Embed Size (px)
Citation preview

Proceedings of the South Dakota Academy of Science, Vol. 90 (2011) 12�
nEW RECORDS OF GASTEROID AnD SECOTIOID FUnGI FROM SAnD DUnES In nORTHWESTERn SOUTH DAKOTA
A. C. Gabel and M. L. Gabel*Biology Department
Black Hills State University1200 University
Spearfish, South Dakota 57799*Corresponding author email: [email protected]
ABSTRACT
Sand dunes in northwestern South Dakota were surveyed for fungi from 2002–2006 in Harding County and from 2003–2005 in Perkins County. In total, eight taxa of gasteroid fungi, Calvatia cyathiformis, C. fragilis, Disciseda bovista, Geastrum campestre, G. pectinatum, G. xerophilum, Tulostoma brumale, and T. fimbriatum var. campestre, and one secotioid fungal species, Montagnea arenaria, were recovered. Calvatia fragilis, Geastrum xerophilum, Montagnea are-naria, and Tulostoma brumale are new records for South Dakota, and M. arenaria is reported east of the Rocky Mountains for the first time. By contributing to the inventory of fungal biota in South Dakota and extending the known range for the taxa reported here, we highlight the importance of basic surveys of fungal diversity in the United States.
Keywords
distribution, gasteroid and secotioid fungi, northwestern South Dakota, sand dunes
INTRODUCTION
Fungal organisms with fruiting bodies consisting of a mass of spores (glebum) enclosed by an outer layer (peridium) have been termed gasteroid fungi. Once considered a growth form of a single class of fungi, the Gasteromycetes, it is now known that gasteroid fungi in the Basidiomycota have evolved several times from their agaricoid (mushroom) ancestors (Hibbett et al. 1997). Intermediate between the agaricoid and gasteroid forms are the secotioid fungi that resemble mushrooms with unexpanded caps and poorly developed gills (Hopple and Vilgalys 1999). Gasteroid and secotioid fungi can be found in a wide range of habitats, from high altitude forests to sea shores, and are particularly well repre-sented in arid areas throughout the world (Coker and Couch 1928; Bates 2004; Bates 2006; Bates et al. 2009).

12� Proceedings of the South Dakota Academy of Science, Vol. 90 (2011)
Orders of epigeous (emerging above ground) gasteroid fungi include the Lyco-perdales and Tulostomatales (Alexopoulos et al.1996; Kendrick 2000). Species in the Lycoperdales are known to inhabit xeric pastures, prairies, and deserts in North America (Coker and Couch 1928; Smith 1951; Miller and Miller 1988; Bates et al. 2009). The gasteroid order Tulostomatales includes species that are cosmopolitan in arid areas such as desert habitats and sand dunes, which occur primarily in the western and mid-continental United States (Wright 1987). Fruitings of saprophytic, secotioid fungi, such as Podaxis pistillaris and agaricus texensis (= Longula texensis), are frequently reported from sandy, arid areas in-cluding the southwestern and western United States and Mexico (Zeller 1943; Bowerman and Groves 1962; Miller 1983; Miller and Miller 1988).
Sand dune areas in northwestern South Dakota have not been well surveyed for fungi and the only gasteroid species reported previously from the area (Hard-ing County) were Disciseda subterranea (Lycoperdaceae), Geastrum drummondii (Geastraceae), and Tulostoma campestre (Tulostomataceae) (Gabel et al. 2004). There are no reports of secotioid fungi, however, from this area. The objective of this study was to collect and document more thoroughly gasteroid and secotioid fungi that occur in sand dunes of northwestern South Dakota. The results of our survey are reported from frequent visits to a sand dune complex in Hard-ing county from 2002–2006, and to a sand dune area in Perkins County from 2003–2005.
