Embed Size (px)
Citation preview

This article was downloaded by [UZH Hauptbibliothek Zentralbibliothek Zuumlrich]On 10 September 2013 At 0551Publisher Taylor amp FrancisInforma Ltd Registered in England and Wales Registered Number 1072954 Registered office Mortimer House37-41 Mortimer Street London W1T 3JH UK
Journal of Vertebrate PaleontologyPublication details including instructions for authors and subscription informationhttpwwwtandfonlinecomloiujvp20
New material of Menadon besairiei (CynodontiaTraversodontidae) from the Triassic of MadagascarChristian F Kammerer a b John J Flynn c Lovasoa Ranivoharimanana d amp Andreacute R Wyss ea Committee on Evolutionary Biology University of Chicago Chicago Illinois 60637 USAE-mailb Department of Geology The Field Museum Chicago Illinois 60605 USAc Division of Paleontology American Museum of Natural History New York New York 10024USA E-maild Deacutepartement de Paleacuteontologie et dAnthropologie Biologique Universiteacute dAntananarivoAntananarivo Madagascare Department of Geological Sciences University of California Santa Barbara California93106 USA E-mailPublished online 02 Aug 2010
To cite this article Christian F Kammerer John J Flynn Lovasoa Ranivoharimanana amp Andreacute R Wyss (2008) New materialof Menadon besairiei (Cynodontia Traversodontidae) from the Triassic of Madagascar Journal of Vertebrate Paleontology282 445-462 DOI 1016710272-4634(2008)28[445NMOMBC]20CO2
To link to this article httpdxdoiorg1016710272-4634(2008)28[445NMOMBC]20CO2
PLEASE SCROLL DOWN FOR ARTICLE
Taylor amp Francis makes every effort to ensure the accuracy of all the information (the ldquoContentrdquo) containedin the publications on our platform However Taylor amp Francis our agents and our licensors make norepresentations or warranties whatsoever as to the accuracy completeness or suitability for any purpose of theContent Any opinions and views expressed in this publication are the opinions and views of the authors andare not the views of or endorsed by Taylor amp Francis The accuracy of the Content should not be relied upon andshould be independently verified with primary sources of information Taylor and Francis shall not be liable forany losses actions claims proceedings demands costs expenses damages and other liabilities whatsoeveror howsoever caused arising directly or indirectly in connection with in relation to or arising out of the use ofthe Content
This article may be used for research teaching and private study purposes Any substantial or systematicreproduction redistribution reselling loan sub-licensing systematic supply or distribution in anyform to anyone is expressly forbidden Terms amp Conditions of access and use can be found at httpwwwtandfonlinecompageterms-and-conditions
ARTICLE
NEW MATERIAL OF MENADON BESAIRIEI (CYNODONTIA TRAVERSODONTIDAE) FROMTHE TRIASSIC OF MADAGASCAR
CHRISTIAN F KAMMERER12 JOHN J FLYNN3 LOVASOA RANIVOHARIMANANA4 and ANDREacute R WYSS5
1Committee on Evolutionary Biology University of Chicago Chicago Illinois 60637 USA cfkammeruchicagoedu2Department of Geology The Field Museum Chicago Illinois 60605 USA
3Division of Paleontology American Museum of Natural History New York New York 10024 USA jflynnamnhorg4Deacutepartement de Paleacuteontologie et drsquoAnthropologie Biologique Universiteacute drsquoAntananarivo Antananarivo Madagascar
5Department of Geological Sciences University of California Santa Barbara California 93106 USA wyssgeolucsbedu
ABSTRACTmdashNew specimens of the traversodontid cynodont Menadon besairiei from the MiddleLate Triassic lsquoIsalo IIrsquobeds of southwestern Madagascar are described The new specimens include a skull and the first postcranial materialdescribed for this taxon Although the referred skull differs from the holotype of M besairiei in some regards thesedifferences are interpreted as intraspecific variation and preservational artifacts The new material allows clarification ofsome aspects of the cranial morphology of Menadon showing that contrary to previous reports this taxon had awell-developed internarial bar and four upper incisors The postcranium of Menadon displays a mixture of advancedfeatures observed in the South American traversodontid Exaeretodon such as tall caudal neural spines and primitivefeatures observed in earlier traversodontids like Luangwa such as heterogeneous rib morphology A phylogeneticanalysis incorporating information from the new specimens supports the placement of Menadon as the sister taxon to theclade Exaeretodon + Scalenodontoides
INTRODUCTION
The Traversodontidae represent a diverse group of non-mammalian cynodonts (henceforth simply referred to as lsquocyn-odontsrsquo) Traversodontids are well represented from Triassic de-posits across the world with specimens known from NorthAmerica (eg Arctotraversodon Boreogomphodon Sues andOlsen 1990 Sues et al 1992) South America (eg Ex-aeretodon Massetognathus Traversodon Gomphodontosuchusvon Huene 1928 1936 Cabrera 1943 Bonaparte 1962 Romer1967 1972 Abdala and Giannini 2000 Abdala et al 2002 Ab-dala and Ribeiro 2003) mainland Africa (eg Scalenodon Sca-lenodontoides Crompton 1955 1972 Crompton and Ellen-berger 1957 Gow and Hancox 1993) Madagascar (MenadonDadadon Flynn et al 1999 2000) India (Exaeretodon Chatter-jee 1982) and Europe (eg Nanogomphodon Hopson andSues 2006) Traversodontids have a characteristic lsquogomphodontrsquodentition in which the postcanines are transversely expandedwhich is generally interpreted as an adaptation for herbivory(Reisz and Sues 2000) Similar expanded postcanines are alsopresent in diademodontid and trirachodontid cynodonts andthese three groups have been hypothesized to form a monophy-letic Gomphodontia (Abdala et al 2006) However controversyconcerning the relationships of traversodontids to other cyn-odonts and even traversodontid monophyly persists (Rowe1993 Hopson and Kitching 2001)
In recent years a wealth of new terrestrial vertebrate fossilsincluding traversodontids has been collected from Triassic de-posits in Madagascar (Flynn et al 1999) Two species of traver-sodontids (Menadon besairiei and Dadadon isaloi) have beendescribed from these deposits on the basis of cranial remains
from the basal Isalo II beds (Flynn et al 2000) Here we reporton new specimens of Menadon besairiei representing cranial andpostcranial material that expand our knowledge of this taxonrsquosmorphology Although partially disarticulated the first of thesenew specimens almost certainly pertains to a single individualand sheds light on several aspects of the skeletal anatomy ofMiddleLate Triassic traversodontids The second new specimenis an isolated pelvis These new specimens bear on the questionof traversodontid phylogeny and provide a fuller appreciation ofthe complexity of character transformations in cynodont phylog-eny
Institutional AbbreviationsmdashBP Bernard Price InstituteUniversity of the Witwatersrand Johannesburg South AfricaFMNH Field Museum of Natural History Chicago IllinoisUSA GPIT Institut und Museum fuumlr Geologie und Palaumlon-tologie der Universitaumlt Tuumlbingen Tuumlbingen Germany MCZMuseum of Comparative Zoology Harvard University Cam-bridge Massachusetts USA MNHN Museacuteum nationaldrsquoHistoire naturelle Paris France NMQR National MuseumBloemfontein South Africa TSK Collection of Prof ThomasKemp Oxford University Museum of Natural History OxfordUK UA Universiteacute drsquoAntananarivo Antananarivo Madagas-car UMZC University Museum of Zoology Cambridge UK
MATERIALS AND METHODS
FMNH PR 2444 was collected at site M-14 (basal Isalo II bedsof Bereketa Madagascar) in three separate plaster jackets giventhe field number 9-4-98-440 In all six cranial elements 20 ver-tebrae nine ribs and three limb bones were recovered fromthese slabs The partial cranium is poorly preserved deformedand practically edentulous Additional elements assigned thefield number 9-2-98-422 and consisting of a nearly complete rightpelvis an isolated vertebra and several rib fragments wereCorresponding author
Journal of Vertebrate Paleontology 28(2)445ndash462 June 2008copy 2008 by the Society of Vertebrate Paleontology
445
Dow
nloa
ded
by [
UZ
H H
aupt
bibl
ioth
ek
Zen
tral
bibl
ioth
ek Z
uumlric
h] a
t 05
51 1
0 Se
ptem
ber
2013
found in close proximity to 9-4-98-440 and are considered topertain to the same individual This decision is based on the factthat 9-2-98-422 was recovered on the same erosional surface asand very near 9-4-98-440 is in the same size range as the mate-rial from 9-4-98-440 and there is no overlap in elements betweenthe two field-numbered specimens The isolated vertebra from9-2-98-422 in particular is identical in size preservational stateand morphology to those present in 9-4-98-440 An isolated frag-mentary right pelvis (field number 8-31-98-387) herein referredto Menadon was collected at a different locality (M-18) than theprevious specimen Specimens were measured using Fowlerregdigital calipers to an accuracy of 001 mm Phylogenetic analyseswere run using a modified version of the data matrix of Abdalaet al (2006) in the parsimony program PAUP v 40b10 (Swof-ford 2003)
Comparisons with other cynodonts were made on the basis offirst hand examination of the following specimens
Menadon besairiei UA 10601 FMNH PR 2104 Dadadon isa-loi UA 10605 10606 Luangwa drysdalli BP13731 TSK 121Scalenodon angustifrons UMZC T907 T914 T918 Traversodonstahleckeri GPITRE7170 Scalenodontoides macrodontesMNHN 1957-23 NMQR 3053 Exaeretodon frenguellii MCZ4469 4470 4472 4483 4486 4493 4505 Gomphodontosuchusbrasiliensis GPIT unnumbered Massetognathus pascuali MCZ3782 3784 3786 3789 3790 3791 3792 3795 3796 3800 38013801-188 3803 3806 3807 3810 3999 4003 4009 4012 4018-164 4018-167 4047 4048 4049 4050 4138 4207 4208 42134214 4217 4218 4219 4221 4222 4230 4246 4253 4258 42654420 Diademodon tetragonus UMZC T492 Cricodon metabo-lus UMZC T905
Other comparisons were based on specimen photographs anddescriptions from the literature (Brink 1963 Bonaparte 19631966 Jenkins 1970 1971 Kemp 1980 Hopson 1985 Flynn etal 2000 Abdala et al 2002 Abdala and Teixeira 2004)
SYSTEMATIC PALEONTOLOGY
SYNAPSIDA Osborn 1903THERAPSIDA Broom 1905CYNODONTIA Owen 1861
EUCYNODONTIA Kemp 1982GOMPHODONTIA Seeley 1894 (sensu Hopson and
Kitching 2001)TRAVERSODONTIDAE von Huene 1936
DefinitionmdashHere we consider Traversodontidae to representthe most inclusive clade containing Traversodon stahleckeri vonHuene 1936132 but not Trirachodon kannemeyeri Seeley 189548 or Diademodon tetragonus Seeley 18941030
MENADON BESAIRIEI Flynn Parrish RakotosamimananaRanivoharimanana Simpson and Wyss 2000
HolotypemdashUA 10601 skull and mandible with eroded leftside
Referred SpecimensmdashFMNH PR 2444 a partial skull andpostcranium field number 8-31-98-387 an isolated right pelvisFMNH PR 2104 an isolated mandible
Horizon and LocalitymdashBasal lsquoIsalo IIrsquo of Besairie equated tothe Makay Formation by Razafimbelo (1987) drainage of theMalio River Morondava Basin Madagascar
AgemdashLate Middle (Ladinian) or early Late (Carnian) Triassic(see Flynn et al 1999 2000)
Revised DiagnosismdashTraversodontid with the followingunique combination of features four large upper incisors firstand second upper incisors and all lower incisors procumbentthird and fourth upper incisors caniniform with fourth upperincisor strongly recurved serrated upper canines small equal in
dorsoventral length to incisors distinctly canted forwards (auta-pomorphy) canine alveolus proportionally narrower than incisoralveoli with a much greater anteroposterior than labiolinguallength (autapomorphy) no diastema between upper incisors andcanine low number of postcanines (8 uppers and 6ndash7 lowers)upper and lower postcanines quadrangular roughly trapezoidalin outline descending process of jugal posterior to the last upperpostcanine axial spine with concave dorsal profile and elongateposterior process overhanging shortened neural spine of subse-quent cervical vertebra expanded ribs present caudal neuralspines very tall anterior edge of the iliac blade sloping upwardsat a sim45deg angle acetabular buttresses of the three pelvic boneslargely confluent posterior process of the iliac blade short anddirected away from the dorsal edge of the ischium ischium andpubis extremely constricted resulting in a large obturator fora-men
CommentsmdashMenadon can be distinguished from all other tra-versodontids by the combination of four large upper incisors andincisor procumbency The presence of four upper incisors isprimitive for traversodontids (Hopson and Kitching 2001) En-largement of the incisors is an advanced feature present in suchtaxa as Exaeretodon Scalenodontoides and lsquoScalenodonrsquo hirsch-soni (Abdala and Ribeiro 2003 Abdala et al 2006) Incisorprocumbency is otherwise only known in Exaeretodon and Sca-lenodontoides among traversodontids but these taxa have onlythree upper incisors (Bonaparte 1962 Hopson 1984) Addition-ally Menadon has a descending process of the jugal that contactsthe pterygoids and excludes the maxilla from the margin of thesubtemporal fenestra which is present in most gomphodonts butnot in Exaeretodon or Scalenodontoides (Abdala et al 2006)
Because the postcranium is known in few traversodontid taxathe combination of pelvic characters listed in the Diagnosis canonly be said to diagnose Menadon in comparison with LuangwaPascualgnathus Massetognathus and Exaeretodon The postcra-nium of Menadon resembles that of Luangwa in the shape of theiliac blade the large obturator foramen and the heterogeneousrib morphology but resembles Exaeretodon in the tall caudalneural spines confluent acetabular buttresses and short poste-rior process of the iliac blade projecting away from the ischium
DESCRIPTION
Skull
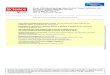
The cranium of FMNH PR 2444 is poorly preserved and nosutures are visible Six portions of the skull are represented theentire rostrum and left orbitalzygomatic region (Figs 1ndash3) partsof both zygomatic arches (Fig 4AndashC) a fragment of the tempo-ral region including the sagittal crest (Fig 4D) and the occiputand basicranium (Figs 4E 5) The following description is basedon the left side of the skull The right side of the rostrum hasbeen sheared moving some cranial landmarks (eg naris al-veoli) out of their original positions The frontal region on bothsides of the skull has also been sheared anterolaterally towardsthe left such that a shelf of bone made up of the nasals leftprefrontal and left lacrimal overhangs the snout (see Figs 12A)
The rostrum is tall and robust as in Gomphodontosuchus(Hopson 1985) and juveniles of Exaeretodon (Abdala et al2002) The external naris is large and teardrop-shaped narrow-ing towards the rear (Fig 1) A well-developed internarial bar ispresent The maxilla curves inward posterior to the root of thecanine so that the postcanine tooth row is medially inset locatedon an axis posterior to the paracanine fossa (Fig 2B) A singlelateral maxillary foramen is present The orbit is large and sub-circular Prominent interorbital depressions are present on theskull roof (Fig 2A)
Other than a fragment of the left fourth incisor the skull lacks
JOURNAL OF VERTEBRATE PALEONTOLOGY VOL 28 NO 2 2008446
Dow
nloa
ded
by [
UZ
H H
aupt
bibl
ioth
ek
Zen
tral
bibl
ioth
ek Z
uumlric
h] a
t 05
51 1
0 Se
ptem
ber
2013
teeth entirely Alveoli are preserved for all tooth loci except theinferred anterior postcanines (five postcanine alveoli are pre-served on the left side) The paracanine fossa is ovoid locatedmedial to the canine alveolus and much larger than all toothalveoli (Fig 3) Paired incisive foramina are located medial tothe paracanine fossae and extend from the anterior edge of thefourth incisor to the posterior edge of the canine The postca-nines are few in number (minimally five in this individual butprobably eight see Discussion) compared to many traversodon-tids (Romer 1967 Crompton 1972 Abdala et al 2002) andtheir alveoli are large and roughly trapezoidal in outline Thisconfiguration is similar to that observed in Gomphodontosuchus(Hopson 1985)
The left zygomatic arch is composed almost entirely of thejugal and as in most traversodontids it is relatively deep (Flynnet al 2000 Hopson and Kitching 2001) The posterior portionwhere the jugal would have articulated with the squamosal isbroken The isolated right jugal is more complete and its pos-terior articulation with the squamosal is intact (Fig 4A B) Thedorsal edge of the jugal is broken and is missing the posteriorextension over the squamosal The squamosals are very delicatebut well preserved bones each consisting of a cranial and zygo-matic ramus (Fig 4AndashC) The cranial ramus is an extremely thinwedge of bone directed dorsally that would have flanked therear of the braincase in the complete skull The zygomatic ramusis subdivided into a process extending anteriorly and one extend-ing ventrally with a pair of facets between them where the squa-mosal receives the posterior end of the jugal (Fig 4A C) Theanterior process is thin much longer than the ventral processand tapers forward The ventral process is short blunt and wideand forms the lateral portion of the external auditory meatus
The occiput and foramen magnum are tall and narrow (Fig4E) The occipital condyles are separated by a prominent groove
and are also taller than they are wide (but see Discussion) Abroken fragment of the proatlas is preserved out-of-place abovethe foramen magnum (Fig 4E) The occipital fragment includesthe posterior part of the basicranium The basicranium (Fig 5) issimilar to that described for Massetognathus (Rougier et al1992) and Exaeretodon (Abdala et al 2002) However FMNHPR 2444 differs from these in the possession of a basipterygoidforamen Such a foramen has been described by Parrington(1946) in the early cynodont Thrinaxodon but is not known intraversodontids This foramen is not evident in the holotype ofMenadon and this character may be variable in cynodonts
Axial Skeleton
A total of 21 vertebrae are preserved in FMNH PR 2444 Theangle of zygapophyseal articulation varies along the length of thevertebral column This angle (from the sagittal plane) is sim45deg forthe axisC3 (cervical 3) articulation sim30deg for the posterior cervi-cals lt15deg for the dorsals and sim20-30deg for the caudals Other thanthe atlas all vertebral centra are amphicoelous
Atlas-Axis ComplexmdashThree intercentra are articulated withthe cervical series IC2 (fused to the atlas-axis complex) IC3(between C2 and C3) and IC4 (between C3 and C4) IC2 makesup the lower part of the anterior face of the atlas-axis complexand extends back to the ventral anterior edge of the axial cen-trum (Fig 6) The free intercentra (IC3 and IC4) are singleirregularly shaped ossicles situated at the ventral margins of theintercervical articulations (Fig 6) The intercentra of FMNH PR2444 differ markedly from those described for Thrinaxodon(Jenkins 1971) but in general shape and position resemble thosefigured by Brink (1955) for Diademodon The intercentra ofThrinaxodon are transversely wide and anteroposteriorly short(Jenkins 1971fig 2) whereas the intercentra of Menadon be-
FIGURE 1 Stereopair and interpretive drawing of main cranial fragment of FMNH PR 2444 in left lateral view Scale bar equals 5 cm Abbre-viationsmdashJU jugal LA lacrimal MA maxilla mfo lateral maxillary foramen NA nasal nar naris orb orbit PF prefrontal PM premaxilla POpostorbital SM septomaxilla Sutures are not visible in this specimen and have been reconstructed based on the holotype of Menadon besairiei
KAMMERER ET ALmdashCYNODONT FROM MADAGASCAR 447
Dow
nloa
ded
by [
UZ
H H
aupt
bibl
ioth
ek
Zen
tral
bibl
ioth
ek Z
uumlric
h] a
t 05
51 1
0 Se
ptem
ber
2013
FIGURE 2 Stereopairs of main cranial fragment of FMNH PR 2444 in A dorsal view and B palatal view Scale bars equal 5 cm
JOURNAL OF VERTEBRATE PALEONTOLOGY VOL 28 NO 2 2008448
Dow
nloa
ded
by [
UZ
H H
aupt
bibl
ioth
ek
Zen
tral
bibl
ioth
ek Z
uumlric
h] a
t 05
51 1
0 Se
ptem
ber
2013
sairiei have equal transverse and anteroposterior dimensions Attheir maximum depth in Thrinaxodon the intercentra barely ex-tend beyond the ventral margins of the cervical pleurocentra InFMNH PR 2444 IC4 extends for half of its total depth below theventral margins of the cervical pleurocentra In IC3 this condi-tion is even more extreme extending below the ventral marginsof the cervical pleurocentra for 80 of its total depth The shapeand position of the intercentra give a strongly arched appearanceto the ventral profile of elements in the cervical series The ven-tral surfaces of the intercentra bear a posteriorly directed process
on each side In IC3 a weak ridge originates between the pairedprocesses and extends posteriorly indicating muscle attachmenton the intercentra The condition in IC4 cannot be determinedbecause of incomplete preservation Although only three inter-centra are known ventral facets in the centra of the isolatedcervicals indicate that others were present
As in other cynodonts the atlas-axis complex of FMNH PR2444 consists of a pair of separate atlantal neural arches articu-lating with the occipital condyles and the atlas centrum Only theleft atlas arch is preserved in this specimen (Fig 7) It is similar
FIGURE 3 Stereopair and interpretive drawing of anterior palatal region of FMNH PR 2444 Scale bar equals 5 cm AbbreviationsmdashC canineI incisor i f incisive foramina PC postcanine pc f paracanine fossa Postcanine numbering based on comparison with the holotype UA 10601
FIGURE 4 Cranial fragments of FMNH PR 2444 A stereopair of right zygomatic arch in lateral view Scale bar equals 5 cm B stereopair of rightzygomatic arch in medial view Scale bar equals 5 cm C stereopair of left squamosal in lateral view Scale bar equals 5 cm D sagittal crest in rightlateral view Scale bar equals 1 cm E occiput in occipital view Scale bar equals 1 cm
KAMMERER ET ALmdashCYNODONT FROM MADAGASCAR 449
Dow
nloa
ded
by [
UZ
H H
aupt
bibl
ioth
ek
Zen
tral
bibl
ioth
ek Z
uumlric
h] a
t 05
51 1
0 Se
ptem
ber
2013
to the atlantal neural arches described for other cynodonts suchas Massetognathus (Kemp 1969 Jenkins 1971) There is a me-dial facet for articulation with the centrum of the atlas (theldquoodontoid facetrdquo of Kemp 1969) and an anterior facet for ar-ticulation with the occipital condyle (Fig 7B) A pair of pro-cesses dominates the atlas arch a dorsal lamina (a roughly rect-angular plate of bone arching above the facets) and a largeposterolaterally directed transverse process An oval tubercle atthe anterior end of the transverse process represents the facet forthe proatlas
The centrum of the atlas is fused to the axis forming a singlelarge vertebral unit (Fig 6) A weak odontoid process is presentat the dorsal limit of the anterior surface of the atlas The ante-rior aspect of the centrum is dominated by a roughly lsquoTrsquo-shapedstructure composed of the two facets for articulation with theatlas arches and the facet for articulation with the atlas intercen-trum The axis bears no prezygapophyses and as such does notarticulate with the atlas arch as in Early Triassic cynodonts suchas Galesaurus (Parrington 1934) There appears to be a trendtowards loss of the axial prezygapophyses in later traversodon-
tids which are present but vestigial in Massetognathus (Jenkins1970 although in the specimens described the prezygapophysesare broken so their reduction is perhaps questionable) and ab-sent in Exaeretodon (Bonaparte 1963) The transverse processesof the axis are large and robust compared to those of the othercervicals The axial spine is saddle-shaped with a concave dorsalprofile unlike previous reconstructions of the traversodontidaxis which depict a straight or convex dorsal profile similar tothat observed in galesaurid cynodonts and early mammals (Jen-kins 1971) The morphology of the axial spine is similar to thatof the Early Triassic cynodont Cynognathus (Seeley 1895) InCynognathus however the neural spine of the third cervical(C3) is tall and canted posteriorly behind the posterior process ofthe axial spine whereas in FMNH PR 2444 the short C3 neuralspine fits below the posterior portion of the axial spine Theposterior process of the axial spine bears a pair of parallel elon-gate grooves on its underside attachment sites for the axial mus-culature
Post-Axial VertebraemdashThree post-axial cervicals are knownfor FMNH PR 2444 (Figs 6 8AndashD) The first of these C3 is
FIGURE 5 Stereopair of ventral view of the occipital fragment of FMNH PR 2444 showing the basicranium Anterior is up
FIGURE 6 Stereopair and interpretive drawing of atlas-axis complex and third cervical of FMNH PR 2444 in left lateral view Scale bar equals5 cm Abbreviationsmdasha n sp axial neural spine d dens IC intercentrum f at a facet for articulation with the atlas arch p parapophysis pozpostzygapophysis t p transverse process
JOURNAL OF VERTEBRATE PALEONTOLOGY VOL 28 NO 2 2008450
Dow
nloa
ded
by [
UZ
H H
aupt
bibl
ioth
ek
Zen
tral
bibl
ioth
ek Z
uumlric
h] a
t 05
51 1
0 Se
ptem
ber
2013
preserved in articulation with the axis (Fig 6) The two isolatedcervicals bear successively taller and anterposteriorly wider neu-ral spines (Fig 8AndashD) Based on their staggered size and failureto articulate we suggest that they represent C5 and C7 (assumingthat this specimen possessed the generalized cynodont cervicalnumber of seven) The cervicals display robust transverse pro-cesses which are directed laterally and slightly anteriorly A par-apophysis lies below the anterior edge of the transverse processalong the antero-lateral edge of the centrum No cervical ribs arepreserved but their presence can be inferred from the par-apophyses The notochordal concavity is as in Massetognathussurrounded by a raised rim at each end of the centrum (Jenkins1970) No anapophyses are present The neural spines for allthree post-axial cervicals are tapering and slightly curved poste-riorly The neural spine in C3 is relatively short and wide(thereby fitting underneath the posterior overhang of the axialspine see Fig 6) while that of C5 is tall and narrow The spinefor C7 is taller and anteroposteriorly wider than that of C5 andthe dorsal edge is more rounded similar to the condition in thedorsal vertebrae
Eight post-cervical presacral vertebrae are known for thisspecimen We will refer to them here simply as dorsal vertebraegiven the incompleteness of the vertebral column in the speci-men and the uncertainty concerning the thoraciclumbar transi-tion in this taxon However it is probable that they representthoracic vertebrae (see below) In most cynodonts the lumbar
vertebrae are synostosed to specialized ribs with overlappingelements In earlier taxa (eg Thrinaxodon Cynognathus Lu-angwa Jenkins 1971 Kemp 1980) expanded costal plates arepresent on both thoracic and lumbar ribs In Massetognathus thelumbar ribs consist of a pair of prongs one directed anteriorlybearing a facet for articulation with the previous rib and theother directed posteriorly (Jenkins 1970) In Exaeretodon allpresacral ribs are identical of simple morphology and not syn-ostosed to the vertebrae (Bonaparte 1963) A single isolated rib(Fig 9B) in FMNH PR 2444 demonstrates that some of the ribsin Menadon besairiei were expanded However all preservedvertebrae in this specimen have facets for articulation with theribs rather than breakage points where they would have beenfused to ribs As such it is probable that all of the vertebraepreserved in this specimen represent thoracics although we can-not discount the possibility that as in Exaeretodon no presacralvertebrae were fused to ribs
The neural spines in all of the dorsal vertebrae are elongateposteriorly inclined and roughly rectangular in lateral view witha rounded laterally expanded platform dorsally (Fig 8EndashJ)Costal foveae are present at the anterior and posterior edges ofeach centrum Of the eight dorsal vertebrae there are two ar-ticulated sets and three isolated elements (two from 9-4-98-440and one from 9-2-98-422) The first isolated vertebra (Fig 8E F)has a taller neural spine and shorter centrum than the otherdorsal vertebrae It is most similar in proportions to the isolated
FIGURE 7 Stereopairs and interpretive drawings of left atlas arch of FMNH PR 2444 in A lateral view Anterior is left B medial view Anterioris right Scale bars equal 1 cm Abbreviationsmdashafa anterior facet for articulation with the occipital condyle dl dorsal lamina mfa medial facet forarticulation with the atlas centrum (odontoid facet) pfa tubercle forming the facet for articulation with the proatlas tp transverse process
KAMMERER ET ALmdashCYNODONT FROM MADAGASCAR 451
Dow
nloa
ded
by [
UZ
H H
aupt
bibl
ioth
ek
Zen
tral
bibl
ioth
ek Z
uumlric
h] a
t 05
51 1
0 Se
ptem
ber
2013
FIGURE 8 Vertebrae of FMNH PR 2444 Cervical vertebra 5 in A left lateral and B anterior views Cervical vertebra 7 in C left lateral andD anterior views Isolated dorsal vertebra from anterior region of the series in E left lateral and F anterior views G stereopair and interpretivedrawing of articulated set of two dorsal vertebrae from near the midpoint of the series in left lateral view Anterior is up H articulated set of threedorsal vertebrae from anterior region of the series in left and right lateral views I isolated vertebra from near the midpoint of the series (from fieldnumber 9-2-98-422) J isolated vertebra from posterior region of the series Sacral vertebra in K dorsal view (the distal region of another sacral ribwhich is identical in shape to that of the intact sacral rib albeit slightly larger is also shown in this figure) and L left lateral view Caudal vertebraeM articulated set of two vertebrae from the anterior region of the series showing the lsquowingrsquo-shaped caudal ribs in dorsal view Anterior is left Nisolated caudal vertebra showing tall neural spine in left lateral view O isolated caudal vertebra showing tall neural spine in posterior view Particulated set of four vertebrae from near the end of the caudal series in dorsal view Anterior is left Scale bars equal 1 cm Abbreviationsmdasha cof anterior costal fovea n sp neural spine p co f posterior costal fovea poz postzygapophysis prz prezygapophysis
JOURNAL OF VERTEBRATE PALEONTOLOGY VOL 28 NO 2 2008452
Dow
nloa
ded
by [
UZ
H H
aupt
bibl
ioth
ek
Zen
tral
bibl
ioth
ek Z
uumlric
h] a
t 05
51 1
0 Se
ptem
ber
2013
cervicals and was probably located near the front of the dorsalseries The first articulated set consists of three vertebrae (Fig8H) The middle vertebra in this set is broken from the anteriorbase of the neural spine to the ventral midpoint and most of theright side of the anterior half of the centrum is missing This setof vertebrae probably also came from the anterior region of theseries as their neural spines are longer and narrower than allvertebrae other than the one described above The second ar-ticulated set (Fig 8G) consists of two well-preserved vertebraethat based on comparisons with Massetognathus and Ex-aeretodon (Jenkins 1970 Bonaparte 1963) probably came fromnear the midpoint of the dorsal series The second isolated ver-tebra (Fig 8I) from 9-2-98-422 is similar to the members of thesecond set The third isolated vertebra (Fig 8J) appears to befrom the posterior end of the series based on the relatively shortneural spine and reduced posterior costal fovea
One sacral vertebra (Fig 8K L) is known consisting of awell-preserved centrum neural spine and the right sacral rib(the left is broken off) The centrum is shorter (both anteropos-teriorly and dorsoventrally) than those of the dorsal vertebraeand the neural spine is shorter and more rounded in profileAlso the neural spine is directed vertically unlike those of thedorsal vertebrae which are canted posteriorly The zygapophy-ses are poorly developed The sacral rib is fused to a singlesynapophysis and projects anterolaterally The rib narrows to-wards the middle and expands distally terminating in a widerectangular facet where it would meet the ilium (Fig 8K)
Seven caudal vertebrae are known The caudal vertebrae aredistinctive in having very tall narrow neural spines Some EarlyTriassic cynodonts possess blunt neural spines of moderateheight on the proximal caudals (see for example Thrinaxodonin Jenkins 1971 and Diademodon in Brink 1955) Massetogna-thus represents an extreme condition wherein the caudal neuralspines are greatly reduced (Jenkins 1970) Very tall neuralspines are seen in Exaeretodon (Bonaparte 1963) and thosespines are unusual in that they do not diminish in height until thelast caudals The caudal vertebrae of FMNH PR 2444 consist ofa pair of articulated vertebrae most likely from near the sacrum(Fig 8M) an isolated caudal lacking postzygapophyses and ribsbut with the neural spine (Fig 8N O) and a quartet of smaller
articulated elements (Fig 8P) The anterior caudals have nar-rower neural spines and shorter zygapophyses than the subse-quent vertebrae in the series The caudal ribs attached to thispair have a crescentic distal region that curves anteriorly pro-ducing a lsquowingrsquo-shape Only a portion of the right rib of theisolated caudal is preserved but the lsquowingrsquo-shaped morphologyof the near-sacral caudal ribs does not appear to have been pre-sent In the posteriormost four vertebrae caudal ribs are pre-served on the left side of the first and the right side of the secondand third members On the first of these vertebrae the rib isshort curved posteriorly (but not lsquowingrsquo-shaped) and taperingRibs of the second and third vertebrae are less curved resem-bling posteriorly directed rods The zygapophyses of both theisolated caudal vertebra and the quartet of small caudals areanteroposteriorly elongate becoming increasingly so posteriorly
Thoracic RibsmdashNine thoracic ribs are known from 9-4-98-440(four left four right and an indeterminate splint of bone Fig9A indeterminate bone not illustrated) These elements re-semble the flattened ribs lacking costal plates of Exaeretodon(Bonaparte 1963) and the anterior thoracic ribs of Massetogna-thus and Luangwa (Jenkins 1970 Kemp 1980) The tuberculumand capitulum are separated by a shallow depression The tuber-culum has a shorter base than the capitulum but their proximalfaces are subequal in size An additional isolated left rib (Fig9B) from 9-2-98-422 is expanded indicating that unlike Ex-aeretodon Menadon rib morphology was heterogeneous alongthe length of the dorsal series This rib closely resembles theposterior thoracic ribs described for Luangwa (Kemp 1980fig16) Pascualgnathus (Bonaparte 1966fig 7) and the trirach-odontid Cricodon (Crompton 1955 Fig 4) This rib is con-stricted at the base of the head then expands distally with an-terior and posterior costal plates As in Luangwa the anteriorcostal plate is shallow and evenly sloping proximally and distallywhile the posterior costal plate is falcate with a concave edgedistally The distal expansion of this rib occurs at an angle of sim30degto the long axis of the rib head Despite their overall similaritythe expanded rib in FMNH PR 2444 does exhibit some differ-ences from the posterior thoracic rib of Luangwa The expandedrib of Menadon has a longer shaft between the rib head and thecostal plates and a shorter capitulum lacks a ridge on the pos-
FIGURE 9 A anterior thoracic ribs of FMNH PR 2444 in anterior view (indeterminate rib not figured) B expanded posterior thoracic rib ofFMNH PR 2444 in anterior view Scale bars equal 1 cm
KAMMERER ET ALmdashCYNODONT FROM MADAGASCAR 453
Dow
nloa
ded
by [
UZ
H H
aupt
bibl
ioth
ek
Zen
tral
bibl
ioth
ek Z
uumlric
h] a
t 05
51 1
0 Se
ptem
ber
2013
terior costal plate and has an anteroposteriorly shorter anteriorcostal plate Additionally the posterior margin of the costal plateis less curved than that of Luangwa These differences couldindicate that the expanded rib of Menadon is from a more pos-terior position in the series than the rib from Luangwa as re-duced curvature and increasing confluence of the tuberculumand capitulum are characteristic of lumbar ribs in traversodon-tids (Jenkins 1970 Kemp 1980) However the length of theshaft is at odds with this interpretation and in Cricodon andPascualgnathus the more posterior ribs are not more similar tothe element from FMNH PR 2444 These differences probablyrepresent legitimate morphological distinctions between the ribsof earlier gomphodonts and Menadon The head of the expandedrib is well defined and unbroken indicating that it was not syn-ostosed to a vertebra
Appendicular Skeleton
The appendicular skeleton is represented by both scapulocora-coids and a fragment of a clavicle from field number 9-4-98-440and a partial right pelvis from field number 9-2-98-422 An ad-ditional partial right pelvis is represented from field number8-31-98-387 The scapulocoracoids (Figs 10 11) are very similarto those described for other traversodontids as well as cynog-nathids and diademodontids (Jenkins 1970 1971) The scapularinfraspinous fossa has out-turned anterior and posterior edgesThe anterior edge of the scapula is rounded and curves laterallyforming the scapular spine At the lower end of the spine theacromion forms a prominent process (broken in the left scapula)Below this process the scapula is constricted and then expandsventrally to form the dorsal margin of the articular glenoid Theglenoid fossa is in general morphology very similar to that ofother Triassic cynodonts (Jenkins 1970 1971 Kemp 1980) It isposteroventrally oriented with roughly equal contributions fromthe scapular and coracoid glenoid facets The coracoid glenoidfacet is roughly triangular while the scapular glenoid facet issemiovoid as in other cynodonts In anterior or posterior viewthe fossa appears smoothly crescentic The procoracoid is aslightly curved trapezoidal plate of bone It does not appear tocontribute to the glenoid fossa The procoracoid foramen is lo-cated midway up the height of the procoracoid at a point directlybelow the base of the acromion process The coracoid is semi-crescentic curving posteriorly from a broad base The coracoidin Menadon is longer and more falcate than those described forExaeretodon (Bonaparte 1963) and Luangwa (Kemp 1980 al-though the distal tip of both coracoids is broken in the specimenof Luangwa [TSK 121] described by Kemp and may have beenlonger than reconstructed) but agrees closely with those of Mas-setognathus and cynognathidsdiademodontids (Jenkins 19701971) The distal coracoid tubercle for attachment to the tricepsis strongly curved upwards similar to the condition in Masseto-gnathus (Jenkins 1970) and Diademodon (Brink 1955)
A small fragment of bone resembling a curved cylinder almostcertainly represents part of a clavicle It has no distinguishingfeatures other than a groove associated with a ridge on one sideHowever the degree of curvature observed precludes this ele-ment from representing a long bone and the circular cross-section excludes it from being a rib
The pelvis of FMNH PR 2444 (Fig 12) resembles those ofother traversodontids in general morphology The ilium bears anarrow anteriorly directed iliac blade curving outward distallyThe anterior edge of the iliac blade slopes upward at a roughly45deg angle similar to Luangwa (Kemp 1980) but unlike Ex-aeretodon in which the anterior edge of the iliac blade curvesbackwards (Bonaparte 1963) or Pascualgnathus and Masseto-gnathus in which it is rounded (Bonaparte 1966 Jenkins 1970)The supraacetabular buttress at the base of the ilium is located inan anterodorsal position relative to the acetabulum as in other
traversodontids but unlike earlier cynodonts in which it is lo-cated dorsally (Jenkins 1970) The acetabular buttresses of thethree pelvic bones are largely confluent forming a fairly uniformlsquoliprsquo around the acetabulum as in Exaeretodon (Bonaparte1963) On the contrary in Luangwa the supraacetabular ischialand pubic buttresses are highly distinct spaced 6ndash8 mm apart andwithout confluence of their acetabular facets The pelvis ofFMNH PR 2444 does have a small depression between the su-praacetabular and ischial buttresses (also present in Ex-aeretodon Bonaparte 1963) but this is not as developed as inLuangwa The posterior extent of the iliac blade is broken inFMNH PR 2444 but this region is preserved in another isolatedright pelvis field number 8-31-98-387 (Fig 13) probably pertain-ing to Menadon besairiei The posterior process of the iliac bladein this specimen is relatively short and directed away from thedorsal edge of the ischium as in Exaeretodon (Bonaparte 1963)but unlike Massetognathus and Luangwa in which the posteriorprocess is long and extends roughly horizontally over the dorsaledge of the ischium (Jenkins 1970 Kemp 1980) and evencurves downward distally in Luangwa The posterior process ofthe iliac blade in Pascualgnathus is directed away from the is-chium (Bonaparte 1966) as in Exaeretodon and Menadon but isrelatively long as in Massetognathus and Luangwa
The proximal portion of the ischium is broad and dominatedby the ischial buttress The ischium constricts posterior to theproximal portion and expands distally into a spatulate platewhich is thickened at its posterior margin This thickening con-tinues along the dorsolateral surface of the ischium forming aridge that slopes ventrally until reaching the edge of the ischialbuttress The downward slope of this ridge produces a prominentgroove at the dorsal edge of the lateral surface of the ischiumthought to represent the origin of the ischio-trochantericusmuscle (Kemp 1980) The constriction of the ischium in Mena-don is like that of Luangwa albeit even more extreme in itsventral concavity but unlike that of Exaeretodon in which themedial and distal ends of the ischium are broadly united Theischium in general is a proportionally shorter element in Ex-aeretodon than in other traversodontids including Menadon(Bonaparte 1963) The anteroventral portion of the ischium thatconnects to the pubis is not preserved in either of the referredMenadon pelves
The pubis is the most incompletely preserved pelvic elementThe acetabular contribution of the pubis is smaller than that ofthe ischium or ilium Distal to the acetabulum the pubis is con-stricted indicating that the connection between the acetabularregion and the pubic plate was considerably narrower than inExaeretodon (Bonaparte 1963) but close akin to the pubis ofLuangwa (Kemp 1980) The pubic plate itself is not preservedThe ventral outline of the ischium and what is known of thepubis indicate that the obturator foramen was relatively larger inMenadon (Fig 14) than in Exaeretodon (Bonaparte 1963)
DISCUSSION
Comparison with the Holotype of Menadon besairiei
FMNH PR 2444 differs in several significant ways from theholotype of Menadon besairiei as previously described most no-tably in the possession of an internarial bar (initially consideredabsent in M besairiei) and four upper incisors (reported as threein M besairiei but with the caveat that the homologies of thefourth and fifth anterior tooth loci were problematic Flynn et al2000) However upon further study we interpret the lack of acontinuous internarial bar in the holotype as breakage ratherthan the natural condition of the skull (a prominent medial pro-tuberance is present on the ventral rim of the nares in UA10601) Regarding the incisor count it should be noted that theholotype of Menadon besairiei displays an unusual set of anterior
JOURNAL OF VERTEBRATE PALEONTOLOGY VOL 28 NO 2 2008454
Dow
nloa
ded
by [
UZ
H H
aupt
bibl
ioth
ek
Zen
tral
bibl
ioth
ek Z
uumlric
h] a
t 05
51 1
0 Se
ptem
ber
2013
FIGURE 10 Right scapulocoracoids of FMNH PR 2444 A stereopair of right scapulocoracoid in medial view B stereopair and interpretivedrawing of right scapulocoracoid in lateral view Scale bars equal 5 cm Abbreviationsmdashac acromion process C coracoid c gl coracoid contributionto the glenoid fossa c tr tubercle for coracoid attachment of triceps PC procoracoid pc f procoracoid foramen S scapula s gl scapularcontribution to the glenoid fossa
KAMMERER ET ALmdashCYNODONT FROM MADAGASCAR 455
Dow
nloa
ded
by [
UZ
H H
aupt
bibl
ioth
ek
Zen
tral
bibl
ioth
ek Z
uumlric
h] a
t 05
51 1
0 Se
ptem
ber
2013
teeth for a traversodontid As mentioned above Flynn et al(2000) noted that the homologies of these teeth were difficult todetermine Of the five anterior teeth preserved in the holotypethe anteriormost three are ldquoundoubtedly incisorsrdquo although I3 isldquounusually large and is caniniformrdquo (Flynn et al 2000423) Therobust recurved fourth tooth in the series was considered torepresent the canine with the subsequent tooth interpreted as anolder canine in the process of replacement Although FMNH PR2444 is edentulous the alveoli on the left side of the skull are wellpreserved and indicate an anterior tooth set identical to the ho-lotype The alveolar morphology indicates that the fifth toothwas canted anteriorly as in the holotype This suggests that themysterious fifth tooth is not the result of a transient period in thedental replacement history of M besairiei as suggested by Flynnet al (2000) but rather is the norm This leaves two possibilities1) M besairiei persistently had two canines This condition isunknown in cynodonts 2) The canine-like fourth tooth actuallyrepresents an incisor and the fifth tooth is the true canine Wefavor the latter explanation In FMNH PR 2444 the first fouralveoli are nearly identical in size and shape However the fifthalveolus while similar in anteroposterior length to its predeces-sors is strongly compressed labiolingually with an anteroposte-rior lengthlabiolingual width ratio of 044 (compared to 080 inthe fourth alveolus and 084ndash85 for I2ndash3) We interpret thismarked difference in alveolar ratio as indicating the separationbetween incisors and canine The canine-like I4 would then rep-resent the endpoint in a series of large increasingly caniniformincisors Supporting this interpretation recurved incisors are alsoobserved in juvenile Exaeretodon (eg MCP 3843 PV see Ab-dala et al 2002)
There are several legitimate differences between FMNH PR2444 and the holotype of Menadon besairiei but we take a con-servative taxonomic stance and argue that these differences rep-resent intraspecific variation and preservational artifacts FMNHPR 2444 represents a larger animal than UA 10601 The snoutlength (measured as the distance between the anterior edge ofthe orbit and a vertical plane through the anterior tip of thepremaxilla) of FMNH PR 2444 is 911 mm while that of UA10601 is 801 mm As such proportional differences between thetwo skulls may be attributable to ontogeny Variations in FMNHPR 2444 compared to the holotype that probably represent on-togentic differences include wider longer and deeper paraca-nine fossae a much deeper ventral zygomatic flange (althoughthe lower border of the flange is slightly abraded in the holotypeof Menadon besairiei there is no evidence that a major flangewas broken off) a proportionally shorter and more circular out-line to the posterior border of the maxilla below the zygomaticprocess and a more deeply emarginated posterior squamosal(ie a more prominent laterally projecting flange)
FMNH PR 2444 has a proportionally shorter more robustsnout and larger orbits than the holotype While these charactersare certainly variable in cynodonts it is unusual that the largerspecimen is shorter-snouted and larger-eyed contra the usualpattern The proportional differences between the two skulls areminor however (orbit diametersnout length is 042 for FMNHPR 2444 and 041 for the holotype) and are partially attributableto deformation (shear in FMNH PR 2444 and breakage of thepostorbital bar in the holotype) The following differences canalso probably be attributed to preservational artifact only fivepostcanine alveoli are preserved in FMNH PR 2444 while theholotype has eight postcanines The anteriormost three in theholotype however are tiny and peg-like As the area of themaxilla between the canine alveolus and the first preserved post-canine alveolus is poorly preserved in FMNH PR 2444 the ab-sence of these postcanines may be artifactual However it is alsopossible that the lower postcanine count of FMNH PR 2444compared to the holotype is an ontogenetic effect as postcaninecount is known to decrease with increasing skull size in Ex-
FIGURE 11 Left scapulocoracoids of FMNH PR 2444 A left scapu-locoracoid in medial view B left scapulocoracoid in lateral view Scalebars equal 5 cm
JOURNAL OF VERTEBRATE PALEONTOLOGY VOL 28 NO 2 2008456
Dow
nloa
ded
by [
UZ
H H
aupt
bibl
ioth
ek
Zen
tral
bibl
ioth
ek Z
uumlric
h] a
t 05
51 1
0 Se
ptem
ber
2013
FIGURE 12 Right pelvis of FMNH PR 2444 A stereopair of right pelvis in medial view B stereopair and interpretive drawing of right pelvis inlateral view Scale bars equal 5 cm Abbreviationsmdashbpp base of posterior process of ilium ib iliac blade IL ilium IS ischium ob f obturatorforamen P pubis
KAMMERER ET ALmdashCYNODONT FROM MADAGASCAR 457
Dow
nloa
ded
by [
UZ
H H
aupt
bibl
ioth
ek
Zen
tral
bibl
ioth
ek Z
uumlric
h] a
t 05
51 1
0 Se
ptem
ber
2013
aeretodon (Abdala et al 2002) The orbit of FMNH PR 2444 isless upwardly directed than that of the holotype (being at anangle of sim65deg to a sagittal plane through the midline of the skullas opposed to sim45deg in the holotype) This difference in orbitalangle is probably attributable to postmortem deformation as theskull of FMNH PR 2444 is noticeably sheared to the front leftThe appearance of a wider squarer incisor arcade and broader
anterior palate in FMNH PR 2444 can also be attributed to thisshear Finally the occipital region of FMNH PR 2444 is alto-gether taller and narrower than that of the holotype For ex-ample the foramen magnum in FMNH PR 2444 approximates arounded isosceles triangle in shape while the foramen magnumof the holotype is more ovoid These differences can also beattributed to deformation Altogether the differences between
FIGURE 13 Stereopairs of field number 8-31-98-387 an isolated right pelvis in A medial and B lateral view Scale bars equal 5 cm
JOURNAL OF VERTEBRATE PALEONTOLOGY VOL 28 NO 2 2008458
Dow
nloa
ded
by [
UZ
H H
aupt
bibl
ioth
ek
Zen
tral
bibl
ioth
ek Z
uumlric
h] a
t 05
51 1
0 Se
ptem
ber
2013
FMNH PR 2444 and the holotype fall within the range of intra-specific variation recorded for other gomphodont cynodontssuch as the traversodontids Massetognathus pascuali (Abdalaand Giannini 2000 Kammerer pers obs) and Exaeretodon(Abdala et al 2002) and the diademodontid Diademodon tetra-gonus (Grine et al 1978 Bradu and Grine 1979)
Phylogenetic Analysis
Utilizing the new information afforded by FMNH PR 2444and a reexamination of the holotype (UA 10601) Menadon be-sairiei was recoded in the Gondwanan gomphodont data matrixof Abdala et al (2006) Abdala et alrsquos (2006) data matrix en-compassed 18 taxa and 43 characters For the present analysis anadditional character ldquoincisor procumbencyrdquo (states 0-absent1-present) was added to Abdala et alrsquos (2006) data set (see
Appendix for coding of this character) Several traversodontidtaxa (eg lsquoScalenodonrsquo hirschsoni) have relatively large incisorsbut only in Menadon Scalenodontoides and Exaeretodon are theincisors procumbent Three of Abdala et alrsquos (2006) originalcodings for Menadon besairiei characters 0 (ldquonumber of upperincisorsrdquo) 5 (ldquoposition of paracanine fossa in relation to theupper caninerdquo) and 21 (ldquointernarial barrdquo) were changed to in-corporate data from the new specimen (see Appendix) Addi-tionally further examination of the holotype (UA 10601) ofMenadon besairiei allowed us to code characters 16ndash18 20 38and 40 (morphology of the lower postcanines all marked as ldquordquoin Abdala et alrsquos analysis) Parsimony searches were performedin PAUP v40b10 using a branch-and-bound search Addition-ally a bootstrap analysis (also using branch-and-bound) was per-formed using 1000 replicates
A total of 34 most parsimonious trees of length 103 werefound with the scores CI0573 RI0725 RC0415 andHI0427 The strict consensus tree (Fig 15A) did not recovera polytomy between Menadon Exaeretodon and Scalenodont-oides as in Abdala and Ribeiro (2003) or a (Menadon + Ex-aeretodon) sister-group relationship as in Abdala et al (2006)Rather Menadon was recovered as the sister taxon of the (Ex-aeretodon + Scalenodontoides) clade supporting Flynn et alrsquos(2000) original placement of this taxon There is strong bootstrapsupport (88) for Traversodontidae but within traversodontidsthe only strongly supported subclade is (Gomphodontosuchus +(Menadon + (Scalenodontoides + Exaeretodon))) with a boot-strap value of 80 Within this clade the (Menadon + (Scalen-odontoides + Exaeretodon)) grouping is supported by a boot-strap value of 83 and the (Scalenodontoides + Exaeretodon)grouping is supported by a bootstrap value of 94 (Fig 15B) Inaddition to the positioning of Menadon the strict consensus tree(Fig 15A) differs from the topology of Abdala et al (2006) inrecovering a sister-group relationship between Massetognathusand Santacruzodon rather than having Massetognathus andSantacruzodon as successive outgroups to the clade containingGomphodontosuchus Menadon Scalenodontoides and Ex-aeretodon The (Massetognathus + Santacruzodon) clade is diag-nosed by the absence of an anterolingual cusp in the upper post-canines denticulated incisor cutting margins and the presence ofan anterior cingulum in the upper postcanines
FIGURE 14 Reconstruction of the pelvis of Menadon besairiei basedon information from FMNH PR 2444 and field number 8-31-98-387Dotted area not preserved in either specimen reconstruction based onPascualgnathus (Bonaparte 1966) and Exaeretodon (Bonaparte 1963)
FIGURE 15 Cladogram of traversodontid relationships based on revised data set of Abdala et al (2006) showing A strict consensus tree and B50 majority rule tree Numbers at nodes of majority rule tree represent bootstrap values
KAMMERER ET ALmdashCYNODONT FROM MADAGASCAR 459
Dow
nloa
ded
by [
UZ
H H
aupt
bibl
ioth
ek
Zen
tral
bibl
ioth
ek Z
uumlric
h] a
t 05
51 1
0 Se
ptem
ber
2013
Relationships of Menadon besairiei
Menadon besairiei is the sister taxon to (Exaeretodon + Sca-lenodontoides) the latter clade diagnosed by the possession ofthree upper incisors a posteromedial position of the paracaninefossae relative to the canines the absence of an internarial barlarge canines a projected zygomatic process of the jugal and amaxilla that contributes to the margin of the subtemporal fenes-tra (the last three character states represent reversals) The clade(Menadon + (Exaeretodon + Scalenodontoides)) is diagnosed bythe possession of large procumbent incisors The presence offour incisors a primitive retention distinguishes Menadon be-sairiei from Exaeretodon Scalenodontoides and an unnamed tra-versodontid (ldquoTraversodontid Type 1rdquo) from the Santa MariaFormation of Brazil (Abdala et al 2001 Abdala and Ribeiro2002) Gomphodontosuchus is recovered as the sister taxon to(Menadon (Exaeretodon + Scalenodontoides)) as has beenfound in previous studies (Flynn et al 2000 Abdala and Ribeiro2003) The taxon name Gomphodontosuchinae Watson andRomer 1956 (ex Gomphodontosuchidae Watson and Romer1956 first given subfamilial rank in Lehman 1961) is availablefor this clade As circumscribed here Gomphodontosuchinaeconsists of the most inclusive clade containing Gomphodontosu-chus brasiliensis von Huene 1928264 but not Massetognathuspascuali Romer 19672
Menadon besairiei displays a mixture of primitive and derivedcharacters within traversodontids The internarial bar and fourincisors are primitive features whereas the enlargement and pro-cumbency of the incisors are derived features shared with Ex-aeretodon and Scalenodontoides The postcranium shows severalsimilarities to that of Exaeretodon (apparent exclusion of theprocoracoid from the glenoid some features of the pelvis tallcaudal neural spines) but in general displays a rather conserva-tive morphology observed throughout Gomphodontia The mor-phology of the scapulocoracoids is closer akin to diademodontidsor trirachodontids than Exaeretodon and the expanded thoracicrib of Menadon has only slight differences from those of trirach-odontids or early traversodontids like Luangwa The atlas-axiscomplex of Menadon is similar to that of the Early Triassic cyn-odont Cynognathus in possessing a concave axial spine with along posterior process Although most gomphodonts have beenreconstructed with a convex axial spine as in early mammals thisspine is broken in most of the best described gomphodont post-crania (eg Diademodon Brink 1955) and the lsquosaddle-shapedrsquoaxial morphology may be more widely distributed among cyn-odonts than previously realized The new specimens of Menadondescribed in this paper demonstrate that a generalized gompho-dont postcranial morphology was retained deep within the tra-versodontid tree and that the derived morphologies of either ofthe well-known taxa Massetognathus or Exaeretodon should notbe taken as representative of traversodontids in general
ACKNOWLEDGMENTS
This work would not have been possible without the continu-ing support and encouragement of our collaborative programwith the Deacutepartement de Paleacuteontologie et drsquoAnthropologie Bi-ologique Universiteacute drsquoAntananarivo Madagascar under the di-rection of Armand Rasoamiaramanana (Department Chairman)and Gisegravele Ravololonarivo Randria (Head of the PaleontologyLaboratories) We are extremely grateful to the National Geo-graphic Society for their long-term support of the fieldwork as-sociated with this project Additional financial or logistical sup-port for fieldwork was provided by The Field Museum the Uni-versity of California-Santa Barbara and the WWF (WorldwideFund for Nature Madagascar) We acknowledge the Ministegraverede LrsquoEneacutergie et des Mines for authorizing field efforts and for
continuing to facilitate our joint US-Madagascar explorationresearch and education program Steve Goodman and AsminaGandhi generously assisted us in innumerable ways includinghospitality nonpareil We especially thank R Andriantompoha-vana L Razafimanantsoa A Toto Volahy JA Rabarison AGandie and P Vavisaro for assistance in the field during theexpedition in which these new specimens were discovered BillSimpson and Gina Wesley-Hunt for their patient excavation ofthis important material Allison Smith Ray Vodden Lisa Berg-wall and Deborah Wagner for excellent preparation of a chal-lenging specimen Jennifer Botha and Elize Butler (NationalMuseum Bloemfontein) Michael Raath and Fernando Abdala(Bernard Price Institute) Thomas Kemp (Oxford UniversityMuseum of Natural History) Ray Symonds (University Museumof Zoology Cambridge) Bernard Battail (Museacuteum nationaldrsquoHistoire naturelle Paris) Michael Maisch (Universitaumlt Tuumlbin-gen) and Charles Schaff and Farish Jenkins (Museum of Com-parative Zoology Harvard) for access to specimens and AgustiacutenMartinelli Fernando Abdala and James Hopson for helpful re-views of an earlier version of the manuscript
LITERATURE CITED
Abdala F M C Barbarena and J Dornelles 2002 A new species of thetraversodontid cynodont Exaeretodon from the Santa Maria Forma-tion (MiddleLate Triassic) of Southern Brazil Journal of Verte-brate Paleontology 22313ndash325
Abdala F and N P Giannini 2000 Gomphodont cynodonts of theChantildeares Formation the analysis of an ontogenetic sequence Jour-nal of Vertebrate Paleontology 20501ndash506
Abdala F J Neveling and J Welman 2006 A new trirachodontidcynodont from the lower levels of the Burgersdorp Formation(Lower Triassic) of the Beaufort Group South Africa and the cla-distic relationships of Gondwanan gomphodonts Zoological Jour-nal of the Linnean Society 147383ndash413
Abdala F A M Ribeiro and C L Schultz 2001 A rich cynodont faunaof Santa Cruz do Sul Santa Maria Formation (Middle-Late Trias-sic) southern Brazil Neues Jahrbuch fuumlr Geologie und Palaumlontolo-gie Monatshefte 2001669ndash687
Abdala F and A M Ribeiro 2002 Nuevos cinodontes traversodoacuten-tidos (Synapsida-Eucynodontia) de la Formacioacuten Santa Maria(Triaacutesico Medio-Superior) Rio Grande do Sul Brasil RevistaEspantildeola de Paleontologiacutea 17237ndash243
Abdala F and A M Ribeiro 2003 A new traversodontid cynodontfrom the Santa Maria Formation (Ladinian-Carnian) of southernBrazil with a phylogenetic analysis of Gondwanan traversodontidsZoological Journal of the Linnaean Society 139529ndash545
Abdala F and A M S Teixeira 2004 A traversodontid cynodont ofAfrican affinity in the South American Triassic Palaeontologia Af-ricana 4011ndash22
Bonaparte J F 1962 Descripcioacuten del craacuteneo y mandiacutebula de Ex-aeretodon frenguellii Cabrera y su comparacioacuten con Diademodon-tidae Tritylodontidae y los cinodontes sudamericanos Publica-ciones del Museo Municipal de Ciencias Naturales y Tradicional deMar del Plata 1135ndash202
Bonaparte J F 1963 Descripcioacuten del esqueleto postcraneano de Ex-aeretodon (CynodontiandashTraversodontidae) Acta Geoloacutegica Lil-loana 45ndash52
Bonaparte J F 1966 Una nueva ldquofaunardquo Triaacutesica de Argentina (Ther-apsida Cynodontia Dicynodontia) Consideracions filogeneacuteticas ypaleobiogeograacuteficas Ameghiniana 8243ndash296
Bradu D and F E Grine 1979 Multivariate analysis of diademodon-tine crania from South Africa and Zambia South African Journal ofScience 79441ndash448
Brink A S 1955 A study of the skeleton of Diademodon Palaeonto-logia Africana 33-39
Brink A S 1963 Two new cynodonts from the Ntawere Formation inthe Luangwa Valley of Northern Rhodesia Palaeontologia Africana877ndash96
Broom R 1905 On the use of the term Anomodontia Records of theAlbany Museum 1266ndash269
Cabrera A 1943 El primer hallazgo de teraacutepsidos en la ArgentinaNotas del Museo de La Plata Paleontologiacutea 55317ndash331
JOURNAL OF VERTEBRATE PALEONTOLOGY VOL 28 NO 2 2008460
Dow
nloa
ded
by [
UZ
H H
aupt
bibl
ioth
ek
Zen
tral
bibl
ioth
ek Z
uumlric
h] a
t 05
51 1
0 Se
ptem
ber
2013
Chatterjee S 1982 A new cynodont reptile from the Triassic of IndiaJournal of Paleontology 56203ndash214
Crompton A W 1955 On some Triassic cynodonts from TanganyikaProceedings of the Zoological Society of London 125617ndash669
Crompton A W 1972 Postcanine occlusion in cynodonts and tritylo-donts Bulletin of the British Museum (Natural History) Geology2129ndash71
Crompton A W and F Ellenberger 1957 On a new cynodont from theMolteno Beds and the origin of the tritylodontids Annals of theSouth African Museum 441ndash13
Flynn J J J M Parrish B Rakotosaminimanana W F Simpson R LWhatley and A R Wyss 1999 A Triassic fauna from Madagascarincluding early dinosaurs Science 286763ndash765
Flynn J J J M Parrish B Rakotosamimanana L RanivoharimananaW F Simpson and A R Wyss 2000 New traversodontids (Syn-apsida Eucynodontia) from the Triassic of Madagascar Journal ofVertebrate Paleontology 20422ndash427
Gow C E and P J Hancox 1993 First complete skull of the LateTriassic Scalenodontoides (Reptilia Cynodontia) from southern Af-rica In Lucas S G and M Morales (eds) The nonmarine TriassicNew Mexico Museum of Natural History amp Science Bulletin 3161ndash168
Grine F E B D Hahn and C E Gow 1978 Aspects of relativegrowth and variability in Diademodon (Reptilia Therapsida) SouthAfrican Journal of Science 7450ndash58
Hopson J A 1984 Late Triassic traversodont cynodonts from NovaScotia and southern Africa Palaeontologia Africana 25181ndash201
Hopson J A 1985 Morphology and relationships of Gomphodontosu-chus brasiliensis von Huene (Synapsida Cynodontia Tritylodontoi-dea) from the Triassic of Brazil Neues Jahrbuch fuumlr Geologie undPalaumlontologie Monatshefte 1985285ndash299
Hopson J A and J W Kitching 2001 A probainognathian cynodontfrom South Africa and the phylogeny of nonmammalian cynodontsBulletin of the Museum of Comparative Zoology 1565ndash35
Hopson J A and H-D Sues 2006 A traversodont cynodont from theMiddle Triassic (Ladinian) of Baden-Wuumlrttemberg (Germany)Palaumlontologische Zeitschrift 80124ndash129
Huene F von 1928 Ein Cynodontier aus der Trias Brasiliens Zentral-blatt fuumlr Mineralogie Geologie und Palaumlontologie 1928251ndash270
Huene F von 1936 Die fossilen Reptilien des suumldamerikanischenGondwanalandes Ergebnisse der Sauriergrabungen in Suumldbrasilien192829 Lieferung 2 (Cynodontia)93ndash159 Franz F Heine Tuumlbin-gen
Jenkins F A 1970 The Chantildeares (Argentina) Triassic reptile faunaVII The postcranial skeleton of the traversodontid Massetognathuspascuali (Therapsida Cynodontia) Breviora 3521ndash28
Jenkins F A 1971 The postcranial skeleton of African cynodonts prob-lems in the early evolution of the mammalian postcranial skeletonBulletin of the Peabody Museum of Natural History 361ndash216
Kemp T S 1969 The atlas-axis complex of mammal-like reptiles Jour-nal of Zoology 159223ndash248
Kemp T S 1980 Aspects of the structure and functional anatomy of theMiddle Triassic cynodont Luangwa Journal of Zoology 191193ndash239
Kemp T S 1982 Mammal-Like Reptiles and the Origin of MammalsAcademic Press New York
Lehman J-P 1961 Cynodontia pp 140ndash191 in Piveteau J (ed) Traiteacutede Paleacuteontologie Masson et Cie Paris
Osborn 1903 On the primary division of the Reptilia into two sub-classes Synapsida and Diapsida Science 17275ndash276
Owen R 1861 Palaeontology or a systematic summary of extinct ani-mals and their geological relations 2nd Ed Edinburgh
Parrington F R 1934 On the cynodont genus Galesaurus with a noteon the functional significance of the changes in the evolution of thetheriodont skull Annals and Magazine of Natural History 10(13)38ndash67
Parrington F R 1946 On the cranial anatomy of cynodonts Proceed-ings of the Zoological Society of London 116181ndash197
Razafimbelo E 1987 Le basin de Morondava (Madagascar) Synthegravesegeacuteologique et structurale PhD dissertation University Louis Pas-teur Strasbourg France 189 pp
Reisz R R and H-D Sues 2000 Herbivory in Late Paleozoic andTriassic terrestrial vertebrates pp 9ndash41 in Sues H-D (ed) Evo-lution of Herbivory in Terrestrial Vertebrates Cambridge Univer-sity Press New York
Romer A S 1967 The Chantildeares (Argentina) Triassic reptile fauna IIITwo new gomphodonts Massetognathus pascuali and M terugiiBreviora 2641ndash25
Romer A S 1972 The Chantildeares (Argentina) Triassic reptile faunaXVII The Chantildeares gomphodonts Breviora 3961ndash9
Rougier G W J R Wible and J A Hopson 1992 Reconstruction ofthe cranial vessels in the Early Cretaceous mammal Vincelestes neu-quenianus implications for the evolution of the mammalian cranialvascular system Journal of Vertebrate Paleontology 12188ndash216
Rowe T 1988 Definition diagnosis and origin of Mammalia Journal ofVertebrate Paleontology 8241ndash264
Rowe T 1993 Early mammal phylogenetic systematics pp 129ndash145 inSzalay F S M J Novacek and M C McKenna (eds) MammalPhylogeny Vol 1 Mesozoic Differentiation MultituberculatesMonotremes Early Therians and Marsupials Springer-Verlag NewYork
Seeley H G 1894 Researches on the structure organization and clas-sification of the fossil Reptilia Part IX Section 3 On DiademodonPhilosophical Transactions of the Royal Society of London 1851029ndash1041
Seeley H G 1895 Researches on the structure organization and clas-sification of the fossil Reptilia Part IX Section 4 On the Gompho-dontia Philosophical Transactions of the Royal Society of London1861ndash57
Seeley H G 1895 Researches on the structure organization and clas-sification of the fossil Reptilia Part IX Section 5 On the skeletonin new Cynodontia from the Karroo rocks Philosophical Transac-tions of the Royal Society of London 18659ndash148
Sues H-D J A Hopson and N H Shubin 1992 Affinities of Sca-lenodontoides plemmyridon Hopson 1984 (Synapsida Cynodontia)from the Upper Triassic of Nova Scotia Journal of Vertebrate Pa-leontology 12168ndash171
Sues H-D and P E Olsen 1990 Triassic vertebrates of Gondwananaspect from the Richmond basin of Virginia Science 2491020ndash1023
Swofford D L 2003 PAUP v 40b10 Sinauer Associates Inc Pub-lishers Sunderland Massachusetts
Watson D M S and A S Romer 1956 A classification of therapsidreptiles Bulletin of the Museum of Comparative Zoology 11435ndash89
Submitted September 20 2007 accepted November 3 2007
KAMMERER ET ALmdashCYNODONT FROM MADAGASCAR 461
Dow
nloa
ded
by [
UZ
H H
aupt
bibl
ioth
ek
Zen
tral
bibl
ioth
ek Z
uumlric
h] a
t 05
51 1
0 Se
ptem
ber
2013
APPENDIX Data matrix used in the phylogenetic analysesThis is a revised version of the matrix of Abdala et al (2006) with the following changes character 43 [Incisor procumbency (0) absent (1) present]added Menadon recoded for characters 0 (Number of upper incisors was 1 ldquothreerdquo changed to 0 ldquofourrdquo) 5 (Position of paracanine fossae in relationto the upper canine was changed to 1 ldquomedialrdquo) 16 (Overall morphology of lower postcanines in occlusal view was changed to 3 ldquoquadran-gularrdquo) 17 (Position of the transverse crest in lower postcanines was changed to 1 ldquoanteriorrdquo) 18 (Number of cusps in the transverse crest of thelower postcanines was changed to 0 ldquotwordquo) 20 (Anterior cingulum in the lower postcanines was changed to 2 ldquoabsentrdquo) 21 (Internarial barwas 1 ldquoabsentrdquo changed to 0 ldquopresentrdquo) 38 (Posterior cingulum in the lower postcanines was changed to 1 ldquoabsentrdquo) and 40 (Deep occlusalbasins in the postcanines was changed to 1 ldquopresentrdquo) Langbergia recoded for character 34 (Incisor cutting margins was 0 ldquoserratedrdquo changedto 1 ldquosmoothly ridgedrdquo) based on the description of the incisors in Abdala et al (2006) Gomphodontosuchus recoded for character 31 (Maxilla inthe margin of the subtemporal fenestra was 1 ldquoincludedrdquo changed to 0 ldquoexcludedrdquo) based on von Huene (1936) and Kammerer (pers obs of theholotype) contra Hopsonrsquos (1985) reconstruction lsquoScalenodonrsquo hirschsoni recoded for character 39 (Anterior cingulum in the upper postcanines was1 ldquoabsentrdquo changed to 0 ldquopresentrdquo) based on Abdala (pers comm 2006) Trirachodon recoded for character 30 (Ectopterygoid was changedto 0 ldquopresentrdquo) based on Abdala (pers comm 2006) and Traversodon recoded for character 15 (Number of cusps in the sectorial border of the upperpostcanines was 0 ldquomore than twordquo changed to 1 ldquotwordquo) based on Abdala (pers comm 2006) and Kammerer (pers obs)
1111111111 2222222222 3333333333 44440123456789 0123456789 0123456789 0123456789 0123
Cynognathus 00000000 000 000000000 00000000 000Diademodon 0000001000 0000100 000010A000 0000000100 0100Trirachodon 000000100B 1A002201 001A100010 0001001200 0110Langbergia 0000001000 10000201 0000101010 0001101200 0110Cricodon 0100001000 11000201 0010100 0001200 0110Andescynodon 0000002001 01013101 101120010 0112311 1100Massetognathus 00111121A2 1110003100 2011200011 1011212300 1100Exaeretodon 1110122112 010103101 2111110201 1111112311 1101Luangwa 0000001002 11010A3100 101010010 01002300 1100Scal angustifrons 000000100B 1111023100 10102101 00002301 1100Scal hirschsoni 11110202 1100113100 1010 1112300 1100Traversodon 0000002012 1101113100 201010 011230 1100Gomphodontosuchus 0011112112 0103101 2010 0112311 1100Pascualgnathus 1000001000 000231 1100001 0011230 1100Scalenodontoides 1112212 01003101 21111021 1212311 1101Menadon 0111112112 01012310 20121020 0102311 1101Dadadon 00002112 111112 013 11231 1100Santacruzodon 00002112 1110003101 201300 212300 1100
A(01) B(02)
JOURNAL OF VERTEBRATE PALEONTOLOGY VOL 28 NO 2 2008462
Dow
nloa
ded
by [
UZ
H H
aupt
bibl
ioth
ek
Zen
tral
bibl
ioth
ek Z
uumlric
h] a
t 05
51 1
0 Se
ptem
ber
2013

ARTICLE
NEW MATERIAL OF MENADON BESAIRIEI (CYNODONTIA TRAVERSODONTIDAE) FROMTHE TRIASSIC OF MADAGASCAR
CHRISTIAN F KAMMERER12 JOHN J FLYNN3 LOVASOA RANIVOHARIMANANA4 and ANDREacute R WYSS5
1Committee on Evolutionary Biology University of Chicago Chicago Illinois 60637 USA cfkammeruchicagoedu2Department of Geology The Field Museum Chicago Illinois 60605 USA
3Division of Paleontology American Museum of Natural History New York New York 10024 USA jflynnamnhorg4Deacutepartement de Paleacuteontologie et drsquoAnthropologie Biologique Universiteacute drsquoAntananarivo Antananarivo Madagascar
5Department of Geological Sciences University of California Santa Barbara California 93106 USA wyssgeolucsbedu
ABSTRACTmdashNew specimens of the traversodontid cynodont Menadon besairiei from the MiddleLate Triassic lsquoIsalo IIrsquobeds of southwestern Madagascar are described The new specimens include a skull and the first postcranial materialdescribed for this taxon Although the referred skull differs from the holotype of M besairiei in some regards thesedifferences are interpreted as intraspecific variation and preservational artifacts The new material allows clarification ofsome aspects of the cranial morphology of Menadon showing that contrary to previous reports this taxon had awell-developed internarial bar and four upper incisors The postcranium of Menadon displays a mixture of advancedfeatures observed in the South American traversodontid Exaeretodon such as tall caudal neural spines and primitivefeatures observed in earlier traversodontids like Luangwa such as heterogeneous rib morphology A phylogeneticanalysis incorporating information from the new specimens supports the placement of Menadon as the sister taxon to theclade Exaeretodon + Scalenodontoides
INTRODUCTION
The Traversodontidae represent a diverse group of non-mammalian cynodonts (henceforth simply referred to as lsquocyn-odontsrsquo) Traversodontids are well represented from Triassic de-posits across the world with specimens known from NorthAmerica (eg Arctotraversodon Boreogomphodon Sues andOlsen 1990 Sues et al 1992) South America (eg Ex-aeretodon Massetognathus Traversodon Gomphodontosuchusvon Huene 1928 1936 Cabrera 1943 Bonaparte 1962 Romer1967 1972 Abdala and Giannini 2000 Abdala et al 2002 Ab-dala and Ribeiro 2003) mainland Africa (eg Scalenodon Sca-lenodontoides Crompton 1955 1972 Crompton and Ellen-berger 1957 Gow and Hancox 1993) Madagascar (MenadonDadadon Flynn et al 1999 2000) India (Exaeretodon Chatter-jee 1982) and Europe (eg Nanogomphodon Hopson andSues 2006) Traversodontids have a characteristic lsquogomphodontrsquodentition in which the postcanines are transversely expandedwhich is generally interpreted as an adaptation for herbivory(Reisz and Sues 2000) Similar expanded postcanines are alsopresent in diademodontid and trirachodontid cynodonts andthese three groups have been hypothesized to form a monophy-letic Gomphodontia (Abdala et al 2006) However controversyconcerning the relationships of traversodontids to other cyn-odonts and even traversodontid monophyly persists (Rowe1993 Hopson and Kitching 2001)
In recent years a wealth of new terrestrial vertebrate fossilsincluding traversodontids has been collected from Triassic de-posits in Madagascar (Flynn et al 1999) Two species of traver-sodontids (Menadon besairiei and Dadadon isaloi) have beendescribed from these deposits on the basis of cranial remains
from the basal Isalo II beds (Flynn et al 2000) Here we reporton new specimens of Menadon besairiei representing cranial andpostcranial material that expand our knowledge of this taxonrsquosmorphology Although partially disarticulated the first of thesenew specimens almost certainly pertains to a single individualand sheds light on several aspects of the skeletal anatomy ofMiddleLate Triassic traversodontids The second new specimenis an isolated pelvis These new specimens bear on the questionof traversodontid phylogeny and provide a fuller appreciation ofthe complexity of character transformations in cynodont phylog-eny
Institutional AbbreviationsmdashBP Bernard Price InstituteUniversity of the Witwatersrand Johannesburg South AfricaFMNH Field Museum of Natural History Chicago IllinoisUSA GPIT Institut und Museum fuumlr Geologie und Palaumlon-tologie der Universitaumlt Tuumlbingen Tuumlbingen Germany MCZMuseum of Comparative Zoology Harvard University Cam-bridge Massachusetts USA MNHN Museacuteum nationaldrsquoHistoire naturelle Paris France NMQR National MuseumBloemfontein South Africa TSK Collection of Prof ThomasKemp Oxford University Museum of Natural History OxfordUK UA Universiteacute drsquoAntananarivo Antananarivo Madagas-car UMZC University Museum of Zoology Cambridge UK
MATERIALS AND METHODS
FMNH PR 2444 was collected at site M-14 (basal Isalo II bedsof Bereketa Madagascar) in three separate plaster jackets giventhe field number 9-4-98-440 In all six cranial elements 20 ver-tebrae nine ribs and three limb bones were recovered fromthese slabs The partial cranium is poorly preserved deformedand practically edentulous Additional elements assigned thefield number 9-2-98-422 and consisting of a nearly complete rightpelvis an isolated vertebra and several rib fragments wereCorresponding author
Journal of Vertebrate Paleontology 28(2)445ndash462 June 2008copy 2008 by the Society of Vertebrate Paleontology
445
Dow
nloa
ded
by [
UZ
H H
aupt
bibl
ioth
ek
Zen
tral
bibl
ioth
ek Z
uumlric
h] a
t 05
51 1
0 Se
ptem
ber
2013
