Embed Size (px)
Citation preview

ME63CH06-Picker ARI 12 December 2011 11:55
New Paradigms forHIV/AIDS VaccineDevelopmentLouis J. Picker,1 Scott G. Hansen,1
and Jeffrey D. Lifson2
1Vaccine and Gene Therapy Institute, Departments of Molecular Microbiology andImmunology and Pathology, and the Oregon National Primate Research Center,Oregon Health & Science University, Beaverton, Oregon 97006; email: [email protected] and Cancer Virus Program, SAIC Frederick, Inc., National CancerInstitute–Frederick, Frederick, Maryland 21702
Annu. Rev. Med. 2012. 63:95–111
First published online as a Review in Advance onSeptember 19, 2011
The Annual Review of Medicine is online atmed.annualreviews.org
This article’s doi:10.1146/annurev-med-042010-085643
Copyright c© 2012 by Annual Reviews.All rights reserved
0066-4219/12/0218-0095$20.00
Keywords
HIV/SIV, T cell vaccines, antibody vaccines, immune evasion,nonhuman primate models
Abstract
HIV-1 and its simian counterpart SIV have been exquisitely tailored byevolution to evade host immunity. By virtue of specific adaptations thatthwart individual innate or adaptive immune mechanisms, and an over-all replication strategy that provides for rapid establishment of a large,systemic viral population, capable of dynamic adaptation to almost allimmune selection pressures, these viruses, once established, almost in-variably stay one step ahead of the host’s immune system, and in the vastmajority of infected individuals, replicate indefinitely. Although manyvaccine approaches tested to date have been able to enhance the mag-nitude of the immune responses to HIV/SIV infection, most of theseresponses, whether cellular or humoral, have largely failed to be botheffectively antiviral and targeted to prevent the emergence of fully func-tional escape variants. Recent advances, however, have provided strongevidence that the initial stages of infection following mucosal transmis-sion of these viruses are more vulnerable to immune intervention, andhave led to the development of vaccine strategies that elicit responsesable to effectively intervene in these early stages of infection, eitherpreventing acquisition of infection or establishing early, stringent, anddurable control. Here, we place HIV/AIDS vaccine development in thecontext of the basic immunobiology of HIV and SIV, review the ev-idence for their vulnerability to immune responses immediately aftermucosal transmission, and discuss how this newly recognized vulnera-bility might be exploited for the development of an effective HIV/AIDSvaccine.
95
Ann
u. R
ev. M
ed. 2
012.
63:9
5-11
1. D
ownl
oade
d fr
om w
ww
.ann
ualr
evie
ws.
org
by U
nive
rsity
of
Cal
ifor
nia
- Sa
n D
iego
on
11/1
1/13
. For
per
sona
l use
onl
y.

ME63CH06-Picker ARI 12 December 2011 11:55
NHP: nonhumanprimate
INTRODUCTION
HIV-1 infections of humans and pathogenicSIV infections of nonhuman primates (NHPs)share a pattern of viral replication and aconstellation of pathologic features that in theabsence of antiretroviral therapy almost alwaysresult in unremitting infection and a progres-sive, ultimately fatal, immune deficiency (1–3).A striking feature of these pathologic lentiviralinfections is their induction of robust cellularand humoral immune responses, which failto clear or, in the vast majority of subjects,even effectively control viral replication (4,5). The AIDS-causing lentiviruses of humansand Asian NHPs derive from SIVs endemicin African NHP populations that typically donot cause disease in their native hosts (6, 7).However, this lack of pathogenicity is due notto the ability of these natural hosts to immuno-logically control SIV replication, but ratherto host adaptations that prevent the adverseeffects of high viral replication on the immunesystem (7). The prevailing view is that extendedcoevolution of these different viruses and theirvarious natural hosts has resulted in variationson the common theme that pathogenesis isavoided through adaptive mechanisms otherthan the achievement of effective immunecontrol of SIV replication. This is in strikingcontrast to the vast majority of viral pathogens,for which immune control of replicationplays the major role in preventing pathogenicconsequences (including even other persistentviruses, such as Herpes family viruses).
These observations (along with the absenceof precedent for natural immunity leadingto clearance of HIV and durable immuneprotection from super-infection) indicate thatdevelopment of an effective HIV/AIDS vaccinewill not consist of identifying an immuniza-tion approach that mimics protective naturalimmunity, as has been the path to successfor most vaccines, but rather will require thatvaccine developers achieve what nature couldnot: a generally protective anti-HIV immuneresponse (8). These considerations underscorethe difficulty in developing an HIV/AIDS
vaccine and largely account for the disappoint-ing progress in this development over the past25 years. However, the substantial investmentin basic and clinical research on the immuno-biology of these AIDS-causing lentiviruses isclearly paying off. These investigations havepainted an increasingly clear picture of themechanisms used by pathogenic lentivirusesto evade immunity, and most importantlyhave revealed that these viruses appear to havean “Achilles heel,” immune vulnerabilitiesthat may allow the host to prevent or controlinfection. Here, we review the strategies usedby HIV/SIV to avoid host immunity, as well asthe weaknesses in these strategies that mightbe exploited for development of an effectiveHIV/AIDS vaccine. We focus on the apparentwindow of viral vulnerability in the early stagesof infection following mucosal transmission.
IMMUNOBIOLOGY OFHIV/SIV INFECTION
A crucial and slowly learned lesson of nearlythree decades of HIV/SIV research has beenthe fact that the interaction of these viruseswith the host immune system frequentlydoes not follow the patterns established forother commonly studied viral pathogens. Allintracellular pathogens have to counter hostimmunity, especially innate immunity, toreplicate and manifest disease, but the breadthand depth of immune evasion achieved by HIVand SIV, given their comparatively meager≤10-kb genomes, are astonishing. Specific viraladaptations targeting innate immunity includethe viral vpu and vif genes countering the hostantiviral proteins tetherin and APOBECs, re-spectively (9). Cytotoxic T lymphocyte (CTL)responses are subverted by nef-mediateddownregulation of HLA-A and HLA-B, whichprevents CTL recognition of infected cells.Notably, HLA-C expression is not down-regulated in such infected cells, preventingtheir efficient killing by natural killer cells (9).Humoral immune responses are counteredby specific adaptations in the viral enve-lope glycoprotein (Env) that prevent the vast
96 Picker · Hansen · Lifson
Ann
u. R
ev. M
ed. 2
012.
63:9
5-11
1. D
ownl
oade
d fr
om w
ww
.ann
ualr
evie
ws.
org
by U
nive
rsity
of
Cal
ifor
nia
- Sa
n D
iego
on
11/1
1/13
. For
per
sona
l use
onl
y.