METHODS
Collecting sites were in Harding and Perkins Counties. The Harding County site was on the Hunsucker Ranch, T18N, R5E, in the north central portion of Section 3, latitude N45o 33.439’, longitude W103o 29.630’, ranging in altitude from 875 to 900 m. The Perkins County site was on the McKenna Ranch, T14 N, R10E, SW 1/4 of Section 24, latitude N45o 9.349’, longitude W102o 51.007’, at an altitude of approximately 800 m. Average annual precipitation recorded from 1948–2005 at the Antelope Station, the nearest recording station to Hunsucker Ranch, was 37.6 cm. The Perkins County (Zeona) Station, the nearest to McKenna Ranch, recorded an average annual precipitation of 38.4 cm from 1949–2002. Average wind velocities for these same time periods were 19.0 kph and 20.1 kph, for the Antelope and Zeona stations respectively.
The sandy prairies of these areas included swales (depressions), dunes (elevated areas), and blowouts. Gabel and Simonson (2008) found vegetation in swales and dunes to be more dense and more diverse (an average of 46 species and 43 species per transect, respectively) than in blowouts (averaging at 18 species per transect). The same study found vegetation of these areas to be dominated by native species, which represented 86–100% of all species reported from the transects, and included forbs, sedges, and grasses. Common forbs of the area include Eriogonum annuum Nutt., Ladeania lanceolata (Pursh) Egan & Reveal (= Psoralidium lanceolatum), and solidago missouriensis Nutt., as well as species of ambrosia and Penstemon. Thirty percent of species reported by Gabel and Simonson (2008) were grasses, such as andropogon hallii Hack., Bouteloua graci-

Proceedings of the South Dakota Academy of Science, Vol. 90 (2011) 12�
lis (Willd. ex Kunth) Lag. ex Griffiths, Calamovilfa longifolia (Hook.) Scribn., Hesperostipa comata (Trin. & Rupr.) Barkw., and sporobolus cryptandrus (Torr.) A. Gray.
Several fungi were collected from transects established for the floristic study of Gabel and Simonson (2008) and others were collected outside transects, result-ing in a repetition of collecting dates for some specimens. Collected specimens were brought to the laboratory and identified using standard microscopic proce-dures. As the identification of the Tulostoma species typically requires detailed ultrastructural information, spore and peristomes of collections from this genus were examined under scanning electron microscopy (SEM) using a JEOL 5600 LV. Date, site, location (latitude/longitude), collection number, habitat, and col-lector were recorded for each specimen, which are vouchered at the Black Hills State University Herbarium (BHSC), Spearfish, South Dakota.
RESULTS
Montagnea arenaria, a secotioid fungus, is newly reported from South Dakota, being the first published record of the species east of the Rocky Mountains. The nearest published records for M. arenaria are from southwest Wyoming and western Montana, approximately 600–650 km distant (Chen 1999). Calvatia fragilis, Geastrum xerophilum, and Tulostoma brumale are likewise new records for South Dakota. A detailed account of the specimens collected is given in Table 1. The following species are reported from Hunsucker Ranch sand dune area of Harding County: Calvatia cyathiformis, C. fragilis, Disciseda bovista (Fig. 1), Geastrum campestre (Fig. 2), G. pectinatum, G. xerophilum, Montagnea arenaria (Figs. 3–4), Tulostoma brumale (Figs. 5–7), and T. fimbriatum var. campestre (Figs. 8–10). The following species are reported from McKenna Ranch sand dune area in Perkins County: C. cyathiformis, C. fragilis, Disciseda bovista, G. pectinatum, G. xerophilum, M. arenaria, T. brumale, and T. fimbriatum var. camp-estre. Where specimens were collected from transects, all species except one were collected from swales or dunes. A single specimen, D. bovista, was collected from a Harding County blowout.
DISCUSSION
Because some disagreement exists in the literature over the salient characters that define particular gasteroid species, we have included details on the morphol-ogy of the taxa collected over the course of our survey to clarify the taxonomic concepts used here.