found in close proximity to 9-4-98-440 and are considered topertain to the same individual This decision is based on the factthat 9-2-98-422 was recovered on the same erosional surface asand very near 9-4-98-440 is in the same size range as the mate-rial from 9-4-98-440 and there is no overlap in elements betweenthe two field-numbered specimens The isolated vertebra from9-2-98-422 in particular is identical in size preservational stateand morphology to those present in 9-4-98-440 An isolated frag-mentary right pelvis (field number 8-31-98-387) herein referredto Menadon was collected at a different locality (M-18) than theprevious specimen Specimens were measured using Fowlerregdigital calipers to an accuracy of 001 mm Phylogenetic analyseswere run using a modified version of the data matrix of Abdalaet al (2006) in the parsimony program PAUP v 40b10 (Swof-ford 2003)
Comparisons with other cynodonts were made on the basis offirst hand examination of the following specimens
Menadon besairiei UA 10601 FMNH PR 2104 Dadadon isa-loi UA 10605 10606 Luangwa drysdalli BP13731 TSK 121Scalenodon angustifrons UMZC T907 T914 T918 Traversodonstahleckeri GPITRE7170 Scalenodontoides macrodontesMNHN 1957-23 NMQR 3053 Exaeretodon frenguellii MCZ4469 4470 4472 4483 4486 4493 4505 Gomphodontosuchusbrasiliensis GPIT unnumbered Massetognathus pascuali MCZ3782 3784 3786 3789 3790 3791 3792 3795 3796 3800 38013801-188 3803 3806 3807 3810 3999 4003 4009 4012 4018-164 4018-167 4047 4048 4049 4050 4138 4207 4208 42134214 4217 4218 4219 4221 4222 4230 4246 4253 4258 42654420 Diademodon tetragonus UMZC T492 Cricodon metabo-lus UMZC T905
Other comparisons were based on specimen photographs anddescriptions from the literature (Brink 1963 Bonaparte 19631966 Jenkins 1970 1971 Kemp 1980 Hopson 1985 Flynn etal 2000 Abdala et al 2002 Abdala and Teixeira 2004)
SYSTEMATIC PALEONTOLOGY
SYNAPSIDA Osborn 1903THERAPSIDA Broom 1905CYNODONTIA Owen 1861
EUCYNODONTIA Kemp 1982GOMPHODONTIA Seeley 1894 (sensu Hopson and
Kitching 2001)TRAVERSODONTIDAE von Huene 1936
DefinitionmdashHere we consider Traversodontidae to representthe most inclusive clade containing Traversodon stahleckeri vonHuene 1936132 but not Trirachodon kannemeyeri Seeley 189548 or Diademodon tetragonus Seeley 18941030
MENADON BESAIRIEI Flynn Parrish RakotosamimananaRanivoharimanana Simpson and Wyss 2000
HolotypemdashUA 10601 skull and mandible with eroded leftside
Referred SpecimensmdashFMNH PR 2444 a partial skull andpostcranium field number 8-31-98-387 an isolated right pelvisFMNH PR 2104 an isolated mandible
Horizon and LocalitymdashBasal lsquoIsalo IIrsquo of Besairie equated tothe Makay Formation by Razafimbelo (1987) drainage of theMalio River Morondava Basin Madagascar
AgemdashLate Middle (Ladinian) or early Late (Carnian) Triassic(see Flynn et al 1999 2000)
Revised DiagnosismdashTraversodontid with the followingunique combination of features four large upper incisors firstand second upper incisors and all lower incisors procumbentthird and fourth upper incisors caniniform with fourth upperincisor strongly recurved serrated upper canines small equal in
dorsoventral length to incisors distinctly canted forwards (auta-pomorphy) canine alveolus proportionally narrower than incisoralveoli with a much greater anteroposterior than labiolinguallength (autapomorphy) no diastema between upper incisors andcanine low number of postcanines (8 uppers and 6ndash7 lowers)upper and lower postcanines quadrangular roughly trapezoidalin outline descending process of jugal posterior to the last upperpostcanine axial spine with concave dorsal profile and elongateposterior process overhanging shortened neural spine of subse-quent cervical vertebra expanded ribs present caudal neuralspines very tall anterior edge of the iliac blade sloping upwardsat a sim45deg angle acetabular buttresses of the three pelvic boneslargely confluent posterior process of the iliac blade short anddirected away from the dorsal edge of the ischium ischium andpubis extremely constricted resulting in a large obturator fora-men
CommentsmdashMenadon can be distinguished from all other tra-versodontids by the combination of four large upper incisors andincisor procumbency The presence of four upper incisors isprimitive for traversodontids (Hopson and Kitching 2001) En-largement of the incisors is an advanced feature present in suchtaxa as Exaeretodon Scalenodontoides and lsquoScalenodonrsquo hirsch-soni (Abdala and Ribeiro 2003 Abdala et al 2006) Incisorprocumbency is otherwise only known in Exaeretodon and Sca-lenodontoides among traversodontids but these taxa have onlythree upper incisors (Bonaparte 1962 Hopson 1984) Addition-ally Menadon has a descending process of the jugal that contactsthe pterygoids and excludes the maxilla from the margin of thesubtemporal fenestra which is present in most gomphodonts butnot in Exaeretodon or Scalenodontoides (Abdala et al 2006)
Because the postcranium is known in few traversodontid taxathe combination of pelvic characters listed in the Diagnosis canonly be said to diagnose Menadon in comparison with LuangwaPascualgnathus Massetognathus and Exaeretodon The postcra-nium of Menadon resembles that of Luangwa in the shape of theiliac blade the large obturator foramen and the heterogeneousrib morphology but resembles Exaeretodon in the tall caudalneural spines confluent acetabular buttresses and short poste-rior process of the iliac blade projecting away from the ischium
DESCRIPTION
Skull
The cranium of FMNH PR 2444 is poorly preserved and nosutures are visible Six portions of the skull are represented theentire rostrum and left orbitalzygomatic region (Figs 1ndash3) partsof both zygomatic arches (Fig 4AndashC) a fragment of the tempo-ral region including the sagittal crest (Fig 4D) and the occiputand basicranium (Figs 4E 5) The following description is basedon the left side of the skull The right side of the rostrum hasbeen sheared moving some cranial landmarks (eg naris al-veoli) out of their original positions The frontal region on bothsides of the skull has also been sheared anterolaterally towardsthe left such that a shelf of bone made up of the nasals leftprefrontal and left lacrimal overhangs the snout (see Figs 12A)
The rostrum is tall and robust as in Gomphodontosuchus(Hopson 1985) and juveniles of Exaeretodon (Abdala et al2002) The external naris is large and teardrop-shaped narrow-ing towards the rear (Fig 1) A well-developed internarial bar ispresent The maxilla curves inward posterior to the root of thecanine so that the postcanine tooth row is medially inset locatedon an axis posterior to the paracanine fossa (Fig 2B) A singlelateral maxillary foramen is present The orbit is large and sub-circular Prominent interorbital depressions are present on theskull roof (Fig 2A)
Other than a fragment of the left fourth incisor the skull lacks
JOURNAL OF VERTEBRATE PALEONTOLOGY VOL 28 NO 2 2008446
Dow
nloa
ded
by [
UZ
H H
aupt
bibl
ioth
ek
Zen
tral
bibl
ioth
ek Z
uumlric
h] a
t 05
51 1
0 Se
ptem
ber
2013
teeth entirely Alveoli are preserved for all tooth loci except theinferred anterior postcanines (five postcanine alveoli are pre-served on the left side) The paracanine fossa is ovoid locatedmedial to the canine alveolus and much larger than all toothalveoli (Fig 3) Paired incisive foramina are located medial tothe paracanine fossae and extend from the anterior edge of thefourth incisor to the posterior edge of the canine The postca-nines are few in number (minimally five in this individual butprobably eight see Discussion) compared to many traversodon-tids (Romer 1967 Crompton 1972 Abdala et al 2002) andtheir alveoli are large and roughly trapezoidal in outline Thisconfiguration is similar to that observed in Gomphodontosuchus(Hopson 1985)
The left zygomatic arch is composed almost entirely of thejugal and as in most traversodontids it is relatively deep (Flynnet al 2000 Hopson and Kitching 2001) The posterior portionwhere the jugal would have articulated with the squamosal isbroken The isolated right jugal is more complete and its pos-terior articulation with the squamosal is intact (Fig 4A B) Thedorsal edge of the jugal is broken and is missing the posteriorextension over the squamosal The squamosals are very delicatebut well preserved bones each consisting of a cranial and zygo-matic ramus (Fig 4AndashC) The cranial ramus is an extremely thinwedge of bone directed dorsally that would have flanked therear of the braincase in the complete skull The zygomatic ramusis subdivided into a process extending anteriorly and one extend-ing ventrally with a pair of facets between them where the squa-mosal receives the posterior end of the jugal (Fig 4A C) Theanterior process is thin much longer than the ventral processand tapers forward The ventral process is short blunt and wideand forms the lateral portion of the external auditory meatus
The occiput and foramen magnum are tall and narrow (Fig4E) The occipital condyles are separated by a prominent groove
and are also taller than they are wide (but see Discussion) Abroken fragment of the proatlas is preserved out-of-place abovethe foramen magnum (Fig 4E) The occipital fragment includesthe posterior part of the basicranium The basicranium (Fig 5) issimilar to that described for Massetognathus (Rougier et al1992) and Exaeretodon (Abdala et al 2002) However FMNHPR 2444 differs from these in the possession of a basipterygoidforamen Such a foramen has been described by Parrington(1946) in the early cynodont Thrinaxodon but is not known intraversodontids This foramen is not evident in the holotype ofMenadon and this character may be variable in cynodonts
Axial Skeleton
A total of 21 vertebrae are preserved in FMNH PR 2444 Theangle of zygapophyseal articulation varies along the length of thevertebral column This angle (from the sagittal plane) is sim45deg forthe axisC3 (cervical 3) articulation sim30deg for the posterior cervi-cals lt15deg for the dorsals and sim20-30deg for the caudals Other thanthe atlas all vertebral centra are amphicoelous
Atlas-Axis ComplexmdashThree intercentra are articulated withthe cervical series IC2 (fused to the atlas-axis complex) IC3(between C2 and C3) and IC4 (between C3 and C4) IC2 makesup the lower part of the anterior face of the atlas-axis complexand extends back to the ventral anterior edge of the axial cen-trum (Fig 6) The free intercentra (IC3 and IC4) are singleirregularly shaped ossicles situated at the ventral margins of theintercervical articulations (Fig 6) The intercentra of FMNH PR2444 differ markedly from those described for Thrinaxodon(Jenkins 1971) but in general shape and position resemble thosefigured by Brink (1955) for Diademodon The intercentra ofThrinaxodon are transversely wide and anteroposteriorly short(Jenkins 1971fig 2) whereas the intercentra of Menadon be-
FIGURE 1 Stereopair and interpretive drawing of main cranial fragment of FMNH PR 2444 in left lateral view Scale bar equals 5 cm Abbre-viationsmdashJU jugal LA lacrimal MA maxilla mfo lateral maxillary foramen NA nasal nar naris orb orbit PF prefrontal PM premaxilla POpostorbital SM septomaxilla Sutures are not visible in this specimen and have been reconstructed based on the holotype of Menadon besairiei
KAMMERER ET ALmdashCYNODONT FROM MADAGASCAR 447
Dow
nloa
ded
by [
UZ
H H
aupt
bibl
ioth
ek
Zen
tral
bibl
ioth
ek Z
uumlric
h] a
t 05
51 1
0 Se
ptem
ber
2013
FIGURE 2 Stereopairs of main cranial fragment of FMNH PR 2444 in A dorsal view and B palatal view Scale bars equal 5 cm
JOURNAL OF VERTEBRATE PALEONTOLOGY VOL 28 NO 2 2008448
Dow
nloa
ded
by [
UZ
H H
aupt
bibl
ioth
ek
Zen
tral
bibl
ioth
ek Z
uumlric
h] a
t 05
51 1
0 Se
ptem
ber
2013
sairiei have equal transverse and anteroposterior dimensions Attheir maximum depth in Thrinaxodon the intercentra barely ex-tend beyond the ventral margins of the cervical pleurocentra InFMNH PR 2444 IC4 extends for half of its total depth below theventral margins of the cervical pleurocentra In IC3 this condi-tion is even more extreme extending below the ventral marginsof the cervical pleurocentra for 80 of its total depth The shapeand position of the intercentra give a strongly arched appearanceto the ventral profile of elements in the cervical series The ven-tral surfaces of the intercentra bear a posteriorly directed process
on each side In IC3 a weak ridge originates between the pairedprocesses and extends posteriorly indicating muscle attachmenton the intercentra The condition in IC4 cannot be determinedbecause of incomplete preservation Although only three inter-centra are known ventral facets in the centra of the isolatedcervicals indicate that others were present
As in other cynodonts the atlas-axis complex of FMNH PR2444 consists of a pair of separate atlantal neural arches articu-lating with the occipital condyles and the atlas centrum Only theleft atlas arch is preserved in this specimen (Fig 7) It is similar
FIGURE 3 Stereopair and interpretive drawing of anterior palatal region of FMNH PR 2444 Scale bar equals 5 cm AbbreviationsmdashC canineI incisor i f incisive foramina PC postcanine pc f paracanine fossa Postcanine numbering based on comparison with the holotype UA 10601
FIGURE 4 Cranial fragments of FMNH PR 2444 A stereopair of right zygomatic arch in lateral view Scale bar equals 5 cm B stereopair of rightzygomatic arch in medial view Scale bar equals 5 cm C stereopair of left squamosal in lateral view Scale bar equals 5 cm D sagittal crest in rightlateral view Scale bar equals 1 cm E occiput in occipital view Scale bar equals 1 cm
KAMMERER ET ALmdashCYNODONT FROM MADAGASCAR 449
Dow
nloa
ded
by [
UZ
H H
aupt
bibl
ioth
ek
Zen
tral
bibl
ioth
ek Z
uumlric
h] a
t 05
51 1
0 Se
ptem
ber
2013
to the atlantal neural arches described for other cynodonts suchas Massetognathus (Kemp 1969 Jenkins 1971) There is a me-dial facet for articulation with the centrum of the atlas (theldquoodontoid facetrdquo of Kemp 1969) and an anterior facet for ar-ticulation with the occipital condyle (Fig 7B) A pair of pro-cesses dominates the atlas arch a dorsal lamina (a roughly rect-angular plate of bone arching above the facets) and a largeposterolaterally directed transverse process An oval tubercle atthe anterior end of the transverse process represents the facet forthe proatlas
The centrum of the atlas is fused to the axis forming a singlelarge vertebral unit (Fig 6) A weak odontoid process is presentat the dorsal limit of the anterior surface of the atlas The ante-rior aspect of the centrum is dominated by a roughly lsquoTrsquo-shapedstructure composed of the two facets for articulation with theatlas arches and the facet for articulation with the atlas intercen-trum The axis bears no prezygapophyses and as such does notarticulate with the atlas arch as in Early Triassic cynodonts suchas Galesaurus (Parrington 1934) There appears to be a trendtowards loss of the axial prezygapophyses in later traversodon-
tids which are present but vestigial in Massetognathus (Jenkins1970 although in the specimens described the prezygapophysesare broken so their reduction is perhaps questionable) and ab-sent in Exaeretodon (Bonaparte 1963) The transverse processesof the axis are large and robust compared to those of the othercervicals The axial spine is saddle-shaped with a concave dorsalprofile unlike previous reconstructions of the traversodontidaxis which depict a straight or convex dorsal profile similar tothat observed in galesaurid cynodonts and early mammals (Jen-kins 1971) The morphology of the axial spine is similar to thatof the Early Triassic cynodont Cynognathus (Seeley 1895) InCynognathus however the neural spine of the third cervical(C3) is tall and canted posteriorly behind the posterior process ofthe axial spine whereas in FMNH PR 2444 the short C3 neuralspine fits below the posterior portion of the axial spine Theposterior process of the axial spine bears a pair of parallel elon-gate grooves on its underside attachment sites for the axial mus-culature
Post-Axial VertebraemdashThree post-axial cervicals are knownfor FMNH PR 2444 (Figs 6 8AndashD) The first of these C3 is
FIGURE 5 Stereopair of ventral view of the occipital fragment of FMNH PR 2444 showing the basicranium Anterior is up
FIGURE 6 Stereopair and interpretive drawing of atlas-axis complex and third cervical of FMNH PR 2444 in left lateral view Scale bar equals5 cm Abbreviationsmdasha n sp axial neural spine d dens IC intercentrum f at a facet for articulation with the atlas arch p parapophysis pozpostzygapophysis t p transverse process
JOURNAL OF VERTEBRATE PALEONTOLOGY VOL 28 NO 2 2008450
Dow
nloa
ded
by [
UZ
H H
aupt
bibl
ioth
ek
Zen
tral
bibl
ioth
ek Z
uumlric
h] a
t 05
51 1
0 Se
ptem
ber
2013
preserved in articulation with the axis (Fig 6) The two isolatedcervicals bear successively taller and anterposteriorly wider neu-ral spines (Fig 8AndashD) Based on their staggered size and failureto articulate we suggest that they represent C5 and C7 (assumingthat this specimen possessed the generalized cynodont cervicalnumber of seven) The cervicals display robust transverse pro-cesses which are directed laterally and slightly anteriorly A par-apophysis lies below the anterior edge of the transverse processalong the antero-lateral edge of the centrum No cervical ribs arepreserved but their presence can be inferred from the par-apophyses The notochordal concavity is as in Massetognathussurrounded by a raised rim at each end of the centrum (Jenkins1970) No anapophyses are present The neural spines for allthree post-axial cervicals are tapering and slightly curved poste-riorly The neural spine in C3 is relatively short and wide(thereby fitting underneath the posterior overhang of the axialspine see Fig 6) while that of C5 is tall and narrow The spinefor C7 is taller and anteroposteriorly wider than that of C5 andthe dorsal edge is more rounded similar to the condition in thedorsal vertebrae
Eight post-cervical presacral vertebrae are known for thisspecimen We will refer to them here simply as dorsal vertebraegiven the incompleteness of the vertebral column in the speci-men and the uncertainty concerning the thoraciclumbar transi-tion in this taxon However it is probable that they representthoracic vertebrae (see below) In most cynodonts the lumbar
vertebrae are synostosed to specialized ribs with overlappingelements In earlier taxa (eg Thrinaxodon Cynognathus Lu-angwa Jenkins 1971 Kemp 1980) expanded costal plates arepresent on both thoracic and lumbar ribs In Massetognathus thelumbar ribs consist of a pair of prongs one directed anteriorlybearing a facet for articulation with the previous rib and theother directed posteriorly (Jenkins 1970) In Exaeretodon allpresacral ribs are identical of simple morphology and not syn-ostosed to the vertebrae (Bonaparte 1963) A single isolated rib(Fig 9B) in FMNH PR 2444 demonstrates that some of the ribsin Menadon besairiei were expanded However all preservedvertebrae in this specimen have facets for articulation with theribs rather than breakage points where they would have beenfused to ribs As such it is probable that all of the vertebraepreserved in this specimen represent thoracics although we can-not discount the possibility that as in Exaeretodon no presacralvertebrae were fused to ribs
The neural spines in all of the dorsal vertebrae are elongateposteriorly inclined and roughly rectangular in lateral view witha rounded laterally expanded platform dorsally (Fig 8EndashJ)Costal foveae are present at the anterior and posterior edges ofeach centrum Of the eight dorsal vertebrae there are two ar-ticulated sets and three isolated elements (two from 9-4-98-440and one from 9-2-98-422) The first isolated vertebra (Fig 8E F)has a taller neural spine and shorter centrum than the otherdorsal vertebrae It is most similar in proportions to the isolated
FIGURE 7 Stereopairs and interpretive drawings of left atlas arch of FMNH PR 2444 in A lateral view Anterior is left B medial view Anterioris right Scale bars equal 1 cm Abbreviationsmdashafa anterior facet for articulation with the occipital condyle dl dorsal lamina mfa medial facet forarticulation with the atlas centrum (odontoid facet) pfa tubercle forming the facet for articulation with the proatlas tp transverse process
KAMMERER ET ALmdashCYNODONT FROM MADAGASCAR 451
Dow
nloa
ded
by [
UZ
H H
aupt
bibl
ioth
ek
Zen
tral
bibl
ioth
ek Z
uumlric
h] a
t 05
51 1
0 Se
ptem
ber
2013
FIGURE 8 Vertebrae of FMNH PR 2444 Cervical vertebra 5 in A left lateral and B anterior views Cervical vertebra 7 in C left lateral andD anterior views Isolated dorsal vertebra from anterior region of the series in E left lateral and F anterior views G stereopair and interpretivedrawing of articulated set of two dorsal vertebrae from near the midpoint of the series in left lateral view Anterior is up H articulated set of threedorsal vertebrae from anterior region of the series in left and right lateral views I isolated vertebra from near the midpoint of the series (from fieldnumber 9-2-98-422) J isolated vertebra from posterior region of the series Sacral vertebra in K dorsal view (the distal region of another sacral ribwhich is identical in shape to that of the intact sacral rib albeit slightly larger is also shown in this figure) and L left lateral view Caudal vertebraeM articulated set of two vertebrae from the anterior region of the series showing the lsquowingrsquo-shaped caudal ribs in dorsal view Anterior is left Nisolated caudal vertebra showing tall neural spine in left lateral view O isolated caudal vertebra showing tall neural spine in posterior view Particulated set of four vertebrae from near the end of the caudal series in dorsal view Anterior is left Scale bars equal 1 cm Abbreviationsmdasha cof anterior costal fovea n sp neural spine p co f posterior costal fovea poz postzygapophysis prz prezygapophysis
JOURNAL OF VERTEBRATE PALEONTOLOGY VOL 28 NO 2 2008452
Dow
nloa
ded
by [
UZ
H H
aupt
bibl
ioth
ek
Zen
tral
bibl
ioth
ek Z
uumlric
h] a
t 05
51 1
0 Se
ptem
ber
2013
cervicals and was probably located near the front of the dorsalseries The first articulated set consists of three vertebrae (Fig8H) The middle vertebra in this set is broken from the anteriorbase of the neural spine to the ventral midpoint and most of theright side of the anterior half of the centrum is missing This setof vertebrae probably also came from the anterior region of theseries as their neural spines are longer and narrower than allvertebrae other than the one described above The second ar-ticulated set (Fig 8G) consists of two well-preserved vertebraethat based on comparisons with Massetognathus and Ex-aeretodon (Jenkins 1970 Bonaparte 1963) probably came fromnear the midpoint of the dorsal series The second isolated ver-tebra (Fig 8I) from 9-2-98-422 is similar to the members of thesecond set The third isolated vertebra (Fig 8J) appears to befrom the posterior end of the series based on the relatively shortneural spine and reduced posterior costal fovea
One sacral vertebra (Fig 8K L) is known consisting of awell-preserved centrum neural spine and the right sacral rib(the left is broken off) The centrum is shorter (both anteropos-teriorly and dorsoventrally) than those of the dorsal vertebraeand the neural spine is shorter and more rounded in profileAlso the neural spine is directed vertically unlike those of thedorsal vertebrae which are canted posteriorly The zygapophy-ses are poorly developed The sacral rib is fused to a singlesynapophysis and projects anterolaterally The rib narrows to-wards the middle and expands distally terminating in a widerectangular facet where it would meet the ilium (Fig 8K)
Seven caudal vertebrae are known The caudal vertebrae aredistinctive in having very tall narrow neural spines Some EarlyTriassic cynodonts possess blunt neural spines of moderateheight on the proximal caudals (see for example Thrinaxodonin Jenkins 1971 and Diademodon in Brink 1955) Massetogna-thus represents an extreme condition wherein the caudal neuralspines are greatly reduced (Jenkins 1970) Very tall neuralspines are seen in Exaeretodon (Bonaparte 1963) and thosespines are unusual in that they do not diminish in height until thelast caudals The caudal vertebrae of FMNH PR 2444 consist ofa pair of articulated vertebrae most likely from near the sacrum(Fig 8M) an isolated caudal lacking postzygapophyses and ribsbut with the neural spine (Fig 8N O) and a quartet of smaller
articulated elements (Fig 8P) The anterior caudals have nar-rower neural spines and shorter zygapophyses than the subse-quent vertebrae in the series The caudal ribs attached to thispair have a crescentic distal region that curves anteriorly pro-ducing a lsquowingrsquo-shape Only a portion of the right rib of theisolated caudal is preserved but the lsquowingrsquo-shaped morphologyof the near-sacral caudal ribs does not appear to have been pre-sent In the posteriormost four vertebrae caudal ribs are pre-served on the left side of the first and the right side of the secondand third members On the first of these vertebrae the rib isshort curved posteriorly (but not lsquowingrsquo-shaped) and taperingRibs of the second and third vertebrae are less curved resem-bling posteriorly directed rods The zygapophyses of both theisolated caudal vertebra and the quartet of small caudals areanteroposteriorly elongate becoming increasingly so posteriorly
Thoracic RibsmdashNine thoracic ribs are known from 9-4-98-440(four left four right and an indeterminate splint of bone Fig9A indeterminate bone not illustrated) These elements re-semble the flattened ribs lacking costal plates of Exaeretodon(Bonaparte 1963) and the anterior thoracic ribs of Massetogna-thus and Luangwa (Jenkins 1970 Kemp 1980) The tuberculumand capitulum are separated by a shallow depression The tuber-culum has a shorter base than the capitulum but their proximalfaces are subequal in size An additional isolated left rib (Fig9B) from 9-2-98-422 is expanded indicating that unlike Ex-aeretodon Menadon rib morphology was heterogeneous alongthe length of the dorsal series This rib closely resembles theposterior thoracic ribs described for Luangwa (Kemp 1980fig16) Pascualgnathus (Bonaparte 1966fig 7) and the trirach-odontid Cricodon (Crompton 1955 Fig 4) This rib is con-stricted at the base of the head then expands distally with an-terior and posterior costal plates As in Luangwa the anteriorcostal plate is shallow and evenly sloping proximally and distallywhile the posterior costal plate is falcate with a concave edgedistally The distal expansion of this rib occurs at an angle of sim30degto the long axis of the rib head Despite their overall similaritythe expanded rib in FMNH PR 2444 does exhibit some differ-ences from the posterior thoracic rib of Luangwa The expandedrib of Menadon has a longer shaft between the rib head and thecostal plates and a shorter capitulum lacks a ridge on the pos-
FIGURE 9 A anterior thoracic ribs of FMNH PR 2444 in anterior view (indeterminate rib not figured) B expanded posterior thoracic rib ofFMNH PR 2444 in anterior view Scale bars equal 1 cm
KAMMERER ET ALmdashCYNODONT FROM MADAGASCAR 453
Dow
nloa
ded
by [
UZ
H H
aupt
bibl
ioth
ek
Zen
tral
bibl
ioth
ek Z
uumlric
h] a
t 05
51 1
0 Se
ptem
ber
2013
terior costal plate and has an anteroposteriorly shorter anteriorcostal plate Additionally the posterior margin of the costal plateis less curved than that of Luangwa These differences couldindicate that the expanded rib of Menadon is from a more pos-terior position in the series than the rib from Luangwa as re-duced curvature and increasing confluence of the tuberculumand capitulum are characteristic of lumbar ribs in traversodon-tids (Jenkins 1970 Kemp 1980) However the length of theshaft is at odds with this interpretation and in Cricodon andPascualgnathus the more posterior ribs are not more similar tothe element from FMNH PR 2444 These differences probablyrepresent legitimate morphological distinctions between the ribsof earlier gomphodonts and Menadon The head of the expandedrib is well defined and unbroken indicating that it was not syn-ostosed to a vertebra
Appendicular Skeleton
The appendicular skeleton is represented by both scapulocora-coids and a fragment of a clavicle from field number 9-4-98-440and a partial right pelvis from field number 9-2-98-422 An ad-ditional partial right pelvis is represented from field number8-31-98-387 The scapulocoracoids (Figs 10 11) are very similarto those described for other traversodontids as well as cynog-nathids and diademodontids (Jenkins 1970 1971) The scapularinfraspinous fossa has out-turned anterior and posterior edgesThe anterior edge of the scapula is rounded and curves laterallyforming the scapular spine At the lower end of the spine theacromion forms a prominent process (broken in the left scapula)Below this process the scapula is constricted and then expandsventrally to form the dorsal margin of the articular glenoid Theglenoid fossa is in general morphology very similar to that ofother Triassic cynodonts (Jenkins 1970 1971 Kemp 1980) It isposteroventrally oriented with roughly equal contributions fromthe scapular and coracoid glenoid facets The coracoid glenoidfacet is roughly triangular while the scapular glenoid facet issemiovoid as in other cynodonts In anterior or posterior viewthe fossa appears smoothly crescentic The procoracoid is aslightly curved trapezoidal plate of bone It does not appear tocontribute to the glenoid fossa The procoracoid foramen is lo-cated midway up the height of the procoracoid at a point directlybelow the base of the acromion process The coracoid is semi-crescentic curving posteriorly from a broad base The coracoidin Menadon is longer and more falcate than those described forExaeretodon (Bonaparte 1963) and Luangwa (Kemp 1980 al-though the distal tip of both coracoids is broken in the specimenof Luangwa [TSK 121] described by Kemp and may have beenlonger than reconstructed) but agrees closely with those of Mas-setognathus and cynognathidsdiademodontids (Jenkins 19701971) The distal coracoid tubercle for attachment to the tricepsis strongly curved upwards similar to the condition in Masseto-gnathus (Jenkins 1970) and Diademodon (Brink 1955)
A small fragment of bone resembling a curved cylinder almostcertainly represents part of a clavicle It has no distinguishingfeatures other than a groove associated with a ridge on one sideHowever the degree of curvature observed precludes this ele-ment from representing a long bone and the circular cross-section excludes it from being a rib
The pelvis of FMNH PR 2444 (Fig 12) resembles those ofother traversodontids in general morphology The ilium bears anarrow anteriorly directed iliac blade curving outward distallyThe anterior edge of the iliac blade slopes upward at a roughly45deg angle similar to Luangwa (Kemp 1980) but unlike Ex-aeretodon in which the anterior edge of the iliac blade curvesbackwards (Bonaparte 1963) or Pascualgnathus and Masseto-gnathus in which it is rounded (Bonaparte 1966 Jenkins 1970)The supraacetabular buttress at the base of the ilium is located inan anterodorsal position relative to the acetabulum as in other
traversodontids but unlike earlier cynodonts in which it is lo-cated dorsally (Jenkins 1970) The acetabular buttresses of thethree pelvic bones are largely confluent forming a fairly uniformlsquoliprsquo around the acetabulum as in Exaeretodon (Bonaparte1963) On the contrary in Luangwa the supraacetabular ischialand pubic buttresses are highly distinct spaced 6ndash8 mm apart andwithout confluence of their acetabular facets The pelvis ofFMNH PR 2444 does have a small depression between the su-praacetabular and ischial buttresses (also present in Ex-aeretodon Bonaparte 1963) but this is not as developed as inLuangwa The posterior extent of the iliac blade is broken inFMNH PR 2444 but this region is preserved in another isolatedright pelvis field number 8-31-98-387 (Fig 13) probably pertain-ing to Menadon besairiei The posterior process of the iliac bladein this specimen is relatively short and directed away from thedorsal edge of the ischium as in Exaeretodon (Bonaparte 1963)but unlike Massetognathus and Luangwa in which the posteriorprocess is long and extends roughly horizontally over the dorsaledge of the ischium (Jenkins 1970 Kemp 1980) and evencurves downward distally in Luangwa The posterior process ofthe iliac blade in Pascualgnathus is directed away from the is-chium (Bonaparte 1966) as in Exaeretodon and Menadon but isrelatively long as in Massetognathus and Luangwa
The proximal portion of the ischium is broad and dominatedby the ischial buttress The ischium constricts posterior to theproximal portion and expands distally into a spatulate platewhich is thickened at its posterior margin This thickening con-tinues along the dorsolateral surface of the ischium forming aridge that slopes ventrally until reaching the edge of the ischialbuttress The downward slope of this ridge produces a prominentgroove at the dorsal edge of the lateral surface of the ischiumthought to represent the origin of the ischio-trochantericusmuscle (Kemp 1980) The constriction of the ischium in Mena-don is like that of Luangwa albeit even more extreme in itsventral concavity but unlike that of Exaeretodon in which themedial and distal ends of the ischium are broadly united Theischium in general is a proportionally shorter element in Ex-aeretodon than in other traversodontids including Menadon(Bonaparte 1963) The anteroventral portion of the ischium thatconnects to the pubis is not preserved in either of the referredMenadon pelves
The pubis is the most incompletely preserved pelvic elementThe acetabular contribution of the pubis is smaller than that ofthe ischium or ilium Distal to the acetabulum the pubis is con-stricted indicating that the connection between the acetabularregion and the pubic plate was considerably narrower than inExaeretodon (Bonaparte 1963) but close akin to the pubis ofLuangwa (Kemp 1980) The pubic plate itself is not preservedThe ventral outline of the ischium and what is known of thepubis indicate that the obturator foramen was relatively larger inMenadon (Fig 14) than in Exaeretodon (Bonaparte 1963)
DISCUSSION
Comparison with the Holotype of Menadon besairiei
FMNH PR 2444 differs in several significant ways from theholotype of Menadon besairiei as previously described most no-tably in the possession of an internarial bar (initially consideredabsent in M besairiei) and four upper incisors (reported as threein M besairiei but with the caveat that the homologies of thefourth and fifth anterior tooth loci were problematic Flynn et al2000) However upon further study we interpret the lack of acontinuous internarial bar in the holotype as breakage ratherthan the natural condition of the skull (a prominent medial pro-tuberance is present on the ventral rim of the nares in UA10601) Regarding the incisor count it should be noted that theholotype of Menadon besairiei displays an unusual set of anterior
JOURNAL OF VERTEBRATE PALEONTOLOGY VOL 28 NO 2 2008454
Dow
nloa
ded
by [
UZ
H H
aupt
bibl
ioth
ek
Zen
tral
bibl
ioth
ek Z
uumlric
h] a
t 05
51 1
0 Se
ptem
ber
2013
FIGURE 10 Right scapulocoracoids of FMNH PR 2444 A stereopair of right scapulocoracoid in medial view B stereopair and interpretivedrawing of right scapulocoracoid in lateral view Scale bars equal 5 cm Abbreviationsmdashac acromion process C coracoid c gl coracoid contributionto the glenoid fossa c tr tubercle for coracoid attachment of triceps PC procoracoid pc f procoracoid foramen S scapula s gl scapularcontribution to the glenoid fossa
KAMMERER ET ALmdashCYNODONT FROM MADAGASCAR 455
Dow
nloa
ded
by [
UZ
H H
aupt
bibl
ioth
ek
Zen
tral
bibl
ioth
ek Z
uumlric
h] a
t 05
51 1
0 Se
ptem
ber
2013
teeth for a traversodontid As mentioned above Flynn et al(2000) noted that the homologies of these teeth were difficult todetermine Of the five anterior teeth preserved in the holotypethe anteriormost three are ldquoundoubtedly incisorsrdquo although I3 isldquounusually large and is caniniformrdquo (Flynn et al 2000423) Therobust recurved fourth tooth in the series was considered torepresent the canine with the subsequent tooth interpreted as anolder canine in the process of replacement Although FMNH PR2444 is edentulous the alveoli on the left side of the skull are wellpreserved and indicate an anterior tooth set identical to the ho-lotype The alveolar morphology indicates that the fifth toothwas canted anteriorly as in the holotype This suggests that themysterious fifth tooth is not the result of a transient period in thedental replacement history of M besairiei as suggested by Flynnet al (2000) but rather is the norm This leaves two possibilities1) M besairiei persistently had two canines This condition isunknown in cynodonts 2) The canine-like fourth tooth actuallyrepresents an incisor and the fifth tooth is the true canine Wefavor the latter explanation In FMNH PR 2444 the first fouralveoli are nearly identical in size and shape However the fifthalveolus while similar in anteroposterior length to its predeces-sors is strongly compressed labiolingually with an anteroposte-rior lengthlabiolingual width ratio of 044 (compared to 080 inthe fourth alveolus and 084ndash85 for I2ndash3) We interpret thismarked difference in alveolar ratio as indicating the separationbetween incisors and canine The canine-like I4 would then rep-resent the endpoint in a series of large increasingly caniniformincisors Supporting this interpretation recurved incisors are alsoobserved in juvenile Exaeretodon (eg MCP 3843 PV see Ab-dala et al 2002)
There are several legitimate differences between FMNH PR2444 and the holotype of Menadon besairiei but we take a con-servative taxonomic stance and argue that these differences rep-resent intraspecific variation and preservational artifacts FMNHPR 2444 represents a larger animal than UA 10601 The snoutlength (measured as the distance between the anterior edge ofthe orbit and a vertical plane through the anterior tip of thepremaxilla) of FMNH PR 2444 is 911 mm while that of UA10601 is 801 mm As such proportional differences between thetwo skulls may be attributable to ontogeny Variations in FMNHPR 2444 compared to the holotype that probably represent on-togentic differences include wider longer and deeper paraca-nine fossae a much deeper ventral zygomatic flange (althoughthe lower border of the flange is slightly abraded in the holotypeof Menadon besairiei there is no evidence that a major flangewas broken off) a proportionally shorter and more circular out-line to the posterior border of the maxilla below the zygomaticprocess and a more deeply emarginated posterior squamosal(ie a more prominent laterally projecting flange)
FMNH PR 2444 has a proportionally shorter more robustsnout and larger orbits than the holotype While these charactersare certainly variable in cynodonts it is unusual that the largerspecimen is shorter-snouted and larger-eyed contra the usualpattern The proportional differences between the two skulls areminor however (orbit diametersnout length is 042 for FMNHPR 2444 and 041 for the holotype) and are partially attributableto deformation (shear in FMNH PR 2444 and breakage of thepostorbital bar in the holotype) The following differences canalso probably be attributed to preservational artifact only fivepostcanine alveoli are preserved in FMNH PR 2444 while theholotype has eight postcanines The anteriormost three in theholotype however are tiny and peg-like As the area of themaxilla between the canine alveolus and the first preserved post-canine alveolus is poorly preserved in FMNH PR 2444 the ab-sence of these postcanines may be artifactual However it is alsopossible that the lower postcanine count of FMNH PR 2444compared to the holotype is an ontogenetic effect as postcaninecount is known to decrease with increasing skull size in Ex-
FIGURE 11 Left scapulocoracoids of FMNH PR 2444 A left scapu-locoracoid in medial view B left scapulocoracoid in lateral view Scalebars equal 5 cm
JOURNAL OF VERTEBRATE PALEONTOLOGY VOL 28 NO 2 2008456
Dow
nloa
ded
by [
UZ
H H
aupt
bibl
ioth
ek
Zen
tral
bibl
ioth
ek Z
uumlric
h] a
t 05
51 1
0 Se
ptem
ber
2013
FIGURE 12 Right pelvis of FMNH PR 2444 A stereopair of right pelvis in medial view B stereopair and interpretive drawing of right pelvis inlateral view Scale bars equal 5 cm Abbreviationsmdashbpp base of posterior process of ilium ib iliac blade IL ilium IS ischium ob f obturatorforamen P pubis
KAMMERER ET ALmdashCYNODONT FROM MADAGASCAR 457
Dow
nloa
ded
by [
UZ
H H
aupt
bibl
ioth
ek
Zen
tral
bibl
ioth
ek Z
uumlric
h] a
t 05
51 1
0 Se
ptem
ber
2013
aeretodon (Abdala et al 2002) The orbit of FMNH PR 2444 isless upwardly directed than that of the holotype (being at anangle of sim65deg to a sagittal plane through the midline of the skullas opposed to sim45deg in the holotype) This difference in orbitalangle is probably attributable to postmortem deformation as theskull of FMNH PR 2444 is noticeably sheared to the front leftThe appearance of a wider squarer incisor arcade and broader
anterior palate in FMNH PR 2444 can also be attributed to thisshear Finally the occipital region of FMNH PR 2444 is alto-gether taller and narrower than that of the holotype For ex-ample the foramen magnum in FMNH PR 2444 approximates arounded isosceles triangle in shape while the foramen magnumof the holotype is more ovoid These differences can also beattributed to deformation Altogether the differences between
FIGURE 13 Stereopairs of field number 8-31-98-387 an isolated right pelvis in A medial and B lateral view Scale bars equal 5 cm
JOURNAL OF VERTEBRATE PALEONTOLOGY VOL 28 NO 2 2008458
Dow
nloa
ded
by [
UZ
H H
aupt
bibl
ioth
ek
Zen
tral
bibl
ioth
ek Z
uumlric
h] a
t 05
51 1
0 Se
ptem
ber
2013
FMNH PR 2444 and the holotype fall within the range of intra-specific variation recorded for other gomphodont cynodontssuch as the traversodontids Massetognathus pascuali (Abdalaand Giannini 2000 Kammerer pers obs) and Exaeretodon(Abdala et al 2002) and the diademodontid Diademodon tetra-gonus (Grine et al 1978 Bradu and Grine 1979)
Phylogenetic Analysis
Utilizing the new information afforded by FMNH PR 2444and a reexamination of the holotype (UA 10601) Menadon be-sairiei was recoded in the Gondwanan gomphodont data matrixof Abdala et al (2006) Abdala et alrsquos (2006) data matrix en-compassed 18 taxa and 43 characters For the present analysis anadditional character ldquoincisor procumbencyrdquo (states 0-absent1-present) was added to Abdala et alrsquos (2006) data set (see
Appendix for coding of this character) Several traversodontidtaxa (eg lsquoScalenodonrsquo hirschsoni) have relatively large incisorsbut only in Menadon Scalenodontoides and Exaeretodon are theincisors procumbent Three of Abdala et alrsquos (2006) originalcodings for Menadon besairiei characters 0 (ldquonumber of upperincisorsrdquo) 5 (ldquoposition of paracanine fossa in relation to theupper caninerdquo) and 21 (ldquointernarial barrdquo) were changed to in-corporate data from the new specimen (see Appendix) Addi-tionally further examination of the holotype (UA 10601) ofMenadon besairiei allowed us to code characters 16ndash18 20 38and 40 (morphology of the lower postcanines all marked as ldquordquoin Abdala et alrsquos analysis) Parsimony searches were performedin PAUP v40b10 using a branch-and-bound search Addition-ally a bootstrap analysis (also using branch-and-bound) was per-formed using 1000 replicates
A total of 34 most parsimonious trees of length 103 werefound with the scores CI0573 RI0725 RC0415 andHI0427 The strict consensus tree (Fig 15A) did not recovera polytomy between Menadon Exaeretodon and Scalenodont-oides as in Abdala and Ribeiro (2003) or a (Menadon + Ex-aeretodon) sister-group relationship as in Abdala et al (2006)Rather Menadon was recovered as the sister taxon of the (Ex-aeretodon + Scalenodontoides) clade supporting Flynn et alrsquos(2000) original placement of this taxon There is strong bootstrapsupport (88) for Traversodontidae but within traversodontidsthe only strongly supported subclade is (Gomphodontosuchus +(Menadon + (Scalenodontoides + Exaeretodon))) with a boot-strap value of 80 Within this clade the (Menadon + (Scalen-odontoides + Exaeretodon)) grouping is supported by a boot-strap value of 83 and the (Scalenodontoides + Exaeretodon)grouping is supported by a bootstrap value of 94 (Fig 15B) Inaddition to the positioning of Menadon the strict consensus tree(Fig 15A) differs from the topology of Abdala et al (2006) inrecovering a sister-group relationship between Massetognathusand Santacruzodon rather than having Massetognathus andSantacruzodon as successive outgroups to the clade containingGomphodontosuchus Menadon Scalenodontoides and Ex-aeretodon The (Massetognathus + Santacruzodon) clade is diag-nosed by the absence of an anterolingual cusp in the upper post-canines denticulated incisor cutting margins and the presence ofan anterior cingulum in the upper postcanines
FIGURE 14 Reconstruction of the pelvis of Menadon besairiei basedon information from FMNH PR 2444 and field number 8-31-98-387Dotted area not preserved in either specimen reconstruction based onPascualgnathus (Bonaparte 1966) and Exaeretodon (Bonaparte 1963)
FIGURE 15 Cladogram of traversodontid relationships based on revised data set of Abdala et al (2006) showing A strict consensus tree and B50 majority rule tree Numbers at nodes of majority rule tree represent bootstrap values
KAMMERER ET ALmdashCYNODONT FROM MADAGASCAR 459
Dow
nloa
ded
by [
UZ
H H
aupt
bibl
ioth
ek
Zen
tral
bibl
ioth
ek Z
uumlric
h] a
t 05
51 1
0 Se
ptem
ber
2013
Relationships of Menadon besairiei
Menadon besairiei is the sister taxon to (Exaeretodon + Sca-lenodontoides) the latter clade diagnosed by the possession ofthree upper incisors a posteromedial position of the paracaninefossae relative to the canines the absence of an internarial barlarge canines a projected zygomatic process of the jugal and amaxilla that contributes to the margin of the subtemporal fenes-tra (the last three character states represent reversals) The clade(Menadon + (Exaeretodon + Scalenodontoides)) is diagnosed bythe possession of large procumbent incisors The presence offour incisors a primitive retention distinguishes Menadon be-sairiei from Exaeretodon Scalenodontoides and an unnamed tra-versodontid (ldquoTraversodontid Type 1rdquo) from the Santa MariaFormation of Brazil (Abdala et al 2001 Abdala and Ribeiro2002) Gomphodontosuchus is recovered as the sister taxon to(Menadon (Exaeretodon + Scalenodontoides)) as has beenfound in previous studies (Flynn et al 2000 Abdala and Ribeiro2003) The taxon name Gomphodontosuchinae Watson andRomer 1956 (ex Gomphodontosuchidae Watson and Romer1956 first given subfamilial rank in Lehman 1961) is availablefor this clade As circumscribed here Gomphodontosuchinaeconsists of the most inclusive clade containing Gomphodontosu-chus brasiliensis von Huene 1928264 but not Massetognathuspascuali Romer 19672
Menadon besairiei displays a mixture of primitive and derivedcharacters within traversodontids The internarial bar and fourincisors are primitive features whereas the enlargement and pro-cumbency of the incisors are derived features shared with Ex-aeretodon and Scalenodontoides The postcranium shows severalsimilarities to that of Exaeretodon (apparent exclusion of theprocoracoid from the glenoid some features of the pelvis tallcaudal neural spines) but in general displays a rather conserva-tive morphology observed throughout Gomphodontia The mor-phology of the scapulocoracoids is closer akin to diademodontidsor trirachodontids than Exaeretodon and the expanded thoracicrib of Menadon has only slight differences from those of trirach-odontids or early traversodontids like Luangwa The atlas-axiscomplex of Menadon is similar to that of the Early Triassic cyn-odont Cynognathus in possessing a concave axial spine with along posterior process Although most gomphodonts have beenreconstructed with a convex axial spine as in early mammals thisspine is broken in most of the best described gomphodont post-crania (eg Diademodon Brink 1955) and the lsquosaddle-shapedrsquoaxial morphology may be more widely distributed among cyn-odonts than previously realized The new specimens of Menadondescribed in this paper demonstrate that a generalized gompho-dont postcranial morphology was retained deep within the tra-versodontid tree and that the derived morphologies of either ofthe well-known taxa Massetognathus or Exaeretodon should notbe taken as representative of traversodontids in general
ACKNOWLEDGMENTS
This work would not have been possible without the continu-ing support and encouragement of our collaborative programwith the Deacutepartement de Paleacuteontologie et drsquoAnthropologie Bi-ologique Universiteacute drsquoAntananarivo Madagascar under the di-rection of Armand Rasoamiaramanana (Department Chairman)and Gisegravele Ravololonarivo Randria (Head of the PaleontologyLaboratories) We are extremely grateful to the National Geo-graphic Society for their long-term support of the fieldwork as-sociated with this project Additional financial or logistical sup-port for fieldwork was provided by The Field Museum the Uni-versity of California-Santa Barbara and the WWF (WorldwideFund for Nature Madagascar) We acknowledge the Ministegraverede LrsquoEneacutergie et des Mines for authorizing field efforts and for
continuing to facilitate our joint US-Madagascar explorationresearch and education program Steve Goodman and AsminaGandhi generously assisted us in innumerable ways includinghospitality nonpareil We especially thank R Andriantompoha-vana L Razafimanantsoa A Toto Volahy JA Rabarison AGandie and P Vavisaro for assistance in the field during theexpedition in which these new specimens were discovered BillSimpson and Gina Wesley-Hunt for their patient excavation ofthis important material Allison Smith Ray Vodden Lisa Berg-wall and Deborah Wagner for excellent preparation of a chal-lenging specimen Jennifer Botha and Elize Butler (NationalMuseum Bloemfontein) Michael Raath and Fernando Abdala(Bernard Price Institute) Thomas Kemp (Oxford UniversityMuseum of Natural History) Ray Symonds (University Museumof Zoology Cambridge) Bernard Battail (Museacuteum nationaldrsquoHistoire naturelle Paris) Michael Maisch (Universitaumlt Tuumlbin-gen) and Charles Schaff and Farish Jenkins (Museum of Com-parative Zoology Harvard) for access to specimens and AgustiacutenMartinelli Fernando Abdala and James Hopson for helpful re-views of an earlier version of the manuscript
LITERATURE CITED
Abdala F M C Barbarena and J Dornelles 2002 A new species of thetraversodontid cynodont Exaeretodon from the Santa Maria Forma-tion (MiddleLate Triassic) of Southern Brazil Journal of Verte-brate Paleontology 22313ndash325
Abdala F and N P Giannini 2000 Gomphodont cynodonts of theChantildeares Formation the analysis of an ontogenetic sequence Jour-nal of Vertebrate Paleontology 20501ndash506
Abdala F J Neveling and J Welman 2006 A new trirachodontidcynodont from the lower levels of the Burgersdorp Formation(Lower Triassic) of the Beaufort Group South Africa and the cla-distic relationships of Gondwanan gomphodonts Zoological Jour-nal of the Linnean Society 147383ndash413
Abdala F A M Ribeiro and C L Schultz 2001 A rich cynodont faunaof Santa Cruz do Sul Santa Maria Formation (Middle-Late Trias-sic) southern Brazil Neues Jahrbuch fuumlr Geologie und Palaumlontolo-gie Monatshefte 2001669ndash687
Abdala F and A M Ribeiro 2002 Nuevos cinodontes traversodoacuten-tidos (Synapsida-Eucynodontia) de la Formacioacuten Santa Maria(Triaacutesico Medio-Superior) Rio Grande do Sul Brasil RevistaEspantildeola de Paleontologiacutea 17237ndash243
Abdala F and A M Ribeiro 2003 A new traversodontid cynodontfrom the Santa Maria Formation (Ladinian-Carnian) of southernBrazil with a phylogenetic analysis of Gondwanan traversodontidsZoological Journal of the Linnaean Society 139529ndash545
Abdala F and A M S Teixeira 2004 A traversodontid cynodont ofAfrican affinity in the South American Triassic Palaeontologia Af-ricana 4011ndash22
Bonaparte J F 1962 Descripcioacuten del craacuteneo y mandiacutebula de Ex-aeretodon frenguellii Cabrera y su comparacioacuten con Diademodon-tidae Tritylodontidae y los cinodontes sudamericanos Publica-ciones del Museo Municipal de Ciencias Naturales y Tradicional deMar del Plata 1135ndash202
Bonaparte J F 1963 Descripcioacuten del esqueleto postcraneano de Ex-aeretodon (CynodontiandashTraversodontidae) Acta Geoloacutegica Lil-loana 45ndash52
Bonaparte J F 1966 Una nueva ldquofaunardquo Triaacutesica de Argentina (Ther-apsida Cynodontia Dicynodontia) Consideracions filogeneacuteticas ypaleobiogeograacuteficas Ameghiniana 8243ndash296
Bradu D and F E Grine 1979 Multivariate analysis of diademodon-tine crania from South Africa and Zambia South African Journal ofScience 79441ndash448
Brink A S 1955 A study of the skeleton of Diademodon Palaeonto-logia Africana 33-39
Brink A S 1963 Two new cynodonts from the Ntawere Formation inthe Luangwa Valley of Northern Rhodesia Palaeontologia Africana877ndash96
Broom R 1905 On the use of the term Anomodontia Records of theAlbany Museum 1266ndash269
Cabrera A 1943 El primer hallazgo de teraacutepsidos en la ArgentinaNotas del Museo de La Plata Paleontologiacutea 55317ndash331
JOURNAL OF VERTEBRATE PALEONTOLOGY VOL 28 NO 2 2008460
Dow
nloa
ded
by [
UZ
H H
aupt
bibl
ioth
ek
Zen
tral
bibl
ioth
ek Z
uumlric
h] a
t 05
51 1
0 Se
ptem
ber
2013
Chatterjee S 1982 A new cynodont reptile from the Triassic of IndiaJournal of Paleontology 56203ndash214
Crompton A W 1955 On some Triassic cynodonts from TanganyikaProceedings of the Zoological Society of London 125617ndash669
Crompton A W 1972 Postcanine occlusion in cynodonts and tritylo-donts Bulletin of the British Museum (Natural History) Geology2129ndash71
Crompton A W and F Ellenberger 1957 On a new cynodont from theMolteno Beds and the origin of the tritylodontids Annals of theSouth African Museum 441ndash13
Flynn J J J M Parrish B Rakotosaminimanana W F Simpson R LWhatley and A R Wyss 1999 A Triassic fauna from Madagascarincluding early dinosaurs Science 286763ndash765
Flynn J J J M Parrish B Rakotosamimanana L RanivoharimananaW F Simpson and A R Wyss 2000 New traversodontids (Syn-apsida Eucynodontia) from the Triassic of Madagascar Journal ofVertebrate Paleontology 20422ndash427
Gow C E and P J Hancox 1993 First complete skull of the LateTriassic Scalenodontoides (Reptilia Cynodontia) from southern Af-rica In Lucas S G and M Morales (eds) The nonmarine TriassicNew Mexico Museum of Natural History amp Science Bulletin 3161ndash168
Grine F E B D Hahn and C E Gow 1978 Aspects of relativegrowth and variability in Diademodon (Reptilia Therapsida) SouthAfrican Journal of Science 7450ndash58
Hopson J A 1984 Late Triassic traversodont cynodonts from NovaScotia and southern Africa Palaeontologia Africana 25181ndash201
Hopson J A 1985 Morphology and relationships of Gomphodontosu-chus brasiliensis von Huene (Synapsida Cynodontia Tritylodontoi-dea) from the Triassic of Brazil Neues Jahrbuch fuumlr Geologie undPalaumlontologie Monatshefte 1985285ndash299
Hopson J A and J W Kitching 2001 A probainognathian cynodontfrom South Africa and the phylogeny of nonmammalian cynodontsBulletin of the Museum of Comparative Zoology 1565ndash35
Hopson J A and H-D Sues 2006 A traversodont cynodont from theMiddle Triassic (Ladinian) of Baden-Wuumlrttemberg (Germany)Palaumlontologische Zeitschrift 80124ndash129
Huene F von 1928 Ein Cynodontier aus der Trias Brasiliens Zentral-blatt fuumlr Mineralogie Geologie und Palaumlontologie 1928251ndash270
Huene F von 1936 Die fossilen Reptilien des suumldamerikanischenGondwanalandes Ergebnisse der Sauriergrabungen in Suumldbrasilien192829 Lieferung 2 (Cynodontia)93ndash159 Franz F Heine Tuumlbin-gen
Jenkins F A 1970 The Chantildeares (Argentina) Triassic reptile faunaVII The postcranial skeleton of the traversodontid Massetognathuspascuali (Therapsida Cynodontia) Breviora 3521ndash28
Jenkins F A 1971 The postcranial skeleton of African cynodonts prob-lems in the early evolution of the mammalian postcranial skeletonBulletin of the Peabody Museum of Natural History 361ndash216
Kemp T S 1969 The atlas-axis complex of mammal-like reptiles Jour-nal of Zoology 159223ndash248
Kemp T S 1980 Aspects of the structure and functional anatomy of theMiddle Triassic cynodont Luangwa Journal of Zoology 191193ndash239
Kemp T S 1982 Mammal-Like Reptiles and the Origin of MammalsAcademic Press New York
Lehman J-P 1961 Cynodontia pp 140ndash191 in Piveteau J (ed) Traiteacutede Paleacuteontologie Masson et Cie Paris
Osborn 1903 On the primary division of the Reptilia into two sub-classes Synapsida and Diapsida Science 17275ndash276
Owen R 1861 Palaeontology or a systematic summary of extinct ani-mals and their geological relations 2nd Ed Edinburgh
Parrington F R 1934 On the cynodont genus Galesaurus with a noteon the functional significance of the changes in the evolution of thetheriodont skull Annals and Magazine of Natural History 10(13)38ndash67
Parrington F R 1946 On the cranial anatomy of cynodonts Proceed-ings of the Zoological Society of London 116181ndash197
Razafimbelo E 1987 Le basin de Morondava (Madagascar) Synthegravesegeacuteologique et structurale PhD dissertation University Louis Pas-teur Strasbourg France 189 pp
Reisz R R and H-D Sues 2000 Herbivory in Late Paleozoic andTriassic terrestrial vertebrates pp 9ndash41 in Sues H-D (ed) Evo-lution of Herbivory in Terrestrial Vertebrates Cambridge Univer-sity Press New York
Romer A S 1967 The Chantildeares (Argentina) Triassic reptile fauna IIITwo new gomphodonts Massetognathus pascuali and M terugiiBreviora 2641ndash25
Romer A S 1972 The Chantildeares (Argentina) Triassic reptile faunaXVII The Chantildeares gomphodonts Breviora 3961ndash9
Rougier G W J R Wible and J A Hopson 1992 Reconstruction ofthe cranial vessels in the Early Cretaceous mammal Vincelestes neu-quenianus implications for the evolution of the mammalian cranialvascular system Journal of Vertebrate Paleontology 12188ndash216
Rowe T 1988 Definition diagnosis and origin of Mammalia Journal ofVertebrate Paleontology 8241ndash264
Rowe T 1993 Early mammal phylogenetic systematics pp 129ndash145 inSzalay F S M J Novacek and M C McKenna (eds) MammalPhylogeny Vol 1 Mesozoic Differentiation MultituberculatesMonotremes Early Therians and Marsupials Springer-Verlag NewYork
Seeley H G 1894 Researches on the structure organization and clas-sification of the fossil Reptilia Part IX Section 3 On DiademodonPhilosophical Transactions of the Royal Society of London 1851029ndash1041
Seeley H G 1895 Researches on the structure organization and clas-sification of the fossil Reptilia Part IX Section 4 On the Gompho-dontia Philosophical Transactions of the Royal Society of London1861ndash57
Seeley H G 1895 Researches on the structure organization and clas-sification of the fossil Reptilia Part IX Section 5 On the skeletonin new Cynodontia from the Karroo rocks Philosophical Transac-tions of the Royal Society of London 18659ndash148
Sues H-D J A Hopson and N H Shubin 1992 Affinities of Sca-lenodontoides plemmyridon Hopson 1984 (Synapsida Cynodontia)from the Upper Triassic of Nova Scotia Journal of Vertebrate Pa-leontology 12168ndash171
Sues H-D and P E Olsen 1990 Triassic vertebrates of Gondwananaspect from the Richmond basin of Virginia Science 2491020ndash1023
Swofford D L 2003 PAUP v 40b10 Sinauer Associates Inc Pub-lishers Sunderland Massachusetts
Watson D M S and A S Romer 1956 A classification of therapsidreptiles Bulletin of the Museum of Comparative Zoology 11435ndash89
Submitted September 20 2007 accepted November 3 2007
KAMMERER ET ALmdashCYNODONT FROM MADAGASCAR 461
Dow
nloa
ded
by [
UZ
H H
aupt
bibl
ioth
ek
Zen
tral
bibl
ioth
ek Z
uumlric
h] a
t 05
51 1
0 Se
ptem
ber
2013
APPENDIX Data matrix used in the phylogenetic analysesThis is a revised version of the matrix of Abdala et al (2006) with the following changes character 43 [Incisor procumbency (0) absent (1) present]added Menadon recoded for characters 0 (Number of upper incisors was 1 ldquothreerdquo changed to 0 ldquofourrdquo) 5 (Position of paracanine fossae in relationto the upper canine was changed to 1 ldquomedialrdquo) 16 (Overall morphology of lower postcanines in occlusal view was changed to 3 ldquoquadran-gularrdquo) 17 (Position of the transverse crest in lower postcanines was changed to 1 ldquoanteriorrdquo) 18 (Number of cusps in the transverse crest of thelower postcanines was changed to 0 ldquotwordquo) 20 (Anterior cingulum in the lower postcanines was changed to 2 ldquoabsentrdquo) 21 (Internarial barwas 1 ldquoabsentrdquo changed to 0 ldquopresentrdquo) 38 (Posterior cingulum in the lower postcanines was changed to 1 ldquoabsentrdquo) and 40 (Deep occlusalbasins in the postcanines was changed to 1 ldquopresentrdquo) Langbergia recoded for character 34 (Incisor cutting margins was 0 ldquoserratedrdquo changedto 1 ldquosmoothly ridgedrdquo) based on the description of the incisors in Abdala et al (2006) Gomphodontosuchus recoded for character 31 (Maxilla inthe margin of the subtemporal fenestra was 1 ldquoincludedrdquo changed to 0 ldquoexcludedrdquo) based on von Huene (1936) and Kammerer (pers obs of theholotype) contra Hopsonrsquos (1985) reconstruction lsquoScalenodonrsquo hirschsoni recoded for character 39 (Anterior cingulum in the upper postcanines was1 ldquoabsentrdquo changed to 0 ldquopresentrdquo) based on Abdala (pers comm 2006) Trirachodon recoded for character 30 (Ectopterygoid was changedto 0 ldquopresentrdquo) based on Abdala (pers comm 2006) and Traversodon recoded for character 15 (Number of cusps in the sectorial border of the upperpostcanines was 0 ldquomore than twordquo changed to 1 ldquotwordquo) based on Abdala (pers comm 2006) and Kammerer (pers obs)
1111111111 2222222222 3333333333 44440123456789 0123456789 0123456789 0123456789 0123
Cynognathus 00000000 000 000000000 00000000 000Diademodon 0000001000 0000100 000010A000 0000000100 0100Trirachodon 000000100B 1A002201 001A100010 0001001200 0110Langbergia 0000001000 10000201 0000101010 0001101200 0110Cricodon 0100001000 11000201 0010100 0001200 0110Andescynodon 0000002001 01013101 101120010 0112311 1100Massetognathus 00111121A2 1110003100 2011200011 1011212300 1100Exaeretodon 1110122112 010103101 2111110201 1111112311 1101Luangwa 0000001002 11010A3100 101010010 01002300 1100Scal angustifrons 000000100B 1111023100 10102101 00002301 1100Scal hirschsoni 11110202 1100113100 1010 1112300 1100Traversodon 0000002012 1101113100 201010 011230 1100Gomphodontosuchus 0011112112 0103101 2010 0112311 1100Pascualgnathus 1000001000 000231 1100001 0011230 1100Scalenodontoides 1112212 01003101 21111021 1212311 1101Menadon 0111112112 01012310 20121020 0102311 1101Dadadon 00002112 111112 013 11231 1100Santacruzodon 00002112 1110003101 201300 212300 1100
A(01) B(02)
JOURNAL OF VERTEBRATE PALEONTOLOGY VOL 28 NO 2 2008462
Dow
nloa
ded
by [
UZ
H H
aupt
bibl
ioth
ek
Zen
tral
bibl
ioth
ek Z
uumlric
h] a
t 05
51 1
0 Se
ptem
ber
2013

found in close proximity to 9-4-98-440 and are considered topertain to the same individual This decision is based on the factthat 9-2-98-422 was recovered on the same erosional surface asand very near 9-4-98-440 is in the same size range as the mate-rial from 9-4-98-440 and there is no overlap in elements betweenthe two field-numbered specimens The isolated vertebra from9-2-98-422 in particular is identical in size preservational stateand morphology to those present in 9-4-98-440 An isolated frag-mentary right pelvis (field number 8-31-98-387) herein referredto Menadon was collected at a different locality (M-18) than theprevious specimen Specimens were measured using Fowlerregdigital calipers to an accuracy of 001 mm Phylogenetic analyseswere run using a modified version of the data matrix of Abdalaet al (2006) in the parsimony program PAUP v 40b10 (Swof-ford 2003)
Comparisons with other cynodonts were made on the basis offirst hand examination of the following specimens
Menadon besairiei UA 10601 FMNH PR 2104 Dadadon isa-loi UA 10605 10606 Luangwa drysdalli BP13731 TSK 121Scalenodon angustifrons UMZC T907 T914 T918 Traversodonstahleckeri GPITRE7170 Scalenodontoides macrodontesMNHN 1957-23 NMQR 3053 Exaeretodon frenguellii MCZ4469 4470 4472 4483 4486 4493 4505 Gomphodontosuchusbrasiliensis GPIT unnumbered Massetognathus pascuali MCZ3782 3784 3786 3789 3790 3791 3792 3795 3796 3800 38013801-188 3803 3806 3807 3810 3999 4003 4009 4012 4018-164 4018-167 4047 4048 4049 4050 4138 4207 4208 42134214 4217 4218 4219 4221 4222 4230 4246 4253 4258 42654420 Diademodon tetragonus UMZC T492 Cricodon metabo-lus UMZC T905
Other comparisons were based on specimen photographs anddescriptions from the literature (Brink 1963 Bonaparte 19631966 Jenkins 1970 1971 Kemp 1980 Hopson 1985 Flynn etal 2000 Abdala et al 2002 Abdala and Teixeira 2004)
SYSTEMATIC PALEONTOLOGY
SYNAPSIDA Osborn 1903THERAPSIDA Broom 1905CYNODONTIA Owen 1861
EUCYNODONTIA Kemp 1982GOMPHODONTIA Seeley 1894 (sensu Hopson and
Kitching 2001)TRAVERSODONTIDAE von Huene 1936
DefinitionmdashHere we consider Traversodontidae to representthe most inclusive clade containing Traversodon stahleckeri vonHuene 1936132 but not Trirachodon kannemeyeri Seeley 189548 or Diademodon tetragonus Seeley 18941030
MENADON BESAIRIEI Flynn Parrish RakotosamimananaRanivoharimanana Simpson and Wyss 2000
HolotypemdashUA 10601 skull and mandible with eroded leftside
Referred SpecimensmdashFMNH PR 2444 a partial skull andpostcranium field number 8-31-98-387 an isolated right pelvisFMNH PR 2104 an isolated mandible
Horizon and LocalitymdashBasal lsquoIsalo IIrsquo of Besairie equated tothe Makay Formation by Razafimbelo (1987) drainage of theMalio River Morondava Basin Madagascar
AgemdashLate Middle (Ladinian) or early Late (Carnian) Triassic(see Flynn et al 1999 2000)
Revised DiagnosismdashTraversodontid with the followingunique combination of features four large upper incisors firstand second upper incisors and all lower incisors procumbentthird and fourth upper incisors caniniform with fourth upperincisor strongly recurved serrated upper canines small equal in
dorsoventral length to incisors distinctly canted forwards (auta-pomorphy) canine alveolus proportionally narrower than incisoralveoli with a much greater anteroposterior than labiolinguallength (autapomorphy) no diastema between upper incisors andcanine low number of postcanines (8 uppers and 6ndash7 lowers)upper and lower postcanines quadrangular roughly trapezoidalin outline descending process of jugal posterior to the last upperpostcanine axial spine with concave dorsal profile and elongateposterior process overhanging shortened neural spine of subse-quent cervical vertebra expanded ribs present caudal neuralspines very tall anterior edge of the iliac blade sloping upwardsat a sim45deg angle acetabular buttresses of the three pelvic boneslargely confluent posterior process of the iliac blade short anddirected away from the dorsal edge of the ischium ischium andpubis extremely constricted resulting in a large obturator fora-men
CommentsmdashMenadon can be distinguished from all other tra-versodontids by the combination of four large upper incisors andincisor procumbency The presence of four upper incisors isprimitive for traversodontids (Hopson and Kitching 2001) En-largement of the incisors is an advanced feature present in suchtaxa as Exaeretodon Scalenodontoides and lsquoScalenodonrsquo hirsch-soni (Abdala and Ribeiro 2003 Abdala et al 2006) Incisorprocumbency is otherwise only known in Exaeretodon and Sca-lenodontoides among traversodontids but these taxa have onlythree upper incisors (Bonaparte 1962 Hopson 1984) Addition-ally Menadon has a descending process of the jugal that contactsthe pterygoids and excludes the maxilla from the margin of thesubtemporal fenestra which is present in most gomphodonts butnot in Exaeretodon or Scalenodontoides (Abdala et al 2006)
Because the postcranium is known in few traversodontid taxathe combination of pelvic characters listed in the Diagnosis canonly be said to diagnose Menadon in comparison with LuangwaPascualgnathus Massetognathus and Exaeretodon The postcra-nium of Menadon resembles that of Luangwa in the shape of theiliac blade the large obturator foramen and the heterogeneousrib morphology but resembles Exaeretodon in the tall caudalneural spines confluent acetabular buttresses and short poste-rior process of the iliac blade projecting away from the ischium
DESCRIPTION
Skull
The cranium of FMNH PR 2444 is poorly preserved and nosutures are visible Six portions of the skull are represented theentire rostrum and left orbitalzygomatic region (Figs 1ndash3) partsof both zygomatic arches (Fig 4AndashC) a fragment of the tempo-ral region including the sagittal crest (Fig 4D) and the occiputand basicranium (Figs 4E 5) The following description is basedon the left side of the skull The right side of the rostrum hasbeen sheared moving some cranial landmarks (eg naris al-veoli) out of their original positions The frontal region on bothsides of the skull has also been sheared anterolaterally towardsthe left such that a shelf of bone made up of the nasals leftprefrontal and left lacrimal overhangs the snout (see Figs 12A)
The rostrum is tall and robust as in Gomphodontosuchus(Hopson 1985) and juveniles of Exaeretodon (Abdala et al2002) The external naris is large and teardrop-shaped narrow-ing towards the rear (Fig 1) A well-developed internarial bar ispresent The maxilla curves inward posterior to the root of thecanine so that the postcanine tooth row is medially inset locatedon an axis posterior to the paracanine fossa (Fig 2B) A singlelateral maxillary foramen is present The orbit is large and sub-circular Prominent interorbital depressions are present on theskull roof (Fig 2A)
Other than a fragment of the left fourth incisor the skull lacks
JOURNAL OF VERTEBRATE PALEONTOLOGY VOL 28 NO 2 2008446
Dow
nloa
ded
by [
UZ
H H
aupt
bibl
ioth
ek
Zen
tral
bibl
ioth
ek Z
uumlric
h] a
t 05
51 1
0 Se
ptem
ber
2013
teeth entirely Alveoli are preserved for all tooth loci except theinferred anterior postcanines (five postcanine alveoli are pre-served on the left side) The paracanine fossa is ovoid locatedmedial to the canine alveolus and much larger than all toothalveoli (Fig 3) Paired incisive foramina are located medial tothe paracanine fossae and extend from the anterior edge of thefourth incisor to the posterior edge of the canine The postca-nines are few in number (minimally five in this individual butprobably eight see Discussion) compared to many traversodon-tids (Romer 1967 Crompton 1972 Abdala et al 2002) andtheir alveoli are large and roughly trapezoidal in outline Thisconfiguration is similar to that observed in Gomphodontosuchus(Hopson 1985)
The left zygomatic arch is composed almost entirely of thejugal and as in most traversodontids it is relatively deep (Flynnet al 2000 Hopson and Kitching 2001) The posterior portionwhere the jugal would have articulated with the squamosal isbroken The isolated right jugal is more complete and its pos-terior articulation with the squamosal is intact (Fig 4A B) Thedorsal edge of the jugal is broken and is missing the posteriorextension over the squamosal The squamosals are very delicatebut well preserved bones each consisting of a cranial and zygo-matic ramus (Fig 4AndashC) The cranial ramus is an extremely thinwedge of bone directed dorsally that would have flanked therear of the braincase in the complete skull The zygomatic ramusis subdivided into a process extending anteriorly and one extend-ing ventrally with a pair of facets between them where the squa-mosal receives the posterior end of the jugal (Fig 4A C) Theanterior process is thin much longer than the ventral processand tapers forward The ventral process is short blunt and wideand forms the lateral portion of the external auditory meatus
The occiput and foramen magnum are tall and narrow (Fig4E) The occipital condyles are separated by a prominent groove
and are also taller than they are wide (but see Discussion) Abroken fragment of the proatlas is preserved out-of-place abovethe foramen magnum (Fig 4E) The occipital fragment includesthe posterior part of the basicranium The basicranium (Fig 5) issimilar to that described for Massetognathus (Rougier et al1992) and Exaeretodon (Abdala et al 2002) However FMNHPR 2444 differs from these in the possession of a basipterygoidforamen Such a foramen has been described by Parrington(1946) in the early cynodont Thrinaxodon but is not known intraversodontids This foramen is not evident in the holotype ofMenadon and this character may be variable in cynodonts
Axial Skeleton
A total of 21 vertebrae are preserved in FMNH PR 2444 Theangle of zygapophyseal articulation varies along the length of thevertebral column This angle (from the sagittal plane) is sim45deg forthe axisC3 (cervical 3) articulation sim30deg for the posterior cervi-cals lt15deg for the dorsals and sim20-30deg for the caudals Other thanthe atlas all vertebral centra are amphicoelous
Atlas-Axis ComplexmdashThree intercentra are articulated withthe cervical series IC2 (fused to the atlas-axis complex) IC3(between C2 and C3) and IC4 (between C3 and C4) IC2 makesup the lower part of the anterior face of the atlas-axis complexand extends back to the ventral anterior edge of the axial cen-trum (Fig 6) The free intercentra (IC3 and IC4) are singleirregularly shaped ossicles situated at the ventral margins of theintercervical articulations (Fig 6) The intercentra of FMNH PR2444 differ markedly from those described for Thrinaxodon(Jenkins 1971) but in general shape and position resemble thosefigured by Brink (1955) for Diademodon The intercentra ofThrinaxodon are transversely wide and anteroposteriorly short(Jenkins 1971fig 2) whereas the intercentra of Menadon be-
FIGURE 1 Stereopair and interpretive drawing of main cranial fragment of FMNH PR 2444 in left lateral view Scale bar equals 5 cm Abbre-viationsmdashJU jugal LA lacrimal MA maxilla mfo lateral maxillary foramen NA nasal nar naris orb orbit PF prefrontal PM premaxilla POpostorbital SM septomaxilla Sutures are not visible in this specimen and have been reconstructed based on the holotype of Menadon besairiei
KAMMERER ET ALmdashCYNODONT FROM MADAGASCAR 447
Dow
nloa
ded
by [
UZ
H H
aupt
bibl
ioth
ek
Zen
tral
bibl
ioth
ek Z
uumlric
h] a
t 05
51 1
0 Se
ptem
ber
2013
FIGURE 2 Stereopairs of main cranial fragment of FMNH PR 2444 in A dorsal view and B palatal view Scale bars equal 5 cm
JOURNAL OF VERTEBRATE PALEONTOLOGY VOL 28 NO 2 2008448
Dow
nloa
ded
by [
UZ
H H
aupt
bibl
ioth
ek
Zen
tral
bibl
ioth
ek Z
uumlric
h] a
t 05
51 1
0 Se
ptem
ber
2013
sairiei have equal transverse and anteroposterior dimensions Attheir maximum depth in Thrinaxodon the intercentra barely ex-tend beyond the ventral margins of the cervical pleurocentra InFMNH PR 2444 IC4 extends for half of its total depth below theventral margins of the cervical pleurocentra In IC3 this condi-tion is even more extreme extending below the ventral marginsof the cervical pleurocentra for 80 of its total depth The shapeand position of the intercentra give a strongly arched appearanceto the ventral profile of elements in the cervical series The ven-tral surfaces of the intercentra bear a posteriorly directed process
on each side In IC3 a weak ridge originates between the pairedprocesses and extends posteriorly indicating muscle attachmenton the intercentra The condition in IC4 cannot be determinedbecause of incomplete preservation Although only three inter-centra are known ventral facets in the centra of the isolatedcervicals indicate that others were present
As in other cynodonts the atlas-axis complex of FMNH PR2444 consists of a pair of separate atlantal neural arches articu-lating with the occipital condyles and the atlas centrum Only theleft atlas arch is preserved in this specimen (Fig 7) It is similar
FIGURE 3 Stereopair and interpretive drawing of anterior palatal region of FMNH PR 2444 Scale bar equals 5 cm AbbreviationsmdashC canineI incisor i f incisive foramina PC postcanine pc f paracanine fossa Postcanine numbering based on comparison with the holotype UA 10601
FIGURE 4 Cranial fragments of FMNH PR 2444 A stereopair of right zygomatic arch in lateral view Scale bar equals 5 cm B stereopair of rightzygomatic arch in medial view Scale bar equals 5 cm C stereopair of left squamosal in lateral view Scale bar equals 5 cm D sagittal crest in rightlateral view Scale bar equals 1 cm E occiput in occipital view Scale bar equals 1 cm
KAMMERER ET ALmdashCYNODONT FROM MADAGASCAR 449
Dow
nloa
ded
by [
UZ
H H
aupt
bibl
ioth
ek
Zen
tral
bibl
ioth
ek Z
uumlric
h] a
t 05
51 1
0 Se
ptem
ber
2013
to the atlantal neural arches described for other cynodonts suchas Massetognathus (Kemp 1969 Jenkins 1971) There is a me-dial facet for articulation with the centrum of the atlas (theldquoodontoid facetrdquo of Kemp 1969) and an anterior facet for ar-ticulation with the occipital condyle (Fig 7B) A pair of pro-cesses dominates the atlas arch a dorsal lamina (a roughly rect-angular plate of bone arching above the facets) and a largeposterolaterally directed transverse process An oval tubercle atthe anterior end of the transverse process represents the facet forthe proatlas