ME63CH06-Picker ARI 12 December 2011 11:55
TEM: effectormemory T cell(s)
majority of anti-Env antibodies from mediatingneutralization, or even recognizing native Envspikes on the HIV/SIV virion surface (10–12).However, although each of these specificmechanisms makes an important contributionto HIV/SIV immune evasion, they are butpieces of a larger overall strategy that startswith the transmission, target cell selection, andreplication mechanisms of these viruses.
Early research identified the CD4 moleculeas an entry receptor for HIV and SIV, ex-plaining the loss of CD4+ T cells noted ininfected subjects with AIDS (13). Depletionof this subset can subvert CD4+ T cell help,potentially leading to suboptimal antiviral im-mune responses that favor persistent viral repli-cation rather than host immune control. How-ever, maximal CD4+ T cell depletion typicallyoccurs after failure of viral control, and initialadaptive immune responses, particularly CD8+
T cell responses, to infection tend to be ro-bust (4), suggesting loss of CD4+ T cell help
does not solely account for the poor adaptiveimmune control of these viruses. The discov-eries that coreceptors are required for viral en-try and that transmitted forms of these virusesutilize CCR5 [a chemokine receptor involvedin T cell migration in effector/inflammatorysites (14)] as coreceptor (15) offered additionalinsight into the complex immunobiology ofthese infections. CCR5 coreceptor use, con-ferring “CCR5-tropism,” ingeniously focusesHIV/SIV infection on differentiated or differ-entiating effector memory CD4+ T cells (TEM)(Figure 1), an enormous, widely distributed,and readily replenished reservoir of potentialviral target cells (16, 17).
This population constitutes the majorCD4+ T cell population in gastrointestinaland genital mucosa, and thus the availability ofthese target cells in such mucosal tissues facil-itates efficient transmission and local spread ofinfection at these sites. The latter is augmentedby the recruitment of additional target cells
TCM
TEM
TCM TEM
“Transitional” effector memory
NaïveThymus
Central memory
Localization: 2° lymphoid tissues Lymphoid and extra-lymphoid effector sites
Expansion capacity: High Low
Antiviral effector function: Low High
HIV/SIV infectivity (CD4+ only): Low High
Effector memory
Ag
Ag
Inflammation
CCR5 expression
Figure 1Schema of peripheral T cell differentiation. The figure shows the coordinate changes in T cell homingbehavior, expansion capacity, and effector function that occur with progressive differentiation in theperipheral immune system. Straight arrows indicate a linear differentiation sequence. Circular arrowsindicate a population with independently regulated, secondary lymphoid tissue–based homeostasis.Because of the CCR5 tropism of transmitted strains of HIV/SIV and the preferential expression ofCCR5 in later stages of memory differentiation, this differentiation process is fundamental to HIV/SIVimmunopathogenesis, providing for selective targeting of later-stage (TEM) CD4+ memory cells whilesparing a progenitor population (TCM) for continued production of new targets. Mechanisms of memoryT cell differentiation are equally important to the activity of vaccine-elicited T cell responses, as theydetermine whether such responses predominantly localize in lymphoid tissue and respond to infection withexpansion, effector differentiation, and delayed migration to effector sites (TCM), or in extra-lymphoideffector sites, allowing immediate interception of transmitted virus in such sites (TEM). Abbreviations:Ag, antigen; CCR5, C-C chemokine receptor 5; HIV, human immunodeficiency virus; SIV, simianimmunodeficiency virus; TCM, central memory T cell; TEM, effector memory T cell.
www.annualreviews.org • New Paradigms for HIV/AIDS Vaccine Development 97
Ann
u. R
ev. M
ed. 2
012.
63:9
5-11
1. D
ownl
oade
d fr
om w
ww
.ann
ualr
evie
ws.
org
by U
nive
rsity
of
Cal
ifor
nia
- Sa
n D
iego
on
11/1
1/13
. For
per
sona
l use
onl
y.
ME63CH06-Picker ARI 12 December 2011 11:55
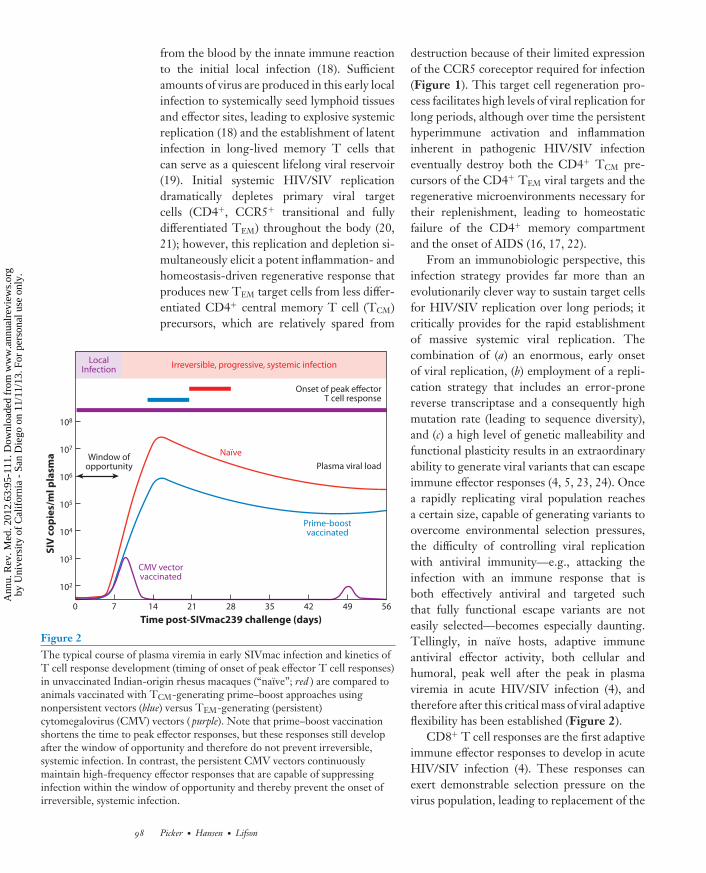
from the blood by the innate immune reactionto the initial local infection (18). Sufficientamounts of virus are produced in this early localinfection to systemically seed lymphoid tissuesand effector sites, leading to explosive systemicreplication (18) and the establishment of latentinfection in long-lived memory T cells thatcan serve as a quiescent lifelong viral reservoir(19). Initial systemic HIV/SIV replicationdramatically depletes primary viral targetcells (CD4+, CCR5+ transitional and fullydifferentiated TEM) throughout the body (20,21); however, this replication and depletion si-multaneously elicit a potent inflammation- andhomeostasis-driven regenerative response thatproduces new TEM target cells from less differ-entiated CD4+ central memory T cell (TCM)precursors, which are relatively spared from
Window ofopportunity
Irreversible, progressive, systemic infectionLocal
Infection
Onset of peak effectorT cell response
Plasma viral load
Naïve
Prime-boostvaccinated
CMV vectorvaccinated
SIV
co
pie
s/m
l p
lasm
a
Time post-SIVmac239 challenge (days)
0
102
103
104
105
106
107
108
7 14 21 28 35 42 49 56
Figure 2The typical course of plasma viremia in early SIVmac infection and kinetics ofT cell response development (timing of onset of peak effector T cell responses)in unvaccinated Indian-origin rhesus macaques (“naıve”; red ) are compared toanimals vaccinated with TCM-generating prime–boost approaches usingnonpersistent vectors (blue) versus TEM-generating (persistent)cytomegalovirus (CMV) vectors ( purple). Note that prime–boost vaccinationshortens the time to peak effector responses, but these responses still developafter the window of opportunity and therefore do not prevent irreversible,systemic infection. In contrast, the persistent CMV vectors continuouslymaintain high-frequency effector responses that are capable of suppressinginfection within the window of opportunity and thereby prevent the onset ofirreversible, systemic infection.
destruction because of their limited expressionof the CCR5 coreceptor required for infection(Figure 1). This target cell regeneration pro-cess facilitates high levels of viral replication forlong periods, although over time the persistenthyperimmune activation and inflammationinherent in pathogenic HIV/SIV infectioneventually destroy both the CD4+ TCM pre-cursors of the CD4+ TEM viral targets and theregenerative microenvironments necessary fortheir replenishment, leading to homeostaticfailure of the CD4+ memory compartmentand the onset of AIDS (16, 17, 22).
From an immunobiologic perspective, thisinfection strategy provides far more than anevolutionarily clever way to sustain target cellsfor HIV/SIV replication over long periods; itcritically provides for the rapid establishmentof massive systemic viral replication. Thecombination of (a) an enormous, early onsetof viral replication, (b) employment of a repli-cation strategy that includes an error-pronereverse transcriptase and a consequently highmutation rate (leading to sequence diversity),and (c) a high level of genetic malleability andfunctional plasticity results in an extraordinaryability to generate viral variants that can escapeimmune effector responses (4, 5, 23, 24). Oncea rapidly replicating viral population reachesa certain size, capable of generating variants toovercome environmental selection pressures,the difficulty of controlling viral replicationwith antiviral immunity—e.g., attacking theinfection with an immune response that isboth effectively antiviral and targeted suchthat fully functional escape variants are noteasily selected—becomes especially daunting.Tellingly, in naıve hosts, adaptive immuneantiviral effector activity, both cellular andhumoral, peak well after the peak in plasmaviremia in acute HIV/SIV infection (4), andtherefore after this critical mass of viral adaptiveflexibility has been established (Figure 2).
CD8+ T cell responses are the first adaptiveimmune effector responses to develop in acuteHIV/SIV infection (4). These responses canexert demonstrable selection pressure on thevirus population, leading to replacement of the
98 Picker · Hansen · Lifson
Ann
u. R
ev. M
ed. 2
012.
63:9
5-11
1. D
ownl
oade
d fr
om w
ww
.ann
ualr
evie
ws.
org
by U
nive
rsity
of
Cal
ifor
nia
- Sa
n D
iego
on
11/1
1/13
. For
per
sona
l use
onl
y.

ME63CH06-Picker ARI 12 December 2011 11:55
founder virus sequence with escape variants. Insome infected individuals, typically a subset ofthose with “protective” major histocompatibil-ity complex (MHC) class I alleles that directCD8+ T cell targeting to conserved regions ofviral gene products (particularly Gag), this canresult in selection of viral variants with signif-icantly reduced replicative fitness, facilitatingvirologic control (25–27). Such individuals canmanifest long-term “elite” virologic control(28), but this outcome is uncommon in bothhumans and NHPs. In most infected subjects,antiviral CD8+ T cell responses are noteffective; either they select for viral variantswith no apparent fitness defect, or they are notassociated with sequence changes at all (4, 29),suggesting that the responses have not exertedbiologically meaningful selection pressure.Such “ignored” responses may representdysfunctional CD8+ T cells (30) or inadequateT cell receptor avidity (5), but more likely theysimply reflect an inability of these responses toachieve high enough effector-to-target ratiosto impact viral replication (18).
The situation with regard to antiviralantibody responses in primary infection iseven bleaker. Owing to intrinsic characteristicsof the Env protein (10–12), antibodies thatcan neutralize autologous virus do not beginto appear until after ∼12 weeks of infection(4). These antibody responses can also exertsufficient selection pressure to lead to escapemutations, and indeed the typical course ofinfection is characterized by the developmentof antibody responses capable of neutralizingvirus, selection of escape variants, development(after a delay) of new responses capable ofneutralizing the new escape variants, andselection of new escape variants, with the hostalways playing catch-up (31, 32). The host’simmunologic pressure appears in most cases tohave little effect on the course of infection (33).
Thus, perhaps the most critical adaptivecharacteristic of HIV/SIV pathobiology isthe speed and dynamism of these viruses(in ironic contrast to their group name oflentiviruses, i.e., “slow viruses,” based on theirtypically protracted clinical course) in both the
ability to rapidly establish a large, geneticallyflexible, adaptive viral population before theonset of effective antiviral immunity, and theresulting ability to rapidly generate replication-competent viral variants in response to envi-ronmental selection pressures. This strategyalmost invariably keeps the virus one stepahead of the host, and the infection permanent.
LESSONS FROM VACCINES:FAILURES OF PROTECTIONAND PARTIAL SUCCESSES
Although the de novo T cell response toprimary HIV/SIV infection brings “too little”antiviral activity to sites of viral replication “toolate” to control infection in most naıve individ-uals (34), it remains possible that a prophylacticvaccine capable of eliciting memory responsesthat respond to HIV/SIV infection with earlier,stronger, and more broadly targeted CD8+ Tcell effector responses than develop in naıvesubjects might more effectively suppress viralreplication in infected individuals. A vaccineof this type would confer both individual andpopulation benefits by limiting pathogenesisand reducing the likelihood of transmission(8, 35). Such anamnestic T cell responseswould not be expected to prevent infection,but by applying stronger immune pressure tomore viral sequences (or, by virtue of targetingmore viral sequences, having a higher likeli-hood of targeting sequences critical to viralfitness) might reduce viral replication by directsuppression or by forcing genetic changesthat compromise viral replication efficiency.Such suppression would be enhanced if theseT cell responses were also able to interceptthe infection prior to the massive systemicreplication that facilitates immune escape.
This concept has guided development ofT-cell-response-targeted HIV/AIDS vaccinesover the past decade and has been extensivelyevaluated in various NHP AIDS models usingincreasingly potent DNA and viral vector im-munogens and combinations of these vectors invarious prime–boost combinations. These vac-cines have typically used replication-impaired
www.annualreviews.org • New Paradigms for HIV/AIDS Vaccine Development 99
Ann
u. R
ev. M
ed. 2
012.
63:9
5-11
1. D
ownl
oade
d fr
om w
ww
.ann
ualr
evie
ws.
org
by U
nive
rsity
of
Cal
ifor
nia
- Sa
n D
iego
on
11/1
1/13
. For
per
sona
l use
onl
y.

