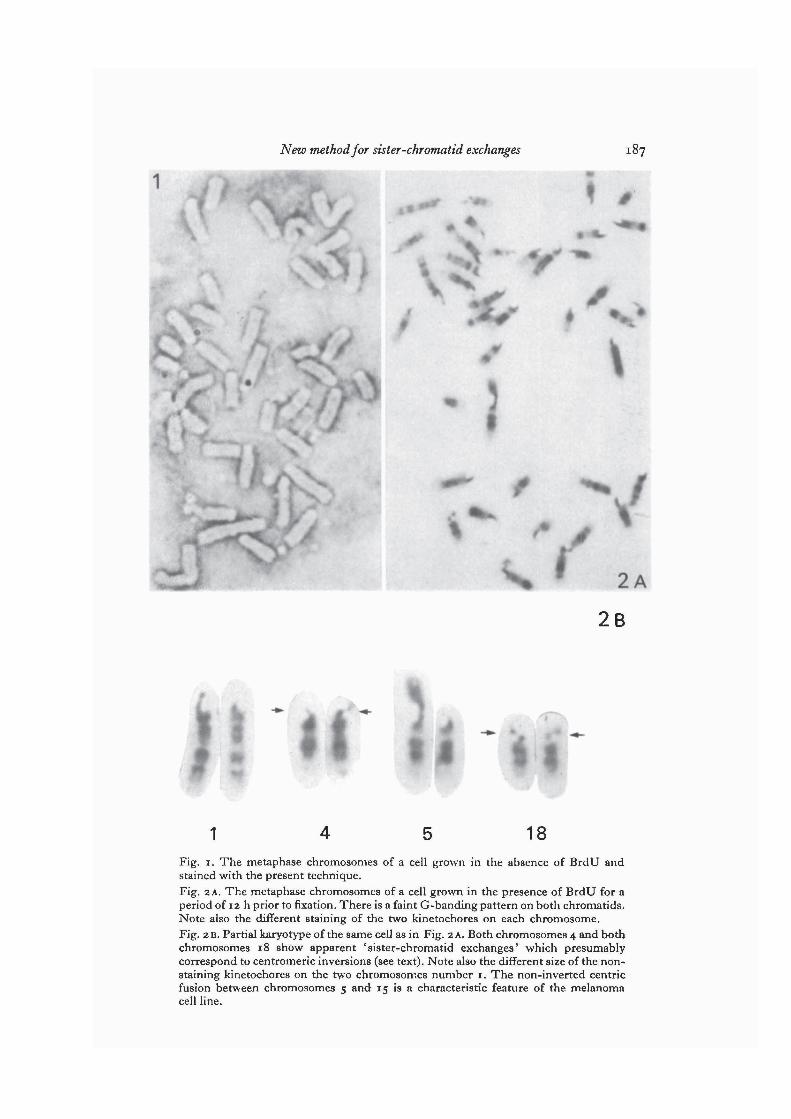
The metaphase chromosomes of cells grown in the absence of BrdU are refractoryto Giemsa staining with the present technique (Fig. 1).
After one complete cycle of DNA replication in the presence of BrdU all chromo-somes appear faintly stained, with a weak G-band differentiation along bothchromatids. One of the two kinetochores on each chromosome is totally resistant tothe Giemsa stain (Fig. 2 A).
A cytologically detectable lateral asymmetry in the centromeric region of mousechromosomes was first described by Lin, Latt & Davidson (1974) after studiesinvolving the BrdU quenching of 33258 Hoechst fluorescence. They put forward thehypothesis that the phenomenon may reflect the unequal distribution of thymidinebetween the two complementary polynucleotide chains of mouse satellite DNA (22vs 45 %) described by Flamm, McCallum & Walker (1967). Replication of such DNAin the presence of BrdU should result in the incorporation of more BrdU into one ofthe daughter helices than the other. This hypothesis can also fully explain our observa-tions, obtained with the present direct Giemsa technique. The parental strand ofDNA with a high thymidine content should in our case be represented by a non-staining kinetochore at the 1st division. The segregation pattern of the non-stainingkinetochores at the 2nd and 3rd divisions is in agreement with this notion (see furtherbelow).
In the present material we also found that after one complete cycle of DNA replica-tion in the presence of BrdU, both chromosomes 4 and both chromosomes 18 wereinvolved in apparent 'sister-chromatid exchanges' within the centromeric region(Fig. 2B) in all cells examined. Sister-chromatid exchanges on other chromosomeswere only rarely seen. The point of breakage of chromosome 18 was always located in
New method for sister-chromatid exchanges
1 18Fig. 1. The metaphase chromosomes of a cell grown in the absence of BrdU andstained with the present technique.Fig. 2 A. The metaphase chromosomes of a cell grown in the presence of BrdU for aperiod of 12 h prior to fixation. There is a faint G-banding pattern on both chromatids.Note also the different staining of the two kinetochores on each chromosome.Fig. 2B. Partial karyotype of the same cell as in Fig. 2 A. Both chromosomes 4 and bothchromosomes 18 show apparent 'sister-chromatid exchanges' which presumablycorrespond to centromeric inversions (see text). Note also the different size of the non-staining kinetochores on the two chromosomes number 1. The non-inverted centricfusion between chromosomes 5 and 15 is a characteristic feature of the melanomacell line.