The centrum of the atlas is fused to the axis forming a singlelarge vertebral unit (Fig 6) A weak odontoid process is presentat the dorsal limit of the anterior surface of the atlas The ante-rior aspect of the centrum is dominated by a roughly lsquoTrsquo-shapedstructure composed of the two facets for articulation with theatlas arches and the facet for articulation with the atlas intercen-trum The axis bears no prezygapophyses and as such does notarticulate with the atlas arch as in Early Triassic cynodonts suchas Galesaurus (Parrington 1934) There appears to be a trendtowards loss of the axial prezygapophyses in later traversodon-
tids which are present but vestigial in Massetognathus (Jenkins1970 although in the specimens described the prezygapophysesare broken so their reduction is perhaps questionable) and ab-sent in Exaeretodon (Bonaparte 1963) The transverse processesof the axis are large and robust compared to those of the othercervicals The axial spine is saddle-shaped with a concave dorsalprofile unlike previous reconstructions of the traversodontidaxis which depict a straight or convex dorsal profile similar tothat observed in galesaurid cynodonts and early mammals (Jen-kins 1971) The morphology of the axial spine is similar to thatof the Early Triassic cynodont Cynognathus (Seeley 1895) InCynognathus however the neural spine of the third cervical(C3) is tall and canted posteriorly behind the posterior process ofthe axial spine whereas in FMNH PR 2444 the short C3 neuralspine fits below the posterior portion of the axial spine Theposterior process of the axial spine bears a pair of parallel elon-gate grooves on its underside attachment sites for the axial mus-culature
Post-Axial VertebraemdashThree post-axial cervicals are knownfor FMNH PR 2444 (Figs 6 8AndashD) The first of these C3 is
FIGURE 5 Stereopair of ventral view of the occipital fragment of FMNH PR 2444 showing the basicranium Anterior is up
FIGURE 6 Stereopair and interpretive drawing of atlas-axis complex and third cervical of FMNH PR 2444 in left lateral view Scale bar equals5 cm Abbreviationsmdasha n sp axial neural spine d dens IC intercentrum f at a facet for articulation with the atlas arch p parapophysis pozpostzygapophysis t p transverse process
JOURNAL OF VERTEBRATE PALEONTOLOGY VOL 28 NO 2 2008450
Dow
nloa
ded
by [
UZ
H H
aupt
bibl
ioth
ek
Zen
tral
bibl
ioth
ek Z
uumlric
h] a
t 05
51 1
0 Se
ptem
ber
2013
preserved in articulation with the axis (Fig 6) The two isolatedcervicals bear successively taller and anterposteriorly wider neu-ral spines (Fig 8AndashD) Based on their staggered size and failureto articulate we suggest that they represent C5 and C7 (assumingthat this specimen possessed the generalized cynodont cervicalnumber of seven) The cervicals display robust transverse pro-cesses which are directed laterally and slightly anteriorly A par-apophysis lies below the anterior edge of the transverse processalong the antero-lateral edge of the centrum No cervical ribs arepreserved but their presence can be inferred from the par-apophyses The notochordal concavity is as in Massetognathussurrounded by a raised rim at each end of the centrum (Jenkins1970) No anapophyses are present The neural spines for allthree post-axial cervicals are tapering and slightly curved poste-riorly The neural spine in C3 is relatively short and wide(thereby fitting underneath the posterior overhang of the axialspine see Fig 6) while that of C5 is tall and narrow The spinefor C7 is taller and anteroposteriorly wider than that of C5 andthe dorsal edge is more rounded similar to the condition in thedorsal vertebrae
Eight post-cervical presacral vertebrae are known for thisspecimen We will refer to them here simply as dorsal vertebraegiven the incompleteness of the vertebral column in the speci-men and the uncertainty concerning the thoraciclumbar transi-tion in this taxon However it is probable that they representthoracic vertebrae (see below) In most cynodonts the lumbar
vertebrae are synostosed to specialized ribs with overlappingelements In earlier taxa (eg Thrinaxodon Cynognathus Lu-angwa Jenkins 1971 Kemp 1980) expanded costal plates arepresent on both thoracic and lumbar ribs In Massetognathus thelumbar ribs consist of a pair of prongs one directed anteriorlybearing a facet for articulation with the previous rib and theother directed posteriorly (Jenkins 1970) In Exaeretodon allpresacral ribs are identical of simple morphology and not syn-ostosed to the vertebrae (Bonaparte 1963) A single isolated rib(Fig 9B) in FMNH PR 2444 demonstrates that some of the ribsin Menadon besairiei were expanded However all preservedvertebrae in this specimen have facets for articulation with theribs rather than breakage points where they would have beenfused to ribs As such it is probable that all of the vertebraepreserved in this specimen represent thoracics although we can-not discount the possibility that as in Exaeretodon no presacralvertebrae were fused to ribs
The neural spines in all of the dorsal vertebrae are elongateposteriorly inclined and roughly rectangular in lateral view witha rounded laterally expanded platform dorsally (Fig 8EndashJ)Costal foveae are present at the anterior and posterior edges ofeach centrum Of the eight dorsal vertebrae there are two ar-ticulated sets and three isolated elements (two from 9-4-98-440and one from 9-2-98-422) The first isolated vertebra (Fig 8E F)has a taller neural spine and shorter centrum than the otherdorsal vertebrae It is most similar in proportions to the isolated
FIGURE 7 Stereopairs and interpretive drawings of left atlas arch of FMNH PR 2444 in A lateral view Anterior is left B medial view Anterioris right Scale bars equal 1 cm Abbreviationsmdashafa anterior facet for articulation with the occipital condyle dl dorsal lamina mfa medial facet forarticulation with the atlas centrum (odontoid facet) pfa tubercle forming the facet for articulation with the proatlas tp transverse process
KAMMERER ET ALmdashCYNODONT FROM MADAGASCAR 451
Dow
nloa
ded
by [
UZ
H H
aupt
bibl
ioth
ek
Zen
tral
bibl
ioth
ek Z
uumlric
h] a
t 05
51 1
0 Se
ptem
ber
2013
FIGURE 8 Vertebrae of FMNH PR 2444 Cervical vertebra 5 in A left lateral and B anterior views Cervical vertebra 7 in C left lateral andD anterior views Isolated dorsal vertebra from anterior region of the series in E left lateral and F anterior views G stereopair and interpretivedrawing of articulated set of two dorsal vertebrae from near the midpoint of the series in left lateral view Anterior is up H articulated set of threedorsal vertebrae from anterior region of the series in left and right lateral views I isolated vertebra from near the midpoint of the series (from fieldnumber 9-2-98-422) J isolated vertebra from posterior region of the series Sacral vertebra in K dorsal view (the distal region of another sacral ribwhich is identical in shape to that of the intact sacral rib albeit slightly larger is also shown in this figure) and L left lateral view Caudal vertebraeM articulated set of two vertebrae from the anterior region of the series showing the lsquowingrsquo-shaped caudal ribs in dorsal view Anterior is left Nisolated caudal vertebra showing tall neural spine in left lateral view O isolated caudal vertebra showing tall neural spine in posterior view Particulated set of four vertebrae from near the end of the caudal series in dorsal view Anterior is left Scale bars equal 1 cm Abbreviationsmdasha cof anterior costal fovea n sp neural spine p co f posterior costal fovea poz postzygapophysis prz prezygapophysis
JOURNAL OF VERTEBRATE PALEONTOLOGY VOL 28 NO 2 2008452
Dow
nloa
ded
by [
UZ
H H
aupt
bibl
ioth
ek
Zen
tral
bibl
ioth
ek Z
uumlric
h] a
t 05
51 1
0 Se
ptem
ber
2013
cervicals and was probably located near the front of the dorsalseries The first articulated set consists of three vertebrae (Fig8H) The middle vertebra in this set is broken from the anteriorbase of the neural spine to the ventral midpoint and most of theright side of the anterior half of the centrum is missing This setof vertebrae probably also came from the anterior region of theseries as their neural spines are longer and narrower than allvertebrae other than the one described above The second ar-ticulated set (Fig 8G) consists of two well-preserved vertebraethat based on comparisons with Massetognathus and Ex-aeretodon (Jenkins 1970 Bonaparte 1963) probably came fromnear the midpoint of the dorsal series The second isolated ver-tebra (Fig 8I) from 9-2-98-422 is similar to the members of thesecond set The third isolated vertebra (Fig 8J) appears to befrom the posterior end of the series based on the relatively shortneural spine and reduced posterior costal fovea
One sacral vertebra (Fig 8K L) is known consisting of awell-preserved centrum neural spine and the right sacral rib(the left is broken off) The centrum is shorter (both anteropos-teriorly and dorsoventrally) than those of the dorsal vertebraeand the neural spine is shorter and more rounded in profileAlso the neural spine is directed vertically unlike those of thedorsal vertebrae which are canted posteriorly The zygapophy-ses are poorly developed The sacral rib is fused to a singlesynapophysis and projects anterolaterally The rib narrows to-wards the middle and expands distally terminating in a widerectangular facet where it would meet the ilium (Fig 8K)
Seven caudal vertebrae are known The caudal vertebrae aredistinctive in having very tall narrow neural spines Some EarlyTriassic cynodonts possess blunt neural spines of moderateheight on the proximal caudals (see for example Thrinaxodonin Jenkins 1971 and Diademodon in Brink 1955) Massetogna-thus represents an extreme condition wherein the caudal neuralspines are greatly reduced (Jenkins 1970) Very tall neuralspines are seen in Exaeretodon (Bonaparte 1963) and thosespines are unusual in that they do not diminish in height until thelast caudals The caudal vertebrae of FMNH PR 2444 consist ofa pair of articulated vertebrae most likely from near the sacrum(Fig 8M) an isolated caudal lacking postzygapophyses and ribsbut with the neural spine (Fig 8N O) and a quartet of smaller
articulated elements (Fig 8P) The anterior caudals have nar-rower neural spines and shorter zygapophyses than the subse-quent vertebrae in the series The caudal ribs attached to thispair have a crescentic distal region that curves anteriorly pro-ducing a lsquowingrsquo-shape Only a portion of the right rib of theisolated caudal is preserved but the lsquowingrsquo-shaped morphologyof the near-sacral caudal ribs does not appear to have been pre-sent In the posteriormost four vertebrae caudal ribs are pre-served on the left side of the first and the right side of the secondand third members On the first of these vertebrae the rib isshort curved posteriorly (but not lsquowingrsquo-shaped) and taperingRibs of the second and third vertebrae are less curved resem-bling posteriorly directed rods The zygapophyses of both theisolated caudal vertebra and the quartet of small caudals areanteroposteriorly elongate becoming increasingly so posteriorly
Thoracic RibsmdashNine thoracic ribs are known from 9-4-98-440(four left four right and an indeterminate splint of bone Fig9A indeterminate bone not illustrated) These elements re-semble the flattened ribs lacking costal plates of Exaeretodon(Bonaparte 1963) and the anterior thoracic ribs of Massetogna-thus and Luangwa (Jenkins 1970 Kemp 1980) The tuberculumand capitulum are separated by a shallow depression The tuber-culum has a shorter base than the capitulum but their proximalfaces are subequal in size An additional isolated left rib (Fig9B) from 9-2-98-422 is expanded indicating that unlike Ex-aeretodon Menadon rib morphology was heterogeneous alongthe length of the dorsal series This rib closely resembles theposterior thoracic ribs described for Luangwa (Kemp 1980fig16) Pascualgnathus (Bonaparte 1966fig 7) and the trirach-odontid Cricodon (Crompton 1955 Fig 4) This rib is con-stricted at the base of the head then expands distally with an-terior and posterior costal plates As in Luangwa the anteriorcostal plate is shallow and evenly sloping proximally and distallywhile the posterior costal plate is falcate with a concave edgedistally The distal expansion of this rib occurs at an angle of sim30degto the long axis of the rib head Despite their overall similaritythe expanded rib in FMNH PR 2444 does exhibit some differ-ences from the posterior thoracic rib of Luangwa The expandedrib of Menadon has a longer shaft between the rib head and thecostal plates and a shorter capitulum lacks a ridge on the pos-
FIGURE 9 A anterior thoracic ribs of FMNH PR 2444 in anterior view (indeterminate rib not figured) B expanded posterior thoracic rib ofFMNH PR 2444 in anterior view Scale bars equal 1 cm
KAMMERER ET ALmdashCYNODONT FROM MADAGASCAR 453
Dow
nloa
ded
by [
UZ
H H
aupt
bibl
ioth
ek
Zen
tral
bibl
ioth
ek Z
uumlric
h] a
t 05
51 1
0 Se
ptem
ber
2013
terior costal plate and has an anteroposteriorly shorter anteriorcostal plate Additionally the posterior margin of the costal plateis less curved than that of Luangwa These differences couldindicate that the expanded rib of Menadon is from a more pos-terior position in the series than the rib from Luangwa as re-duced curvature and increasing confluence of the tuberculumand capitulum are characteristic of lumbar ribs in traversodon-tids (Jenkins 1970 Kemp 1980) However the length of theshaft is at odds with this interpretation and in Cricodon andPascualgnathus the more posterior ribs are not more similar tothe element from FMNH PR 2444 These differences probablyrepresent legitimate morphological distinctions between the ribsof earlier gomphodonts and Menadon The head of the expandedrib is well defined and unbroken indicating that it was not syn-ostosed to a vertebra
Appendicular Skeleton
The appendicular skeleton is represented by both scapulocora-coids and a fragment of a clavicle from field number 9-4-98-440and a partial right pelvis from field number 9-2-98-422 An ad-ditional partial right pelvis is represented from field number8-31-98-387 The scapulocoracoids (Figs 10 11) are very similarto those described for other traversodontids as well as cynog-nathids and diademodontids (Jenkins 1970 1971) The scapularinfraspinous fossa has out-turned anterior and posterior edgesThe anterior edge of the scapula is rounded and curves laterallyforming the scapular spine At the lower end of the spine theacromion forms a prominent process (broken in the left scapula)Below this process the scapula is constricted and then expandsventrally to form the dorsal margin of the articular glenoid Theglenoid fossa is in general morphology very similar to that ofother Triassic cynodonts (Jenkins 1970 1971 Kemp 1980) It isposteroventrally oriented with roughly equal contributions fromthe scapular and coracoid glenoid facets The coracoid glenoidfacet is roughly triangular while the scapular glenoid facet issemiovoid as in other cynodonts In anterior or posterior viewthe fossa appears smoothly crescentic The procoracoid is aslightly curved trapezoidal plate of bone It does not appear tocontribute to the glenoid fossa The procoracoid foramen is lo-cated midway up the height of the procoracoid at a point directlybelow the base of the acromion process The coracoid is semi-crescentic curving posteriorly from a broad base The coracoidin Menadon is longer and more falcate than those described forExaeretodon (Bonaparte 1963) and Luangwa (Kemp 1980 al-though the distal tip of both coracoids is broken in the specimenof Luangwa [TSK 121] described by Kemp and may have beenlonger than reconstructed) but agrees closely with those of Mas-setognathus and cynognathidsdiademodontids (Jenkins 19701971) The distal coracoid tubercle for attachment to the tricepsis strongly curved upwards similar to the condition in Masseto-gnathus (Jenkins 1970) and Diademodon (Brink 1955)
A small fragment of bone resembling a curved cylinder almostcertainly represents part of a clavicle It has no distinguishingfeatures other than a groove associated with a ridge on one sideHowever the degree of curvature observed precludes this ele-ment from representing a long bone and the circular cross-section excludes it from being a rib
The pelvis of FMNH PR 2444 (Fig 12) resembles those ofother traversodontids in general morphology The ilium bears anarrow anteriorly directed iliac blade curving outward distallyThe anterior edge of the iliac blade slopes upward at a roughly45deg angle similar to Luangwa (Kemp 1980) but unlike Ex-aeretodon in which the anterior edge of the iliac blade curvesbackwards (Bonaparte 1963) or Pascualgnathus and Masseto-gnathus in which it is rounded (Bonaparte 1966 Jenkins 1970)The supraacetabular buttress at the base of the ilium is located inan anterodorsal position relative to the acetabulum as in other
traversodontids but unlike earlier cynodonts in which it is lo-cated dorsally (Jenkins 1970) The acetabular buttresses of thethree pelvic bones are largely confluent forming a fairly uniformlsquoliprsquo around the acetabulum as in Exaeretodon (Bonaparte1963) On the contrary in Luangwa the supraacetabular ischialand pubic buttresses are highly distinct spaced 6ndash8 mm apart andwithout confluence of their acetabular facets The pelvis ofFMNH PR 2444 does have a small depression between the su-praacetabular and ischial buttresses (also present in Ex-aeretodon Bonaparte 1963) but this is not as developed as inLuangwa The posterior extent of the iliac blade is broken inFMNH PR 2444 but this region is preserved in another isolatedright pelvis field number 8-31-98-387 (Fig 13) probably pertain-ing to Menadon besairiei The posterior process of the iliac bladein this specimen is relatively short and directed away from thedorsal edge of the ischium as in Exaeretodon (Bonaparte 1963)but unlike Massetognathus and Luangwa in which the posteriorprocess is long and extends roughly horizontally over the dorsaledge of the ischium (Jenkins 1970 Kemp 1980) and evencurves downward distally in Luangwa The posterior process ofthe iliac blade in Pascualgnathus is directed away from the is-chium (Bonaparte 1966) as in Exaeretodon and Menadon but isrelatively long as in Massetognathus and Luangwa
The proximal portion of the ischium is broad and dominatedby the ischial buttress The ischium constricts posterior to theproximal portion and expands distally into a spatulate platewhich is thickened at its posterior margin This thickening con-tinues along the dorsolateral surface of the ischium forming aridge that slopes ventrally until reaching the edge of the ischialbuttress The downward slope of this ridge produces a prominentgroove at the dorsal edge of the lateral surface of the ischiumthought to represent the origin of the ischio-trochantericusmuscle (Kemp 1980) The constriction of the ischium in Mena-don is like that of Luangwa albeit even more extreme in itsventral concavity but unlike that of Exaeretodon in which themedial and distal ends of the ischium are broadly united Theischium in general is a proportionally shorter element in Ex-aeretodon than in other traversodontids including Menadon(Bonaparte 1963) The anteroventral portion of the ischium thatconnects to the pubis is not preserved in either of the referredMenadon pelves
The pubis is the most incompletely preserved pelvic elementThe acetabular contribution of the pubis is smaller than that ofthe ischium or ilium Distal to the acetabulum the pubis is con-stricted indicating that the connection between the acetabularregion and the pubic plate was considerably narrower than inExaeretodon (Bonaparte 1963) but close akin to the pubis ofLuangwa (Kemp 1980) The pubic plate itself is not preservedThe ventral outline of the ischium and what is known of thepubis indicate that the obturator foramen was relatively larger inMenadon (Fig 14) than in Exaeretodon (Bonaparte 1963)
DISCUSSION
Comparison with the Holotype of Menadon besairiei
FMNH PR 2444 differs in several significant ways from theholotype of Menadon besairiei as previously described most no-tably in the possession of an internarial bar (initially consideredabsent in M besairiei) and four upper incisors (reported as threein M besairiei but with the caveat that the homologies of thefourth and fifth anterior tooth loci were problematic Flynn et al2000) However upon further study we interpret the lack of acontinuous internarial bar in the holotype as breakage ratherthan the natural condition of the skull (a prominent medial pro-tuberance is present on the ventral rim of the nares in UA10601) Regarding the incisor count it should be noted that theholotype of Menadon besairiei displays an unusual set of anterior
JOURNAL OF VERTEBRATE PALEONTOLOGY VOL 28 NO 2 2008454
Dow
nloa
ded
by [
UZ
H H
aupt
bibl
ioth
ek
Zen
tral
bibl
ioth
ek Z
uumlric
h] a
t 05
51 1
0 Se
ptem
ber
2013
FIGURE 10 Right scapulocoracoids of FMNH PR 2444 A stereopair of right scapulocoracoid in medial view B stereopair and interpretivedrawing of right scapulocoracoid in lateral view Scale bars equal 5 cm Abbreviationsmdashac acromion process C coracoid c gl coracoid contributionto the glenoid fossa c tr tubercle for coracoid attachment of triceps PC procoracoid pc f procoracoid foramen S scapula s gl scapularcontribution to the glenoid fossa
KAMMERER ET ALmdashCYNODONT FROM MADAGASCAR 455
Dow
nloa
ded
by [
UZ
H H
aupt
bibl
ioth
ek
Zen
tral
bibl
ioth
ek Z
uumlric
h] a
t 05
51 1
0 Se
ptem
ber
2013
teeth for a traversodontid As mentioned above Flynn et al(2000) noted that the homologies of these teeth were difficult todetermine Of the five anterior teeth preserved in the holotypethe anteriormost three are ldquoundoubtedly incisorsrdquo although I3 isldquounusually large and is caniniformrdquo (Flynn et al 2000423) Therobust recurved fourth tooth in the series was considered torepresent the canine with the subsequent tooth interpreted as anolder canine in the process of replacement Although FMNH PR2444 is edentulous the alveoli on the left side of the skull are wellpreserved and indicate an anterior tooth set identical to the ho-lotype The alveolar morphology indicates that the fifth toothwas canted anteriorly as in the holotype This suggests that themysterious fifth tooth is not the result of a transient period in thedental replacement history of M besairiei as suggested by Flynnet al (2000) but rather is the norm This leaves two possibilities1) M besairiei persistently had two canines This condition isunknown in cynodonts 2) The canine-like fourth tooth actuallyrepresents an incisor and the fifth tooth is the true canine Wefavor the latter explanation In FMNH PR 2444 the first fouralveoli are nearly identical in size and shape However the fifthalveolus while similar in anteroposterior length to its predeces-sors is strongly compressed labiolingually with an anteroposte-rior lengthlabiolingual width ratio of 044 (compared to 080 inthe fourth alveolus and 084ndash85 for I2ndash3) We interpret thismarked difference in alveolar ratio as indicating the separationbetween incisors and canine The canine-like I4 would then rep-resent the endpoint in a series of large increasingly caniniformincisors Supporting this interpretation recurved incisors are alsoobserved in juvenile Exaeretodon (eg MCP 3843 PV see Ab-dala et al 2002)
There are several legitimate differences between FMNH PR2444 and the holotype of Menadon besairiei but we take a con-servative taxonomic stance and argue that these differences rep-resent intraspecific variation and preservational artifacts FMNHPR 2444 represents a larger animal than UA 10601 The snoutlength (measured as the distance between the anterior edge ofthe orbit and a vertical plane through the anterior tip of thepremaxilla) of FMNH PR 2444 is 911 mm while that of UA10601 is 801 mm As such proportional differences between thetwo skulls may be attributable to ontogeny Variations in FMNHPR 2444 compared to the holotype that probably represent on-togentic differences include wider longer and deeper paraca-nine fossae a much deeper ventral zygomatic flange (althoughthe lower border of the flange is slightly abraded in the holotypeof Menadon besairiei there is no evidence that a major flangewas broken off) a proportionally shorter and more circular out-line to the posterior border of the maxilla below the zygomaticprocess and a more deeply emarginated posterior squamosal(ie a more prominent laterally projecting flange)
FMNH PR 2444 has a proportionally shorter more robustsnout and larger orbits than the holotype While these charactersare certainly variable in cynodonts it is unusual that the largerspecimen is shorter-snouted and larger-eyed contra the usualpattern The proportional differences between the two skulls areminor however (orbit diametersnout length is 042 for FMNHPR 2444 and 041 for the holotype) and are partially attributableto deformation (shear in FMNH PR 2444 and breakage of thepostorbital bar in the holotype) The following differences canalso probably be attributed to preservational artifact only fivepostcanine alveoli are preserved in FMNH PR 2444 while theholotype has eight postcanines The anteriormost three in theholotype however are tiny and peg-like As the area of themaxilla between the canine alveolus and the first preserved post-canine alveolus is poorly preserved in FMNH PR 2444 the ab-sence of these postcanines may be artifactual However it is alsopossible that the lower postcanine count of FMNH PR 2444compared to the holotype is an ontogenetic effect as postcaninecount is known to decrease with increasing skull size in Ex-
FIGURE 11 Left scapulocoracoids of FMNH PR 2444 A left scapu-locoracoid in medial view B left scapulocoracoid in lateral view Scalebars equal 5 cm
JOURNAL OF VERTEBRATE PALEONTOLOGY VOL 28 NO 2 2008456
Dow
nloa
ded
by [
UZ
H H
aupt
bibl
ioth
ek
Zen
tral
bibl
ioth
ek Z
uumlric
h] a
t 05
51 1
0 Se
ptem
ber
2013
FIGURE 12 Right pelvis of FMNH PR 2444 A stereopair of right pelvis in medial view B stereopair and interpretive drawing of right pelvis inlateral view Scale bars equal 5 cm Abbreviationsmdashbpp base of posterior process of ilium ib iliac blade IL ilium IS ischium ob f obturatorforamen P pubis
KAMMERER ET ALmdashCYNODONT FROM MADAGASCAR 457
Dow
nloa
ded
by [
UZ
H H
aupt
bibl
ioth
ek
Zen
tral
bibl
ioth
ek Z
uumlric
h] a
t 05
51 1
0 Se
ptem
ber
2013
aeretodon (Abdala et al 2002) The orbit of FMNH PR 2444 isless upwardly directed than that of the holotype (being at anangle of sim65deg to a sagittal plane through the midline of the skullas opposed to sim45deg in the holotype) This difference in orbitalangle is probably attributable to postmortem deformation as theskull of FMNH PR 2444 is noticeably sheared to the front leftThe appearance of a wider squarer incisor arcade and broader
anterior palate in FMNH PR 2444 can also be attributed to thisshear Finally the occipital region of FMNH PR 2444 is alto-gether taller and narrower than that of the holotype For ex-ample the foramen magnum in FMNH PR 2444 approximates arounded isosceles triangle in shape while the foramen magnumof the holotype is more ovoid These differences can also beattributed to deformation Altogether the differences between
FIGURE 13 Stereopairs of field number 8-31-98-387 an isolated right pelvis in A medial and B lateral view Scale bars equal 5 cm
JOURNAL OF VERTEBRATE PALEONTOLOGY VOL 28 NO 2 2008458
Dow
nloa
ded
by [
UZ
H H
aupt
bibl
ioth
ek
Zen
tral
bibl
ioth
ek Z
uumlric
h] a
t 05
51 1
0 Se
ptem
ber
2013
FMNH PR 2444 and the holotype fall within the range of intra-specific variation recorded for other gomphodont cynodontssuch as the traversodontids Massetognathus pascuali (Abdalaand Giannini 2000 Kammerer pers obs) and Exaeretodon(Abdala et al 2002) and the diademodontid Diademodon tetra-gonus (Grine et al 1978 Bradu and Grine 1979)
Phylogenetic Analysis
Utilizing the new information afforded by FMNH PR 2444and a reexamination of the holotype (UA 10601) Menadon be-sairiei was recoded in the Gondwanan gomphodont data matrixof Abdala et al (2006) Abdala et alrsquos (2006) data matrix en-compassed 18 taxa and 43 characters For the present analysis anadditional character ldquoincisor procumbencyrdquo (states 0-absent1-present) was added to Abdala et alrsquos (2006) data set (see
Appendix for coding of this character) Several traversodontidtaxa (eg lsquoScalenodonrsquo hirschsoni) have relatively large incisorsbut only in Menadon Scalenodontoides and Exaeretodon are theincisors procumbent Three of Abdala et alrsquos (2006) originalcodings for Menadon besairiei characters 0 (ldquonumber of upperincisorsrdquo) 5 (ldquoposition of paracanine fossa in relation to theupper caninerdquo) and 21 (ldquointernarial barrdquo) were changed to in-corporate data from the new specimen (see Appendix) Addi-tionally further examination of the holotype (UA 10601) ofMenadon besairiei allowed us to code characters 16ndash18 20 38and 40 (morphology of the lower postcanines all marked as ldquordquoin Abdala et alrsquos analysis) Parsimony searches were performedin PAUP v40b10 using a branch-and-bound search Addition-ally a bootstrap analysis (also using branch-and-bound) was per-formed using 1000 replicates
A total of 34 most parsimonious trees of length 103 werefound with the scores CI0573 RI0725 RC0415 andHI0427 The strict consensus tree (Fig 15A) did not recovera polytomy between Menadon Exaeretodon and Scalenodont-oides as in Abdala and Ribeiro (2003) or a (Menadon + Ex-aeretodon) sister-group relationship as in Abdala et al (2006)Rather Menadon was recovered as the sister taxon of the (Ex-aeretodon + Scalenodontoides) clade supporting Flynn et alrsquos(2000) original placement of this taxon There is strong bootstrapsupport (88) for Traversodontidae but within traversodontidsthe only strongly supported subclade is (Gomphodontosuchus +(Menadon + (Scalenodontoides + Exaeretodon))) with a boot-strap value of 80 Within this clade the (Menadon + (Scalen-odontoides + Exaeretodon)) grouping is supported by a boot-strap value of 83 and the (Scalenodontoides + Exaeretodon)grouping is supported by a bootstrap value of 94 (Fig 15B) Inaddition to the positioning of Menadon the strict consensus tree(Fig 15A) differs from the topology of Abdala et al (2006) inrecovering a sister-group relationship between Massetognathusand Santacruzodon rather than having Massetognathus andSantacruzodon as successive outgroups to the clade containingGomphodontosuchus Menadon Scalenodontoides and Ex-aeretodon The (Massetognathus + Santacruzodon) clade is diag-nosed by the absence of an anterolingual cusp in the upper post-canines denticulated incisor cutting margins and the presence ofan anterior cingulum in the upper postcanines
FIGURE 14 Reconstruction of the pelvis of Menadon besairiei basedon information from FMNH PR 2444 and field number 8-31-98-387Dotted area not preserved in either specimen reconstruction based onPascualgnathus (Bonaparte 1966) and Exaeretodon (Bonaparte 1963)
FIGURE 15 Cladogram of traversodontid relationships based on revised data set of Abdala et al (2006) showing A strict consensus tree and B50 majority rule tree Numbers at nodes of majority rule tree represent bootstrap values
KAMMERER ET ALmdashCYNODONT FROM MADAGASCAR 459
Dow
nloa
ded
by [
UZ
H H
aupt
bibl
ioth
ek
Zen
tral
bibl
ioth
ek Z
uumlric
h] a
t 05
51 1
0 Se
ptem
ber
2013
Relationships of Menadon besairiei
Menadon besairiei is the sister taxon to (Exaeretodon + Sca-lenodontoides) the latter clade diagnosed by the possession ofthree upper incisors a posteromedial position of the paracaninefossae relative to the canines the absence of an internarial barlarge canines a projected zygomatic process of the jugal and amaxilla that contributes to the margin of the subtemporal fenes-tra (the last three character states represent reversals) The clade(Menadon + (Exaeretodon + Scalenodontoides)) is diagnosed bythe possession of large procumbent incisors The presence offour incisors a primitive retention distinguishes Menadon be-sairiei from Exaeretodon Scalenodontoides and an unnamed tra-versodontid (ldquoTraversodontid Type 1rdquo) from the Santa MariaFormation of Brazil (Abdala et al 2001 Abdala and Ribeiro2002) Gomphodontosuchus is recovered as the sister taxon to(Menadon (Exaeretodon + Scalenodontoides)) as has beenfound in previous studies (Flynn et al 2000 Abdala and Ribeiro2003) The taxon name Gomphodontosuchinae Watson andRomer 1956 (ex Gomphodontosuchidae Watson and Romer1956 first given subfamilial rank in Lehman 1961) is availablefor this clade As circumscribed here Gomphodontosuchinaeconsists of the most inclusive clade containing Gomphodontosu-chus brasiliensis von Huene 1928264 but not Massetognathuspascuali Romer 19672
Menadon besairiei displays a mixture of primitive and derivedcharacters within traversodontids The internarial bar and fourincisors are primitive features whereas the enlargement and pro-cumbency of the incisors are derived features shared with Ex-aeretodon and Scalenodontoides The postcranium shows severalsimilarities to that of Exaeretodon (apparent exclusion of theprocoracoid from the glenoid some features of the pelvis tallcaudal neural spines) but in general displays a rather conserva-tive morphology observed throughout Gomphodontia The mor-phology of the scapulocoracoids is closer akin to diademodontidsor trirachodontids than Exaeretodon and the expanded thoracicrib of Menadon has only slight differences from those of trirach-odontids or early traversodontids like Luangwa The atlas-axiscomplex of Menadon is similar to that of the Early Triassic cyn-odont Cynognathus in possessing a concave axial spine with along posterior process Although most gomphodonts have beenreconstructed with a convex axial spine as in early mammals thisspine is broken in most of the best described gomphodont post-crania (eg Diademodon Brink 1955) and the lsquosaddle-shapedrsquoaxial morphology may be more widely distributed among cyn-odonts than previously realized The new specimens of Menadondescribed in this paper demonstrate that a generalized gompho-dont postcranial morphology was retained deep within the tra-versodontid tree and that the derived morphologies of either ofthe well-known taxa Massetognathus or Exaeretodon should notbe taken as representative of traversodontids in general
ACKNOWLEDGMENTS
This work would not have been possible without the continu-ing support and encouragement of our collaborative programwith the Deacutepartement de Paleacuteontologie et drsquoAnthropologie Bi-ologique Universiteacute drsquoAntananarivo Madagascar under the di-rection of Armand Rasoamiaramanana (Department Chairman)and Gisegravele Ravololonarivo Randria (Head of the PaleontologyLaboratories) We are extremely grateful to the National Geo-graphic Society for their long-term support of the fieldwork as-sociated with this project Additional financial or logistical sup-port for fieldwork was provided by The Field Museum the Uni-versity of California-Santa Barbara and the WWF (WorldwideFund for Nature Madagascar) We acknowledge the Ministegraverede LrsquoEneacutergie et des Mines for authorizing field efforts and for
continuing to facilitate our joint US-Madagascar explorationresearch and education program Steve Goodman and AsminaGandhi generously assisted us in innumerable ways includinghospitality nonpareil We especially thank R Andriantompoha-vana L Razafimanantsoa A Toto Volahy JA Rabarison AGandie and P Vavisaro for assistance in the field during theexpedition in which these new specimens were discovered BillSimpson and Gina Wesley-Hunt for their patient excavation ofthis important material Allison Smith Ray Vodden Lisa Berg-wall and Deborah Wagner for excellent preparation of a chal-lenging specimen Jennifer Botha and Elize Butler (NationalMuseum Bloemfontein) Michael Raath and Fernando Abdala(Bernard Price Institute) Thomas Kemp (Oxford UniversityMuseum of Natural History) Ray Symonds (University Museumof Zoology Cambridge) Bernard Battail (Museacuteum nationaldrsquoHistoire naturelle Paris) Michael Maisch (Universitaumlt Tuumlbin-gen) and Charles Schaff and Farish Jenkins (Museum of Com-parative Zoology Harvard) for access to specimens and AgustiacutenMartinelli Fernando Abdala and James Hopson for helpful re-views of an earlier version of the manuscript
LITERATURE CITED
Abdala F M C Barbarena and J Dornelles 2002 A new species of thetraversodontid cynodont Exaeretodon from the Santa Maria Forma-tion (MiddleLate Triassic) of Southern Brazil Journal of Verte-brate Paleontology 22313ndash325
Abdala F and N P Giannini 2000 Gomphodont cynodonts of theChantildeares Formation the analysis of an ontogenetic sequence Jour-nal of Vertebrate Paleontology 20501ndash506
Abdala F J Neveling and J Welman 2006 A new trirachodontidcynodont from the lower levels of the Burgersdorp Formation(Lower Triassic) of the Beaufort Group South Africa and the cla-distic relationships of Gondwanan gomphodonts Zoological Jour-nal of the Linnean Society 147383ndash413
Abdala F A M Ribeiro and C L Schultz 2001 A rich cynodont faunaof Santa Cruz do Sul Santa Maria Formation (Middle-Late Trias-sic) southern Brazil Neues Jahrbuch fuumlr Geologie und Palaumlontolo-gie Monatshefte 2001669ndash687
Abdala F and A M Ribeiro 2002 Nuevos cinodontes traversodoacuten-tidos (Synapsida-Eucynodontia) de la Formacioacuten Santa Maria(Triaacutesico Medio-Superior) Rio Grande do Sul Brasil RevistaEspantildeola de Paleontologiacutea 17237ndash243
Abdala F and A M Ribeiro 2003 A new traversodontid cynodontfrom the Santa Maria Formation (Ladinian-Carnian) of southernBrazil with a phylogenetic analysis of Gondwanan traversodontidsZoological Journal of the Linnaean Society 139529ndash545
Abdala F and A M S Teixeira 2004 A traversodontid cynodont ofAfrican affinity in the South American Triassic Palaeontologia Af-ricana 4011ndash22
Bonaparte J F 1962 Descripcioacuten del craacuteneo y mandiacutebula de Ex-aeretodon frenguellii Cabrera y su comparacioacuten con Diademodon-tidae Tritylodontidae y los cinodontes sudamericanos Publica-ciones del Museo Municipal de Ciencias Naturales y Tradicional deMar del Plata 1135ndash202
Bonaparte J F 1963 Descripcioacuten del esqueleto postcraneano de Ex-aeretodon (CynodontiandashTraversodontidae) Acta Geoloacutegica Lil-loana 45ndash52
Bonaparte J F 1966 Una nueva ldquofaunardquo Triaacutesica de Argentina (Ther-apsida Cynodontia Dicynodontia) Consideracions filogeneacuteticas ypaleobiogeograacuteficas Ameghiniana 8243ndash296
Bradu D and F E Grine 1979 Multivariate analysis of diademodon-tine crania from South Africa and Zambia South African Journal ofScience 79441ndash448
Brink A S 1955 A study of the skeleton of Diademodon Palaeonto-logia Africana 33-39
Brink A S 1963 Two new cynodonts from the Ntawere Formation inthe Luangwa Valley of Northern Rhodesia Palaeontologia Africana877ndash96
Broom R 1905 On the use of the term Anomodontia Records of theAlbany Museum 1266ndash269
Cabrera A 1943 El primer hallazgo de teraacutepsidos en la ArgentinaNotas del Museo de La Plata Paleontologiacutea 55317ndash331
JOURNAL OF VERTEBRATE PALEONTOLOGY VOL 28 NO 2 2008460
Dow
nloa
ded
by [
UZ
H H
aupt
bibl
ioth
ek
Zen
tral
bibl
ioth
ek Z
uumlric
h] a
t 05
51 1
0 Se
ptem
ber
2013
Chatterjee S 1982 A new cynodont reptile from the Triassic of IndiaJournal of Paleontology 56203ndash214
Crompton A W 1955 On some Triassic cynodonts from TanganyikaProceedings of the Zoological Society of London 125617ndash669
Crompton A W 1972 Postcanine occlusion in cynodonts and tritylo-donts Bulletin of the British Museum (Natural History) Geology2129ndash71
Crompton A W and F Ellenberger 1957 On a new cynodont from theMolteno Beds and the origin of the tritylodontids Annals of theSouth African Museum 441ndash13
Flynn J J J M Parrish B Rakotosaminimanana W F Simpson R LWhatley and A R Wyss 1999 A Triassic fauna from Madagascarincluding early dinosaurs Science 286763ndash765
Flynn J J J M Parrish B Rakotosamimanana L RanivoharimananaW F Simpson and A R Wyss 2000 New traversodontids (Syn-apsida Eucynodontia) from the Triassic of Madagascar Journal ofVertebrate Paleontology 20422ndash427
Gow C E and P J Hancox 1993 First complete skull of the LateTriassic Scalenodontoides (Reptilia Cynodontia) from southern Af-rica In Lucas S G and M Morales (eds) The nonmarine TriassicNew Mexico Museum of Natural History amp Science Bulletin 3161ndash168
Grine F E B D Hahn and C E Gow 1978 Aspects of relativegrowth and variability in Diademodon (Reptilia Therapsida) SouthAfrican Journal of Science 7450ndash58
Hopson J A 1984 Late Triassic traversodont cynodonts from NovaScotia and southern Africa Palaeontologia Africana 25181ndash201
Hopson J A 1985 Morphology and relationships of Gomphodontosu-chus brasiliensis von Huene (Synapsida Cynodontia Tritylodontoi-dea) from the Triassic of Brazil Neues Jahrbuch fuumlr Geologie undPalaumlontologie Monatshefte 1985285ndash299
Hopson J A and J W Kitching 2001 A probainognathian cynodontfrom South Africa and the phylogeny of nonmammalian cynodontsBulletin of the Museum of Comparative Zoology 1565ndash35
Hopson J A and H-D Sues 2006 A traversodont cynodont from theMiddle Triassic (Ladinian) of Baden-Wuumlrttemberg (Germany)Palaumlontologische Zeitschrift 80124ndash129
Huene F von 1928 Ein Cynodontier aus der Trias Brasiliens Zentral-blatt fuumlr Mineralogie Geologie und Palaumlontologie 1928251ndash270
Huene F von 1936 Die fossilen Reptilien des suumldamerikanischenGondwanalandes Ergebnisse der Sauriergrabungen in Suumldbrasilien192829 Lieferung 2 (Cynodontia)93ndash159 Franz F Heine Tuumlbin-gen
Jenkins F A 1970 The Chantildeares (Argentina) Triassic reptile faunaVII The postcranial skeleton of the traversodontid Massetognathuspascuali (Therapsida Cynodontia) Breviora 3521ndash28
Jenkins F A 1971 The postcranial skeleton of African cynodonts prob-lems in the early evolution of the mammalian postcranial skeletonBulletin of the Peabody Museum of Natural History 361ndash216
Kemp T S 1969 The atlas-axis complex of mammal-like reptiles Jour-nal of Zoology 159223ndash248
Kemp T S 1980 Aspects of the structure and functional anatomy of theMiddle Triassic cynodont Luangwa Journal of Zoology 191193ndash239
Kemp T S 1982 Mammal-Like Reptiles and the Origin of MammalsAcademic Press New York
Lehman J-P 1961 Cynodontia pp 140ndash191 in Piveteau J (ed) Traiteacutede Paleacuteontologie Masson et Cie Paris
Osborn 1903 On the primary division of the Reptilia into two sub-classes Synapsida and Diapsida Science 17275ndash276
Owen R 1861 Palaeontology or a systematic summary of extinct ani-mals and their geological relations 2nd Ed Edinburgh
Parrington F R 1934 On the cynodont genus Galesaurus with a noteon the functional significance of the changes in the evolution of thetheriodont skull Annals and Magazine of Natural History 10(13)38ndash67
Parrington F R 1946 On the cranial anatomy of cynodonts Proceed-ings of the Zoological Society of London 116181ndash197
Razafimbelo E 1987 Le basin de Morondava (Madagascar) Synthegravesegeacuteologique et structurale PhD dissertation University Louis Pas-teur Strasbourg France 189 pp
Reisz R R and H-D Sues 2000 Herbivory in Late Paleozoic andTriassic terrestrial vertebrates pp 9ndash41 in Sues H-D (ed) Evo-lution of Herbivory in Terrestrial Vertebrates Cambridge Univer-sity Press New York
Romer A S 1967 The Chantildeares (Argentina) Triassic reptile fauna IIITwo new gomphodonts Massetognathus pascuali and M terugiiBreviora 2641ndash25
Romer A S 1972 The Chantildeares (Argentina) Triassic reptile faunaXVII The Chantildeares gomphodonts Breviora 3961ndash9
Rougier G W J R Wible and J A Hopson 1992 Reconstruction ofthe cranial vessels in the Early Cretaceous mammal Vincelestes neu-quenianus implications for the evolution of the mammalian cranialvascular system Journal of Vertebrate Paleontology 12188ndash216
Rowe T 1988 Definition diagnosis and origin of Mammalia Journal ofVertebrate Paleontology 8241ndash264
Rowe T 1993 Early mammal phylogenetic systematics pp 129ndash145 inSzalay F S M J Novacek and M C McKenna (eds) MammalPhylogeny Vol 1 Mesozoic Differentiation MultituberculatesMonotremes Early Therians and Marsupials Springer-Verlag NewYork
Seeley H G 1894 Researches on the structure organization and clas-sification of the fossil Reptilia Part IX Section 3 On DiademodonPhilosophical Transactions of the Royal Society of London 1851029ndash1041
Seeley H G 1895 Researches on the structure organization and clas-sification of the fossil Reptilia Part IX Section 4 On the Gompho-dontia Philosophical Transactions of the Royal Society of London1861ndash57
Seeley H G 1895 Researches on the structure organization and clas-sification of the fossil Reptilia Part IX Section 5 On the skeletonin new Cynodontia from the Karroo rocks Philosophical Transac-tions of the Royal Society of London 18659ndash148
Sues H-D J A Hopson and N H Shubin 1992 Affinities of Sca-lenodontoides plemmyridon Hopson 1984 (Synapsida Cynodontia)from the Upper Triassic of Nova Scotia Journal of Vertebrate Pa-leontology 12168ndash171
Sues H-D and P E Olsen 1990 Triassic vertebrates of Gondwananaspect from the Richmond basin of Virginia Science 2491020ndash1023
Swofford D L 2003 PAUP v 40b10 Sinauer Associates Inc Pub-lishers Sunderland Massachusetts
Watson D M S and A S Romer 1956 A classification of therapsidreptiles Bulletin of the Museum of Comparative Zoology 11435ndash89
Submitted September 20 2007 accepted November 3 2007
KAMMERER ET ALmdashCYNODONT FROM MADAGASCAR 461
Dow
nloa
ded
by [
UZ
H H
aupt
bibl
ioth
ek
Zen
tral
bibl
ioth
ek Z
uumlric
h] a
t 05
51 1
0 Se
ptem
ber
2013
APPENDIX Data matrix used in the phylogenetic analysesThis is a revised version of the matrix of Abdala et al (2006) with the following changes character 43 [Incisor procumbency (0) absent (1) present]added Menadon recoded for characters 0 (Number of upper incisors was 1 ldquothreerdquo changed to 0 ldquofourrdquo) 5 (Position of paracanine fossae in relationto the upper canine was changed to 1 ldquomedialrdquo) 16 (Overall morphology of lower postcanines in occlusal view was changed to 3 ldquoquadran-gularrdquo) 17 (Position of the transverse crest in lower postcanines was changed to 1 ldquoanteriorrdquo) 18 (Number of cusps in the transverse crest of thelower postcanines was changed to 0 ldquotwordquo) 20 (Anterior cingulum in the lower postcanines was changed to 2 ldquoabsentrdquo) 21 (Internarial barwas 1 ldquoabsentrdquo changed to 0 ldquopresentrdquo) 38 (Posterior cingulum in the lower postcanines was changed to 1 ldquoabsentrdquo) and 40 (Deep occlusalbasins in the postcanines was changed to 1 ldquopresentrdquo) Langbergia recoded for character 34 (Incisor cutting margins was 0 ldquoserratedrdquo changedto 1 ldquosmoothly ridgedrdquo) based on the description of the incisors in Abdala et al (2006) Gomphodontosuchus recoded for character 31 (Maxilla inthe margin of the subtemporal fenestra was 1 ldquoincludedrdquo changed to 0 ldquoexcludedrdquo) based on von Huene (1936) and Kammerer (pers obs of theholotype) contra Hopsonrsquos (1985) reconstruction lsquoScalenodonrsquo hirschsoni recoded for character 39 (Anterior cingulum in the upper postcanines was1 ldquoabsentrdquo changed to 0 ldquopresentrdquo) based on Abdala (pers comm 2006) Trirachodon recoded for character 30 (Ectopterygoid was changedto 0 ldquopresentrdquo) based on Abdala (pers comm 2006) and Traversodon recoded for character 15 (Number of cusps in the sectorial border of the upperpostcanines was 0 ldquomore than twordquo changed to 1 ldquotwordquo) based on Abdala (pers comm 2006) and Kammerer (pers obs)
1111111111 2222222222 3333333333 44440123456789 0123456789 0123456789 0123456789 0123
Cynognathus 00000000 000 000000000 00000000 000Diademodon 0000001000 0000100 000010A000 0000000100 0100Trirachodon 000000100B 1A002201 001A100010 0001001200 0110Langbergia 0000001000 10000201 0000101010 0001101200 0110Cricodon 0100001000 11000201 0010100 0001200 0110Andescynodon 0000002001 01013101 101120010 0112311 1100Massetognathus 00111121A2 1110003100 2011200011 1011212300 1100Exaeretodon 1110122112 010103101 2111110201 1111112311 1101Luangwa 0000001002 11010A3100 101010010 01002300 1100Scal angustifrons 000000100B 1111023100 10102101 00002301 1100Scal hirschsoni 11110202 1100113100 1010 1112300 1100Traversodon 0000002012 1101113100 201010 011230 1100Gomphodontosuchus 0011112112 0103101 2010 0112311 1100Pascualgnathus 1000001000 000231 1100001 0011230 1100Scalenodontoides 1112212 01003101 21111021 1212311 1101Menadon 0111112112 01012310 20121020 0102311 1101Dadadon 00002112 111112 013 11231 1100Santacruzodon 00002112 1110003101 201300 212300 1100
A(01) B(02)
JOURNAL OF VERTEBRATE PALEONTOLOGY VOL 28 NO 2 2008462
Dow
nloa
ded
by [
UZ
H H
aupt
bibl
ioth
ek
Zen
tral
bibl
ioth
ek Z
uumlric
h] a
t 05
51 1
0 Se
ptem
ber
2013
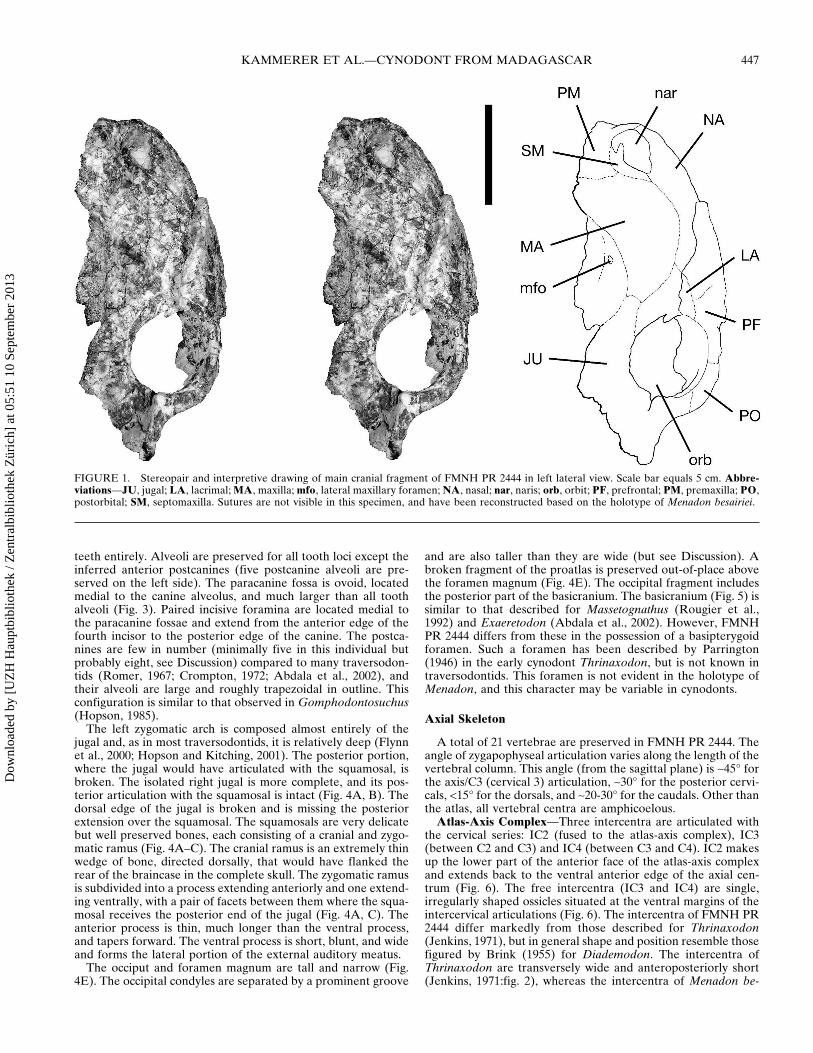
teeth entirely Alveoli are preserved for all tooth loci except theinferred anterior postcanines (five postcanine alveoli are pre-served on the left side) The paracanine fossa is ovoid locatedmedial to the canine alveolus and much larger than all toothalveoli (Fig 3) Paired incisive foramina are located medial tothe paracanine fossae and extend from the anterior edge of thefourth incisor to the posterior edge of the canine The postca-nines are few in number (minimally five in this individual butprobably eight see Discussion) compared to many traversodon-tids (Romer 1967 Crompton 1972 Abdala et al 2002) andtheir alveoli are large and roughly trapezoidal in outline Thisconfiguration is similar to that observed in Gomphodontosuchus(Hopson 1985)
The left zygomatic arch is composed almost entirely of thejugal and as in most traversodontids it is relatively deep (Flynnet al 2000 Hopson and Kitching 2001) The posterior portionwhere the jugal would have articulated with the squamosal isbroken The isolated right jugal is more complete and its pos-terior articulation with the squamosal is intact (Fig 4A B) Thedorsal edge of the jugal is broken and is missing the posteriorextension over the squamosal The squamosals are very delicatebut well preserved bones each consisting of a cranial and zygo-matic ramus (Fig 4AndashC) The cranial ramus is an extremely thinwedge of bone directed dorsally that would have flanked therear of the braincase in the complete skull The zygomatic ramusis subdivided into a process extending anteriorly and one extend-ing ventrally with a pair of facets between them where the squa-mosal receives the posterior end of the jugal (Fig 4A C) Theanterior process is thin much longer than the ventral processand tapers forward The ventral process is short blunt and wideand forms the lateral portion of the external auditory meatus
The occiput and foramen magnum are tall and narrow (Fig4E) The occipital condyles are separated by a prominent groove
and are also taller than they are wide (but see Discussion) Abroken fragment of the proatlas is preserved out-of-place abovethe foramen magnum (Fig 4E) The occipital fragment includesthe posterior part of the basicranium The basicranium (Fig 5) issimilar to that described for Massetognathus (Rougier et al1992) and Exaeretodon (Abdala et al 2002) However FMNHPR 2444 differs from these in the possession of a basipterygoidforamen Such a foramen has been described by Parrington(1946) in the early cynodont Thrinaxodon but is not known intraversodontids This foramen is not evident in the holotype ofMenadon and this character may be variable in cynodonts
Axial Skeleton
A total of 21 vertebrae are preserved in FMNH PR 2444 Theangle of zygapophyseal articulation varies along the length of thevertebral column This angle (from the sagittal plane) is sim45deg forthe axisC3 (cervical 3) articulation sim30deg for the posterior cervi-cals lt15deg for the dorsals and sim20-30deg for the caudals Other thanthe atlas all vertebral centra are amphicoelous
Atlas-Axis ComplexmdashThree intercentra are articulated withthe cervical series IC2 (fused to the atlas-axis complex) IC3(between C2 and C3) and IC4 (between C3 and C4) IC2 makesup the lower part of the anterior face of the atlas-axis complexand extends back to the ventral anterior edge of the axial cen-trum (Fig 6) The free intercentra (IC3 and IC4) are singleirregularly shaped ossicles situated at the ventral margins of theintercervical articulations (Fig 6) The intercentra of FMNH PR2444 differ markedly from those described for Thrinaxodon(Jenkins 1971) but in general shape and position resemble thosefigured by Brink (1955) for Diademodon The intercentra ofThrinaxodon are transversely wide and anteroposteriorly short(Jenkins 1971fig 2) whereas the intercentra of Menadon be-
FIGURE 1 Stereopair and interpretive drawing of main cranial fragment of FMNH PR 2444 in left lateral view Scale bar equals 5 cm Abbre-viationsmdashJU jugal LA lacrimal MA maxilla mfo lateral maxillary foramen NA nasal nar naris orb orbit PF prefrontal PM premaxilla POpostorbital SM septomaxilla Sutures are not visible in this specimen and have been reconstructed based on the holotype of Menadon besairiei
KAMMERER ET ALmdashCYNODONT FROM MADAGASCAR 447
Dow
nloa
ded
by [
UZ
H H
aupt
bibl
ioth
ek
Zen
tral
bibl
ioth
ek Z
uumlric
h] a
t 05
51 1
0 Se
ptem
ber
2013
FIGURE 2 Stereopairs of main cranial fragment of FMNH PR 2444 in A dorsal view and B palatal view Scale bars equal 5 cm
JOURNAL OF VERTEBRATE PALEONTOLOGY VOL 28 NO 2 2008448
Dow
nloa
ded
by [
UZ
H H
aupt
bibl
ioth
ek
Zen
tral
bibl
ioth
ek Z
uumlric
h] a
t 05
51 1
0 Se
ptem
ber
2013
sairiei have equal transverse and anteroposterior dimensions Attheir maximum depth in Thrinaxodon the intercentra barely ex-tend beyond the ventral margins of the cervical pleurocentra InFMNH PR 2444 IC4 extends for half of its total depth below theventral margins of the cervical pleurocentra In IC3 this condi-tion is even more extreme extending below the ventral marginsof the cervical pleurocentra for 80 of its total depth The shapeand position of the intercentra give a strongly arched appearanceto the ventral profile of elements in the cervical series The ven-tral surfaces of the intercentra bear a posteriorly directed process
on each side In IC3 a weak ridge originates between the pairedprocesses and extends posteriorly indicating muscle attachmenton the intercentra The condition in IC4 cannot be determinedbecause of incomplete preservation Although only three inter-centra are known ventral facets in the centra of the isolatedcervicals indicate that others were present
As in other cynodonts the atlas-axis complex of FMNH PR2444 consists of a pair of separate atlantal neural arches articu-lating with the occipital condyles and the atlas centrum Only theleft atlas arch is preserved in this specimen (Fig 7) It is similar
FIGURE 3 Stereopair and interpretive drawing of anterior palatal region of FMNH PR 2444 Scale bar equals 5 cm AbbreviationsmdashC canineI incisor i f incisive foramina PC postcanine pc f paracanine fossa Postcanine numbering based on comparison with the holotype UA 10601
FIGURE 4 Cranial fragments of FMNH PR 2444 A stereopair of right zygomatic arch in lateral view Scale bar equals 5 cm B stereopair of rightzygomatic arch in medial view Scale bar equals 5 cm C stereopair of left squamosal in lateral view Scale bar equals 5 cm D sagittal crest in rightlateral view Scale bar equals 1 cm E occiput in occipital view Scale bar equals 1 cm
KAMMERER ET ALmdashCYNODONT FROM MADAGASCAR 449
Dow
nloa
ded
by [
UZ
H H
aupt
bibl
ioth
ek
Zen
tral
bibl
ioth
ek Z
uumlric
h] a
t 05
51 1
0 Se
ptem
ber
2013
to the atlantal neural arches described for other cynodonts suchas Massetognathus (Kemp 1969 Jenkins 1971) There is a me-dial facet for articulation with the centrum of the atlas (theldquoodontoid facetrdquo of Kemp 1969) and an anterior facet for ar-ticulation with the occipital condyle (Fig 7B) A pair of pro-cesses dominates the atlas arch a dorsal lamina (a roughly rect-angular plate of bone arching above the facets) and a largeposterolaterally directed transverse process An oval tubercle atthe anterior end of the transverse process represents the facet forthe proatlas
The centrum of the atlas is fused to the axis forming a singlelarge vertebral unit (Fig 6) A weak odontoid process is presentat the dorsal limit of the anterior surface of the atlas The ante-rior aspect of the centrum is dominated by a roughly lsquoTrsquo-shapedstructure composed of the two facets for articulation with theatlas arches and the facet for articulation with the atlas intercen-trum The axis bears no prezygapophyses and as such does notarticulate with the atlas arch as in Early Triassic cynodonts suchas Galesaurus (Parrington 1934) There appears to be a trendtowards loss of the axial prezygapophyses in later traversodon-
tids which are present but vestigial in Massetognathus (Jenkins1970 although in the specimens described the prezygapophysesare broken so their reduction is perhaps questionable) and ab-sent in Exaeretodon (Bonaparte 1963) The transverse processesof the axis are large and robust compared to those of the othercervicals The axial spine is saddle-shaped with a concave dorsalprofile unlike previous reconstructions of the traversodontidaxis which depict a straight or convex dorsal profile similar tothat observed in galesaurid cynodonts and early mammals (Jen-kins 1971) The morphology of the axial spine is similar to thatof the Early Triassic cynodont Cynognathus (Seeley 1895) InCynognathus however the neural spine of the third cervical(C3) is tall and canted posteriorly behind the posterior process ofthe axial spine whereas in FMNH PR 2444 the short C3 neuralspine fits below the posterior portion of the axial spine Theposterior process of the axial spine bears a pair of parallel elon-gate grooves on its underside attachment sites for the axial mus-culature
Post-Axial VertebraemdashThree post-axial cervicals are knownfor FMNH PR 2444 (Figs 6 8AndashD) The first of these C3 is
FIGURE 5 Stereopair of ventral view of the occipital fragment of FMNH PR 2444 showing the basicranium Anterior is up
FIGURE 6 Stereopair and interpretive drawing of atlas-axis complex and third cervical of FMNH PR 2444 in left lateral view Scale bar equals5 cm Abbreviationsmdasha n sp axial neural spine d dens IC intercentrum f at a facet for articulation with the atlas arch p parapophysis pozpostzygapophysis t p transverse process
JOURNAL OF VERTEBRATE PALEONTOLOGY VOL 28 NO 2 2008450
Dow
nloa
ded
by [
UZ
H H
aupt
bibl
ioth
ek
Zen
tral
bibl
ioth
ek Z
uumlric
h] a
t 05
51 1
0 Se
ptem
ber
2013
preserved in articulation with the axis (Fig 6) The two isolatedcervicals bear successively taller and anterposteriorly wider neu-ral spines (Fig 8AndashD) Based on their staggered size and failureto articulate we suggest that they represent C5 and C7 (assumingthat this specimen possessed the generalized cynodont cervicalnumber of seven) The cervicals display robust transverse pro-cesses which are directed laterally and slightly anteriorly A par-apophysis lies below the anterior edge of the transverse processalong the antero-lateral edge of the centrum No cervical ribs arepreserved but their presence can be inferred from the par-apophyses The notochordal concavity is as in Massetognathussurrounded by a raised rim at each end of the centrum (Jenkins1970) No anapophyses are present The neural spines for allthree post-axial cervicals are tapering and slightly curved poste-riorly The neural spine in C3 is relatively short and wide(thereby fitting underneath the posterior overhang of the axialspine see Fig 6) while that of C5 is tall and narrow The spinefor C7 is taller and anteroposteriorly wider than that of C5 andthe dorsal edge is more rounded similar to the condition in thedorsal vertebrae
Eight post-cervical presacral vertebrae are known for thisspecimen We will refer to them here simply as dorsal vertebraegiven the incompleteness of the vertebral column in the speci-men and the uncertainty concerning the thoraciclumbar transi-tion in this taxon However it is probable that they representthoracic vertebrae (see below) In most cynodonts the lumbar
vertebrae are synostosed to specialized ribs with overlappingelements In earlier taxa (eg Thrinaxodon Cynognathus Lu-angwa Jenkins 1971 Kemp 1980) expanded costal plates arepresent on both thoracic and lumbar ribs In Massetognathus thelumbar ribs consist of a pair of prongs one directed anteriorlybearing a facet for articulation with the previous rib and theother directed posteriorly (Jenkins 1970) In Exaeretodon allpresacral ribs are identical of simple morphology and not syn-ostosed to the vertebrae (Bonaparte 1963) A single isolated rib(Fig 9B) in FMNH PR 2444 demonstrates that some of the ribsin Menadon besairiei were expanded However all preservedvertebrae in this specimen have facets for articulation with theribs rather than breakage points where they would have beenfused to ribs As such it is probable that all of the vertebraepreserved in this specimen represent thoracics although we can-not discount the possibility that as in Exaeretodon no presacralvertebrae were fused to ribs
The neural spines in all of the dorsal vertebrae are elongateposteriorly inclined and roughly rectangular in lateral view witha rounded laterally expanded platform dorsally (Fig 8EndashJ)Costal foveae are present at the anterior and posterior edges ofeach centrum Of the eight dorsal vertebrae there are two ar-ticulated sets and three isolated elements (two from 9-4-98-440and one from 9-2-98-422) The first isolated vertebra (Fig 8E F)has a taller neural spine and shorter centrum than the otherdorsal vertebrae It is most similar in proportions to the isolated
FIGURE 7 Stereopairs and interpretive drawings of left atlas arch of FMNH PR 2444 in A lateral view Anterior is left B medial view Anterioris right Scale bars equal 1 cm Abbreviationsmdashafa anterior facet for articulation with the occipital condyle dl dorsal lamina mfa medial facet forarticulation with the atlas centrum (odontoid facet) pfa tubercle forming the facet for articulation with the proatlas tp transverse process
KAMMERER ET ALmdashCYNODONT FROM MADAGASCAR 451
Dow
nloa
ded
by [
UZ
H H
aupt
bibl
ioth
ek
Zen
tral
bibl
ioth
ek Z
uumlric
h] a
t 05
51 1
0 Se
ptem
ber
2013
FIGURE 8 Vertebrae of FMNH PR 2444 Cervical vertebra 5 in A left lateral and B anterior views Cervical vertebra 7 in C left lateral andD anterior views Isolated dorsal vertebra from anterior region of the series in E left lateral and F anterior views G stereopair and interpretivedrawing of articulated set of two dorsal vertebrae from near the midpoint of the series in left lateral view Anterior is up H articulated set of threedorsal vertebrae from anterior region of the series in left and right lateral views I isolated vertebra from near the midpoint of the series (from fieldnumber 9-2-98-422) J isolated vertebra from posterior region of the series Sacral vertebra in K dorsal view (the distal region of another sacral ribwhich is identical in shape to that of the intact sacral rib albeit slightly larger is also shown in this figure) and L left lateral view Caudal vertebraeM articulated set of two vertebrae from the anterior region of the series showing the lsquowingrsquo-shaped caudal ribs in dorsal view Anterior is left Nisolated caudal vertebra showing tall neural spine in left lateral view O isolated caudal vertebra showing tall neural spine in posterior view Particulated set of four vertebrae from near the end of the caudal series in dorsal view Anterior is left Scale bars equal 1 cm Abbreviationsmdasha cof anterior costal fovea n sp neural spine p co f posterior costal fovea poz postzygapophysis prz prezygapophysis
JOURNAL OF VERTEBRATE PALEONTOLOGY VOL 28 NO 2 2008452
Dow
nloa
ded
by [
UZ
H H
aupt
bibl
ioth
ek
Zen
tral
bibl
ioth
ek Z
uumlric
h] a
t 05
51 1
0 Se
ptem
ber
2013
cervicals and was probably located near the front of the dorsalseries The first articulated set consists of three vertebrae (Fig8H) The middle vertebra in this set is broken from the anteriorbase of the neural spine to the ventral midpoint and most of theright side of the anterior half of the centrum is missing This setof vertebrae probably also came from the anterior region of theseries as their neural spines are longer and narrower than allvertebrae other than the one described above The second ar-ticulated set (Fig 8G) consists of two well-preserved vertebraethat based on comparisons with Massetognathus and Ex-aeretodon (Jenkins 1970 Bonaparte 1963) probably came fromnear the midpoint of the dorsal series The second isolated ver-tebra (Fig 8I) from 9-2-98-422 is similar to the members of thesecond set The third isolated vertebra (Fig 8J) appears to befrom the posterior end of the series based on the relatively shortneural spine and reduced posterior costal fovea
One sacral vertebra (Fig 8K L) is known consisting of awell-preserved centrum neural spine and the right sacral rib(the left is broken off) The centrum is shorter (both anteropos-teriorly and dorsoventrally) than those of the dorsal vertebraeand the neural spine is shorter and more rounded in profileAlso the neural spine is directed vertically unlike those of thedorsal vertebrae which are canted posteriorly The zygapophy-ses are poorly developed The sacral rib is fused to a singlesynapophysis and projects anterolaterally The rib narrows to-wards the middle and expands distally terminating in a widerectangular facet where it would meet the ilium (Fig 8K)
Seven caudal vertebrae are known The caudal vertebrae aredistinctive in having very tall narrow neural spines Some EarlyTriassic cynodonts possess blunt neural spines of moderateheight on the proximal caudals (see for example Thrinaxodonin Jenkins 1971 and Diademodon in Brink 1955) Massetogna-thus represents an extreme condition wherein the caudal neuralspines are greatly reduced (Jenkins 1970) Very tall neuralspines are seen in Exaeretodon (Bonaparte 1963) and thosespines are unusual in that they do not diminish in height until thelast caudals The caudal vertebrae of FMNH PR 2444 consist ofa pair of articulated vertebrae most likely from near the sacrum(Fig 8M) an isolated caudal lacking postzygapophyses and ribsbut with the neural spine (Fig 8N O) and a quartet of smaller
articulated elements (Fig 8P) The anterior caudals have nar-rower neural spines and shorter zygapophyses than the subse-quent vertebrae in the series The caudal ribs attached to thispair have a crescentic distal region that curves anteriorly pro-ducing a lsquowingrsquo-shape Only a portion of the right rib of theisolated caudal is preserved but the lsquowingrsquo-shaped morphologyof the near-sacral caudal ribs does not appear to have been pre-sent In the posteriormost four vertebrae caudal ribs are pre-served on the left side of the first and the right side of the secondand third members On the first of these vertebrae the rib isshort curved posteriorly (but not lsquowingrsquo-shaped) and taperingRibs of the second and third vertebrae are less curved resem-bling posteriorly directed rods The zygapophyses of both theisolated caudal vertebra and the quartet of small caudals areanteroposteriorly elongate becoming increasingly so posteriorly
Thoracic RibsmdashNine thoracic ribs are known from 9-4-98-440(four left four right and an indeterminate splint of bone Fig9A indeterminate bone not illustrated) These elements re-semble the flattened ribs lacking costal plates of Exaeretodon(Bonaparte 1963) and the anterior thoracic ribs of Massetogna-thus and Luangwa (Jenkins 1970 Kemp 1980) The tuberculumand capitulum are separated by a shallow depression The tuber-culum has a shorter base than the capitulum but their proximalfaces are subequal in size An additional isolated left rib (Fig9B) from 9-2-98-422 is expanded indicating that unlike Ex-aeretodon Menadon rib morphology was heterogeneous alongthe length of the dorsal series This rib closely resembles theposterior thoracic ribs described for Luangwa (Kemp 1980fig16) Pascualgnathus (Bonaparte 1966fig 7) and the trirach-odontid Cricodon (Crompton 1955 Fig 4) This rib is con-stricted at the base of the head then expands distally with an-terior and posterior costal plates As in Luangwa the anteriorcostal plate is shallow and evenly sloping proximally and distallywhile the posterior costal plate is falcate with a concave edgedistally The distal expansion of this rib occurs at an angle of sim30degto the long axis of the rib head Despite their overall similaritythe expanded rib in FMNH PR 2444 does exhibit some differ-ences from the posterior thoracic rib of Luangwa The expandedrib of Menadon has a longer shaft between the rib head and thecostal plates and a shorter capitulum lacks a ridge on the pos-
FIGURE 9 A anterior thoracic ribs of FMNH PR 2444 in anterior view (indeterminate rib not figured) B expanded posterior thoracic rib ofFMNH PR 2444 in anterior view Scale bars equal 1 cm
KAMMERER ET ALmdashCYNODONT FROM MADAGASCAR 453
Dow
nloa
ded
by [
UZ
H H
aupt
bibl
ioth
ek
Zen
tral
bibl
ioth
ek Z
uumlric
h] a
t 05
51 1
0 Se
ptem
ber
2013
terior costal plate and has an anteroposteriorly shorter anteriorcostal plate Additionally the posterior margin of the costal plateis less curved than that of Luangwa These differences couldindicate that the expanded rib of Menadon is from a more pos-terior position in the series than the rib from Luangwa as re-duced curvature and increasing confluence of the tuberculumand capitulum are characteristic of lumbar ribs in traversodon-tids (Jenkins 1970 Kemp 1980) However the length of theshaft is at odds with this interpretation and in Cricodon andPascualgnathus the more posterior ribs are not more similar tothe element from FMNH PR 2444 These differences probablyrepresent legitimate morphological distinctions between the ribsof earlier gomphodonts and Menadon The head of the expandedrib is well defined and unbroken indicating that it was not syn-ostosed to a vertebra
Appendicular Skeleton
The appendicular skeleton is represented by both scapulocora-coids and a fragment of a clavicle from field number 9-4-98-440and a partial right pelvis from field number 9-2-98-422 An ad-ditional partial right pelvis is represented from field number8-31-98-387 The scapulocoracoids (Figs 10 11) are very similarto those described for other traversodontids as well as cynog-nathids and diademodontids (Jenkins 1970 1971) The scapularinfraspinous fossa has out-turned anterior and posterior edgesThe anterior edge of the scapula is rounded and curves laterallyforming the scapular spine At the lower end of the spine theacromion forms a prominent process (broken in the left scapula)Below this process the scapula is constricted and then expandsventrally to form the dorsal margin of the articular glenoid Theglenoid fossa is in general morphology very similar to that ofother Triassic cynodonts (Jenkins 1970 1971 Kemp 1980) It isposteroventrally oriented with roughly equal contributions fromthe scapular and coracoid glenoid facets The coracoid glenoidfacet is roughly triangular while the scapular glenoid facet issemiovoid as in other cynodonts In anterior or posterior viewthe fossa appears smoothly crescentic The procoracoid is aslightly curved trapezoidal plate of bone It does not appear tocontribute to the glenoid fossa The procoracoid foramen is lo-cated midway up the height of the procoracoid at a point directlybelow the base of the acromion process The coracoid is semi-crescentic curving posteriorly from a broad base The coracoidin Menadon is longer and more falcate than those described forExaeretodon (Bonaparte 1963) and Luangwa (Kemp 1980 al-though the distal tip of both coracoids is broken in the specimenof Luangwa [TSK 121] described by Kemp and may have beenlonger than reconstructed) but agrees closely with those of Mas-setognathus and cynognathidsdiademodontids (Jenkins 19701971) The distal coracoid tubercle for attachment to the tricepsis strongly curved upwards similar to the condition in Masseto-gnathus (Jenkins 1970) and Diademodon (Brink 1955)
A small fragment of bone resembling a curved cylinder almostcertainly represents part of a clavicle It has no distinguishingfeatures other than a groove associated with a ridge on one sideHowever the degree of curvature observed precludes this ele-ment from representing a long bone and the circular cross-section excludes it from being a rib
The pelvis of FMNH PR 2444 (Fig 12) resembles those ofother traversodontids in general morphology The ilium bears anarrow anteriorly directed iliac blade curving outward distallyThe anterior edge of the iliac blade slopes upward at a roughly45deg angle similar to Luangwa (Kemp 1980) but unlike Ex-aeretodon in which the anterior edge of the iliac blade curvesbackwards (Bonaparte 1963) or Pascualgnathus and Masseto-gnathus in which it is rounded (Bonaparte 1966 Jenkins 1970)The supraacetabular buttress at the base of the ilium is located inan anterodorsal position relative to the acetabulum as in other
traversodontids but unlike earlier cynodonts in which it is lo-cated dorsally (Jenkins 1970) The acetabular buttresses of thethree pelvic bones are largely confluent forming a fairly uniformlsquoliprsquo around the acetabulum as in Exaeretodon (Bonaparte1963) On the contrary in Luangwa the supraacetabular ischialand pubic buttresses are highly distinct spaced 6ndash8 mm apart andwithout confluence of their acetabular facets The pelvis ofFMNH PR 2444 does have a small depression between the su-praacetabular and ischial buttresses (also present in Ex-aeretodon Bonaparte 1963) but this is not as developed as inLuangwa The posterior extent of the iliac blade is broken inFMNH PR 2444 but this region is preserved in another isolatedright pelvis field number 8-31-98-387 (Fig 13) probably pertain-ing to Menadon besairiei The posterior process of the iliac bladein this specimen is relatively short and directed away from thedorsal edge of the ischium as in Exaeretodon (Bonaparte 1963)but unlike Massetognathus and Luangwa in which the posteriorprocess is long and extends roughly horizontally over the dorsaledge of the ischium (Jenkins 1970 Kemp 1980) and evencurves downward distally in Luangwa The posterior process ofthe iliac blade in Pascualgnathus is directed away from the is-chium (Bonaparte 1966) as in Exaeretodon and Menadon but isrelatively long as in Massetognathus and Luangwa
The proximal portion of the ischium is broad and dominatedby the ischial buttress The ischium constricts posterior to theproximal portion and expands distally into a spatulate platewhich is thickened at its posterior margin This thickening con-tinues along the dorsolateral surface of the ischium forming aridge that slopes ventrally until reaching the edge of the ischialbuttress The downward slope of this ridge produces a prominentgroove at the dorsal edge of the lateral surface of the ischiumthought to represent the origin of the ischio-trochantericusmuscle (Kemp 1980) The constriction of the ischium in Mena-don is like that of Luangwa albeit even more extreme in itsventral concavity but unlike that of Exaeretodon in which themedial and distal ends of the ischium are broadly united Theischium in general is a proportionally shorter element in Ex-aeretodon than in other traversodontids including Menadon(Bonaparte 1963) The anteroventral portion of the ischium thatconnects to the pubis is not preserved in either of the referredMenadon pelves
The pubis is the most incompletely preserved pelvic elementThe acetabular contribution of the pubis is smaller than that ofthe ischium or ilium Distal to the acetabulum the pubis is con-stricted indicating that the connection between the acetabularregion and the pubic plate was considerably narrower than inExaeretodon (Bonaparte 1963) but close akin to the pubis ofLuangwa (Kemp 1980) The pubic plate itself is not preservedThe ventral outline of the ischium and what is known of thepubis indicate that the obturator foramen was relatively larger inMenadon (Fig 14) than in Exaeretodon (Bonaparte 1963)
DISCUSSION
Comparison with the Holotype of Menadon besairiei
FMNH PR 2444 differs in several significant ways from theholotype of Menadon besairiei as previously described most no-tably in the possession of an internarial bar (initially consideredabsent in M besairiei) and four upper incisors (reported as threein M besairiei but with the caveat that the homologies of thefourth and fifth anterior tooth loci were problematic Flynn et al2000) However upon further study we interpret the lack of acontinuous internarial bar in the holotype as breakage ratherthan the natural condition of the skull (a prominent medial pro-tuberance is present on the ventral rim of the nares in UA10601) Regarding the incisor count it should be noted that theholotype of Menadon besairiei displays an unusual set of anterior
JOURNAL OF VERTEBRATE PALEONTOLOGY VOL 28 NO 2 2008454
Dow
nloa
ded
by [
UZ
H H
aupt
bibl
ioth
ek
Zen
tral
bibl
ioth
ek Z
uumlric
h] a
t 05
51 1
0 Se
ptem
ber
2013
FIGURE 10 Right scapulocoracoids of FMNH PR 2444 A stereopair of right scapulocoracoid in medial view B stereopair and interpretivedrawing of right scapulocoracoid in lateral view Scale bars equal 5 cm Abbreviationsmdashac acromion process C coracoid c gl coracoid contributionto the glenoid fossa c tr tubercle for coracoid attachment of triceps PC procoracoid pc f procoracoid foramen S scapula s gl scapularcontribution to the glenoid fossa