ME63CH06-Picker ARI 12 December 2011 11:55
or replication-incompetent viral vectors(particularly vectors based on poxviruses andadenoviruses) that safely provide high-level,but relatively transient, exposure to viral anti-gens, along with the necessary innate immunestimulation, to elicit potent conventionalCD8+ T cell memory responses—that is,TCM-biased populations capable of robustexpansion and effector cell production afterantigen restimulation (Figure 1).
Although the results of these studies varysomewhat with the vaccine approaches, NHPspecies, and challenge viruses utilized, the ex-perience is now extensive enough that a numberof consistent themes have emerged. First, thebest of these vaccines, particularly prime–boostcombination vaccines, can in fact dramaticallyenhance the magnitude of antiviral CD8+ Tcell responses after infection (often >10-fold)(36–45). Moreover, these anamnestic responsesgenerally peak somewhat earlier than primaryresponses in naıve animals, although these peakresponses still follow, rather than precede,peak viral replication (Figure 2). Second,the most effective regimens can also reducepeak and early chronic phase viral loads andextend median survival after challenge withhighly pathogenic, CCR5-tropic SIV (SIVmacand SIVsm derivatives), the challenge modelsthought to best recapitulate HIV infectionof humans (35, 46). However, the protectionobserved with these vaccines has been found tobe uneven within identically vaccinated rhesusmonkey cohorts. Protection is often correlatedwith protective MHC alleles, and, for SIVsm-derived viruses, TRIM5 alleles that influencetarget cell permissiveness for these viruses.The protective effect is usually limited to anapproximately 1.5–2-log median reduction inpeak and plateau phase plasma viral loads withSIVmac challenge and is subject to loss ofcontrol over time (36–45, 47–50). This patternof heterogeneous outcomes, with most vaccinesmanifesting limited and/or transient protec-tion, has been observed with both intravenousand mucosal challenges [the latter includingboth high (single) and low (repeated) doses].This is the pattern expected of an antiviral
response that depends on anamnestic expan-sion, effector differentiation, and homing ofCD8+ memory T cells, and therefore can act tocontain the infection only after systemic spreadand massive viral replication have alreadyoccurred.
Arguably, infection with the highlypathogenic, CCR5-tropic SIV challengeused in these NHP vaccine efficacy studies(especially SIVmac-derived challenge viruses)is more aggressive than typical clinical HIVinfections, attaining peak plasma viremia abouta week earlier and manifesting 10–100-foldhigher viral replication rates at set point.This difference raises the possibility that suchmodels might represent an unduly stringentchallenge for vaccines, and that these vaccineapproaches might be more effective againstHIV infection in humans than in SIVmacinfection in macaques. Unfortunately, this wasnot the case in the first clinical assessment ofthis CD8+ memory T cell vaccine concept,the phase IIb Merck STEP trial (HVTN502), which used Adenovirus (Ad) 5 vectorsexpressing HIV Gag, Nef, and Pol. Despitedetection of HIV-specific CD8+ T cells in73% of vaccinated subjects, this trial gave aclear negative result—there was no evidenceof vaccine-induced protection in terms of pre-venting acquisition of infection or facilitatingcontrol of postinfection HIV replication in thevaccinated group (51, 52). Post-hoc analysis re-vealed a statistically significant “sieving” effectof vaccine-elicited responses on the sequenceof the viruses that established infection invaccinees (53), suggesting that vaccine-elicitedT cells did exert weak selection pressure, par-ticularly in individuals with protective MHC Ialleles, but this was much less than was hopedfor with this vaccine. In a further complication,the results raised the question of whether thevaccine might have increased the incidence ofinfection in uncircumcised men (54).
The vaccine used in the STEP trial (whichconsisted of Ad5 vectors only, not the morepotent prime–boost approach) appears, in ret-rospect, to have been insufficiently potent inboth the magnitude and breadth of responses
100 Picker · Hansen · Lifson
Ann
u. R
ev. M
ed. 2
012.
63:9
5-11
1. D
ownl
oade
d fr
om w
ww
.ann
ualr
evie
ws.
org
by U
nive
rsity
of
Cal
ifor
nia
- Sa
n D
iego
on
11/1
1/13
. For
per
sona
l use
onl
y.

ME63CH06-Picker ARI 12 December 2011 11:55
induced to achieve significant protection (55). Itis noteworthy that analogous “Ad5 vector only”vaccines did not provide significant protectionin NHPs against intrarectal SIVmac (44) orSIVsmE660 challenge (M. Reynolds and D.Watkins, personal communication). Althoughit remains possible that a more potent prime–boost regimen would yield better results in ahuman trial, taken together, the NHP and hu-man data illustrate the difficulty in attaining ef-ficacy against HIV/SIV with a vaccine approachdesigned to elicit protection via an anamnes-tic CD8+ memory T cell response, as even thebest of these responses still confront HIV/SIVinfection only after its systemic establishment,and therefore must contend with immune eva-sion enabled by an established virus populationcapable of extensive adaptive variation.
The development of HIV/AIDS vaccinestargeting humoral immunity has encounteredequally formidable obstacles. The initialcharacterization of HIV-1 as a retrovirus withEnv protein–mediated entry brought highhopes that an Env-based vaccination approachwould yield neutralizing antibodies that wouldprovide protection against either acquisitionor progression of infection. These hopes werespurred on by the relative susceptibility oflaboratory passaged HIV strains to neutraliza-tion by anti-Env antibodies. However, it wassubsequently determined that clinical isolateswere highly resistant to neutralization andthat in fact the vast majority of antibodieselicited by immunization with monomeric Envproteins were directed at “decoy” epitopeswith little or no neutralization activity (10–12).As might be expected from these findings, twolarge, well-conducted phase III efficacy trials(VAX003 and VAX004) of alum-adjuvantedEnv protein (gp120) showed no efficacy againstHIV-1 acquisition or postinfection viremia(56, 57). Additional studies have demonstratedthat neutralization-sensitive epitopes (con-served functional parts of the Env protein) areeffectively concealed in native Env trimers andare poorly immunogenic. Epitopes presentedby denatured Env dominate the anti-Envresponse but do not bind the critical regions
of the timeric Env spikes of infectious virionsrequired for neutralization (10–12, 58). Asindicated above, HIV-infected individuals dostart making neutralizing antibodies to theirautologous virus after ∼12 weeks of infection,and ∼20% of infected individuals eventuallydevelop HIV-specific antibodies capable ofneutralizing heterologous HIV strains (4), withrare individuals manifesting antibodies thatrecognize conserved regions of Env capable ofbroad neutralization (59). Although these an-tibodies, developing relatively late in infection,confer little or no benefit to these individualsowing to viral escape (33), monoclonal antibod-ies with broadly neutralizing activity can, whenpresent in appropriate concentration at the timeof challenge, completely protect NHPs fromchallenge with pathogenic SHIV (SIV-HIVhybrid virus) expressing HIV Env (60, 61). Todate, however, despite extensive effort, no im-munogen/vaccine approach has been capable ofreliably eliciting such broadly neutralizing anti-bodies, and the prospects for such immunogensremain uncertain. Thus, HIV/AIDS vaccinestargeting humoral immunity have beenstymied by the inability to achieve responseswith sufficient antiviral activity, as defined bybroad, high-titer neutralization activity.
IMMUNE VULNERABILITIESREVEALED
These considerations suggest that develop-ment of vaccines that prevent infection orconsistently mediate effective immune controlof HIV/SIV will be very difficult to achieve.Indeed, in the wake of the negative results fromthe STEP trial, there were calls from somequarters for abandoning vaccine developmentin favor of pursuing other HIV preventionstrategies. However, more recent developmentsprovide evidence of effective, vaccine-elicitedanti-HIV/SIV immunity and point to brighterprospects for the development of an effectivevaccine. Foremost among these developmentsis the outcome of the RV144 AIDS vaccine trial,which evaluated the ability of a prime–boostvaccination versus placebo to protect against
www.annualreviews.org • New Paradigms for HIV/AIDS Vaccine Development 101
Ann
u. R
ev. M
ed. 2
012.
63:9
5-11
1. D
ownl
oade
d fr
om w
ww
.ann
ualr
evie
ws.
org
by U
nive
rsity
of
Cal
ifor
nia
- Sa
n D
iego
on
11/1
1/13
. For
per
sona
l use
onl
y.