Calvatia cyathiformis and C. fragilis are distinguished by the morphology of the sterile portion of tissue below the gleba (termed a subglebum), as well as by their size, shape, and degree of spore ornamentation (Smith 1951; Bowerman and Groves 1962; Kuo 2008; Bates et al. 2009). Specimens in this study were identified as Calvatia fragilis if the gasterocarps were less than 6 × 6 cm, had a

128 Proceedings of the South Dakota Academy of Science, Vol. 90 (2011)
tapered base, and the subgleba was absent or less than 1/3 to 1/2 of the gastero-carp height.
Geastrum campestre specimens were small earthstars with a sulcate peristome (mouth area) that exhibited thick, fleshy, hygroscopic exoperidial rays (outer layer) that entirely enclosed the asperulate, sessile to nearly sessile endoperidium (spore containing area) when specimens were dry (Smith 1951; Cunningham 1979; Smith et al. 1981; Pegler et al. 1995). Geastrum pectinatum specimens exhibited a lead-purple endoperidium, a narrow, striate pedicel, and a sulcate peristome (Smith 1951; Cunningham 1979; Smith et al. 1981; Pegler et al. 1995; Miller and Miller 2006). Specimens of G. xerophilum had a sulcate peri-stome and a saccate endoperidium that is attached to a pedicel and surrounded by involute exoperidial rays (Ponce de Leon 1968).
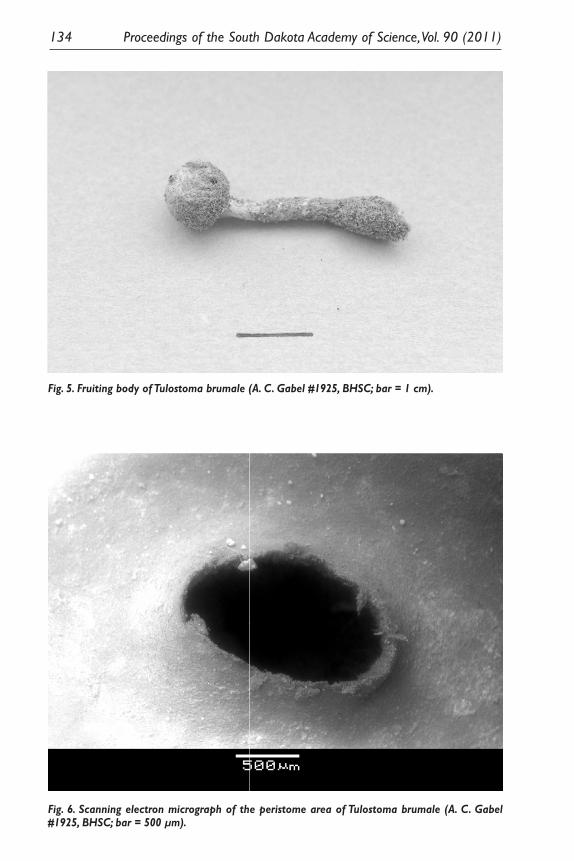
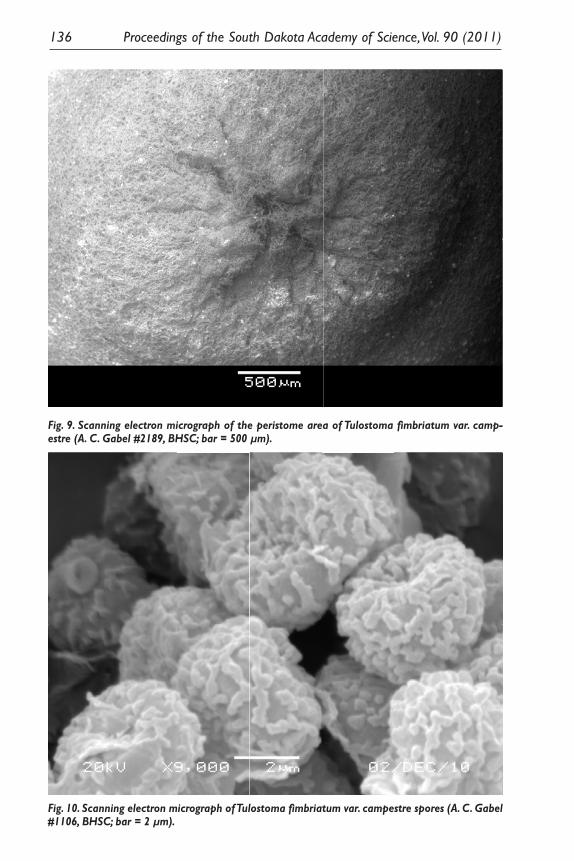
Scanning electron microscopy of spores from five specimens of each of the two Tulostoma species collected all exhibited distinct verrucae that coalesced into ridges or crests of different lengths (Figs. 7 and 10); therefore, other characteris-tic were important in distinguishing the species. Wright (1955) cites peristome morphology as an important character used to delimit species in the genus Tu-lostoma. Tulostoma brumale has a raised, smooth, circular ostiole (opening) that is typically darker than the surrounding endoperidium (Smith et al. 1981; Pegler et al. 1995). Wright (1987) recognized the combination T. fimbriatum Fries var. campestre (Morgan) Moreno, and indicated this was the most common variety in North America. The peristome of this species exhibits a lobed or mammose margin with lacerated openings (Wright 1987), and the specimens collected over the course of this study were consistent with Wright’s circumscription of this va-riety, having endoperidia with a persistent, basal sand case, a ribbed upper stalk, and stalks of 2.0–20.0 cm in height × 0.3–0.8 cm in diam.