KAMMERER ET ALmdashCYNODONT FROM MADAGASCAR 455
Dow
nloa
ded
by [
UZ
H H
aupt
bibl
ioth
ek
Zen
tral
bibl
ioth
ek Z
uumlric
h] a
t 05
51 1
0 Se
ptem
ber
2013
teeth for a traversodontid As mentioned above Flynn et al(2000) noted that the homologies of these teeth were difficult todetermine Of the five anterior teeth preserved in the holotypethe anteriormost three are ldquoundoubtedly incisorsrdquo although I3 isldquounusually large and is caniniformrdquo (Flynn et al 2000423) Therobust recurved fourth tooth in the series was considered torepresent the canine with the subsequent tooth interpreted as anolder canine in the process of replacement Although FMNH PR2444 is edentulous the alveoli on the left side of the skull are wellpreserved and indicate an anterior tooth set identical to the ho-lotype The alveolar morphology indicates that the fifth toothwas canted anteriorly as in the holotype This suggests that themysterious fifth tooth is not the result of a transient period in thedental replacement history of M besairiei as suggested by Flynnet al (2000) but rather is the norm This leaves two possibilities1) M besairiei persistently had two canines This condition isunknown in cynodonts 2) The canine-like fourth tooth actuallyrepresents an incisor and the fifth tooth is the true canine Wefavor the latter explanation In FMNH PR 2444 the first fouralveoli are nearly identical in size and shape However the fifthalveolus while similar in anteroposterior length to its predeces-sors is strongly compressed labiolingually with an anteroposte-rior lengthlabiolingual width ratio of 044 (compared to 080 inthe fourth alveolus and 084ndash85 for I2ndash3) We interpret thismarked difference in alveolar ratio as indicating the separationbetween incisors and canine The canine-like I4 would then rep-resent the endpoint in a series of large increasingly caniniformincisors Supporting this interpretation recurved incisors are alsoobserved in juvenile Exaeretodon (eg MCP 3843 PV see Ab-dala et al 2002)
There are several legitimate differences between FMNH PR2444 and the holotype of Menadon besairiei but we take a con-servative taxonomic stance and argue that these differences rep-resent intraspecific variation and preservational artifacts FMNHPR 2444 represents a larger animal than UA 10601 The snoutlength (measured as the distance between the anterior edge ofthe orbit and a vertical plane through the anterior tip of thepremaxilla) of FMNH PR 2444 is 911 mm while that of UA10601 is 801 mm As such proportional differences between thetwo skulls may be attributable to ontogeny Variations in FMNHPR 2444 compared to the holotype that probably represent on-togentic differences include wider longer and deeper paraca-nine fossae a much deeper ventral zygomatic flange (althoughthe lower border of the flange is slightly abraded in the holotypeof Menadon besairiei there is no evidence that a major flangewas broken off) a proportionally shorter and more circular out-line to the posterior border of the maxilla below the zygomaticprocess and a more deeply emarginated posterior squamosal(ie a more prominent laterally projecting flange)
FMNH PR 2444 has a proportionally shorter more robustsnout and larger orbits than the holotype While these charactersare certainly variable in cynodonts it is unusual that the largerspecimen is shorter-snouted and larger-eyed contra the usualpattern The proportional differences between the two skulls areminor however (orbit diametersnout length is 042 for FMNHPR 2444 and 041 for the holotype) and are partially attributableto deformation (shear in FMNH PR 2444 and breakage of thepostorbital bar in the holotype) The following differences canalso probably be attributed to preservational artifact only fivepostcanine alveoli are preserved in FMNH PR 2444 while theholotype has eight postcanines The anteriormost three in theholotype however are tiny and peg-like As the area of themaxilla between the canine alveolus and the first preserved post-canine alveolus is poorly preserved in FMNH PR 2444 the ab-sence of these postcanines may be artifactual However it is alsopossible that the lower postcanine count of FMNH PR 2444compared to the holotype is an ontogenetic effect as postcaninecount is known to decrease with increasing skull size in Ex-
FIGURE 11 Left scapulocoracoids of FMNH PR 2444 A left scapu-locoracoid in medial view B left scapulocoracoid in lateral view Scalebars equal 5 cm
JOURNAL OF VERTEBRATE PALEONTOLOGY VOL 28 NO 2 2008456
Dow
nloa
ded
by [
UZ
H H
aupt
bibl
ioth
ek
Zen
tral
bibl
ioth
ek Z
uumlric
h] a
t 05
51 1
0 Se
ptem
ber
2013
FIGURE 12 Right pelvis of FMNH PR 2444 A stereopair of right pelvis in medial view B stereopair and interpretive drawing of right pelvis inlateral view Scale bars equal 5 cm Abbreviationsmdashbpp base of posterior process of ilium ib iliac blade IL ilium IS ischium ob f obturatorforamen P pubis
KAMMERER ET ALmdashCYNODONT FROM MADAGASCAR 457
Dow
nloa
ded
by [
UZ
H H
aupt
bibl
ioth
ek
Zen
tral
bibl
ioth
ek Z
uumlric
h] a
t 05
51 1
0 Se
ptem
ber
2013
aeretodon (Abdala et al 2002) The orbit of FMNH PR 2444 isless upwardly directed than that of the holotype (being at anangle of sim65deg to a sagittal plane through the midline of the skullas opposed to sim45deg in the holotype) This difference in orbitalangle is probably attributable to postmortem deformation as theskull of FMNH PR 2444 is noticeably sheared to the front leftThe appearance of a wider squarer incisor arcade and broader
anterior palate in FMNH PR 2444 can also be attributed to thisshear Finally the occipital region of FMNH PR 2444 is alto-gether taller and narrower than that of the holotype For ex-ample the foramen magnum in FMNH PR 2444 approximates arounded isosceles triangle in shape while the foramen magnumof the holotype is more ovoid These differences can also beattributed to deformation Altogether the differences between
FIGURE 13 Stereopairs of field number 8-31-98-387 an isolated right pelvis in A medial and B lateral view Scale bars equal 5 cm
JOURNAL OF VERTEBRATE PALEONTOLOGY VOL 28 NO 2 2008458
Dow
nloa
ded
by [
UZ
H H
aupt
bibl
ioth
ek
Zen
tral
bibl
ioth
ek Z
uumlric
h] a
t 05
51 1
0 Se
ptem
ber
2013
FMNH PR 2444 and the holotype fall within the range of intra-specific variation recorded for other gomphodont cynodontssuch as the traversodontids Massetognathus pascuali (Abdalaand Giannini 2000 Kammerer pers obs) and Exaeretodon(Abdala et al 2002) and the diademodontid Diademodon tetra-gonus (Grine et al 1978 Bradu and Grine 1979)
Phylogenetic Analysis
Utilizing the new information afforded by FMNH PR 2444and a reexamination of the holotype (UA 10601) Menadon be-sairiei was recoded in the Gondwanan gomphodont data matrixof Abdala et al (2006) Abdala et alrsquos (2006) data matrix en-compassed 18 taxa and 43 characters For the present analysis anadditional character ldquoincisor procumbencyrdquo (states 0-absent1-present) was added to Abdala et alrsquos (2006) data set (see
Appendix for coding of this character) Several traversodontidtaxa (eg lsquoScalenodonrsquo hirschsoni) have relatively large incisorsbut only in Menadon Scalenodontoides and Exaeretodon are theincisors procumbent Three of Abdala et alrsquos (2006) originalcodings for Menadon besairiei characters 0 (ldquonumber of upperincisorsrdquo) 5 (ldquoposition of paracanine fossa in relation to theupper caninerdquo) and 21 (ldquointernarial barrdquo) were changed to in-corporate data from the new specimen (see Appendix) Addi-tionally further examination of the holotype (UA 10601) ofMenadon besairiei allowed us to code characters 16ndash18 20 38and 40 (morphology of the lower postcanines all marked as ldquordquoin Abdala et alrsquos analysis) Parsimony searches were performedin PAUP v40b10 using a branch-and-bound search Addition-ally a bootstrap analysis (also using branch-and-bound) was per-formed using 1000 replicates
A total of 34 most parsimonious trees of length 103 werefound with the scores CI0573 RI0725 RC0415 andHI0427 The strict consensus tree (Fig 15A) did not recovera polytomy between Menadon Exaeretodon and Scalenodont-oides as in Abdala and Ribeiro (2003) or a (Menadon + Ex-aeretodon) sister-group relationship as in Abdala et al (2006)Rather Menadon was recovered as the sister taxon of the (Ex-aeretodon + Scalenodontoides) clade supporting Flynn et alrsquos(2000) original placement of this taxon There is strong bootstrapsupport (88) for Traversodontidae but within traversodontidsthe only strongly supported subclade is (Gomphodontosuchus +(Menadon + (Scalenodontoides + Exaeretodon))) with a boot-strap value of 80 Within this clade the (Menadon + (Scalen-odontoides + Exaeretodon)) grouping is supported by a boot-strap value of 83 and the (Scalenodontoides + Exaeretodon)grouping is supported by a bootstrap value of 94 (Fig 15B) Inaddition to the positioning of Menadon the strict consensus tree(Fig 15A) differs from the topology of Abdala et al (2006) inrecovering a sister-group relationship between Massetognathusand Santacruzodon rather than having Massetognathus andSantacruzodon as successive outgroups to the clade containingGomphodontosuchus Menadon Scalenodontoides and Ex-aeretodon The (Massetognathus + Santacruzodon) clade is diag-nosed by the absence of an anterolingual cusp in the upper post-canines denticulated incisor cutting margins and the presence ofan anterior cingulum in the upper postcanines
FIGURE 14 Reconstruction of the pelvis of Menadon besairiei basedon information from FMNH PR 2444 and field number 8-31-98-387Dotted area not preserved in either specimen reconstruction based onPascualgnathus (Bonaparte 1966) and Exaeretodon (Bonaparte 1963)
FIGURE 15 Cladogram of traversodontid relationships based on revised data set of Abdala et al (2006) showing A strict consensus tree and B50 majority rule tree Numbers at nodes of majority rule tree represent bootstrap values
KAMMERER ET ALmdashCYNODONT FROM MADAGASCAR 459
Dow
nloa
ded
by [
UZ
H H
aupt
bibl
ioth
ek
Zen
tral
bibl
ioth
ek Z
uumlric
h] a
t 05
51 1
0 Se
ptem
ber
2013
Relationships of Menadon besairiei
Menadon besairiei is the sister taxon to (Exaeretodon + Sca-lenodontoides) the latter clade diagnosed by the possession ofthree upper incisors a posteromedial position of the paracaninefossae relative to the canines the absence of an internarial barlarge canines a projected zygomatic process of the jugal and amaxilla that contributes to the margin of the subtemporal fenes-tra (the last three character states represent reversals) The clade(Menadon + (Exaeretodon + Scalenodontoides)) is diagnosed bythe possession of large procumbent incisors The presence offour incisors a primitive retention distinguishes Menadon be-sairiei from Exaeretodon Scalenodontoides and an unnamed tra-versodontid (ldquoTraversodontid Type 1rdquo) from the Santa MariaFormation of Brazil (Abdala et al 2001 Abdala and Ribeiro2002) Gomphodontosuchus is recovered as the sister taxon to(Menadon (Exaeretodon + Scalenodontoides)) as has beenfound in previous studies (Flynn et al 2000 Abdala and Ribeiro2003) The taxon name Gomphodontosuchinae Watson andRomer 1956 (ex Gomphodontosuchidae Watson and Romer1956 first given subfamilial rank in Lehman 1961) is availablefor this clade As circumscribed here Gomphodontosuchinaeconsists of the most inclusive clade containing Gomphodontosu-chus brasiliensis von Huene 1928264 but not Massetognathuspascuali Romer 19672
Menadon besairiei displays a mixture of primitive and derivedcharacters within traversodontids The internarial bar and fourincisors are primitive features whereas the enlargement and pro-cumbency of the incisors are derived features shared with Ex-aeretodon and Scalenodontoides The postcranium shows severalsimilarities to that of Exaeretodon (apparent exclusion of theprocoracoid from the glenoid some features of the pelvis tallcaudal neural spines) but in general displays a rather conserva-tive morphology observed throughout Gomphodontia The mor-phology of the scapulocoracoids is closer akin to diademodontidsor trirachodontids than Exaeretodon and the expanded thoracicrib of Menadon has only slight differences from those of trirach-odontids or early traversodontids like Luangwa The atlas-axiscomplex of Menadon is similar to that of the Early Triassic cyn-odont Cynognathus in possessing a concave axial spine with along posterior process Although most gomphodonts have beenreconstructed with a convex axial spine as in early mammals thisspine is broken in most of the best described gomphodont post-crania (eg Diademodon Brink 1955) and the lsquosaddle-shapedrsquoaxial morphology may be more widely distributed among cyn-odonts than previously realized The new specimens of Menadondescribed in this paper demonstrate that a generalized gompho-dont postcranial morphology was retained deep within the tra-versodontid tree and that the derived morphologies of either ofthe well-known taxa Massetognathus or Exaeretodon should notbe taken as representative of traversodontids in general
ACKNOWLEDGMENTS
This work would not have been possible without the continu-ing support and encouragement of our collaborative programwith the Deacutepartement de Paleacuteontologie et drsquoAnthropologie Bi-ologique Universiteacute drsquoAntananarivo Madagascar under the di-rection of Armand Rasoamiaramanana (Department Chairman)and Gisegravele Ravololonarivo Randria (Head of the PaleontologyLaboratories) We are extremely grateful to the National Geo-graphic Society for their long-term support of the fieldwork as-sociated with this project Additional financial or logistical sup-port for fieldwork was provided by The Field Museum the Uni-versity of California-Santa Barbara and the WWF (WorldwideFund for Nature Madagascar) We acknowledge the Ministegraverede LrsquoEneacutergie et des Mines for authorizing field efforts and for
continuing to facilitate our joint US-Madagascar explorationresearch and education program Steve Goodman and AsminaGandhi generously assisted us in innumerable ways includinghospitality nonpareil We especially thank R Andriantompoha-vana L Razafimanantsoa A Toto Volahy JA Rabarison AGandie and P Vavisaro for assistance in the field during theexpedition in which these new specimens were discovered BillSimpson and Gina Wesley-Hunt for their patient excavation ofthis important material Allison Smith Ray Vodden Lisa Berg-wall and Deborah Wagner for excellent preparation of a chal-lenging specimen Jennifer Botha and Elize Butler (NationalMuseum Bloemfontein) Michael Raath and Fernando Abdala(Bernard Price Institute) Thomas Kemp (Oxford UniversityMuseum of Natural History) Ray Symonds (University Museumof Zoology Cambridge) Bernard Battail (Museacuteum nationaldrsquoHistoire naturelle Paris) Michael Maisch (Universitaumlt Tuumlbin-gen) and Charles Schaff and Farish Jenkins (Museum of Com-parative Zoology Harvard) for access to specimens and AgustiacutenMartinelli Fernando Abdala and James Hopson for helpful re-views of an earlier version of the manuscript
LITERATURE CITED
Abdala F M C Barbarena and J Dornelles 2002 A new species of thetraversodontid cynodont Exaeretodon from the Santa Maria Forma-tion (MiddleLate Triassic) of Southern Brazil Journal of Verte-brate Paleontology 22313ndash325
Abdala F and N P Giannini 2000 Gomphodont cynodonts of theChantildeares Formation the analysis of an ontogenetic sequence Jour-nal of Vertebrate Paleontology 20501ndash506
Abdala F J Neveling and J Welman 2006 A new trirachodontidcynodont from the lower levels of the Burgersdorp Formation(Lower Triassic) of the Beaufort Group South Africa and the cla-distic relationships of Gondwanan gomphodonts Zoological Jour-nal of the Linnean Society 147383ndash413
Abdala F A M Ribeiro and C L Schultz 2001 A rich cynodont faunaof Santa Cruz do Sul Santa Maria Formation (Middle-Late Trias-sic) southern Brazil Neues Jahrbuch fuumlr Geologie und Palaumlontolo-gie Monatshefte 2001669ndash687
Abdala F and A M Ribeiro 2002 Nuevos cinodontes traversodoacuten-tidos (Synapsida-Eucynodontia) de la Formacioacuten Santa Maria(Triaacutesico Medio-Superior) Rio Grande do Sul Brasil RevistaEspantildeola de Paleontologiacutea 17237ndash243
Abdala F and A M Ribeiro 2003 A new traversodontid cynodontfrom the Santa Maria Formation (Ladinian-Carnian) of southernBrazil with a phylogenetic analysis of Gondwanan traversodontidsZoological Journal of the Linnaean Society 139529ndash545
Abdala F and A M S Teixeira 2004 A traversodontid cynodont ofAfrican affinity in the South American Triassic Palaeontologia Af-ricana 4011ndash22
Bonaparte J F 1962 Descripcioacuten del craacuteneo y mandiacutebula de Ex-aeretodon frenguellii Cabrera y su comparacioacuten con Diademodon-tidae Tritylodontidae y los cinodontes sudamericanos Publica-ciones del Museo Municipal de Ciencias Naturales y Tradicional deMar del Plata 1135ndash202
Bonaparte J F 1963 Descripcioacuten del esqueleto postcraneano de Ex-aeretodon (CynodontiandashTraversodontidae) Acta Geoloacutegica Lil-loana 45ndash52
Bonaparte J F 1966 Una nueva ldquofaunardquo Triaacutesica de Argentina (Ther-apsida Cynodontia Dicynodontia) Consideracions filogeneacuteticas ypaleobiogeograacuteficas Ameghiniana 8243ndash296
Bradu D and F E Grine 1979 Multivariate analysis of diademodon-tine crania from South Africa and Zambia South African Journal ofScience 79441ndash448
Brink A S 1955 A study of the skeleton of Diademodon Palaeonto-logia Africana 33-39
Brink A S 1963 Two new cynodonts from the Ntawere Formation inthe Luangwa Valley of Northern Rhodesia Palaeontologia Africana877ndash96
Broom R 1905 On the use of the term Anomodontia Records of theAlbany Museum 1266ndash269
Cabrera A 1943 El primer hallazgo de teraacutepsidos en la ArgentinaNotas del Museo de La Plata Paleontologiacutea 55317ndash331
JOURNAL OF VERTEBRATE PALEONTOLOGY VOL 28 NO 2 2008460
Dow
nloa
ded
by [
UZ
H H
aupt
bibl
ioth
ek
Zen
tral
bibl
ioth
ek Z
uumlric
h] a
t 05
51 1
0 Se
ptem
ber
2013
Chatterjee S 1982 A new cynodont reptile from the Triassic of IndiaJournal of Paleontology 56203ndash214
Crompton A W 1955 On some Triassic cynodonts from TanganyikaProceedings of the Zoological Society of London 125617ndash669
Crompton A W 1972 Postcanine occlusion in cynodonts and tritylo-donts Bulletin of the British Museum (Natural History) Geology2129ndash71
Crompton A W and F Ellenberger 1957 On a new cynodont from theMolteno Beds and the origin of the tritylodontids Annals of theSouth African Museum 441ndash13
Flynn J J J M Parrish B Rakotosaminimanana W F Simpson R LWhatley and A R Wyss 1999 A Triassic fauna from Madagascarincluding early dinosaurs Science 286763ndash765
Flynn J J J M Parrish B Rakotosamimanana L RanivoharimananaW F Simpson and A R Wyss 2000 New traversodontids (Syn-apsida Eucynodontia) from the Triassic of Madagascar Journal ofVertebrate Paleontology 20422ndash427
Gow C E and P J Hancox 1993 First complete skull of the LateTriassic Scalenodontoides (Reptilia Cynodontia) from southern Af-rica In Lucas S G and M Morales (eds) The nonmarine TriassicNew Mexico Museum of Natural History amp Science Bulletin 3161ndash168
Grine F E B D Hahn and C E Gow 1978 Aspects of relativegrowth and variability in Diademodon (Reptilia Therapsida) SouthAfrican Journal of Science 7450ndash58
Hopson J A 1984 Late Triassic traversodont cynodonts from NovaScotia and southern Africa Palaeontologia Africana 25181ndash201
Hopson J A 1985 Morphology and relationships of Gomphodontosu-chus brasiliensis von Huene (Synapsida Cynodontia Tritylodontoi-dea) from the Triassic of Brazil Neues Jahrbuch fuumlr Geologie undPalaumlontologie Monatshefte 1985285ndash299
Hopson J A and J W Kitching 2001 A probainognathian cynodontfrom South Africa and the phylogeny of nonmammalian cynodontsBulletin of the Museum of Comparative Zoology 1565ndash35
Hopson J A and H-D Sues 2006 A traversodont cynodont from theMiddle Triassic (Ladinian) of Baden-Wuumlrttemberg (Germany)Palaumlontologische Zeitschrift 80124ndash129
Huene F von 1928 Ein Cynodontier aus der Trias Brasiliens Zentral-blatt fuumlr Mineralogie Geologie und Palaumlontologie 1928251ndash270
Huene F von 1936 Die fossilen Reptilien des suumldamerikanischenGondwanalandes Ergebnisse der Sauriergrabungen in Suumldbrasilien192829 Lieferung 2 (Cynodontia)93ndash159 Franz F Heine Tuumlbin-gen
Jenkins F A 1970 The Chantildeares (Argentina) Triassic reptile faunaVII The postcranial skeleton of the traversodontid Massetognathuspascuali (Therapsida Cynodontia) Breviora 3521ndash28
Jenkins F A 1971 The postcranial skeleton of African cynodonts prob-lems in the early evolution of the mammalian postcranial skeletonBulletin of the Peabody Museum of Natural History 361ndash216
Kemp T S 1969 The atlas-axis complex of mammal-like reptiles Jour-nal of Zoology 159223ndash248
Kemp T S 1980 Aspects of the structure and functional anatomy of theMiddle Triassic cynodont Luangwa Journal of Zoology 191193ndash239
Kemp T S 1982 Mammal-Like Reptiles and the Origin of MammalsAcademic Press New York
Lehman J-P 1961 Cynodontia pp 140ndash191 in Piveteau J (ed) Traiteacutede Paleacuteontologie Masson et Cie Paris
Osborn 1903 On the primary division of the Reptilia into two sub-classes Synapsida and Diapsida Science 17275ndash276
Owen R 1861 Palaeontology or a systematic summary of extinct ani-mals and their geological relations 2nd Ed Edinburgh
Parrington F R 1934 On the cynodont genus Galesaurus with a noteon the functional significance of the changes in the evolution of thetheriodont skull Annals and Magazine of Natural History 10(13)38ndash67
Parrington F R 1946 On the cranial anatomy of cynodonts Proceed-ings of the Zoological Society of London 116181ndash197
Razafimbelo E 1987 Le basin de Morondava (Madagascar) Synthegravesegeacuteologique et structurale PhD dissertation University Louis Pas-teur Strasbourg France 189 pp
Reisz R R and H-D Sues 2000 Herbivory in Late Paleozoic andTriassic terrestrial vertebrates pp 9ndash41 in Sues H-D (ed) Evo-lution of Herbivory in Terrestrial Vertebrates Cambridge Univer-sity Press New York
Romer A S 1967 The Chantildeares (Argentina) Triassic reptile fauna IIITwo new gomphodonts Massetognathus pascuali and M terugiiBreviora 2641ndash25
Romer A S 1972 The Chantildeares (Argentina) Triassic reptile faunaXVII The Chantildeares gomphodonts Breviora 3961ndash9
Rougier G W J R Wible and J A Hopson 1992 Reconstruction ofthe cranial vessels in the Early Cretaceous mammal Vincelestes neu-quenianus implications for the evolution of the mammalian cranialvascular system Journal of Vertebrate Paleontology 12188ndash216
Rowe T 1988 Definition diagnosis and origin of Mammalia Journal ofVertebrate Paleontology 8241ndash264
Rowe T 1993 Early mammal phylogenetic systematics pp 129ndash145 inSzalay F S M J Novacek and M C McKenna (eds) MammalPhylogeny Vol 1 Mesozoic Differentiation MultituberculatesMonotremes Early Therians and Marsupials Springer-Verlag NewYork
Seeley H G 1894 Researches on the structure organization and clas-sification of the fossil Reptilia Part IX Section 3 On DiademodonPhilosophical Transactions of the Royal Society of London 1851029ndash1041
Seeley H G 1895 Researches on the structure organization and clas-sification of the fossil Reptilia Part IX Section 4 On the Gompho-dontia Philosophical Transactions of the Royal Society of London1861ndash57
Seeley H G 1895 Researches on the structure organization and clas-sification of the fossil Reptilia Part IX Section 5 On the skeletonin new Cynodontia from the Karroo rocks Philosophical Transac-tions of the Royal Society of London 18659ndash148
Sues H-D J A Hopson and N H Shubin 1992 Affinities of Sca-lenodontoides plemmyridon Hopson 1984 (Synapsida Cynodontia)from the Upper Triassic of Nova Scotia Journal of Vertebrate Pa-leontology 12168ndash171
Sues H-D and P E Olsen 1990 Triassic vertebrates of Gondwananaspect from the Richmond basin of Virginia Science 2491020ndash1023
Swofford D L 2003 PAUP v 40b10 Sinauer Associates Inc Pub-lishers Sunderland Massachusetts
Watson D M S and A S Romer 1956 A classification of therapsidreptiles Bulletin of the Museum of Comparative Zoology 11435ndash89
Submitted September 20 2007 accepted November 3 2007
KAMMERER ET ALmdashCYNODONT FROM MADAGASCAR 461
Dow
nloa
ded
by [
UZ
H H
aupt
bibl
ioth
ek
Zen
tral
bibl
ioth
ek Z
uumlric
h] a
t 05
51 1
0 Se
ptem
ber
2013
APPENDIX Data matrix used in the phylogenetic analysesThis is a revised version of the matrix of Abdala et al (2006) with the following changes character 43 [Incisor procumbency (0) absent (1) present]added Menadon recoded for characters 0 (Number of upper incisors was 1 ldquothreerdquo changed to 0 ldquofourrdquo) 5 (Position of paracanine fossae in relationto the upper canine was changed to 1 ldquomedialrdquo) 16 (Overall morphology of lower postcanines in occlusal view was changed to 3 ldquoquadran-gularrdquo) 17 (Position of the transverse crest in lower postcanines was changed to 1 ldquoanteriorrdquo) 18 (Number of cusps in the transverse crest of thelower postcanines was changed to 0 ldquotwordquo) 20 (Anterior cingulum in the lower postcanines was changed to 2 ldquoabsentrdquo) 21 (Internarial barwas 1 ldquoabsentrdquo changed to 0 ldquopresentrdquo) 38 (Posterior cingulum in the lower postcanines was changed to 1 ldquoabsentrdquo) and 40 (Deep occlusalbasins in the postcanines was changed to 1 ldquopresentrdquo) Langbergia recoded for character 34 (Incisor cutting margins was 0 ldquoserratedrdquo changedto 1 ldquosmoothly ridgedrdquo) based on the description of the incisors in Abdala et al (2006) Gomphodontosuchus recoded for character 31 (Maxilla inthe margin of the subtemporal fenestra was 1 ldquoincludedrdquo changed to 0 ldquoexcludedrdquo) based on von Huene (1936) and Kammerer (pers obs of theholotype) contra Hopsonrsquos (1985) reconstruction lsquoScalenodonrsquo hirschsoni recoded for character 39 (Anterior cingulum in the upper postcanines was1 ldquoabsentrdquo changed to 0 ldquopresentrdquo) based on Abdala (pers comm 2006) Trirachodon recoded for character 30 (Ectopterygoid was changedto 0 ldquopresentrdquo) based on Abdala (pers comm 2006) and Traversodon recoded for character 15 (Number of cusps in the sectorial border of the upperpostcanines was 0 ldquomore than twordquo changed to 1 ldquotwordquo) based on Abdala (pers comm 2006) and Kammerer (pers obs)
1111111111 2222222222 3333333333 44440123456789 0123456789 0123456789 0123456789 0123
Cynognathus 00000000 000 000000000 00000000 000Diademodon 0000001000 0000100 000010A000 0000000100 0100Trirachodon 000000100B 1A002201 001A100010 0001001200 0110Langbergia 0000001000 10000201 0000101010 0001101200 0110Cricodon 0100001000 11000201 0010100 0001200 0110Andescynodon 0000002001 01013101 101120010 0112311 1100Massetognathus 00111121A2 1110003100 2011200011 1011212300 1100Exaeretodon 1110122112 010103101 2111110201 1111112311 1101Luangwa 0000001002 11010A3100 101010010 01002300 1100Scal angustifrons 000000100B 1111023100 10102101 00002301 1100Scal hirschsoni 11110202 1100113100 1010 1112300 1100Traversodon 0000002012 1101113100 201010 011230 1100Gomphodontosuchus 0011112112 0103101 2010 0112311 1100Pascualgnathus 1000001000 000231 1100001 0011230 1100Scalenodontoides 1112212 01003101 21111021 1212311 1101Menadon 0111112112 01012310 20121020 0102311 1101Dadadon 00002112 111112 013 11231 1100Santacruzodon 00002112 1110003101 201300 212300 1100
A(01) B(02)
JOURNAL OF VERTEBRATE PALEONTOLOGY VOL 28 NO 2 2008462
Dow
nloa
ded
by [
UZ
H H
aupt
bibl
ioth
ek
Zen
tral
bibl
ioth
ek Z
uumlric
h] a
t 05
51 1
0 Se
ptem
ber
2013
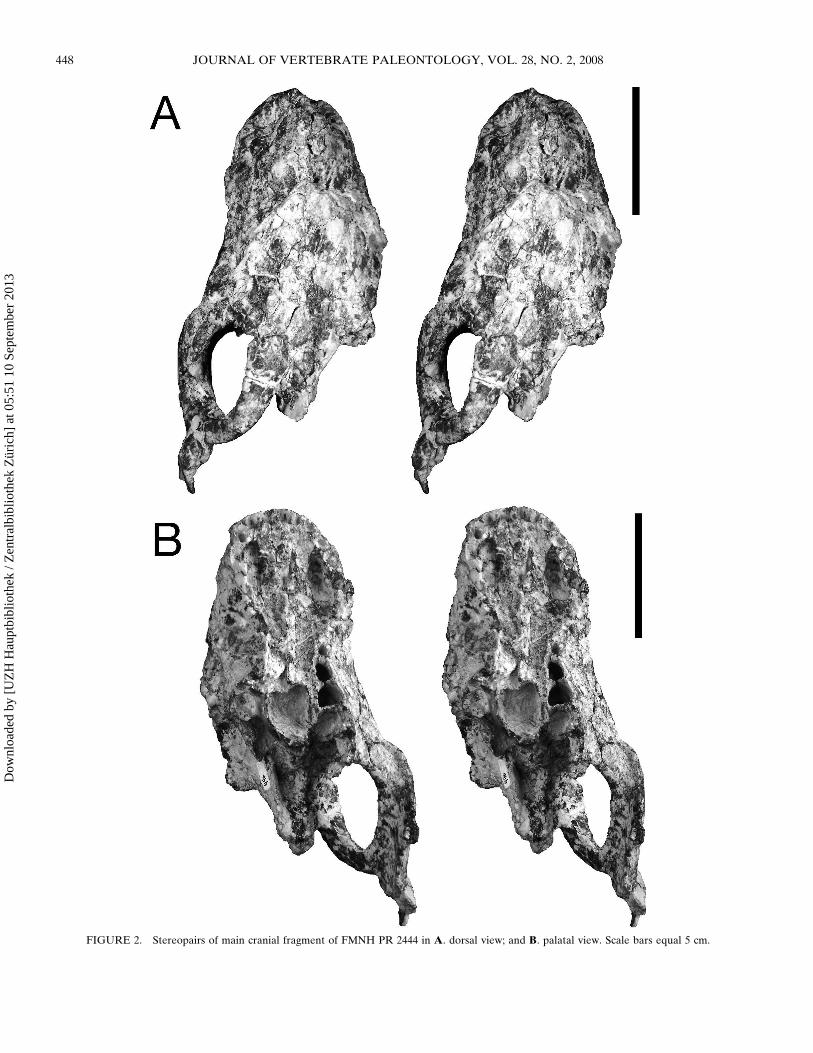
FIGURE 2 Stereopairs of main cranial fragment of FMNH PR 2444 in A dorsal view and B palatal view Scale bars equal 5 cm
JOURNAL OF VERTEBRATE PALEONTOLOGY VOL 28 NO 2 2008448
Dow
nloa
ded
by [
UZ
H H
aupt
bibl
ioth
ek
Zen
tral
bibl
ioth
ek Z
uumlric
h] a
t 05
51 1
0 Se
ptem
ber
2013
sairiei have equal transverse and anteroposterior dimensions Attheir maximum depth in Thrinaxodon the intercentra barely ex-tend beyond the ventral margins of the cervical pleurocentra InFMNH PR 2444 IC4 extends for half of its total depth below theventral margins of the cervical pleurocentra In IC3 this condi-tion is even more extreme extending below the ventral marginsof the cervical pleurocentra for 80 of its total depth The shapeand position of the intercentra give a strongly arched appearanceto the ventral profile of elements in the cervical series The ven-tral surfaces of the intercentra bear a posteriorly directed process
on each side In IC3 a weak ridge originates between the pairedprocesses and extends posteriorly indicating muscle attachmenton the intercentra The condition in IC4 cannot be determinedbecause of incomplete preservation Although only three inter-centra are known ventral facets in the centra of the isolatedcervicals indicate that others were present
As in other cynodonts the atlas-axis complex of FMNH PR2444 consists of a pair of separate atlantal neural arches articu-lating with the occipital condyles and the atlas centrum Only theleft atlas arch is preserved in this specimen (Fig 7) It is similar
FIGURE 3 Stereopair and interpretive drawing of anterior palatal region of FMNH PR 2444 Scale bar equals 5 cm AbbreviationsmdashC canineI incisor i f incisive foramina PC postcanine pc f paracanine fossa Postcanine numbering based on comparison with the holotype UA 10601
FIGURE 4 Cranial fragments of FMNH PR 2444 A stereopair of right zygomatic arch in lateral view Scale bar equals 5 cm B stereopair of rightzygomatic arch in medial view Scale bar equals 5 cm C stereopair of left squamosal in lateral view Scale bar equals 5 cm D sagittal crest in rightlateral view Scale bar equals 1 cm E occiput in occipital view Scale bar equals 1 cm
KAMMERER ET ALmdashCYNODONT FROM MADAGASCAR 449
Dow
nloa
ded
by [
UZ
H H
aupt
bibl
ioth
ek
Zen
tral
bibl
ioth
ek Z
uumlric
h] a
t 05
51 1
0 Se
ptem
ber
2013
to the atlantal neural arches described for other cynodonts suchas Massetognathus (Kemp 1969 Jenkins 1971) There is a me-dial facet for articulation with the centrum of the atlas (theldquoodontoid facetrdquo of Kemp 1969) and an anterior facet for ar-ticulation with the occipital condyle (Fig 7B) A pair of pro-cesses dominates the atlas arch a dorsal lamina (a roughly rect-angular plate of bone arching above the facets) and a largeposterolaterally directed transverse process An oval tubercle atthe anterior end of the transverse process represents the facet forthe proatlas
The centrum of the atlas is fused to the axis forming a singlelarge vertebral unit (Fig 6) A weak odontoid process is presentat the dorsal limit of the anterior surface of the atlas The ante-rior aspect of the centrum is dominated by a roughly lsquoTrsquo-shapedstructure composed of the two facets for articulation with theatlas arches and the facet for articulation with the atlas intercen-trum The axis bears no prezygapophyses and as such does notarticulate with the atlas arch as in Early Triassic cynodonts suchas Galesaurus (Parrington 1934) There appears to be a trendtowards loss of the axial prezygapophyses in later traversodon-
tids which are present but vestigial in Massetognathus (Jenkins1970 although in the specimens described the prezygapophysesare broken so their reduction is perhaps questionable) and ab-sent in Exaeretodon (Bonaparte 1963) The transverse processesof the axis are large and robust compared to those of the othercervicals The axial spine is saddle-shaped with a concave dorsalprofile unlike previous reconstructions of the traversodontidaxis which depict a straight or convex dorsal profile similar tothat observed in galesaurid cynodonts and early mammals (Jen-kins 1971) The morphology of the axial spine is similar to thatof the Early Triassic cynodont Cynognathus (Seeley 1895) InCynognathus however the neural spine of the third cervical(C3) is tall and canted posteriorly behind the posterior process ofthe axial spine whereas in FMNH PR 2444 the short C3 neuralspine fits below the posterior portion of the axial spine Theposterior process of the axial spine bears a pair of parallel elon-gate grooves on its underside attachment sites for the axial mus-culature
Post-Axial VertebraemdashThree post-axial cervicals are knownfor FMNH PR 2444 (Figs 6 8AndashD) The first of these C3 is
FIGURE 5 Stereopair of ventral view of the occipital fragment of FMNH PR 2444 showing the basicranium Anterior is up
FIGURE 6 Stereopair and interpretive drawing of atlas-axis complex and third cervical of FMNH PR 2444 in left lateral view Scale bar equals5 cm Abbreviationsmdasha n sp axial neural spine d dens IC intercentrum f at a facet for articulation with the atlas arch p parapophysis pozpostzygapophysis t p transverse process
JOURNAL OF VERTEBRATE PALEONTOLOGY VOL 28 NO 2 2008450
Dow
nloa
ded
by [
UZ
H H
aupt
bibl
ioth
ek
Zen
tral
bibl
ioth
ek Z
uumlric
h] a
t 05
51 1
0 Se
ptem
ber
2013
preserved in articulation with the axis (Fig 6) The two isolatedcervicals bear successively taller and anterposteriorly wider neu-ral spines (Fig 8AndashD) Based on their staggered size and failureto articulate we suggest that they represent C5 and C7 (assumingthat this specimen possessed the generalized cynodont cervicalnumber of seven) The cervicals display robust transverse pro-cesses which are directed laterally and slightly anteriorly A par-apophysis lies below the anterior edge of the transverse processalong the antero-lateral edge of the centrum No cervical ribs arepreserved but their presence can be inferred from the par-apophyses The notochordal concavity is as in Massetognathussurrounded by a raised rim at each end of the centrum (Jenkins1970) No anapophyses are present The neural spines for allthree post-axial cervicals are tapering and slightly curved poste-riorly The neural spine in C3 is relatively short and wide(thereby fitting underneath the posterior overhang of the axialspine see Fig 6) while that of C5 is tall and narrow The spinefor C7 is taller and anteroposteriorly wider than that of C5 andthe dorsal edge is more rounded similar to the condition in thedorsal vertebrae
Eight post-cervical presacral vertebrae are known for thisspecimen We will refer to them here simply as dorsal vertebraegiven the incompleteness of the vertebral column in the speci-men and the uncertainty concerning the thoraciclumbar transi-tion in this taxon However it is probable that they representthoracic vertebrae (see below) In most cynodonts the lumbar
vertebrae are synostosed to specialized ribs with overlappingelements In earlier taxa (eg Thrinaxodon Cynognathus Lu-angwa Jenkins 1971 Kemp 1980) expanded costal plates arepresent on both thoracic and lumbar ribs In Massetognathus thelumbar ribs consist of a pair of prongs one directed anteriorlybearing a facet for articulation with the previous rib and theother directed posteriorly (Jenkins 1970) In Exaeretodon allpresacral ribs are identical of simple morphology and not syn-ostosed to the vertebrae (Bonaparte 1963) A single isolated rib(Fig 9B) in FMNH PR 2444 demonstrates that some of the ribsin Menadon besairiei were expanded However all preservedvertebrae in this specimen have facets for articulation with theribs rather than breakage points where they would have beenfused to ribs As such it is probable that all of the vertebraepreserved in this specimen represent thoracics although we can-not discount the possibility that as in Exaeretodon no presacralvertebrae were fused to ribs
The neural spines in all of the dorsal vertebrae are elongateposteriorly inclined and roughly rectangular in lateral view witha rounded laterally expanded platform dorsally (Fig 8EndashJ)Costal foveae are present at the anterior and posterior edges ofeach centrum Of the eight dorsal vertebrae there are two ar-ticulated sets and three isolated elements (two from 9-4-98-440and one from 9-2-98-422) The first isolated vertebra (Fig 8E F)has a taller neural spine and shorter centrum than the otherdorsal vertebrae It is most similar in proportions to the isolated
FIGURE 7 Stereopairs and interpretive drawings of left atlas arch of FMNH PR 2444 in A lateral view Anterior is left B medial view Anterioris right Scale bars equal 1 cm Abbreviationsmdashafa anterior facet for articulation with the occipital condyle dl dorsal lamina mfa medial facet forarticulation with the atlas centrum (odontoid facet) pfa tubercle forming the facet for articulation with the proatlas tp transverse process
KAMMERER ET ALmdashCYNODONT FROM MADAGASCAR 451
Dow
nloa
ded
by [
UZ
H H
aupt
bibl
ioth
ek
Zen
tral
bibl
ioth
ek Z
uumlric
h] a
t 05
51 1
0 Se
ptem
ber
2013
FIGURE 8 Vertebrae of FMNH PR 2444 Cervical vertebra 5 in A left lateral and B anterior views Cervical vertebra 7 in C left lateral andD anterior views Isolated dorsal vertebra from anterior region of the series in E left lateral and F anterior views G stereopair and interpretivedrawing of articulated set of two dorsal vertebrae from near the midpoint of the series in left lateral view Anterior is up H articulated set of threedorsal vertebrae from anterior region of the series in left and right lateral views I isolated vertebra from near the midpoint of the series (from fieldnumber 9-2-98-422) J isolated vertebra from posterior region of the series Sacral vertebra in K dorsal view (the distal region of another sacral ribwhich is identical in shape to that of the intact sacral rib albeit slightly larger is also shown in this figure) and L left lateral view Caudal vertebraeM articulated set of two vertebrae from the anterior region of the series showing the lsquowingrsquo-shaped caudal ribs in dorsal view Anterior is left Nisolated caudal vertebra showing tall neural spine in left lateral view O isolated caudal vertebra showing tall neural spine in posterior view Particulated set of four vertebrae from near the end of the caudal series in dorsal view Anterior is left Scale bars equal 1 cm Abbreviationsmdasha cof anterior costal fovea n sp neural spine p co f posterior costal fovea poz postzygapophysis prz prezygapophysis
JOURNAL OF VERTEBRATE PALEONTOLOGY VOL 28 NO 2 2008452
Dow
nloa
ded
by [
UZ
H H
aupt
bibl
ioth
ek
Zen
tral
bibl
ioth
ek Z
uumlric
h] a
t 05
51 1
0 Se
ptem
ber
2013
cervicals and was probably located near the front of the dorsalseries The first articulated set consists of three vertebrae (Fig8H) The middle vertebra in this set is broken from the anteriorbase of the neural spine to the ventral midpoint and most of theright side of the anterior half of the centrum is missing This setof vertebrae probably also came from the anterior region of theseries as their neural spines are longer and narrower than allvertebrae other than the one described above The second ar-ticulated set (Fig 8G) consists of two well-preserved vertebraethat based on comparisons with Massetognathus and Ex-aeretodon (Jenkins 1970 Bonaparte 1963) probably came fromnear the midpoint of the dorsal series The second isolated ver-tebra (Fig 8I) from 9-2-98-422 is similar to the members of thesecond set The third isolated vertebra (Fig 8J) appears to befrom the posterior end of the series based on the relatively shortneural spine and reduced posterior costal fovea
One sacral vertebra (Fig 8K L) is known consisting of awell-preserved centrum neural spine and the right sacral rib(the left is broken off) The centrum is shorter (both anteropos-teriorly and dorsoventrally) than those of the dorsal vertebraeand the neural spine is shorter and more rounded in profileAlso the neural spine is directed vertically unlike those of thedorsal vertebrae which are canted posteriorly The zygapophy-ses are poorly developed The sacral rib is fused to a singlesynapophysis and projects anterolaterally The rib narrows to-wards the middle and expands distally terminating in a widerectangular facet where it would meet the ilium (Fig 8K)
Seven caudal vertebrae are known The caudal vertebrae aredistinctive in having very tall narrow neural spines Some EarlyTriassic cynodonts possess blunt neural spines of moderateheight on the proximal caudals (see for example Thrinaxodonin Jenkins 1971 and Diademodon in Brink 1955) Massetogna-thus represents an extreme condition wherein the caudal neuralspines are greatly reduced (Jenkins 1970) Very tall neuralspines are seen in Exaeretodon (Bonaparte 1963) and thosespines are unusual in that they do not diminish in height until thelast caudals The caudal vertebrae of FMNH PR 2444 consist ofa pair of articulated vertebrae most likely from near the sacrum(Fig 8M) an isolated caudal lacking postzygapophyses and ribsbut with the neural spine (Fig 8N O) and a quartet of smaller
articulated elements (Fig 8P) The anterior caudals have nar-rower neural spines and shorter zygapophyses than the subse-quent vertebrae in the series The caudal ribs attached to thispair have a crescentic distal region that curves anteriorly pro-ducing a lsquowingrsquo-shape Only a portion of the right rib of theisolated caudal is preserved but the lsquowingrsquo-shaped morphologyof the near-sacral caudal ribs does not appear to have been pre-sent In the posteriormost four vertebrae caudal ribs are pre-served on the left side of the first and the right side of the secondand third members On the first of these vertebrae the rib isshort curved posteriorly (but not lsquowingrsquo-shaped) and taperingRibs of the second and third vertebrae are less curved resem-bling posteriorly directed rods The zygapophyses of both theisolated caudal vertebra and the quartet of small caudals areanteroposteriorly elongate becoming increasingly so posteriorly
Thoracic RibsmdashNine thoracic ribs are known from 9-4-98-440(four left four right and an indeterminate splint of bone Fig9A indeterminate bone not illustrated) These elements re-semble the flattened ribs lacking costal plates of Exaeretodon(Bonaparte 1963) and the anterior thoracic ribs of Massetogna-thus and Luangwa (Jenkins 1970 Kemp 1980) The tuberculumand capitulum are separated by a shallow depression The tuber-culum has a shorter base than the capitulum but their proximalfaces are subequal in size An additional isolated left rib (Fig9B) from 9-2-98-422 is expanded indicating that unlike Ex-aeretodon Menadon rib morphology was heterogeneous alongthe length of the dorsal series This rib closely resembles theposterior thoracic ribs described for Luangwa (Kemp 1980fig16) Pascualgnathus (Bonaparte 1966fig 7) and the trirach-odontid Cricodon (Crompton 1955 Fig 4) This rib is con-stricted at the base of the head then expands distally with an-terior and posterior costal plates As in Luangwa the anteriorcostal plate is shallow and evenly sloping proximally and distallywhile the posterior costal plate is falcate with a concave edgedistally The distal expansion of this rib occurs at an angle of sim30degto the long axis of the rib head Despite their overall similaritythe expanded rib in FMNH PR 2444 does exhibit some differ-ences from the posterior thoracic rib of Luangwa The expandedrib of Menadon has a longer shaft between the rib head and thecostal plates and a shorter capitulum lacks a ridge on the pos-
FIGURE 9 A anterior thoracic ribs of FMNH PR 2444 in anterior view (indeterminate rib not figured) B expanded posterior thoracic rib ofFMNH PR 2444 in anterior view Scale bars equal 1 cm
KAMMERER ET ALmdashCYNODONT FROM MADAGASCAR 453
Dow
nloa
ded
by [
UZ
H H
aupt
bibl
ioth
ek
Zen
tral
bibl
ioth
ek Z
uumlric
h] a
t 05
51 1
0 Se
ptem
ber
2013
terior costal plate and has an anteroposteriorly shorter anteriorcostal plate Additionally the posterior margin of the costal plateis less curved than that of Luangwa These differences couldindicate that the expanded rib of Menadon is from a more pos-terior position in the series than the rib from Luangwa as re-duced curvature and increasing confluence of the tuberculumand capitulum are characteristic of lumbar ribs in traversodon-tids (Jenkins 1970 Kemp 1980) However the length of theshaft is at odds with this interpretation and in Cricodon andPascualgnathus the more posterior ribs are not more similar tothe element from FMNH PR 2444 These differences probablyrepresent legitimate morphological distinctions between the ribsof earlier gomphodonts and Menadon The head of the expandedrib is well defined and unbroken indicating that it was not syn-ostosed to a vertebra
Appendicular Skeleton
The appendicular skeleton is represented by both scapulocora-coids and a fragment of a clavicle from field number 9-4-98-440and a partial right pelvis from field number 9-2-98-422 An ad-ditional partial right pelvis is represented from field number8-31-98-387 The scapulocoracoids (Figs 10 11) are very similarto those described for other traversodontids as well as cynog-nathids and diademodontids (Jenkins 1970 1971) The scapularinfraspinous fossa has out-turned anterior and posterior edgesThe anterior edge of the scapula is rounded and curves laterallyforming the scapular spine At the lower end of the spine theacromion forms a prominent process (broken in the left scapula)Below this process the scapula is constricted and then expandsventrally to form the dorsal margin of the articular glenoid Theglenoid fossa is in general morphology very similar to that ofother Triassic cynodonts (Jenkins 1970 1971 Kemp 1980) It isposteroventrally oriented with roughly equal contributions fromthe scapular and coracoid glenoid facets The coracoid glenoidfacet is roughly triangular while the scapular glenoid facet issemiovoid as in other cynodonts In anterior or posterior viewthe fossa appears smoothly crescentic The procoracoid is aslightly curved trapezoidal plate of bone It does not appear tocontribute to the glenoid fossa The procoracoid foramen is lo-cated midway up the height of the procoracoid at a point directlybelow the base of the acromion process The coracoid is semi-crescentic curving posteriorly from a broad base The coracoidin Menadon is longer and more falcate than those described forExaeretodon (Bonaparte 1963) and Luangwa (Kemp 1980 al-though the distal tip of both coracoids is broken in the specimenof Luangwa [TSK 121] described by Kemp and may have beenlonger than reconstructed) but agrees closely with those of Mas-setognathus and cynognathidsdiademodontids (Jenkins 19701971) The distal coracoid tubercle for attachment to the tricepsis strongly curved upwards similar to the condition in Masseto-gnathus (Jenkins 1970) and Diademodon (Brink 1955)
A small fragment of bone resembling a curved cylinder almostcertainly represents part of a clavicle It has no distinguishingfeatures other than a groove associated with a ridge on one sideHowever the degree of curvature observed precludes this ele-ment from representing a long bone and the circular cross-section excludes it from being a rib
The pelvis of FMNH PR 2444 (Fig 12) resembles those ofother traversodontids in general morphology The ilium bears anarrow anteriorly directed iliac blade curving outward distallyThe anterior edge of the iliac blade slopes upward at a roughly45deg angle similar to Luangwa (Kemp 1980) but unlike Ex-aeretodon in which the anterior edge of the iliac blade curvesbackwards (Bonaparte 1963) or Pascualgnathus and Masseto-gnathus in which it is rounded (Bonaparte 1966 Jenkins 1970)The supraacetabular buttress at the base of the ilium is located inan anterodorsal position relative to the acetabulum as in other
traversodontids but unlike earlier cynodonts in which it is lo-cated dorsally (Jenkins 1970) The acetabular buttresses of thethree pelvic bones are largely confluent forming a fairly uniformlsquoliprsquo around the acetabulum as in Exaeretodon (Bonaparte1963) On the contrary in Luangwa the supraacetabular ischialand pubic buttresses are highly distinct spaced 6ndash8 mm apart andwithout confluence of their acetabular facets The pelvis ofFMNH PR 2444 does have a small depression between the su-praacetabular and ischial buttresses (also present in Ex-aeretodon Bonaparte 1963) but this is not as developed as inLuangwa The posterior extent of the iliac blade is broken inFMNH PR 2444 but this region is preserved in another isolatedright pelvis field number 8-31-98-387 (Fig 13) probably pertain-ing to Menadon besairiei The posterior process of the iliac bladein this specimen is relatively short and directed away from thedorsal edge of the ischium as in Exaeretodon (Bonaparte 1963)but unlike Massetognathus and Luangwa in which the posteriorprocess is long and extends roughly horizontally over the dorsaledge of the ischium (Jenkins 1970 Kemp 1980) and evencurves downward distally in Luangwa The posterior process ofthe iliac blade in Pascualgnathus is directed away from the is-chium (Bonaparte 1966) as in Exaeretodon and Menadon but isrelatively long as in Massetognathus and Luangwa
The proximal portion of the ischium is broad and dominatedby the ischial buttress The ischium constricts posterior to theproximal portion and expands distally into a spatulate platewhich is thickened at its posterior margin This thickening con-tinues along the dorsolateral surface of the ischium forming aridge that slopes ventrally until reaching the edge of the ischialbuttress The downward slope of this ridge produces a prominentgroove at the dorsal edge of the lateral surface of the ischiumthought to represent the origin of the ischio-trochantericusmuscle (Kemp 1980) The constriction of the ischium in Mena-don is like that of Luangwa albeit even more extreme in itsventral concavity but unlike that of Exaeretodon in which themedial and distal ends of the ischium are broadly united Theischium in general is a proportionally shorter element in Ex-aeretodon than in other traversodontids including Menadon(Bonaparte 1963) The anteroventral portion of the ischium thatconnects to the pubis is not preserved in either of the referredMenadon pelves
The pubis is the most incompletely preserved pelvic elementThe acetabular contribution of the pubis is smaller than that ofthe ischium or ilium Distal to the acetabulum the pubis is con-stricted indicating that the connection between the acetabularregion and the pubic plate was considerably narrower than inExaeretodon (Bonaparte 1963) but close akin to the pubis ofLuangwa (Kemp 1980) The pubic plate itself is not preservedThe ventral outline of the ischium and what is known of thepubis indicate that the obturator foramen was relatively larger inMenadon (Fig 14) than in Exaeretodon (Bonaparte 1963)
DISCUSSION
Comparison with the Holotype of Menadon besairiei
FMNH PR 2444 differs in several significant ways from theholotype of Menadon besairiei as previously described most no-tably in the possession of an internarial bar (initially consideredabsent in M besairiei) and four upper incisors (reported as threein M besairiei but with the caveat that the homologies of thefourth and fifth anterior tooth loci were problematic Flynn et al2000) However upon further study we interpret the lack of acontinuous internarial bar in the holotype as breakage ratherthan the natural condition of the skull (a prominent medial pro-tuberance is present on the ventral rim of the nares in UA10601) Regarding the incisor count it should be noted that theholotype of Menadon besairiei displays an unusual set of anterior
JOURNAL OF VERTEBRATE PALEONTOLOGY VOL 28 NO 2 2008454
Dow
nloa
ded
by [
UZ
H H
aupt
bibl
ioth
ek
Zen
tral
bibl
ioth
ek Z
uumlric
h] a
t 05
51 1
0 Se
ptem
ber
2013
FIGURE 10 Right scapulocoracoids of FMNH PR 2444 A stereopair of right scapulocoracoid in medial view B stereopair and interpretivedrawing of right scapulocoracoid in lateral view Scale bars equal 5 cm Abbreviationsmdashac acromion process C coracoid c gl coracoid contributionto the glenoid fossa c tr tubercle for coracoid attachment of triceps PC procoracoid pc f procoracoid foramen S scapula s gl scapularcontribution to the glenoid fossa
KAMMERER ET ALmdashCYNODONT FROM MADAGASCAR 455
Dow
nloa
ded
by [
UZ
H H
aupt
bibl
ioth
ek
Zen
tral
bibl
ioth
ek Z
uumlric
h] a
t 05
51 1
0 Se
ptem
ber
2013
teeth for a traversodontid As mentioned above Flynn et al(2000) noted that the homologies of these teeth were difficult todetermine Of the five anterior teeth preserved in the holotypethe anteriormost three are ldquoundoubtedly incisorsrdquo although I3 isldquounusually large and is caniniformrdquo (Flynn et al 2000423) Therobust recurved fourth tooth in the series was considered torepresent the canine with the subsequent tooth interpreted as anolder canine in the process of replacement Although FMNH PR2444 is edentulous the alveoli on the left side of the skull are wellpreserved and indicate an anterior tooth set identical to the ho-lotype The alveolar morphology indicates that the fifth toothwas canted anteriorly as in the holotype This suggests that themysterious fifth tooth is not the result of a transient period in thedental replacement history of M besairiei as suggested by Flynnet al (2000) but rather is the norm This leaves two possibilities1) M besairiei persistently had two canines This condition isunknown in cynodonts 2) The canine-like fourth tooth actuallyrepresents an incisor and the fifth tooth is the true canine Wefavor the latter explanation In FMNH PR 2444 the first fouralveoli are nearly identical in size and shape However the fifthalveolus while similar in anteroposterior length to its predeces-sors is strongly compressed labiolingually with an anteroposte-rior lengthlabiolingual width ratio of 044 (compared to 080 inthe fourth alveolus and 084ndash85 for I2ndash3) We interpret thismarked difference in alveolar ratio as indicating the separationbetween incisors and canine The canine-like I4 would then rep-resent the endpoint in a series of large increasingly caniniformincisors Supporting this interpretation recurved incisors are alsoobserved in juvenile Exaeretodon (eg MCP 3843 PV see Ab-dala et al 2002)
There are several legitimate differences between FMNH PR2444 and the holotype of Menadon besairiei but we take a con-servative taxonomic stance and argue that these differences rep-resent intraspecific variation and preservational artifacts FMNHPR 2444 represents a larger animal than UA 10601 The snoutlength (measured as the distance between the anterior edge ofthe orbit and a vertical plane through the anterior tip of thepremaxilla) of FMNH PR 2444 is 911 mm while that of UA10601 is 801 mm As such proportional differences between thetwo skulls may be attributable to ontogeny Variations in FMNHPR 2444 compared to the holotype that probably represent on-togentic differences include wider longer and deeper paraca-nine fossae a much deeper ventral zygomatic flange (althoughthe lower border of the flange is slightly abraded in the holotypeof Menadon besairiei there is no evidence that a major flangewas broken off) a proportionally shorter and more circular out-line to the posterior border of the maxilla below the zygomaticprocess and a more deeply emarginated posterior squamosal(ie a more prominent laterally projecting flange)
FMNH PR 2444 has a proportionally shorter more robustsnout and larger orbits than the holotype While these charactersare certainly variable in cynodonts it is unusual that the largerspecimen is shorter-snouted and larger-eyed contra the usualpattern The proportional differences between the two skulls areminor however (orbit diametersnout length is 042 for FMNHPR 2444 and 041 for the holotype) and are partially attributableto deformation (shear in FMNH PR 2444 and breakage of thepostorbital bar in the holotype) The following differences canalso probably be attributed to preservational artifact only fivepostcanine alveoli are preserved in FMNH PR 2444 while theholotype has eight postcanines The anteriormost three in theholotype however are tiny and peg-like As the area of themaxilla between the canine alveolus and the first preserved post-canine alveolus is poorly preserved in FMNH PR 2444 the ab-sence of these postcanines may be artifactual However it is alsopossible that the lower postcanine count of FMNH PR 2444compared to the holotype is an ontogenetic effect as postcaninecount is known to decrease with increasing skull size in Ex-
FIGURE 11 Left scapulocoracoids of FMNH PR 2444 A left scapu-locoracoid in medial view B left scapulocoracoid in lateral view Scalebars equal 5 cm
JOURNAL OF VERTEBRATE PALEONTOLOGY VOL 28 NO 2 2008456
Dow
nloa
ded
by [
UZ
H H
aupt
bibl
ioth
ek
Zen
tral
bibl
ioth
ek Z
uumlric
h] a
t 05
51 1
0 Se
ptem
ber
2013
FIGURE 12 Right pelvis of FMNH PR 2444 A stereopair of right pelvis in medial view B stereopair and interpretive drawing of right pelvis inlateral view Scale bars equal 5 cm Abbreviationsmdashbpp base of posterior process of ilium ib iliac blade IL ilium IS ischium ob f obturatorforamen P pubis
KAMMERER ET ALmdashCYNODONT FROM MADAGASCAR 457
Dow
nloa
ded
by [
UZ
H H
aupt
bibl
ioth
ek
Zen
tral
bibl
ioth
ek Z
uumlric
h] a
t 05
51 1
0 Se
ptem
ber
2013
aeretodon (Abdala et al 2002) The orbit of FMNH PR 2444 isless upwardly directed than that of the holotype (being at anangle of sim65deg to a sagittal plane through the midline of the skullas opposed to sim45deg in the holotype) This difference in orbitalangle is probably attributable to postmortem deformation as theskull of FMNH PR 2444 is noticeably sheared to the front leftThe appearance of a wider squarer incisor arcade and broader
anterior palate in FMNH PR 2444 can also be attributed to thisshear Finally the occipital region of FMNH PR 2444 is alto-gether taller and narrower than that of the holotype For ex-ample the foramen magnum in FMNH PR 2444 approximates arounded isosceles triangle in shape while the foramen magnumof the holotype is more ovoid These differences can also beattributed to deformation Altogether the differences between
FIGURE 13 Stereopairs of field number 8-31-98-387 an isolated right pelvis in A medial and B lateral view Scale bars equal 5 cm
JOURNAL OF VERTEBRATE PALEONTOLOGY VOL 28 NO 2 2008458
Dow
nloa
ded
by [
UZ
H H
aupt
bibl
ioth
ek
Zen
tral
bibl
ioth
ek Z
uumlric
h] a
t 05
51 1
0 Se
ptem
ber
2013
FMNH PR 2444 and the holotype fall within the range of intra-specific variation recorded for other gomphodont cynodontssuch as the traversodontids Massetognathus pascuali (Abdalaand Giannini 2000 Kammerer pers obs) and Exaeretodon(Abdala et al 2002) and the diademodontid Diademodon tetra-gonus (Grine et al 1978 Bradu and Grine 1979)
Phylogenetic Analysis
Utilizing the new information afforded by FMNH PR 2444and a reexamination of the holotype (UA 10601) Menadon be-sairiei was recoded in the Gondwanan gomphodont data matrixof Abdala et al (2006) Abdala et alrsquos (2006) data matrix en-compassed 18 taxa and 43 characters For the present analysis anadditional character ldquoincisor procumbencyrdquo (states 0-absent1-present) was added to Abdala et alrsquos (2006) data set (see
Appendix for coding of this character) Several traversodontidtaxa (eg lsquoScalenodonrsquo hirschsoni) have relatively large incisorsbut only in Menadon Scalenodontoides and Exaeretodon are theincisors procumbent Three of Abdala et alrsquos (2006) originalcodings for Menadon besairiei characters 0 (ldquonumber of upperincisorsrdquo) 5 (ldquoposition of paracanine fossa in relation to theupper caninerdquo) and 21 (ldquointernarial barrdquo) were changed to in-corporate data from the new specimen (see Appendix) Addi-tionally further examination of the holotype (UA 10601) ofMenadon besairiei allowed us to code characters 16ndash18 20 38and 40 (morphology of the lower postcanines all marked as ldquordquoin Abdala et alrsquos analysis) Parsimony searches were performedin PAUP v40b10 using a branch-and-bound search Addition-ally a bootstrap analysis (also using branch-and-bound) was per-formed using 1000 replicates
A total of 34 most parsimonious trees of length 103 werefound with the scores CI0573 RI0725 RC0415 andHI0427 The strict consensus tree (Fig 15A) did not recovera polytomy between Menadon Exaeretodon and Scalenodont-oides as in Abdala and Ribeiro (2003) or a (Menadon + Ex-aeretodon) sister-group relationship as in Abdala et al (2006)Rather Menadon was recovered as the sister taxon of the (Ex-aeretodon + Scalenodontoides) clade supporting Flynn et alrsquos(2000) original placement of this taxon There is strong bootstrapsupport (88) for Traversodontidae but within traversodontidsthe only strongly supported subclade is (Gomphodontosuchus +(Menadon + (Scalenodontoides + Exaeretodon))) with a boot-strap value of 80 Within this clade the (Menadon + (Scalen-odontoides + Exaeretodon)) grouping is supported by a boot-strap value of 83 and the (Scalenodontoides + Exaeretodon)grouping is supported by a bootstrap value of 94 (Fig 15B) Inaddition to the positioning of Menadon the strict consensus tree(Fig 15A) differs from the topology of Abdala et al (2006) inrecovering a sister-group relationship between Massetognathusand Santacruzodon rather than having Massetognathus andSantacruzodon as successive outgroups to the clade containingGomphodontosuchus Menadon Scalenodontoides and Ex-aeretodon The (Massetognathus + Santacruzodon) clade is diag-nosed by the absence of an anterolingual cusp in the upper post-canines denticulated incisor cutting margins and the presence ofan anterior cingulum in the upper postcanines
FIGURE 14 Reconstruction of the pelvis of Menadon besairiei basedon information from FMNH PR 2444 and field number 8-31-98-387Dotted area not preserved in either specimen reconstruction based onPascualgnathus (Bonaparte 1966) and Exaeretodon (Bonaparte 1963)
FIGURE 15 Cladogram of traversodontid relationships based on revised data set of Abdala et al (2006) showing A strict consensus tree and B50 majority rule tree Numbers at nodes of majority rule tree represent bootstrap values
KAMMERER ET ALmdashCYNODONT FROM MADAGASCAR 459
Dow
nloa
ded
by [
UZ
H H
aupt
bibl
ioth
ek
Zen
tral
bibl
ioth
ek Z
uumlric
h] a
t 05
51 1
0 Se
ptem
ber
2013
Relationships of Menadon besairiei
Menadon besairiei is the sister taxon to (Exaeretodon + Sca-lenodontoides) the latter clade diagnosed by the possession ofthree upper incisors a posteromedial position of the paracaninefossae relative to the canines the absence of an internarial barlarge canines a projected zygomatic process of the jugal and amaxilla that contributes to the margin of the subtemporal fenes-tra (the last three character states represent reversals) The clade(Menadon + (Exaeretodon + Scalenodontoides)) is diagnosed bythe possession of large procumbent incisors The presence offour incisors a primitive retention distinguishes Menadon be-sairiei from Exaeretodon Scalenodontoides and an unnamed tra-versodontid (ldquoTraversodontid Type 1rdquo) from the Santa MariaFormation of Brazil (Abdala et al 2001 Abdala and Ribeiro2002) Gomphodontosuchus is recovered as the sister taxon to(Menadon (Exaeretodon + Scalenodontoides)) as has beenfound in previous studies (Flynn et al 2000 Abdala and Ribeiro2003) The taxon name Gomphodontosuchinae Watson andRomer 1956 (ex Gomphodontosuchidae Watson and Romer1956 first given subfamilial rank in Lehman 1961) is availablefor this clade As circumscribed here Gomphodontosuchinaeconsists of the most inclusive clade containing Gomphodontosu-chus brasiliensis von Huene 1928264 but not Massetognathuspascuali Romer 19672
Menadon besairiei displays a mixture of primitive and derivedcharacters within traversodontids The internarial bar and fourincisors are primitive features whereas the enlargement and pro-cumbency of the incisors are derived features shared with Ex-aeretodon and Scalenodontoides The postcranium shows severalsimilarities to that of Exaeretodon (apparent exclusion of theprocoracoid from the glenoid some features of the pelvis tallcaudal neural spines) but in general displays a rather conserva-tive morphology observed throughout Gomphodontia The mor-phology of the scapulocoracoids is closer akin to diademodontidsor trirachodontids than Exaeretodon and the expanded thoracicrib of Menadon has only slight differences from those of trirach-odontids or early traversodontids like Luangwa The atlas-axiscomplex of Menadon is similar to that of the Early Triassic cyn-odont Cynognathus in possessing a concave axial spine with along posterior process Although most gomphodonts have beenreconstructed with a convex axial spine as in early mammals thisspine is broken in most of the best described gomphodont post-crania (eg Diademodon Brink 1955) and the lsquosaddle-shapedrsquoaxial morphology may be more widely distributed among cyn-odonts than previously realized The new specimens of Menadondescribed in this paper demonstrate that a generalized gompho-dont postcranial morphology was retained deep within the tra-versodontid tree and that the derived morphologies of either ofthe well-known taxa Massetognathus or Exaeretodon should notbe taken as representative of traversodontids in general
ACKNOWLEDGMENTS
This work would not have been possible without the continu-ing support and encouragement of our collaborative programwith the Deacutepartement de Paleacuteontologie et drsquoAnthropologie Bi-ologique Universiteacute drsquoAntananarivo Madagascar under the di-rection of Armand Rasoamiaramanana (Department Chairman)and Gisegravele Ravololonarivo Randria (Head of the PaleontologyLaboratories) We are extremely grateful to the National Geo-graphic Society for their long-term support of the fieldwork as-sociated with this project Additional financial or logistical sup-port for fieldwork was provided by The Field Museum the Uni-versity of California-Santa Barbara and the WWF (WorldwideFund for Nature Madagascar) We acknowledge the Ministegraverede LrsquoEneacutergie et des Mines for authorizing field efforts and for
continuing to facilitate our joint US-Madagascar explorationresearch and education program Steve Goodman and AsminaGandhi generously assisted us in innumerable ways includinghospitality nonpareil We especially thank R Andriantompoha-vana L Razafimanantsoa A Toto Volahy JA Rabarison AGandie and P Vavisaro for assistance in the field during theexpedition in which these new specimens were discovered BillSimpson and Gina Wesley-Hunt for their patient excavation ofthis important material Allison Smith Ray Vodden Lisa Berg-wall and Deborah Wagner for excellent preparation of a chal-lenging specimen Jennifer Botha and Elize Butler (NationalMuseum Bloemfontein) Michael Raath and Fernando Abdala(Bernard Price Institute) Thomas Kemp (Oxford UniversityMuseum of Natural History) Ray Symonds (University Museumof Zoology Cambridge) Bernard Battail (Museacuteum nationaldrsquoHistoire naturelle Paris) Michael Maisch (Universitaumlt Tuumlbin-gen) and Charles Schaff and Farish Jenkins (Museum of Com-parative Zoology Harvard) for access to specimens and AgustiacutenMartinelli Fernando Abdala and James Hopson for helpful re-views of an earlier version of the manuscript
LITERATURE CITED
Abdala F M C Barbarena and J Dornelles 2002 A new species of thetraversodontid cynodont Exaeretodon from the Santa Maria Forma-tion (MiddleLate Triassic) of Southern Brazil Journal of Verte-brate Paleontology 22313ndash325
Abdala F and N P Giannini 2000 Gomphodont cynodonts of theChantildeares Formation the analysis of an ontogenetic sequence Jour-nal of Vertebrate Paleontology 20501ndash506
Abdala F J Neveling and J Welman 2006 A new trirachodontidcynodont from the lower levels of the Burgersdorp Formation(Lower Triassic) of the Beaufort Group South Africa and the cla-distic relationships of Gondwanan gomphodonts Zoological Jour-nal of the Linnean Society 147383ndash413
Abdala F A M Ribeiro and C L Schultz 2001 A rich cynodont faunaof Santa Cruz do Sul Santa Maria Formation (Middle-Late Trias-sic) southern Brazil Neues Jahrbuch fuumlr Geologie und Palaumlontolo-gie Monatshefte 2001669ndash687
Abdala F and A M Ribeiro 2002 Nuevos cinodontes traversodoacuten-tidos (Synapsida-Eucynodontia) de la Formacioacuten Santa Maria(Triaacutesico Medio-Superior) Rio Grande do Sul Brasil RevistaEspantildeola de Paleontologiacutea 17237ndash243
Abdala F and A M Ribeiro 2003 A new traversodontid cynodontfrom the Santa Maria Formation (Ladinian-Carnian) of southernBrazil with a phylogenetic analysis of Gondwanan traversodontidsZoological Journal of the Linnaean Society 139529ndash545
Abdala F and A M S Teixeira 2004 A traversodontid cynodont ofAfrican affinity in the South American Triassic Palaeontologia Af-ricana 4011ndash22
Bonaparte J F 1962 Descripcioacuten del craacuteneo y mandiacutebula de Ex-aeretodon frenguellii Cabrera y su comparacioacuten con Diademodon-tidae Tritylodontidae y los cinodontes sudamericanos Publica-ciones del Museo Municipal de Ciencias Naturales y Tradicional deMar del Plata 1135ndash202
Bonaparte J F 1963 Descripcioacuten del esqueleto postcraneano de Ex-aeretodon (CynodontiandashTraversodontidae) Acta Geoloacutegica Lil-loana 45ndash52
Bonaparte J F 1966 Una nueva ldquofaunardquo Triaacutesica de Argentina (Ther-apsida Cynodontia Dicynodontia) Consideracions filogeneacuteticas ypaleobiogeograacuteficas Ameghiniana 8243ndash296
Bradu D and F E Grine 1979 Multivariate analysis of diademodon-tine crania from South Africa and Zambia South African Journal ofScience 79441ndash448
Brink A S 1955 A study of the skeleton of Diademodon Palaeonto-logia Africana 33-39
Brink A S 1963 Two new cynodonts from the Ntawere Formation inthe Luangwa Valley of Northern Rhodesia Palaeontologia Africana877ndash96
Broom R 1905 On the use of the term Anomodontia Records of theAlbany Museum 1266ndash269
Cabrera A 1943 El primer hallazgo de teraacutepsidos en la ArgentinaNotas del Museo de La Plata Paleontologiacutea 55317ndash331
JOURNAL OF VERTEBRATE PALEONTOLOGY VOL 28 NO 2 2008460
Dow
nloa
ded
by [
UZ
H H
aupt
bibl
ioth
ek
Zen
tral
bibl
ioth
ek Z
uumlric
h] a
t 05
51 1
0 Se
ptem
ber
2013
Chatterjee S 1982 A new cynodont reptile from the Triassic of IndiaJournal of Paleontology 56203ndash214
Crompton A W 1955 On some Triassic cynodonts from TanganyikaProceedings of the Zoological Society of London 125617ndash669
Crompton A W 1972 Postcanine occlusion in cynodonts and tritylo-donts Bulletin of the British Museum (Natural History) Geology2129ndash71
Crompton A W and F Ellenberger 1957 On a new cynodont from theMolteno Beds and the origin of the tritylodontids Annals of theSouth African Museum 441ndash13
Flynn J J J M Parrish B Rakotosaminimanana W F Simpson R LWhatley and A R Wyss 1999 A Triassic fauna from Madagascarincluding early dinosaurs Science 286763ndash765
Flynn J J J M Parrish B Rakotosamimanana L RanivoharimananaW F Simpson and A R Wyss 2000 New traversodontids (Syn-apsida Eucynodontia) from the Triassic of Madagascar Journal ofVertebrate Paleontology 20422ndash427
Gow C E and P J Hancox 1993 First complete skull of the LateTriassic Scalenodontoides (Reptilia Cynodontia) from southern Af-rica In Lucas S G and M Morales (eds) The nonmarine TriassicNew Mexico Museum of Natural History amp Science Bulletin 3161ndash168
Grine F E B D Hahn and C E Gow 1978 Aspects of relativegrowth and variability in Diademodon (Reptilia Therapsida) SouthAfrican Journal of Science 7450ndash58
Hopson J A 1984 Late Triassic traversodont cynodonts from NovaScotia and southern Africa Palaeontologia Africana 25181ndash201
Hopson J A 1985 Morphology and relationships of Gomphodontosu-chus brasiliensis von Huene (Synapsida Cynodontia Tritylodontoi-dea) from the Triassic of Brazil Neues Jahrbuch fuumlr Geologie undPalaumlontologie Monatshefte 1985285ndash299
Hopson J A and J W Kitching 2001 A probainognathian cynodontfrom South Africa and the phylogeny of nonmammalian cynodontsBulletin of the Museum of Comparative Zoology 1565ndash35
Hopson J A and H-D Sues 2006 A traversodont cynodont from theMiddle Triassic (Ladinian) of Baden-Wuumlrttemberg (Germany)Palaumlontologische Zeitschrift 80124ndash129
Huene F von 1928 Ein Cynodontier aus der Trias Brasiliens Zentral-blatt fuumlr Mineralogie Geologie und Palaumlontologie 1928251ndash270
Huene F von 1936 Die fossilen Reptilien des suumldamerikanischenGondwanalandes Ergebnisse der Sauriergrabungen in Suumldbrasilien192829 Lieferung 2 (Cynodontia)93ndash159 Franz F Heine Tuumlbin-gen
Jenkins F A 1970 The Chantildeares (Argentina) Triassic reptile faunaVII The postcranial skeleton of the traversodontid Massetognathuspascuali (Therapsida Cynodontia) Breviora 3521ndash28
Jenkins F A 1971 The postcranial skeleton of African cynodonts prob-lems in the early evolution of the mammalian postcranial skeletonBulletin of the Peabody Museum of Natural History 361ndash216
Kemp T S 1969 The atlas-axis complex of mammal-like reptiles Jour-nal of Zoology 159223ndash248
Kemp T S 1980 Aspects of the structure and functional anatomy of theMiddle Triassic cynodont Luangwa Journal of Zoology 191193ndash239
Kemp T S 1982 Mammal-Like Reptiles and the Origin of MammalsAcademic Press New York
Lehman J-P 1961 Cynodontia pp 140ndash191 in Piveteau J (ed) Traiteacutede Paleacuteontologie Masson et Cie Paris
Osborn 1903 On the primary division of the Reptilia into two sub-classes Synapsida and Diapsida Science 17275ndash276
Owen R 1861 Palaeontology or a systematic summary of extinct ani-mals and their geological relations 2nd Ed Edinburgh
Parrington F R 1934 On the cynodont genus Galesaurus with a noteon the functional significance of the changes in the evolution of thetheriodont skull Annals and Magazine of Natural History 10(13)38ndash67
Parrington F R 1946 On the cranial anatomy of cynodonts Proceed-ings of the Zoological Society of London 116181ndash197
Razafimbelo E 1987 Le basin de Morondava (Madagascar) Synthegravesegeacuteologique et structurale PhD dissertation University Louis Pas-teur Strasbourg France 189 pp
Reisz R R and H-D Sues 2000 Herbivory in Late Paleozoic andTriassic terrestrial vertebrates pp 9ndash41 in Sues H-D (ed) Evo-lution of Herbivory in Terrestrial Vertebrates Cambridge Univer-sity Press New York
Romer A S 1967 The Chantildeares (Argentina) Triassic reptile fauna IIITwo new gomphodonts Massetognathus pascuali and M terugiiBreviora 2641ndash25
Romer A S 1972 The Chantildeares (Argentina) Triassic reptile faunaXVII The Chantildeares gomphodonts Breviora 3961ndash9
Rougier G W J R Wible and J A Hopson 1992 Reconstruction ofthe cranial vessels in the Early Cretaceous mammal Vincelestes neu-quenianus implications for the evolution of the mammalian cranialvascular system Journal of Vertebrate Paleontology 12188ndash216
Rowe T 1988 Definition diagnosis and origin of Mammalia Journal ofVertebrate Paleontology 8241ndash264
Rowe T 1993 Early mammal phylogenetic systematics pp 129ndash145 inSzalay F S M J Novacek and M C McKenna (eds) MammalPhylogeny Vol 1 Mesozoic Differentiation MultituberculatesMonotremes Early Therians and Marsupials Springer-Verlag NewYork
Seeley H G 1894 Researches on the structure organization and clas-sification of the fossil Reptilia Part IX Section 3 On DiademodonPhilosophical Transactions of the Royal Society of London 1851029ndash1041
Seeley H G 1895 Researches on the structure organization and clas-sification of the fossil Reptilia Part IX Section 4 On the Gompho-dontia Philosophical Transactions of the Royal Society of London1861ndash57
Seeley H G 1895 Researches on the structure organization and clas-sification of the fossil Reptilia Part IX Section 5 On the skeletonin new Cynodontia from the Karroo rocks Philosophical Transac-tions of the Royal Society of London 18659ndash148
Sues H-D J A Hopson and N H Shubin 1992 Affinities of Sca-lenodontoides plemmyridon Hopson 1984 (Synapsida Cynodontia)from the Upper Triassic of Nova Scotia Journal of Vertebrate Pa-leontology 12168ndash171
Sues H-D and P E Olsen 1990 Triassic vertebrates of Gondwananaspect from the Richmond basin of Virginia Science 2491020ndash1023
Swofford D L 2003 PAUP v 40b10 Sinauer Associates Inc Pub-lishers Sunderland Massachusetts
Watson D M S and A S Romer 1956 A classification of therapsidreptiles Bulletin of the Museum of Comparative Zoology 11435ndash89
Submitted September 20 2007 accepted November 3 2007
KAMMERER ET ALmdashCYNODONT FROM MADAGASCAR 461
Dow
nloa
ded
by [
UZ
H H
aupt
bibl
ioth
ek
Zen
tral
bibl
ioth
ek Z
uumlric
h] a
t 05
51 1
0 Se
ptem
ber
2013
APPENDIX Data matrix used in the phylogenetic analysesThis is a revised version of the matrix of Abdala et al (2006) with the following changes character 43 [Incisor procumbency (0) absent (1) present]added Menadon recoded for characters 0 (Number of upper incisors was 1 ldquothreerdquo changed to 0 ldquofourrdquo) 5 (Position of paracanine fossae in relationto the upper canine was changed to 1 ldquomedialrdquo) 16 (Overall morphology of lower postcanines in occlusal view was changed to 3 ldquoquadran-gularrdquo) 17 (Position of the transverse crest in lower postcanines was changed to 1 ldquoanteriorrdquo) 18 (Number of cusps in the transverse crest of thelower postcanines was changed to 0 ldquotwordquo) 20 (Anterior cingulum in the lower postcanines was changed to 2 ldquoabsentrdquo) 21 (Internarial barwas 1 ldquoabsentrdquo changed to 0 ldquopresentrdquo) 38 (Posterior cingulum in the lower postcanines was changed to 1 ldquoabsentrdquo) and 40 (Deep occlusalbasins in the postcanines was changed to 1 ldquopresentrdquo) Langbergia recoded for character 34 (Incisor cutting margins was 0 ldquoserratedrdquo changedto 1 ldquosmoothly ridgedrdquo) based on the description of the incisors in Abdala et al (2006) Gomphodontosuchus recoded for character 31 (Maxilla inthe margin of the subtemporal fenestra was 1 ldquoincludedrdquo changed to 0 ldquoexcludedrdquo) based on von Huene (1936) and Kammerer (pers obs of theholotype) contra Hopsonrsquos (1985) reconstruction lsquoScalenodonrsquo hirschsoni recoded for character 39 (Anterior cingulum in the upper postcanines was1 ldquoabsentrdquo changed to 0 ldquopresentrdquo) based on Abdala (pers comm 2006) Trirachodon recoded for character 30 (Ectopterygoid was changedto 0 ldquopresentrdquo) based on Abdala (pers comm 2006) and Traversodon recoded for character 15 (Number of cusps in the sectorial border of the upperpostcanines was 0 ldquomore than twordquo changed to 1 ldquotwordquo) based on Abdala (pers comm 2006) and Kammerer (pers obs)