ME63CH06-Picker ARI 12 December 2011 11:55
HIV infection in 16,402 relatively low-risk sub-jects in Thailand (62). The vaccinated groupreceived a canarypox virus prime (ALVAC-HIV) and an Env protein (gp120 subunit)boost. Compared to the placebo group, thevaccinated group manifested a 31% reductionin infection acquisition, which was significantat a p-value of 0.04 (modified intent-to-treatanalysis). The apparent vaccination-associatedefficacy seemed to be greatest in the first yearafter vaccination and then decreased thereafter.In study subjects who were infected with HIV,there was no difference between vaccinees andcontrols with respect to viral load. Immunolog-ically, this vaccine approach elicited strong, buttransient, Env-specific antibody responses andEnv-specific CD4+ T cell lymphoproliferativeresponses, but no significant HIV-specificCD8+ T cell responses (55, 58, 62). TheEnv-specific antibody responses includedneutralization of tissue-culture-adapted virusesand the ability to mediate antibody-dependentcell-mediated cytotoxicity, but not the broad,potent neutralization of transmitted HIVstrains previously thought to be required formeaningful protection.
Although the immunologic correlate orcorrelates of the modest protection observedin the RV144 trial have not been preciselydetermined, the nature of protection (againstacquisition of infection only, without controlof virus postacquisition) and the nature of theelicited responses (humoral immune domi-nated) strongly suggest that antibody-mediatedmechanisms may be responsible. This hypothe-sis is in keeping with an increasing body of datashowing the ability of antibodies to mediateanti-HIV/SIV activity by mechanisms otherthan potent, broad virus neutralization, in-cluding antibody-dependent cell-mediated cy-totoxicity, antibody-dependent cell-mediatedviral inhibition, antibody-mediated trappingof virions in mucus, or inhibition of viraltranslocation across epithelia (63–67).
Importantly, RV144-like protection againstacquisition of infection has been recapitulatedin the NHP models of infection. In the largestNHP study, analogous to the RV144 trial, a
prime–boost vaccination regimen protected∼50% of vaccinated monkeys from acquisi-tion of SIVsmE660 after repeated low-doseintrarectal challenge but had a minimal effecton peak viral loads of those animals that didbecome infected (50). Immunologic correlatesof this protection included low neutralizingantibody titers and Env-specific CD4+ T cellresponses. In another recent study, mucosalvaccination of NHPs with gp41 expressing vi-rosomes showed protection against acquisitionof SHIV infection after repeated, limiting-dose intravaginal challenge. This protectioncorrelated with antiviral IgAs in the vagina andoccurred in the absence of plasma IgGs withneutralization activity (68).
The significance of these observations lies inthe facts that (a) for the first time, a vaccine hasbeen shown to significantly, albeit modestly,reduce acquisition of HIV infection in the field;(b) such protection can be elicited by prime–boost vaccines and other approaches usingrelatively simple immunogens, almost certainlythrough an antibody-mediated mechanism;and (c) protection against infection acquisitionappears to be achievable without the broad,potent, transmitted-strain neutralization re-sponse that has been so difficult to achievewith vaccines. Although these results arequite promising, caveats include the facts that(a) the modest RV144 protection appeared tobe time limited and was achieved in a low-riskpopulation, and it may not be recapitulated inupcoming studies in higher-risk populationssuch as those in sub-Saharan Africa; and(b) the acquisition protection achieved withSIVsmE660 was not recapitulated in a parallelstudy employing repeated low-dose intrarectalchallenge of monkeys with a different SIVstrain (SIVmac251) given the same vaccine(50). Nevertheless, taken together, these dataindicate that significant protection againstinfection acquisition is attainable with vaccina-tion and strongly suggest that such protectioncan be mediated by antiviral Abs that lackbroadly neutralizing activity. A key implicationof this latter conclusion is that the immunologicrequirements for prevention or early control of
102 Picker · Hansen · Lifson
Ann
u. R
ev. M
ed. 2
012.
63:9
5-11
1. D
ownl
oade
d fr
om w
ww
.ann
ualr
evie
ws.
org
by U
nive
rsity
of
Cal
ifor
nia
- Sa
n D
iego
on
11/1
1/13
. For
per
sona
l use
onl
y.

ME63CH06-Picker ARI 12 December 2011 11:55
infection may be less stringent than previouslythought.
A second hopeful development forHIV/AIDS vaccine research is the demonstra-tion that early SIV infection at mucosal portalsof entry appears to be more vulnerable to Tcell–mediated control than the massive sys-temic replication characteristic of later stagesof infection. In this regard, it is now well doc-umented that sexually/mucosally transmittedHIV infections in humans and experimentalSIV infections in NHPs after limiting-dosemucosal challenge are typically initiated by oneor very few transmitted/founder viral variants,and that in vaginal SIV transmission modelsin NHPs, the establishment of systemic,progressive infection requires up to a week oflocal amplification and spread at the site ofinitial transmission before disseminated, sys-temic productive infection is established (18)(Figure 2). Conceptually, if vaccine-elicitedimmune responses were able to intercept thedeveloping infection in this early period (the“window of opportunity”), antiviral effectormechanisms would act on a much smaller, lo-calized, and less diverse viral population, whichis probably less able to evade T cell–mediatedsuppression.
As described above, almost all T cell–targeted HIV/SIV vaccines studied to date arebased on nonpersistent vectors and induce con-ventional (TCM-biased) CD8+ memory T cellresponses that depend on an anamnestic expan-sion to provide antiviral effectors in sufficientnumbers to combat infection (Figures 1 and2). Although for antibody-mediated vaccineapproaches it is assumed that long-lived, effec-tive antibody responses will be present at thetime of exposure and able to deal directly withthe incoming viral inoculum, the delay inherentin developing antiviral effectors from the TCM
populations induced by typical vaccines resultsin such anamnestic response-derived effectorsonly confronting primary HIV/SIV infectionafter systemic establishment of the infection.Thus, the responses elicited by most of the Tcell–based vaccines studied to date have beenunable to exploit the potential window of viral
vulnerability in early infection. Advances inour understanding of the biology of memoryT cells over the past decade have offered analternative approach: development of vaccinesthat establish and maintain high-frequency,tissue-based, functionally differentiated TEM,especially CD8+ TEM, at potential sites ofinfection (Figure 1). Because CD4+ TEM, theprimary targets of CCR5-tropic HIV/SIV,are invariably colocalized with their CD8+
counterparts, this strategy would provide thepotential to have in place antiviral effectorsin the immediate vicinity of the likeliestviral targets. This would enable immediateinterception, and potentially, suppression ofinfection by either cytolysis of infected cells orsuppression of viral spread by soluble factors.However, to the extent that virus-specificCD4+ TEM are also generated by a candidateTEM targeting vaccine, this approach does runthe risk of providing a higher frequency ofactivated target cells at sites of early infec-tion. Determination of the balance betweenthese potentially infection-suppressing andinfection-facilitating mechanisms, and thuspredicting the efficacy of TEM responsesin HIV/SIV infection, requires a vaccineapproach that can establish and maintain suchresponses.
TEM-biased responses are the hallmark ofpersistent pathogens, and therefore the testingof this concept necessitates a change in vaccinestrategy from vectors that provide high, buttransient, antigen exposure to vectors capableof providing lower, but persistent, levels ofantigen (Figure 1). The first evidence thatpersistent vectors might have a superior abilityto control pathogenic lentiviral infectioncame from analysis of live attenuated SIVvaccines. Although clinical development ofattenuated HIV vaccines is precluded by theinherent potential of an attenuated primatelentivirus for pathogenicity (69), the ability ofsuch vaccines to mediate stringent protectionagainst pathogenic SIV challenge, especiallyhomologous challenge, has been demonstratedby multiple groups (70, 71). Persistence of thevector and associated antigen exposure appear
www.annualreviews.org • New Paradigms for HIV/AIDS Vaccine Development 103
Ann
u. R
ev. M
ed. 2
012.
63:9
5-11
1. D
ownl
oade
d fr
om w
ww
.ann
ualr
evie
ws.
org
by U
nive
rsity
of
Cal
ifor
nia
- Sa
n D
iego
on
11/1
1/13
. For
per
sona
l use
onl
y.