188 P. Alves andj. Jonasson
the middle of the kinetochore, whereas the corresponding point on chromosome 4was always closer to the tip of the chromosome (Fig. 2 B).
This consistent feature of homozygosity for 'sister-chromatid exchanges' withinthe centromeric region of chromosomes 4 and 18 can easily be explained on the basisof the hypothesis of Lin et al. (1974) if one assumes that an inversion of the DNAdouble helix has occurred within the centromeric region of these chromosomes.(Throughout this work, we will use the concept of the chromatid as a continuous DNAdouble helix of which the centromeric satellite DNA is an interspersed and integralpart.) The thymidine-rich chain of satellite DNA of the inverted segment will thenbe linked to the opposite DNA chain of neighbouring segments (see e.g. Angell &Jacobs, 1975). The morphology of chromosome 18 in the present material is in factcompatible with such an inversion being present in the 'lower' segment of the centro-meric region (Fig. 2B). Under favourable conditions, a second break point can bedetected close to the edge of the centromeric region. On the other hand, the rearrangedchromosome 4 does not show any sign of having three segments, which one shouldexpect for an inversion.
Since chromosome 4 cannot positively be identified as having a true inversion oneshould also, perhaps, consider the possibility that it might have arisen by a process ofcentric fusion followed by centric fission. This would give the right chromosomalappearance, using the present method, if the thymidine-rich chain of satellite DNAwas associated with the same DNA chain in both interacting chromosomes. Alter-natively, if it was associated with the opposite DNA chains in the two chromosomes,a reciprocal translocation between the centromeric regions would produce the observedphenomenon. In any event, the chromosome will undoubtedly carry a segment ofsatellite DNA that is inverted relative to that of the rest of the chromosome. Therefore,we think it is appropriate to talk about 'centromeric inversions' being present evenif some of them may eventually turn out to represent centric fusion/fission or reci-procal translocation events rather than true inversions.
The same two 'centromeric inversions' as those observed in the present materialwere also identified in normal lymphocytes from C57Black/6J mice (kindly providedby Dr H. Wigzell) from which the melanoma was originally derived. We have seenthat polymorphism for such inversions exists between different inbred strains of mice.
Fig. 3 A. The metaphase chromosomes of a cell grown in the presence of BrdU for aperiod of 12 h prior to fixation. The slide was preincubated in the buffer before staining(see Materials and methods). A 'centromeric dot' (see text) can be seen on the palechromatid on most chromosomes. Its position generally agrees with the 'narrowestpoint' of the centromeric region. This can be seen on some chromosomes which haveconsistently got more than one 'centromeric dot', e.g. chromosome 19, which is thesmall one at the bottom of the photograph. This chromosome has got two dots wideapart. An unidentified chromosome in the centre has got a tandemly repeated dot. A'centromeric dot' is also present on the small centric fragment in the centre of thephotograph. Note also that the 'centromeric inversion' present on chromosome 18 isclearly visualized with its two break points in the bottom left corner.Fig. 3B. The metaphase chromosomes of a normal diploid C67Black/6J lymphocytegrown in the presence of BrdU for a period of 48 h prior to fixation. This cell is shownonly for the comparison with Fig. 3 A.