Montagnea arenaria is a secotioid fungus that exhibits a fruiting body with a stalk, and cap-like head resembling mushrooms in the Agaricales; however, in this species the gills are reduced to plate-like structures called gussets containing spores that are not forcibly discharged (Smith 1973). The specimens collected in our survey, all exhibited these features. Hopple and Vilgalys (1999) confirmed the close relationship between M. arenaria and Coprinus comatus from their stud-ies of restriction site mapping of nuclear ribosomal DNA.
Thiers (1984) discussed the evolution of secotioid fungi and argued that this growth habit of possessing a closed or partially closed pileus, and basidiospores that are not forcibly discharged are responses to dry and inhospitable growing conditions. Discovery of the specimens of M. arenaria in the xeric conditions of northwestern South Dakota sand dunes would support Thiers’ (1984) hypoth-esis; however, the recent discovery of a wood inhabiting secotioid fungus from tropical forests of Guyana (Henkel et al. 2010) suggests otherwise.
Previous studies (Gabel and Ebbert 2004; Gabel et al. 2004) have recorded numerous gasteroid taxa from the Black Hills in western South Dakota that were not found in the dune sites. These include Bovista plumbea Pers., Calvatia booni-ana A. H. Sm., C. craniformis (Bosc.) Morgan, Crucibulum leave (Huds.) Kamb., Cyathis stercoreus (Schw.) de Toni, C. striatus Huds. ex Pers., Geastrum fornicatum (Huds.) Hook., G. quadrifidum Pers., G. saccatum Fr., G. schmidelii Vittad. (= G. nanuum Pers.), Lycoperdon foetidum Bonard., L. gunnii Berk., L. marginatum

Proceedings of the South Dakota Academy of Science, Vol. 90 (2011) 129
Vitt., L. pedicellatum Pk., L. perlatum Pers., and L. pyriforme Pers. Only three species that were collected from the dunes in this study, Calvatia cyathiformis, Geastrum campestre, and G. pectinatum, have been recorded previously from the Black Hills in western South Dakota (Gabel et al. 2004; Gabel and Gabel 2007). These numbers suggest a diverse flora of gasteroid taxa exist in western South Dakota; however, more collecting, including sand dunes or xeric areas, will be required before the true extent of gasteroid and secotioid fungal diversity from this area can be determined. Beyond adding to the knowledge of fungal diversity in this region, such survey efforts will ultimately help to better define the fungal flora of the United States, which is likewise poorly understood.