ME63CH06-Picker ARI 12 December 2011 11:55
to be required for this type of protection,as too high a level of attenuation results invaccines that are considerably less effective (70,72). Because these attenuated SIV vectors alsoinfect CD4+ T cells and elicit both cellular andhumoral adaptive immunity, as well as innateimmunity, the mechanisms responsible fortheir high-level protection are hard to defineunambiguously and remain controversial, butpersistent SIV-specific T cell responses havebeen implicated in several studies (73–77).
The ability of persistent vectors and theirassociated TEM responses to mediate a quan-titatively and qualitatively unique pattern ofprotection consistent with control of early viralreplication has been more definitively shown bythe analysis of SIV protein-expressing vectorsbased on the persistent β-herpesvirus CMV[RhCMV in the rhesus macaque model (78)].RhCMV/SIV vectors can repeatedly superin-fect RhCMV+ monkeys in a clinically silentmanner, and, in the process of superinfection(unaffected by pre-existing CMV-specific im-munity), reliably elicit and indefinitely maintainhigh-frequency SIV-specific CD4+ and CD8+
TEM responses at potential sites of early viralreplication after mucosal challenge (49, 79, 80).RhCMV/SIV vector designs analyzed to dateelicit these TEM responses in the absence ofa significant anti-SIV antibody response, andthus appear to allow delineation of the efficacyof SIV-specific TEM without the potentiallyconfounding effects of humoral responses.
Strikingly, ∼50% of monkeys vaccinatedwith RhCMV/SIV vectors have manifestedearly, stringent control of intrarectally admin-istered highly pathogenic SIVmac. This pro-tection was characterized by an initial peak ofviremia of variable, but usually low, magnitude,followed by nearly immediate control to belowquantifiable levels (Figure 2). Although manyof the protected monkeys showed periodic, low-level “blips” of measurable viremia during thefirst 30 weeks of follow-up (with blips approxi-mately once every 6–7 weeks on average), theseblips gradually waned (49). Overall viral controlwas sufficiently early and stringent to precludeany measurable CD4+ target cell depletion,
as well as to prevent seroconversion to SIVEnv. This early-onset CMV-vector-mediatedprotection occurred without an anamnesticresponse, and, consistent with this, was “all ornone”—there was no peak or postpeak sup-pression of viral replication in nonprotectedmonkeys. There was also no measureable en-hancement of infection in these nonprotectedvaccinees, suggesting that vaccine-mediatedinduction of SIV-specific CD4+ T cells didnot increase the pathogenicity of the infection.
The outcome of challenge in CMV-vector-vaccinated monkeys (e.g., protection versusnonprotection) was predicted by peak SIV-specific CD8+ T cell response frequenciesmeasurable in peripheral blood during the vac-cine phase (prechallenge), which likely reflectsthe degree of seeding of SIV-specific TEM invarious effector sites, but was not predicted byCD8+ T cell responses associated with protec-tive MHC I alleles. Most significantly, stringentSIV control has been stable for >1 year inall but 1 of the 17 protected monkeys withlong-term follow-up (in association with stableRhCMV/SIV-vector-maintained, SIV-specificTEM response frequencies). Comprehensiveanalysis of lymphoid tissues and effector sites of4 long-term RhCMV/SIV-vector-vaccinatedcontrollers (necropsied after >1 year of con-trol) with ultrasensitive nested PCR/RT-PCRassays has only rarely detected SIV nucleicacid at levels that are >3 logs lower thanthose measured in rhesus macaques studied inparallel that had well-controlled SIVmac239infection, as assessed by conventional criteria(49). Moreover, all lymphoid tissues from theseRhCMV/SIV-vector-vaccinated controllerswere negative for “rescuable” SIV by coculture(versus easily coculturable SIV in conventionalcontrollers). Lastly, long-term CMV-vector-mediated protection was unperturbed by CD8+
lymphocyte depletion—in contrast to conven-tional protection, which is almost invariablydiminished (e.g., viral replication increased)by this treatment. These data demonstratean unprecedentedly low level of virus com-pared to any controlled or pharmacologicallysuppressed HIV or SIV infection reported to
104 Picker · Hansen · Lifson
Ann
u. R
ev. M
ed. 2
012.
63:9
5-11
1. D
ownl
oade
d fr
om w
ww
.ann
ualr
evie
ws.
org
by U
nive
rsity
of
Cal
ifor
nia
- Sa
n D
iego
on
11/1
1/13
. For
per
sona
l use
onl
y.

ME63CH06-Picker ARI 12 December 2011 11:55
date, and they raise the remarkable possibilitythat the SIV infection in RhCMV/SIV-vector-vaccinated controllers may ultimately becleared through ongoing immune surveillance.
The mechanisms by which CMV-vector-elicited, TEM-dominated immune responsesmediate this remarkable protection remain tobe precisely determined, but the ability of theseresponses to arrest, control, and perhaps evenclear highly pathogenic SIV infection prior toirreversible systemic infection provides strongevidence that lentiviral immune evasion capa-bility is limited when early-stage infection isconfronted by potent pre-existing antiviral ef-fector responses that do not require anamnes-tic expansion (49). A corollary of this premiseis that the mechanisms involved in mediatingelite HIV/SIV control and long-term nonpro-gression of established, chronic infection maynot be required, or may be distinct from thosenecessary for early control and therefore shouldnot serve as a strict guide for prophylactic vac-cine development.
PROSPECTS FOR AN HIV/AIDSVACCINE—REVISITED
With the negative result of the STEP trialand the realization that even the most potentvaccines designed to elicit conventional CD8+
memory T cell responses are unlikely toconfer elite-controller status to the majorityof infected vaccinees, the HIV/AIDS vaccinedevelopment field was left with no clear path-way to an effective vaccine—only the hope thatincreasingly detailed and sophisticated struc-tural analysis of the interaction between Envand a growing panel of broadly neutralizingmonoclonal antibodies isolated from HIV-infected patients would lead to advances in Envimmunogen design, and eventually to a vaccinecapable of eliciting high-titer, broadly neutral-izing antibodies (11, 12, 58). Although muchhas been learned about the epitopes recognizedby these different broadly neutralizing antibod-ies, applying this information on antigenicityto the development of useful immunogens ca-pable of inducing similar responses has proven
Stages of primary infection Vaccine intervention
Abs
TEM responses
(CMV vectors)
TCM responses
(nonpersistent vectors)
Local replication at portalof entry
Step 2
Mucosal translocation andinitial infection
Step 1
Generalized systemicreplication
Step 4
Early spread (amplificationand broadcast of infection)
Step 3
Figure 3Stages of early HIV/SIV infection and the point of effective intercept of thedesignated vaccine approaches.
an especially daunting challenge. There is littleargument over the importance of fully explor-ing this strategy, but at the time of this writing,the outcome of this effort remains uncertain.
Recent data offer a new approach toHIV/AIDS vaccine development based on ex-ploiting the immune vulnerabilities of the virusduring the early stage of infection (Figure 3).Mucosal infection might be prevented by moreprosaic antibody-targeted vaccines that induceantibody responses capable of binding to a virusor infected cell, but not necessarily capable ofbroadly neutralizing activity. Alternatively, mu-cosal infection might be prevented by stringentcontrol of infection through TEM-generatingvaccines. Although neither of these vaccinestrategies is sufficiently optimized for clinicaluse in its current form, it is not unreasonable tosuggest that both empirical optimization andrational design can improve the efficacy of eachapproach, and that the combination of thesedisparate and independent approaches mightresult in additive or potentially synergistic in-creases in overall efficacy. The RV144 vaccineapproach might be empirically improved by op-timization of priming vectors, their Env in-serts, and the Env protein immunogens usedin the boost, as well as the use of more po-tent adjuvants with the protein boost (58, 81).
www.annualreviews.org • New Paradigms for HIV/AIDS Vaccine Development 105
Ann
u. R
ev. M
ed. 2
012.
63:9
5-11
1. D
ownl
oade
d fr
om w
ww
.ann
ualr
evie
ws.
org
by U
nive
rsity
of
Cal
ifor
nia
- Sa
n D
iego
on
11/1
1/13
. For
per
sona
l use
onl
y.

ME63CH06-Picker ARI 12 December 2011 11:55
Obviously, novel immunogens derived fromstructure–function analysis of broadly neutral-izing monoclonal antibodies could be incorpo-rated into these antibody-targeted prime–boostdesigns as they are shown to be effective. Thisoptimization might be accomplished using bothNHP models and adaptive clinical trials (82).
The TEM vaccine concept, which, to date,has only been tested in an NHP SIV model, hasa more complex development path. CMV vec-tors, the prototype TEM vaccine approach, arespecies-specific, and therefore the translation ofthese vectors to the clinic requires developmentof human CMV/HIV vectors based on RhCMVdesigns validated in the NHP model. Althoughthe rhesus and human forms of CMV are ge-netically distinct, they are closely related, andthere is sufficient functional homology amongkey genes to translate design concepts fromthe former to the latter (78). Although CMVinfection is ubiquitous and nonpathogenic inthe vast majority of infected individuals, other-wise wildtype CMV vectors would pose a possi-ble risk to certain vulnerable populations, suchas pregnant women and CMV-naıve individu-als with unsuspected immune deficiency. Thesevectors would therefore not be ideal candidatesfor inclusion in a prophylactic vaccine. How-ever, in both NHP and mouse models, CMVimmunogenicity does not depend on full repli-cation competence; genetically modified CMVconstructs with an inability or a highly reducedability to spread after the initial round of in-fection are capable of eliciting and maintaininghigh-frequency TEM responses that are essen-tially indistinguishable from wildtype responses(83; L. Picker, S. Hansen, and K. Frueh, unpub-lished data). This capability likely reflects theability of CMV-infected cells to avoid immuneelimination and persist for prolonged periodsdespite ongoing antigen expression, a biologythat strongly favors the development of safe, yethighly immunogenic, CMV vectors. Such vec-tors might be used subsequent to a heterologousprime, potentially increasing overall immuno-genicity while retaining the TEM character ofthe elicited responses.
Given variability in the magnitude, qual-ity, and duration of vaccine-elicited immuneresponses in humans and both the immuneevasion capabilities and diversity of HIV, itis unlikely that any single vaccine approach—not even the HIV/AIDS vaccinologist’s “HolyGrail” of an approach that elicits potent,durable, broadly neutralizing antibodies—willbe effective (i.e., prevent or stringently con-trol infection) in all potential transmissions.And, of course, when infection occurs in theface of significant immunologic pressure thatdoes not confer solid protection, it almost in-evitably leads to immune escape, and poten-tially, generation of transmissible viruses thatare no longer sensitive to the involved immuno-logic mechanism. Strategies such as mosaic vac-cine insert/immunogen designs can broadenvaccine-elicited immune responses, and helpovercome the sequence diversity of transmittedHIV strains (58), but a more general solutionto this issue may lie in the development of mul-timodal vaccines that target different immunevulnerabilities (much like the need for multiple,differentially targeted antiretroviral drugs in ef-fective combination antiretroviral chemother-apy regimens).
This strategy is subtly different from theconventional-wisdom mantra that an optimallyeffective AIDS vaccine should induce bothhumoral and cellular immunity, in the hopethat one arm of the adaptive immune systemcan confront the fraction of virus not effectivelydealt with by the other. Here, we suggest com-bining independent vaccine elements (designedto be noninterfering) that will work togetherin a strategically complementary fashion forenhanced overall efficacy (Figure 3). Forexample, an optimized prime–boost regimenfocused on HIV Env and designed to generateacquisition-blocking antibodies might be com-bined with CMV vectors focused on the rest ofthe HIV proteome and designed to elicit long-lasting TEM-based cellular responses for earlycontrol and long-term immune surveillance ofany residual infection. If the former compo-nent is 50% effective in blocking acquisition,
106 Picker · Hansen · Lifson
Ann
u. R
ev. M
ed. 2
012.
63:9
5-11
1. D
ownl
oade
d fr
om w
ww
.ann
ualr
evie
ws.
org
by U
nive
rsity
of
Cal
ifor
nia
- Sa
n D
iego
on
11/1
1/13
. For
per
sona
l use
onl
y.