New method for sister-chromatid exchanges
32

190 P. Alves andj. Jonasson
One can therefore conveniently use these ' centromeric inversions' as chromosomalmarkers (unpublished results).
In this context it should also be mentioned that Lin & Davidson (1974) observedthat all metacentric chromosomes of the tobacco mouse, and also of other mouse cells,show contralateral symmetry in the centromeric region. They concluded: 'Theseresults suggest that centric fusion involves the maintenance of DNA polarity throughthe centromere and that the thymidine-rich chain of satellite DNA in the centromericregion is associated with the same DNA chain in every mouse chromosome'. Holm-quist & Comings (1975) confirmed these results.
Obviously, one cannot take their interpretations literally, but their results mightindicate that for functional reasons complete inversions of the centromeric regionpossibly do not occur in the mouse. To this matter, we should like to add the followingobservations. Two metacentric chromosomes are regularly present in the PG19 cellline. One of them is a homologous Robertsonian translocation of the two chromosomes12, which does show the expected contralateral symmetry, whereas the other is aRobertsonian translocation between chromosomes 5 and 15, which does not showcontralateral symmetry in spite of the bipartite nature of its centromeric region (Fig.2 B). If one supposes that completely inverted kinetochores do not exist in the mouse,the interpretation of this structure in accordance with the hypothesis of Lin et al.(1974) would require a centromeric inversion and the subsequent loss of non-invertedsegments, if any occurred at the microscopical level. This would in a great majorityof the cases be contradictory to the model for the formation of Robertsonian transloca-tions that Dancis & Holmquist (1977) recently have proposed, since their modelpredicts that no loss of chromosome material occurs at centric fusions. If, on the otherhand, completely inverted kinetochores do exist in the mouse and, therefore, oneshould explain the centric fusion between chromosomes 5 and 15 accordingly, thebasis of their model (viz. a strict conservation of' satellite polarity' within the telomere)is suspended.
In our preparations, under certain conditions, it is possible to assign a morpho-logical direction to each kinetochore in the following way. The centromeric region isprovisionally divided in two main segments, viz. a telomere proximal segment, and atelomere distal segment. The borderline between these two segments is demarcatedby the 'centromeric dot', a minute structure which does not show any lateral asym-metry, which we think may very well correspond to the attachment site of the spindlefibres (Fig. 3). (We propose to use this nomenclature even if this dot may not beidentical to the centromeric dots in Nigella described by Marks (1977).) The 'proximalsegment' of the centromeric region, which includes the end of the chromosome,
Fig. 4. Detail of a cell stained under the same conditions as in Fig. 3 A, where the only'centromeric dot' (arrow) present on the centric fusion chromosome 5 and 15 (vertical)can be clearly seen (see text).Fig. 5. The metaphase chromosomes of a cell grown in the presence of BrdU for aperiod of 24 h prior to fixation. There is one dark and one pale chromatid on eachchromosome. Approximately half of the pale ones have a non-staining kinetochore.Sister-chromatid exchanges can be seen on some chromosomes.