1111111111 2222222222 3333333333 44440123456789 0123456789 0123456789 0123456789 0123
Cynognathus 00000000 000 000000000 00000000 000Diademodon 0000001000 0000100 000010A000 0000000100 0100Trirachodon 000000100B 1A002201 001A100010 0001001200 0110Langbergia 0000001000 10000201 0000101010 0001101200 0110Cricodon 0100001000 11000201 0010100 0001200 0110Andescynodon 0000002001 01013101 101120010 0112311 1100Massetognathus 00111121A2 1110003100 2011200011 1011212300 1100Exaeretodon 1110122112 010103101 2111110201 1111112311 1101Luangwa 0000001002 11010A3100 101010010 01002300 1100Scal angustifrons 000000100B 1111023100 10102101 00002301 1100Scal hirschsoni 11110202 1100113100 1010 1112300 1100Traversodon 0000002012 1101113100 201010 011230 1100Gomphodontosuchus 0011112112 0103101 2010 0112311 1100Pascualgnathus 1000001000 000231 1100001 0011230 1100Scalenodontoides 1112212 01003101 21111021 1212311 1101Menadon 0111112112 01012310 20121020 0102311 1101Dadadon 00002112 111112 013 11231 1100Santacruzodon 00002112 1110003101 201300 212300 1100
A(01) B(02)
JOURNAL OF VERTEBRATE PALEONTOLOGY VOL 28 NO 2 2008462
Dow
nloa
ded
by [
UZ
H H
aupt
bibl
ioth
ek
Zen
tral
bibl
ioth
ek Z
uumlric
h] a
t 05
51 1
0 Se
ptem
ber
2013
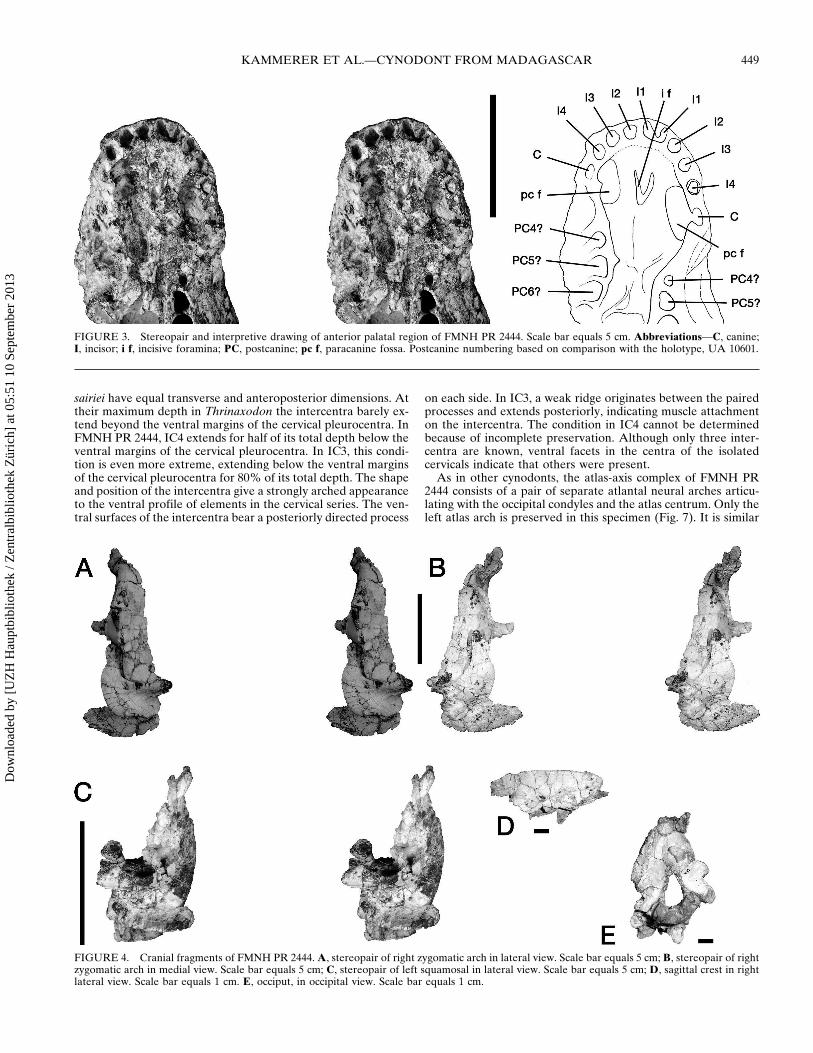
sairiei have equal transverse and anteroposterior dimensions Attheir maximum depth in Thrinaxodon the intercentra barely ex-tend beyond the ventral margins of the cervical pleurocentra InFMNH PR 2444 IC4 extends for half of its total depth below theventral margins of the cervical pleurocentra In IC3 this condi-tion is even more extreme extending below the ventral marginsof the cervical pleurocentra for 80 of its total depth The shapeand position of the intercentra give a strongly arched appearanceto the ventral profile of elements in the cervical series The ven-tral surfaces of the intercentra bear a posteriorly directed process
on each side In IC3 a weak ridge originates between the pairedprocesses and extends posteriorly indicating muscle attachmenton the intercentra The condition in IC4 cannot be determinedbecause of incomplete preservation Although only three inter-centra are known ventral facets in the centra of the isolatedcervicals indicate that others were present
As in other cynodonts the atlas-axis complex of FMNH PR2444 consists of a pair of separate atlantal neural arches articu-lating with the occipital condyles and the atlas centrum Only theleft atlas arch is preserved in this specimen (Fig 7) It is similar
FIGURE 3 Stereopair and interpretive drawing of anterior palatal region of FMNH PR 2444 Scale bar equals 5 cm AbbreviationsmdashC canineI incisor i f incisive foramina PC postcanine pc f paracanine fossa Postcanine numbering based on comparison with the holotype UA 10601
FIGURE 4 Cranial fragments of FMNH PR 2444 A stereopair of right zygomatic arch in lateral view Scale bar equals 5 cm B stereopair of rightzygomatic arch in medial view Scale bar equals 5 cm C stereopair of left squamosal in lateral view Scale bar equals 5 cm D sagittal crest in rightlateral view Scale bar equals 1 cm E occiput in occipital view Scale bar equals 1 cm
KAMMERER ET ALmdashCYNODONT FROM MADAGASCAR 449
Dow
nloa
ded
by [
UZ
H H
aupt
bibl
ioth
ek
Zen
tral
bibl
ioth
ek Z
uumlric
h] a
t 05
51 1
0 Se
ptem
ber
2013
to the atlantal neural arches described for other cynodonts suchas Massetognathus (Kemp 1969 Jenkins 1971) There is a me-dial facet for articulation with the centrum of the atlas (theldquoodontoid facetrdquo of Kemp 1969) and an anterior facet for ar-ticulation with the occipital condyle (Fig 7B) A pair of pro-cesses dominates the atlas arch a dorsal lamina (a roughly rect-angular plate of bone arching above the facets) and a largeposterolaterally directed transverse process An oval tubercle atthe anterior end of the transverse process represents the facet forthe proatlas
The centrum of the atlas is fused to the axis forming a singlelarge vertebral unit (Fig 6) A weak odontoid process is presentat the dorsal limit of the anterior surface of the atlas The ante-rior aspect of the centrum is dominated by a roughly lsquoTrsquo-shapedstructure composed of the two facets for articulation with theatlas arches and the facet for articulation with the atlas intercen-trum The axis bears no prezygapophyses and as such does notarticulate with the atlas arch as in Early Triassic cynodonts suchas Galesaurus (Parrington 1934) There appears to be a trendtowards loss of the axial prezygapophyses in later traversodon-
tids which are present but vestigial in Massetognathus (Jenkins1970 although in the specimens described the prezygapophysesare broken so their reduction is perhaps questionable) and ab-sent in Exaeretodon (Bonaparte 1963) The transverse processesof the axis are large and robust compared to those of the othercervicals The axial spine is saddle-shaped with a concave dorsalprofile unlike previous reconstructions of the traversodontidaxis which depict a straight or convex dorsal profile similar tothat observed in galesaurid cynodonts and early mammals (Jen-kins 1971) The morphology of the axial spine is similar to thatof the Early Triassic cynodont Cynognathus (Seeley 1895) InCynognathus however the neural spine of the third cervical(C3) is tall and canted posteriorly behind the posterior process ofthe axial spine whereas in FMNH PR 2444 the short C3 neuralspine fits below the posterior portion of the axial spine Theposterior process of the axial spine bears a pair of parallel elon-gate grooves on its underside attachment sites for the axial mus-culature
Post-Axial VertebraemdashThree post-axial cervicals are knownfor FMNH PR 2444 (Figs 6 8AndashD) The first of these C3 is
FIGURE 5 Stereopair of ventral view of the occipital fragment of FMNH PR 2444 showing the basicranium Anterior is up
FIGURE 6 Stereopair and interpretive drawing of atlas-axis complex and third cervical of FMNH PR 2444 in left lateral view Scale bar equals5 cm Abbreviationsmdasha n sp axial neural spine d dens IC intercentrum f at a facet for articulation with the atlas arch p parapophysis pozpostzygapophysis t p transverse process
JOURNAL OF VERTEBRATE PALEONTOLOGY VOL 28 NO 2 2008450
Dow
nloa
ded
by [
UZ
H H
aupt
bibl
ioth
ek
Zen
tral
bibl
ioth
ek Z
uumlric
h] a
t 05
51 1
0 Se
ptem
ber
2013
preserved in articulation with the axis (Fig 6) The two isolatedcervicals bear successively taller and anterposteriorly wider neu-ral spines (Fig 8AndashD) Based on their staggered size and failureto articulate we suggest that they represent C5 and C7 (assumingthat this specimen possessed the generalized cynodont cervicalnumber of seven) The cervicals display robust transverse pro-cesses which are directed laterally and slightly anteriorly A par-apophysis lies below the anterior edge of the transverse processalong the antero-lateral edge of the centrum No cervical ribs arepreserved but their presence can be inferred from the par-apophyses The notochordal concavity is as in Massetognathussurrounded by a raised rim at each end of the centrum (Jenkins1970) No anapophyses are present The neural spines for allthree post-axial cervicals are tapering and slightly curved poste-riorly The neural spine in C3 is relatively short and wide(thereby fitting underneath the posterior overhang of the axialspine see Fig 6) while that of C5 is tall and narrow The spinefor C7 is taller and anteroposteriorly wider than that of C5 andthe dorsal edge is more rounded similar to the condition in thedorsal vertebrae
Eight post-cervical presacral vertebrae are known for thisspecimen We will refer to them here simply as dorsal vertebraegiven the incompleteness of the vertebral column in the speci-men and the uncertainty concerning the thoraciclumbar transi-tion in this taxon However it is probable that they representthoracic vertebrae (see below) In most cynodonts the lumbar
vertebrae are synostosed to specialized ribs with overlappingelements In earlier taxa (eg Thrinaxodon Cynognathus Lu-angwa Jenkins 1971 Kemp 1980) expanded costal plates arepresent on both thoracic and lumbar ribs In Massetognathus thelumbar ribs consist of a pair of prongs one directed anteriorlybearing a facet for articulation with the previous rib and theother directed posteriorly (Jenkins 1970) In Exaeretodon allpresacral ribs are identical of simple morphology and not syn-ostosed to the vertebrae (Bonaparte 1963) A single isolated rib(Fig 9B) in FMNH PR 2444 demonstrates that some of the ribsin Menadon besairiei were expanded However all preservedvertebrae in this specimen have facets for articulation with theribs rather than breakage points where they would have beenfused to ribs As such it is probable that all of the vertebraepreserved in this specimen represent thoracics although we can-not discount the possibility that as in Exaeretodon no presacralvertebrae were fused to ribs
The neural spines in all of the dorsal vertebrae are elongateposteriorly inclined and roughly rectangular in lateral view witha rounded laterally expanded platform dorsally (Fig 8EndashJ)Costal foveae are present at the anterior and posterior edges ofeach centrum Of the eight dorsal vertebrae there are two ar-ticulated sets and three isolated elements (two from 9-4-98-440and one from 9-2-98-422) The first isolated vertebra (Fig 8E F)has a taller neural spine and shorter centrum than the otherdorsal vertebrae It is most similar in proportions to the isolated
FIGURE 7 Stereopairs and interpretive drawings of left atlas arch of FMNH PR 2444 in A lateral view Anterior is left B medial view Anterioris right Scale bars equal 1 cm Abbreviationsmdashafa anterior facet for articulation with the occipital condyle dl dorsal lamina mfa medial facet forarticulation with the atlas centrum (odontoid facet) pfa tubercle forming the facet for articulation with the proatlas tp transverse process
KAMMERER ET ALmdashCYNODONT FROM MADAGASCAR 451
Dow
nloa
ded
by [
UZ
H H
aupt
bibl
ioth
ek
Zen
tral
bibl
ioth
ek Z
uumlric
h] a
t 05
51 1
0 Se
ptem
ber
2013
FIGURE 8 Vertebrae of FMNH PR 2444 Cervical vertebra 5 in A left lateral and B anterior views Cervical vertebra 7 in C left lateral andD anterior views Isolated dorsal vertebra from anterior region of the series in E left lateral and F anterior views G stereopair and interpretivedrawing of articulated set of two dorsal vertebrae from near the midpoint of the series in left lateral view Anterior is up H articulated set of threedorsal vertebrae from anterior region of the series in left and right lateral views I isolated vertebra from near the midpoint of the series (from fieldnumber 9-2-98-422) J isolated vertebra from posterior region of the series Sacral vertebra in K dorsal view (the distal region of another sacral ribwhich is identical in shape to that of the intact sacral rib albeit slightly larger is also shown in this figure) and L left lateral view Caudal vertebraeM articulated set of two vertebrae from the anterior region of the series showing the lsquowingrsquo-shaped caudal ribs in dorsal view Anterior is left Nisolated caudal vertebra showing tall neural spine in left lateral view O isolated caudal vertebra showing tall neural spine in posterior view Particulated set of four vertebrae from near the end of the caudal series in dorsal view Anterior is left Scale bars equal 1 cm Abbreviationsmdasha cof anterior costal fovea n sp neural spine p co f posterior costal fovea poz postzygapophysis prz prezygapophysis
JOURNAL OF VERTEBRATE PALEONTOLOGY VOL 28 NO 2 2008452
Dow
nloa
ded
by [
UZ
H H
aupt
bibl
ioth
ek
Zen
tral
bibl
ioth
ek Z
uumlric
h] a
t 05
51 1
0 Se
ptem
ber
2013
cervicals and was probably located near the front of the dorsalseries The first articulated set consists of three vertebrae (Fig8H) The middle vertebra in this set is broken from the anteriorbase of the neural spine to the ventral midpoint and most of theright side of the anterior half of the centrum is missing This setof vertebrae probably also came from the anterior region of theseries as their neural spines are longer and narrower than allvertebrae other than the one described above The second ar-ticulated set (Fig 8G) consists of two well-preserved vertebraethat based on comparisons with Massetognathus and Ex-aeretodon (Jenkins 1970 Bonaparte 1963) probably came fromnear the midpoint of the dorsal series The second isolated ver-tebra (Fig 8I) from 9-2-98-422 is similar to the members of thesecond set The third isolated vertebra (Fig 8J) appears to befrom the posterior end of the series based on the relatively shortneural spine and reduced posterior costal fovea
One sacral vertebra (Fig 8K L) is known consisting of awell-preserved centrum neural spine and the right sacral rib(the left is broken off) The centrum is shorter (both anteropos-teriorly and dorsoventrally) than those of the dorsal vertebraeand the neural spine is shorter and more rounded in profileAlso the neural spine is directed vertically unlike those of thedorsal vertebrae which are canted posteriorly The zygapophy-ses are poorly developed The sacral rib is fused to a singlesynapophysis and projects anterolaterally The rib narrows to-wards the middle and expands distally terminating in a widerectangular facet where it would meet the ilium (Fig 8K)
Seven caudal vertebrae are known The caudal vertebrae aredistinctive in having very tall narrow neural spines Some EarlyTriassic cynodonts possess blunt neural spines of moderateheight on the proximal caudals (see for example Thrinaxodonin Jenkins 1971 and Diademodon in Brink 1955) Massetogna-thus represents an extreme condition wherein the caudal neuralspines are greatly reduced (Jenkins 1970) Very tall neuralspines are seen in Exaeretodon (Bonaparte 1963) and thosespines are unusual in that they do not diminish in height until thelast caudals The caudal vertebrae of FMNH PR 2444 consist ofa pair of articulated vertebrae most likely from near the sacrum(Fig 8M) an isolated caudal lacking postzygapophyses and ribsbut with the neural spine (Fig 8N O) and a quartet of smaller
articulated elements (Fig 8P) The anterior caudals have nar-rower neural spines and shorter zygapophyses than the subse-quent vertebrae in the series The caudal ribs attached to thispair have a crescentic distal region that curves anteriorly pro-ducing a lsquowingrsquo-shape Only a portion of the right rib of theisolated caudal is preserved but the lsquowingrsquo-shaped morphologyof the near-sacral caudal ribs does not appear to have been pre-sent In the posteriormost four vertebrae caudal ribs are pre-served on the left side of the first and the right side of the secondand third members On the first of these vertebrae the rib isshort curved posteriorly (but not lsquowingrsquo-shaped) and taperingRibs of the second and third vertebrae are less curved resem-bling posteriorly directed rods The zygapophyses of both theisolated caudal vertebra and the quartet of small caudals areanteroposteriorly elongate becoming increasingly so posteriorly
Thoracic RibsmdashNine thoracic ribs are known from 9-4-98-440(four left four right and an indeterminate splint of bone Fig9A indeterminate bone not illustrated) These elements re-semble the flattened ribs lacking costal plates of Exaeretodon(Bonaparte 1963) and the anterior thoracic ribs of Massetogna-thus and Luangwa (Jenkins 1970 Kemp 1980) The tuberculumand capitulum are separated by a shallow depression The tuber-culum has a shorter base than the capitulum but their proximalfaces are subequal in size An additional isolated left rib (Fig9B) from 9-2-98-422 is expanded indicating that unlike Ex-aeretodon Menadon rib morphology was heterogeneous alongthe length of the dorsal series This rib closely resembles theposterior thoracic ribs described for Luangwa (Kemp 1980fig16) Pascualgnathus (Bonaparte 1966fig 7) and the trirach-odontid Cricodon (Crompton 1955 Fig 4) This rib is con-stricted at the base of the head then expands distally with an-terior and posterior costal plates As in Luangwa the anteriorcostal plate is shallow and evenly sloping proximally and distallywhile the posterior costal plate is falcate with a concave edgedistally The distal expansion of this rib occurs at an angle of sim30degto the long axis of the rib head Despite their overall similaritythe expanded rib in FMNH PR 2444 does exhibit some differ-ences from the posterior thoracic rib of Luangwa The expandedrib of Menadon has a longer shaft between the rib head and thecostal plates and a shorter capitulum lacks a ridge on the pos-
FIGURE 9 A anterior thoracic ribs of FMNH PR 2444 in anterior view (indeterminate rib not figured) B expanded posterior thoracic rib ofFMNH PR 2444 in anterior view Scale bars equal 1 cm
KAMMERER ET ALmdashCYNODONT FROM MADAGASCAR 453
Dow
nloa
ded
by [
UZ
H H
aupt
bibl
ioth
ek
Zen
tral
bibl
ioth
ek Z
uumlric
h] a
t 05
51 1
0 Se
ptem
ber
2013
terior costal plate and has an anteroposteriorly shorter anteriorcostal plate Additionally the posterior margin of the costal plateis less curved than that of Luangwa These differences couldindicate that the expanded rib of Menadon is from a more pos-terior position in the series than the rib from Luangwa as re-duced curvature and increasing confluence of the tuberculumand capitulum are characteristic of lumbar ribs in traversodon-tids (Jenkins 1970 Kemp 1980) However the length of theshaft is at odds with this interpretation and in Cricodon andPascualgnathus the more posterior ribs are not more similar tothe element from FMNH PR 2444 These differences probablyrepresent legitimate morphological distinctions between the ribsof earlier gomphodonts and Menadon The head of the expandedrib is well defined and unbroken indicating that it was not syn-ostosed to a vertebra
Appendicular Skeleton
The appendicular skeleton is represented by both scapulocora-coids and a fragment of a clavicle from field number 9-4-98-440and a partial right pelvis from field number 9-2-98-422 An ad-ditional partial right pelvis is represented from field number8-31-98-387 The scapulocoracoids (Figs 10 11) are very similarto those described for other traversodontids as well as cynog-nathids and diademodontids (Jenkins 1970 1971) The scapularinfraspinous fossa has out-turned anterior and posterior edgesThe anterior edge of the scapula is rounded and curves laterallyforming the scapular spine At the lower end of the spine theacromion forms a prominent process (broken in the left scapula)Below this process the scapula is constricted and then expandsventrally to form the dorsal margin of the articular glenoid Theglenoid fossa is in general morphology very similar to that ofother Triassic cynodonts (Jenkins 1970 1971 Kemp 1980) It isposteroventrally oriented with roughly equal contributions fromthe scapular and coracoid glenoid facets The coracoid glenoidfacet is roughly triangular while the scapular glenoid facet issemiovoid as in other cynodonts In anterior or posterior viewthe fossa appears smoothly crescentic The procoracoid is aslightly curved trapezoidal plate of bone It does not appear tocontribute to the glenoid fossa The procoracoid foramen is lo-cated midway up the height of the procoracoid at a point directlybelow the base of the acromion process The coracoid is semi-crescentic curving posteriorly from a broad base The coracoidin Menadon is longer and more falcate than those described forExaeretodon (Bonaparte 1963) and Luangwa (Kemp 1980 al-though the distal tip of both coracoids is broken in the specimenof Luangwa [TSK 121] described by Kemp and may have beenlonger than reconstructed) but agrees closely with those of Mas-setognathus and cynognathidsdiademodontids (Jenkins 19701971) The distal coracoid tubercle for attachment to the tricepsis strongly curved upwards similar to the condition in Masseto-gnathus (Jenkins 1970) and Diademodon (Brink 1955)
A small fragment of bone resembling a curved cylinder almostcertainly represents part of a clavicle It has no distinguishingfeatures other than a groove associated with a ridge on one sideHowever the degree of curvature observed precludes this ele-ment from representing a long bone and the circular cross-section excludes it from being a rib
The pelvis of FMNH PR 2444 (Fig 12) resembles those ofother traversodontids in general morphology The ilium bears anarrow anteriorly directed iliac blade curving outward distallyThe anterior edge of the iliac blade slopes upward at a roughly45deg angle similar to Luangwa (Kemp 1980) but unlike Ex-aeretodon in which the anterior edge of the iliac blade curvesbackwards (Bonaparte 1963) or Pascualgnathus and Masseto-gnathus in which it is rounded (Bonaparte 1966 Jenkins 1970)The supraacetabular buttress at the base of the ilium is located inan anterodorsal position relative to the acetabulum as in other
traversodontids but unlike earlier cynodonts in which it is lo-cated dorsally (Jenkins 1970) The acetabular buttresses of thethree pelvic bones are largely confluent forming a fairly uniformlsquoliprsquo around the acetabulum as in Exaeretodon (Bonaparte1963) On the contrary in Luangwa the supraacetabular ischialand pubic buttresses are highly distinct spaced 6ndash8 mm apart andwithout confluence of their acetabular facets The pelvis ofFMNH PR 2444 does have a small depression between the su-praacetabular and ischial buttresses (also present in Ex-aeretodon Bonaparte 1963) but this is not as developed as inLuangwa The posterior extent of the iliac blade is broken inFMNH PR 2444 but this region is preserved in another isolatedright pelvis field number 8-31-98-387 (Fig 13) probably pertain-ing to Menadon besairiei The posterior process of the iliac bladein this specimen is relatively short and directed away from thedorsal edge of the ischium as in Exaeretodon (Bonaparte 1963)but unlike Massetognathus and Luangwa in which the posteriorprocess is long and extends roughly horizontally over the dorsaledge of the ischium (Jenkins 1970 Kemp 1980) and evencurves downward distally in Luangwa The posterior process ofthe iliac blade in Pascualgnathus is directed away from the is-chium (Bonaparte 1966) as in Exaeretodon and Menadon but isrelatively long as in Massetognathus and Luangwa
The proximal portion of the ischium is broad and dominatedby the ischial buttress The ischium constricts posterior to theproximal portion and expands distally into a spatulate platewhich is thickened at its posterior margin This thickening con-tinues along the dorsolateral surface of the ischium forming aridge that slopes ventrally until reaching the edge of the ischialbuttress The downward slope of this ridge produces a prominentgroove at the dorsal edge of the lateral surface of the ischiumthought to represent the origin of the ischio-trochantericusmuscle (Kemp 1980) The constriction of the ischium in Mena-don is like that of Luangwa albeit even more extreme in itsventral concavity but unlike that of Exaeretodon in which themedial and distal ends of the ischium are broadly united Theischium in general is a proportionally shorter element in Ex-aeretodon than in other traversodontids including Menadon(Bonaparte 1963) The anteroventral portion of the ischium thatconnects to the pubis is not preserved in either of the referredMenadon pelves
The pubis is the most incompletely preserved pelvic elementThe acetabular contribution of the pubis is smaller than that ofthe ischium or ilium Distal to the acetabulum the pubis is con-stricted indicating that the connection between the acetabularregion and the pubic plate was considerably narrower than inExaeretodon (Bonaparte 1963) but close akin to the pubis ofLuangwa (Kemp 1980) The pubic plate itself is not preservedThe ventral outline of the ischium and what is known of thepubis indicate that the obturator foramen was relatively larger inMenadon (Fig 14) than in Exaeretodon (Bonaparte 1963)
DISCUSSION
Comparison with the Holotype of Menadon besairiei
FMNH PR 2444 differs in several significant ways from theholotype of Menadon besairiei as previously described most no-tably in the possession of an internarial bar (initially consideredabsent in M besairiei) and four upper incisors (reported as threein M besairiei but with the caveat that the homologies of thefourth and fifth anterior tooth loci were problematic Flynn et al2000) However upon further study we interpret the lack of acontinuous internarial bar in the holotype as breakage ratherthan the natural condition of the skull (a prominent medial pro-tuberance is present on the ventral rim of the nares in UA10601) Regarding the incisor count it should be noted that theholotype of Menadon besairiei displays an unusual set of anterior
JOURNAL OF VERTEBRATE PALEONTOLOGY VOL 28 NO 2 2008454
Dow
nloa
ded
by [
UZ
H H
aupt
bibl
ioth
ek
Zen
tral
bibl
ioth
ek Z
uumlric
h] a
t 05
51 1
0 Se
ptem
ber
2013
FIGURE 10 Right scapulocoracoids of FMNH PR 2444 A stereopair of right scapulocoracoid in medial view B stereopair and interpretivedrawing of right scapulocoracoid in lateral view Scale bars equal 5 cm Abbreviationsmdashac acromion process C coracoid c gl coracoid contributionto the glenoid fossa c tr tubercle for coracoid attachment of triceps PC procoracoid pc f procoracoid foramen S scapula s gl scapularcontribution to the glenoid fossa
KAMMERER ET ALmdashCYNODONT FROM MADAGASCAR 455
Dow
nloa
ded
by [
UZ
H H
aupt
bibl
ioth
ek
Zen
tral
bibl
ioth
ek Z
uumlric
h] a
t 05
51 1
0 Se
ptem
ber
2013
teeth for a traversodontid As mentioned above Flynn et al(2000) noted that the homologies of these teeth were difficult todetermine Of the five anterior teeth preserved in the holotypethe anteriormost three are ldquoundoubtedly incisorsrdquo although I3 isldquounusually large and is caniniformrdquo (Flynn et al 2000423) Therobust recurved fourth tooth in the series was considered torepresent the canine with the subsequent tooth interpreted as anolder canine in the process of replacement Although FMNH PR2444 is edentulous the alveoli on the left side of the skull are wellpreserved and indicate an anterior tooth set identical to the ho-lotype The alveolar morphology indicates that the fifth toothwas canted anteriorly as in the holotype This suggests that themysterious fifth tooth is not the result of a transient period in thedental replacement history of M besairiei as suggested by Flynnet al (2000) but rather is the norm This leaves two possibilities1) M besairiei persistently had two canines This condition isunknown in cynodonts 2) The canine-like fourth tooth actuallyrepresents an incisor and the fifth tooth is the true canine Wefavor the latter explanation In FMNH PR 2444 the first fouralveoli are nearly identical in size and shape However the fifthalveolus while similar in anteroposterior length to its predeces-sors is strongly compressed labiolingually with an anteroposte-rior lengthlabiolingual width ratio of 044 (compared to 080 inthe fourth alveolus and 084ndash85 for I2ndash3) We interpret thismarked difference in alveolar ratio as indicating the separationbetween incisors and canine The canine-like I4 would then rep-resent the endpoint in a series of large increasingly caniniformincisors Supporting this interpretation recurved incisors are alsoobserved in juvenile Exaeretodon (eg MCP 3843 PV see Ab-dala et al 2002)
There are several legitimate differences between FMNH PR2444 and the holotype of Menadon besairiei but we take a con-servative taxonomic stance and argue that these differences rep-resent intraspecific variation and preservational artifacts FMNHPR 2444 represents a larger animal than UA 10601 The snoutlength (measured as the distance between the anterior edge ofthe orbit and a vertical plane through the anterior tip of thepremaxilla) of FMNH PR 2444 is 911 mm while that of UA10601 is 801 mm As such proportional differences between thetwo skulls may be attributable to ontogeny Variations in FMNHPR 2444 compared to the holotype that probably represent on-togentic differences include wider longer and deeper paraca-nine fossae a much deeper ventral zygomatic flange (althoughthe lower border of the flange is slightly abraded in the holotypeof Menadon besairiei there is no evidence that a major flangewas broken off) a proportionally shorter and more circular out-line to the posterior border of the maxilla below the zygomaticprocess and a more deeply emarginated posterior squamosal(ie a more prominent laterally projecting flange)
FMNH PR 2444 has a proportionally shorter more robustsnout and larger orbits than the holotype While these charactersare certainly variable in cynodonts it is unusual that the largerspecimen is shorter-snouted and larger-eyed contra the usualpattern The proportional differences between the two skulls areminor however (orbit diametersnout length is 042 for FMNHPR 2444 and 041 for the holotype) and are partially attributableto deformation (shear in FMNH PR 2444 and breakage of thepostorbital bar in the holotype) The following differences canalso probably be attributed to preservational artifact only fivepostcanine alveoli are preserved in FMNH PR 2444 while theholotype has eight postcanines The anteriormost three in theholotype however are tiny and peg-like As the area of themaxilla between the canine alveolus and the first preserved post-canine alveolus is poorly preserved in FMNH PR 2444 the ab-sence of these postcanines may be artifactual However it is alsopossible that the lower postcanine count of FMNH PR 2444compared to the holotype is an ontogenetic effect as postcaninecount is known to decrease with increasing skull size in Ex-
FIGURE 11 Left scapulocoracoids of FMNH PR 2444 A left scapu-locoracoid in medial view B left scapulocoracoid in lateral view Scalebars equal 5 cm
JOURNAL OF VERTEBRATE PALEONTOLOGY VOL 28 NO 2 2008456
Dow
nloa
ded
by [
UZ
H H
aupt
bibl
ioth
ek
Zen
tral
bibl
ioth
ek Z
uumlric
h] a
t 05
51 1
0 Se
ptem
ber
2013
FIGURE 12 Right pelvis of FMNH PR 2444 A stereopair of right pelvis in medial view B stereopair and interpretive drawing of right pelvis inlateral view Scale bars equal 5 cm Abbreviationsmdashbpp base of posterior process of ilium ib iliac blade IL ilium IS ischium ob f obturatorforamen P pubis
KAMMERER ET ALmdashCYNODONT FROM MADAGASCAR 457
Dow
nloa
ded
by [
UZ
H H
aupt
bibl
ioth
ek
Zen
tral
bibl
ioth
ek Z
uumlric
h] a
t 05
51 1
0 Se
ptem
ber
2013
aeretodon (Abdala et al 2002) The orbit of FMNH PR 2444 isless upwardly directed than that of the holotype (being at anangle of sim65deg to a sagittal plane through the midline of the skullas opposed to sim45deg in the holotype) This difference in orbitalangle is probably attributable to postmortem deformation as theskull of FMNH PR 2444 is noticeably sheared to the front leftThe appearance of a wider squarer incisor arcade and broader
anterior palate in FMNH PR 2444 can also be attributed to thisshear Finally the occipital region of FMNH PR 2444 is alto-gether taller and narrower than that of the holotype For ex-ample the foramen magnum in FMNH PR 2444 approximates arounded isosceles triangle in shape while the foramen magnumof the holotype is more ovoid These differences can also beattributed to deformation Altogether the differences between
FIGURE 13 Stereopairs of field number 8-31-98-387 an isolated right pelvis in A medial and B lateral view Scale bars equal 5 cm
JOURNAL OF VERTEBRATE PALEONTOLOGY VOL 28 NO 2 2008458
Dow
nloa
ded
by [
UZ
H H
aupt
bibl
ioth
ek
Zen
tral
bibl
ioth
ek Z
uumlric
h] a
t 05
51 1
0 Se
ptem
ber
2013
FMNH PR 2444 and the holotype fall within the range of intra-specific variation recorded for other gomphodont cynodontssuch as the traversodontids Massetognathus pascuali (Abdalaand Giannini 2000 Kammerer pers obs) and Exaeretodon(Abdala et al 2002) and the diademodontid Diademodon tetra-gonus (Grine et al 1978 Bradu and Grine 1979)
Phylogenetic Analysis
Utilizing the new information afforded by FMNH PR 2444and a reexamination of the holotype (UA 10601) Menadon be-sairiei was recoded in the Gondwanan gomphodont data matrixof Abdala et al (2006) Abdala et alrsquos (2006) data matrix en-compassed 18 taxa and 43 characters For the present analysis anadditional character ldquoincisor procumbencyrdquo (states 0-absent1-present) was added to Abdala et alrsquos (2006) data set (see
Appendix for coding of this character) Several traversodontidtaxa (eg lsquoScalenodonrsquo hirschsoni) have relatively large incisorsbut only in Menadon Scalenodontoides and Exaeretodon are theincisors procumbent Three of Abdala et alrsquos (2006) originalcodings for Menadon besairiei characters 0 (ldquonumber of upperincisorsrdquo) 5 (ldquoposition of paracanine fossa in relation to theupper caninerdquo) and 21 (ldquointernarial barrdquo) were changed to in-corporate data from the new specimen (see Appendix) Addi-tionally further examination of the holotype (UA 10601) ofMenadon besairiei allowed us to code characters 16ndash18 20 38and 40 (morphology of the lower postcanines all marked as ldquordquoin Abdala et alrsquos analysis) Parsimony searches were performedin PAUP v40b10 using a branch-and-bound search Addition-ally a bootstrap analysis (also using branch-and-bound) was per-formed using 1000 replicates
A total of 34 most parsimonious trees of length 103 werefound with the scores CI0573 RI0725 RC0415 andHI0427 The strict consensus tree (Fig 15A) did not recovera polytomy between Menadon Exaeretodon and Scalenodont-oides as in Abdala and Ribeiro (2003) or a (Menadon + Ex-aeretodon) sister-group relationship as in Abdala et al (2006)Rather Menadon was recovered as the sister taxon of the (Ex-aeretodon + Scalenodontoides) clade supporting Flynn et alrsquos(2000) original placement of this taxon There is strong bootstrapsupport (88) for Traversodontidae but within traversodontidsthe only strongly supported subclade is (Gomphodontosuchus +(Menadon + (Scalenodontoides + Exaeretodon))) with a boot-strap value of 80 Within this clade the (Menadon + (Scalen-odontoides + Exaeretodon)) grouping is supported by a boot-strap value of 83 and the (Scalenodontoides + Exaeretodon)grouping is supported by a bootstrap value of 94 (Fig 15B) Inaddition to the positioning of Menadon the strict consensus tree(Fig 15A) differs from the topology of Abdala et al (2006) inrecovering a sister-group relationship between Massetognathusand Santacruzodon rather than having Massetognathus andSantacruzodon as successive outgroups to the clade containingGomphodontosuchus Menadon Scalenodontoides and Ex-aeretodon The (Massetognathus + Santacruzodon) clade is diag-nosed by the absence of an anterolingual cusp in the upper post-canines denticulated incisor cutting margins and the presence ofan anterior cingulum in the upper postcanines
FIGURE 14 Reconstruction of the pelvis of Menadon besairiei basedon information from FMNH PR 2444 and field number 8-31-98-387Dotted area not preserved in either specimen reconstruction based onPascualgnathus (Bonaparte 1966) and Exaeretodon (Bonaparte 1963)
FIGURE 15 Cladogram of traversodontid relationships based on revised data set of Abdala et al (2006) showing A strict consensus tree and B50 majority rule tree Numbers at nodes of majority rule tree represent bootstrap values
KAMMERER ET ALmdashCYNODONT FROM MADAGASCAR 459
Dow
nloa
ded
by [
UZ
H H
aupt
bibl
ioth
ek
Zen
tral
bibl
ioth
ek Z
uumlric
h] a
t 05
51 1
0 Se
ptem
ber
2013
Relationships of Menadon besairiei
Menadon besairiei is the sister taxon to (Exaeretodon + Sca-lenodontoides) the latter clade diagnosed by the possession ofthree upper incisors a posteromedial position of the paracaninefossae relative to the canines the absence of an internarial barlarge canines a projected zygomatic process of the jugal and amaxilla that contributes to the margin of the subtemporal fenes-tra (the last three character states represent reversals) The clade(Menadon + (Exaeretodon + Scalenodontoides)) is diagnosed bythe possession of large procumbent incisors The presence offour incisors a primitive retention distinguishes Menadon be-sairiei from Exaeretodon Scalenodontoides and an unnamed tra-versodontid (ldquoTraversodontid Type 1rdquo) from the Santa MariaFormation of Brazil (Abdala et al 2001 Abdala and Ribeiro2002) Gomphodontosuchus is recovered as the sister taxon to(Menadon (Exaeretodon + Scalenodontoides)) as has beenfound in previous studies (Flynn et al 2000 Abdala and Ribeiro2003) The taxon name Gomphodontosuchinae Watson andRomer 1956 (ex Gomphodontosuchidae Watson and Romer1956 first given subfamilial rank in Lehman 1961) is availablefor this clade As circumscribed here Gomphodontosuchinaeconsists of the most inclusive clade containing Gomphodontosu-chus brasiliensis von Huene 1928264 but not Massetognathuspascuali Romer 19672
Menadon besairiei displays a mixture of primitive and derivedcharacters within traversodontids The internarial bar and fourincisors are primitive features whereas the enlargement and pro-cumbency of the incisors are derived features shared with Ex-aeretodon and Scalenodontoides The postcranium shows severalsimilarities to that of Exaeretodon (apparent exclusion of theprocoracoid from the glenoid some features of the pelvis tallcaudal neural spines) but in general displays a rather conserva-tive morphology observed throughout Gomphodontia The mor-phology of the scapulocoracoids is closer akin to diademodontidsor trirachodontids than Exaeretodon and the expanded thoracicrib of Menadon has only slight differences from those of trirach-odontids or early traversodontids like Luangwa The atlas-axiscomplex of Menadon is similar to that of the Early Triassic cyn-odont Cynognathus in possessing a concave axial spine with along posterior process Although most gomphodonts have beenreconstructed with a convex axial spine as in early mammals thisspine is broken in most of the best described gomphodont post-crania (eg Diademodon Brink 1955) and the lsquosaddle-shapedrsquoaxial morphology may be more widely distributed among cyn-odonts than previously realized The new specimens of Menadondescribed in this paper demonstrate that a generalized gompho-dont postcranial morphology was retained deep within the tra-versodontid tree and that the derived morphologies of either ofthe well-known taxa Massetognathus or Exaeretodon should notbe taken as representative of traversodontids in general
ACKNOWLEDGMENTS
This work would not have been possible without the continu-ing support and encouragement of our collaborative programwith the Deacutepartement de Paleacuteontologie et drsquoAnthropologie Bi-ologique Universiteacute drsquoAntananarivo Madagascar under the di-rection of Armand Rasoamiaramanana (Department Chairman)and Gisegravele Ravololonarivo Randria (Head of the PaleontologyLaboratories) We are extremely grateful to the National Geo-graphic Society for their long-term support of the fieldwork as-sociated with this project Additional financial or logistical sup-port for fieldwork was provided by The Field Museum the Uni-versity of California-Santa Barbara and the WWF (WorldwideFund for Nature Madagascar) We acknowledge the Ministegraverede LrsquoEneacutergie et des Mines for authorizing field efforts and for
continuing to facilitate our joint US-Madagascar explorationresearch and education program Steve Goodman and AsminaGandhi generously assisted us in innumerable ways includinghospitality nonpareil We especially thank R Andriantompoha-vana L Razafimanantsoa A Toto Volahy JA Rabarison AGandie and P Vavisaro for assistance in the field during theexpedition in which these new specimens were discovered BillSimpson and Gina Wesley-Hunt for their patient excavation ofthis important material Allison Smith Ray Vodden Lisa Berg-wall and Deborah Wagner for excellent preparation of a chal-lenging specimen Jennifer Botha and Elize Butler (NationalMuseum Bloemfontein) Michael Raath and Fernando Abdala(Bernard Price Institute) Thomas Kemp (Oxford UniversityMuseum of Natural History) Ray Symonds (University Museumof Zoology Cambridge) Bernard Battail (Museacuteum nationaldrsquoHistoire naturelle Paris) Michael Maisch (Universitaumlt Tuumlbin-gen) and Charles Schaff and Farish Jenkins (Museum of Com-parative Zoology Harvard) for access to specimens and AgustiacutenMartinelli Fernando Abdala and James Hopson for helpful re-views of an earlier version of the manuscript
LITERATURE CITED
Abdala F M C Barbarena and J Dornelles 2002 A new species of thetraversodontid cynodont Exaeretodon from the Santa Maria Forma-tion (MiddleLate Triassic) of Southern Brazil Journal of Verte-brate Paleontology 22313ndash325
Abdala F and N P Giannini 2000 Gomphodont cynodonts of theChantildeares Formation the analysis of an ontogenetic sequence Jour-nal of Vertebrate Paleontology 20501ndash506
Abdala F J Neveling and J Welman 2006 A new trirachodontidcynodont from the lower levels of the Burgersdorp Formation(Lower Triassic) of the Beaufort Group South Africa and the cla-distic relationships of Gondwanan gomphodonts Zoological Jour-nal of the Linnean Society 147383ndash413
Abdala F A M Ribeiro and C L Schultz 2001 A rich cynodont faunaof Santa Cruz do Sul Santa Maria Formation (Middle-Late Trias-sic) southern Brazil Neues Jahrbuch fuumlr Geologie und Palaumlontolo-gie Monatshefte 2001669ndash687
Abdala F and A M Ribeiro 2002 Nuevos cinodontes traversodoacuten-tidos (Synapsida-Eucynodontia) de la Formacioacuten Santa Maria(Triaacutesico Medio-Superior) Rio Grande do Sul Brasil RevistaEspantildeola de Paleontologiacutea 17237ndash243
Abdala F and A M Ribeiro 2003 A new traversodontid cynodontfrom the Santa Maria Formation (Ladinian-Carnian) of southernBrazil with a phylogenetic analysis of Gondwanan traversodontidsZoological Journal of the Linnaean Society 139529ndash545
Abdala F and A M S Teixeira 2004 A traversodontid cynodont ofAfrican affinity in the South American Triassic Palaeontologia Af-ricana 4011ndash22
Bonaparte J F 1962 Descripcioacuten del craacuteneo y mandiacutebula de Ex-aeretodon frenguellii Cabrera y su comparacioacuten con Diademodon-tidae Tritylodontidae y los cinodontes sudamericanos Publica-ciones del Museo Municipal de Ciencias Naturales y Tradicional deMar del Plata 1135ndash202
Bonaparte J F 1963 Descripcioacuten del esqueleto postcraneano de Ex-aeretodon (CynodontiandashTraversodontidae) Acta Geoloacutegica Lil-loana 45ndash52
Bonaparte J F 1966 Una nueva ldquofaunardquo Triaacutesica de Argentina (Ther-apsida Cynodontia Dicynodontia) Consideracions filogeneacuteticas ypaleobiogeograacuteficas Ameghiniana 8243ndash296
Bradu D and F E Grine 1979 Multivariate analysis of diademodon-tine crania from South Africa and Zambia South African Journal ofScience 79441ndash448
Brink A S 1955 A study of the skeleton of Diademodon Palaeonto-logia Africana 33-39
Brink A S 1963 Two new cynodonts from the Ntawere Formation inthe Luangwa Valley of Northern Rhodesia Palaeontologia Africana877ndash96
Broom R 1905 On the use of the term Anomodontia Records of theAlbany Museum 1266ndash269
Cabrera A 1943 El primer hallazgo de teraacutepsidos en la ArgentinaNotas del Museo de La Plata Paleontologiacutea 55317ndash331
JOURNAL OF VERTEBRATE PALEONTOLOGY VOL 28 NO 2 2008460
Dow
nloa
ded
by [
UZ
H H
aupt
bibl
ioth
ek
Zen
tral
bibl
ioth
ek Z
uumlric
h] a
t 05
51 1
0 Se
ptem
ber
2013
Chatterjee S 1982 A new cynodont reptile from the Triassic of IndiaJournal of Paleontology 56203ndash214
Crompton A W 1955 On some Triassic cynodonts from TanganyikaProceedings of the Zoological Society of London 125617ndash669
Crompton A W 1972 Postcanine occlusion in cynodonts and tritylo-donts Bulletin of the British Museum (Natural History) Geology2129ndash71
Crompton A W and F Ellenberger 1957 On a new cynodont from theMolteno Beds and the origin of the tritylodontids Annals of theSouth African Museum 441ndash13
Flynn J J J M Parrish B Rakotosaminimanana W F Simpson R LWhatley and A R Wyss 1999 A Triassic fauna from Madagascarincluding early dinosaurs Science 286763ndash765
Flynn J J J M Parrish B Rakotosamimanana L RanivoharimananaW F Simpson and A R Wyss 2000 New traversodontids (Syn-apsida Eucynodontia) from the Triassic of Madagascar Journal ofVertebrate Paleontology 20422ndash427
Gow C E and P J Hancox 1993 First complete skull of the LateTriassic Scalenodontoides (Reptilia Cynodontia) from southern Af-rica In Lucas S G and M Morales (eds) The nonmarine TriassicNew Mexico Museum of Natural History amp Science Bulletin 3161ndash168
Grine F E B D Hahn and C E Gow 1978 Aspects of relativegrowth and variability in Diademodon (Reptilia Therapsida) SouthAfrican Journal of Science 7450ndash58
Hopson J A 1984 Late Triassic traversodont cynodonts from NovaScotia and southern Africa Palaeontologia Africana 25181ndash201
Hopson J A 1985 Morphology and relationships of Gomphodontosu-chus brasiliensis von Huene (Synapsida Cynodontia Tritylodontoi-dea) from the Triassic of Brazil Neues Jahrbuch fuumlr Geologie undPalaumlontologie Monatshefte 1985285ndash299
Hopson J A and J W Kitching 2001 A probainognathian cynodontfrom South Africa and the phylogeny of nonmammalian cynodontsBulletin of the Museum of Comparative Zoology 1565ndash35
Hopson J A and H-D Sues 2006 A traversodont cynodont from theMiddle Triassic (Ladinian) of Baden-Wuumlrttemberg (Germany)Palaumlontologische Zeitschrift 80124ndash129
Huene F von 1928 Ein Cynodontier aus der Trias Brasiliens Zentral-blatt fuumlr Mineralogie Geologie und Palaumlontologie 1928251ndash270
Huene F von 1936 Die fossilen Reptilien des suumldamerikanischenGondwanalandes Ergebnisse der Sauriergrabungen in Suumldbrasilien192829 Lieferung 2 (Cynodontia)93ndash159 Franz F Heine Tuumlbin-gen
Jenkins F A 1970 The Chantildeares (Argentina) Triassic reptile faunaVII The postcranial skeleton of the traversodontid Massetognathuspascuali (Therapsida Cynodontia) Breviora 3521ndash28
Jenkins F A 1971 The postcranial skeleton of African cynodonts prob-lems in the early evolution of the mammalian postcranial skeletonBulletin of the Peabody Museum of Natural History 361ndash216
Kemp T S 1969 The atlas-axis complex of mammal-like reptiles Jour-nal of Zoology 159223ndash248
Kemp T S 1980 Aspects of the structure and functional anatomy of theMiddle Triassic cynodont Luangwa Journal of Zoology 191193ndash239
Kemp T S 1982 Mammal-Like Reptiles and the Origin of MammalsAcademic Press New York
Lehman J-P 1961 Cynodontia pp 140ndash191 in Piveteau J (ed) Traiteacutede Paleacuteontologie Masson et Cie Paris
Osborn 1903 On the primary division of the Reptilia into two sub-classes Synapsida and Diapsida Science 17275ndash276
Owen R 1861 Palaeontology or a systematic summary of extinct ani-mals and their geological relations 2nd Ed Edinburgh
Parrington F R 1934 On the cynodont genus Galesaurus with a noteon the functional significance of the changes in the evolution of thetheriodont skull Annals and Magazine of Natural History 10(13)38ndash67
Parrington F R 1946 On the cranial anatomy of cynodonts Proceed-ings of the Zoological Society of London 116181ndash197
Razafimbelo E 1987 Le basin de Morondava (Madagascar) Synthegravesegeacuteologique et structurale PhD dissertation University Louis Pas-teur Strasbourg France 189 pp
Reisz R R and H-D Sues 2000 Herbivory in Late Paleozoic andTriassic terrestrial vertebrates pp 9ndash41 in Sues H-D (ed) Evo-lution of Herbivory in Terrestrial Vertebrates Cambridge Univer-sity Press New York
Romer A S 1967 The Chantildeares (Argentina) Triassic reptile fauna IIITwo new gomphodonts Massetognathus pascuali and M terugiiBreviora 2641ndash25
Romer A S 1972 The Chantildeares (Argentina) Triassic reptile faunaXVII The Chantildeares gomphodonts Breviora 3961ndash9
Rougier G W J R Wible and J A Hopson 1992 Reconstruction ofthe cranial vessels in the Early Cretaceous mammal Vincelestes neu-quenianus implications for the evolution of the mammalian cranialvascular system Journal of Vertebrate Paleontology 12188ndash216
Rowe T 1988 Definition diagnosis and origin of Mammalia Journal ofVertebrate Paleontology 8241ndash264
Rowe T 1993 Early mammal phylogenetic systematics pp 129ndash145 inSzalay F S M J Novacek and M C McKenna (eds) MammalPhylogeny Vol 1 Mesozoic Differentiation MultituberculatesMonotremes Early Therians and Marsupials Springer-Verlag NewYork
Seeley H G 1894 Researches on the structure organization and clas-sification of the fossil Reptilia Part IX Section 3 On DiademodonPhilosophical Transactions of the Royal Society of London 1851029ndash1041
Seeley H G 1895 Researches on the structure organization and clas-sification of the fossil Reptilia Part IX Section 4 On the Gompho-dontia Philosophical Transactions of the Royal Society of London1861ndash57
Seeley H G 1895 Researches on the structure organization and clas-sification of the fossil Reptilia Part IX Section 5 On the skeletonin new Cynodontia from the Karroo rocks Philosophical Transac-tions of the Royal Society of London 18659ndash148
Sues H-D J A Hopson and N H Shubin 1992 Affinities of Sca-lenodontoides plemmyridon Hopson 1984 (Synapsida Cynodontia)from the Upper Triassic of Nova Scotia Journal of Vertebrate Pa-leontology 12168ndash171
Sues H-D and P E Olsen 1990 Triassic vertebrates of Gondwananaspect from the Richmond basin of Virginia Science 2491020ndash1023
Swofford D L 2003 PAUP v 40b10 Sinauer Associates Inc Pub-lishers Sunderland Massachusetts
Watson D M S and A S Romer 1956 A classification of therapsidreptiles Bulletin of the Museum of Comparative Zoology 11435ndash89
Submitted September 20 2007 accepted November 3 2007
KAMMERER ET ALmdashCYNODONT FROM MADAGASCAR 461
Dow
nloa
ded
by [
UZ
H H
aupt
bibl
ioth
ek
Zen
tral
bibl
ioth
ek Z
uumlric
h] a
t 05
51 1
0 Se
ptem
ber
2013
APPENDIX Data matrix used in the phylogenetic analysesThis is a revised version of the matrix of Abdala et al (2006) with the following changes character 43 [Incisor procumbency (0) absent (1) present]added Menadon recoded for characters 0 (Number of upper incisors was 1 ldquothreerdquo changed to 0 ldquofourrdquo) 5 (Position of paracanine fossae in relationto the upper canine was changed to 1 ldquomedialrdquo) 16 (Overall morphology of lower postcanines in occlusal view was changed to 3 ldquoquadran-gularrdquo) 17 (Position of the transverse crest in lower postcanines was changed to 1 ldquoanteriorrdquo) 18 (Number of cusps in the transverse crest of thelower postcanines was changed to 0 ldquotwordquo) 20 (Anterior cingulum in the lower postcanines was changed to 2 ldquoabsentrdquo) 21 (Internarial barwas 1 ldquoabsentrdquo changed to 0 ldquopresentrdquo) 38 (Posterior cingulum in the lower postcanines was changed to 1 ldquoabsentrdquo) and 40 (Deep occlusalbasins in the postcanines was changed to 1 ldquopresentrdquo) Langbergia recoded for character 34 (Incisor cutting margins was 0 ldquoserratedrdquo changedto 1 ldquosmoothly ridgedrdquo) based on the description of the incisors in Abdala et al (2006) Gomphodontosuchus recoded for character 31 (Maxilla inthe margin of the subtemporal fenestra was 1 ldquoincludedrdquo changed to 0 ldquoexcludedrdquo) based on von Huene (1936) and Kammerer (pers obs of theholotype) contra Hopsonrsquos (1985) reconstruction lsquoScalenodonrsquo hirschsoni recoded for character 39 (Anterior cingulum in the upper postcanines was1 ldquoabsentrdquo changed to 0 ldquopresentrdquo) based on Abdala (pers comm 2006) Trirachodon recoded for character 30 (Ectopterygoid was changedto 0 ldquopresentrdquo) based on Abdala (pers comm 2006) and Traversodon recoded for character 15 (Number of cusps in the sectorial border of the upperpostcanines was 0 ldquomore than twordquo changed to 1 ldquotwordquo) based on Abdala (pers comm 2006) and Kammerer (pers obs)
1111111111 2222222222 3333333333 44440123456789 0123456789 0123456789 0123456789 0123
Cynognathus 00000000 000 000000000 00000000 000Diademodon 0000001000 0000100 000010A000 0000000100 0100Trirachodon 000000100B 1A002201 001A100010 0001001200 0110Langbergia 0000001000 10000201 0000101010 0001101200 0110Cricodon 0100001000 11000201 0010100 0001200 0110Andescynodon 0000002001 01013101 101120010 0112311 1100Massetognathus 00111121A2 1110003100 2011200011 1011212300 1100Exaeretodon 1110122112 010103101 2111110201 1111112311 1101Luangwa 0000001002 11010A3100 101010010 01002300 1100Scal angustifrons 000000100B 1111023100 10102101 00002301 1100Scal hirschsoni 11110202 1100113100 1010 1112300 1100Traversodon 0000002012 1101113100 201010 011230 1100Gomphodontosuchus 0011112112 0103101 2010 0112311 1100Pascualgnathus 1000001000 000231 1100001 0011230 1100Scalenodontoides 1112212 01003101 21111021 1212311 1101Menadon 0111112112 01012310 20121020 0102311 1101Dadadon 00002112 111112 013 11231 1100Santacruzodon 00002112 1110003101 201300 212300 1100
A(01) B(02)
JOURNAL OF VERTEBRATE PALEONTOLOGY VOL 28 NO 2 2008462
Dow
nloa
ded
by [
UZ
H H
aupt
bibl
ioth
ek
Zen
tral
bibl
ioth
ek Z
uumlric
h] a
t 05
51 1
0 Se
ptem
ber
2013
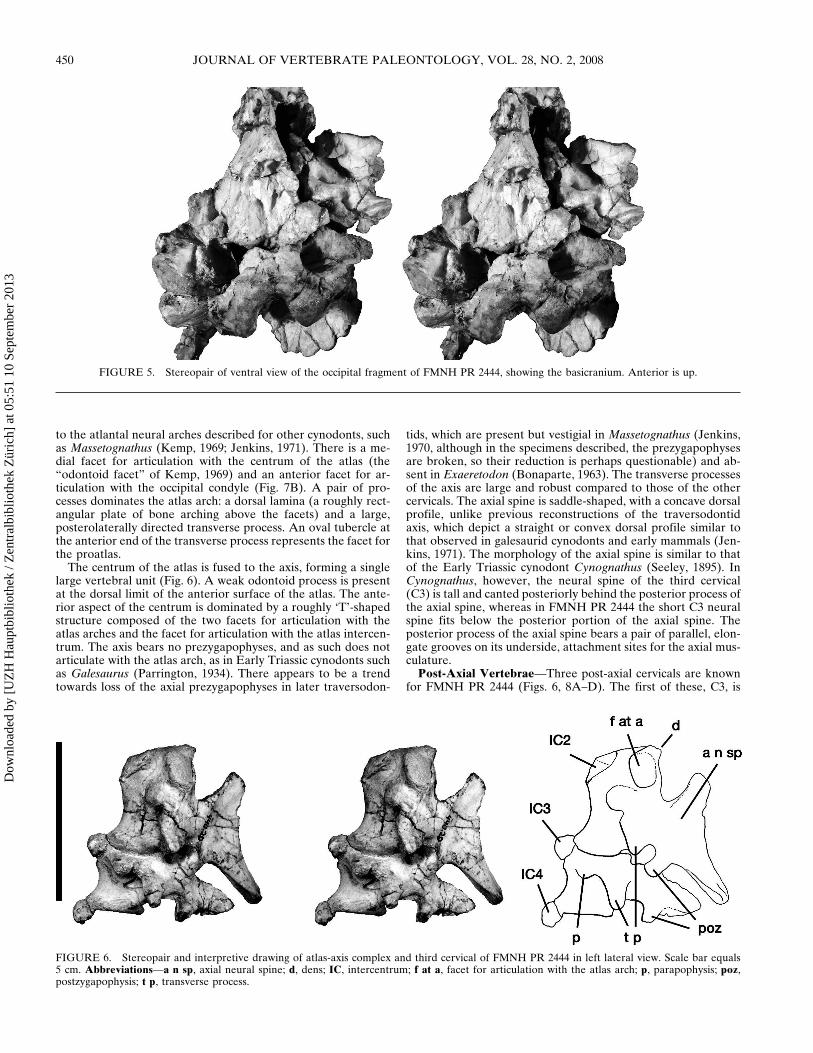
to the atlantal neural arches described for other cynodonts suchas Massetognathus (Kemp 1969 Jenkins 1971) There is a me-dial facet for articulation with the centrum of the atlas (theldquoodontoid facetrdquo of Kemp 1969) and an anterior facet for ar-ticulation with the occipital condyle (Fig 7B) A pair of pro-cesses dominates the atlas arch a dorsal lamina (a roughly rect-angular plate of bone arching above the facets) and a largeposterolaterally directed transverse process An oval tubercle atthe anterior end of the transverse process represents the facet forthe proatlas
The centrum of the atlas is fused to the axis forming a singlelarge vertebral unit (Fig 6) A weak odontoid process is presentat the dorsal limit of the anterior surface of the atlas The ante-rior aspect of the centrum is dominated by a roughly lsquoTrsquo-shapedstructure composed of the two facets for articulation with theatlas arches and the facet for articulation with the atlas intercen-trum The axis bears no prezygapophyses and as such does notarticulate with the atlas arch as in Early Triassic cynodonts suchas Galesaurus (Parrington 1934) There appears to be a trendtowards loss of the axial prezygapophyses in later traversodon-
tids which are present but vestigial in Massetognathus (Jenkins1970 although in the specimens described the prezygapophysesare broken so their reduction is perhaps questionable) and ab-sent in Exaeretodon (Bonaparte 1963) The transverse processesof the axis are large and robust compared to those of the othercervicals The axial spine is saddle-shaped with a concave dorsalprofile unlike previous reconstructions of the traversodontidaxis which depict a straight or convex dorsal profile similar tothat observed in galesaurid cynodonts and early mammals (Jen-kins 1971) The morphology of the axial spine is similar to thatof the Early Triassic cynodont Cynognathus (Seeley 1895) InCynognathus however the neural spine of the third cervical(C3) is tall and canted posteriorly behind the posterior process ofthe axial spine whereas in FMNH PR 2444 the short C3 neuralspine fits below the posterior portion of the axial spine Theposterior process of the axial spine bears a pair of parallel elon-gate grooves on its underside attachment sites for the axial mus-culature
Post-Axial VertebraemdashThree post-axial cervicals are knownfor FMNH PR 2444 (Figs 6 8AndashD) The first of these C3 is
FIGURE 5 Stereopair of ventral view of the occipital fragment of FMNH PR 2444 showing the basicranium Anterior is up
FIGURE 6 Stereopair and interpretive drawing of atlas-axis complex and third cervical of FMNH PR 2444 in left lateral view Scale bar equals5 cm Abbreviationsmdasha n sp axial neural spine d dens IC intercentrum f at a facet for articulation with the atlas arch p parapophysis pozpostzygapophysis t p transverse process
JOURNAL OF VERTEBRATE PALEONTOLOGY VOL 28 NO 2 2008450
Dow
nloa
ded
by [
UZ
H H
aupt
bibl
ioth
ek
Zen
tral
bibl
ioth
ek Z
uumlric
h] a
t 05
51 1
0 Se
ptem
ber
2013
preserved in articulation with the axis (Fig 6) The two isolatedcervicals bear successively taller and anterposteriorly wider neu-ral spines (Fig 8AndashD) Based on their staggered size and failureto articulate we suggest that they represent C5 and C7 (assumingthat this specimen possessed the generalized cynodont cervicalnumber of seven) The cervicals display robust transverse pro-cesses which are directed laterally and slightly anteriorly A par-apophysis lies below the anterior edge of the transverse processalong the antero-lateral edge of the centrum No cervical ribs arepreserved but their presence can be inferred from the par-apophyses The notochordal concavity is as in Massetognathussurrounded by a raised rim at each end of the centrum (Jenkins1970) No anapophyses are present The neural spines for allthree post-axial cervicals are tapering and slightly curved poste-riorly The neural spine in C3 is relatively short and wide(thereby fitting underneath the posterior overhang of the axialspine see Fig 6) while that of C5 is tall and narrow The spinefor C7 is taller and anteroposteriorly wider than that of C5 andthe dorsal edge is more rounded similar to the condition in thedorsal vertebrae
Eight post-cervical presacral vertebrae are known for thisspecimen We will refer to them here simply as dorsal vertebraegiven the incompleteness of the vertebral column in the speci-men and the uncertainty concerning the thoraciclumbar transi-tion in this taxon However it is probable that they representthoracic vertebrae (see below) In most cynodonts the lumbar
vertebrae are synostosed to specialized ribs with overlappingelements In earlier taxa (eg Thrinaxodon Cynognathus Lu-angwa Jenkins 1971 Kemp 1980) expanded costal plates arepresent on both thoracic and lumbar ribs In Massetognathus thelumbar ribs consist of a pair of prongs one directed anteriorlybearing a facet for articulation with the previous rib and theother directed posteriorly (Jenkins 1970) In Exaeretodon allpresacral ribs are identical of simple morphology and not syn-ostosed to the vertebrae (Bonaparte 1963) A single isolated rib(Fig 9B) in FMNH PR 2444 demonstrates that some of the ribsin Menadon besairiei were expanded However all preservedvertebrae in this specimen have facets for articulation with theribs rather than breakage points where they would have beenfused to ribs As such it is probable that all of the vertebraepreserved in this specimen represent thoracics although we can-not discount the possibility that as in Exaeretodon no presacralvertebrae were fused to ribs
The neural spines in all of the dorsal vertebrae are elongateposteriorly inclined and roughly rectangular in lateral view witha rounded laterally expanded platform dorsally (Fig 8EndashJ)Costal foveae are present at the anterior and posterior edges ofeach centrum Of the eight dorsal vertebrae there are two ar-ticulated sets and three isolated elements (two from 9-4-98-440and one from 9-2-98-422) The first isolated vertebra (Fig 8E F)has a taller neural spine and shorter centrum than the otherdorsal vertebrae It is most similar in proportions to the isolated
FIGURE 7 Stereopairs and interpretive drawings of left atlas arch of FMNH PR 2444 in A lateral view Anterior is left B medial view Anterioris right Scale bars equal 1 cm Abbreviationsmdashafa anterior facet for articulation with the occipital condyle dl dorsal lamina mfa medial facet forarticulation with the atlas centrum (odontoid facet) pfa tubercle forming the facet for articulation with the proatlas tp transverse process
KAMMERER ET ALmdashCYNODONT FROM MADAGASCAR 451
Dow
nloa
ded
by [
UZ
H H
aupt
bibl
ioth
ek
Zen
tral
bibl
ioth
ek Z
uumlric
h] a
t 05
51 1
0 Se
ptem
ber
2013
FIGURE 8 Vertebrae of FMNH PR 2444 Cervical vertebra 5 in A left lateral and B anterior views Cervical vertebra 7 in C left lateral andD anterior views Isolated dorsal vertebra from anterior region of the series in E left lateral and F anterior views G stereopair and interpretivedrawing of articulated set of two dorsal vertebrae from near the midpoint of the series in left lateral view Anterior is up H articulated set of threedorsal vertebrae from anterior region of the series in left and right lateral views I isolated vertebra from near the midpoint of the series (from fieldnumber 9-2-98-422) J isolated vertebra from posterior region of the series Sacral vertebra in K dorsal view (the distal region of another sacral ribwhich is identical in shape to that of the intact sacral rib albeit slightly larger is also shown in this figure) and L left lateral view Caudal vertebraeM articulated set of two vertebrae from the anterior region of the series showing the lsquowingrsquo-shaped caudal ribs in dorsal view Anterior is left Nisolated caudal vertebra showing tall neural spine in left lateral view O isolated caudal vertebra showing tall neural spine in posterior view Particulated set of four vertebrae from near the end of the caudal series in dorsal view Anterior is left Scale bars equal 1 cm Abbreviationsmdasha cof anterior costal fovea n sp neural spine p co f posterior costal fovea poz postzygapophysis prz prezygapophysis
JOURNAL OF VERTEBRATE PALEONTOLOGY VOL 28 NO 2 2008452
Dow
nloa
ded
by [
UZ
H H
aupt
bibl
ioth
ek
Zen
tral
bibl
ioth
ek Z
uumlric
h] a
t 05
51 1
0 Se
ptem
ber
2013
cervicals and was probably located near the front of the dorsalseries The first articulated set consists of three vertebrae (Fig8H) The middle vertebra in this set is broken from the anteriorbase of the neural spine to the ventral midpoint and most of theright side of the anterior half of the centrum is missing This setof vertebrae probably also came from the anterior region of theseries as their neural spines are longer and narrower than allvertebrae other than the one described above The second ar-ticulated set (Fig 8G) consists of two well-preserved vertebraethat based on comparisons with Massetognathus and Ex-aeretodon (Jenkins 1970 Bonaparte 1963) probably came fromnear the midpoint of the dorsal series The second isolated ver-tebra (Fig 8I) from 9-2-98-422 is similar to the members of thesecond set The third isolated vertebra (Fig 8J) appears to befrom the posterior end of the series based on the relatively shortneural spine and reduced posterior costal fovea
One sacral vertebra (Fig 8K L) is known consisting of awell-preserved centrum neural spine and the right sacral rib(the left is broken off) The centrum is shorter (both anteropos-teriorly and dorsoventrally) than those of the dorsal vertebraeand the neural spine is shorter and more rounded in profileAlso the neural spine is directed vertically unlike those of thedorsal vertebrae which are canted posteriorly The zygapophy-ses are poorly developed The sacral rib is fused to a singlesynapophysis and projects anterolaterally The rib narrows to-wards the middle and expands distally terminating in a widerectangular facet where it would meet the ilium (Fig 8K)
Seven caudal vertebrae are known The caudal vertebrae aredistinctive in having very tall narrow neural spines Some EarlyTriassic cynodonts possess blunt neural spines of moderateheight on the proximal caudals (see for example Thrinaxodonin Jenkins 1971 and Diademodon in Brink 1955) Massetogna-thus represents an extreme condition wherein the caudal neuralspines are greatly reduced (Jenkins 1970) Very tall neuralspines are seen in Exaeretodon (Bonaparte 1963) and thosespines are unusual in that they do not diminish in height until thelast caudals The caudal vertebrae of FMNH PR 2444 consist ofa pair of articulated vertebrae most likely from near the sacrum(Fig 8M) an isolated caudal lacking postzygapophyses and ribsbut with the neural spine (Fig 8N O) and a quartet of smaller
articulated elements (Fig 8P) The anterior caudals have nar-rower neural spines and shorter zygapophyses than the subse-quent vertebrae in the series The caudal ribs attached to thispair have a crescentic distal region that curves anteriorly pro-ducing a lsquowingrsquo-shape Only a portion of the right rib of theisolated caudal is preserved but the lsquowingrsquo-shaped morphologyof the near-sacral caudal ribs does not appear to have been pre-sent In the posteriormost four vertebrae caudal ribs are pre-served on the left side of the first and the right side of the secondand third members On the first of these vertebrae the rib isshort curved posteriorly (but not lsquowingrsquo-shaped) and taperingRibs of the second and third vertebrae are less curved resem-bling posteriorly directed rods The zygapophyses of both theisolated caudal vertebra and the quartet of small caudals areanteroposteriorly elongate becoming increasingly so posteriorly
Thoracic RibsmdashNine thoracic ribs are known from 9-4-98-440(four left four right and an indeterminate splint of bone Fig9A indeterminate bone not illustrated) These elements re-semble the flattened ribs lacking costal plates of Exaeretodon(Bonaparte 1963) and the anterior thoracic ribs of Massetogna-thus and Luangwa (Jenkins 1970 Kemp 1980) The tuberculumand capitulum are separated by a shallow depression The tuber-culum has a shorter base than the capitulum but their proximalfaces are subequal in size An additional isolated left rib (Fig9B) from 9-2-98-422 is expanded indicating that unlike Ex-aeretodon Menadon rib morphology was heterogeneous alongthe length of the dorsal series This rib closely resembles theposterior thoracic ribs described for Luangwa (Kemp 1980fig16) Pascualgnathus (Bonaparte 1966fig 7) and the trirach-odontid Cricodon (Crompton 1955 Fig 4) This rib is con-stricted at the base of the head then expands distally with an-terior and posterior costal plates As in Luangwa the anteriorcostal plate is shallow and evenly sloping proximally and distallywhile the posterior costal plate is falcate with a concave edgedistally The distal expansion of this rib occurs at an angle of sim30degto the long axis of the rib head Despite their overall similaritythe expanded rib in FMNH PR 2444 does exhibit some differ-ences from the posterior thoracic rib of Luangwa The expandedrib of Menadon has a longer shaft between the rib head and thecostal plates and a shorter capitulum lacks a ridge on the pos-
FIGURE 9 A anterior thoracic ribs of FMNH PR 2444 in anterior view (indeterminate rib not figured) B expanded posterior thoracic rib ofFMNH PR 2444 in anterior view Scale bars equal 1 cm
KAMMERER ET ALmdashCYNODONT FROM MADAGASCAR 453
Dow
nloa
ded
by [
UZ
H H
aupt
bibl
ioth
ek
Zen
tral
bibl
ioth
ek Z
uumlric
h] a
t 05
51 1
0 Se
ptem
ber
2013
terior costal plate and has an anteroposteriorly shorter anteriorcostal plate Additionally the posterior margin of the costal plateis less curved than that of Luangwa These differences couldindicate that the expanded rib of Menadon is from a more pos-terior position in the series than the rib from Luangwa as re-duced curvature and increasing confluence of the tuberculumand capitulum are characteristic of lumbar ribs in traversodon-tids (Jenkins 1970 Kemp 1980) However the length of theshaft is at odds with this interpretation and in Cricodon andPascualgnathus the more posterior ribs are not more similar tothe element from FMNH PR 2444 These differences probablyrepresent legitimate morphological distinctions between the ribsof earlier gomphodonts and Menadon The head of the expandedrib is well defined and unbroken indicating that it was not syn-ostosed to a vertebra
Appendicular Skeleton
The appendicular skeleton is represented by both scapulocora-coids and a fragment of a clavicle from field number 9-4-98-440and a partial right pelvis from field number 9-2-98-422 An ad-ditional partial right pelvis is represented from field number8-31-98-387 The scapulocoracoids (Figs 10 11) are very similarto those described for other traversodontids as well as cynog-nathids and diademodontids (Jenkins 1970 1971) The scapularinfraspinous fossa has out-turned anterior and posterior edgesThe anterior edge of the scapula is rounded and curves laterallyforming the scapular spine At the lower end of the spine theacromion forms a prominent process (broken in the left scapula)Below this process the scapula is constricted and then expandsventrally to form the dorsal margin of the articular glenoid Theglenoid fossa is in general morphology very similar to that ofother Triassic cynodonts (Jenkins 1970 1971 Kemp 1980) It isposteroventrally oriented with roughly equal contributions fromthe scapular and coracoid glenoid facets The coracoid glenoidfacet is roughly triangular while the scapular glenoid facet issemiovoid as in other cynodonts In anterior or posterior viewthe fossa appears smoothly crescentic The procoracoid is aslightly curved trapezoidal plate of bone It does not appear tocontribute to the glenoid fossa The procoracoid foramen is lo-cated midway up the height of the procoracoid at a point directlybelow the base of the acromion process The coracoid is semi-crescentic curving posteriorly from a broad base The coracoidin Menadon is longer and more falcate than those described forExaeretodon (Bonaparte 1963) and Luangwa (Kemp 1980 al-though the distal tip of both coracoids is broken in the specimenof Luangwa [TSK 121] described by Kemp and may have beenlonger than reconstructed) but agrees closely with those of Mas-setognathus and cynognathidsdiademodontids (Jenkins 19701971) The distal coracoid tubercle for attachment to the tricepsis strongly curved upwards similar to the condition in Masseto-gnathus (Jenkins 1970) and Diademodon (Brink 1955)
A small fragment of bone resembling a curved cylinder almostcertainly represents part of a clavicle It has no distinguishingfeatures other than a groove associated with a ridge on one sideHowever the degree of curvature observed precludes this ele-ment from representing a long bone and the circular cross-section excludes it from being a rib
The pelvis of FMNH PR 2444 (Fig 12) resembles those ofother traversodontids in general morphology The ilium bears anarrow anteriorly directed iliac blade curving outward distallyThe anterior edge of the iliac blade slopes upward at a roughly45deg angle similar to Luangwa (Kemp 1980) but unlike Ex-aeretodon in which the anterior edge of the iliac blade curvesbackwards (Bonaparte 1963) or Pascualgnathus and Masseto-gnathus in which it is rounded (Bonaparte 1966 Jenkins 1970)The supraacetabular buttress at the base of the ilium is located inan anterodorsal position relative to the acetabulum as in other
traversodontids but unlike earlier cynodonts in which it is lo-cated dorsally (Jenkins 1970) The acetabular buttresses of thethree pelvic bones are largely confluent forming a fairly uniformlsquoliprsquo around the acetabulum as in Exaeretodon (Bonaparte1963) On the contrary in Luangwa the supraacetabular ischialand pubic buttresses are highly distinct spaced 6ndash8 mm apart andwithout confluence of their acetabular facets The pelvis ofFMNH PR 2444 does have a small depression between the su-praacetabular and ischial buttresses (also present in Ex-aeretodon Bonaparte 1963) but this is not as developed as inLuangwa The posterior extent of the iliac blade is broken inFMNH PR 2444 but this region is preserved in another isolatedright pelvis field number 8-31-98-387 (Fig 13) probably pertain-ing to Menadon besairiei The posterior process of the iliac bladein this specimen is relatively short and directed away from thedorsal edge of the ischium as in Exaeretodon (Bonaparte 1963)but unlike Massetognathus and Luangwa in which the posteriorprocess is long and extends roughly horizontally over the dorsaledge of the ischium (Jenkins 1970 Kemp 1980) and evencurves downward distally in Luangwa The posterior process ofthe iliac blade in Pascualgnathus is directed away from the is-chium (Bonaparte 1966) as in Exaeretodon and Menadon but isrelatively long as in Massetognathus and Luangwa
The proximal portion of the ischium is broad and dominatedby the ischial buttress The ischium constricts posterior to theproximal portion and expands distally into a spatulate platewhich is thickened at its posterior margin This thickening con-tinues along the dorsolateral surface of the ischium forming aridge that slopes ventrally until reaching the edge of the ischialbuttress The downward slope of this ridge produces a prominentgroove at the dorsal edge of the lateral surface of the ischiumthought to represent the origin of the ischio-trochantericusmuscle (Kemp 1980) The constriction of the ischium in Mena-don is like that of Luangwa albeit even more extreme in itsventral concavity but unlike that of Exaeretodon in which themedial and distal ends of the ischium are broadly united Theischium in general is a proportionally shorter element in Ex-aeretodon than in other traversodontids including Menadon(Bonaparte 1963) The anteroventral portion of the ischium thatconnects to the pubis is not preserved in either of the referredMenadon pelves
The pubis is the most incompletely preserved pelvic elementThe acetabular contribution of the pubis is smaller than that ofthe ischium or ilium Distal to the acetabulum the pubis is con-stricted indicating that the connection between the acetabularregion and the pubic plate was considerably narrower than inExaeretodon (Bonaparte 1963) but close akin to the pubis ofLuangwa (Kemp 1980) The pubic plate itself is not preservedThe ventral outline of the ischium and what is known of thepubis indicate that the obturator foramen was relatively larger inMenadon (Fig 14) than in Exaeretodon (Bonaparte 1963)
DISCUSSION
Comparison with the Holotype of Menadon besairiei
FMNH PR 2444 differs in several significant ways from theholotype of Menadon besairiei as previously described most no-tably in the possession of an internarial bar (initially consideredabsent in M besairiei) and four upper incisors (reported as threein M besairiei but with the caveat that the homologies of thefourth and fifth anterior tooth loci were problematic Flynn et al2000) However upon further study we interpret the lack of acontinuous internarial bar in the holotype as breakage ratherthan the natural condition of the skull (a prominent medial pro-tuberance is present on the ventral rim of the nares in UA10601) Regarding the incisor count it should be noted that theholotype of Menadon besairiei displays an unusual set of anterior
JOURNAL OF VERTEBRATE PALEONTOLOGY VOL 28 NO 2 2008454
Dow
nloa
ded
by [
UZ
H H
aupt
bibl
ioth
ek
Zen
tral
bibl
ioth
ek Z
uumlric
h] a
t 05
51 1
0 Se
ptem
ber
2013
FIGURE 10 Right scapulocoracoids of FMNH PR 2444 A stereopair of right scapulocoracoid in medial view B stereopair and interpretivedrawing of right scapulocoracoid in lateral view Scale bars equal 5 cm Abbreviationsmdashac acromion process C coracoid c gl coracoid contributionto the glenoid fossa c tr tubercle for coracoid attachment of triceps PC procoracoid pc f procoracoid foramen S scapula s gl scapularcontribution to the glenoid fossa
KAMMERER ET ALmdashCYNODONT FROM MADAGASCAR 455
Dow
nloa
ded
by [
UZ
H H
aupt
bibl
ioth
ek
Zen
tral
bibl
ioth
ek Z
uumlric
h] a
t 05
51 1
0 Se
ptem
ber
2013
teeth for a traversodontid As mentioned above Flynn et al(2000) noted that the homologies of these teeth were difficult todetermine Of the five anterior teeth preserved in the holotypethe anteriormost three are ldquoundoubtedly incisorsrdquo although I3 isldquounusually large and is caniniformrdquo (Flynn et al 2000423) Therobust recurved fourth tooth in the series was considered torepresent the canine with the subsequent tooth interpreted as anolder canine in the process of replacement Although FMNH PR2444 is edentulous the alveoli on the left side of the skull are wellpreserved and indicate an anterior tooth set identical to the ho-lotype The alveolar morphology indicates that the fifth toothwas canted anteriorly as in the holotype This suggests that themysterious fifth tooth is not the result of a transient period in thedental replacement history of M besairiei as suggested by Flynnet al (2000) but rather is the norm This leaves two possibilities1) M besairiei persistently had two canines This condition isunknown in cynodonts 2) The canine-like fourth tooth actuallyrepresents an incisor and the fifth tooth is the true canine Wefavor the latter explanation In FMNH PR 2444 the first fouralveoli are nearly identical in size and shape However the fifthalveolus while similar in anteroposterior length to its predeces-sors is strongly compressed labiolingually with an anteroposte-rior lengthlabiolingual width ratio of 044 (compared to 080 inthe fourth alveolus and 084ndash85 for I2ndash3) We interpret thismarked difference in alveolar ratio as indicating the separationbetween incisors and canine The canine-like I4 would then rep-resent the endpoint in a series of large increasingly caniniformincisors Supporting this interpretation recurved incisors are alsoobserved in juvenile Exaeretodon (eg MCP 3843 PV see Ab-dala et al 2002)