ME63CH06-Picker ARI 12 December 2011 11:55
and the latter component is 50% effectivein stringently controlling early infection, theoverall efficacy, without synergy, would be arespectable 75%. However, it is plausible thatsynergy would occur—for example, amongthose vaccinees that acquire infection, the Env-specific antibody response might decrease thenumber of infective foci and/or hamper earlycell-to-cell transmission, and thereby increasethe likelihood that CMV-vector-generatedTEM would effectively control the infection. Inaddition, to the extent that initial protection isnonsterilizing, the ability of CMV vectors tomaintain high-frequency TEM over the longterm could potentially clear residual infectionor subject any rebound infection to stringentcontrol, even if the antibody componenteventually waned.
In conclusion, continued basic and clinicalresearch on HIV/AIDS immunobiology and
vaccinology has, in the wake of the disappoint-ing outcome of the STEP trial, reinvigoratedHIV/AIDS vaccine development. Coupledwith the surprising positive results from theRV144 study, this work has unequivocallyshown that the initial take of HIV/SIV infec-tion after mucosal exposure and the early viralreplication prior to irreversible disseminatedinfection are more vulnerable to immune inter-vention than previously appreciated. Vaccinesthat can place appropriate immune effectorresponses at these early sites appear to providemeaningful protection. Although there remainsmuch work to be done to optimize these ap-proaches and translate this information into li-censable vaccines, the HIV/AIDS vaccine field,for the first time, has a pathway to follow thatis based on solid observations of efficacy andthe foundation of an increasingly sophisticatedunderstanding of lentiviral immunobiology.
DISCLOSURE STATEMENT
The authors declare the following competing financial interests: Oregon Health & ScienceUniversity has licensed CMV vector technology, for which L.J.P. and S.G.H. are inventors, tothe International AIDS Vaccine Initiative (IAVI).
ACKNOWLEDGMENTS
L.J.P. and S.G.H. acknowledge support from the National Institutes of Health (AI060392,AI095113, DE021291, RR00163); the International AIDS Vaccine Initiative (IAVI) and its donors,particularly the United States Agency for International Development (USAID); and the Collabo-ration for AIDS Vaccine Discovery (CAVD), supported by the Bill & Melinda Gates Foundation.J.D.L. acknowledges support from the National Cancer Institute (#HHSN261200800001E).
LITERATURE CITED
1. Grovit-Ferbas K, Pappas T, O’Brien WA. 1999. Human immunodeficiency virus. In Persistent ViralInfections, ed. R Ahmed, AI Chen, pp. 3–45. Chichester, UK: Wiley
2. Cohen OJ, Fauci AS. 2001. Pathogenesis and medical aspects of HIV-1 infection. In Fields Virology, ed.DM Knipe, PM Howley, pp. 2043–94. Philadelphia: Lippincott Williams & Wilkins. 4th ed.
3. Hirsch VM, Lifson JD. 2000. Simian immunodeficiency virus infection of monkeys as a model system forthe study of AIDS pathogenesis, treatment, and prevention. Adv. Pharmacol. 49:437–77
4. McMichael AJ, Borrow P, Tomaras GD, et al. 2010. The immune response during acute HIV-1 infection:clues for vaccine development. Nat. Rev. Immunol. 10:11–23
5. O’Connor DH, Allen TM, Vogel TU, et al. 2002. Acute phase cytotoxic T lymphocyte escape is a hallmarkof simian immunodeficiency virus infection. Nat. Med. 8:493–99
www.annualreviews.org • New Paradigms for HIV/AIDS Vaccine Development 107
Ann
u. R
ev. M
ed. 2
012.
63:9
5-11
1. D
ownl
oade
d fr
om w
ww
.ann
ualr
evie
ws.
org
by U
nive
rsity
of
Cal
ifor
nia
- Sa
n D
iego
on
11/1
1/13
. For
per
sona
l use
onl
y.

ME63CH06-Picker ARI 12 December 2011 11:55
6. Hahn BH, Shaw GM, De Cock KM, et al. 2000. AIDS as a zoonosis: scientific and public health impli-cations. Science 287:607–14
7. Brenchley JM, Silvestri G, Douek DC. 2010. Nonprogressive and progressive primate immunodeficiencylentivirus infections. Immunity 32:737–42
8. Walker BD, Burton DR. 2008. Toward an AIDS vaccine. Science 320:760–649. Kirchhoff F. 2010. Immune evasion and counteraction of restriction factors by HIV-1 and other primate
lentiviruses. Cell Host Microbe 8:55–6710. Burton DR, Stanfield RL, Wilson IA. 2005. Antibody versus HIV in a clash of evolutionary titans.
Proc. Natl. Acad. Sci. USA 102:14943–4811. Mascola JR, Montefiori DC. 2010. The role of antibodies in HIV vaccines. Annu. Rev. Immunol. 28:413–4412. Hoxie JA. 2010. Toward an antibody-based HIV-1 vaccine. Annu. Rev. Med. 61:135–5213. Levy JA. 1993. Pathogenesis of human immunodeficiency virus infection. Microbiol. Rev. 57:183–28914. Kunkel EJ, Boisvert J, Murphy K, et al. 2002. Expression of the chemokine receptors CCR4, CCR5, and
CXCR3 by human tissue-infiltrating lymphocytes. Am. J. Pathol. 160:347–5515. Simmons G, Reeves JD, Hibbitts S, et al. 2000. Co-receptor use by HIV and inhibition of HIV infection
by chemokine receptor ligands. Immunol. Rev. 177:112–2616. Grossman Z, Meier-Schellersheim M, Paul WE, et al. 2006. Pathogenesis of HIV infection: what the
virus spares is as important as what it destroys. Nat. Med. 12:289–9517. Grossman Z, Picker LJ. 2008. Pathogenic mechanisms in simian immunodeficiency virus infection.
Curr. Opin. HIV AIDS 3:380–8618. Haase AT. 2011. Early events in sexual transmission of HIV and SIV and opportunities for interventions.
Annu. Rev. Med. 62:127–3919. Chun TW, Engel D, Berrey MM, et al. 1998. Early establishment of a pool of latently infected, resting
CD4+ T cells during primary HIV-1 infection. Proc. Natl. Acad. Sci. USA 95:8869–7320. Mattapallil JJ, Douek DC, Hill B, et al. 2005. Massive infection and loss of memory CD4+ T cells in
multiple tissues during acute SIV infection. Nature 434:1093–9721. Picker LJ, Hagen SI, Lum R, et al. 2004. Insufficient production and tissue delivery of CD4+ memory
T cells in rapidly progressive simian immunodeficiency virus infection. J. Exp. Med. 200:1299–31422. Zeng M, Smith AJ, Wietgrefe SW, et al. 2011. Cumulative mechanisms of lymphoid tissue fibrosis and
T cell depletion in HIV-1 and SIV infections. J. Clin. Invest. 121:998–100823. Bailey J, Blankson JN, Wind-Rotolo M, et al. 2004. Mechanisms of HIV-1 escape from immune responses
and antiretroviral drugs. Curr. Opin. Immunol. 16:470–7624. Boutwell CL, Rolland MM, Herbeck JT, et al. 2010. Viral evolution and escape during acute HIV-1
infection. J. Infect. Dis. 202(Suppl. 2):S309–1425. Goulder PJ, Watkins DI. 2008. Impact of MHC class I diversity on immune control of immunodeficiency
virus replication. Nat. Rev. Immunol. 8:619–3026. Dinges WL, Richardt J, Friedrich D, et al. 2010. Virus-specific CD8+ T-cell responses better define HIV
disease progression than HLA genotype. J. Virol. 84:4461–6827. Martins MA, Wilson NA, Reed JS, et al. 2010. T-cell correlates of vaccine efficacy after a heterologous
simian immunodeficiency virus challenge. J. Virol. 84:4352–6528. Walker BD. 2007. Elite control of HIV infection: implications for vaccines and treatment. Top. HIV Med.
15:134–3629. Goonetilleke N, Liu MK, Salazar-Gonzalez JF, et al. 2009. The first T cell response to transmitted/founder
virus contributes to the control of acute viremia in HIV-1 infection. J. Exp. Med. 206:1253–7230. Douek DC, Roederer M, Koup RA. 2009. Emerging concepts in the immunopathogenesis of AIDS. Annu.
Rev. Med. 60:471–8431. Wei X, Decker JM, Wang S, et al. 2003. Antibody neutralization and escape by HIV-1. Nature 422:307–1232. Richman DD, Wrin T, Little SJ, et al. 2003. Rapid evolution of the neutralizing antibody response to
HIV type 1 infection. Proc. Natl. Acad. Sci. USA 100:4144–4933. Euler Z, van Gils MJ, Bunnik EM, et al. 2010. Cross-reactive neutralizing humoral immunity does not
protect from HIV type 1 disease progression. J. Infect. Dis. 201:1045–53
108 Picker · Hansen · Lifson
Ann
u. R
ev. M
ed. 2
012.
63:9
5-11
1. D
ownl
oade
d fr
om w
ww
.ann
ualr
evie
ws.
org
by U
nive
rsity
of
Cal
ifor
nia
- Sa
n D
iego
on
11/1
1/13
. For
per
sona
l use
onl
y.