New method for sister -chromatid exchanges
/i* X*

192 P. Alves andj.jonasson
differs in its staining properties from the 'distal segment' and sometimes does notshow any lateral asymmetry, whereas the lateral asymmetry of the ' distal segment'seems to be constitutive. This difference is more pronounced in prometaphase cellswith relatively long chromosomes than in highly contracted c-mitoses, and it maywell reflect an organizational and functional difference between the two segments.Therefore, a complete inversion of the whole centromeric region should manifestitself, but we have not yet been able to obtain evidence of such an inversion of eitherchromosome 5 or 15 or any other chromosome in the present material. However, thereis evidence for structural rearrangements on several chromosomes including the centricfusion chromosome 5 and 15. This chromosome has got one centromeric dot insteadof the two that should be expected (Fig. 4). Consequently, it seems unnecessary tointroduce the concept of a kinetochore having a normal morphological direction butcompletely inverted satellite DNA. In the melanoma material there is a variationbetween the individual chromosomes in the amount of centromeric heterochromatinthat is present. Each segment within the kinetochore can occur small or large and issometimes repeated (compare e.g. the two chromosomes 1 in Fig. 2B, and see alsoFig. 3). Polymorphism in the mouse for the amount of centromeric heterochromatinis a well-known phenomenon (see e.g. Dev et al. 1975), which may be related to thisheterogeneity.
Some of the results, presented in the previous two paragraphs, are of preliminarycharacter. Therefore, it should perhaps be pointed out that they are used only toillustrate the possible applications of the present method. However, we are currentlyengaged in the further characterization of the kinetochores of both the melanoma cellline and normal diploid cells.
Sister-chromatid exchanges outside the centromeric region can be detected in themetaphase chromosomes of cells that have been grown for two replication cycles inthe presence of BrdU (Fig. 5). The fully BrdU-substituted chromatid stains intenselymagenta, whereas the half-substituted chromatid remains pale with a barely visibleG-banding pattern. Approximately half of the chromosomes receive a non-stainingkinetochore, which is nearly always located on the pale chromatid as expected, althoughoccasional transpositions may occur by sister-chromatid exchange (Fig. 5).
There is a difficulty with the conventional FPG technique to discriminate betweencells that were in the late 5-period when the BrdU was introduced (and thus may havechromosomal regions replicated three times in the presence of BrdU when observed)
Fig. 6. Detail of a metaphase cell grown in the presence of BrdU for a period of 36 hprior to fixation. There are short segments of isolabelling of the chromatids. This ispresumably due to the partial completion of three replication cycles in the presenceof BrdU.Fig. 7. The metaphase chromosomes of a cell grown in the presence of BrdU for aperiod of 36 h prior to fixation. Approximately half of the chromosomes appear withboth chromatids intensely staining. A few of the pale chromatids have a non-stainingkinetochore. A non-staining kinetochore is also found on a chromosome with bothchromatids intensely staining present in the upper left quadrant. There are severalsegmented chromosomes which presumably have arisen from sister-chromatidexchanges in previous cell cycles.

New method for sister-chromatid exchanges 193
s

194 P. Alves andj. Jonasson
and those cells that were actually out of the S-period. Such cells that have beenactively synthesizing DNA at the beginning of the BrdU-labelling period easily revealthemselves with the present technique. An example of a cell that has partially com-pleted three replication cycles in the presence of BrdU is given in Fig. 6. If, on theother hand, BrdU was present for less than two complete replication cycles the intenselystaining chromatid appears with unstained segments. These phenomena are not easilyseen with the conventional FPG technique.
After three replication cycles in the presence of BrdU. on the average half of thechromosomes appear with both chromatids intensely staining. There is still a faintG-banding pattern superimposed that often allows for their identification. The otherremaining chromosomes have got one pale and one intensely staining chromatid(Fig. 7). The actual number of each kind in the individual cell should be dependenton the orientation of dark and pale chromatids at the metaphase following the secondreplication cycle. From theoretical considerations one should expect on the averageone-eighth of the chromatids to have a non-staining kinetochore and these should bepale ones. The actual number in this material is slightly higher due to the productsof the 'centromeric inversions' described above. The exceptional case where a non-staining kinetochore is found on a chromosome having two intensely staining chroma-tids (Fig. 7) probably represents a sister-chromatid exchange event in a previous cellcycle. Such events also produce hybrid chromosomes which in one segment of thechromosome have got both chromatids intensely staining and in another segment havegot one pale and one intensely staining chromatid (Fig. 7).
It is not within the aim of the present communication to give an account for thefrequency of sister-chromatid exchanges in the material. This is to be presentedseparately.
CONCLUSIONS
It is concluded that the present Giemsa-staining technique has got many character-istics that in our opinion make it the method of choice for sister-chromatid exchangeanalysis: (1) It is a quick and simple method, and it gives excellent results. (2) Thereis more information to be gained from the direct staining of BrdU-substituted chromatinwith the present method than from other methods presently used which all givenegative staining of the BrdU-substituted chromatin.
Furthermore, we conclude from the present data that ' centromeric inversions' inthe mouse (which to our knowledge have not been described before) can be easilyrevealed by the use of the present Giemsa-staining technique.
This investigation was supported by grants from the Marcus Borgstrdm Fund and theSwedish Cancer Society (1082-B77-01X).