ACKNOWLEDGEMENTS
We regret to report that Audrey Gabel died 05 Jul 2011 of Creutzfeldt-Jakob disease. We appreciate the excellent comments and suggestions of our reviewers, Grace Kostel and especially Scott Bates who spent much time and effort to help with the timely completion of the manuscript. Black Hills State University pro-vided laboratory space for the study, and specimens are maintained in the Black Hills State University Herbarium (BHSC).
LITERATURE CITED
Alexopoulos, C. J., C. W. Mims and M. Blackwell. 1996. Introductory Mycol-ogy. John Wiley & Sons, New York, NY. 869 pp.
Bates, S. T. 2004. Arizona members of the Geastraceae and Lycopodiaceae (Ba-sidiomycota, fungi), M.S. Thesis, Arizona State University, Tempe, AZ. 445 pp.
Bates, S. T. 2006. Preliminary checklist of Arizona macrofungi. Canotia 2:47–78.
Bates, S. T., R. W. Roberson and D. E. Desjardin. 2009. Arizona gasteroid fungi I: Lycoperdaceae (Agaricales, Basidiomycota). Fungal Diversity 37:153–207.
Bowerman, C. A. and J. W. Groves. 1962. Notes on fungi from northern Canada V. Gasteromycetes. Canadian Journal of Botany 40:239–254.
Chen, C. 1999. Genetical and Molecular Systematic Study on the Genus Mon-tagnea Fr., A Desert Adapted Gasteromycete. M. S. Thesis, Virginia Poly-technic Institute and State University, Blacksburg, VA. 59 pp.
Coker, W. C. and J. N. Couch. 1928. The Gasteromycetes of the Eastern United States and Canada. Dover Publications, New York, NY. 201 pp.
Cunningham, G. H. 1979. The Gasteromycetes of Australia and New Zealand. J. Cramer, Dunedin, New Zealand. 236 pp.
Gabel, A. and E. Ebbert. 2004. Mushrooms and Other Fungi of the Black Hills and Surrounding Area. Black Hills State University Press, Spearfish, South Dakota. 164 pp.

1�0 Proceedings of the South Dakota Academy of Science, Vol. 90 (2011)
Gabel, A., E. Ebbert, and K. Lovett. 2004. Macrofungi collected from the Black Hills of South Dakota and Bear Lodge Mountains of Wyoming. The Ameri-can Midland Naturalist Journal 152:43–62.
Gabel, A. C. and M. L. Gabel. 2007. Comparison of diversity of macrofungi and vascular plants at seven sites in the Black Hills of South Dakota. The American Midland Naturalist 157:258–296.
Gabel, M. L. and D. L. Simonson. 2008. A comparison of the vegetation of two sand dune areas in northwestern South Dakota, U. S. A. Proceedings of the South Dakota Academy of Science 87:205–221.
Henkel, T. W., M. E. Smith, and M. C. Aime. 2010. Guyanagaster, a new wood-decaying sequestrate fungal genus related to armillaria (Physalacriaceae, Agaricales, Basidiomycota). American Journal of Botany 97:1474–1484.
Hibbett, D. S., E. M. Pine, E. Langer, G. Langer, and M. J. Donoghue. 1997. Evolution of gilled mushrooms and puffballs inferred from ribosomal DNA sequences. Proceedings of the National Academy of Science. 94:12002–12006.
Hopple, J. S. and R. Vilgalys. 1999. Phylogenetic relationships among coprinoid taxa and allies based on data from restriction site mapping of nucleus rDNA. Mycologia 86:96–107.
Kendrick, B. 2000. The Fifth Kingdom. Focus Publishing, R. Pullins Co., New-buryport, MA. 373 pp.
Kuo, M. 2008. Calvatia fragilis. Retrieved from the MushroomExpert.Com Web site: http://www.mushroomexpert.com/calvatia_fragilis.html (Cited 2 Febr. 2011).
Miller, O. K. Jr. 1983. Ectomycorrhizae in the Agaricales and Gasteromycetes. Canadian Journal of Botany 61:909–916.