ME63CH06-Picker ARI 12 December 2011 11:55
34. Reynolds MR, Rakasz E, Skinner PJ, et al. 2005. CD8+ T-lymphocyte response to major immunodom-inant epitopes after vaginal exposure to simian immunodeficiency virus: too late and too little. J. Virol.79:9228–35
35. Watkins DI, Burton DR, Kallas EG, et al. 2008. Nonhuman primate models and the failure of the MerckHIV-1 vaccine in humans. Nat. Med. 14:617–21
36. Wilson NA, Reed J, Napoe GS, et al. 2006. Vaccine-induced cellular immune responses reduce plasmaviral concentrations after repeated low-dose challenge with pathogenic simian immunodeficiency virusSIVmac239. J. Virol. 80:5875–85
37. Rosati M, von Gegerfelt A, Roth P, et al. 2005. DNA vaccines expressing different forms of simianimmunodeficiency virus antigens decrease viremia upon SIVmac251 challenge. J. Virol. 79:8480–92
38. Mattapallil JJ, Douek DC, Buckler-White A, et al. 2006. Vaccination preserves CD4 memory T cellsduring acute simian immunodeficiency virus challenge. J. Exp. Med. 203:1533–41
39. Liu J, O’Brien KL, Lynch DM, et al. 2009. Immune control of an SIV challenge by a T-cell-based vaccinein rhesus monkeys. Nature 457:87–91
40. Horton H, Vogel TU, Carter DK, et al. 2002. Immunization of rhesus macaques with a DNAprime/modified vaccinia virus Ankara boost regimen induces broad simian immunodeficiency virus (SIV)-specific T-cell responses and reduces initial viral replication but does not prevent disease progressionfollowing challenge with pathogenic SIVmac239. J. Virol. 76:7187–202
41. Hel Z, Nacsa J, Tryniszewska E, et al. 2002. Containment of simian immunodeficiency virus infection invaccinated macaques: correlation with the magnitude of virus-specific pre- and postchallenge CD4+ andCD8+ T cell responses. J. Immunol. 169:4778–87
42. Hel Z, Tsai WP, Tryniszewska E, et al. 2006. Improved vaccine protection from simian AIDS by theaddition of nonstructural simian immunodeficiency virus genes. J. Immunol. 176:85–96
43. Engram JC, Dunham RM, Makedonas G, et al. 2009. Vaccine-induced, simian immunodeficiency virus-specific CD8+ T cells reduce virus replication but do not protect from simian immunodeficiency virusdisease progression. J. Immunol. 183:706–17
44. Casimiro DR, Wang F, Schleif WA, et al. 2005. Attenuation of simian immunodeficiency virus SIVmac239infection by prophylactic immunization with DNA and recombinant adenoviral vaccine vectors expressingGag. J. Virol. 79:15547–55
45. Barouch DH, Liu J, Lynch DM, et al. 2009. Protective efficacy of a single immunization of a chimericadenovirus vector-based vaccine against simian immunodeficiency virus challenge in rhesus monkeys.J. Virol. 83:9584–90
46. Valentine LE, Watkins DI. 2008. Relevance of studying T cell responses in SIV-infected rhesus macaques.Trends Microbiol. 16:605–11
47. McDermott AB, O’Connor DH, Fuenger S, et al. 2005. Cytotoxic T-lymphocyte escape does not alwaysexplain the transient control of simian immunodeficiency virus SIVmac239 viremia in adenovirus-boostedand DNA-primed Mamu-A∗01-positive rhesus macaques. J. Virol. 79:15556–66
48. Letvin NL, Mascola JR, Sun Y, et al. 2006. Preserved CD4+ central memory T cells and survival invaccinated SIV-challenged monkeys. Science 312:1530–33
49. Hansen SG, Ford JC, Lewis MS, et al. 2011. Profound early control of highly pathogenic SIV by aneffector memory T-cell vaccine. Nature 473:523–27
50. Letvin NL, Rao SS, Montefiori DC, et al. 2011. Immune and genetic correlates of vaccine protectionagainst mucosal infection by SIV in monkeys. Sci. Transl. Med. 3:81ra36
51. Buchbinder SP, Mehrotra DV, Duerr A, et al. 2008. Efficacy assessment of a cell-mediated immunityHIV-1 vaccine (the Step Study): a double-blind, randomised, placebo-controlled, test-of-concept trial.Lancet 372:1881–93
52. McElrath MJ, De Rosa SC, Moodie Z, et al. 2008. HIV-1 vaccine-induced immunity in the test-of-conceptStep Study: a case-cohort analysis. Lancet 372:1894–905
53. Rolland M, Tovanabutra S, deCamp AC, et al. 2011. Genetic impact of vaccination on breakthroughHIV-1 sequences from the STEP trial. Nat. Med. 17:366–71
54. Gray G, Buchbinder S, Duerr A. 2010. Overview of STEP and Phambili trial results: two phase IIbtest-of-concept studies investigating the efficacy of MRK adenovirus type 5 gag/pol/nef subtype B HIVvaccine. Curr. Opin. HIV AIDS 5:357–61
www.annualreviews.org • New Paradigms for HIV/AIDS Vaccine Development 109
Ann
u. R
ev. M
ed. 2
012.
63:9
5-11
1. D
ownl
oade
d fr
om w
ww
.ann
ualr
evie
ws.
org
by U
nive
rsity
of
Cal
ifor
nia
- Sa
n D
iego
on
11/1
1/13
. For
per
sona
l use
onl
y.

ME63CH06-Picker ARI 12 December 2011 11:55
55. McElrath MJ. 2010. Immune responses to HIV vaccines and potential impact on control of acute HIV-1infection. J. Infect. Dis. 202(Suppl 2):S323–26
56. Flynn NM, Forthal DN, Harro CD, et al. 2005. Placebo-controlled phase 3 trial of a recombinantglycoprotein 120 vaccine to prevent HIV-1 infection. J. Infect. Dis. 191:654–65
57. Pitisuttithum P, Gilbert P, Gurwith M, et al. 2006. Randomized, double-blind, placebo-controlled efficacytrial of a bivalent recombinant glycoprotein 120 HIV-1 vaccine among injection drug users in Bangkok,Thailand. J. Infect. Dis. 194:1661–71
58. McElrath MJ, Haynes BF. 2010. Induction of immunity to human immunodeficiency virus type-1 byvaccination. Immunity 33:542–54
59. Simek MD, Rida W, Priddy FH, et al. 2009. Human immunodeficiency virus type 1 elite neutralizers:individuals with broad and potent neutralizing activity identified by using a high-throughput neutralizationassay together with an analytical selection algorithm. J. Virol. 83:7337–48
60. Parren PW, Marx PA, Hessell AJ, et al. 2001. Antibody protects macaques against vaginal challenge witha pathogenic R5 simian/human immunodeficiency virus at serum levels giving complete neutralization invitro. J. Virol. 75:8340–47
61. Hessell AJ, Poignard P, Hunter M, et al. 2009. Effective, low-titer antibody protection against low-doserepeated mucosal SHIV challenge in macaques. Nat. Med. 15:951–54
62. Rerks-Ngarm S, Pitisuttithum P, Nitayaphan S, et al. 2009. Vaccination with ALVAC and AIDSVAX toprevent HIV-1 infection in Thailand. N. Engl. J. Med. 361:2209–20
63. Florese RH, Demberg T, Xiao P, et al. 2009. Contribution of nonneutralizing vaccine-elicited antibodyactivities to improved protective efficacy in rhesus macaques immunized with Tat/Env compared withmultigenic vaccines. J. Immunol. 182:3718–27
64. Forthal DN, Landucci G, Phan TB, et al. 2005. Interactions between natural killer cells and antibody Fcresult in enhanced antibody neutralization of human immunodeficiency virus type 1. J. Virol. 79:2042–49
65. Asmal M, Sun Y, Lane S, et al. 2011. Antibody-dependent cell-mediated viral inhibition emerges aftersimian immunodeficiency virus SIVmac251 infection of rhesus monkeys coincident with gp140-bindingantibodies and is effective against neutralization-resistant viruses. J. Virol. 85:5465–75
66. Hladik F, Hope TJ. 2009. HIV infection of the genital mucosa in women. Curr. HIV/AIDS Rep. 6:20–2867. Tudor D, Derrien M, Diomede L, et al. 2009. HIV-1 gp41-specific monoclonal mucosal IgAs derived
from highly exposed but IgG-seronegative individuals block HIV-1 epithelial transcytosis and neutralizeCD4+ cell infection: an IgA gene and functional analysis. Mucosal Immunol. 2:412–26
68. Bomsel M, Tudor D, Drillet AS, et al. 2011. Immunization with HIV-1 gp41 subunit virosomes inducesmucosal antibodies protecting nonhuman primates against vaginal SHIV challenges. Immunity 34:269–80
69. Ruprecht RM. 1999. Live attenuated AIDS viruses as vaccines: promise or peril? Immunol. Rev. 170:135–4970. Johnson RP, Desrosiers RC. 1998. Protective immunity induced by live attenuated simian immunodefi-
ciency virus. Curr. Opin. Immunol. 10:436–4371. Koff WC, Johnson PR, Watkins DI, et al. 2006. HIV vaccine design: insights from live attenuated SIV
vaccines. Nat. Immunol. 7:19–2372. Jia B, Ng SK, DeGottardi MQ, et al. 2009. Immunization with single-cycle SIV significantly reduces viral
loads after an intravenous challenge with SIV(mac)239. PLoS Pathog. 5:e100027273. Genesca M, McChesney MB, Miller CJ. 2009. Antiviral CD8+ T cells in the genital tract control viral
replication and delay progression to AIDS after vaginal SIV challenge in rhesus macaques immunizedwith virulence attenuated SHIV 89.6. J. Intern. Med. 265:67–77
74. Schmitz JE, Johnson RP, McClure HM, et al. 2005. Effect of CD8+ lymphocyte depletion on virus con-tainment after simian immunodeficiency virus SIVmac251 challenge of live attenuated SIVmac239delta3-vaccinated rhesus macaques. J. Virol. 79:8131–41
75. Reynolds MR, Weiler AM, Weisgrau KL, et al. 2008. Macaques vaccinated with live-attenuated SIVcontrol replication of heterologous virus. J. Exp. Med. 205:2537–50
76. Gauduin MC, Yu Y, Barabasz A, et al. 2006. Induction of a virus-specific effector-memory CD4+ T cellresponse by attenuated SIV infection. J. Exp. Med. 203:2661–72
77. Reynolds MR, Weiler AM, Piaskowski SM, et al. 2010. Macaques vaccinated with simian immuno-deficiency virus SIVmac239Delta nef delay acquisition and control replication after repeated low-doseheterologous SIV challenge. J. Virol. 84:9190–99
110 Picker · Hansen · Lifson
Ann
u. R
ev. M
ed. 2
012.
63:9
5-11
1. D
ownl
oade
d fr
om w
ww
.ann
ualr
evie
ws.
org
by U
nive
rsity
of
Cal
ifor
nia
- Sa
n D
iego
on
11/1
1/13
. For
per
sona
l use
onl
y.