New method for sister-chromatid exchanges 195
REFERENCES
ANGELL, R. R. & JACOBS, P. A. (1975). Lateral asymmetry in human constitutive hetero-chromatin. Chromosoma 51, 301-310.
BOBROW, M., MADAN, K. & PEARSON, P. L. (1972). Staining of some specific regions of humanchromosomes, particularly the secondary constriction of No. 9. Nature, New Biol. 238,122-124.
DANCIS, B. M. & HOLMQUIST, G. P. (1977). Fusion model of telomere replication and itsimplications for chromosomal rearrangements. In Chromosomes Today, vol. 6 (ed. A. de laChapelle & M. Sorea), pp. 95-104. Amsterdam: Elsevier/North Holland Biomedical Press.
DEV, V. G., MILLER, D. A., TANTRAVAHI, R., SCHBECK, R. R., RODERICK, T . H., ERLANGER,
B. F. & MILLER, O. J. (1975). Chromosome markers in Mus musculus: differences in C-banding between the subspecies M.m. musculus and M.m. molossinus. Cltromosoina 53, 335-344.
EAGLE, H., OYAMA, V. I., LEVY, M., HORTON, C. L. & FLEISCHMANN, R. (1956). The growthresponse of mammalian cells in tissue culture to L-glutamine and L-glutamic acid. J. biol.Chem. 218, 607-616.
FLAMM, W. G., MCCALLUM, M. & WALKER, P. M. B. (1967). The isolation of complementarystrands from a mouse DNA fraction. Proc. vatn. Acad. Set. U.S.A. 57, 1729-1734.
HOLMQUIST, G. P. & COMINGS, D. E. (1975). Sister chromatid exchange and chromosomeorganization based on a bromodeoxyuridine Giemsa-C-banding technique (TC'-banding).Cftromosoma 52, 245-259.
JONASSON, J., POVEY, S. & HARRIS, H. (1977). The analysis of malignancy by cell fusion. VII.Cytogenetic analysis of hybrids between malignant and diploid cells and of tumours derivedfrom them.y. Cell Set. 24, 217-254.
KLEIN, G., BRECULA, U., WIENER, F. & HARRIS, H. (1971). The analysis of malignancy by cellfusion. I. Hybrids between tumour cells and L cell derivatives.^. Cell Set. 8, 659-672.
KORENBERG, J. R. & FREEDLENDER, E. F. (1974). Giemsa technique for the detection of sister-chromatid exchanges. Chromosoma 48, 355-360.
LATT, S. A. (1973). Microfluorometric detection of deoxyribonucleic acid replication in humanmetaphase chromosomes. Proc. natn. Acad. Set. U.S.A. 70, 3395-3399.
L I N , M. S. & DAVIDSON, R. L. (1974). Centric fusion, satellite DNA and DNA polarity inmouse chromosomes. Science, N.Y. 185, 1179-1181.
L I N , M. S., LATT, S. A. & DAVIDSON, R. L. (1974). Microfluorometric detection of asymmetryin the centromeric region of mouse chromosomes. Expl Cell Res. 86, 392-395.
MARKS, G. E. (1977). The nature of centromeric dots in Nigella chromosomes. Chromosoma62, 36o-373-
PERRY, P. & WOLFF, S. (1974). New Giemsa method for the differential staining of sisterchromatids. Nature, Lond. 251, 156-158.
(Received 28 November 1977)














![Catch and release: how do kinetochores hook the …...their biochemical affinity for [9,12], and promoting their detachment from, microtubules [12–16]. If the de-tachment-promoting](https://img.pdfslide.us/doc/110x75/5f288d866ae18939334091d8/catch-and-release-how-do-kinetochores-hook-the-their-biochemical-afinity.jpg)