Miller, O. K. and H. H. Miller. 1988. Gasteromycetes - Morphological and De-velopmental Features with Keys to the Orders, Families, and Genera. Mad River Press Inc., Eureka CA. 157 pp.
Miller, O. K. and H. H. Miller. 2006. North American Mushrooms, a Field Guide to Edible and Inedible Fungi. Falcon Guide, Helena, MT. 584 pp.
Mitchel, D. H., S. W. Chapman, and G. Grimes. 1975. Studies of Disciseda (Gasteromycetes) in Colorado. Mycologia 67:586–596.
Pegler, D. N., T. Laessoe and B. M. Spooner. 1995. British Puffballs, Earthstars and Stinkhorns: An Account of the British Gasteroid Fungi. Royal Botanic Gardens, Kew, England. 254 pp.
Ponce de Leon, P. 1968. A revision of the family Geastraceae. Fieldiana 31:303–349.
Smith, A. H. 1951. Puffballs and Their Allies in Michigan. University of Michi-gan Press, Ann Arbor, MI. 131 pp + 43 plates.
Smith, A. H. 1973. Agaricales and related secotioid Gasteromycetes, pp 421–450 In Ainsworth, G. C., F. K. Sparrow, and A. S. Sussman, editors. The fungi Vol. IVB, A Taxonomic Review with Keys: Basidiomycetes. Academic Press, New York, NY.
Smith, A. H., H. V. Smith and N. S. Weber. 1981. How to Know the Non-Gilled Mushrooms. Wm. C. Brown, Dubuque, IA. 324 pp.
Thiers, H. D. 1984. The secotioid syndrome. Mycologia 76:1–8.
Proceedings of the South Dakota Academy of Science, Vol. 90 (2011) 1�1
Wright, J. E. 1955. Evaluation of specific characters in the genus Tulostoma Pers. Papers of the Michigan Academy of Science, Arts and Letters 40:79–87.
Wright, J. E. 1987. The Genus Tulostoma (Gasteromycetes) – A World Mono-graph. J. Cramer, Bibliotheca Mycologica, Berlin, Germany 338 pp. + 50 plates.
Zeller, S. M. 1943. North American species of Galeropsis, Gyrophragmium, Lon-gia, and Montagnea. Mycologia 35:409–421.
Table 1. Specimen data for species collected over the course of the survey (2002–2006) from sand dune areas of northwestern South Dakota. All specimens were collected by A. C. Gabel, and data include collection number and date (multiple collections of a single species for a given date rep-resent specimens gathered at different transects). Localities are indicated for Hunsucker Ranch, Harding Co. (H) or McKenna Ranch, Perkins Co. (M). All specimens are deposited in the Black Hills State University Herbarium (BHSC).
Species Collection DataCalvatia cyathiformis(Bosc) Morgan
1501 (18 Jun 03; M), 2172 (01 Sep 04; M), 2173 (01 Sep 04; M),2169 (01 Sep 04; H)
C. fragilis(Vittad.) Morgan
1521 (27 Jun 03; H), 2168 (01 Sep 04; H), 2170 (01 Sep 04; M),2399 (30 Oct 05; M)
Disciseda bovista(Klotzsch) P. Henn
1105 (20 Jun 02; H), 1147 (02 Aug 02; H), 2203 (29 Sep 02; H),1922 (23 May 03; H), 2198 (23 May 03; H), 2194 (27 Jun 03; H),2186 (09 Apr 04; M), 2199 (09 Apr 04; M), 1927 (26 May 04; H),1936 (30 May 04; M), 1930 (30 May 04; M), 1934 (30 May 04; M), 2070 (29 Jul 04; H), 2074 (29 Jul 04; H), 2174 (01 Sep 04; M),2324 (20 Jul 05; H) 2341 (31 Jul 05; H), 2388 (27 Sep 05; H),2398 (10 Oct 05; H), 2403 (30 Oct 05; M), 2431 (7 Jun 06; H)
Geastrum campestre(Morgan) Kambly & Lee
1148 (02 Aug 02; H), 2200 (29 Sep 02; H), 2193 (27 Jun 03; H),2185b (09 Apr 04; H), 1926 (26 May 04; H), 1928 (26 May 04; H),1924 (26 May 04; H), 2073 (29 Jul 04; H), 2706 (29 Jul 04; H),2078 (29 Jul 04; M), 2175 (01 Sep 04; H), 2176 (01 Sep 04; H),2434 (07 Jun 06; H)
Geastrum pectinatumPersoon 2190 (09 Apr 04; H), 2197 (09 Apr 04; H), 1933 (30 May 04; M)
Geastrum xerophilum(Long) P. Ponce.