ME63CH06-Picker ARI 12 December 2011 11:55
78. Powers C, Fruh K. 2008. Rhesus CMV: an emerging animal model for human CMV. Med. Microbiol.Immunol. 197:109–15
79. Hansen SG, Vieville C, Whizin N, et al. 2009. Effector memory T cell responses are associated withprotection of rhesus monkeys from mucosal simian immunodeficiency virus challenge. Nat. Med. 15:293–99
80. Hansen SG, Powers CJ, Richards R, et al. 2010. Evasion of CD8+ T cells is critical for superinfection bycytomegalovirus. Science 328:102–6
81. Kim JH, Rerks-Ngarm S, Excler JL, et al. 2010. HIV vaccines: lessons learned and the way forward. Curr.Opin. HIV AIDS 5:428–34
82. Corey L, Nabel GJ, Dieffenbach C, et al. 2011. HIV-1 vaccines and adaptive trial designs. Sci. Transl.Med. 3:79ps13
83. Mohr CA, Arapovic J, Muhlbach H, et al. 2010. A spread-deficient cytomegalovirus for assessment offirst-target cells in vaccination. J. Virol. 84:7730–42
www.annualreviews.org • New Paradigms for HIV/AIDS Vaccine Development 111
Ann
u. R
ev. M
ed. 2
012.
63:9
5-11
1. D
ownl
oade
d fr
om w
ww
.ann
ualr
evie
ws.
org
by U
nive
rsity
of
Cal
ifor
nia
- Sa
n D
iego
on
11/1
1/13
. For
per
sona
l use
onl
y.

ME63-frontmatter ARI 20 December 2011 12:3
Annual Review ofMedicine
Volume 63, 2012Contents
Huntington’s Disease: Advocacy Driving ScienceNancy S. Wexler � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � 1
Direct-to-Consumer Genetic Testing: Perceptions, Problems,and Policy ResponsesTimothy Caulfield and Amy L. McGuire � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � �23
Human Genome Sequencing in Health and DiseaseClaudia Gonzaga-Jauregui, James R. Lupski, and Richard A. Gibbs � � � � � � � � � � � � � � � � � � � � �35
The Genetic Architecture of Schizophrenia: New Mutationsand Emerging ParadigmsLaura Rodriguez-Murillo, Joseph A. Gogos, and Maria Karayiorgou � � � � � � � � � � � � � � � � � � � �63
CCR5 Antagonism in HIV Infection: Current Conceptsand Future OpportunitiesTimothy J. Wilkin and Roy M. Gulick � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � �81
New Paradigms for HIV/AIDS Vaccine DevelopmentLouis J. Picker, Scott G. Hansen, and Jeffrey D. Lifson � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � �95
Emerging Concepts on the Role of Innate Immunity in the Preventionand Control of HIV InfectionMargaret E. Ackerman, Anne-Sophie Dugast, and Galit Alter � � � � � � � � � � � � � � � � � � � � � � � � � 113
Immunogenetics of Spontaneous Control of HIVMary Carrington and Bruce D. Walker � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � 131
Recent Progress in HIV-Associated NephropathyChristina M. Wyatt, Kristin Meliambro, and Paul E. Klotman � � � � � � � � � � � � � � � � � � � � � � � � 147
Screening for Prostate Cancer: Early Detection or Overdetection?Andrew J. Vickers, Monique J. Roobol, and Hans Lilja � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � 161
Targeting Metastatic MelanomaKeith T. Flaherty � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � 171
Nanoparticle Delivery of Cancer DrugsAndrew Z. Wang, Robert Langer, and Omid C. Farokhzad � � � � � � � � � � � � � � � � � � � � � � � � � � � � 185
v
Ann
u. R
ev. M
ed. 2
012.
63:9
5-11
1. D
ownl
oade
d fr
om w
ww
.ann
ualr
evie
ws.
org
by U
nive
rsity
of
Cal
ifor
nia
- Sa
n D
iego
on
11/1
1/13
. For
per
sona
l use
onl
y.

ME63-frontmatter ARI 20 December 2011 12:3
Circulating Tumor Cells and Circulating Tumor DNACatherine Alix-Panabieres, Heidi Schwarzenbach, and Klaus Pantel � � � � � � � � � � � � � � � � � � � 199
Translation of Near-Infrared Fluorescence Imaging Technologies:Emerging Clinical ApplicationsE.M. Sevick-Muraca � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � 217
Familial and Acquired Hemophagocytic LymphohistiocytosisG.E. Janka � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � 233
The Management of Gastrointestinal Stromal Tumors: A Modelfor Targeted and Multidisciplinary Therapy of MalignancyHeikki Joensuu and Ronald P. DeMatteo � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � 247
Carotid Stenting Versus EndarterectomyDavid Doig and Martin M. Brown � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � 259
Mitral Valve ProlapseT. Sloane Guy and Arthur C. Hill � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � 277
Telomeres, Atherosclerosis, and the Hemothelium: The Longer ViewAbraham Aviv and Daniel Levy � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � 293
Aquaporins in Clinical MedicineA.S. Verkman � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � 303
Role of Endoplasmic Reticulum Stress in Metabolic Diseaseand Other DisordersLale Ozcan and Ira Tabas � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � 317
Role of Fructose-Containing Sugars in the Epidemics of Obesityand Metabolic SyndromeKimber L. Stanhope � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � 329
Vaccines for Malaria: How Close Are We?Mahamadou A. Thera and Christopher V. Plowe � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � 345
Crisis in Hospital-Acquired, Healthcare-Associated InfectionsDavid P. Calfee � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � 359
Novel Therapies for Hepatitis C: Insights from the Structureof the VirusDahlene N. Fusco and Raymond T. Chung � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � 373
Multiple Sclerosis: New Insights in Pathogenesis and NovelTherapeuticsDaniel Ontaneda, Megan Hyland, and Jeffrey A. Cohen � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � 389
Traumatic Brain Injury and Its Neuropsychiatric Sequelaein War VeteransNina A. Sayer � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � 405
vi Contents
Ann
u. R
ev. M
ed. 2
012.
63:9
5-11
1. D
ownl
oade
d fr
om w
ww
.ann
ualr
evie
ws.
org
by U
nive
rsity
of
Cal
ifor
nia
- Sa
n D
iego
on
11/1
1/13
. For
per
sona
l use
onl
y.

ME63-frontmatter ARI 20 December 2011 12:3
Eosinophilic Esophagitis: Rapidly Advancing InsightsJ. Pablo Abonia and Marc E. Rothenberg � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � 421
Physician Workforce Projections in an Era of Health Care ReformDarrell G. Kirch, Mackenzie K. Henderson, and Michael J. Dill � � � � � � � � � � � � � � � � � � � � � � � 435
Reducing Medical Errors and Adverse EventsJulius Cuong Pham, Monica S. Aswani, Michael Rosen, HeeWon Lee,
Matthew Huddle, Kristina Weeks, and Peter J. Pronovost � � � � � � � � � � � � � � � � � � � � � � � � � � � � 447
Relationships Between Medicine and Industry: Approaches to theProblem of Conflicts of InterestRaymond Raad and Paul S. Appelbaum � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � 465
Wireless Technology in Disease Management and MedicineGari D. Clifford and David Clifton � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � 479
Geographic Variation in Health CareTom Rosenthal � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � 493
Deep Brain Stimulation for Intractable Psychiatric DisordersWayne K. Goodman and Ron L. Alterman � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � 511
Contemporary Management of Male InfertilityPeter J. Stahl, Doron S. Stember, and Marc Goldstein � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � 525
Indexes
Cumulative Index of Contributing Authors, Volumes 59–63 � � � � � � � � � � � � � � � � � � � � � � � � � � � 541
Cumulative Index of Chapter Titles, Volumes 59–63 � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � 545
Errata
An online log of corrections to Annual Review of Medicine articles may be found athttp://med.annualreviews.org/errata.shtml
Contents vii
Ann
u. R
ev. M
ed. 2
012.
63:9
5-11
1. D
ownl
oade
d fr
om w
ww
.ann
ualr
evie
ws.
org
by U
nive
rsity
of
Cal
ifor
nia
- Sa
n D
iego
on
11/1
1/13
. For
per
sona
l use
onl
y.





