There are several legitimate differences between FMNH PR2444 and the holotype of Menadon besairiei but we take a con-servative taxonomic stance and argue that these differences rep-resent intraspecific variation and preservational artifacts FMNHPR 2444 represents a larger animal than UA 10601 The snoutlength (measured as the distance between the anterior edge ofthe orbit and a vertical plane through the anterior tip of thepremaxilla) of FMNH PR 2444 is 911 mm while that of UA10601 is 801 mm As such proportional differences between thetwo skulls may be attributable to ontogeny Variations in FMNHPR 2444 compared to the holotype that probably represent on-togentic differences include wider longer and deeper paraca-nine fossae a much deeper ventral zygomatic flange (althoughthe lower border of the flange is slightly abraded in the holotypeof Menadon besairiei there is no evidence that a major flangewas broken off) a proportionally shorter and more circular out-line to the posterior border of the maxilla below the zygomaticprocess and a more deeply emarginated posterior squamosal(ie a more prominent laterally projecting flange)
FMNH PR 2444 has a proportionally shorter more robustsnout and larger orbits than the holotype While these charactersare certainly variable in cynodonts it is unusual that the largerspecimen is shorter-snouted and larger-eyed contra the usualpattern The proportional differences between the two skulls areminor however (orbit diametersnout length is 042 for FMNHPR 2444 and 041 for the holotype) and are partially attributableto deformation (shear in FMNH PR 2444 and breakage of thepostorbital bar in the holotype) The following differences canalso probably be attributed to preservational artifact only fivepostcanine alveoli are preserved in FMNH PR 2444 while theholotype has eight postcanines The anteriormost three in theholotype however are tiny and peg-like As the area of themaxilla between the canine alveolus and the first preserved post-canine alveolus is poorly preserved in FMNH PR 2444 the ab-sence of these postcanines may be artifactual However it is alsopossible that the lower postcanine count of FMNH PR 2444compared to the holotype is an ontogenetic effect as postcaninecount is known to decrease with increasing skull size in Ex-
FIGURE 11 Left scapulocoracoids of FMNH PR 2444 A left scapu-locoracoid in medial view B left scapulocoracoid in lateral view Scalebars equal 5 cm
JOURNAL OF VERTEBRATE PALEONTOLOGY VOL 28 NO 2 2008456
Dow
nloa
ded
by [
UZ
H H
aupt
bibl
ioth
ek
Zen
tral
bibl
ioth
ek Z
uumlric
h] a
t 05
51 1
0 Se
ptem
ber
2013
FIGURE 12 Right pelvis of FMNH PR 2444 A stereopair of right pelvis in medial view B stereopair and interpretive drawing of right pelvis inlateral view Scale bars equal 5 cm Abbreviationsmdashbpp base of posterior process of ilium ib iliac blade IL ilium IS ischium ob f obturatorforamen P pubis
KAMMERER ET ALmdashCYNODONT FROM MADAGASCAR 457
Dow
nloa
ded
by [
UZ
H H
aupt
bibl
ioth
ek
Zen
tral
bibl
ioth
ek Z
uumlric
h] a
t 05
51 1
0 Se
ptem
ber
2013
aeretodon (Abdala et al 2002) The orbit of FMNH PR 2444 isless upwardly directed than that of the holotype (being at anangle of sim65deg to a sagittal plane through the midline of the skullas opposed to sim45deg in the holotype) This difference in orbitalangle is probably attributable to postmortem deformation as theskull of FMNH PR 2444 is noticeably sheared to the front leftThe appearance of a wider squarer incisor arcade and broader
anterior palate in FMNH PR 2444 can also be attributed to thisshear Finally the occipital region of FMNH PR 2444 is alto-gether taller and narrower than that of the holotype For ex-ample the foramen magnum in FMNH PR 2444 approximates arounded isosceles triangle in shape while the foramen magnumof the holotype is more ovoid These differences can also beattributed to deformation Altogether the differences between
FIGURE 13 Stereopairs of field number 8-31-98-387 an isolated right pelvis in A medial and B lateral view Scale bars equal 5 cm
JOURNAL OF VERTEBRATE PALEONTOLOGY VOL 28 NO 2 2008458
Dow
nloa
ded
by [
UZ
H H
aupt
bibl
ioth
ek
Zen
tral
bibl
ioth
ek Z
uumlric
h] a
t 05
51 1
0 Se
ptem
ber
2013
FMNH PR 2444 and the holotype fall within the range of intra-specific variation recorded for other gomphodont cynodontssuch as the traversodontids Massetognathus pascuali (Abdalaand Giannini 2000 Kammerer pers obs) and Exaeretodon(Abdala et al 2002) and the diademodontid Diademodon tetra-gonus (Grine et al 1978 Bradu and Grine 1979)
Phylogenetic Analysis
Utilizing the new information afforded by FMNH PR 2444and a reexamination of the holotype (UA 10601) Menadon be-sairiei was recoded in the Gondwanan gomphodont data matrixof Abdala et al (2006) Abdala et alrsquos (2006) data matrix en-compassed 18 taxa and 43 characters For the present analysis anadditional character ldquoincisor procumbencyrdquo (states 0-absent1-present) was added to Abdala et alrsquos (2006) data set (see
Appendix for coding of this character) Several traversodontidtaxa (eg lsquoScalenodonrsquo hirschsoni) have relatively large incisorsbut only in Menadon Scalenodontoides and Exaeretodon are theincisors procumbent Three of Abdala et alrsquos (2006) originalcodings for Menadon besairiei characters 0 (ldquonumber of upperincisorsrdquo) 5 (ldquoposition of paracanine fossa in relation to theupper caninerdquo) and 21 (ldquointernarial barrdquo) were changed to in-corporate data from the new specimen (see Appendix) Addi-tionally further examination of the holotype (UA 10601) ofMenadon besairiei allowed us to code characters 16ndash18 20 38and 40 (morphology of the lower postcanines all marked as ldquordquoin Abdala et alrsquos analysis) Parsimony searches were performedin PAUP v40b10 using a branch-and-bound search Addition-ally a bootstrap analysis (also using branch-and-bound) was per-formed using 1000 replicates
A total of 34 most parsimonious trees of length 103 werefound with the scores CI0573 RI0725 RC0415 andHI0427 The strict consensus tree (Fig 15A) did not recovera polytomy between Menadon Exaeretodon and Scalenodont-oides as in Abdala and Ribeiro (2003) or a (Menadon + Ex-aeretodon) sister-group relationship as in Abdala et al (2006)Rather Menadon was recovered as the sister taxon of the (Ex-aeretodon + Scalenodontoides) clade supporting Flynn et alrsquos(2000) original placement of this taxon There is strong bootstrapsupport (88) for Traversodontidae but within traversodontidsthe only strongly supported subclade is (Gomphodontosuchus +(Menadon + (Scalenodontoides + Exaeretodon))) with a boot-strap value of 80 Within this clade the (Menadon + (Scalen-odontoides + Exaeretodon)) grouping is supported by a boot-strap value of 83 and the (Scalenodontoides + Exaeretodon)grouping is supported by a bootstrap value of 94 (Fig 15B) Inaddition to the positioning of Menadon the strict consensus tree(Fig 15A) differs from the topology of Abdala et al (2006) inrecovering a sister-group relationship between Massetognathusand Santacruzodon rather than having Massetognathus andSantacruzodon as successive outgroups to the clade containingGomphodontosuchus Menadon Scalenodontoides and Ex-aeretodon The (Massetognathus + Santacruzodon) clade is diag-nosed by the absence of an anterolingual cusp in the upper post-canines denticulated incisor cutting margins and the presence ofan anterior cingulum in the upper postcanines
FIGURE 14 Reconstruction of the pelvis of Menadon besairiei basedon information from FMNH PR 2444 and field number 8-31-98-387Dotted area not preserved in either specimen reconstruction based onPascualgnathus (Bonaparte 1966) and Exaeretodon (Bonaparte 1963)
FIGURE 15 Cladogram of traversodontid relationships based on revised data set of Abdala et al (2006) showing A strict consensus tree and B50 majority rule tree Numbers at nodes of majority rule tree represent bootstrap values
KAMMERER ET ALmdashCYNODONT FROM MADAGASCAR 459
Dow
nloa
ded
by [
UZ
H H
aupt
bibl
ioth
ek
Zen
tral
bibl
ioth
ek Z
uumlric
h] a
t 05
51 1
0 Se
ptem
ber
2013
Relationships of Menadon besairiei
Menadon besairiei is the sister taxon to (Exaeretodon + Sca-lenodontoides) the latter clade diagnosed by the possession ofthree upper incisors a posteromedial position of the paracaninefossae relative to the canines the absence of an internarial barlarge canines a projected zygomatic process of the jugal and amaxilla that contributes to the margin of the subtemporal fenes-tra (the last three character states represent reversals) The clade(Menadon + (Exaeretodon + Scalenodontoides)) is diagnosed bythe possession of large procumbent incisors The presence offour incisors a primitive retention distinguishes Menadon be-sairiei from Exaeretodon Scalenodontoides and an unnamed tra-versodontid (ldquoTraversodontid Type 1rdquo) from the Santa MariaFormation of Brazil (Abdala et al 2001 Abdala and Ribeiro2002) Gomphodontosuchus is recovered as the sister taxon to(Menadon (Exaeretodon + Scalenodontoides)) as has beenfound in previous studies (Flynn et al 2000 Abdala and Ribeiro2003) The taxon name Gomphodontosuchinae Watson andRomer 1956 (ex Gomphodontosuchidae Watson and Romer1956 first given subfamilial rank in Lehman 1961) is availablefor this clade As circumscribed here Gomphodontosuchinaeconsists of the most inclusive clade containing Gomphodontosu-chus brasiliensis von Huene 1928264 but not Massetognathuspascuali Romer 19672
Menadon besairiei displays a mixture of primitive and derivedcharacters within traversodontids The internarial bar and fourincisors are primitive features whereas the enlargement and pro-cumbency of the incisors are derived features shared with Ex-aeretodon and Scalenodontoides The postcranium shows severalsimilarities to that of Exaeretodon (apparent exclusion of theprocoracoid from the glenoid some features of the pelvis tallcaudal neural spines) but in general displays a rather conserva-tive morphology observed throughout Gomphodontia The mor-phology of the scapulocoracoids is closer akin to diademodontidsor trirachodontids than Exaeretodon and the expanded thoracicrib of Menadon has only slight differences from those of trirach-odontids or early traversodontids like Luangwa The atlas-axiscomplex of Menadon is similar to that of the Early Triassic cyn-odont Cynognathus in possessing a concave axial spine with along posterior process Although most gomphodonts have beenreconstructed with a convex axial spine as in early mammals thisspine is broken in most of the best described gomphodont post-crania (eg Diademodon Brink 1955) and the lsquosaddle-shapedrsquoaxial morphology may be more widely distributed among cyn-odonts than previously realized The new specimens of Menadondescribed in this paper demonstrate that a generalized gompho-dont postcranial morphology was retained deep within the tra-versodontid tree and that the derived morphologies of either ofthe well-known taxa Massetognathus or Exaeretodon should notbe taken as representative of traversodontids in general
ACKNOWLEDGMENTS
This work would not have been possible without the continu-ing support and encouragement of our collaborative programwith the Deacutepartement de Paleacuteontologie et drsquoAnthropologie Bi-ologique Universiteacute drsquoAntananarivo Madagascar under the di-rection of Armand Rasoamiaramanana (Department Chairman)and Gisegravele Ravololonarivo Randria (Head of the PaleontologyLaboratories) We are extremely grateful to the National Geo-graphic Society for their long-term support of the fieldwork as-sociated with this project Additional financial or logistical sup-port for fieldwork was provided by The Field Museum the Uni-versity of California-Santa Barbara and the WWF (WorldwideFund for Nature Madagascar) We acknowledge the Ministegraverede LrsquoEneacutergie et des Mines for authorizing field efforts and for
continuing to facilitate our joint US-Madagascar explorationresearch and education program Steve Goodman and AsminaGandhi generously assisted us in innumerable ways includinghospitality nonpareil We especially thank R Andriantompoha-vana L Razafimanantsoa A Toto Volahy JA Rabarison AGandie and P Vavisaro for assistance in the field during theexpedition in which these new specimens were discovered BillSimpson and Gina Wesley-Hunt for their patient excavation ofthis important material Allison Smith Ray Vodden Lisa Berg-wall and Deborah Wagner for excellent preparation of a chal-lenging specimen Jennifer Botha and Elize Butler (NationalMuseum Bloemfontein) Michael Raath and Fernando Abdala(Bernard Price Institute) Thomas Kemp (Oxford UniversityMuseum of Natural History) Ray Symonds (University Museumof Zoology Cambridge) Bernard Battail (Museacuteum nationaldrsquoHistoire naturelle Paris) Michael Maisch (Universitaumlt Tuumlbin-gen) and Charles Schaff and Farish Jenkins (Museum of Com-parative Zoology Harvard) for access to specimens and AgustiacutenMartinelli Fernando Abdala and James Hopson for helpful re-views of an earlier version of the manuscript
LITERATURE CITED
Abdala F M C Barbarena and J Dornelles 2002 A new species of thetraversodontid cynodont Exaeretodon from the Santa Maria Forma-tion (MiddleLate Triassic) of Southern Brazil Journal of Verte-brate Paleontology 22313ndash325
Abdala F and N P Giannini 2000 Gomphodont cynodonts of theChantildeares Formation the analysis of an ontogenetic sequence Jour-nal of Vertebrate Paleontology 20501ndash506
Abdala F J Neveling and J Welman 2006 A new trirachodontidcynodont from the lower levels of the Burgersdorp Formation(Lower Triassic) of the Beaufort Group South Africa and the cla-distic relationships of Gondwanan gomphodonts Zoological Jour-nal of the Linnean Society 147383ndash413
Abdala F A M Ribeiro and C L Schultz 2001 A rich cynodont faunaof Santa Cruz do Sul Santa Maria Formation (Middle-Late Trias-sic) southern Brazil Neues Jahrbuch fuumlr Geologie und Palaumlontolo-gie Monatshefte 2001669ndash687
Abdala F and A M Ribeiro 2002 Nuevos cinodontes traversodoacuten-tidos (Synapsida-Eucynodontia) de la Formacioacuten Santa Maria(Triaacutesico Medio-Superior) Rio Grande do Sul Brasil RevistaEspantildeola de Paleontologiacutea 17237ndash243
Abdala F and A M Ribeiro 2003 A new traversodontid cynodontfrom the Santa Maria Formation (Ladinian-Carnian) of southernBrazil with a phylogenetic analysis of Gondwanan traversodontidsZoological Journal of the Linnaean Society 139529ndash545
Abdala F and A M S Teixeira 2004 A traversodontid cynodont ofAfrican affinity in the South American Triassic Palaeontologia Af-ricana 4011ndash22
Bonaparte J F 1962 Descripcioacuten del craacuteneo y mandiacutebula de Ex-aeretodon frenguellii Cabrera y su comparacioacuten con Diademodon-tidae Tritylodontidae y los cinodontes sudamericanos Publica-ciones del Museo Municipal de Ciencias Naturales y Tradicional deMar del Plata 1135ndash202
Bonaparte J F 1963 Descripcioacuten del esqueleto postcraneano de Ex-aeretodon (CynodontiandashTraversodontidae) Acta Geoloacutegica Lil-loana 45ndash52
Bonaparte J F 1966 Una nueva ldquofaunardquo Triaacutesica de Argentina (Ther-apsida Cynodontia Dicynodontia) Consideracions filogeneacuteticas ypaleobiogeograacuteficas Ameghiniana 8243ndash296
Bradu D and F E Grine 1979 Multivariate analysis of diademodon-tine crania from South Africa and Zambia South African Journal ofScience 79441ndash448
Brink A S 1955 A study of the skeleton of Diademodon Palaeonto-logia Africana 33-39
Brink A S 1963 Two new cynodonts from the Ntawere Formation inthe Luangwa Valley of Northern Rhodesia Palaeontologia Africana877ndash96
Broom R 1905 On the use of the term Anomodontia Records of theAlbany Museum 1266ndash269
Cabrera A 1943 El primer hallazgo de teraacutepsidos en la ArgentinaNotas del Museo de La Plata Paleontologiacutea 55317ndash331
JOURNAL OF VERTEBRATE PALEONTOLOGY VOL 28 NO 2 2008460
Dow
nloa
ded
by [
UZ
H H
aupt
bibl
ioth
ek
Zen
tral
bibl
ioth
ek Z
uumlric
h] a
t 05
51 1
0 Se
ptem
ber
2013
Chatterjee S 1982 A new cynodont reptile from the Triassic of IndiaJournal of Paleontology 56203ndash214
Crompton A W 1955 On some Triassic cynodonts from TanganyikaProceedings of the Zoological Society of London 125617ndash669
Crompton A W 1972 Postcanine occlusion in cynodonts and tritylo-donts Bulletin of the British Museum (Natural History) Geology2129ndash71
Crompton A W and F Ellenberger 1957 On a new cynodont from theMolteno Beds and the origin of the tritylodontids Annals of theSouth African Museum 441ndash13
Flynn J J J M Parrish B Rakotosaminimanana W F Simpson R LWhatley and A R Wyss 1999 A Triassic fauna from Madagascarincluding early dinosaurs Science 286763ndash765
Flynn J J J M Parrish B Rakotosamimanana L RanivoharimananaW F Simpson and A R Wyss 2000 New traversodontids (Syn-apsida Eucynodontia) from the Triassic of Madagascar Journal ofVertebrate Paleontology 20422ndash427
Gow C E and P J Hancox 1993 First complete skull of the LateTriassic Scalenodontoides (Reptilia Cynodontia) from southern Af-rica In Lucas S G and M Morales (eds) The nonmarine TriassicNew Mexico Museum of Natural History amp Science Bulletin 3161ndash168
Grine F E B D Hahn and C E Gow 1978 Aspects of relativegrowth and variability in Diademodon (Reptilia Therapsida) SouthAfrican Journal of Science 7450ndash58
Hopson J A 1984 Late Triassic traversodont cynodonts from NovaScotia and southern Africa Palaeontologia Africana 25181ndash201
Hopson J A 1985 Morphology and relationships of Gomphodontosu-chus brasiliensis von Huene (Synapsida Cynodontia Tritylodontoi-dea) from the Triassic of Brazil Neues Jahrbuch fuumlr Geologie undPalaumlontologie Monatshefte 1985285ndash299
Hopson J A and J W Kitching 2001 A probainognathian cynodontfrom South Africa and the phylogeny of nonmammalian cynodontsBulletin of the Museum of Comparative Zoology 1565ndash35
Hopson J A and H-D Sues 2006 A traversodont cynodont from theMiddle Triassic (Ladinian) of Baden-Wuumlrttemberg (Germany)Palaumlontologische Zeitschrift 80124ndash129
Huene F von 1928 Ein Cynodontier aus der Trias Brasiliens Zentral-blatt fuumlr Mineralogie Geologie und Palaumlontologie 1928251ndash270
Huene F von 1936 Die fossilen Reptilien des suumldamerikanischenGondwanalandes Ergebnisse der Sauriergrabungen in Suumldbrasilien192829 Lieferung 2 (Cynodontia)93ndash159 Franz F Heine Tuumlbin-gen
Jenkins F A 1970 The Chantildeares (Argentina) Triassic reptile faunaVII The postcranial skeleton of the traversodontid Massetognathuspascuali (Therapsida Cynodontia) Breviora 3521ndash28
Jenkins F A 1971 The postcranial skeleton of African cynodonts prob-lems in the early evolution of the mammalian postcranial skeletonBulletin of the Peabody Museum of Natural History 361ndash216
Kemp T S 1969 The atlas-axis complex of mammal-like reptiles Jour-nal of Zoology 159223ndash248
Kemp T S 1980 Aspects of the structure and functional anatomy of theMiddle Triassic cynodont Luangwa Journal of Zoology 191193ndash239
Kemp T S 1982 Mammal-Like Reptiles and the Origin of MammalsAcademic Press New York
Lehman J-P 1961 Cynodontia pp 140ndash191 in Piveteau J (ed) Traiteacutede Paleacuteontologie Masson et Cie Paris
Osborn 1903 On the primary division of the Reptilia into two sub-classes Synapsida and Diapsida Science 17275ndash276
Owen R 1861 Palaeontology or a systematic summary of extinct ani-mals and their geological relations 2nd Ed Edinburgh
Parrington F R 1934 On the cynodont genus Galesaurus with a noteon the functional significance of the changes in the evolution of thetheriodont skull Annals and Magazine of Natural History 10(13)38ndash67
Parrington F R 1946 On the cranial anatomy of cynodonts Proceed-ings of the Zoological Society of London 116181ndash197
Razafimbelo E 1987 Le basin de Morondava (Madagascar) Synthegravesegeacuteologique et structurale PhD dissertation University Louis Pas-teur Strasbourg France 189 pp
Reisz R R and H-D Sues 2000 Herbivory in Late Paleozoic andTriassic terrestrial vertebrates pp 9ndash41 in Sues H-D (ed) Evo-lution of Herbivory in Terrestrial Vertebrates Cambridge Univer-sity Press New York
Romer A S 1967 The Chantildeares (Argentina) Triassic reptile fauna IIITwo new gomphodonts Massetognathus pascuali and M terugiiBreviora 2641ndash25
Romer A S 1972 The Chantildeares (Argentina) Triassic reptile faunaXVII The Chantildeares gomphodonts Breviora 3961ndash9
Rougier G W J R Wible and J A Hopson 1992 Reconstruction ofthe cranial vessels in the Early Cretaceous mammal Vincelestes neu-quenianus implications for the evolution of the mammalian cranialvascular system Journal of Vertebrate Paleontology 12188ndash216
Rowe T 1988 Definition diagnosis and origin of Mammalia Journal ofVertebrate Paleontology 8241ndash264
Rowe T 1993 Early mammal phylogenetic systematics pp 129ndash145 inSzalay F S M J Novacek and M C McKenna (eds) MammalPhylogeny Vol 1 Mesozoic Differentiation MultituberculatesMonotremes Early Therians and Marsupials Springer-Verlag NewYork
Seeley H G 1894 Researches on the structure organization and clas-sification of the fossil Reptilia Part IX Section 3 On DiademodonPhilosophical Transactions of the Royal Society of London 1851029ndash1041
Seeley H G 1895 Researches on the structure organization and clas-sification of the fossil Reptilia Part IX Section 4 On the Gompho-dontia Philosophical Transactions of the Royal Society of London1861ndash57
Seeley H G 1895 Researches on the structure organization and clas-sification of the fossil Reptilia Part IX Section 5 On the skeletonin new Cynodontia from the Karroo rocks Philosophical Transac-tions of the Royal Society of London 18659ndash148
Sues H-D J A Hopson and N H Shubin 1992 Affinities of Sca-lenodontoides plemmyridon Hopson 1984 (Synapsida Cynodontia)from the Upper Triassic of Nova Scotia Journal of Vertebrate Pa-leontology 12168ndash171
Sues H-D and P E Olsen 1990 Triassic vertebrates of Gondwananaspect from the Richmond basin of Virginia Science 2491020ndash1023
Swofford D L 2003 PAUP v 40b10 Sinauer Associates Inc Pub-lishers Sunderland Massachusetts
Watson D M S and A S Romer 1956 A classification of therapsidreptiles Bulletin of the Museum of Comparative Zoology 11435ndash89
Submitted September 20 2007 accepted November 3 2007
KAMMERER ET ALmdashCYNODONT FROM MADAGASCAR 461
Dow
nloa
ded
by [
UZ
H H
aupt
bibl
ioth
ek
Zen
tral
bibl
ioth
ek Z
uumlric
h] a
t 05
51 1
0 Se
ptem
ber
2013
APPENDIX Data matrix used in the phylogenetic analysesThis is a revised version of the matrix of Abdala et al (2006) with the following changes character 43 [Incisor procumbency (0) absent (1) present]added Menadon recoded for characters 0 (Number of upper incisors was 1 ldquothreerdquo changed to 0 ldquofourrdquo) 5 (Position of paracanine fossae in relationto the upper canine was changed to 1 ldquomedialrdquo) 16 (Overall morphology of lower postcanines in occlusal view was changed to 3 ldquoquadran-gularrdquo) 17 (Position of the transverse crest in lower postcanines was changed to 1 ldquoanteriorrdquo) 18 (Number of cusps in the transverse crest of thelower postcanines was changed to 0 ldquotwordquo) 20 (Anterior cingulum in the lower postcanines was changed to 2 ldquoabsentrdquo) 21 (Internarial barwas 1 ldquoabsentrdquo changed to 0 ldquopresentrdquo) 38 (Posterior cingulum in the lower postcanines was changed to 1 ldquoabsentrdquo) and 40 (Deep occlusalbasins in the postcanines was changed to 1 ldquopresentrdquo) Langbergia recoded for character 34 (Incisor cutting margins was 0 ldquoserratedrdquo changedto 1 ldquosmoothly ridgedrdquo) based on the description of the incisors in Abdala et al (2006) Gomphodontosuchus recoded for character 31 (Maxilla inthe margin of the subtemporal fenestra was 1 ldquoincludedrdquo changed to 0 ldquoexcludedrdquo) based on von Huene (1936) and Kammerer (pers obs of theholotype) contra Hopsonrsquos (1985) reconstruction lsquoScalenodonrsquo hirschsoni recoded for character 39 (Anterior cingulum in the upper postcanines was1 ldquoabsentrdquo changed to 0 ldquopresentrdquo) based on Abdala (pers comm 2006) Trirachodon recoded for character 30 (Ectopterygoid was changedto 0 ldquopresentrdquo) based on Abdala (pers comm 2006) and Traversodon recoded for character 15 (Number of cusps in the sectorial border of the upperpostcanines was 0 ldquomore than twordquo changed to 1 ldquotwordquo) based on Abdala (pers comm 2006) and Kammerer (pers obs)
1111111111 2222222222 3333333333 44440123456789 0123456789 0123456789 0123456789 0123
Cynognathus 00000000 000 000000000 00000000 000Diademodon 0000001000 0000100 000010A000 0000000100 0100Trirachodon 000000100B 1A002201 001A100010 0001001200 0110Langbergia 0000001000 10000201 0000101010 0001101200 0110Cricodon 0100001000 11000201 0010100 0001200 0110Andescynodon 0000002001 01013101 101120010 0112311 1100Massetognathus 00111121A2 1110003100 2011200011 1011212300 1100Exaeretodon 1110122112 010103101 2111110201 1111112311 1101Luangwa 0000001002 11010A3100 101010010 01002300 1100Scal angustifrons 000000100B 1111023100 10102101 00002301 1100Scal hirschsoni 11110202 1100113100 1010 1112300 1100Traversodon 0000002012 1101113100 201010 011230 1100Gomphodontosuchus 0011112112 0103101 2010 0112311 1100Pascualgnathus 1000001000 000231 1100001 0011230 1100Scalenodontoides 1112212 01003101 21111021 1212311 1101Menadon 0111112112 01012310 20121020 0102311 1101Dadadon 00002112 111112 013 11231 1100Santacruzodon 00002112 1110003101 201300 212300 1100
A(01) B(02)
JOURNAL OF VERTEBRATE PALEONTOLOGY VOL 28 NO 2 2008462
Dow
nloa
ded
by [
UZ
H H
aupt
bibl
ioth
ek
Zen
tral
bibl
ioth
ek Z
uumlric
h] a
t 05
51 1
0 Se
ptem
ber
2013
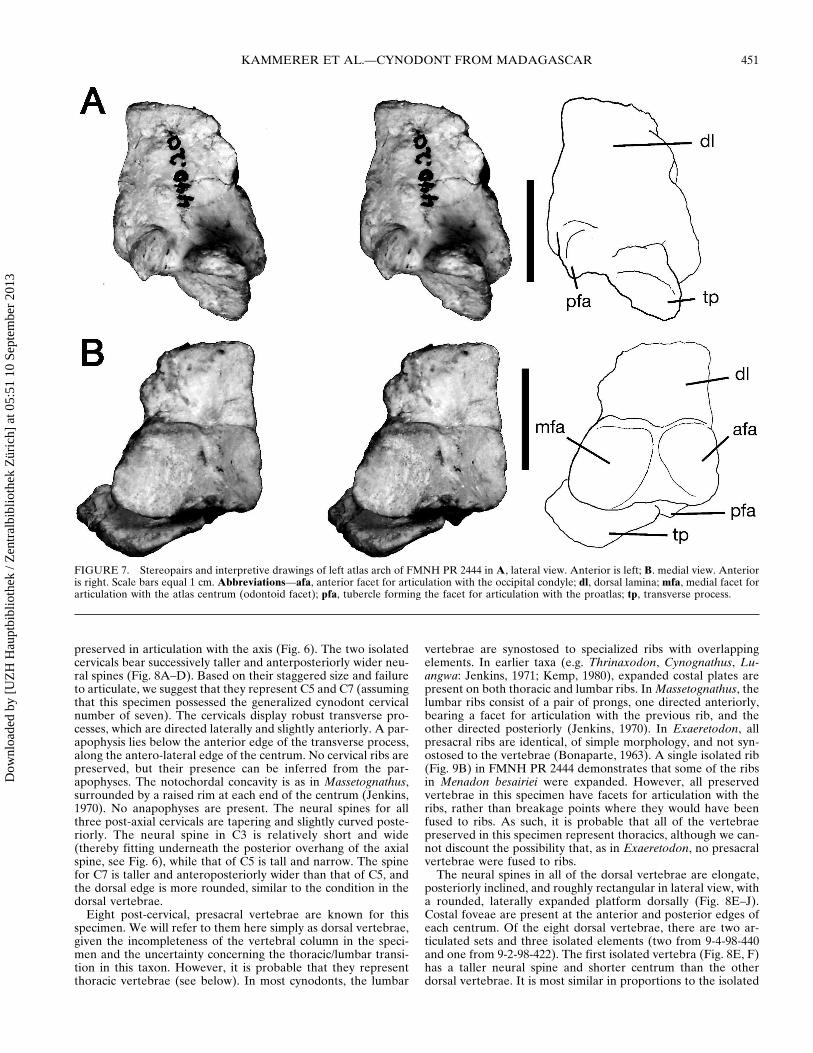
preserved in articulation with the axis (Fig 6) The two isolatedcervicals bear successively taller and anterposteriorly wider neu-ral spines (Fig 8AndashD) Based on their staggered size and failureto articulate we suggest that they represent C5 and C7 (assumingthat this specimen possessed the generalized cynodont cervicalnumber of seven) The cervicals display robust transverse pro-cesses which are directed laterally and slightly anteriorly A par-apophysis lies below the anterior edge of the transverse processalong the antero-lateral edge of the centrum No cervical ribs arepreserved but their presence can be inferred from the par-apophyses The notochordal concavity is as in Massetognathussurrounded by a raised rim at each end of the centrum (Jenkins1970) No anapophyses are present The neural spines for allthree post-axial cervicals are tapering and slightly curved poste-riorly The neural spine in C3 is relatively short and wide(thereby fitting underneath the posterior overhang of the axialspine see Fig 6) while that of C5 is tall and narrow The spinefor C7 is taller and anteroposteriorly wider than that of C5 andthe dorsal edge is more rounded similar to the condition in thedorsal vertebrae
Eight post-cervical presacral vertebrae are known for thisspecimen We will refer to them here simply as dorsal vertebraegiven the incompleteness of the vertebral column in the speci-men and the uncertainty concerning the thoraciclumbar transi-tion in this taxon However it is probable that they representthoracic vertebrae (see below) In most cynodonts the lumbar
vertebrae are synostosed to specialized ribs with overlappingelements In earlier taxa (eg Thrinaxodon Cynognathus Lu-angwa Jenkins 1971 Kemp 1980) expanded costal plates arepresent on both thoracic and lumbar ribs In Massetognathus thelumbar ribs consist of a pair of prongs one directed anteriorlybearing a facet for articulation with the previous rib and theother directed posteriorly (Jenkins 1970) In Exaeretodon allpresacral ribs are identical of simple morphology and not syn-ostosed to the vertebrae (Bonaparte 1963) A single isolated rib(Fig 9B) in FMNH PR 2444 demonstrates that some of the ribsin Menadon besairiei were expanded However all preservedvertebrae in this specimen have facets for articulation with theribs rather than breakage points where they would have beenfused to ribs As such it is probable that all of the vertebraepreserved in this specimen represent thoracics although we can-not discount the possibility that as in Exaeretodon no presacralvertebrae were fused to ribs
The neural spines in all of the dorsal vertebrae are elongateposteriorly inclined and roughly rectangular in lateral view witha rounded laterally expanded platform dorsally (Fig 8EndashJ)Costal foveae are present at the anterior and posterior edges ofeach centrum Of the eight dorsal vertebrae there are two ar-ticulated sets and three isolated elements (two from 9-4-98-440and one from 9-2-98-422) The first isolated vertebra (Fig 8E F)has a taller neural spine and shorter centrum than the otherdorsal vertebrae It is most similar in proportions to the isolated
FIGURE 7 Stereopairs and interpretive drawings of left atlas arch of FMNH PR 2444 in A lateral view Anterior is left B medial view Anterioris right Scale bars equal 1 cm Abbreviationsmdashafa anterior facet for articulation with the occipital condyle dl dorsal lamina mfa medial facet forarticulation with the atlas centrum (odontoid facet) pfa tubercle forming the facet for articulation with the proatlas tp transverse process
KAMMERER ET ALmdashCYNODONT FROM MADAGASCAR 451
Dow
nloa
ded
by [
UZ
H H
aupt
bibl
ioth
ek
Zen
tral
bibl
ioth
ek Z
uumlric
h] a
t 05
51 1
0 Se
ptem
ber
2013
FIGURE 8 Vertebrae of FMNH PR 2444 Cervical vertebra 5 in A left lateral and B anterior views Cervical vertebra 7 in C left lateral andD anterior views Isolated dorsal vertebra from anterior region of the series in E left lateral and F anterior views G stereopair and interpretivedrawing of articulated set of two dorsal vertebrae from near the midpoint of the series in left lateral view Anterior is up H articulated set of threedorsal vertebrae from anterior region of the series in left and right lateral views I isolated vertebra from near the midpoint of the series (from fieldnumber 9-2-98-422) J isolated vertebra from posterior region of the series Sacral vertebra in K dorsal view (the distal region of another sacral ribwhich is identical in shape to that of the intact sacral rib albeit slightly larger is also shown in this figure) and L left lateral view Caudal vertebraeM articulated set of two vertebrae from the anterior region of the series showing the lsquowingrsquo-shaped caudal ribs in dorsal view Anterior is left Nisolated caudal vertebra showing tall neural spine in left lateral view O isolated caudal vertebra showing tall neural spine in posterior view Particulated set of four vertebrae from near the end of the caudal series in dorsal view Anterior is left Scale bars equal 1 cm Abbreviationsmdasha cof anterior costal fovea n sp neural spine p co f posterior costal fovea poz postzygapophysis prz prezygapophysis
JOURNAL OF VERTEBRATE PALEONTOLOGY VOL 28 NO 2 2008452
Dow
nloa
ded
by [
UZ
H H
aupt
bibl
ioth
ek
Zen
tral
bibl
ioth
ek Z
uumlric
h] a
t 05
51 1
0 Se
ptem
ber
2013
cervicals and was probably located near the front of the dorsalseries The first articulated set consists of three vertebrae (Fig8H) The middle vertebra in this set is broken from the anteriorbase of the neural spine to the ventral midpoint and most of theright side of the anterior half of the centrum is missing This setof vertebrae probably also came from the anterior region of theseries as their neural spines are longer and narrower than allvertebrae other than the one described above The second ar-ticulated set (Fig 8G) consists of two well-preserved vertebraethat based on comparisons with Massetognathus and Ex-aeretodon (Jenkins 1970 Bonaparte 1963) probably came fromnear the midpoint of the dorsal series The second isolated ver-tebra (Fig 8I) from 9-2-98-422 is similar to the members of thesecond set The third isolated vertebra (Fig 8J) appears to befrom the posterior end of the series based on the relatively shortneural spine and reduced posterior costal fovea
One sacral vertebra (Fig 8K L) is known consisting of awell-preserved centrum neural spine and the right sacral rib(the left is broken off) The centrum is shorter (both anteropos-teriorly and dorsoventrally) than those of the dorsal vertebraeand the neural spine is shorter and more rounded in profileAlso the neural spine is directed vertically unlike those of thedorsal vertebrae which are canted posteriorly The zygapophy-ses are poorly developed The sacral rib is fused to a singlesynapophysis and projects anterolaterally The rib narrows to-wards the middle and expands distally terminating in a widerectangular facet where it would meet the ilium (Fig 8K)
Seven caudal vertebrae are known The caudal vertebrae aredistinctive in having very tall narrow neural spines Some EarlyTriassic cynodonts possess blunt neural spines of moderateheight on the proximal caudals (see for example Thrinaxodonin Jenkins 1971 and Diademodon in Brink 1955) Massetogna-thus represents an extreme condition wherein the caudal neuralspines are greatly reduced (Jenkins 1970) Very tall neuralspines are seen in Exaeretodon (Bonaparte 1963) and thosespines are unusual in that they do not diminish in height until thelast caudals The caudal vertebrae of FMNH PR 2444 consist ofa pair of articulated vertebrae most likely from near the sacrum(Fig 8M) an isolated caudal lacking postzygapophyses and ribsbut with the neural spine (Fig 8N O) and a quartet of smaller
articulated elements (Fig 8P) The anterior caudals have nar-rower neural spines and shorter zygapophyses than the subse-quent vertebrae in the series The caudal ribs attached to thispair have a crescentic distal region that curves anteriorly pro-ducing a lsquowingrsquo-shape Only a portion of the right rib of theisolated caudal is preserved but the lsquowingrsquo-shaped morphologyof the near-sacral caudal ribs does not appear to have been pre-sent In the posteriormost four vertebrae caudal ribs are pre-served on the left side of the first and the right side of the secondand third members On the first of these vertebrae the rib isshort curved posteriorly (but not lsquowingrsquo-shaped) and taperingRibs of the second and third vertebrae are less curved resem-bling posteriorly directed rods The zygapophyses of both theisolated caudal vertebra and the quartet of small caudals areanteroposteriorly elongate becoming increasingly so posteriorly
Thoracic RibsmdashNine thoracic ribs are known from 9-4-98-440(four left four right and an indeterminate splint of bone Fig9A indeterminate bone not illustrated) These elements re-semble the flattened ribs lacking costal plates of Exaeretodon(Bonaparte 1963) and the anterior thoracic ribs of Massetogna-thus and Luangwa (Jenkins 1970 Kemp 1980) The tuberculumand capitulum are separated by a shallow depression The tuber-culum has a shorter base than the capitulum but their proximalfaces are subequal in size An additional isolated left rib (Fig9B) from 9-2-98-422 is expanded indicating that unlike Ex-aeretodon Menadon rib morphology was heterogeneous alongthe length of the dorsal series This rib closely resembles theposterior thoracic ribs described for Luangwa (Kemp 1980fig16) Pascualgnathus (Bonaparte 1966fig 7) and the trirach-odontid Cricodon (Crompton 1955 Fig 4) This rib is con-stricted at the base of the head then expands distally with an-terior and posterior costal plates As in Luangwa the anteriorcostal plate is shallow and evenly sloping proximally and distallywhile the posterior costal plate is falcate with a concave edgedistally The distal expansion of this rib occurs at an angle of sim30degto the long axis of the rib head Despite their overall similaritythe expanded rib in FMNH PR 2444 does exhibit some differ-ences from the posterior thoracic rib of Luangwa The expandedrib of Menadon has a longer shaft between the rib head and thecostal plates and a shorter capitulum lacks a ridge on the pos-
FIGURE 9 A anterior thoracic ribs of FMNH PR 2444 in anterior view (indeterminate rib not figured) B expanded posterior thoracic rib ofFMNH PR 2444 in anterior view Scale bars equal 1 cm
KAMMERER ET ALmdashCYNODONT FROM MADAGASCAR 453
Dow
nloa
ded
by [
UZ
H H
aupt
bibl
ioth
ek
Zen
tral
bibl
ioth
ek Z
uumlric
h] a
t 05
51 1
0 Se
ptem
ber
2013
terior costal plate and has an anteroposteriorly shorter anteriorcostal plate Additionally the posterior margin of the costal plateis less curved than that of Luangwa These differences couldindicate that the expanded rib of Menadon is from a more pos-terior position in the series than the rib from Luangwa as re-duced curvature and increasing confluence of the tuberculumand capitulum are characteristic of lumbar ribs in traversodon-tids (Jenkins 1970 Kemp 1980) However the length of theshaft is at odds with this interpretation and in Cricodon andPascualgnathus the more posterior ribs are not more similar tothe element from FMNH PR 2444 These differences probablyrepresent legitimate morphological distinctions between the ribsof earlier gomphodonts and Menadon The head of the expandedrib is well defined and unbroken indicating that it was not syn-ostosed to a vertebra
Appendicular Skeleton
The appendicular skeleton is represented by both scapulocora-coids and a fragment of a clavicle from field number 9-4-98-440and a partial right pelvis from field number 9-2-98-422 An ad-ditional partial right pelvis is represented from field number8-31-98-387 The scapulocoracoids (Figs 10 11) are very similarto those described for other traversodontids as well as cynog-nathids and diademodontids (Jenkins 1970 1971) The scapularinfraspinous fossa has out-turned anterior and posterior edgesThe anterior edge of the scapula is rounded and curves laterallyforming the scapular spine At the lower end of the spine theacromion forms a prominent process (broken in the left scapula)Below this process the scapula is constricted and then expandsventrally to form the dorsal margin of the articular glenoid Theglenoid fossa is in general morphology very similar to that ofother Triassic cynodonts (Jenkins 1970 1971 Kemp 1980) It isposteroventrally oriented with roughly equal contributions fromthe scapular and coracoid glenoid facets The coracoid glenoidfacet is roughly triangular while the scapular glenoid facet issemiovoid as in other cynodonts In anterior or posterior viewthe fossa appears smoothly crescentic The procoracoid is aslightly curved trapezoidal plate of bone It does not appear tocontribute to the glenoid fossa The procoracoid foramen is lo-cated midway up the height of the procoracoid at a point directlybelow the base of the acromion process The coracoid is semi-crescentic curving posteriorly from a broad base The coracoidin Menadon is longer and more falcate than those described forExaeretodon (Bonaparte 1963) and Luangwa (Kemp 1980 al-though the distal tip of both coracoids is broken in the specimenof Luangwa [TSK 121] described by Kemp and may have beenlonger than reconstructed) but agrees closely with those of Mas-setognathus and cynognathidsdiademodontids (Jenkins 19701971) The distal coracoid tubercle for attachment to the tricepsis strongly curved upwards similar to the condition in Masseto-gnathus (Jenkins 1970) and Diademodon (Brink 1955)
A small fragment of bone resembling a curved cylinder almostcertainly represents part of a clavicle It has no distinguishingfeatures other than a groove associated with a ridge on one sideHowever the degree of curvature observed precludes this ele-ment from representing a long bone and the circular cross-section excludes it from being a rib
The pelvis of FMNH PR 2444 (Fig 12) resembles those ofother traversodontids in general morphology The ilium bears anarrow anteriorly directed iliac blade curving outward distallyThe anterior edge of the iliac blade slopes upward at a roughly45deg angle similar to Luangwa (Kemp 1980) but unlike Ex-aeretodon in which the anterior edge of the iliac blade curvesbackwards (Bonaparte 1963) or Pascualgnathus and Masseto-gnathus in which it is rounded (Bonaparte 1966 Jenkins 1970)The supraacetabular buttress at the base of the ilium is located inan anterodorsal position relative to the acetabulum as in other
traversodontids but unlike earlier cynodonts in which it is lo-cated dorsally (Jenkins 1970) The acetabular buttresses of thethree pelvic bones are largely confluent forming a fairly uniformlsquoliprsquo around the acetabulum as in Exaeretodon (Bonaparte1963) On the contrary in Luangwa the supraacetabular ischialand pubic buttresses are highly distinct spaced 6ndash8 mm apart andwithout confluence of their acetabular facets The pelvis ofFMNH PR 2444 does have a small depression between the su-praacetabular and ischial buttresses (also present in Ex-aeretodon Bonaparte 1963) but this is not as developed as inLuangwa The posterior extent of the iliac blade is broken inFMNH PR 2444 but this region is preserved in another isolatedright pelvis field number 8-31-98-387 (Fig 13) probably pertain-ing to Menadon besairiei The posterior process of the iliac bladein this specimen is relatively short and directed away from thedorsal edge of the ischium as in Exaeretodon (Bonaparte 1963)but unlike Massetognathus and Luangwa in which the posteriorprocess is long and extends roughly horizontally over the dorsaledge of the ischium (Jenkins 1970 Kemp 1980) and evencurves downward distally in Luangwa The posterior process ofthe iliac blade in Pascualgnathus is directed away from the is-chium (Bonaparte 1966) as in Exaeretodon and Menadon but isrelatively long as in Massetognathus and Luangwa
The proximal portion of the ischium is broad and dominatedby the ischial buttress The ischium constricts posterior to theproximal portion and expands distally into a spatulate platewhich is thickened at its posterior margin This thickening con-tinues along the dorsolateral surface of the ischium forming aridge that slopes ventrally until reaching the edge of the ischialbuttress The downward slope of this ridge produces a prominentgroove at the dorsal edge of the lateral surface of the ischiumthought to represent the origin of the ischio-trochantericusmuscle (Kemp 1980) The constriction of the ischium in Mena-don is like that of Luangwa albeit even more extreme in itsventral concavity but unlike that of Exaeretodon in which themedial and distal ends of the ischium are broadly united Theischium in general is a proportionally shorter element in Ex-aeretodon than in other traversodontids including Menadon(Bonaparte 1963) The anteroventral portion of the ischium thatconnects to the pubis is not preserved in either of the referredMenadon pelves
The pubis is the most incompletely preserved pelvic elementThe acetabular contribution of the pubis is smaller than that ofthe ischium or ilium Distal to the acetabulum the pubis is con-stricted indicating that the connection between the acetabularregion and the pubic plate was considerably narrower than inExaeretodon (Bonaparte 1963) but close akin to the pubis ofLuangwa (Kemp 1980) The pubic plate itself is not preservedThe ventral outline of the ischium and what is known of thepubis indicate that the obturator foramen was relatively larger inMenadon (Fig 14) than in Exaeretodon (Bonaparte 1963)
DISCUSSION
Comparison with the Holotype of Menadon besairiei
FMNH PR 2444 differs in several significant ways from theholotype of Menadon besairiei as previously described most no-tably in the possession of an internarial bar (initially consideredabsent in M besairiei) and four upper incisors (reported as threein M besairiei but with the caveat that the homologies of thefourth and fifth anterior tooth loci were problematic Flynn et al2000) However upon further study we interpret the lack of acontinuous internarial bar in the holotype as breakage ratherthan the natural condition of the skull (a prominent medial pro-tuberance is present on the ventral rim of the nares in UA10601) Regarding the incisor count it should be noted that theholotype of Menadon besairiei displays an unusual set of anterior
JOURNAL OF VERTEBRATE PALEONTOLOGY VOL 28 NO 2 2008454
Dow
nloa
ded
by [
UZ
H H
aupt
bibl
ioth
ek
Zen
tral
bibl
ioth
ek Z
uumlric
h] a
t 05
51 1
0 Se
ptem
ber
2013
FIGURE 10 Right scapulocoracoids of FMNH PR 2444 A stereopair of right scapulocoracoid in medial view B stereopair and interpretivedrawing of right scapulocoracoid in lateral view Scale bars equal 5 cm Abbreviationsmdashac acromion process C coracoid c gl coracoid contributionto the glenoid fossa c tr tubercle for coracoid attachment of triceps PC procoracoid pc f procoracoid foramen S scapula s gl scapularcontribution to the glenoid fossa
KAMMERER ET ALmdashCYNODONT FROM MADAGASCAR 455
Dow
nloa
ded
by [
UZ
H H
aupt
bibl
ioth
ek
Zen
tral
bibl
ioth
ek Z
uumlric
h] a
t 05
51 1
0 Se
ptem
ber
2013
teeth for a traversodontid As mentioned above Flynn et al(2000) noted that the homologies of these teeth were difficult todetermine Of the five anterior teeth preserved in the holotypethe anteriormost three are ldquoundoubtedly incisorsrdquo although I3 isldquounusually large and is caniniformrdquo (Flynn et al 2000423) Therobust recurved fourth tooth in the series was considered torepresent the canine with the subsequent tooth interpreted as anolder canine in the process of replacement Although FMNH PR2444 is edentulous the alveoli on the left side of the skull are wellpreserved and indicate an anterior tooth set identical to the ho-lotype The alveolar morphology indicates that the fifth toothwas canted anteriorly as in the holotype This suggests that themysterious fifth tooth is not the result of a transient period in thedental replacement history of M besairiei as suggested by Flynnet al (2000) but rather is the norm This leaves two possibilities1) M besairiei persistently had two canines This condition isunknown in cynodonts 2) The canine-like fourth tooth actuallyrepresents an incisor and the fifth tooth is the true canine Wefavor the latter explanation In FMNH PR 2444 the first fouralveoli are nearly identical in size and shape However the fifthalveolus while similar in anteroposterior length to its predeces-sors is strongly compressed labiolingually with an anteroposte-rior lengthlabiolingual width ratio of 044 (compared to 080 inthe fourth alveolus and 084ndash85 for I2ndash3) We interpret thismarked difference in alveolar ratio as indicating the separationbetween incisors and canine The canine-like I4 would then rep-resent the endpoint in a series of large increasingly caniniformincisors Supporting this interpretation recurved incisors are alsoobserved in juvenile Exaeretodon (eg MCP 3843 PV see Ab-dala et al 2002)
There are several legitimate differences between FMNH PR2444 and the holotype of Menadon besairiei but we take a con-servative taxonomic stance and argue that these differences rep-resent intraspecific variation and preservational artifacts FMNHPR 2444 represents a larger animal than UA 10601 The snoutlength (measured as the distance between the anterior edge ofthe orbit and a vertical plane through the anterior tip of thepremaxilla) of FMNH PR 2444 is 911 mm while that of UA10601 is 801 mm As such proportional differences between thetwo skulls may be attributable to ontogeny Variations in FMNHPR 2444 compared to the holotype that probably represent on-togentic differences include wider longer and deeper paraca-nine fossae a much deeper ventral zygomatic flange (althoughthe lower border of the flange is slightly abraded in the holotypeof Menadon besairiei there is no evidence that a major flangewas broken off) a proportionally shorter and more circular out-line to the posterior border of the maxilla below the zygomaticprocess and a more deeply emarginated posterior squamosal(ie a more prominent laterally projecting flange)
FMNH PR 2444 has a proportionally shorter more robustsnout and larger orbits than the holotype While these charactersare certainly variable in cynodonts it is unusual that the largerspecimen is shorter-snouted and larger-eyed contra the usualpattern The proportional differences between the two skulls areminor however (orbit diametersnout length is 042 for FMNHPR 2444 and 041 for the holotype) and are partially attributableto deformation (shear in FMNH PR 2444 and breakage of thepostorbital bar in the holotype) The following differences canalso probably be attributed to preservational artifact only fivepostcanine alveoli are preserved in FMNH PR 2444 while theholotype has eight postcanines The anteriormost three in theholotype however are tiny and peg-like As the area of themaxilla between the canine alveolus and the first preserved post-canine alveolus is poorly preserved in FMNH PR 2444 the ab-sence of these postcanines may be artifactual However it is alsopossible that the lower postcanine count of FMNH PR 2444compared to the holotype is an ontogenetic effect as postcaninecount is known to decrease with increasing skull size in Ex-
FIGURE 11 Left scapulocoracoids of FMNH PR 2444 A left scapu-locoracoid in medial view B left scapulocoracoid in lateral view Scalebars equal 5 cm
JOURNAL OF VERTEBRATE PALEONTOLOGY VOL 28 NO 2 2008456
Dow
nloa
ded
by [
UZ
H H
aupt
bibl
ioth
ek
Zen
tral
bibl
ioth
ek Z
uumlric
h] a
t 05
51 1
0 Se
ptem
ber
2013
FIGURE 12 Right pelvis of FMNH PR 2444 A stereopair of right pelvis in medial view B stereopair and interpretive drawing of right pelvis inlateral view Scale bars equal 5 cm Abbreviationsmdashbpp base of posterior process of ilium ib iliac blade IL ilium IS ischium ob f obturatorforamen P pubis
KAMMERER ET ALmdashCYNODONT FROM MADAGASCAR 457
Dow
nloa
ded
by [
UZ
H H
aupt
bibl
ioth
ek
Zen
tral
bibl
ioth
ek Z
uumlric
h] a
t 05
51 1
0 Se
ptem
ber
2013
aeretodon (Abdala et al 2002) The orbit of FMNH PR 2444 isless upwardly directed than that of the holotype (being at anangle of sim65deg to a sagittal plane through the midline of the skullas opposed to sim45deg in the holotype) This difference in orbitalangle is probably attributable to postmortem deformation as theskull of FMNH PR 2444 is noticeably sheared to the front leftThe appearance of a wider squarer incisor arcade and broader
anterior palate in FMNH PR 2444 can also be attributed to thisshear Finally the occipital region of FMNH PR 2444 is alto-gether taller and narrower than that of the holotype For ex-ample the foramen magnum in FMNH PR 2444 approximates arounded isosceles triangle in shape while the foramen magnumof the holotype is more ovoid These differences can also beattributed to deformation Altogether the differences between
FIGURE 13 Stereopairs of field number 8-31-98-387 an isolated right pelvis in A medial and B lateral view Scale bars equal 5 cm
JOURNAL OF VERTEBRATE PALEONTOLOGY VOL 28 NO 2 2008458
Dow
nloa
ded
by [
UZ
H H
aupt
bibl
ioth
ek
Zen
tral
bibl
ioth
ek Z
uumlric
h] a
t 05
51 1
0 Se
ptem
ber
2013
FMNH PR 2444 and the holotype fall within the range of intra-specific variation recorded for other gomphodont cynodontssuch as the traversodontids Massetognathus pascuali (Abdalaand Giannini 2000 Kammerer pers obs) and Exaeretodon(Abdala et al 2002) and the diademodontid Diademodon tetra-gonus (Grine et al 1978 Bradu and Grine 1979)
Phylogenetic Analysis
Utilizing the new information afforded by FMNH PR 2444and a reexamination of the holotype (UA 10601) Menadon be-sairiei was recoded in the Gondwanan gomphodont data matrixof Abdala et al (2006) Abdala et alrsquos (2006) data matrix en-compassed 18 taxa and 43 characters For the present analysis anadditional character ldquoincisor procumbencyrdquo (states 0-absent1-present) was added to Abdala et alrsquos (2006) data set (see
Appendix for coding of this character) Several traversodontidtaxa (eg lsquoScalenodonrsquo hirschsoni) have relatively large incisorsbut only in Menadon Scalenodontoides and Exaeretodon are theincisors procumbent Three of Abdala et alrsquos (2006) originalcodings for Menadon besairiei characters 0 (ldquonumber of upperincisorsrdquo) 5 (ldquoposition of paracanine fossa in relation to theupper caninerdquo) and 21 (ldquointernarial barrdquo) were changed to in-corporate data from the new specimen (see Appendix) Addi-tionally further examination of the holotype (UA 10601) ofMenadon besairiei allowed us to code characters 16ndash18 20 38and 40 (morphology of the lower postcanines all marked as ldquordquoin Abdala et alrsquos analysis) Parsimony searches were performedin PAUP v40b10 using a branch-and-bound search Addition-ally a bootstrap analysis (also using branch-and-bound) was per-formed using 1000 replicates
A total of 34 most parsimonious trees of length 103 werefound with the scores CI0573 RI0725 RC0415 andHI0427 The strict consensus tree (Fig 15A) did not recovera polytomy between Menadon Exaeretodon and Scalenodont-oides as in Abdala and Ribeiro (2003) or a (Menadon + Ex-aeretodon) sister-group relationship as in Abdala et al (2006)Rather Menadon was recovered as the sister taxon of the (Ex-aeretodon + Scalenodontoides) clade supporting Flynn et alrsquos(2000) original placement of this taxon There is strong bootstrapsupport (88) for Traversodontidae but within traversodontidsthe only strongly supported subclade is (Gomphodontosuchus +(Menadon + (Scalenodontoides + Exaeretodon))) with a boot-strap value of 80 Within this clade the (Menadon + (Scalen-odontoides + Exaeretodon)) grouping is supported by a boot-strap value of 83 and the (Scalenodontoides + Exaeretodon)grouping is supported by a bootstrap value of 94 (Fig 15B) Inaddition to the positioning of Menadon the strict consensus tree(Fig 15A) differs from the topology of Abdala et al (2006) inrecovering a sister-group relationship between Massetognathusand Santacruzodon rather than having Massetognathus andSantacruzodon as successive outgroups to the clade containingGomphodontosuchus Menadon Scalenodontoides and Ex-aeretodon The (Massetognathus + Santacruzodon) clade is diag-nosed by the absence of an anterolingual cusp in the upper post-canines denticulated incisor cutting margins and the presence ofan anterior cingulum in the upper postcanines
FIGURE 14 Reconstruction of the pelvis of Menadon besairiei basedon information from FMNH PR 2444 and field number 8-31-98-387Dotted area not preserved in either specimen reconstruction based onPascualgnathus (Bonaparte 1966) and Exaeretodon (Bonaparte 1963)
FIGURE 15 Cladogram of traversodontid relationships based on revised data set of Abdala et al (2006) showing A strict consensus tree and B50 majority rule tree Numbers at nodes of majority rule tree represent bootstrap values
KAMMERER ET ALmdashCYNODONT FROM MADAGASCAR 459
Dow
nloa
ded
by [
UZ
H H
aupt
bibl
ioth
ek
Zen
tral
bibl
ioth
ek Z
uumlric
h] a
t 05
51 1
0 Se
ptem
ber
2013
Relationships of Menadon besairiei
Menadon besairiei is the sister taxon to (Exaeretodon + Sca-lenodontoides) the latter clade diagnosed by the possession ofthree upper incisors a posteromedial position of the paracaninefossae relative to the canines the absence of an internarial barlarge canines a projected zygomatic process of the jugal and amaxilla that contributes to the margin of the subtemporal fenes-tra (the last three character states represent reversals) The clade(Menadon + (Exaeretodon + Scalenodontoides)) is diagnosed bythe possession of large procumbent incisors The presence offour incisors a primitive retention distinguishes Menadon be-sairiei from Exaeretodon Scalenodontoides and an unnamed tra-versodontid (ldquoTraversodontid Type 1rdquo) from the Santa MariaFormation of Brazil (Abdala et al 2001 Abdala and Ribeiro2002) Gomphodontosuchus is recovered as the sister taxon to(Menadon (Exaeretodon + Scalenodontoides)) as has beenfound in previous studies (Flynn et al 2000 Abdala and Ribeiro2003) The taxon name Gomphodontosuchinae Watson andRomer 1956 (ex Gomphodontosuchidae Watson and Romer1956 first given subfamilial rank in Lehman 1961) is availablefor this clade As circumscribed here Gomphodontosuchinaeconsists of the most inclusive clade containing Gomphodontosu-chus brasiliensis von Huene 1928264 but not Massetognathuspascuali Romer 19672
Menadon besairiei displays a mixture of primitive and derivedcharacters within traversodontids The internarial bar and fourincisors are primitive features whereas the enlargement and pro-cumbency of the incisors are derived features shared with Ex-aeretodon and Scalenodontoides The postcranium shows severalsimilarities to that of Exaeretodon (apparent exclusion of theprocoracoid from the glenoid some features of the pelvis tallcaudal neural spines) but in general displays a rather conserva-tive morphology observed throughout Gomphodontia The mor-phology of the scapulocoracoids is closer akin to diademodontidsor trirachodontids than Exaeretodon and the expanded thoracicrib of Menadon has only slight differences from those of trirach-odontids or early traversodontids like Luangwa The atlas-axiscomplex of Menadon is similar to that of the Early Triassic cyn-odont Cynognathus in possessing a concave axial spine with along posterior process Although most gomphodonts have beenreconstructed with a convex axial spine as in early mammals thisspine is broken in most of the best described gomphodont post-crania (eg Diademodon Brink 1955) and the lsquosaddle-shapedrsquoaxial morphology may be more widely distributed among cyn-odonts than previously realized The new specimens of Menadondescribed in this paper demonstrate that a generalized gompho-dont postcranial morphology was retained deep within the tra-versodontid tree and that the derived morphologies of either ofthe well-known taxa Massetognathus or Exaeretodon should notbe taken as representative of traversodontids in general
ACKNOWLEDGMENTS
This work would not have been possible without the continu-ing support and encouragement of our collaborative programwith the Deacutepartement de Paleacuteontologie et drsquoAnthropologie Bi-ologique Universiteacute drsquoAntananarivo Madagascar under the di-rection of Armand Rasoamiaramanana (Department Chairman)and Gisegravele Ravololonarivo Randria (Head of the PaleontologyLaboratories) We are extremely grateful to the National Geo-graphic Society for their long-term support of the fieldwork as-sociated with this project Additional financial or logistical sup-port for fieldwork was provided by The Field Museum the Uni-versity of California-Santa Barbara and the WWF (WorldwideFund for Nature Madagascar) We acknowledge the Ministegraverede LrsquoEneacutergie et des Mines for authorizing field efforts and for
continuing to facilitate our joint US-Madagascar explorationresearch and education program Steve Goodman and AsminaGandhi generously assisted us in innumerable ways includinghospitality nonpareil We especially thank R Andriantompoha-vana L Razafimanantsoa A Toto Volahy JA Rabarison AGandie and P Vavisaro for assistance in the field during theexpedition in which these new specimens were discovered BillSimpson and Gina Wesley-Hunt for their patient excavation ofthis important material Allison Smith Ray Vodden Lisa Berg-wall and Deborah Wagner for excellent preparation of a chal-lenging specimen Jennifer Botha and Elize Butler (NationalMuseum Bloemfontein) Michael Raath and Fernando Abdala(Bernard Price Institute) Thomas Kemp (Oxford UniversityMuseum of Natural History) Ray Symonds (University Museumof Zoology Cambridge) Bernard Battail (Museacuteum nationaldrsquoHistoire naturelle Paris) Michael Maisch (Universitaumlt Tuumlbin-gen) and Charles Schaff and Farish Jenkins (Museum of Com-parative Zoology Harvard) for access to specimens and AgustiacutenMartinelli Fernando Abdala and James Hopson for helpful re-views of an earlier version of the manuscript
LITERATURE CITED
Abdala F M C Barbarena and J Dornelles 2002 A new species of thetraversodontid cynodont Exaeretodon from the Santa Maria Forma-tion (MiddleLate Triassic) of Southern Brazil Journal of Verte-brate Paleontology 22313ndash325
Abdala F and N P Giannini 2000 Gomphodont cynodonts of theChantildeares Formation the analysis of an ontogenetic sequence Jour-nal of Vertebrate Paleontology 20501ndash506
Abdala F J Neveling and J Welman 2006 A new trirachodontidcynodont from the lower levels of the Burgersdorp Formation(Lower Triassic) of the Beaufort Group South Africa and the cla-distic relationships of Gondwanan gomphodonts Zoological Jour-nal of the Linnean Society 147383ndash413
Abdala F A M Ribeiro and C L Schultz 2001 A rich cynodont faunaof Santa Cruz do Sul Santa Maria Formation (Middle-Late Trias-sic) southern Brazil Neues Jahrbuch fuumlr Geologie und Palaumlontolo-gie Monatshefte 2001669ndash687
Abdala F and A M Ribeiro 2002 Nuevos cinodontes traversodoacuten-tidos (Synapsida-Eucynodontia) de la Formacioacuten Santa Maria(Triaacutesico Medio-Superior) Rio Grande do Sul Brasil RevistaEspantildeola de Paleontologiacutea 17237ndash243
Abdala F and A M Ribeiro 2003 A new traversodontid cynodontfrom the Santa Maria Formation (Ladinian-Carnian) of southernBrazil with a phylogenetic analysis of Gondwanan traversodontidsZoological Journal of the Linnaean Society 139529ndash545
Abdala F and A M S Teixeira 2004 A traversodontid cynodont ofAfrican affinity in the South American Triassic Palaeontologia Af-ricana 4011ndash22
Bonaparte J F 1962 Descripcioacuten del craacuteneo y mandiacutebula de Ex-aeretodon frenguellii Cabrera y su comparacioacuten con Diademodon-tidae Tritylodontidae y los cinodontes sudamericanos Publica-ciones del Museo Municipal de Ciencias Naturales y Tradicional deMar del Plata 1135ndash202
Bonaparte J F 1963 Descripcioacuten del esqueleto postcraneano de Ex-aeretodon (CynodontiandashTraversodontidae) Acta Geoloacutegica Lil-loana 45ndash52
Bonaparte J F 1966 Una nueva ldquofaunardquo Triaacutesica de Argentina (Ther-apsida Cynodontia Dicynodontia) Consideracions filogeneacuteticas ypaleobiogeograacuteficas Ameghiniana 8243ndash296
Bradu D and F E Grine 1979 Multivariate analysis of diademodon-tine crania from South Africa and Zambia South African Journal ofScience 79441ndash448
Brink A S 1955 A study of the skeleton of Diademodon Palaeonto-logia Africana 33-39
Brink A S 1963 Two new cynodonts from the Ntawere Formation inthe Luangwa Valley of Northern Rhodesia Palaeontologia Africana877ndash96
Broom R 1905 On the use of the term Anomodontia Records of theAlbany Museum 1266ndash269
Cabrera A 1943 El primer hallazgo de teraacutepsidos en la ArgentinaNotas del Museo de La Plata Paleontologiacutea 55317ndash331
JOURNAL OF VERTEBRATE PALEONTOLOGY VOL 28 NO 2 2008460
Dow
nloa
ded
by [
UZ
H H
aupt
bibl
ioth
ek
Zen
tral
bibl
ioth
ek Z
uumlric
h] a
t 05
51 1
0 Se
ptem
ber
2013
Chatterjee S 1982 A new cynodont reptile from the Triassic of IndiaJournal of Paleontology 56203ndash214
Crompton A W 1955 On some Triassic cynodonts from TanganyikaProceedings of the Zoological Society of London 125617ndash669
Crompton A W 1972 Postcanine occlusion in cynodonts and tritylo-donts Bulletin of the British Museum (Natural History) Geology2129ndash71
Crompton A W and F Ellenberger 1957 On a new cynodont from theMolteno Beds and the origin of the tritylodontids Annals of theSouth African Museum 441ndash13
Flynn J J J M Parrish B Rakotosaminimanana W F Simpson R LWhatley and A R Wyss 1999 A Triassic fauna from Madagascarincluding early dinosaurs Science 286763ndash765
Flynn J J J M Parrish B Rakotosamimanana L RanivoharimananaW F Simpson and A R Wyss 2000 New traversodontids (Syn-apsida Eucynodontia) from the Triassic of Madagascar Journal ofVertebrate Paleontology 20422ndash427
Gow C E and P J Hancox 1993 First complete skull of the LateTriassic Scalenodontoides (Reptilia Cynodontia) from southern Af-rica In Lucas S G and M Morales (eds) The nonmarine TriassicNew Mexico Museum of Natural History amp Science Bulletin 3161ndash168
Grine F E B D Hahn and C E Gow 1978 Aspects of relativegrowth and variability in Diademodon (Reptilia Therapsida) SouthAfrican Journal of Science 7450ndash58
Hopson J A 1984 Late Triassic traversodont cynodonts from NovaScotia and southern Africa Palaeontologia Africana 25181ndash201
Hopson J A 1985 Morphology and relationships of Gomphodontosu-chus brasiliensis von Huene (Synapsida Cynodontia Tritylodontoi-dea) from the Triassic of Brazil Neues Jahrbuch fuumlr Geologie undPalaumlontologie Monatshefte 1985285ndash299
Hopson J A and J W Kitching 2001 A probainognathian cynodontfrom South Africa and the phylogeny of nonmammalian cynodontsBulletin of the Museum of Comparative Zoology 1565ndash35
Hopson J A and H-D Sues 2006 A traversodont cynodont from theMiddle Triassic (Ladinian) of Baden-Wuumlrttemberg (Germany)Palaumlontologische Zeitschrift 80124ndash129
Huene F von 1928 Ein Cynodontier aus der Trias Brasiliens Zentral-blatt fuumlr Mineralogie Geologie und Palaumlontologie 1928251ndash270
Huene F von 1936 Die fossilen Reptilien des suumldamerikanischenGondwanalandes Ergebnisse der Sauriergrabungen in Suumldbrasilien192829 Lieferung 2 (Cynodontia)93ndash159 Franz F Heine Tuumlbin-gen
Jenkins F A 1970 The Chantildeares (Argentina) Triassic reptile faunaVII The postcranial skeleton of the traversodontid Massetognathuspascuali (Therapsida Cynodontia) Breviora 3521ndash28
Jenkins F A 1971 The postcranial skeleton of African cynodonts prob-lems in the early evolution of the mammalian postcranial skeletonBulletin of the Peabody Museum of Natural History 361ndash216
Kemp T S 1969 The atlas-axis complex of mammal-like reptiles Jour-nal of Zoology 159223ndash248
Kemp T S 1980 Aspects of the structure and functional anatomy of theMiddle Triassic cynodont Luangwa Journal of Zoology 191193ndash239
Kemp T S 1982 Mammal-Like Reptiles and the Origin of MammalsAcademic Press New York
Lehman J-P 1961 Cynodontia pp 140ndash191 in Piveteau J (ed) Traiteacutede Paleacuteontologie Masson et Cie Paris
Osborn 1903 On the primary division of the Reptilia into two sub-classes Synapsida and Diapsida Science 17275ndash276
Owen R 1861 Palaeontology or a systematic summary of extinct ani-mals and their geological relations 2nd Ed Edinburgh
Parrington F R 1934 On the cynodont genus Galesaurus with a noteon the functional significance of the changes in the evolution of thetheriodont skull Annals and Magazine of Natural History 10(13)38ndash67
Parrington F R 1946 On the cranial anatomy of cynodonts Proceed-ings of the Zoological Society of London 116181ndash197
Razafimbelo E 1987 Le basin de Morondava (Madagascar) Synthegravesegeacuteologique et structurale PhD dissertation University Louis Pas-teur Strasbourg France 189 pp
Reisz R R and H-D Sues 2000 Herbivory in Late Paleozoic andTriassic terrestrial vertebrates pp 9ndash41 in Sues H-D (ed) Evo-lution of Herbivory in Terrestrial Vertebrates Cambridge Univer-sity Press New York
Romer A S 1967 The Chantildeares (Argentina) Triassic reptile fauna IIITwo new gomphodonts Massetognathus pascuali and M terugiiBreviora 2641ndash25
Romer A S 1972 The Chantildeares (Argentina) Triassic reptile faunaXVII The Chantildeares gomphodonts Breviora 3961ndash9
Rougier G W J R Wible and J A Hopson 1992 Reconstruction ofthe cranial vessels in the Early Cretaceous mammal Vincelestes neu-quenianus implications for the evolution of the mammalian cranialvascular system Journal of Vertebrate Paleontology 12188ndash216
Rowe T 1988 Definition diagnosis and origin of Mammalia Journal ofVertebrate Paleontology 8241ndash264
Rowe T 1993 Early mammal phylogenetic systematics pp 129ndash145 inSzalay F S M J Novacek and M C McKenna (eds) MammalPhylogeny Vol 1 Mesozoic Differentiation MultituberculatesMonotremes Early Therians and Marsupials Springer-Verlag NewYork
Seeley H G 1894 Researches on the structure organization and clas-sification of the fossil Reptilia Part IX Section 3 On DiademodonPhilosophical Transactions of the Royal Society of London 1851029ndash1041
Seeley H G 1895 Researches on the structure organization and clas-sification of the fossil Reptilia Part IX Section 4 On the Gompho-dontia Philosophical Transactions of the Royal Society of London1861ndash57
Seeley H G 1895 Researches on the structure organization and clas-sification of the fossil Reptilia Part IX Section 5 On the skeletonin new Cynodontia from the Karroo rocks Philosophical Transac-tions of the Royal Society of London 18659ndash148
Sues H-D J A Hopson and N H Shubin 1992 Affinities of Sca-lenodontoides plemmyridon Hopson 1984 (Synapsida Cynodontia)from the Upper Triassic of Nova Scotia Journal of Vertebrate Pa-leontology 12168ndash171
Sues H-D and P E Olsen 1990 Triassic vertebrates of Gondwananaspect from the Richmond basin of Virginia Science 2491020ndash1023
Swofford D L 2003 PAUP v 40b10 Sinauer Associates Inc Pub-lishers Sunderland Massachusetts
Watson D M S and A S Romer 1956 A classification of therapsidreptiles Bulletin of the Museum of Comparative Zoology 11435ndash89
Submitted September 20 2007 accepted November 3 2007
KAMMERER ET ALmdashCYNODONT FROM MADAGASCAR 461
Dow
nloa
ded
by [
UZ
H H
aupt
bibl
ioth
ek
Zen
tral
bibl
ioth
ek Z
uumlric
h] a
t 05
51 1
0 Se
ptem
ber
2013
APPENDIX Data matrix used in the phylogenetic analysesThis is a revised version of the matrix of Abdala et al (2006) with the following changes character 43 [Incisor procumbency (0) absent (1) present]added Menadon recoded for characters 0 (Number of upper incisors was 1 ldquothreerdquo changed to 0 ldquofourrdquo) 5 (Position of paracanine fossae in relationto the upper canine was changed to 1 ldquomedialrdquo) 16 (Overall morphology of lower postcanines in occlusal view was changed to 3 ldquoquadran-gularrdquo) 17 (Position of the transverse crest in lower postcanines was changed to 1 ldquoanteriorrdquo) 18 (Number of cusps in the transverse crest of thelower postcanines was changed to 0 ldquotwordquo) 20 (Anterior cingulum in the lower postcanines was changed to 2 ldquoabsentrdquo) 21 (Internarial barwas 1 ldquoabsentrdquo changed to 0 ldquopresentrdquo) 38 (Posterior cingulum in the lower postcanines was changed to 1 ldquoabsentrdquo) and 40 (Deep occlusalbasins in the postcanines was changed to 1 ldquopresentrdquo) Langbergia recoded for character 34 (Incisor cutting margins was 0 ldquoserratedrdquo changedto 1 ldquosmoothly ridgedrdquo) based on the description of the incisors in Abdala et al (2006) Gomphodontosuchus recoded for character 31 (Maxilla inthe margin of the subtemporal fenestra was 1 ldquoincludedrdquo changed to 0 ldquoexcludedrdquo) based on von Huene (1936) and Kammerer (pers obs of theholotype) contra Hopsonrsquos (1985) reconstruction lsquoScalenodonrsquo hirschsoni recoded for character 39 (Anterior cingulum in the upper postcanines was1 ldquoabsentrdquo changed to 0 ldquopresentrdquo) based on Abdala (pers comm 2006) Trirachodon recoded for character 30 (Ectopterygoid was changedto 0 ldquopresentrdquo) based on Abdala (pers comm 2006) and Traversodon recoded for character 15 (Number of cusps in the sectorial border of the upperpostcanines was 0 ldquomore than twordquo changed to 1 ldquotwordquo) based on Abdala (pers comm 2006) and Kammerer (pers obs)
1111111111 2222222222 3333333333 44440123456789 0123456789 0123456789 0123456789 0123
Cynognathus 00000000 000 000000000 00000000 000Diademodon 0000001000 0000100 000010A000 0000000100 0100Trirachodon 000000100B 1A002201 001A100010 0001001200 0110Langbergia 0000001000 10000201 0000101010 0001101200 0110Cricodon 0100001000 11000201 0010100 0001200 0110Andescynodon 0000002001 01013101 101120010 0112311 1100Massetognathus 00111121A2 1110003100 2011200011 1011212300 1100Exaeretodon 1110122112 010103101 2111110201 1111112311 1101Luangwa 0000001002 11010A3100 101010010 01002300 1100Scal angustifrons 000000100B 1111023100 10102101 00002301 1100Scal hirschsoni 11110202 1100113100 1010 1112300 1100Traversodon 0000002012 1101113100 201010 011230 1100Gomphodontosuchus 0011112112 0103101 2010 0112311 1100Pascualgnathus 1000001000 000231 1100001 0011230 1100Scalenodontoides 1112212 01003101 21111021 1212311 1101Menadon 0111112112 01012310 20121020 0102311 1101Dadadon 00002112 111112 013 11231 1100Santacruzodon 00002112 1110003101 201300 212300 1100
A(01) B(02)
JOURNAL OF VERTEBRATE PALEONTOLOGY VOL 28 NO 2 2008462
Dow
nloa
ded
by [
UZ
H H
aupt
bibl
ioth
ek
Zen
tral
bibl
ioth
ek Z
uumlric
h] a
t 05
51 1
0 Se
ptem
ber
2013
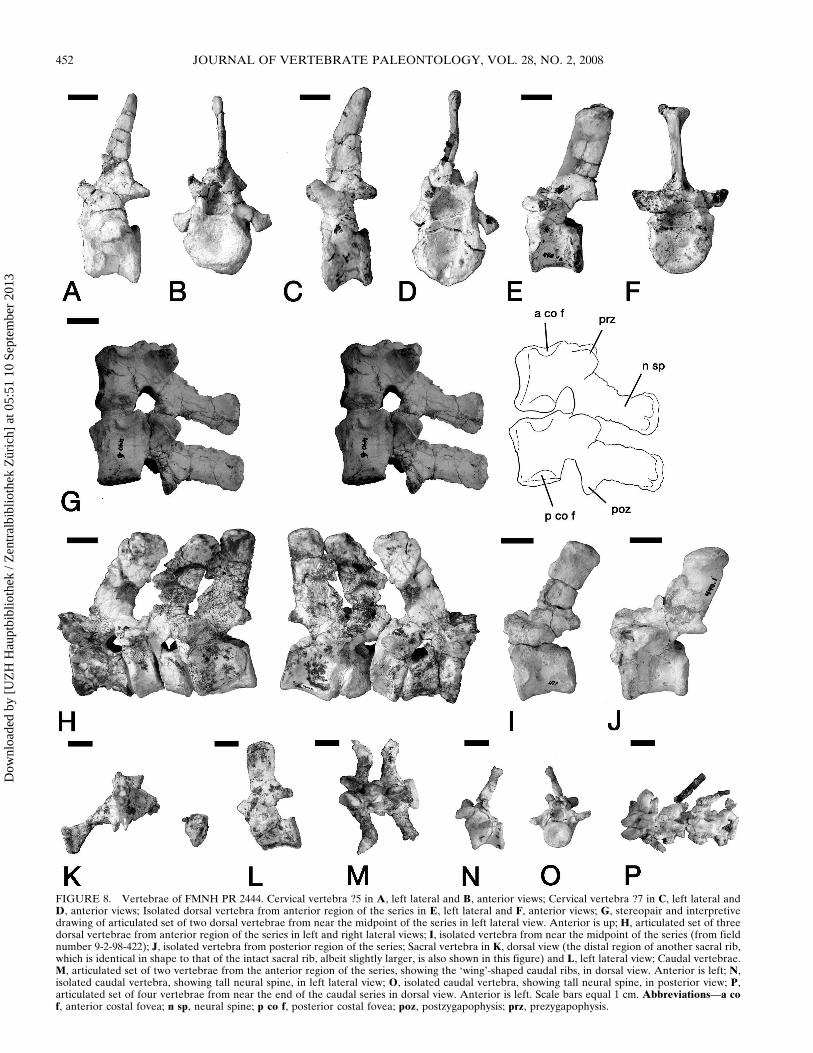
FIGURE 8 Vertebrae of FMNH PR 2444 Cervical vertebra 5 in A left lateral and B anterior views Cervical vertebra 7 in C left lateral andD anterior views Isolated dorsal vertebra from anterior region of the series in E left lateral and F anterior views G stereopair and interpretivedrawing of articulated set of two dorsal vertebrae from near the midpoint of the series in left lateral view Anterior is up H articulated set of threedorsal vertebrae from anterior region of the series in left and right lateral views I isolated vertebra from near the midpoint of the series (from fieldnumber 9-2-98-422) J isolated vertebra from posterior region of the series Sacral vertebra in K dorsal view (the distal region of another sacral ribwhich is identical in shape to that of the intact sacral rib albeit slightly larger is also shown in this figure) and L left lateral view Caudal vertebraeM articulated set of two vertebrae from the anterior region of the series showing the lsquowingrsquo-shaped caudal ribs in dorsal view Anterior is left Nisolated caudal vertebra showing tall neural spine in left lateral view O isolated caudal vertebra showing tall neural spine in posterior view Particulated set of four vertebrae from near the end of the caudal series in dorsal view Anterior is left Scale bars equal 1 cm Abbreviationsmdasha cof anterior costal fovea n sp neural spine p co f posterior costal fovea poz postzygapophysis prz prezygapophysis
JOURNAL OF VERTEBRATE PALEONTOLOGY VOL 28 NO 2 2008452
Dow
nloa
ded
by [
UZ
H H
aupt
bibl
ioth
ek
Zen
tral
bibl
ioth
ek Z
uumlric
h] a
t 05
51 1
0 Se
ptem
ber
2013
cervicals and was probably located near the front of the dorsalseries The first articulated set consists of three vertebrae (Fig8H) The middle vertebra in this set is broken from the anteriorbase of the neural spine to the ventral midpoint and most of theright side of the anterior half of the centrum is missing This setof vertebrae probably also came from the anterior region of theseries as their neural spines are longer and narrower than allvertebrae other than the one described above The second ar-ticulated set (Fig 8G) consists of two well-preserved vertebraethat based on comparisons with Massetognathus and Ex-aeretodon (Jenkins 1970 Bonaparte 1963) probably came fromnear the midpoint of the dorsal series The second isolated ver-tebra (Fig 8I) from 9-2-98-422 is similar to the members of thesecond set The third isolated vertebra (Fig 8J) appears to befrom the posterior end of the series based on the relatively shortneural spine and reduced posterior costal fovea
One sacral vertebra (Fig 8K L) is known consisting of awell-preserved centrum neural spine and the right sacral rib(the left is broken off) The centrum is shorter (both anteropos-teriorly and dorsoventrally) than those of the dorsal vertebraeand the neural spine is shorter and more rounded in profileAlso the neural spine is directed vertically unlike those of thedorsal vertebrae which are canted posteriorly The zygapophy-ses are poorly developed The sacral rib is fused to a singlesynapophysis and projects anterolaterally The rib narrows to-wards the middle and expands distally terminating in a widerectangular facet where it would meet the ilium (Fig 8K)
Seven caudal vertebrae are known The caudal vertebrae aredistinctive in having very tall narrow neural spines Some EarlyTriassic cynodonts possess blunt neural spines of moderateheight on the proximal caudals (see for example Thrinaxodonin Jenkins 1971 and Diademodon in Brink 1955) Massetogna-thus represents an extreme condition wherein the caudal neuralspines are greatly reduced (Jenkins 1970) Very tall neuralspines are seen in Exaeretodon (Bonaparte 1963) and thosespines are unusual in that they do not diminish in height until thelast caudals The caudal vertebrae of FMNH PR 2444 consist ofa pair of articulated vertebrae most likely from near the sacrum(Fig 8M) an isolated caudal lacking postzygapophyses and ribsbut with the neural spine (Fig 8N O) and a quartet of smaller
articulated elements (Fig 8P) The anterior caudals have nar-rower neural spines and shorter zygapophyses than the subse-quent vertebrae in the series The caudal ribs attached to thispair have a crescentic distal region that curves anteriorly pro-ducing a lsquowingrsquo-shape Only a portion of the right rib of theisolated caudal is preserved but the lsquowingrsquo-shaped morphologyof the near-sacral caudal ribs does not appear to have been pre-sent In the posteriormost four vertebrae caudal ribs are pre-served on the left side of the first and the right side of the secondand third members On the first of these vertebrae the rib isshort curved posteriorly (but not lsquowingrsquo-shaped) and taperingRibs of the second and third vertebrae are less curved resem-bling posteriorly directed rods The zygapophyses of both theisolated caudal vertebra and the quartet of small caudals areanteroposteriorly elongate becoming increasingly so posteriorly