1921 (26 May 04; H), 2071 (29 Jul 04; H), 2389 (27 Sep 05; H),2402 (30 Oct 05; M)
Montagnea arenaria(D.C.) Zeller
1488 (18 Jun 03; M), 1929 (30 May 04; M), 2002 (21 Jun 04; M),2080 (29 Jul 04; M), 2322 (20 Jul 05; H), 2387 (27 Sep 05; H)
Tulostoma brumalePersoon
1146b (20 Jun 02; H), 1151b (02 Aug 02; H), 2191 (09 Apr 04; H),1925 (26 May 04;H), 2343 (31 Jul 05; H), 2396 (30 Oct 05;H),2400 (30 Oct 05; M), 1433 (07 Jun 06; H), 2433 (07 Jun 06; H)
T. fimbriatum Fries var.campestre (Morgan) Moreno
1106 (28 Jun 02; H), 1151a (02 Aug 02; H), 2201 (29 Sep 02; H),2197 (23 May 03; H), 2195 (27 Jun 03; H), 2189 (09 Apr 04; H),2188 (09 Apr 04; H), 2192 (23 May 04; H), 1923 (26 May 04; H),1931 (30 May 04; M), 2077 (29 Jul 04; M), 2075 (29 Jul 04; H),2323 (20 Jul 05; H), 2397 (30 Oct 05; H), 2401 (30 Oct 05; M),2432 (07 Jun 06; H)

1�2 Proceedings of the South Dakota Academy of Science, Vol. 90 (2011)
Fig. 1. Fruiting bodies of Disciseda bovista (A. C. Gabel #1147, BHSC; bar = 1 cm).
Fig. 2. Fruiting body of Geastrum campestre with hygroscopic rays (A. C. Gabel #1148, BHSC; bar = 1 cm).

Proceedings of the South Dakota Academy of Science, Vol. 90 (2011) 1��
Fig. 3. Fruiting body of Montagnea arenaria (A. C. Gabel #2002, BHSC; bar = 1 cm).
Fig. 4. Pileus (central area) surrounded by gussets of Montagnea arenaria (A. C. Gabel #2002, BHSC; bar = 1 cm).
1�� Proceedings of the South Dakota Academy of Science, Vol. 90 (2011)
Fig. 5. Fruiting body of Tulostoma brumale (A. C. Gabel #1925, BHSC; bar = 1 cm).
Fig. 6. Scanning electron micrograph of the peristome area of Tulostoma brumale (A. C. Gabel #1925, BHSC; bar = 500 µm).

Proceedings of the South Dakota Academy of Science, Vol. 90 (2011) 1��
Fig. 7. Scanning electron micrograph of Tulostoma brumale spores (A. C. Gabel #1151b, BHSC; bar = 2 µm).
Fig. 8. Fruiting bodies of Tulostoma fimbriatum var. campestre (A. C. Gabel #2189, BHSC; bar = 1 cm).
1�� Proceedings of the South Dakota Academy of Science, Vol. 90 (2011)
Fig. 9. Scanning electron micrograph of the peristome area of Tulostoma fimbriatum var. camp-estre (A. C. Gabel #2189, BHSC; bar = 500 µm).
Fig. 10. Scanning electron micrograph of Tulostoma fimbriatum var. campestre spores (A. C. Gabel #1106, BHSC; bar = 2 µm).





