Thoracic RibsmdashNine thoracic ribs are known from 9-4-98-440(four left four right and an indeterminate splint of bone Fig9A indeterminate bone not illustrated) These elements re-semble the flattened ribs lacking costal plates of Exaeretodon(Bonaparte 1963) and the anterior thoracic ribs of Massetogna-thus and Luangwa (Jenkins 1970 Kemp 1980) The tuberculumand capitulum are separated by a shallow depression The tuber-culum has a shorter base than the capitulum but their proximalfaces are subequal in size An additional isolated left rib (Fig9B) from 9-2-98-422 is expanded indicating that unlike Ex-aeretodon Menadon rib morphology was heterogeneous alongthe length of the dorsal series This rib closely resembles theposterior thoracic ribs described for Luangwa (Kemp 1980fig16) Pascualgnathus (Bonaparte 1966fig 7) and the trirach-odontid Cricodon (Crompton 1955 Fig 4) This rib is con-stricted at the base of the head then expands distally with an-terior and posterior costal plates As in Luangwa the anteriorcostal plate is shallow and evenly sloping proximally and distallywhile the posterior costal plate is falcate with a concave edgedistally The distal expansion of this rib occurs at an angle of sim30degto the long axis of the rib head Despite their overall similaritythe expanded rib in FMNH PR 2444 does exhibit some differ-ences from the posterior thoracic rib of Luangwa The expandedrib of Menadon has a longer shaft between the rib head and thecostal plates and a shorter capitulum lacks a ridge on the pos-
FIGURE 9 A anterior thoracic ribs of FMNH PR 2444 in anterior view (indeterminate rib not figured) B expanded posterior thoracic rib ofFMNH PR 2444 in anterior view Scale bars equal 1 cm
KAMMERER ET ALmdashCYNODONT FROM MADAGASCAR 453
Dow
nloa
ded
by [
UZ
H H
aupt
bibl
ioth
ek
Zen
tral
bibl
ioth
ek Z
uumlric
h] a
t 05
51 1
0 Se
ptem
ber
2013
terior costal plate and has an anteroposteriorly shorter anteriorcostal plate Additionally the posterior margin of the costal plateis less curved than that of Luangwa These differences couldindicate that the expanded rib of Menadon is from a more pos-terior position in the series than the rib from Luangwa as re-duced curvature and increasing confluence of the tuberculumand capitulum are characteristic of lumbar ribs in traversodon-tids (Jenkins 1970 Kemp 1980) However the length of theshaft is at odds with this interpretation and in Cricodon andPascualgnathus the more posterior ribs are not more similar tothe element from FMNH PR 2444 These differences probablyrepresent legitimate morphological distinctions between the ribsof earlier gomphodonts and Menadon The head of the expandedrib is well defined and unbroken indicating that it was not syn-ostosed to a vertebra
Appendicular Skeleton
The appendicular skeleton is represented by both scapulocora-coids and a fragment of a clavicle from field number 9-4-98-440and a partial right pelvis from field number 9-2-98-422 An ad-ditional partial right pelvis is represented from field number8-31-98-387 The scapulocoracoids (Figs 10 11) are very similarto those described for other traversodontids as well as cynog-nathids and diademodontids (Jenkins 1970 1971) The scapularinfraspinous fossa has out-turned anterior and posterior edgesThe anterior edge of the scapula is rounded and curves laterallyforming the scapular spine At the lower end of the spine theacromion forms a prominent process (broken in the left scapula)Below this process the scapula is constricted and then expandsventrally to form the dorsal margin of the articular glenoid Theglenoid fossa is in general morphology very similar to that ofother Triassic cynodonts (Jenkins 1970 1971 Kemp 1980) It isposteroventrally oriented with roughly equal contributions fromthe scapular and coracoid glenoid facets The coracoid glenoidfacet is roughly triangular while the scapular glenoid facet issemiovoid as in other cynodonts In anterior or posterior viewthe fossa appears smoothly crescentic The procoracoid is aslightly curved trapezoidal plate of bone It does not appear tocontribute to the glenoid fossa The procoracoid foramen is lo-cated midway up the height of the procoracoid at a point directlybelow the base of the acromion process The coracoid is semi-crescentic curving posteriorly from a broad base The coracoidin Menadon is longer and more falcate than those described forExaeretodon (Bonaparte 1963) and Luangwa (Kemp 1980 al-though the distal tip of both coracoids is broken in the specimenof Luangwa [TSK 121] described by Kemp and may have beenlonger than reconstructed) but agrees closely with those of Mas-setognathus and cynognathidsdiademodontids (Jenkins 19701971) The distal coracoid tubercle for attachment to the tricepsis strongly curved upwards similar to the condition in Masseto-gnathus (Jenkins 1970) and Diademodon (Brink 1955)
A small fragment of bone resembling a curved cylinder almostcertainly represents part of a clavicle It has no distinguishingfeatures other than a groove associated with a ridge on one sideHowever the degree of curvature observed precludes this ele-ment from representing a long bone and the circular cross-section excludes it from being a rib
The pelvis of FMNH PR 2444 (Fig 12) resembles those ofother traversodontids in general morphology The ilium bears anarrow anteriorly directed iliac blade curving outward distallyThe anterior edge of the iliac blade slopes upward at a roughly45deg angle similar to Luangwa (Kemp 1980) but unlike Ex-aeretodon in which the anterior edge of the iliac blade curvesbackwards (Bonaparte 1963) or Pascualgnathus and Masseto-gnathus in which it is rounded (Bonaparte 1966 Jenkins 1970)The supraacetabular buttress at the base of the ilium is located inan anterodorsal position relative to the acetabulum as in other
traversodontids but unlike earlier cynodonts in which it is lo-cated dorsally (Jenkins 1970) The acetabular buttresses of thethree pelvic bones are largely confluent forming a fairly uniformlsquoliprsquo around the acetabulum as in Exaeretodon (Bonaparte1963) On the contrary in Luangwa the supraacetabular ischialand pubic buttresses are highly distinct spaced 6ndash8 mm apart andwithout confluence of their acetabular facets The pelvis ofFMNH PR 2444 does have a small depression between the su-praacetabular and ischial buttresses (also present in Ex-aeretodon Bonaparte 1963) but this is not as developed as inLuangwa The posterior extent of the iliac blade is broken inFMNH PR 2444 but this region is preserved in another isolatedright pelvis field number 8-31-98-387 (Fig 13) probably pertain-ing to Menadon besairiei The posterior process of the iliac bladein this specimen is relatively short and directed away from thedorsal edge of the ischium as in Exaeretodon (Bonaparte 1963)but unlike Massetognathus and Luangwa in which the posteriorprocess is long and extends roughly horizontally over the dorsaledge of the ischium (Jenkins 1970 Kemp 1980) and evencurves downward distally in Luangwa The posterior process ofthe iliac blade in Pascualgnathus is directed away from the is-chium (Bonaparte 1966) as in Exaeretodon and Menadon but isrelatively long as in Massetognathus and Luangwa
The proximal portion of the ischium is broad and dominatedby the ischial buttress The ischium constricts posterior to theproximal portion and expands distally into a spatulate platewhich is thickened at its posterior margin This thickening con-tinues along the dorsolateral surface of the ischium forming aridge that slopes ventrally until reaching the edge of the ischialbuttress The downward slope of this ridge produces a prominentgroove at the dorsal edge of the lateral surface of the ischiumthought to represent the origin of the ischio-trochantericusmuscle (Kemp 1980) The constriction of the ischium in Mena-don is like that of Luangwa albeit even more extreme in itsventral concavity but unlike that of Exaeretodon in which themedial and distal ends of the ischium are broadly united Theischium in general is a proportionally shorter element in Ex-aeretodon than in other traversodontids including Menadon(Bonaparte 1963) The anteroventral portion of the ischium thatconnects to the pubis is not preserved in either of the referredMenadon pelves
The pubis is the most incompletely preserved pelvic elementThe acetabular contribution of the pubis is smaller than that ofthe ischium or ilium Distal to the acetabulum the pubis is con-stricted indicating that the connection between the acetabularregion and the pubic plate was considerably narrower than inExaeretodon (Bonaparte 1963) but close akin to the pubis ofLuangwa (Kemp 1980) The pubic plate itself is not preservedThe ventral outline of the ischium and what is known of thepubis indicate that the obturator foramen was relatively larger inMenadon (Fig 14) than in Exaeretodon (Bonaparte 1963)
DISCUSSION
Comparison with the Holotype of Menadon besairiei
FMNH PR 2444 differs in several significant ways from theholotype of Menadon besairiei as previously described most no-tably in the possession of an internarial bar (initially consideredabsent in M besairiei) and four upper incisors (reported as threein M besairiei but with the caveat that the homologies of thefourth and fifth anterior tooth loci were problematic Flynn et al2000) However upon further study we interpret the lack of acontinuous internarial bar in the holotype as breakage ratherthan the natural condition of the skull (a prominent medial pro-tuberance is present on the ventral rim of the nares in UA10601) Regarding the incisor count it should be noted that theholotype of Menadon besairiei displays an unusual set of anterior
JOURNAL OF VERTEBRATE PALEONTOLOGY VOL 28 NO 2 2008454
Dow
nloa
ded
by [
UZ
H H
aupt
bibl
ioth
ek
Zen
tral
bibl
ioth
ek Z
uumlric
h] a
t 05
51 1
0 Se
ptem
ber
2013
FIGURE 10 Right scapulocoracoids of FMNH PR 2444 A stereopair of right scapulocoracoid in medial view B stereopair and interpretivedrawing of right scapulocoracoid in lateral view Scale bars equal 5 cm Abbreviationsmdashac acromion process C coracoid c gl coracoid contributionto the glenoid fossa c tr tubercle for coracoid attachment of triceps PC procoracoid pc f procoracoid foramen S scapula s gl scapularcontribution to the glenoid fossa
KAMMERER ET ALmdashCYNODONT FROM MADAGASCAR 455
Dow
nloa
ded
by [
UZ
H H
aupt
bibl
ioth
ek
Zen
tral
bibl
ioth
ek Z
uumlric
h] a
t 05
51 1
0 Se
ptem
ber
2013
teeth for a traversodontid As mentioned above Flynn et al(2000) noted that the homologies of these teeth were difficult todetermine Of the five anterior teeth preserved in the holotypethe anteriormost three are ldquoundoubtedly incisorsrdquo although I3 isldquounusually large and is caniniformrdquo (Flynn et al 2000423) Therobust recurved fourth tooth in the series was considered torepresent the canine with the subsequent tooth interpreted as anolder canine in the process of replacement Although FMNH PR2444 is edentulous the alveoli on the left side of the skull are wellpreserved and indicate an anterior tooth set identical to the ho-lotype The alveolar morphology indicates that the fifth toothwas canted anteriorly as in the holotype This suggests that themysterious fifth tooth is not the result of a transient period in thedental replacement history of M besairiei as suggested by Flynnet al (2000) but rather is the norm This leaves two possibilities1) M besairiei persistently had two canines This condition isunknown in cynodonts 2) The canine-like fourth tooth actuallyrepresents an incisor and the fifth tooth is the true canine Wefavor the latter explanation In FMNH PR 2444 the first fouralveoli are nearly identical in size and shape However the fifthalveolus while similar in anteroposterior length to its predeces-sors is strongly compressed labiolingually with an anteroposte-rior lengthlabiolingual width ratio of 044 (compared to 080 inthe fourth alveolus and 084ndash85 for I2ndash3) We interpret thismarked difference in alveolar ratio as indicating the separationbetween incisors and canine The canine-like I4 would then rep-resent the endpoint in a series of large increasingly caniniformincisors Supporting this interpretation recurved incisors are alsoobserved in juvenile Exaeretodon (eg MCP 3843 PV see Ab-dala et al 2002)
There are several legitimate differences between FMNH PR2444 and the holotype of Menadon besairiei but we take a con-servative taxonomic stance and argue that these differences rep-resent intraspecific variation and preservational artifacts FMNHPR 2444 represents a larger animal than UA 10601 The snoutlength (measured as the distance between the anterior edge ofthe orbit and a vertical plane through the anterior tip of thepremaxilla) of FMNH PR 2444 is 911 mm while that of UA10601 is 801 mm As such proportional differences between thetwo skulls may be attributable to ontogeny Variations in FMNHPR 2444 compared to the holotype that probably represent on-togentic differences include wider longer and deeper paraca-nine fossae a much deeper ventral zygomatic flange (althoughthe lower border of the flange is slightly abraded in the holotypeof Menadon besairiei there is no evidence that a major flangewas broken off) a proportionally shorter and more circular out-line to the posterior border of the maxilla below the zygomaticprocess and a more deeply emarginated posterior squamosal(ie a more prominent laterally projecting flange)
FMNH PR 2444 has a proportionally shorter more robustsnout and larger orbits than the holotype While these charactersare certainly variable in cynodonts it is unusual that the largerspecimen is shorter-snouted and larger-eyed contra the usualpattern The proportional differences between the two skulls areminor however (orbit diametersnout length is 042 for FMNHPR 2444 and 041 for the holotype) and are partially attributableto deformation (shear in FMNH PR 2444 and breakage of thepostorbital bar in the holotype) The following differences canalso probably be attributed to preservational artifact only fivepostcanine alveoli are preserved in FMNH PR 2444 while theholotype has eight postcanines The anteriormost three in theholotype however are tiny and peg-like As the area of themaxilla between the canine alveolus and the first preserved post-canine alveolus is poorly preserved in FMNH PR 2444 the ab-sence of these postcanines may be artifactual However it is alsopossible that the lower postcanine count of FMNH PR 2444compared to the holotype is an ontogenetic effect as postcaninecount is known to decrease with increasing skull size in Ex-
FIGURE 11 Left scapulocoracoids of FMNH PR 2444 A left scapu-locoracoid in medial view B left scapulocoracoid in lateral view Scalebars equal 5 cm
JOURNAL OF VERTEBRATE PALEONTOLOGY VOL 28 NO 2 2008456
Dow
nloa
ded
by [
UZ
H H
aupt
bibl
ioth
ek
Zen
tral
bibl
ioth
ek Z
uumlric
h] a
t 05
51 1
0 Se
ptem
ber
2013
FIGURE 12 Right pelvis of FMNH PR 2444 A stereopair of right pelvis in medial view B stereopair and interpretive drawing of right pelvis inlateral view Scale bars equal 5 cm Abbreviationsmdashbpp base of posterior process of ilium ib iliac blade IL ilium IS ischium ob f obturatorforamen P pubis
KAMMERER ET ALmdashCYNODONT FROM MADAGASCAR 457
Dow
nloa
ded
by [
UZ
H H
aupt
bibl
ioth
ek
Zen
tral
bibl
ioth
ek Z
uumlric
h] a
t 05
51 1
0 Se
ptem
ber
2013
aeretodon (Abdala et al 2002) The orbit of FMNH PR 2444 isless upwardly directed than that of the holotype (being at anangle of sim65deg to a sagittal plane through the midline of the skullas opposed to sim45deg in the holotype) This difference in orbitalangle is probably attributable to postmortem deformation as theskull of FMNH PR 2444 is noticeably sheared to the front leftThe appearance of a wider squarer incisor arcade and broader
anterior palate in FMNH PR 2444 can also be attributed to thisshear Finally the occipital region of FMNH PR 2444 is alto-gether taller and narrower than that of the holotype For ex-ample the foramen magnum in FMNH PR 2444 approximates arounded isosceles triangle in shape while the foramen magnumof the holotype is more ovoid These differences can also beattributed to deformation Altogether the differences between
FIGURE 13 Stereopairs of field number 8-31-98-387 an isolated right pelvis in A medial and B lateral view Scale bars equal 5 cm
JOURNAL OF VERTEBRATE PALEONTOLOGY VOL 28 NO 2 2008458
Dow
nloa
ded
by [
UZ
H H
aupt
bibl
ioth
ek
Zen
tral
bibl
ioth
ek Z
uumlric
h] a
t 05
51 1
0 Se
ptem
ber
2013
FMNH PR 2444 and the holotype fall within the range of intra-specific variation recorded for other gomphodont cynodontssuch as the traversodontids Massetognathus pascuali (Abdalaand Giannini 2000 Kammerer pers obs) and Exaeretodon(Abdala et al 2002) and the diademodontid Diademodon tetra-gonus (Grine et al 1978 Bradu and Grine 1979)
Phylogenetic Analysis
Utilizing the new information afforded by FMNH PR 2444and a reexamination of the holotype (UA 10601) Menadon be-sairiei was recoded in the Gondwanan gomphodont data matrixof Abdala et al (2006) Abdala et alrsquos (2006) data matrix en-compassed 18 taxa and 43 characters For the present analysis anadditional character ldquoincisor procumbencyrdquo (states 0-absent1-present) was added to Abdala et alrsquos (2006) data set (see
Appendix for coding of this character) Several traversodontidtaxa (eg lsquoScalenodonrsquo hirschsoni) have relatively large incisorsbut only in Menadon Scalenodontoides and Exaeretodon are theincisors procumbent Three of Abdala et alrsquos (2006) originalcodings for Menadon besairiei characters 0 (ldquonumber of upperincisorsrdquo) 5 (ldquoposition of paracanine fossa in relation to theupper caninerdquo) and 21 (ldquointernarial barrdquo) were changed to in-corporate data from the new specimen (see Appendix) Addi-tionally further examination of the holotype (UA 10601) ofMenadon besairiei allowed us to code characters 16ndash18 20 38and 40 (morphology of the lower postcanines all marked as ldquordquoin Abdala et alrsquos analysis) Parsimony searches were performedin PAUP v40b10 using a branch-and-bound search Addition-ally a bootstrap analysis (also using branch-and-bound) was per-formed using 1000 replicates
A total of 34 most parsimonious trees of length 103 werefound with the scores CI0573 RI0725 RC0415 andHI0427 The strict consensus tree (Fig 15A) did not recovera polytomy between Menadon Exaeretodon and Scalenodont-oides as in Abdala and Ribeiro (2003) or a (Menadon + Ex-aeretodon) sister-group relationship as in Abdala et al (2006)Rather Menadon was recovered as the sister taxon of the (Ex-aeretodon + Scalenodontoides) clade supporting Flynn et alrsquos(2000) original placement of this taxon There is strong bootstrapsupport (88) for Traversodontidae but within traversodontidsthe only strongly supported subclade is (Gomphodontosuchus +(Menadon + (Scalenodontoides + Exaeretodon))) with a boot-strap value of 80 Within this clade the (Menadon + (Scalen-odontoides + Exaeretodon)) grouping is supported by a boot-strap value of 83 and the (Scalenodontoides + Exaeretodon)grouping is supported by a bootstrap value of 94 (Fig 15B) Inaddition to the positioning of Menadon the strict consensus tree(Fig 15A) differs from the topology of Abdala et al (2006) inrecovering a sister-group relationship between Massetognathusand Santacruzodon rather than having Massetognathus andSantacruzodon as successive outgroups to the clade containingGomphodontosuchus Menadon Scalenodontoides and Ex-aeretodon The (Massetognathus + Santacruzodon) clade is diag-nosed by the absence of an anterolingual cusp in the upper post-canines denticulated incisor cutting margins and the presence ofan anterior cingulum in the upper postcanines
FIGURE 14 Reconstruction of the pelvis of Menadon besairiei basedon information from FMNH PR 2444 and field number 8-31-98-387Dotted area not preserved in either specimen reconstruction based onPascualgnathus (Bonaparte 1966) and Exaeretodon (Bonaparte 1963)
FIGURE 15 Cladogram of traversodontid relationships based on revised data set of Abdala et al (2006) showing A strict consensus tree and B50 majority rule tree Numbers at nodes of majority rule tree represent bootstrap values
KAMMERER ET ALmdashCYNODONT FROM MADAGASCAR 459
Dow
nloa
ded
by [
UZ
H H
aupt
bibl
ioth
ek
Zen
tral
bibl
ioth
ek Z
uumlric
h] a
t 05
51 1
0 Se
ptem
ber
2013
Relationships of Menadon besairiei
Menadon besairiei is the sister taxon to (Exaeretodon + Sca-lenodontoides) the latter clade diagnosed by the possession ofthree upper incisors a posteromedial position of the paracaninefossae relative to the canines the absence of an internarial barlarge canines a projected zygomatic process of the jugal and amaxilla that contributes to the margin of the subtemporal fenes-tra (the last three character states represent reversals) The clade(Menadon + (Exaeretodon + Scalenodontoides)) is diagnosed bythe possession of large procumbent incisors The presence offour incisors a primitive retention distinguishes Menadon be-sairiei from Exaeretodon Scalenodontoides and an unnamed tra-versodontid (ldquoTraversodontid Type 1rdquo) from the Santa MariaFormation of Brazil (Abdala et al 2001 Abdala and Ribeiro2002) Gomphodontosuchus is recovered as the sister taxon to(Menadon (Exaeretodon + Scalenodontoides)) as has beenfound in previous studies (Flynn et al 2000 Abdala and Ribeiro2003) The taxon name Gomphodontosuchinae Watson andRomer 1956 (ex Gomphodontosuchidae Watson and Romer1956 first given subfamilial rank in Lehman 1961) is availablefor this clade As circumscribed here Gomphodontosuchinaeconsists of the most inclusive clade containing Gomphodontosu-chus brasiliensis von Huene 1928264 but not Massetognathuspascuali Romer 19672
Menadon besairiei displays a mixture of primitive and derivedcharacters within traversodontids The internarial bar and fourincisors are primitive features whereas the enlargement and pro-cumbency of the incisors are derived features shared with Ex-aeretodon and Scalenodontoides The postcranium shows severalsimilarities to that of Exaeretodon (apparent exclusion of theprocoracoid from the glenoid some features of the pelvis tallcaudal neural spines) but in general displays a rather conserva-tive morphology observed throughout Gomphodontia The mor-phology of the scapulocoracoids is closer akin to diademodontidsor trirachodontids than Exaeretodon and the expanded thoracicrib of Menadon has only slight differences from those of trirach-odontids or early traversodontids like Luangwa The atlas-axiscomplex of Menadon is similar to that of the Early Triassic cyn-odont Cynognathus in possessing a concave axial spine with along posterior process Although most gomphodonts have beenreconstructed with a convex axial spine as in early mammals thisspine is broken in most of the best described gomphodont post-crania (eg Diademodon Brink 1955) and the lsquosaddle-shapedrsquoaxial morphology may be more widely distributed among cyn-odonts than previously realized The new specimens of Menadondescribed in this paper demonstrate that a generalized gompho-dont postcranial morphology was retained deep within the tra-versodontid tree and that the derived morphologies of either ofthe well-known taxa Massetognathus or Exaeretodon should notbe taken as representative of traversodontids in general
ACKNOWLEDGMENTS
This work would not have been possible without the continu-ing support and encouragement of our collaborative programwith the Deacutepartement de Paleacuteontologie et drsquoAnthropologie Bi-ologique Universiteacute drsquoAntananarivo Madagascar under the di-rection of Armand Rasoamiaramanana (Department Chairman)and Gisegravele Ravololonarivo Randria (Head of the PaleontologyLaboratories) We are extremely grateful to the National Geo-graphic Society for their long-term support of the fieldwork as-sociated with this project Additional financial or logistical sup-port for fieldwork was provided by The Field Museum the Uni-versity of California-Santa Barbara and the WWF (WorldwideFund for Nature Madagascar) We acknowledge the Ministegraverede LrsquoEneacutergie et des Mines for authorizing field efforts and for
continuing to facilitate our joint US-Madagascar explorationresearch and education program Steve Goodman and AsminaGandhi generously assisted us in innumerable ways includinghospitality nonpareil We especially thank R Andriantompoha-vana L Razafimanantsoa A Toto Volahy JA Rabarison AGandie and P Vavisaro for assistance in the field during theexpedition in which these new specimens were discovered BillSimpson and Gina Wesley-Hunt for their patient excavation ofthis important material Allison Smith Ray Vodden Lisa Berg-wall and Deborah Wagner for excellent preparation of a chal-lenging specimen Jennifer Botha and Elize Butler (NationalMuseum Bloemfontein) Michael Raath and Fernando Abdala(Bernard Price Institute) Thomas Kemp (Oxford UniversityMuseum of Natural History) Ray Symonds (University Museumof Zoology Cambridge) Bernard Battail (Museacuteum nationaldrsquoHistoire naturelle Paris) Michael Maisch (Universitaumlt Tuumlbin-gen) and Charles Schaff and Farish Jenkins (Museum of Com-parative Zoology Harvard) for access to specimens and AgustiacutenMartinelli Fernando Abdala and James Hopson for helpful re-views of an earlier version of the manuscript
LITERATURE CITED
Abdala F M C Barbarena and J Dornelles 2002 A new species of thetraversodontid cynodont Exaeretodon from the Santa Maria Forma-tion (MiddleLate Triassic) of Southern Brazil Journal of Verte-brate Paleontology 22313ndash325
Abdala F and N P Giannini 2000 Gomphodont cynodonts of theChantildeares Formation the analysis of an ontogenetic sequence Jour-nal of Vertebrate Paleontology 20501ndash506
Abdala F J Neveling and J Welman 2006 A new trirachodontidcynodont from the lower levels of the Burgersdorp Formation(Lower Triassic) of the Beaufort Group South Africa and the cla-distic relationships of Gondwanan gomphodonts Zoological Jour-nal of the Linnean Society 147383ndash413
Abdala F A M Ribeiro and C L Schultz 2001 A rich cynodont faunaof Santa Cruz do Sul Santa Maria Formation (Middle-Late Trias-sic) southern Brazil Neues Jahrbuch fuumlr Geologie und Palaumlontolo-gie Monatshefte 2001669ndash687
Abdala F and A M Ribeiro 2002 Nuevos cinodontes traversodoacuten-tidos (Synapsida-Eucynodontia) de la Formacioacuten Santa Maria(Triaacutesico Medio-Superior) Rio Grande do Sul Brasil RevistaEspantildeola de Paleontologiacutea 17237ndash243
Abdala F and A M Ribeiro 2003 A new traversodontid cynodontfrom the Santa Maria Formation (Ladinian-Carnian) of southernBrazil with a phylogenetic analysis of Gondwanan traversodontidsZoological Journal of the Linnaean Society 139529ndash545
Abdala F and A M S Teixeira 2004 A traversodontid cynodont ofAfrican affinity in the South American Triassic Palaeontologia Af-ricana 4011ndash22
Bonaparte J F 1962 Descripcioacuten del craacuteneo y mandiacutebula de Ex-aeretodon frenguellii Cabrera y su comparacioacuten con Diademodon-tidae Tritylodontidae y los cinodontes sudamericanos Publica-ciones del Museo Municipal de Ciencias Naturales y Tradicional deMar del Plata 1135ndash202
Bonaparte J F 1963 Descripcioacuten del esqueleto postcraneano de Ex-aeretodon (CynodontiandashTraversodontidae) Acta Geoloacutegica Lil-loana 45ndash52
Bonaparte J F 1966 Una nueva ldquofaunardquo Triaacutesica de Argentina (Ther-apsida Cynodontia Dicynodontia) Consideracions filogeneacuteticas ypaleobiogeograacuteficas Ameghiniana 8243ndash296
Bradu D and F E Grine 1979 Multivariate analysis of diademodon-tine crania from South Africa and Zambia South African Journal ofScience 79441ndash448
Brink A S 1955 A study of the skeleton of Diademodon Palaeonto-logia Africana 33-39
Brink A S 1963 Two new cynodonts from the Ntawere Formation inthe Luangwa Valley of Northern Rhodesia Palaeontologia Africana877ndash96
Broom R 1905 On the use of the term Anomodontia Records of theAlbany Museum 1266ndash269
Cabrera A 1943 El primer hallazgo de teraacutepsidos en la ArgentinaNotas del Museo de La Plata Paleontologiacutea 55317ndash331
JOURNAL OF VERTEBRATE PALEONTOLOGY VOL 28 NO 2 2008460
Dow
nloa
ded
by [
UZ
H H
aupt
bibl
ioth
ek
Zen
tral
bibl
ioth
ek Z
uumlric
h] a
t 05
51 1
0 Se
ptem
ber
2013
Chatterjee S 1982 A new cynodont reptile from the Triassic of IndiaJournal of Paleontology 56203ndash214
Crompton A W 1955 On some Triassic cynodonts from TanganyikaProceedings of the Zoological Society of London 125617ndash669
Crompton A W 1972 Postcanine occlusion in cynodonts and tritylo-donts Bulletin of the British Museum (Natural History) Geology2129ndash71
Crompton A W and F Ellenberger 1957 On a new cynodont from theMolteno Beds and the origin of the tritylodontids Annals of theSouth African Museum 441ndash13
Flynn J J J M Parrish B Rakotosaminimanana W F Simpson R LWhatley and A R Wyss 1999 A Triassic fauna from Madagascarincluding early dinosaurs Science 286763ndash765
Flynn J J J M Parrish B Rakotosamimanana L RanivoharimananaW F Simpson and A R Wyss 2000 New traversodontids (Syn-apsida Eucynodontia) from the Triassic of Madagascar Journal ofVertebrate Paleontology 20422ndash427
Gow C E and P J Hancox 1993 First complete skull of the LateTriassic Scalenodontoides (Reptilia Cynodontia) from southern Af-rica In Lucas S G and M Morales (eds) The nonmarine TriassicNew Mexico Museum of Natural History amp Science Bulletin 3161ndash168
Grine F E B D Hahn and C E Gow 1978 Aspects of relativegrowth and variability in Diademodon (Reptilia Therapsida) SouthAfrican Journal of Science 7450ndash58
Hopson J A 1984 Late Triassic traversodont cynodonts from NovaScotia and southern Africa Palaeontologia Africana 25181ndash201
Hopson J A 1985 Morphology and relationships of Gomphodontosu-chus brasiliensis von Huene (Synapsida Cynodontia Tritylodontoi-dea) from the Triassic of Brazil Neues Jahrbuch fuumlr Geologie undPalaumlontologie Monatshefte 1985285ndash299
Hopson J A and J W Kitching 2001 A probainognathian cynodontfrom South Africa and the phylogeny of nonmammalian cynodontsBulletin of the Museum of Comparative Zoology 1565ndash35
Hopson J A and H-D Sues 2006 A traversodont cynodont from theMiddle Triassic (Ladinian) of Baden-Wuumlrttemberg (Germany)Palaumlontologische Zeitschrift 80124ndash129
Huene F von 1928 Ein Cynodontier aus der Trias Brasiliens Zentral-blatt fuumlr Mineralogie Geologie und Palaumlontologie 1928251ndash270
Huene F von 1936 Die fossilen Reptilien des suumldamerikanischenGondwanalandes Ergebnisse der Sauriergrabungen in Suumldbrasilien192829 Lieferung 2 (Cynodontia)93ndash159 Franz F Heine Tuumlbin-gen
Jenkins F A 1970 The Chantildeares (Argentina) Triassic reptile faunaVII The postcranial skeleton of the traversodontid Massetognathuspascuali (Therapsida Cynodontia) Breviora 3521ndash28
Jenkins F A 1971 The postcranial skeleton of African cynodonts prob-lems in the early evolution of the mammalian postcranial skeletonBulletin of the Peabody Museum of Natural History 361ndash216
Kemp T S 1969 The atlas-axis complex of mammal-like reptiles Jour-nal of Zoology 159223ndash248
Kemp T S 1980 Aspects of the structure and functional anatomy of theMiddle Triassic cynodont Luangwa Journal of Zoology 191193ndash239
Kemp T S 1982 Mammal-Like Reptiles and the Origin of MammalsAcademic Press New York
Lehman J-P 1961 Cynodontia pp 140ndash191 in Piveteau J (ed) Traiteacutede Paleacuteontologie Masson et Cie Paris
Osborn 1903 On the primary division of the Reptilia into two sub-classes Synapsida and Diapsida Science 17275ndash276
Owen R 1861 Palaeontology or a systematic summary of extinct ani-mals and their geological relations 2nd Ed Edinburgh
Parrington F R 1934 On the cynodont genus Galesaurus with a noteon the functional significance of the changes in the evolution of thetheriodont skull Annals and Magazine of Natural History 10(13)38ndash67
Parrington F R 1946 On the cranial anatomy of cynodonts Proceed-ings of the Zoological Society of London 116181ndash197
Razafimbelo E 1987 Le basin de Morondava (Madagascar) Synthegravesegeacuteologique et structurale PhD dissertation University Louis Pas-teur Strasbourg France 189 pp
Reisz R R and H-D Sues 2000 Herbivory in Late Paleozoic andTriassic terrestrial vertebrates pp 9ndash41 in Sues H-D (ed) Evo-lution of Herbivory in Terrestrial Vertebrates Cambridge Univer-sity Press New York
Romer A S 1967 The Chantildeares (Argentina) Triassic reptile fauna IIITwo new gomphodonts Massetognathus pascuali and M terugiiBreviora 2641ndash25
Romer A S 1972 The Chantildeares (Argentina) Triassic reptile faunaXVII The Chantildeares gomphodonts Breviora 3961ndash9
Rougier G W J R Wible and J A Hopson 1992 Reconstruction ofthe cranial vessels in the Early Cretaceous mammal Vincelestes neu-quenianus implications for the evolution of the mammalian cranialvascular system Journal of Vertebrate Paleontology 12188ndash216
Rowe T 1988 Definition diagnosis and origin of Mammalia Journal ofVertebrate Paleontology 8241ndash264
Rowe T 1993 Early mammal phylogenetic systematics pp 129ndash145 inSzalay F S M J Novacek and M C McKenna (eds) MammalPhylogeny Vol 1 Mesozoic Differentiation MultituberculatesMonotremes Early Therians and Marsupials Springer-Verlag NewYork
Seeley H G 1894 Researches on the structure organization and clas-sification of the fossil Reptilia Part IX Section 3 On DiademodonPhilosophical Transactions of the Royal Society of London 1851029ndash1041
Seeley H G 1895 Researches on the structure organization and clas-sification of the fossil Reptilia Part IX Section 4 On the Gompho-dontia Philosophical Transactions of the Royal Society of London1861ndash57
Seeley H G 1895 Researches on the structure organization and clas-sification of the fossil Reptilia Part IX Section 5 On the skeletonin new Cynodontia from the Karroo rocks Philosophical Transac-tions of the Royal Society of London 18659ndash148
Sues H-D J A Hopson and N H Shubin 1992 Affinities of Sca-lenodontoides plemmyridon Hopson 1984 (Synapsida Cynodontia)from the Upper Triassic of Nova Scotia Journal of Vertebrate Pa-leontology 12168ndash171
Sues H-D and P E Olsen 1990 Triassic vertebrates of Gondwananaspect from the Richmond basin of Virginia Science 2491020ndash1023
Swofford D L 2003 PAUP v 40b10 Sinauer Associates Inc Pub-lishers Sunderland Massachusetts
Watson D M S and A S Romer 1956 A classification of therapsidreptiles Bulletin of the Museum of Comparative Zoology 11435ndash89
Submitted September 20 2007 accepted November 3 2007
KAMMERER ET ALmdashCYNODONT FROM MADAGASCAR 461
Dow
nloa
ded
by [
UZ
H H
aupt
bibl
ioth
ek
Zen
tral
bibl
ioth
ek Z
uumlric
h] a
t 05
51 1
0 Se
ptem
ber
2013
APPENDIX Data matrix used in the phylogenetic analysesThis is a revised version of the matrix of Abdala et al (2006) with the following changes character 43 [Incisor procumbency (0) absent (1) present]added Menadon recoded for characters 0 (Number of upper incisors was 1 ldquothreerdquo changed to 0 ldquofourrdquo) 5 (Position of paracanine fossae in relationto the upper canine was changed to 1 ldquomedialrdquo) 16 (Overall morphology of lower postcanines in occlusal view was changed to 3 ldquoquadran-gularrdquo) 17 (Position of the transverse crest in lower postcanines was changed to 1 ldquoanteriorrdquo) 18 (Number of cusps in the transverse crest of thelower postcanines was changed to 0 ldquotwordquo) 20 (Anterior cingulum in the lower postcanines was changed to 2 ldquoabsentrdquo) 21 (Internarial barwas 1 ldquoabsentrdquo changed to 0 ldquopresentrdquo) 38 (Posterior cingulum in the lower postcanines was changed to 1 ldquoabsentrdquo) and 40 (Deep occlusalbasins in the postcanines was changed to 1 ldquopresentrdquo) Langbergia recoded for character 34 (Incisor cutting margins was 0 ldquoserratedrdquo changedto 1 ldquosmoothly ridgedrdquo) based on the description of the incisors in Abdala et al (2006) Gomphodontosuchus recoded for character 31 (Maxilla inthe margin of the subtemporal fenestra was 1 ldquoincludedrdquo changed to 0 ldquoexcludedrdquo) based on von Huene (1936) and Kammerer (pers obs of theholotype) contra Hopsonrsquos (1985) reconstruction lsquoScalenodonrsquo hirschsoni recoded for character 39 (Anterior cingulum in the upper postcanines was1 ldquoabsentrdquo changed to 0 ldquopresentrdquo) based on Abdala (pers comm 2006) Trirachodon recoded for character 30 (Ectopterygoid was changedto 0 ldquopresentrdquo) based on Abdala (pers comm 2006) and Traversodon recoded for character 15 (Number of cusps in the sectorial border of the upperpostcanines was 0 ldquomore than twordquo changed to 1 ldquotwordquo) based on Abdala (pers comm 2006) and Kammerer (pers obs)
1111111111 2222222222 3333333333 44440123456789 0123456789 0123456789 0123456789 0123
Cynognathus 00000000 000 000000000 00000000 000Diademodon 0000001000 0000100 000010A000 0000000100 0100Trirachodon 000000100B 1A002201 001A100010 0001001200 0110Langbergia 0000001000 10000201 0000101010 0001101200 0110Cricodon 0100001000 11000201 0010100 0001200 0110Andescynodon 0000002001 01013101 101120010 0112311 1100Massetognathus 00111121A2 1110003100 2011200011 1011212300 1100Exaeretodon 1110122112 010103101 2111110201 1111112311 1101Luangwa 0000001002 11010A3100 101010010 01002300 1100Scal angustifrons 000000100B 1111023100 10102101 00002301 1100Scal hirschsoni 11110202 1100113100 1010 1112300 1100Traversodon 0000002012 1101113100 201010 011230 1100Gomphodontosuchus 0011112112 0103101 2010 0112311 1100Pascualgnathus 1000001000 000231 1100001 0011230 1100Scalenodontoides 1112212 01003101 21111021 1212311 1101Menadon 0111112112 01012310 20121020 0102311 1101Dadadon 00002112 111112 013 11231 1100Santacruzodon 00002112 1110003101 201300 212300 1100
A(01) B(02)
JOURNAL OF VERTEBRATE PALEONTOLOGY VOL 28 NO 2 2008462
Dow
nloa
ded
by [
UZ
H H
aupt
bibl
ioth
ek
Zen
tral
bibl
ioth
ek Z
uumlric
h] a
t 05
51 1
0 Se
ptem
ber
2013
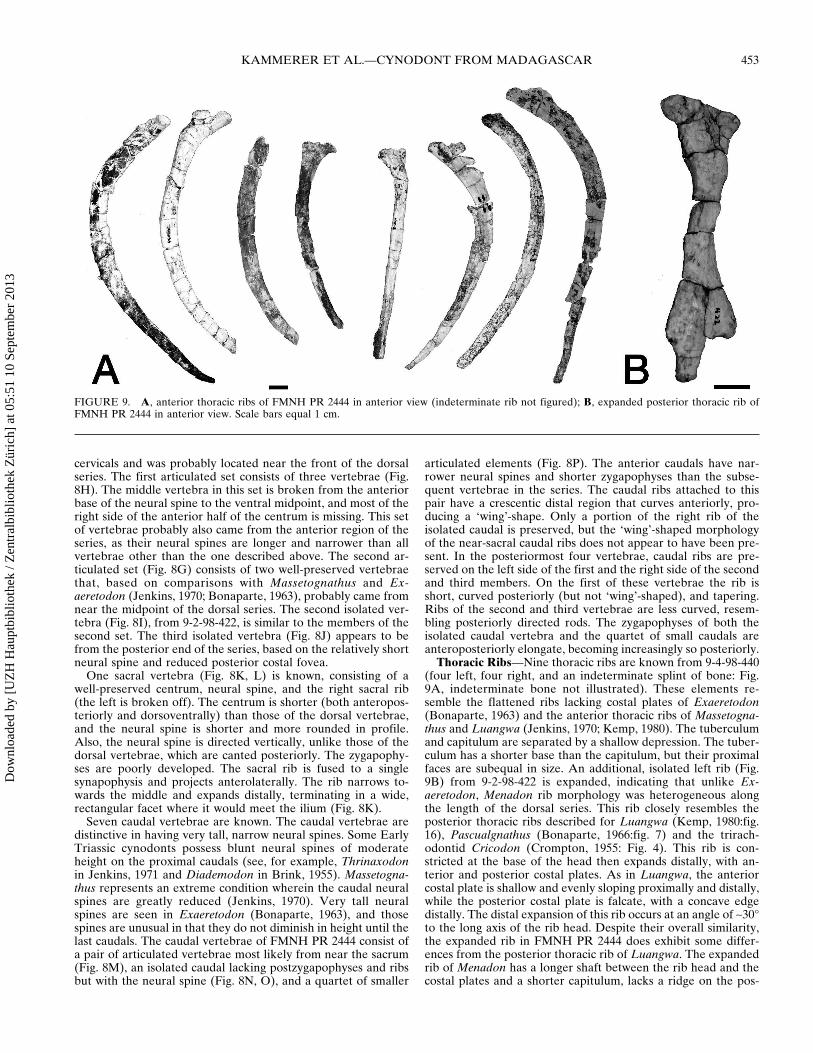
cervicals and was probably located near the front of the dorsalseries The first articulated set consists of three vertebrae (Fig8H) The middle vertebra in this set is broken from the anteriorbase of the neural spine to the ventral midpoint and most of theright side of the anterior half of the centrum is missing This setof vertebrae probably also came from the anterior region of theseries as their neural spines are longer and narrower than allvertebrae other than the one described above The second ar-ticulated set (Fig 8G) consists of two well-preserved vertebraethat based on comparisons with Massetognathus and Ex-aeretodon (Jenkins 1970 Bonaparte 1963) probably came fromnear the midpoint of the dorsal series The second isolated ver-tebra (Fig 8I) from 9-2-98-422 is similar to the members of thesecond set The third isolated vertebra (Fig 8J) appears to befrom the posterior end of the series based on the relatively shortneural spine and reduced posterior costal fovea
One sacral vertebra (Fig 8K L) is known consisting of awell-preserved centrum neural spine and the right sacral rib(the left is broken off) The centrum is shorter (both anteropos-teriorly and dorsoventrally) than those of the dorsal vertebraeand the neural spine is shorter and more rounded in profileAlso the neural spine is directed vertically unlike those of thedorsal vertebrae which are canted posteriorly The zygapophy-ses are poorly developed The sacral rib is fused to a singlesynapophysis and projects anterolaterally The rib narrows to-wards the middle and expands distally terminating in a widerectangular facet where it would meet the ilium (Fig 8K)
Seven caudal vertebrae are known The caudal vertebrae aredistinctive in having very tall narrow neural spines Some EarlyTriassic cynodonts possess blunt neural spines of moderateheight on the proximal caudals (see for example Thrinaxodonin Jenkins 1971 and Diademodon in Brink 1955) Massetogna-thus represents an extreme condition wherein the caudal neuralspines are greatly reduced (Jenkins 1970) Very tall neuralspines are seen in Exaeretodon (Bonaparte 1963) and thosespines are unusual in that they do not diminish in height until thelast caudals The caudal vertebrae of FMNH PR 2444 consist ofa pair of articulated vertebrae most likely from near the sacrum(Fig 8M) an isolated caudal lacking postzygapophyses and ribsbut with the neural spine (Fig 8N O) and a quartet of smaller
articulated elements (Fig 8P) The anterior caudals have nar-rower neural spines and shorter zygapophyses than the subse-quent vertebrae in the series The caudal ribs attached to thispair have a crescentic distal region that curves anteriorly pro-ducing a lsquowingrsquo-shape Only a portion of the right rib of theisolated caudal is preserved but the lsquowingrsquo-shaped morphologyof the near-sacral caudal ribs does not appear to have been pre-sent In the posteriormost four vertebrae caudal ribs are pre-served on the left side of the first and the right side of the secondand third members On the first of these vertebrae the rib isshort curved posteriorly (but not lsquowingrsquo-shaped) and taperingRibs of the second and third vertebrae are less curved resem-bling posteriorly directed rods The zygapophyses of both theisolated caudal vertebra and the quartet of small caudals areanteroposteriorly elongate becoming increasingly so posteriorly
Thoracic RibsmdashNine thoracic ribs are known from 9-4-98-440(four left four right and an indeterminate splint of bone Fig9A indeterminate bone not illustrated) These elements re-semble the flattened ribs lacking costal plates of Exaeretodon(Bonaparte 1963) and the anterior thoracic ribs of Massetogna-thus and Luangwa (Jenkins 1970 Kemp 1980) The tuberculumand capitulum are separated by a shallow depression The tuber-culum has a shorter base than the capitulum but their proximalfaces are subequal in size An additional isolated left rib (Fig9B) from 9-2-98-422 is expanded indicating that unlike Ex-aeretodon Menadon rib morphology was heterogeneous alongthe length of the dorsal series This rib closely resembles theposterior thoracic ribs described for Luangwa (Kemp 1980fig16) Pascualgnathus (Bonaparte 1966fig 7) and the trirach-odontid Cricodon (Crompton 1955 Fig 4) This rib is con-stricted at the base of the head then expands distally with an-terior and posterior costal plates As in Luangwa the anteriorcostal plate is shallow and evenly sloping proximally and distallywhile the posterior costal plate is falcate with a concave edgedistally The distal expansion of this rib occurs at an angle of sim30degto the long axis of the rib head Despite their overall similaritythe expanded rib in FMNH PR 2444 does exhibit some differ-ences from the posterior thoracic rib of Luangwa The expandedrib of Menadon has a longer shaft between the rib head and thecostal plates and a shorter capitulum lacks a ridge on the pos-
FIGURE 9 A anterior thoracic ribs of FMNH PR 2444 in anterior view (indeterminate rib not figured) B expanded posterior thoracic rib ofFMNH PR 2444 in anterior view Scale bars equal 1 cm
KAMMERER ET ALmdashCYNODONT FROM MADAGASCAR 453
Dow
nloa
ded
by [
UZ
H H
aupt
bibl
ioth
ek
Zen
tral
bibl
ioth
ek Z
uumlric
h] a
t 05
51 1
0 Se
ptem
ber
2013
terior costal plate and has an anteroposteriorly shorter anteriorcostal plate Additionally the posterior margin of the costal plateis less curved than that of Luangwa These differences couldindicate that the expanded rib of Menadon is from a more pos-terior position in the series than the rib from Luangwa as re-duced curvature and increasing confluence of the tuberculumand capitulum are characteristic of lumbar ribs in traversodon-tids (Jenkins 1970 Kemp 1980) However the length of theshaft is at odds with this interpretation and in Cricodon andPascualgnathus the more posterior ribs are not more similar tothe element from FMNH PR 2444 These differences probablyrepresent legitimate morphological distinctions between the ribsof earlier gomphodonts and Menadon The head of the expandedrib is well defined and unbroken indicating that it was not syn-ostosed to a vertebra
Appendicular Skeleton
The appendicular skeleton is represented by both scapulocora-coids and a fragment of a clavicle from field number 9-4-98-440and a partial right pelvis from field number 9-2-98-422 An ad-ditional partial right pelvis is represented from field number8-31-98-387 The scapulocoracoids (Figs 10 11) are very similarto those described for other traversodontids as well as cynog-nathids and diademodontids (Jenkins 1970 1971) The scapularinfraspinous fossa has out-turned anterior and posterior edgesThe anterior edge of the scapula is rounded and curves laterallyforming the scapular spine At the lower end of the spine theacromion forms a prominent process (broken in the left scapula)Below this process the scapula is constricted and then expandsventrally to form the dorsal margin of the articular glenoid Theglenoid fossa is in general morphology very similar to that ofother Triassic cynodonts (Jenkins 1970 1971 Kemp 1980) It isposteroventrally oriented with roughly equal contributions fromthe scapular and coracoid glenoid facets The coracoid glenoidfacet is roughly triangular while the scapular glenoid facet issemiovoid as in other cynodonts In anterior or posterior viewthe fossa appears smoothly crescentic The procoracoid is aslightly curved trapezoidal plate of bone It does not appear tocontribute to the glenoid fossa The procoracoid foramen is lo-cated midway up the height of the procoracoid at a point directlybelow the base of the acromion process The coracoid is semi-crescentic curving posteriorly from a broad base The coracoidin Menadon is longer and more falcate than those described forExaeretodon (Bonaparte 1963) and Luangwa (Kemp 1980 al-though the distal tip of both coracoids is broken in the specimenof Luangwa [TSK 121] described by Kemp and may have beenlonger than reconstructed) but agrees closely with those of Mas-setognathus and cynognathidsdiademodontids (Jenkins 19701971) The distal coracoid tubercle for attachment to the tricepsis strongly curved upwards similar to the condition in Masseto-gnathus (Jenkins 1970) and Diademodon (Brink 1955)
A small fragment of bone resembling a curved cylinder almostcertainly represents part of a clavicle It has no distinguishingfeatures other than a groove associated with a ridge on one sideHowever the degree of curvature observed precludes this ele-ment from representing a long bone and the circular cross-section excludes it from being a rib
The pelvis of FMNH PR 2444 (Fig 12) resembles those ofother traversodontids in general morphology The ilium bears anarrow anteriorly directed iliac blade curving outward distallyThe anterior edge of the iliac blade slopes upward at a roughly45deg angle similar to Luangwa (Kemp 1980) but unlike Ex-aeretodon in which the anterior edge of the iliac blade curvesbackwards (Bonaparte 1963) or Pascualgnathus and Masseto-gnathus in which it is rounded (Bonaparte 1966 Jenkins 1970)The supraacetabular buttress at the base of the ilium is located inan anterodorsal position relative to the acetabulum as in other
traversodontids but unlike earlier cynodonts in which it is lo-cated dorsally (Jenkins 1970) The acetabular buttresses of thethree pelvic bones are largely confluent forming a fairly uniformlsquoliprsquo around the acetabulum as in Exaeretodon (Bonaparte1963) On the contrary in Luangwa the supraacetabular ischialand pubic buttresses are highly distinct spaced 6ndash8 mm apart andwithout confluence of their acetabular facets The pelvis ofFMNH PR 2444 does have a small depression between the su-praacetabular and ischial buttresses (also present in Ex-aeretodon Bonaparte 1963) but this is not as developed as inLuangwa The posterior extent of the iliac blade is broken inFMNH PR 2444 but this region is preserved in another isolatedright pelvis field number 8-31-98-387 (Fig 13) probably pertain-ing to Menadon besairiei The posterior process of the iliac bladein this specimen is relatively short and directed away from thedorsal edge of the ischium as in Exaeretodon (Bonaparte 1963)but unlike Massetognathus and Luangwa in which the posteriorprocess is long and extends roughly horizontally over the dorsaledge of the ischium (Jenkins 1970 Kemp 1980) and evencurves downward distally in Luangwa The posterior process ofthe iliac blade in Pascualgnathus is directed away from the is-chium (Bonaparte 1966) as in Exaeretodon and Menadon but isrelatively long as in Massetognathus and Luangwa
The proximal portion of the ischium is broad and dominatedby the ischial buttress The ischium constricts posterior to theproximal portion and expands distally into a spatulate platewhich is thickened at its posterior margin This thickening con-tinues along the dorsolateral surface of the ischium forming aridge that slopes ventrally until reaching the edge of the ischialbuttress The downward slope of this ridge produces a prominentgroove at the dorsal edge of the lateral surface of the ischiumthought to represent the origin of the ischio-trochantericusmuscle (Kemp 1980) The constriction of the ischium in Mena-don is like that of Luangwa albeit even more extreme in itsventral concavity but unlike that of Exaeretodon in which themedial and distal ends of the ischium are broadly united Theischium in general is a proportionally shorter element in Ex-aeretodon than in other traversodontids including Menadon(Bonaparte 1963) The anteroventral portion of the ischium thatconnects to the pubis is not preserved in either of the referredMenadon pelves
The pubis is the most incompletely preserved pelvic elementThe acetabular contribution of the pubis is smaller than that ofthe ischium or ilium Distal to the acetabulum the pubis is con-stricted indicating that the connection between the acetabularregion and the pubic plate was considerably narrower than inExaeretodon (Bonaparte 1963) but close akin to the pubis ofLuangwa (Kemp 1980) The pubic plate itself is not preservedThe ventral outline of the ischium and what is known of thepubis indicate that the obturator foramen was relatively larger inMenadon (Fig 14) than in Exaeretodon (Bonaparte 1963)
DISCUSSION
Comparison with the Holotype of Menadon besairiei
FMNH PR 2444 differs in several significant ways from theholotype of Menadon besairiei as previously described most no-tably in the possession of an internarial bar (initially consideredabsent in M besairiei) and four upper incisors (reported as threein M besairiei but with the caveat that the homologies of thefourth and fifth anterior tooth loci were problematic Flynn et al2000) However upon further study we interpret the lack of acontinuous internarial bar in the holotype as breakage ratherthan the natural condition of the skull (a prominent medial pro-tuberance is present on the ventral rim of the nares in UA10601) Regarding the incisor count it should be noted that theholotype of Menadon besairiei displays an unusual set of anterior
JOURNAL OF VERTEBRATE PALEONTOLOGY VOL 28 NO 2 2008454
Dow
nloa
ded
by [
UZ
H H
aupt
bibl
ioth
ek
Zen
tral
bibl
ioth
ek Z
uumlric
h] a
t 05
51 1
0 Se
ptem
ber
2013
FIGURE 10 Right scapulocoracoids of FMNH PR 2444 A stereopair of right scapulocoracoid in medial view B stereopair and interpretivedrawing of right scapulocoracoid in lateral view Scale bars equal 5 cm Abbreviationsmdashac acromion process C coracoid c gl coracoid contributionto the glenoid fossa c tr tubercle for coracoid attachment of triceps PC procoracoid pc f procoracoid foramen S scapula s gl scapularcontribution to the glenoid fossa
KAMMERER ET ALmdashCYNODONT FROM MADAGASCAR 455
Dow
nloa
ded
by [
UZ
H H
aupt
bibl
ioth
ek
Zen
tral
bibl
ioth
ek Z
uumlric
h] a
t 05
51 1
0 Se
ptem
ber
2013
teeth for a traversodontid As mentioned above Flynn et al(2000) noted that the homologies of these teeth were difficult todetermine Of the five anterior teeth preserved in the holotypethe anteriormost three are ldquoundoubtedly incisorsrdquo although I3 isldquounusually large and is caniniformrdquo (Flynn et al 2000423) Therobust recurved fourth tooth in the series was considered torepresent the canine with the subsequent tooth interpreted as anolder canine in the process of replacement Although FMNH PR2444 is edentulous the alveoli on the left side of the skull are wellpreserved and indicate an anterior tooth set identical to the ho-lotype The alveolar morphology indicates that the fifth toothwas canted anteriorly as in the holotype This suggests that themysterious fifth tooth is not the result of a transient period in thedental replacement history of M besairiei as suggested by Flynnet al (2000) but rather is the norm This leaves two possibilities1) M besairiei persistently had two canines This condition isunknown in cynodonts 2) The canine-like fourth tooth actuallyrepresents an incisor and the fifth tooth is the true canine Wefavor the latter explanation In FMNH PR 2444 the first fouralveoli are nearly identical in size and shape However the fifthalveolus while similar in anteroposterior length to its predeces-sors is strongly compressed labiolingually with an anteroposte-rior lengthlabiolingual width ratio of 044 (compared to 080 inthe fourth alveolus and 084ndash85 for I2ndash3) We interpret thismarked difference in alveolar ratio as indicating the separationbetween incisors and canine The canine-like I4 would then rep-resent the endpoint in a series of large increasingly caniniformincisors Supporting this interpretation recurved incisors are alsoobserved in juvenile Exaeretodon (eg MCP 3843 PV see Ab-dala et al 2002)
There are several legitimate differences between FMNH PR2444 and the holotype of Menadon besairiei but we take a con-servative taxonomic stance and argue that these differences rep-resent intraspecific variation and preservational artifacts FMNHPR 2444 represents a larger animal than UA 10601 The snoutlength (measured as the distance between the anterior edge ofthe orbit and a vertical plane through the anterior tip of thepremaxilla) of FMNH PR 2444 is 911 mm while that of UA10601 is 801 mm As such proportional differences between thetwo skulls may be attributable to ontogeny Variations in FMNHPR 2444 compared to the holotype that probably represent on-togentic differences include wider longer and deeper paraca-nine fossae a much deeper ventral zygomatic flange (althoughthe lower border of the flange is slightly abraded in the holotypeof Menadon besairiei there is no evidence that a major flangewas broken off) a proportionally shorter and more circular out-line to the posterior border of the maxilla below the zygomaticprocess and a more deeply emarginated posterior squamosal(ie a more prominent laterally projecting flange)
FMNH PR 2444 has a proportionally shorter more robustsnout and larger orbits than the holotype While these charactersare certainly variable in cynodonts it is unusual that the largerspecimen is shorter-snouted and larger-eyed contra the usualpattern The proportional differences between the two skulls areminor however (orbit diametersnout length is 042 for FMNHPR 2444 and 041 for the holotype) and are partially attributableto deformation (shear in FMNH PR 2444 and breakage of thepostorbital bar in the holotype) The following differences canalso probably be attributed to preservational artifact only fivepostcanine alveoli are preserved in FMNH PR 2444 while theholotype has eight postcanines The anteriormost three in theholotype however are tiny and peg-like As the area of themaxilla between the canine alveolus and the first preserved post-canine alveolus is poorly preserved in FMNH PR 2444 the ab-sence of these postcanines may be artifactual However it is alsopossible that the lower postcanine count of FMNH PR 2444compared to the holotype is an ontogenetic effect as postcaninecount is known to decrease with increasing skull size in Ex-
FIGURE 11 Left scapulocoracoids of FMNH PR 2444 A left scapu-locoracoid in medial view B left scapulocoracoid in lateral view Scalebars equal 5 cm
JOURNAL OF VERTEBRATE PALEONTOLOGY VOL 28 NO 2 2008456
Dow
nloa
ded
by [
UZ
H H
aupt
bibl
ioth
ek
Zen
tral
bibl
ioth
ek Z
uumlric
h] a
t 05
51 1
0 Se
ptem
ber
2013
FIGURE 12 Right pelvis of FMNH PR 2444 A stereopair of right pelvis in medial view B stereopair and interpretive drawing of right pelvis inlateral view Scale bars equal 5 cm Abbreviationsmdashbpp base of posterior process of ilium ib iliac blade IL ilium IS ischium ob f obturatorforamen P pubis
KAMMERER ET ALmdashCYNODONT FROM MADAGASCAR 457
Dow
nloa
ded
by [
UZ
H H
aupt
bibl
ioth
ek
Zen
tral
bibl
ioth
ek Z
uumlric
h] a
t 05
51 1
0 Se
ptem
ber
2013
aeretodon (Abdala et al 2002) The orbit of FMNH PR 2444 isless upwardly directed than that of the holotype (being at anangle of sim65deg to a sagittal plane through the midline of the skullas opposed to sim45deg in the holotype) This difference in orbitalangle is probably attributable to postmortem deformation as theskull of FMNH PR 2444 is noticeably sheared to the front leftThe appearance of a wider squarer incisor arcade and broader
anterior palate in FMNH PR 2444 can also be attributed to thisshear Finally the occipital region of FMNH PR 2444 is alto-gether taller and narrower than that of the holotype For ex-ample the foramen magnum in FMNH PR 2444 approximates arounded isosceles triangle in shape while the foramen magnumof the holotype is more ovoid These differences can also beattributed to deformation Altogether the differences between
FIGURE 13 Stereopairs of field number 8-31-98-387 an isolated right pelvis in A medial and B lateral view Scale bars equal 5 cm
JOURNAL OF VERTEBRATE PALEONTOLOGY VOL 28 NO 2 2008458
Dow
nloa
ded
by [
UZ
H H
aupt
bibl
ioth
ek
Zen
tral
bibl
ioth
ek Z
uumlric
h] a
t 05
51 1
0 Se
ptem
ber
2013
FMNH PR 2444 and the holotype fall within the range of intra-specific variation recorded for other gomphodont cynodontssuch as the traversodontids Massetognathus pascuali (Abdalaand Giannini 2000 Kammerer pers obs) and Exaeretodon(Abdala et al 2002) and the diademodontid Diademodon tetra-gonus (Grine et al 1978 Bradu and Grine 1979)
Phylogenetic Analysis
Utilizing the new information afforded by FMNH PR 2444and a reexamination of the holotype (UA 10601) Menadon be-sairiei was recoded in the Gondwanan gomphodont data matrixof Abdala et al (2006) Abdala et alrsquos (2006) data matrix en-compassed 18 taxa and 43 characters For the present analysis anadditional character ldquoincisor procumbencyrdquo (states 0-absent1-present) was added to Abdala et alrsquos (2006) data set (see
Appendix for coding of this character) Several traversodontidtaxa (eg lsquoScalenodonrsquo hirschsoni) have relatively large incisorsbut only in Menadon Scalenodontoides and Exaeretodon are theincisors procumbent Three of Abdala et alrsquos (2006) originalcodings for Menadon besairiei characters 0 (ldquonumber of upperincisorsrdquo) 5 (ldquoposition of paracanine fossa in relation to theupper caninerdquo) and 21 (ldquointernarial barrdquo) were changed to in-corporate data from the new specimen (see Appendix) Addi-tionally further examination of the holotype (UA 10601) ofMenadon besairiei allowed us to code characters 16ndash18 20 38and 40 (morphology of the lower postcanines all marked as ldquordquoin Abdala et alrsquos analysis) Parsimony searches were performedin PAUP v40b10 using a branch-and-bound search Addition-ally a bootstrap analysis (also using branch-and-bound) was per-formed using 1000 replicates
A total of 34 most parsimonious trees of length 103 werefound with the scores CI0573 RI0725 RC0415 andHI0427 The strict consensus tree (Fig 15A) did not recovera polytomy between Menadon Exaeretodon and Scalenodont-oides as in Abdala and Ribeiro (2003) or a (Menadon + Ex-aeretodon) sister-group relationship as in Abdala et al (2006)Rather Menadon was recovered as the sister taxon of the (Ex-aeretodon + Scalenodontoides) clade supporting Flynn et alrsquos(2000) original placement of this taxon There is strong bootstrapsupport (88) for Traversodontidae but within traversodontidsthe only strongly supported subclade is (Gomphodontosuchus +(Menadon + (Scalenodontoides + Exaeretodon))) with a boot-strap value of 80 Within this clade the (Menadon + (Scalen-odontoides + Exaeretodon)) grouping is supported by a boot-strap value of 83 and the (Scalenodontoides + Exaeretodon)grouping is supported by a bootstrap value of 94 (Fig 15B) Inaddition to the positioning of Menadon the strict consensus tree(Fig 15A) differs from the topology of Abdala et al (2006) inrecovering a sister-group relationship between Massetognathusand Santacruzodon rather than having Massetognathus andSantacruzodon as successive outgroups to the clade containingGomphodontosuchus Menadon Scalenodontoides and Ex-aeretodon The (Massetognathus + Santacruzodon) clade is diag-nosed by the absence of an anterolingual cusp in the upper post-canines denticulated incisor cutting margins and the presence ofan anterior cingulum in the upper postcanines
FIGURE 14 Reconstruction of the pelvis of Menadon besairiei basedon information from FMNH PR 2444 and field number 8-31-98-387Dotted area not preserved in either specimen reconstruction based onPascualgnathus (Bonaparte 1966) and Exaeretodon (Bonaparte 1963)
FIGURE 15 Cladogram of traversodontid relationships based on revised data set of Abdala et al (2006) showing A strict consensus tree and B50 majority rule tree Numbers at nodes of majority rule tree represent bootstrap values
KAMMERER ET ALmdashCYNODONT FROM MADAGASCAR 459
Dow
nloa
ded
by [
UZ
H H
aupt
bibl
ioth
ek
Zen
tral
bibl
ioth
ek Z
uumlric
h] a
t 05
51 1
0 Se
ptem
ber
2013
Relationships of Menadon besairiei
Menadon besairiei is the sister taxon to (Exaeretodon + Sca-lenodontoides) the latter clade diagnosed by the possession ofthree upper incisors a posteromedial position of the paracaninefossae relative to the canines the absence of an internarial barlarge canines a projected zygomatic process of the jugal and amaxilla that contributes to the margin of the subtemporal fenes-tra (the last three character states represent reversals) The clade(Menadon + (Exaeretodon + Scalenodontoides)) is diagnosed bythe possession of large procumbent incisors The presence offour incisors a primitive retention distinguishes Menadon be-sairiei from Exaeretodon Scalenodontoides and an unnamed tra-versodontid (ldquoTraversodontid Type 1rdquo) from the Santa MariaFormation of Brazil (Abdala et al 2001 Abdala and Ribeiro2002) Gomphodontosuchus is recovered as the sister taxon to(Menadon (Exaeretodon + Scalenodontoides)) as has beenfound in previous studies (Flynn et al 2000 Abdala and Ribeiro2003) The taxon name Gomphodontosuchinae Watson andRomer 1956 (ex Gomphodontosuchidae Watson and Romer1956 first given subfamilial rank in Lehman 1961) is availablefor this clade As circumscribed here Gomphodontosuchinaeconsists of the most inclusive clade containing Gomphodontosu-chus brasiliensis von Huene 1928264 but not Massetognathuspascuali Romer 19672
Menadon besairiei displays a mixture of primitive and derivedcharacters within traversodontids The internarial bar and fourincisors are primitive features whereas the enlargement and pro-cumbency of the incisors are derived features shared with Ex-aeretodon and Scalenodontoides The postcranium shows severalsimilarities to that of Exaeretodon (apparent exclusion of theprocoracoid from the glenoid some features of the pelvis tallcaudal neural spines) but in general displays a rather conserva-tive morphology observed throughout Gomphodontia The mor-phology of the scapulocoracoids is closer akin to diademodontidsor trirachodontids than Exaeretodon and the expanded thoracicrib of Menadon has only slight differences from those of trirach-odontids or early traversodontids like Luangwa The atlas-axiscomplex of Menadon is similar to that of the Early Triassic cyn-odont Cynognathus in possessing a concave axial spine with along posterior process Although most gomphodonts have beenreconstructed with a convex axial spine as in early mammals thisspine is broken in most of the best described gomphodont post-crania (eg Diademodon Brink 1955) and the lsquosaddle-shapedrsquoaxial morphology may be more widely distributed among cyn-odonts than previously realized The new specimens of Menadondescribed in this paper demonstrate that a generalized gompho-dont postcranial morphology was retained deep within the tra-versodontid tree and that the derived morphologies of either ofthe well-known taxa Massetognathus or Exaeretodon should notbe taken as representative of traversodontids in general
ACKNOWLEDGMENTS
This work would not have been possible without the continu-ing support and encouragement of our collaborative programwith the Deacutepartement de Paleacuteontologie et drsquoAnthropologie Bi-ologique Universiteacute drsquoAntananarivo Madagascar under the di-rection of Armand Rasoamiaramanana (Department Chairman)and Gisegravele Ravololonarivo Randria (Head of the PaleontologyLaboratories) We are extremely grateful to the National Geo-graphic Society for their long-term support of the fieldwork as-sociated with this project Additional financial or logistical sup-port for fieldwork was provided by The Field Museum the Uni-versity of California-Santa Barbara and the WWF (WorldwideFund for Nature Madagascar) We acknowledge the Ministegraverede LrsquoEneacutergie et des Mines for authorizing field efforts and for
continuing to facilitate our joint US-Madagascar explorationresearch and education program Steve Goodman and AsminaGandhi generously assisted us in innumerable ways includinghospitality nonpareil We especially thank R Andriantompoha-vana L Razafimanantsoa A Toto Volahy JA Rabarison AGandie and P Vavisaro for assistance in the field during theexpedition in which these new specimens were discovered BillSimpson and Gina Wesley-Hunt for their patient excavation ofthis important material Allison Smith Ray Vodden Lisa Berg-wall and Deborah Wagner for excellent preparation of a chal-lenging specimen Jennifer Botha and Elize Butler (NationalMuseum Bloemfontein) Michael Raath and Fernando Abdala(Bernard Price Institute) Thomas Kemp (Oxford UniversityMuseum of Natural History) Ray Symonds (University Museumof Zoology Cambridge) Bernard Battail (Museacuteum nationaldrsquoHistoire naturelle Paris) Michael Maisch (Universitaumlt Tuumlbin-gen) and Charles Schaff and Farish Jenkins (Museum of Com-parative Zoology Harvard) for access to specimens and AgustiacutenMartinelli Fernando Abdala and James Hopson for helpful re-views of an earlier version of the manuscript
LITERATURE CITED
Abdala F M C Barbarena and J Dornelles 2002 A new species of thetraversodontid cynodont Exaeretodon from the Santa Maria Forma-tion (MiddleLate Triassic) of Southern Brazil Journal of Verte-brate Paleontology 22313ndash325
Abdala F and N P Giannini 2000 Gomphodont cynodonts of theChantildeares Formation the analysis of an ontogenetic sequence Jour-nal of Vertebrate Paleontology 20501ndash506
Abdala F J Neveling and J Welman 2006 A new trirachodontidcynodont from the lower levels of the Burgersdorp Formation(Lower Triassic) of the Beaufort Group South Africa and the cla-distic relationships of Gondwanan gomphodonts Zoological Jour-nal of the Linnean Society 147383ndash413
Abdala F A M Ribeiro and C L Schultz 2001 A rich cynodont faunaof Santa Cruz do Sul Santa Maria Formation (Middle-Late Trias-sic) southern Brazil Neues Jahrbuch fuumlr Geologie und Palaumlontolo-gie Monatshefte 2001669ndash687
Abdala F and A M Ribeiro 2002 Nuevos cinodontes traversodoacuten-tidos (Synapsida-Eucynodontia) de la Formacioacuten Santa Maria(Triaacutesico Medio-Superior) Rio Grande do Sul Brasil RevistaEspantildeola de Paleontologiacutea 17237ndash243
Abdala F and A M Ribeiro 2003 A new traversodontid cynodontfrom the Santa Maria Formation (Ladinian-Carnian) of southernBrazil with a phylogenetic analysis of Gondwanan traversodontidsZoological Journal of the Linnaean Society 139529ndash545
Abdala F and A M S Teixeira 2004 A traversodontid cynodont ofAfrican affinity in the South American Triassic Palaeontologia Af-ricana 4011ndash22
Bonaparte J F 1962 Descripcioacuten del craacuteneo y mandiacutebula de Ex-aeretodon frenguellii Cabrera y su comparacioacuten con Diademodon-tidae Tritylodontidae y los cinodontes sudamericanos Publica-ciones del Museo Municipal de Ciencias Naturales y Tradicional deMar del Plata 1135ndash202
Bonaparte J F 1963 Descripcioacuten del esqueleto postcraneano de Ex-aeretodon (CynodontiandashTraversodontidae) Acta Geoloacutegica Lil-loana 45ndash52
Bonaparte J F 1966 Una nueva ldquofaunardquo Triaacutesica de Argentina (Ther-apsida Cynodontia Dicynodontia) Consideracions filogeneacuteticas ypaleobiogeograacuteficas Ameghiniana 8243ndash296
Bradu D and F E Grine 1979 Multivariate analysis of diademodon-tine crania from South Africa and Zambia South African Journal ofScience 79441ndash448
Brink A S 1955 A study of the skeleton of Diademodon Palaeonto-logia Africana 33-39
Brink A S 1963 Two new cynodonts from the Ntawere Formation inthe Luangwa Valley of Northern Rhodesia Palaeontologia Africana877ndash96
Broom R 1905 On the use of the term Anomodontia Records of theAlbany Museum 1266ndash269
Cabrera A 1943 El primer hallazgo de teraacutepsidos en la ArgentinaNotas del Museo de La Plata Paleontologiacutea 55317ndash331
JOURNAL OF VERTEBRATE PALEONTOLOGY VOL 28 NO 2 2008460
Dow
nloa
ded
by [
UZ
H H
aupt
bibl
ioth
ek
Zen
tral
bibl
ioth
ek Z
uumlric
h] a
t 05
51 1
0 Se
ptem
ber
2013
Chatterjee S 1982 A new cynodont reptile from the Triassic of IndiaJournal of Paleontology 56203ndash214
Crompton A W 1955 On some Triassic cynodonts from TanganyikaProceedings of the Zoological Society of London 125617ndash669
Crompton A W 1972 Postcanine occlusion in cynodonts and tritylo-donts Bulletin of the British Museum (Natural History) Geology2129ndash71
Crompton A W and F Ellenberger 1957 On a new cynodont from theMolteno Beds and the origin of the tritylodontids Annals of theSouth African Museum 441ndash13
Flynn J J J M Parrish B Rakotosaminimanana W F Simpson R LWhatley and A R Wyss 1999 A Triassic fauna from Madagascarincluding early dinosaurs Science 286763ndash765
Flynn J J J M Parrish B Rakotosamimanana L RanivoharimananaW F Simpson and A R Wyss 2000 New traversodontids (Syn-apsida Eucynodontia) from the Triassic of Madagascar Journal ofVertebrate Paleontology 20422ndash427
Gow C E and P J Hancox 1993 First complete skull of the LateTriassic Scalenodontoides (Reptilia Cynodontia) from southern Af-rica In Lucas S G and M Morales (eds) The nonmarine TriassicNew Mexico Museum of Natural History amp Science Bulletin 3161ndash168
Grine F E B D Hahn and C E Gow 1978 Aspects of relativegrowth and variability in Diademodon (Reptilia Therapsida) SouthAfrican Journal of Science 7450ndash58
Hopson J A 1984 Late Triassic traversodont cynodonts from NovaScotia and southern Africa Palaeontologia Africana 25181ndash201
Hopson J A 1985 Morphology and relationships of Gomphodontosu-chus brasiliensis von Huene (Synapsida Cynodontia Tritylodontoi-dea) from the Triassic of Brazil Neues Jahrbuch fuumlr Geologie undPalaumlontologie Monatshefte 1985285ndash299
Hopson J A and J W Kitching 2001 A probainognathian cynodontfrom South Africa and the phylogeny of nonmammalian cynodontsBulletin of the Museum of Comparative Zoology 1565ndash35
Hopson J A and H-D Sues 2006 A traversodont cynodont from theMiddle Triassic (Ladinian) of Baden-Wuumlrttemberg (Germany)Palaumlontologische Zeitschrift 80124ndash129
Huene F von 1928 Ein Cynodontier aus der Trias Brasiliens Zentral-blatt fuumlr Mineralogie Geologie und Palaumlontologie 1928251ndash270
Huene F von 1936 Die fossilen Reptilien des suumldamerikanischenGondwanalandes Ergebnisse der Sauriergrabungen in Suumldbrasilien192829 Lieferung 2 (Cynodontia)93ndash159 Franz F Heine Tuumlbin-gen
Jenkins F A 1970 The Chantildeares (Argentina) Triassic reptile faunaVII The postcranial skeleton of the traversodontid Massetognathuspascuali (Therapsida Cynodontia) Breviora 3521ndash28
Jenkins F A 1971 The postcranial skeleton of African cynodonts prob-lems in the early evolution of the mammalian postcranial skeletonBulletin of the Peabody Museum of Natural History 361ndash216
Kemp T S 1969 The atlas-axis complex of mammal-like reptiles Jour-nal of Zoology 159223ndash248
Kemp T S 1980 Aspects of the structure and functional anatomy of theMiddle Triassic cynodont Luangwa Journal of Zoology 191193ndash239
Kemp T S 1982 Mammal-Like Reptiles and the Origin of MammalsAcademic Press New York
Lehman J-P 1961 Cynodontia pp 140ndash191 in Piveteau J (ed) Traiteacutede Paleacuteontologie Masson et Cie Paris
Osborn 1903 On the primary division of the Reptilia into two sub-classes Synapsida and Diapsida Science 17275ndash276
Owen R 1861 Palaeontology or a systematic summary of extinct ani-mals and their geological relations 2nd Ed Edinburgh
Parrington F R 1934 On the cynodont genus Galesaurus with a noteon the functional significance of the changes in the evolution of thetheriodont skull Annals and Magazine of Natural History 10(13)38ndash67
Parrington F R 1946 On the cranial anatomy of cynodonts Proceed-ings of the Zoological Society of London 116181ndash197
Razafimbelo E 1987 Le basin de Morondava (Madagascar) Synthegravesegeacuteologique et structurale PhD dissertation University Louis Pas-teur Strasbourg France 189 pp
Reisz R R and H-D Sues 2000 Herbivory in Late Paleozoic andTriassic terrestrial vertebrates pp 9ndash41 in Sues H-D (ed) Evo-lution of Herbivory in Terrestrial Vertebrates Cambridge Univer-sity Press New York
Romer A S 1967 The Chantildeares (Argentina) Triassic reptile fauna IIITwo new gomphodonts Massetognathus pascuali and M terugiiBreviora 2641ndash25
Romer A S 1972 The Chantildeares (Argentina) Triassic reptile faunaXVII The Chantildeares gomphodonts Breviora 3961ndash9
Rougier G W J R Wible and J A Hopson 1992 Reconstruction ofthe cranial vessels in the Early Cretaceous mammal Vincelestes neu-quenianus implications for the evolution of the mammalian cranialvascular system Journal of Vertebrate Paleontology 12188ndash216
Rowe T 1988 Definition diagnosis and origin of Mammalia Journal ofVertebrate Paleontology 8241ndash264
Rowe T 1993 Early mammal phylogenetic systematics pp 129ndash145 inSzalay F S M J Novacek and M C McKenna (eds) MammalPhylogeny Vol 1 Mesozoic Differentiation MultituberculatesMonotremes Early Therians and Marsupials Springer-Verlag NewYork
Seeley H G 1894 Researches on the structure organization and clas-sification of the fossil Reptilia Part IX Section 3 On DiademodonPhilosophical Transactions of the Royal Society of London 1851029ndash1041
Seeley H G 1895 Researches on the structure organization and clas-sification of the fossil Reptilia Part IX Section 4 On the Gompho-dontia Philosophical Transactions of the Royal Society of London1861ndash57
Seeley H G 1895 Researches on the structure organization and clas-sification of the fossil Reptilia Part IX Section 5 On the skeletonin new Cynodontia from the Karroo rocks Philosophical Transac-tions of the Royal Society of London 18659ndash148
Sues H-D J A Hopson and N H Shubin 1992 Affinities of Sca-lenodontoides plemmyridon Hopson 1984 (Synapsida Cynodontia)from the Upper Triassic of Nova Scotia Journal of Vertebrate Pa-leontology 12168ndash171
Sues H-D and P E Olsen 1990 Triassic vertebrates of Gondwananaspect from the Richmond basin of Virginia Science 2491020ndash1023
Swofford D L 2003 PAUP v 40b10 Sinauer Associates Inc Pub-lishers Sunderland Massachusetts
Watson D M S and A S Romer 1956 A classification of therapsidreptiles Bulletin of the Museum of Comparative Zoology 11435ndash89
Submitted September 20 2007 accepted November 3 2007
KAMMERER ET ALmdashCYNODONT FROM MADAGASCAR 461
Dow
nloa
ded
by [
UZ
H H
aupt
bibl
ioth
ek
Zen
tral
bibl
ioth
ek Z
uumlric
h] a
t 05
51 1
0 Se
ptem
ber
2013
APPENDIX Data matrix used in the phylogenetic analysesThis is a revised version of the matrix of Abdala et al (2006) with the following changes character 43 [Incisor procumbency (0) absent (1) present]added Menadon recoded for characters 0 (Number of upper incisors was 1 ldquothreerdquo changed to 0 ldquofourrdquo) 5 (Position of paracanine fossae in relationto the upper canine was changed to 1 ldquomedialrdquo) 16 (Overall morphology of lower postcanines in occlusal view was changed to 3 ldquoquadran-gularrdquo) 17 (Position of the transverse crest in lower postcanines was changed to 1 ldquoanteriorrdquo) 18 (Number of cusps in the transverse crest of thelower postcanines was changed to 0 ldquotwordquo) 20 (Anterior cingulum in the lower postcanines was changed to 2 ldquoabsentrdquo) 21 (Internarial barwas 1 ldquoabsentrdquo changed to 0 ldquopresentrdquo) 38 (Posterior cingulum in the lower postcanines was changed to 1 ldquoabsentrdquo) and 40 (Deep occlusalbasins in the postcanines was changed to 1 ldquopresentrdquo) Langbergia recoded for character 34 (Incisor cutting margins was 0 ldquoserratedrdquo changedto 1 ldquosmoothly ridgedrdquo) based on the description of the incisors in Abdala et al (2006) Gomphodontosuchus recoded for character 31 (Maxilla inthe margin of the subtemporal fenestra was 1 ldquoincludedrdquo changed to 0 ldquoexcludedrdquo) based on von Huene (1936) and Kammerer (pers obs of theholotype) contra Hopsonrsquos (1985) reconstruction lsquoScalenodonrsquo hirschsoni recoded for character 39 (Anterior cingulum in the upper postcanines was1 ldquoabsentrdquo changed to 0 ldquopresentrdquo) based on Abdala (pers comm 2006) Trirachodon recoded for character 30 (Ectopterygoid was changedto 0 ldquopresentrdquo) based on Abdala (pers comm 2006) and Traversodon recoded for character 15 (Number of cusps in the sectorial border of the upperpostcanines was 0 ldquomore than twordquo changed to 1 ldquotwordquo) based on Abdala (pers comm 2006) and Kammerer (pers obs)
1111111111 2222222222 3333333333 44440123456789 0123456789 0123456789 0123456789 0123
Cynognathus 00000000 000 000000000 00000000 000Diademodon 0000001000 0000100 000010A000 0000000100 0100Trirachodon 000000100B 1A002201 001A100010 0001001200 0110Langbergia 0000001000 10000201 0000101010 0001101200 0110Cricodon 0100001000 11000201 0010100 0001200 0110Andescynodon 0000002001 01013101 101120010 0112311 1100Massetognathus 00111121A2 1110003100 2011200011 1011212300 1100Exaeretodon 1110122112 010103101 2111110201 1111112311 1101Luangwa 0000001002 11010A3100 101010010 01002300 1100Scal angustifrons 000000100B 1111023100 10102101 00002301 1100Scal hirschsoni 11110202 1100113100 1010 1112300 1100Traversodon 0000002012 1101113100 201010 011230 1100Gomphodontosuchus 0011112112 0103101 2010 0112311 1100Pascualgnathus 1000001000 000231 1100001 0011230 1100Scalenodontoides 1112212 01003101 21111021 1212311 1101Menadon 0111112112 01012310 20121020 0102311 1101Dadadon 00002112 111112 013 11231 1100Santacruzodon 00002112 1110003101 201300 212300 1100
A(01) B(02)
JOURNAL OF VERTEBRATE PALEONTOLOGY VOL 28 NO 2 2008462
Dow
nloa
ded
by [
UZ
H H
aupt
bibl
ioth
ek
Zen
tral
bibl
ioth
ek Z
uumlric
h] a
t 05
51 1
0 Se
ptem
ber
2013

terior costal plate and has an anteroposteriorly shorter anteriorcostal plate Additionally the posterior margin of the costal plateis less curved than that of Luangwa These differences couldindicate that the expanded rib of Menadon is from a more pos-terior position in the series than the rib from Luangwa as re-duced curvature and increasing confluence of the tuberculumand capitulum are characteristic of lumbar ribs in traversodon-tids (Jenkins 1970 Kemp 1980) However the length of theshaft is at odds with this interpretation and in Cricodon andPascualgnathus the more posterior ribs are not more similar tothe element from FMNH PR 2444 These differences probablyrepresent legitimate morphological distinctions between the ribsof earlier gomphodonts and Menadon The head of the expandedrib is well defined and unbroken indicating that it was not syn-ostosed to a vertebra
Appendicular Skeleton
The appendicular skeleton is represented by both scapulocora-coids and a fragment of a clavicle from field number 9-4-98-440and a partial right pelvis from field number 9-2-98-422 An ad-ditional partial right pelvis is represented from field number8-31-98-387 The scapulocoracoids (Figs 10 11) are very similarto those described for other traversodontids as well as cynog-nathids and diademodontids (Jenkins 1970 1971) The scapularinfraspinous fossa has out-turned anterior and posterior edgesThe anterior edge of the scapula is rounded and curves laterallyforming the scapular spine At the lower end of the spine theacromion forms a prominent process (broken in the left scapula)Below this process the scapula is constricted and then expandsventrally to form the dorsal margin of the articular glenoid Theglenoid fossa is in general morphology very similar to that ofother Triassic cynodonts (Jenkins 1970 1971 Kemp 1980) It isposteroventrally oriented with roughly equal contributions fromthe scapular and coracoid glenoid facets The coracoid glenoidfacet is roughly triangular while the scapular glenoid facet issemiovoid as in other cynodonts In anterior or posterior viewthe fossa appears smoothly crescentic The procoracoid is aslightly curved trapezoidal plate of bone It does not appear tocontribute to the glenoid fossa The procoracoid foramen is lo-cated midway up the height of the procoracoid at a point directlybelow the base of the acromion process The coracoid is semi-crescentic curving posteriorly from a broad base The coracoidin Menadon is longer and more falcate than those described forExaeretodon (Bonaparte 1963) and Luangwa (Kemp 1980 al-though the distal tip of both coracoids is broken in the specimenof Luangwa [TSK 121] described by Kemp and may have beenlonger than reconstructed) but agrees closely with those of Mas-setognathus and cynognathidsdiademodontids (Jenkins 19701971) The distal coracoid tubercle for attachment to the tricepsis strongly curved upwards similar to the condition in Masseto-gnathus (Jenkins 1970) and Diademodon (Brink 1955)
A small fragment of bone resembling a curved cylinder almostcertainly represents part of a clavicle It has no distinguishingfeatures other than a groove associated with a ridge on one sideHowever the degree of curvature observed precludes this ele-ment from representing a long bone and the circular cross-section excludes it from being a rib
The pelvis of FMNH PR 2444 (Fig 12) resembles those ofother traversodontids in general morphology The ilium bears anarrow anteriorly directed iliac blade curving outward distallyThe anterior edge of the iliac blade slopes upward at a roughly45deg angle similar to Luangwa (Kemp 1980) but unlike Ex-aeretodon in which the anterior edge of the iliac blade curvesbackwards (Bonaparte 1963) or Pascualgnathus and Masseto-gnathus in which it is rounded (Bonaparte 1966 Jenkins 1970)The supraacetabular buttress at the base of the ilium is located inan anterodorsal position relative to the acetabulum as in other
traversodontids but unlike earlier cynodonts in which it is lo-cated dorsally (Jenkins 1970) The acetabular buttresses of thethree pelvic bones are largely confluent forming a fairly uniformlsquoliprsquo around the acetabulum as in Exaeretodon (Bonaparte1963) On the contrary in Luangwa the supraacetabular ischialand pubic buttresses are highly distinct spaced 6ndash8 mm apart andwithout confluence of their acetabular facets The pelvis ofFMNH PR 2444 does have a small depression between the su-praacetabular and ischial buttresses (also present in Ex-aeretodon Bonaparte 1963) but this is not as developed as inLuangwa The posterior extent of the iliac blade is broken inFMNH PR 2444 but this region is preserved in another isolatedright pelvis field number 8-31-98-387 (Fig 13) probably pertain-ing to Menadon besairiei The posterior process of the iliac bladein this specimen is relatively short and directed away from thedorsal edge of the ischium as in Exaeretodon (Bonaparte 1963)but unlike Massetognathus and Luangwa in which the posteriorprocess is long and extends roughly horizontally over the dorsaledge of the ischium (Jenkins 1970 Kemp 1980) and evencurves downward distally in Luangwa The posterior process ofthe iliac blade in Pascualgnathus is directed away from the is-chium (Bonaparte 1966) as in Exaeretodon and Menadon but isrelatively long as in Massetognathus and Luangwa
The proximal portion of the ischium is broad and dominatedby the ischial buttress The ischium constricts posterior to theproximal portion and expands distally into a spatulate platewhich is thickened at its posterior margin This thickening con-tinues along the dorsolateral surface of the ischium forming aridge that slopes ventrally until reaching the edge of the ischialbuttress The downward slope of this ridge produces a prominentgroove at the dorsal edge of the lateral surface of the ischiumthought to represent the origin of the ischio-trochantericusmuscle (Kemp 1980) The constriction of the ischium in Mena-don is like that of Luangwa albeit even more extreme in itsventral concavity but unlike that of Exaeretodon in which themedial and distal ends of the ischium are broadly united Theischium in general is a proportionally shorter element in Ex-aeretodon than in other traversodontids including Menadon(Bonaparte 1963) The anteroventral portion of the ischium thatconnects to the pubis is not preserved in either of the referredMenadon pelves
The pubis is the most incompletely preserved pelvic elementThe acetabular contribution of the pubis is smaller than that ofthe ischium or ilium Distal to the acetabulum the pubis is con-stricted indicating that the connection between the acetabularregion and the pubic plate was considerably narrower than inExaeretodon (Bonaparte 1963) but close akin to the pubis ofLuangwa (Kemp 1980) The pubic plate itself is not preservedThe ventral outline of the ischium and what is known of thepubis indicate that the obturator foramen was relatively larger inMenadon (Fig 14) than in Exaeretodon (Bonaparte 1963)
DISCUSSION
Comparison with the Holotype of Menadon besairiei
FMNH PR 2444 differs in several significant ways from theholotype of Menadon besairiei as previously described most no-tably in the possession of an internarial bar (initially consideredabsent in M besairiei) and four upper incisors (reported as threein M besairiei but with the caveat that the homologies of thefourth and fifth anterior tooth loci were problematic Flynn et al2000) However upon further study we interpret the lack of acontinuous internarial bar in the holotype as breakage ratherthan the natural condition of the skull (a prominent medial pro-tuberance is present on the ventral rim of the nares in UA10601) Regarding the incisor count it should be noted that theholotype of Menadon besairiei displays an unusual set of anterior
JOURNAL OF VERTEBRATE PALEONTOLOGY VOL 28 NO 2 2008454
Dow
nloa
ded
by [
UZ
H H
aupt
bibl
ioth
ek
Zen
tral
bibl
ioth
ek Z
uumlric
h] a
t 05
51 1
0 Se
ptem
ber
2013
FIGURE 10 Right scapulocoracoids of FMNH PR 2444 A stereopair of right scapulocoracoid in medial view B stereopair and interpretivedrawing of right scapulocoracoid in lateral view Scale bars equal 5 cm Abbreviationsmdashac acromion process C coracoid c gl coracoid contributionto the glenoid fossa c tr tubercle for coracoid attachment of triceps PC procoracoid pc f procoracoid foramen S scapula s gl scapularcontribution to the glenoid fossa
KAMMERER ET ALmdashCYNODONT FROM MADAGASCAR 455
Dow
nloa
ded
by [
UZ
H H
aupt
bibl
ioth
ek
Zen
tral
bibl
ioth
ek Z
uumlric
h] a
t 05
51 1
0 Se
ptem
ber
2013
teeth for a traversodontid As mentioned above Flynn et al(2000) noted that the homologies of these teeth were difficult todetermine Of the five anterior teeth preserved in the holotypethe anteriormost three are ldquoundoubtedly incisorsrdquo although I3 isldquounusually large and is caniniformrdquo (Flynn et al 2000423) Therobust recurved fourth tooth in the series was considered torepresent the canine with the subsequent tooth interpreted as anolder canine in the process of replacement Although FMNH PR2444 is edentulous the alveoli on the left side of the skull are wellpreserved and indicate an anterior tooth set identical to the ho-lotype The alveolar morphology indicates that the fifth toothwas canted anteriorly as in the holotype This suggests that themysterious fifth tooth is not the result of a transient period in thedental replacement history of M besairiei as suggested by Flynnet al (2000) but rather is the norm This leaves two possibilities1) M besairiei persistently had two canines This condition isunknown in cynodonts 2) The canine-like fourth tooth actuallyrepresents an incisor and the fifth tooth is the true canine Wefavor the latter explanation In FMNH PR 2444 the first fouralveoli are nearly identical in size and shape However the fifthalveolus while similar in anteroposterior length to its predeces-sors is strongly compressed labiolingually with an anteroposte-rior lengthlabiolingual width ratio of 044 (compared to 080 inthe fourth alveolus and 084ndash85 for I2ndash3) We interpret thismarked difference in alveolar ratio as indicating the separationbetween incisors and canine The canine-like I4 would then rep-resent the endpoint in a series of large increasingly caniniformincisors Supporting this interpretation recurved incisors are alsoobserved in juvenile Exaeretodon (eg MCP 3843 PV see Ab-dala et al 2002)
There are several legitimate differences between FMNH PR2444 and the holotype of Menadon besairiei but we take a con-servative taxonomic stance and argue that these differences rep-resent intraspecific variation and preservational artifacts FMNHPR 2444 represents a larger animal than UA 10601 The snoutlength (measured as the distance between the anterior edge ofthe orbit and a vertical plane through the anterior tip of thepremaxilla) of FMNH PR 2444 is 911 mm while that of UA10601 is 801 mm As such proportional differences between thetwo skulls may be attributable to ontogeny Variations in FMNHPR 2444 compared to the holotype that probably represent on-togentic differences include wider longer and deeper paraca-nine fossae a much deeper ventral zygomatic flange (althoughthe lower border of the flange is slightly abraded in the holotypeof Menadon besairiei there is no evidence that a major flangewas broken off) a proportionally shorter and more circular out-line to the posterior border of the maxilla below the zygomaticprocess and a more deeply emarginated posterior squamosal(ie a more prominent laterally projecting flange)
FMNH PR 2444 has a proportionally shorter more robustsnout and larger orbits than the holotype While these charactersare certainly variable in cynodonts it is unusual that the largerspecimen is shorter-snouted and larger-eyed contra the usualpattern The proportional differences between the two skulls areminor however (orbit diametersnout length is 042 for FMNHPR 2444 and 041 for the holotype) and are partially attributableto deformation (shear in FMNH PR 2444 and breakage of thepostorbital bar in the holotype) The following differences canalso probably be attributed to preservational artifact only fivepostcanine alveoli are preserved in FMNH PR 2444 while theholotype has eight postcanines The anteriormost three in theholotype however are tiny and peg-like As the area of themaxilla between the canine alveolus and the first preserved post-canine alveolus is poorly preserved in FMNH PR 2444 the ab-sence of these postcanines may be artifactual However it is alsopossible that the lower postcanine count of FMNH PR 2444compared to the holotype is an ontogenetic effect as postcaninecount is known to decrease with increasing skull size in Ex-
FIGURE 11 Left scapulocoracoids of FMNH PR 2444 A left scapu-locoracoid in medial view B left scapulocoracoid in lateral view Scalebars equal 5 cm
JOURNAL OF VERTEBRATE PALEONTOLOGY VOL 28 NO 2 2008456
Dow
nloa
ded
by [
UZ
H H
aupt
bibl
ioth
ek
Zen
tral
bibl
ioth
ek Z
uumlric
h] a
t 05
51 1
0 Se
ptem
ber
2013
FIGURE 12 Right pelvis of FMNH PR 2444 A stereopair of right pelvis in medial view B stereopair and interpretive drawing of right pelvis inlateral view Scale bars equal 5 cm Abbreviationsmdashbpp base of posterior process of ilium ib iliac blade IL ilium IS ischium ob f obturatorforamen P pubis
KAMMERER ET ALmdashCYNODONT FROM MADAGASCAR 457
Dow
nloa
ded
by [
UZ
H H
aupt
bibl
ioth
ek
Zen
tral
bibl
ioth
ek Z
uumlric
h] a
t 05
51 1
0 Se
ptem
ber
2013
aeretodon (Abdala et al 2002) The orbit of FMNH PR 2444 isless upwardly directed than that of the holotype (being at anangle of sim65deg to a sagittal plane through the midline of the skullas opposed to sim45deg in the holotype) This difference in orbitalangle is probably attributable to postmortem deformation as theskull of FMNH PR 2444 is noticeably sheared to the front leftThe appearance of a wider squarer incisor arcade and broader
anterior palate in FMNH PR 2444 can also be attributed to thisshear Finally the occipital region of FMNH PR 2444 is alto-gether taller and narrower than that of the holotype For ex-ample the foramen magnum in FMNH PR 2444 approximates arounded isosceles triangle in shape while the foramen magnumof the holotype is more ovoid These differences can also beattributed to deformation Altogether the differences between
FIGURE 13 Stereopairs of field number 8-31-98-387 an isolated right pelvis in A medial and B lateral view Scale bars equal 5 cm
JOURNAL OF VERTEBRATE PALEONTOLOGY VOL 28 NO 2 2008458
Dow
nloa
ded
by [
UZ
H H
aupt
bibl
ioth
ek
Zen
tral
bibl
ioth
ek Z
uumlric
h] a
t 05
51 1
0 Se
ptem
ber
2013
FMNH PR 2444 and the holotype fall within the range of intra-specific variation recorded for other gomphodont cynodontssuch as the traversodontids Massetognathus pascuali (Abdalaand Giannini 2000 Kammerer pers obs) and Exaeretodon(Abdala et al 2002) and the diademodontid Diademodon tetra-gonus (Grine et al 1978 Bradu and Grine 1979)
Phylogenetic Analysis
Utilizing the new information afforded by FMNH PR 2444and a reexamination of the holotype (UA 10601) Menadon be-sairiei was recoded in the Gondwanan gomphodont data matrixof Abdala et al (2006) Abdala et alrsquos (2006) data matrix en-compassed 18 taxa and 43 characters For the present analysis anadditional character ldquoincisor procumbencyrdquo (states 0-absent1-present) was added to Abdala et alrsquos (2006) data set (see
Appendix for coding of this character) Several traversodontidtaxa (eg lsquoScalenodonrsquo hirschsoni) have relatively large incisorsbut only in Menadon Scalenodontoides and Exaeretodon are theincisors procumbent Three of Abdala et alrsquos (2006) originalcodings for Menadon besairiei characters 0 (ldquonumber of upperincisorsrdquo) 5 (ldquoposition of paracanine fossa in relation to theupper caninerdquo) and 21 (ldquointernarial barrdquo) were changed to in-corporate data from the new specimen (see Appendix) Addi-tionally further examination of the holotype (UA 10601) ofMenadon besairiei allowed us to code characters 16ndash18 20 38and 40 (morphology of the lower postcanines all marked as ldquordquoin Abdala et alrsquos analysis) Parsimony searches were performedin PAUP v40b10 using a branch-and-bound search Addition-ally a bootstrap analysis (also using branch-and-bound) was per-formed using 1000 replicates
A total of 34 most parsimonious trees of length 103 werefound with the scores CI0573 RI0725 RC0415 andHI0427 The strict consensus tree (Fig 15A) did not recovera polytomy between Menadon Exaeretodon and Scalenodont-oides as in Abdala and Ribeiro (2003) or a (Menadon + Ex-aeretodon) sister-group relationship as in Abdala et al (2006)Rather Menadon was recovered as the sister taxon of the (Ex-aeretodon + Scalenodontoides) clade supporting Flynn et alrsquos(2000) original placement of this taxon There is strong bootstrapsupport (88) for Traversodontidae but within traversodontidsthe only strongly supported subclade is (Gomphodontosuchus +(Menadon + (Scalenodontoides + Exaeretodon))) with a boot-strap value of 80 Within this clade the (Menadon + (Scalen-odontoides + Exaeretodon)) grouping is supported by a boot-strap value of 83 and the (Scalenodontoides + Exaeretodon)grouping is supported by a bootstrap value of 94 (Fig 15B) Inaddition to the positioning of Menadon the strict consensus tree(Fig 15A) differs from the topology of Abdala et al (2006) inrecovering a sister-group relationship between Massetognathusand Santacruzodon rather than having Massetognathus andSantacruzodon as successive outgroups to the clade containingGomphodontosuchus Menadon Scalenodontoides and Ex-aeretodon The (Massetognathus + Santacruzodon) clade is diag-nosed by the absence of an anterolingual cusp in the upper post-canines denticulated incisor cutting margins and the presence ofan anterior cingulum in the upper postcanines
FIGURE 14 Reconstruction of the pelvis of Menadon besairiei basedon information from FMNH PR 2444 and field number 8-31-98-387Dotted area not preserved in either specimen reconstruction based onPascualgnathus (Bonaparte 1966) and Exaeretodon (Bonaparte 1963)
FIGURE 15 Cladogram of traversodontid relationships based on revised data set of Abdala et al (2006) showing A strict consensus tree and B50 majority rule tree Numbers at nodes of majority rule tree represent bootstrap values
KAMMERER ET ALmdashCYNODONT FROM MADAGASCAR 459
Dow
nloa
ded
by [
UZ
H H
aupt
bibl
ioth
ek
Zen
tral
bibl
ioth
ek Z
uumlric
h] a
t 05
51 1
0 Se
ptem
ber
2013
Relationships of Menadon besairiei
Menadon besairiei is the sister taxon to (Exaeretodon + Sca-lenodontoides) the latter clade diagnosed by the possession ofthree upper incisors a posteromedial position of the paracaninefossae relative to the canines the absence of an internarial barlarge canines a projected zygomatic process of the jugal and amaxilla that contributes to the margin of the subtemporal fenes-tra (the last three character states represent reversals) The clade(Menadon + (Exaeretodon + Scalenodontoides)) is diagnosed bythe possession of large procumbent incisors The presence offour incisors a primitive retention distinguishes Menadon be-sairiei from Exaeretodon Scalenodontoides and an unnamed tra-versodontid (ldquoTraversodontid Type 1rdquo) from the Santa MariaFormation of Brazil (Abdala et al 2001 Abdala and Ribeiro2002) Gomphodontosuchus is recovered as the sister taxon to(Menadon (Exaeretodon + Scalenodontoides)) as has beenfound in previous studies (Flynn et al 2000 Abdala and Ribeiro2003) The taxon name Gomphodontosuchinae Watson andRomer 1956 (ex Gomphodontosuchidae Watson and Romer1956 first given subfamilial rank in Lehman 1961) is availablefor this clade As circumscribed here Gomphodontosuchinaeconsists of the most inclusive clade containing Gomphodontosu-chus brasiliensis von Huene 1928264 but not Massetognathuspascuali Romer 19672
Menadon besairiei displays a mixture of primitive and derivedcharacters within traversodontids The internarial bar and fourincisors are primitive features whereas the enlargement and pro-cumbency of the incisors are derived features shared with Ex-aeretodon and Scalenodontoides The postcranium shows severalsimilarities to that of Exaeretodon (apparent exclusion of theprocoracoid from the glenoid some features of the pelvis tallcaudal neural spines) but in general displays a rather conserva-tive morphology observed throughout Gomphodontia The mor-phology of the scapulocoracoids is closer akin to diademodontidsor trirachodontids than Exaeretodon and the expanded thoracicrib of Menadon has only slight differences from those of trirach-odontids or early traversodontids like Luangwa The atlas-axiscomplex of Menadon is similar to that of the Early Triassic cyn-odont Cynognathus in possessing a concave axial spine with along posterior process Although most gomphodonts have beenreconstructed with a convex axial spine as in early mammals thisspine is broken in most of the best described gomphodont post-crania (eg Diademodon Brink 1955) and the lsquosaddle-shapedrsquoaxial morphology may be more widely distributed among cyn-odonts than previously realized The new specimens of Menadondescribed in this paper demonstrate that a generalized gompho-dont postcranial morphology was retained deep within the tra-versodontid tree and that the derived morphologies of either ofthe well-known taxa Massetognathus or Exaeretodon should notbe taken as representative of traversodontids in general
ACKNOWLEDGMENTS
This work would not have been possible without the continu-ing support and encouragement of our collaborative programwith the Deacutepartement de Paleacuteontologie et drsquoAnthropologie Bi-ologique Universiteacute drsquoAntananarivo Madagascar under the di-rection of Armand Rasoamiaramanana (Department Chairman)and Gisegravele Ravololonarivo Randria (Head of the PaleontologyLaboratories) We are extremely grateful to the National Geo-graphic Society for their long-term support of the fieldwork as-sociated with this project Additional financial or logistical sup-port for fieldwork was provided by The Field Museum the Uni-versity of California-Santa Barbara and the WWF (WorldwideFund for Nature Madagascar) We acknowledge the Ministegraverede LrsquoEneacutergie et des Mines for authorizing field efforts and for
continuing to facilitate our joint US-Madagascar explorationresearch and education program Steve Goodman and AsminaGandhi generously assisted us in innumerable ways includinghospitality nonpareil We especially thank R Andriantompoha-vana L Razafimanantsoa A Toto Volahy JA Rabarison AGandie and P Vavisaro for assistance in the field during theexpedition in which these new specimens were discovered BillSimpson and Gina Wesley-Hunt for their patient excavation ofthis important material Allison Smith Ray Vodden Lisa Berg-wall and Deborah Wagner for excellent preparation of a chal-lenging specimen Jennifer Botha and Elize Butler (NationalMuseum Bloemfontein) Michael Raath and Fernando Abdala(Bernard Price Institute) Thomas Kemp (Oxford UniversityMuseum of Natural History) Ray Symonds (University Museumof Zoology Cambridge) Bernard Battail (Museacuteum nationaldrsquoHistoire naturelle Paris) Michael Maisch (Universitaumlt Tuumlbin-gen) and Charles Schaff and Farish Jenkins (Museum of Com-parative Zoology Harvard) for access to specimens and AgustiacutenMartinelli Fernando Abdala and James Hopson for helpful re-views of an earlier version of the manuscript
LITERATURE CITED
Abdala F M C Barbarena and J Dornelles 2002 A new species of thetraversodontid cynodont Exaeretodon from the Santa Maria Forma-tion (MiddleLate Triassic) of Southern Brazil Journal of Verte-brate Paleontology 22313ndash325
Abdala F and N P Giannini 2000 Gomphodont cynodonts of theChantildeares Formation the analysis of an ontogenetic sequence Jour-nal of Vertebrate Paleontology 20501ndash506
Abdala F J Neveling and J Welman 2006 A new trirachodontidcynodont from the lower levels of the Burgersdorp Formation(Lower Triassic) of the Beaufort Group South Africa and the cla-distic relationships of Gondwanan gomphodonts Zoological Jour-nal of the Linnean Society 147383ndash413
Abdala F A M Ribeiro and C L Schultz 2001 A rich cynodont faunaof Santa Cruz do Sul Santa Maria Formation (Middle-Late Trias-sic) southern Brazil Neues Jahrbuch fuumlr Geologie und Palaumlontolo-gie Monatshefte 2001669ndash687
Abdala F and A M Ribeiro 2002 Nuevos cinodontes traversodoacuten-tidos (Synapsida-Eucynodontia) de la Formacioacuten Santa Maria(Triaacutesico Medio-Superior) Rio Grande do Sul Brasil RevistaEspantildeola de Paleontologiacutea 17237ndash243
Abdala F and A M Ribeiro 2003 A new traversodontid cynodontfrom the Santa Maria Formation (Ladinian-Carnian) of southernBrazil with a phylogenetic analysis of Gondwanan traversodontidsZoological Journal of the Linnaean Society 139529ndash545
Abdala F and A M S Teixeira 2004 A traversodontid cynodont ofAfrican affinity in the South American Triassic Palaeontologia Af-ricana 4011ndash22
Bonaparte J F 1962 Descripcioacuten del craacuteneo y mandiacutebula de Ex-aeretodon frenguellii Cabrera y su comparacioacuten con Diademodon-tidae Tritylodontidae y los cinodontes sudamericanos Publica-ciones del Museo Municipal de Ciencias Naturales y Tradicional deMar del Plata 1135ndash202
Bonaparte J F 1963 Descripcioacuten del esqueleto postcraneano de Ex-aeretodon (CynodontiandashTraversodontidae) Acta Geoloacutegica Lil-loana 45ndash52
Bonaparte J F 1966 Una nueva ldquofaunardquo Triaacutesica de Argentina (Ther-apsida Cynodontia Dicynodontia) Consideracions filogeneacuteticas ypaleobiogeograacuteficas Ameghiniana 8243ndash296
Bradu D and F E Grine 1979 Multivariate analysis of diademodon-tine crania from South Africa and Zambia South African Journal ofScience 79441ndash448
Brink A S 1955 A study of the skeleton of Diademodon Palaeonto-logia Africana 33-39
Brink A S 1963 Two new cynodonts from the Ntawere Formation inthe Luangwa Valley of Northern Rhodesia Palaeontologia Africana877ndash96
Broom R 1905 On the use of the term Anomodontia Records of theAlbany Museum 1266ndash269
Cabrera A 1943 El primer hallazgo de teraacutepsidos en la ArgentinaNotas del Museo de La Plata Paleontologiacutea 55317ndash331
JOURNAL OF VERTEBRATE PALEONTOLOGY VOL 28 NO 2 2008460
Dow
nloa
ded
by [
UZ
H H
aupt
bibl
ioth
ek
Zen
tral
bibl
ioth
ek Z
uumlric
h] a
t 05
51 1
0 Se
ptem
ber
2013
Chatterjee S 1982 A new cynodont reptile from the Triassic of IndiaJournal of Paleontology 56203ndash214
Crompton A W 1955 On some Triassic cynodonts from TanganyikaProceedings of the Zoological Society of London 125617ndash669
Crompton A W 1972 Postcanine occlusion in cynodonts and tritylo-donts Bulletin of the British Museum (Natural History) Geology2129ndash71
Crompton A W and F Ellenberger 1957 On a new cynodont from theMolteno Beds and the origin of the tritylodontids Annals of theSouth African Museum 441ndash13
Flynn J J J M Parrish B Rakotosaminimanana W F Simpson R LWhatley and A R Wyss 1999 A Triassic fauna from Madagascarincluding early dinosaurs Science 286763ndash765
Flynn J J J M Parrish B Rakotosamimanana L RanivoharimananaW F Simpson and A R Wyss 2000 New traversodontids (Syn-apsida Eucynodontia) from the Triassic of Madagascar Journal ofVertebrate Paleontology 20422ndash427
Gow C E and P J Hancox 1993 First complete skull of the LateTriassic Scalenodontoides (Reptilia Cynodontia) from southern Af-rica In Lucas S G and M Morales (eds) The nonmarine TriassicNew Mexico Museum of Natural History amp Science Bulletin 3161ndash168
Grine F E B D Hahn and C E Gow 1978 Aspects of relativegrowth and variability in Diademodon (Reptilia Therapsida) SouthAfrican Journal of Science 7450ndash58
Hopson J A 1984 Late Triassic traversodont cynodonts from NovaScotia and southern Africa Palaeontologia Africana 25181ndash201
Hopson J A 1985 Morphology and relationships of Gomphodontosu-chus brasiliensis von Huene (Synapsida Cynodontia Tritylodontoi-dea) from the Triassic of Brazil Neues Jahrbuch fuumlr Geologie undPalaumlontologie Monatshefte 1985285ndash299
Hopson J A and J W Kitching 2001 A probainognathian cynodontfrom South Africa and the phylogeny of nonmammalian cynodontsBulletin of the Museum of Comparative Zoology 1565ndash35
Hopson J A and H-D Sues 2006 A traversodont cynodont from theMiddle Triassic (Ladinian) of Baden-Wuumlrttemberg (Germany)Palaumlontologische Zeitschrift 80124ndash129
Huene F von 1928 Ein Cynodontier aus der Trias Brasiliens Zentral-blatt fuumlr Mineralogie Geologie und Palaumlontologie 1928251ndash270
Huene F von 1936 Die fossilen Reptilien des suumldamerikanischenGondwanalandes Ergebnisse der Sauriergrabungen in Suumldbrasilien192829 Lieferung 2 (Cynodontia)93ndash159 Franz F Heine Tuumlbin-gen
Jenkins F A 1970 The Chantildeares (Argentina) Triassic reptile faunaVII The postcranial skeleton of the traversodontid Massetognathuspascuali (Therapsida Cynodontia) Breviora 3521ndash28
Jenkins F A 1971 The postcranial skeleton of African cynodonts prob-lems in the early evolution of the mammalian postcranial skeletonBulletin of the Peabody Museum of Natural History 361ndash216
Kemp T S 1969 The atlas-axis complex of mammal-like reptiles Jour-nal of Zoology 159223ndash248
Kemp T S 1980 Aspects of the structure and functional anatomy of theMiddle Triassic cynodont Luangwa Journal of Zoology 191193ndash239
Kemp T S 1982 Mammal-Like Reptiles and the Origin of MammalsAcademic Press New York
Lehman J-P 1961 Cynodontia pp 140ndash191 in Piveteau J (ed) Traiteacutede Paleacuteontologie Masson et Cie Paris
Osborn 1903 On the primary division of the Reptilia into two sub-classes Synapsida and Diapsida Science 17275ndash276
Owen R 1861 Palaeontology or a systematic summary of extinct ani-mals and their geological relations 2nd Ed Edinburgh
Parrington F R 1934 On the cynodont genus Galesaurus with a noteon the functional significance of the changes in the evolution of thetheriodont skull Annals and Magazine of Natural History 10(13)38ndash67
Parrington F R 1946 On the cranial anatomy of cynodonts Proceed-ings of the Zoological Society of London 116181ndash197
Razafimbelo E 1987 Le basin de Morondava (Madagascar) Synthegravesegeacuteologique et structurale PhD dissertation University Louis Pas-teur Strasbourg France 189 pp
Reisz R R and H-D Sues 2000 Herbivory in Late Paleozoic andTriassic terrestrial vertebrates pp 9ndash41 in Sues H-D (ed) Evo-lution of Herbivory in Terrestrial Vertebrates Cambridge Univer-sity Press New York
Romer A S 1967 The Chantildeares (Argentina) Triassic reptile fauna IIITwo new gomphodonts Massetognathus pascuali and M terugiiBreviora 2641ndash25
Romer A S 1972 The Chantildeares (Argentina) Triassic reptile faunaXVII The Chantildeares gomphodonts Breviora 3961ndash9
Rougier G W J R Wible and J A Hopson 1992 Reconstruction ofthe cranial vessels in the Early Cretaceous mammal Vincelestes neu-quenianus implications for the evolution of the mammalian cranialvascular system Journal of Vertebrate Paleontology 12188ndash216
Rowe T 1988 Definition diagnosis and origin of Mammalia Journal ofVertebrate Paleontology 8241ndash264
Rowe T 1993 Early mammal phylogenetic systematics pp 129ndash145 inSzalay F S M J Novacek and M C McKenna (eds) MammalPhylogeny Vol 1 Mesozoic Differentiation MultituberculatesMonotremes Early Therians and Marsupials Springer-Verlag NewYork
Seeley H G 1894 Researches on the structure organization and clas-sification of the fossil Reptilia Part IX Section 3 On DiademodonPhilosophical Transactions of the Royal Society of London 1851029ndash1041
Seeley H G 1895 Researches on the structure organization and clas-sification of the fossil Reptilia Part IX Section 4 On the Gompho-dontia Philosophical Transactions of the Royal Society of London1861ndash57
Seeley H G 1895 Researches on the structure organization and clas-sification of the fossil Reptilia Part IX Section 5 On the skeletonin new Cynodontia from the Karroo rocks Philosophical Transac-tions of the Royal Society of London 18659ndash148
Sues H-D J A Hopson and N H Shubin 1992 Affinities of Sca-lenodontoides plemmyridon Hopson 1984 (Synapsida Cynodontia)from the Upper Triassic of Nova Scotia Journal of Vertebrate Pa-leontology 12168ndash171
Sues H-D and P E Olsen 1990 Triassic vertebrates of Gondwananaspect from the Richmond basin of Virginia Science 2491020ndash1023
Swofford D L 2003 PAUP v 40b10 Sinauer Associates Inc Pub-lishers Sunderland Massachusetts
Watson D M S and A S Romer 1956 A classification of therapsidreptiles Bulletin of the Museum of Comparative Zoology 11435ndash89
Submitted September 20 2007 accepted November 3 2007
KAMMERER ET ALmdashCYNODONT FROM MADAGASCAR 461
Dow
nloa
ded
by [
UZ
H H
aupt
bibl
ioth
ek
Zen
tral
bibl
ioth
ek Z
uumlric
h] a
t 05
51 1
0 Se
ptem
ber
2013
APPENDIX Data matrix used in the phylogenetic analysesThis is a revised version of the matrix of Abdala et al (2006) with the following changes character 43 [Incisor procumbency (0) absent (1) present]added Menadon recoded for characters 0 (Number of upper incisors was 1 ldquothreerdquo changed to 0 ldquofourrdquo) 5 (Position of paracanine fossae in relationto the upper canine was changed to 1 ldquomedialrdquo) 16 (Overall morphology of lower postcanines in occlusal view was changed to 3 ldquoquadran-gularrdquo) 17 (Position of the transverse crest in lower postcanines was changed to 1 ldquoanteriorrdquo) 18 (Number of cusps in the transverse crest of thelower postcanines was changed to 0 ldquotwordquo) 20 (Anterior cingulum in the lower postcanines was changed to 2 ldquoabsentrdquo) 21 (Internarial barwas 1 ldquoabsentrdquo changed to 0 ldquopresentrdquo) 38 (Posterior cingulum in the lower postcanines was changed to 1 ldquoabsentrdquo) and 40 (Deep occlusalbasins in the postcanines was changed to 1 ldquopresentrdquo) Langbergia recoded for character 34 (Incisor cutting margins was 0 ldquoserratedrdquo changedto 1 ldquosmoothly ridgedrdquo) based on the description of the incisors in Abdala et al (2006) Gomphodontosuchus recoded for character 31 (Maxilla inthe margin of the subtemporal fenestra was 1 ldquoincludedrdquo changed to 0 ldquoexcludedrdquo) based on von Huene (1936) and Kammerer (pers obs of theholotype) contra Hopsonrsquos (1985) reconstruction lsquoScalenodonrsquo hirschsoni recoded for character 39 (Anterior cingulum in the upper postcanines was1 ldquoabsentrdquo changed to 0 ldquopresentrdquo) based on Abdala (pers comm 2006) Trirachodon recoded for character 30 (Ectopterygoid was changedto 0 ldquopresentrdquo) based on Abdala (pers comm 2006) and Traversodon recoded for character 15 (Number of cusps in the sectorial border of the upperpostcanines was 0 ldquomore than twordquo changed to 1 ldquotwordquo) based on Abdala (pers comm 2006) and Kammerer (pers obs)
1111111111 2222222222 3333333333 44440123456789 0123456789 0123456789 0123456789 0123
Cynognathus 00000000 000 000000000 00000000 000Diademodon 0000001000 0000100 000010A000 0000000100 0100Trirachodon 000000100B 1A002201 001A100010 0001001200 0110Langbergia 0000001000 10000201 0000101010 0001101200 0110Cricodon 0100001000 11000201 0010100 0001200 0110Andescynodon 0000002001 01013101 101120010 0112311 1100Massetognathus 00111121A2 1110003100 2011200011 1011212300 1100Exaeretodon 1110122112 010103101 2111110201 1111112311 1101Luangwa 0000001002 11010A3100 101010010 01002300 1100Scal angustifrons 000000100B 1111023100 10102101 00002301 1100Scal hirschsoni 11110202 1100113100 1010 1112300 1100Traversodon 0000002012 1101113100 201010 011230 1100Gomphodontosuchus 0011112112 0103101 2010 0112311 1100Pascualgnathus 1000001000 000231 1100001 0011230 1100Scalenodontoides 1112212 01003101 21111021 1212311 1101Menadon 0111112112 01012310 20121020 0102311 1101Dadadon 00002112 111112 013 11231 1100Santacruzodon 00002112 1110003101 201300 212300 1100
A(01) B(02)
JOURNAL OF VERTEBRATE PALEONTOLOGY VOL 28 NO 2 2008462
Dow
nloa
ded
by [
UZ
H H
aupt
bibl
ioth
ek
Zen
tral
bibl
ioth
ek Z
uumlric
h] a
t 05
51 1
0 Se
ptem
ber
2013
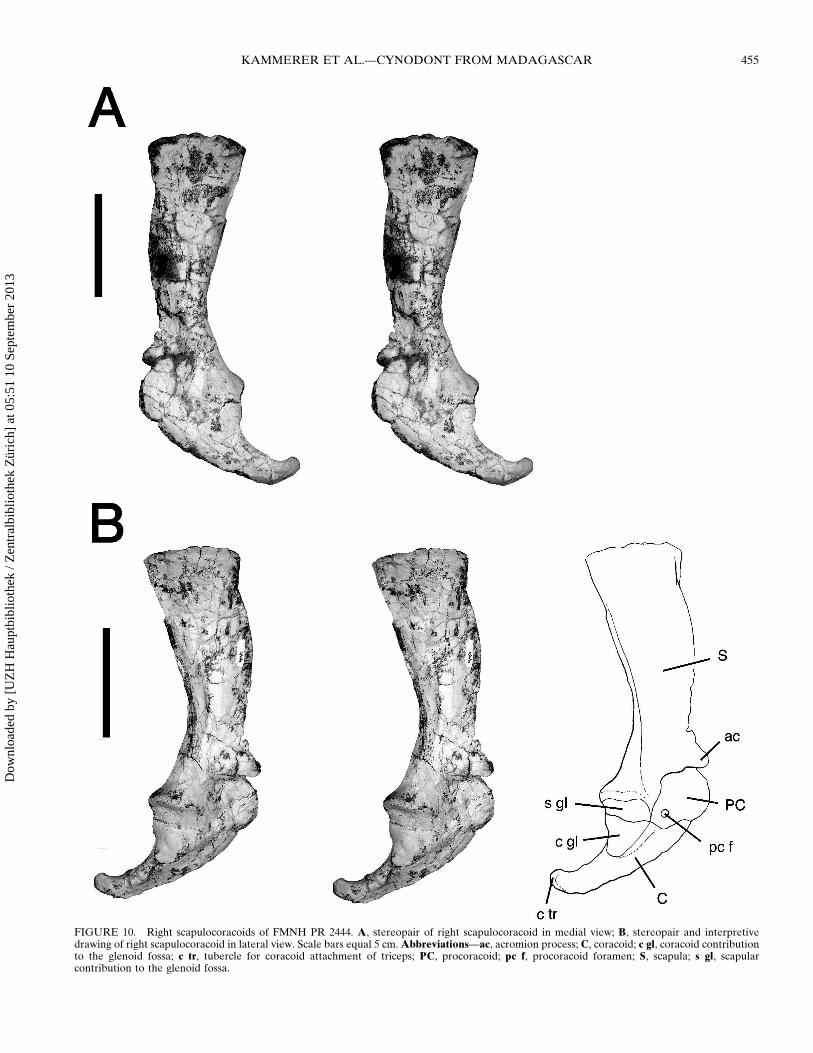
FIGURE 10 Right scapulocoracoids of FMNH PR 2444 A stereopair of right scapulocoracoid in medial view B stereopair and interpretivedrawing of right scapulocoracoid in lateral view Scale bars equal 5 cm Abbreviationsmdashac acromion process C coracoid c gl coracoid contributionto the glenoid fossa c tr tubercle for coracoid attachment of triceps PC procoracoid pc f procoracoid foramen S scapula s gl scapularcontribution to the glenoid fossa
KAMMERER ET ALmdashCYNODONT FROM MADAGASCAR 455
Dow
nloa
ded
by [
UZ
H H
aupt
bibl
ioth
ek
Zen
tral
bibl
ioth
ek Z
uumlric
h] a
t 05
51 1
0 Se
ptem
ber
2013
teeth for a traversodontid As mentioned above Flynn et al(2000) noted that the homologies of these teeth were difficult todetermine Of the five anterior teeth preserved in the holotypethe anteriormost three are ldquoundoubtedly incisorsrdquo although I3 isldquounusually large and is caniniformrdquo (Flynn et al 2000423) Therobust recurved fourth tooth in the series was considered torepresent the canine with the subsequent tooth interpreted as anolder canine in the process of replacement Although FMNH PR2444 is edentulous the alveoli on the left side of the skull are wellpreserved and indicate an anterior tooth set identical to the ho-lotype The alveolar morphology indicates that the fifth toothwas canted anteriorly as in the holotype This suggests that themysterious fifth tooth is not the result of a transient period in thedental replacement history of M besairiei as suggested by Flynnet al (2000) but rather is the norm This leaves two possibilities1) M besairiei persistently had two canines This condition isunknown in cynodonts 2) The canine-like fourth tooth actuallyrepresents an incisor and the fifth tooth is the true canine Wefavor the latter explanation In FMNH PR 2444 the first fouralveoli are nearly identical in size and shape However the fifthalveolus while similar in anteroposterior length to its predeces-sors is strongly compressed labiolingually with an anteroposte-rior lengthlabiolingual width ratio of 044 (compared to 080 inthe fourth alveolus and 084ndash85 for I2ndash3) We interpret thismarked difference in alveolar ratio as indicating the separationbetween incisors and canine The canine-like I4 would then rep-resent the endpoint in a series of large increasingly caniniformincisors Supporting this interpretation recurved incisors are alsoobserved in juvenile Exaeretodon (eg MCP 3843 PV see Ab-dala et al 2002)
There are several legitimate differences between FMNH PR2444 and the holotype of Menadon besairiei but we take a con-servative taxonomic stance and argue that these differences rep-resent intraspecific variation and preservational artifacts FMNHPR 2444 represents a larger animal than UA 10601 The snoutlength (measured as the distance between the anterior edge ofthe orbit and a vertical plane through the anterior tip of thepremaxilla) of FMNH PR 2444 is 911 mm while that of UA10601 is 801 mm As such proportional differences between thetwo skulls may be attributable to ontogeny Variations in FMNHPR 2444 compared to the holotype that probably represent on-togentic differences include wider longer and deeper paraca-nine fossae a much deeper ventral zygomatic flange (althoughthe lower border of the flange is slightly abraded in the holotypeof Menadon besairiei there is no evidence that a major flangewas broken off) a proportionally shorter and more circular out-line to the posterior border of the maxilla below the zygomaticprocess and a more deeply emarginated posterior squamosal(ie a more prominent laterally projecting flange)
FMNH PR 2444 has a proportionally shorter more robustsnout and larger orbits than the holotype While these charactersare certainly variable in cynodonts it is unusual that the largerspecimen is shorter-snouted and larger-eyed contra the usualpattern The proportional differences between the two skulls areminor however (orbit diametersnout length is 042 for FMNHPR 2444 and 041 for the holotype) and are partially attributableto deformation (shear in FMNH PR 2444 and breakage of thepostorbital bar in the holotype) The following differences canalso probably be attributed to preservational artifact only fivepostcanine alveoli are preserved in FMNH PR 2444 while theholotype has eight postcanines The anteriormost three in theholotype however are tiny and peg-like As the area of themaxilla between the canine alveolus and the first preserved post-canine alveolus is poorly preserved in FMNH PR 2444 the ab-sence of these postcanines may be artifactual However it is alsopossible that the lower postcanine count of FMNH PR 2444compared to the holotype is an ontogenetic effect as postcaninecount is known to decrease with increasing skull size in Ex-
FIGURE 11 Left scapulocoracoids of FMNH PR 2444 A left scapu-locoracoid in medial view B left scapulocoracoid in lateral view Scalebars equal 5 cm
JOURNAL OF VERTEBRATE PALEONTOLOGY VOL 28 NO 2 2008456
Dow
nloa
ded
by [
UZ
H H
aupt
bibl
ioth
ek
Zen
tral
bibl
ioth
ek Z
uumlric
h] a
t 05
51 1
0 Se
ptem
ber
2013
FIGURE 12 Right pelvis of FMNH PR 2444 A stereopair of right pelvis in medial view B stereopair and interpretive drawing of right pelvis inlateral view Scale bars equal 5 cm Abbreviationsmdashbpp base of posterior process of ilium ib iliac blade IL ilium IS ischium ob f obturatorforamen P pubis
KAMMERER ET ALmdashCYNODONT FROM MADAGASCAR 457
Dow
nloa
ded
by [
UZ
H H
aupt
bibl
ioth
ek
Zen
tral
bibl
ioth
ek Z
uumlric
h] a
t 05
51 1
0 Se
ptem
ber
2013
aeretodon (Abdala et al 2002) The orbit of FMNH PR 2444 isless upwardly directed than that of the holotype (being at anangle of sim65deg to a sagittal plane through the midline of the skullas opposed to sim45deg in the holotype) This difference in orbitalangle is probably attributable to postmortem deformation as theskull of FMNH PR 2444 is noticeably sheared to the front leftThe appearance of a wider squarer incisor arcade and broader
anterior palate in FMNH PR 2444 can also be attributed to thisshear Finally the occipital region of FMNH PR 2444 is alto-gether taller and narrower than that of the holotype For ex-ample the foramen magnum in FMNH PR 2444 approximates arounded isosceles triangle in shape while the foramen magnumof the holotype is more ovoid These differences can also beattributed to deformation Altogether the differences between
FIGURE 13 Stereopairs of field number 8-31-98-387 an isolated right pelvis in A medial and B lateral view Scale bars equal 5 cm
JOURNAL OF VERTEBRATE PALEONTOLOGY VOL 28 NO 2 2008458
Dow
nloa
ded
by [
UZ
H H
aupt
bibl
ioth
ek
Zen
tral
bibl
ioth
ek Z
uumlric
h] a
t 05
51 1
0 Se
ptem
ber
2013
FMNH PR 2444 and the holotype fall within the range of intra-specific variation recorded for other gomphodont cynodontssuch as the traversodontids Massetognathus pascuali (Abdalaand Giannini 2000 Kammerer pers obs) and Exaeretodon(Abdala et al 2002) and the diademodontid Diademodon tetra-gonus (Grine et al 1978 Bradu and Grine 1979)
Phylogenetic Analysis
Utilizing the new information afforded by FMNH PR 2444and a reexamination of the holotype (UA 10601) Menadon be-sairiei was recoded in the Gondwanan gomphodont data matrixof Abdala et al (2006) Abdala et alrsquos (2006) data matrix en-compassed 18 taxa and 43 characters For the present analysis anadditional character ldquoincisor procumbencyrdquo (states 0-absent1-present) was added to Abdala et alrsquos (2006) data set (see
Appendix for coding of this character) Several traversodontidtaxa (eg lsquoScalenodonrsquo hirschsoni) have relatively large incisorsbut only in Menadon Scalenodontoides and Exaeretodon are theincisors procumbent Three of Abdala et alrsquos (2006) originalcodings for Menadon besairiei characters 0 (ldquonumber of upperincisorsrdquo) 5 (ldquoposition of paracanine fossa in relation to theupper caninerdquo) and 21 (ldquointernarial barrdquo) were changed to in-corporate data from the new specimen (see Appendix) Addi-tionally further examination of the holotype (UA 10601) ofMenadon besairiei allowed us to code characters 16ndash18 20 38and 40 (morphology of the lower postcanines all marked as ldquordquoin Abdala et alrsquos analysis) Parsimony searches were performedin PAUP v40b10 using a branch-and-bound search Addition-ally a bootstrap analysis (also using branch-and-bound) was per-formed using 1000 replicates
A total of 34 most parsimonious trees of length 103 werefound with the scores CI0573 RI0725 RC0415 andHI0427 The strict consensus tree (Fig 15A) did not recovera polytomy between Menadon Exaeretodon and Scalenodont-oides as in Abdala and Ribeiro (2003) or a (Menadon + Ex-aeretodon) sister-group relationship as in Abdala et al (2006)Rather Menadon was recovered as the sister taxon of the (Ex-aeretodon + Scalenodontoides) clade supporting Flynn et alrsquos(2000) original placement of this taxon There is strong bootstrapsupport (88) for Traversodontidae but within traversodontidsthe only strongly supported subclade is (Gomphodontosuchus +(Menadon + (Scalenodontoides + Exaeretodon))) with a boot-strap value of 80 Within this clade the (Menadon + (Scalen-odontoides + Exaeretodon)) grouping is supported by a boot-strap value of 83 and the (Scalenodontoides + Exaeretodon)grouping is supported by a bootstrap value of 94 (Fig 15B) Inaddition to the positioning of Menadon the strict consensus tree(Fig 15A) differs from the topology of Abdala et al (2006) inrecovering a sister-group relationship between Massetognathusand Santacruzodon rather than having Massetognathus andSantacruzodon as successive outgroups to the clade containingGomphodontosuchus Menadon Scalenodontoides and Ex-aeretodon The (Massetognathus + Santacruzodon) clade is diag-nosed by the absence of an anterolingual cusp in the upper post-canines denticulated incisor cutting margins and the presence ofan anterior cingulum in the upper postcanines
FIGURE 14 Reconstruction of the pelvis of Menadon besairiei basedon information from FMNH PR 2444 and field number 8-31-98-387Dotted area not preserved in either specimen reconstruction based onPascualgnathus (Bonaparte 1966) and Exaeretodon (Bonaparte 1963)
FIGURE 15 Cladogram of traversodontid relationships based on revised data set of Abdala et al (2006) showing A strict consensus tree and B50 majority rule tree Numbers at nodes of majority rule tree represent bootstrap values
KAMMERER ET ALmdashCYNODONT FROM MADAGASCAR 459
Dow
nloa
ded
by [
UZ
H H
aupt
bibl
ioth
ek
Zen
tral
bibl
ioth
ek Z
uumlric
h] a
t 05
51 1
0 Se
ptem
ber
2013
Relationships of Menadon besairiei
Menadon besairiei is the sister taxon to (Exaeretodon + Sca-lenodontoides) the latter clade diagnosed by the possession ofthree upper incisors a posteromedial position of the paracaninefossae relative to the canines the absence of an internarial barlarge canines a projected zygomatic process of the jugal and amaxilla that contributes to the margin of the subtemporal fenes-tra (the last three character states represent reversals) The clade(Menadon + (Exaeretodon + Scalenodontoides)) is diagnosed bythe possession of large procumbent incisors The presence offour incisors a primitive retention distinguishes Menadon be-sairiei from Exaeretodon Scalenodontoides and an unnamed tra-versodontid (ldquoTraversodontid Type 1rdquo) from the Santa MariaFormation of Brazil (Abdala et al 2001 Abdala and Ribeiro2002) Gomphodontosuchus is recovered as the sister taxon to(Menadon (Exaeretodon + Scalenodontoides)) as has beenfound in previous studies (Flynn et al 2000 Abdala and Ribeiro2003) The taxon name Gomphodontosuchinae Watson andRomer 1956 (ex Gomphodontosuchidae Watson and Romer1956 first given subfamilial rank in Lehman 1961) is availablefor this clade As circumscribed here Gomphodontosuchinaeconsists of the most inclusive clade containing Gomphodontosu-chus brasiliensis von Huene 1928264 but not Massetognathuspascuali Romer 19672
Menadon besairiei displays a mixture of primitive and derivedcharacters within traversodontids The internarial bar and fourincisors are primitive features whereas the enlargement and pro-cumbency of the incisors are derived features shared with Ex-aeretodon and Scalenodontoides The postcranium shows severalsimilarities to that of Exaeretodon (apparent exclusion of theprocoracoid from the glenoid some features of the pelvis tallcaudal neural spines) but in general displays a rather conserva-tive morphology observed throughout Gomphodontia The mor-phology of the scapulocoracoids is closer akin to diademodontidsor trirachodontids than Exaeretodon and the expanded thoracicrib of Menadon has only slight differences from those of trirach-odontids or early traversodontids like Luangwa The atlas-axiscomplex of Menadon is similar to that of the Early Triassic cyn-odont Cynognathus in possessing a concave axial spine with along posterior process Although most gomphodonts have beenreconstructed with a convex axial spine as in early mammals thisspine is broken in most of the best described gomphodont post-crania (eg Diademodon Brink 1955) and the lsquosaddle-shapedrsquoaxial morphology may be more widely distributed among cyn-odonts than previously realized The new specimens of Menadondescribed in this paper demonstrate that a generalized gompho-dont postcranial morphology was retained deep within the tra-versodontid tree and that the derived morphologies of either ofthe well-known taxa Massetognathus or Exaeretodon should notbe taken as representative of traversodontids in general
ACKNOWLEDGMENTS
This work would not have been possible without the continu-ing support and encouragement of our collaborative programwith the Deacutepartement de Paleacuteontologie et drsquoAnthropologie Bi-ologique Universiteacute drsquoAntananarivo Madagascar under the di-rection of Armand Rasoamiaramanana (Department Chairman)and Gisegravele Ravololonarivo Randria (Head of the PaleontologyLaboratories) We are extremely grateful to the National Geo-graphic Society for their long-term support of the fieldwork as-sociated with this project Additional financial or logistical sup-port for fieldwork was provided by The Field Museum the Uni-versity of California-Santa Barbara and the WWF (WorldwideFund for Nature Madagascar) We acknowledge the Ministegraverede LrsquoEneacutergie et des Mines for authorizing field efforts and for
continuing to facilitate our joint US-Madagascar explorationresearch and education program Steve Goodman and AsminaGandhi generously assisted us in innumerable ways includinghospitality nonpareil We especially thank R Andriantompoha-vana L Razafimanantsoa A Toto Volahy JA Rabarison AGandie and P Vavisaro for assistance in the field during theexpedition in which these new specimens were discovered BillSimpson and Gina Wesley-Hunt for their patient excavation ofthis important material Allison Smith Ray Vodden Lisa Berg-wall and Deborah Wagner for excellent preparation of a chal-lenging specimen Jennifer Botha and Elize Butler (NationalMuseum Bloemfontein) Michael Raath and Fernando Abdala(Bernard Price Institute) Thomas Kemp (Oxford UniversityMuseum of Natural History) Ray Symonds (University Museumof Zoology Cambridge) Bernard Battail (Museacuteum nationaldrsquoHistoire naturelle Paris) Michael Maisch (Universitaumlt Tuumlbin-gen) and Charles Schaff and Farish Jenkins (Museum of Com-parative Zoology Harvard) for access to specimens and AgustiacutenMartinelli Fernando Abdala and James Hopson for helpful re-views of an earlier version of the manuscript
LITERATURE CITED
Abdala F M C Barbarena and J Dornelles 2002 A new species of thetraversodontid cynodont Exaeretodon from the Santa Maria Forma-tion (MiddleLate Triassic) of Southern Brazil Journal of Verte-brate Paleontology 22313ndash325
Abdala F and N P Giannini 2000 Gomphodont cynodonts of theChantildeares Formation the analysis of an ontogenetic sequence Jour-nal of Vertebrate Paleontology 20501ndash506
Abdala F J Neveling and J Welman 2006 A new trirachodontidcynodont from the lower levels of the Burgersdorp Formation(Lower Triassic) of the Beaufort Group South Africa and the cla-distic relationships of Gondwanan gomphodonts Zoological Jour-nal of the Linnean Society 147383ndash413
Abdala F A M Ribeiro and C L Schultz 2001 A rich cynodont faunaof Santa Cruz do Sul Santa Maria Formation (Middle-Late Trias-sic) southern Brazil Neues Jahrbuch fuumlr Geologie und Palaumlontolo-gie Monatshefte 2001669ndash687
Abdala F and A M Ribeiro 2002 Nuevos cinodontes traversodoacuten-tidos (Synapsida-Eucynodontia) de la Formacioacuten Santa Maria(Triaacutesico Medio-Superior) Rio Grande do Sul Brasil RevistaEspantildeola de Paleontologiacutea 17237ndash243
Abdala F and A M Ribeiro 2003 A new traversodontid cynodontfrom the Santa Maria Formation (Ladinian-Carnian) of southernBrazil with a phylogenetic analysis of Gondwanan traversodontidsZoological Journal of the Linnaean Society 139529ndash545
Abdala F and A M S Teixeira 2004 A traversodontid cynodont ofAfrican affinity in the South American Triassic Palaeontologia Af-ricana 4011ndash22
Bonaparte J F 1962 Descripcioacuten del craacuteneo y mandiacutebula de Ex-aeretodon frenguellii Cabrera y su comparacioacuten con Diademodon-tidae Tritylodontidae y los cinodontes sudamericanos Publica-ciones del Museo Municipal de Ciencias Naturales y Tradicional deMar del Plata 1135ndash202
Bonaparte J F 1963 Descripcioacuten del esqueleto postcraneano de Ex-aeretodon (CynodontiandashTraversodontidae) Acta Geoloacutegica Lil-loana 45ndash52
Bonaparte J F 1966 Una nueva ldquofaunardquo Triaacutesica de Argentina (Ther-apsida Cynodontia Dicynodontia) Consideracions filogeneacuteticas ypaleobiogeograacuteficas Ameghiniana 8243ndash296
Bradu D and F E Grine 1979 Multivariate analysis of diademodon-tine crania from South Africa and Zambia South African Journal ofScience 79441ndash448
Brink A S 1955 A study of the skeleton of Diademodon Palaeonto-logia Africana 33-39
Brink A S 1963 Two new cynodonts from the Ntawere Formation inthe Luangwa Valley of Northern Rhodesia Palaeontologia Africana877ndash96
Broom R 1905 On the use of the term Anomodontia Records of theAlbany Museum 1266ndash269
Cabrera A 1943 El primer hallazgo de teraacutepsidos en la ArgentinaNotas del Museo de La Plata Paleontologiacutea 55317ndash331
JOURNAL OF VERTEBRATE PALEONTOLOGY VOL 28 NO 2 2008460
Dow
nloa
ded
by [
UZ
H H
aupt
bibl
ioth
ek
Zen
tral
bibl
ioth
ek Z
uumlric
h] a
t 05
51 1
0 Se
ptem
ber
2013
Chatterjee S 1982 A new cynodont reptile from the Triassic of IndiaJournal of Paleontology 56203ndash214
Crompton A W 1955 On some Triassic cynodonts from TanganyikaProceedings of the Zoological Society of London 125617ndash669
Crompton A W 1972 Postcanine occlusion in cynodonts and tritylo-donts Bulletin of the British Museum (Natural History) Geology2129ndash71
Crompton A W and F Ellenberger 1957 On a new cynodont from theMolteno Beds and the origin of the tritylodontids Annals of theSouth African Museum 441ndash13
Flynn J J J M Parrish B Rakotosaminimanana W F Simpson R LWhatley and A R Wyss 1999 A Triassic fauna from Madagascarincluding early dinosaurs Science 286763ndash765
Flynn J J J M Parrish B Rakotosamimanana L RanivoharimananaW F Simpson and A R Wyss 2000 New traversodontids (Syn-apsida Eucynodontia) from the Triassic of Madagascar Journal ofVertebrate Paleontology 20422ndash427
Gow C E and P J Hancox 1993 First complete skull of the LateTriassic Scalenodontoides (Reptilia Cynodontia) from southern Af-rica In Lucas S G and M Morales (eds) The nonmarine TriassicNew Mexico Museum of Natural History amp Science Bulletin 3161ndash168
Grine F E B D Hahn and C E Gow 1978 Aspects of relativegrowth and variability in Diademodon (Reptilia Therapsida) SouthAfrican Journal of Science 7450ndash58
Hopson J A 1984 Late Triassic traversodont cynodonts from NovaScotia and southern Africa Palaeontologia Africana 25181ndash201
Hopson J A 1985 Morphology and relationships of Gomphodontosu-chus brasiliensis von Huene (Synapsida Cynodontia Tritylodontoi-dea) from the Triassic of Brazil Neues Jahrbuch fuumlr Geologie undPalaumlontologie Monatshefte 1985285ndash299
Hopson J A and J W Kitching 2001 A probainognathian cynodontfrom South Africa and the phylogeny of nonmammalian cynodontsBulletin of the Museum of Comparative Zoology 1565ndash35
Hopson J A and H-D Sues 2006 A traversodont cynodont from theMiddle Triassic (Ladinian) of Baden-Wuumlrttemberg (Germany)Palaumlontologische Zeitschrift 80124ndash129
Huene F von 1928 Ein Cynodontier aus der Trias Brasiliens Zentral-blatt fuumlr Mineralogie Geologie und Palaumlontologie 1928251ndash270
Huene F von 1936 Die fossilen Reptilien des suumldamerikanischenGondwanalandes Ergebnisse der Sauriergrabungen in Suumldbrasilien192829 Lieferung 2 (Cynodontia)93ndash159 Franz F Heine Tuumlbin-gen
Jenkins F A 1970 The Chantildeares (Argentina) Triassic reptile faunaVII The postcranial skeleton of the traversodontid Massetognathuspascuali (Therapsida Cynodontia) Breviora 3521ndash28
Jenkins F A 1971 The postcranial skeleton of African cynodonts prob-lems in the early evolution of the mammalian postcranial skeletonBulletin of the Peabody Museum of Natural History 361ndash216
Kemp T S 1969 The atlas-axis complex of mammal-like reptiles Jour-nal of Zoology 159223ndash248
Kemp T S 1980 Aspects of the structure and functional anatomy of theMiddle Triassic cynodont Luangwa Journal of Zoology 191193ndash239
Kemp T S 1982 Mammal-Like Reptiles and the Origin of MammalsAcademic Press New York
Lehman J-P 1961 Cynodontia pp 140ndash191 in Piveteau J (ed) Traiteacutede Paleacuteontologie Masson et Cie Paris
Osborn 1903 On the primary division of the Reptilia into two sub-classes Synapsida and Diapsida Science 17275ndash276
Owen R 1861 Palaeontology or a systematic summary of extinct ani-mals and their geological relations 2nd Ed Edinburgh
Parrington F R 1934 On the cynodont genus Galesaurus with a noteon the functional significance of the changes in the evolution of thetheriodont skull Annals and Magazine of Natural History 10(13)38ndash67
Parrington F R 1946 On the cranial anatomy of cynodonts Proceed-ings of the Zoological Society of London 116181ndash197
Razafimbelo E 1987 Le basin de Morondava (Madagascar) Synthegravesegeacuteologique et structurale PhD dissertation University Louis Pas-teur Strasbourg France 189 pp
Reisz R R and H-D Sues 2000 Herbivory in Late Paleozoic andTriassic terrestrial vertebrates pp 9ndash41 in Sues H-D (ed) Evo-lution of Herbivory in Terrestrial Vertebrates Cambridge Univer-sity Press New York
Romer A S 1967 The Chantildeares (Argentina) Triassic reptile fauna IIITwo new gomphodonts Massetognathus pascuali and M terugiiBreviora 2641ndash25
Romer A S 1972 The Chantildeares (Argentina) Triassic reptile faunaXVII The Chantildeares gomphodonts Breviora 3961ndash9
Rougier G W J R Wible and J A Hopson 1992 Reconstruction ofthe cranial vessels in the Early Cretaceous mammal Vincelestes neu-quenianus implications for the evolution of the mammalian cranialvascular system Journal of Vertebrate Paleontology 12188ndash216
Rowe T 1988 Definition diagnosis and origin of Mammalia Journal ofVertebrate Paleontology 8241ndash264
Rowe T 1993 Early mammal phylogenetic systematics pp 129ndash145 inSzalay F S M J Novacek and M C McKenna (eds) MammalPhylogeny Vol 1 Mesozoic Differentiation MultituberculatesMonotremes Early Therians and Marsupials Springer-Verlag NewYork
Seeley H G 1894 Researches on the structure organization and clas-sification of the fossil Reptilia Part IX Section 3 On DiademodonPhilosophical Transactions of the Royal Society of London 1851029ndash1041
Seeley H G 1895 Researches on the structure organization and clas-sification of the fossil Reptilia Part IX Section 4 On the Gompho-dontia Philosophical Transactions of the Royal Society of London1861ndash57
Seeley H G 1895 Researches on the structure organization and clas-sification of the fossil Reptilia Part IX Section 5 On the skeletonin new Cynodontia from the Karroo rocks Philosophical Transac-tions of the Royal Society of London 18659ndash148
Sues H-D J A Hopson and N H Shubin 1992 Affinities of Sca-lenodontoides plemmyridon Hopson 1984 (Synapsida Cynodontia)from the Upper Triassic of Nova Scotia Journal of Vertebrate Pa-leontology 12168ndash171
Sues H-D and P E Olsen 1990 Triassic vertebrates of Gondwananaspect from the Richmond basin of Virginia Science 2491020ndash1023
Swofford D L 2003 PAUP v 40b10 Sinauer Associates Inc Pub-lishers Sunderland Massachusetts
Watson D M S and A S Romer 1956 A classification of therapsidreptiles Bulletin of the Museum of Comparative Zoology 11435ndash89
Submitted September 20 2007 accepted November 3 2007
KAMMERER ET ALmdashCYNODONT FROM MADAGASCAR 461
Dow
nloa
ded
by [
UZ
H H
aupt
bibl
ioth
ek
Zen
tral
bibl
ioth
ek Z
uumlric
h] a
t 05
51 1
0 Se
ptem
ber
2013
APPENDIX Data matrix used in the phylogenetic analysesThis is a revised version of the matrix of Abdala et al (2006) with the following changes character 43 [Incisor procumbency (0) absent (1) present]added Menadon recoded for characters 0 (Number of upper incisors was 1 ldquothreerdquo changed to 0 ldquofourrdquo) 5 (Position of paracanine fossae in relationto the upper canine was changed to 1 ldquomedialrdquo) 16 (Overall morphology of lower postcanines in occlusal view was changed to 3 ldquoquadran-gularrdquo) 17 (Position of the transverse crest in lower postcanines was changed to 1 ldquoanteriorrdquo) 18 (Number of cusps in the transverse crest of thelower postcanines was changed to 0 ldquotwordquo) 20 (Anterior cingulum in the lower postcanines was changed to 2 ldquoabsentrdquo) 21 (Internarial barwas 1 ldquoabsentrdquo changed to 0 ldquopresentrdquo) 38 (Posterior cingulum in the lower postcanines was changed to 1 ldquoabsentrdquo) and 40 (Deep occlusalbasins in the postcanines was changed to 1 ldquopresentrdquo) Langbergia recoded for character 34 (Incisor cutting margins was 0 ldquoserratedrdquo changedto 1 ldquosmoothly ridgedrdquo) based on the description of the incisors in Abdala et al (2006) Gomphodontosuchus recoded for character 31 (Maxilla inthe margin of the subtemporal fenestra was 1 ldquoincludedrdquo changed to 0 ldquoexcludedrdquo) based on von Huene (1936) and Kammerer (pers obs of theholotype) contra Hopsonrsquos (1985) reconstruction lsquoScalenodonrsquo hirschsoni recoded for character 39 (Anterior cingulum in the upper postcanines was1 ldquoabsentrdquo changed to 0 ldquopresentrdquo) based on Abdala (pers comm 2006) Trirachodon recoded for character 30 (Ectopterygoid was changedto 0 ldquopresentrdquo) based on Abdala (pers comm 2006) and Traversodon recoded for character 15 (Number of cusps in the sectorial border of the upperpostcanines was 0 ldquomore than twordquo changed to 1 ldquotwordquo) based on Abdala (pers comm 2006) and Kammerer (pers obs)
1111111111 2222222222 3333333333 44440123456789 0123456789 0123456789 0123456789 0123
Cynognathus 00000000 000 000000000 00000000 000Diademodon 0000001000 0000100 000010A000 0000000100 0100Trirachodon 000000100B 1A002201 001A100010 0001001200 0110Langbergia 0000001000 10000201 0000101010 0001101200 0110Cricodon 0100001000 11000201 0010100 0001200 0110Andescynodon 0000002001 01013101 101120010 0112311 1100Massetognathus 00111121A2 1110003100 2011200011 1011212300 1100Exaeretodon 1110122112 010103101 2111110201 1111112311 1101Luangwa 0000001002 11010A3100 101010010 01002300 1100Scal angustifrons 000000100B 1111023100 10102101 00002301 1100Scal hirschsoni 11110202 1100113100 1010 1112300 1100Traversodon 0000002012 1101113100 201010 011230 1100Gomphodontosuchus 0011112112 0103101 2010 0112311 1100Pascualgnathus 1000001000 000231 1100001 0011230 1100Scalenodontoides 1112212 01003101 21111021 1212311 1101Menadon 0111112112 01012310 20121020 0102311 1101Dadadon 00002112 111112 013 11231 1100Santacruzodon 00002112 1110003101 201300 212300 1100
A(01) B(02)
JOURNAL OF VERTEBRATE PALEONTOLOGY VOL 28 NO 2 2008462
Dow
nloa
ded
by [
UZ
H H
aupt
bibl
ioth
ek
Zen
tral
bibl
ioth
ek Z
uumlric
h] a
t 05
51 1
0 Se
ptem
ber
2013
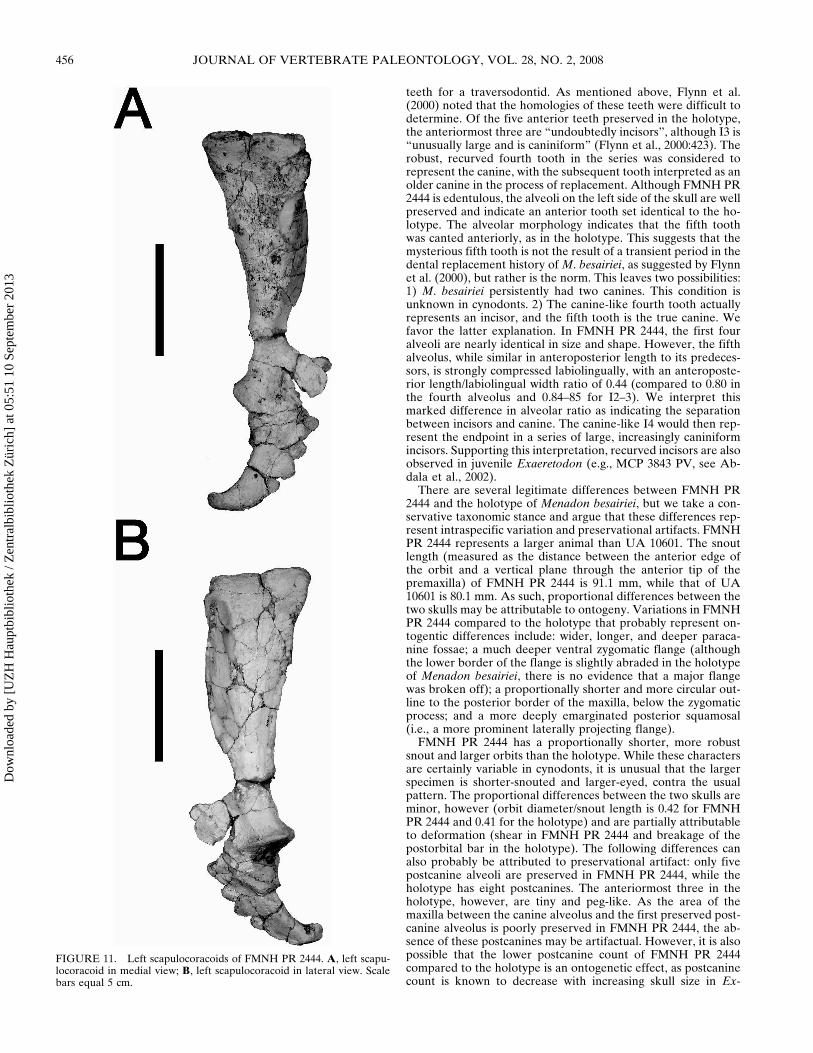
teeth for a traversodontid As mentioned above Flynn et al(2000) noted that the homologies of these teeth were difficult todetermine Of the five anterior teeth preserved in the holotypethe anteriormost three are ldquoundoubtedly incisorsrdquo although I3 isldquounusually large and is caniniformrdquo (Flynn et al 2000423) Therobust recurved fourth tooth in the series was considered torepresent the canine with the subsequent tooth interpreted as anolder canine in the process of replacement Although FMNH PR2444 is edentulous the alveoli on the left side of the skull are wellpreserved and indicate an anterior tooth set identical to the ho-lotype The alveolar morphology indicates that the fifth toothwas canted anteriorly as in the holotype This suggests that themysterious fifth tooth is not the result of a transient period in thedental replacement history of M besairiei as suggested by Flynnet al (2000) but rather is the norm This leaves two possibilities1) M besairiei persistently had two canines This condition isunknown in cynodonts 2) The canine-like fourth tooth actuallyrepresents an incisor and the fifth tooth is the true canine Wefavor the latter explanation In FMNH PR 2444 the first fouralveoli are nearly identical in size and shape However the fifthalveolus while similar in anteroposterior length to its predeces-sors is strongly compressed labiolingually with an anteroposte-rior lengthlabiolingual width ratio of 044 (compared to 080 inthe fourth alveolus and 084ndash85 for I2ndash3) We interpret thismarked difference in alveolar ratio as indicating the separationbetween incisors and canine The canine-like I4 would then rep-resent the endpoint in a series of large increasingly caniniformincisors Supporting this interpretation recurved incisors are alsoobserved in juvenile Exaeretodon (eg MCP 3843 PV see Ab-dala et al 2002)
There are several legitimate differences between FMNH PR2444 and the holotype of Menadon besairiei but we take a con-servative taxonomic stance and argue that these differences rep-resent intraspecific variation and preservational artifacts FMNHPR 2444 represents a larger animal than UA 10601 The snoutlength (measured as the distance between the anterior edge ofthe orbit and a vertical plane through the anterior tip of thepremaxilla) of FMNH PR 2444 is 911 mm while that of UA10601 is 801 mm As such proportional differences between thetwo skulls may be attributable to ontogeny Variations in FMNHPR 2444 compared to the holotype that probably represent on-togentic differences include wider longer and deeper paraca-nine fossae a much deeper ventral zygomatic flange (althoughthe lower border of the flange is slightly abraded in the holotypeof Menadon besairiei there is no evidence that a major flangewas broken off) a proportionally shorter and more circular out-line to the posterior border of the maxilla below the zygomaticprocess and a more deeply emarginated posterior squamosal(ie a more prominent laterally projecting flange)
FMNH PR 2444 has a proportionally shorter more robustsnout and larger orbits than the holotype While these charactersare certainly variable in cynodonts it is unusual that the largerspecimen is shorter-snouted and larger-eyed contra the usualpattern The proportional differences between the two skulls areminor however (orbit diametersnout length is 042 for FMNHPR 2444 and 041 for the holotype) and are partially attributableto deformation (shear in FMNH PR 2444 and breakage of thepostorbital bar in the holotype) The following differences canalso probably be attributed to preservational artifact only fivepostcanine alveoli are preserved in FMNH PR 2444 while theholotype has eight postcanines The anteriormost three in theholotype however are tiny and peg-like As the area of themaxilla between the canine alveolus and the first preserved post-canine alveolus is poorly preserved in FMNH PR 2444 the ab-sence of these postcanines may be artifactual However it is alsopossible that the lower postcanine count of FMNH PR 2444compared to the holotype is an ontogenetic effect as postcaninecount is known to decrease with increasing skull size in Ex-
FIGURE 11 Left scapulocoracoids of FMNH PR 2444 A left scapu-locoracoid in medial view B left scapulocoracoid in lateral view Scalebars equal 5 cm
JOURNAL OF VERTEBRATE PALEONTOLOGY VOL 28 NO 2 2008456
Dow
nloa
ded
by [
UZ
H H
aupt
bibl
ioth
ek
Zen
tral
bibl
ioth
ek Z
uumlric
h] a
t 05
51 1
0 Se
ptem
ber
2013
FIGURE 12 Right pelvis of FMNH PR 2444 A stereopair of right pelvis in medial view B stereopair and interpretive drawing of right pelvis inlateral view Scale bars equal 5 cm Abbreviationsmdashbpp base of posterior process of ilium ib iliac blade IL ilium IS ischium ob f obturatorforamen P pubis
KAMMERER ET ALmdashCYNODONT FROM MADAGASCAR 457
Dow
nloa
ded
by [
UZ
H H
aupt
bibl
ioth
ek
Zen
tral
bibl
ioth
ek Z
uumlric
h] a
t 05
51 1
0 Se
ptem
ber
2013
aeretodon (Abdala et al 2002) The orbit of FMNH PR 2444 isless upwardly directed than that of the holotype (being at anangle of sim65deg to a sagittal plane through the midline of the skullas opposed to sim45deg in the holotype) This difference in orbitalangle is probably attributable to postmortem deformation as theskull of FMNH PR 2444 is noticeably sheared to the front leftThe appearance of a wider squarer incisor arcade and broader
anterior palate in FMNH PR 2444 can also be attributed to thisshear Finally the occipital region of FMNH PR 2444 is alto-gether taller and narrower than that of the holotype For ex-ample the foramen magnum in FMNH PR 2444 approximates arounded isosceles triangle in shape while the foramen magnumof the holotype is more ovoid These differences can also beattributed to deformation Altogether the differences between
FIGURE 13 Stereopairs of field number 8-31-98-387 an isolated right pelvis in A medial and B lateral view Scale bars equal 5 cm
JOURNAL OF VERTEBRATE PALEONTOLOGY VOL 28 NO 2 2008458
Dow
nloa
ded
by [
UZ
H H
aupt
bibl
ioth
ek
Zen
tral
bibl
ioth
ek Z
uumlric
h] a
t 05
51 1
0 Se
ptem
ber
2013
FMNH PR 2444 and the holotype fall within the range of intra-specific variation recorded for other gomphodont cynodontssuch as the traversodontids Massetognathus pascuali (Abdalaand Giannini 2000 Kammerer pers obs) and Exaeretodon(Abdala et al 2002) and the diademodontid Diademodon tetra-gonus (Grine et al 1978 Bradu and Grine 1979)
Phylogenetic Analysis
Utilizing the new information afforded by FMNH PR 2444and a reexamination of the holotype (UA 10601) Menadon be-sairiei was recoded in the Gondwanan gomphodont data matrixof Abdala et al (2006) Abdala et alrsquos (2006) data matrix en-compassed 18 taxa and 43 characters For the present analysis anadditional character ldquoincisor procumbencyrdquo (states 0-absent1-present) was added to Abdala et alrsquos (2006) data set (see
Appendix for coding of this character) Several traversodontidtaxa (eg lsquoScalenodonrsquo hirschsoni) have relatively large incisorsbut only in Menadon Scalenodontoides and Exaeretodon are theincisors procumbent Three of Abdala et alrsquos (2006) originalcodings for Menadon besairiei characters 0 (ldquonumber of upperincisorsrdquo) 5 (ldquoposition of paracanine fossa in relation to theupper caninerdquo) and 21 (ldquointernarial barrdquo) were changed to in-corporate data from the new specimen (see Appendix) Addi-tionally further examination of the holotype (UA 10601) ofMenadon besairiei allowed us to code characters 16ndash18 20 38and 40 (morphology of the lower postcanines all marked as ldquordquoin Abdala et alrsquos analysis) Parsimony searches were performedin PAUP v40b10 using a branch-and-bound search Addition-ally a bootstrap analysis (also using branch-and-bound) was per-formed using 1000 replicates
A total of 34 most parsimonious trees of length 103 werefound with the scores CI0573 RI0725 RC0415 andHI0427 The strict consensus tree (Fig 15A) did not recovera polytomy between Menadon Exaeretodon and Scalenodont-oides as in Abdala and Ribeiro (2003) or a (Menadon + Ex-aeretodon) sister-group relationship as in Abdala et al (2006)Rather Menadon was recovered as the sister taxon of the (Ex-aeretodon + Scalenodontoides) clade supporting Flynn et alrsquos(2000) original placement of this taxon There is strong bootstrapsupport (88) for Traversodontidae but within traversodontidsthe only strongly supported subclade is (Gomphodontosuchus +(Menadon + (Scalenodontoides + Exaeretodon))) with a boot-strap value of 80 Within this clade the (Menadon + (Scalen-odontoides + Exaeretodon)) grouping is supported by a boot-strap value of 83 and the (Scalenodontoides + Exaeretodon)grouping is supported by a bootstrap value of 94 (Fig 15B) Inaddition to the positioning of Menadon the strict consensus tree(Fig 15A) differs from the topology of Abdala et al (2006) inrecovering a sister-group relationship between Massetognathusand Santacruzodon rather than having Massetognathus andSantacruzodon as successive outgroups to the clade containingGomphodontosuchus Menadon Scalenodontoides and Ex-aeretodon The (Massetognathus + Santacruzodon) clade is diag-nosed by the absence of an anterolingual cusp in the upper post-canines denticulated incisor cutting margins and the presence ofan anterior cingulum in the upper postcanines
FIGURE 14 Reconstruction of the pelvis of Menadon besairiei basedon information from FMNH PR 2444 and field number 8-31-98-387Dotted area not preserved in either specimen reconstruction based onPascualgnathus (Bonaparte 1966) and Exaeretodon (Bonaparte 1963)
FIGURE 15 Cladogram of traversodontid relationships based on revised data set of Abdala et al (2006) showing A strict consensus tree and B50 majority rule tree Numbers at nodes of majority rule tree represent bootstrap values
KAMMERER ET ALmdashCYNODONT FROM MADAGASCAR 459
Dow
nloa
ded
by [
UZ
H H
aupt
bibl
ioth
ek
Zen
tral
bibl
ioth
ek Z
uumlric
h] a
t 05
51 1
0 Se
ptem
ber
2013
Relationships of Menadon besairiei
Menadon besairiei is the sister taxon to (Exaeretodon + Sca-lenodontoides) the latter clade diagnosed by the possession ofthree upper incisors a posteromedial position of the paracaninefossae relative to the canines the absence of an internarial barlarge canines a projected zygomatic process of the jugal and amaxilla that contributes to the margin of the subtemporal fenes-tra (the last three character states represent reversals) The clade(Menadon + (Exaeretodon + Scalenodontoides)) is diagnosed bythe possession of large procumbent incisors The presence offour incisors a primitive retention distinguishes Menadon be-sairiei from Exaeretodon Scalenodontoides and an unnamed tra-versodontid (ldquoTraversodontid Type 1rdquo) from the Santa MariaFormation of Brazil (Abdala et al 2001 Abdala and Ribeiro2002) Gomphodontosuchus is recovered as the sister taxon to(Menadon (Exaeretodon + Scalenodontoides)) as has beenfound in previous studies (Flynn et al 2000 Abdala and Ribeiro2003) The taxon name Gomphodontosuchinae Watson andRomer 1956 (ex Gomphodontosuchidae Watson and Romer1956 first given subfamilial rank in Lehman 1961) is availablefor this clade As circumscribed here Gomphodontosuchinaeconsists of the most inclusive clade containing Gomphodontosu-chus brasiliensis von Huene 1928264 but not Massetognathuspascuali Romer 19672
Menadon besairiei displays a mixture of primitive and derivedcharacters within traversodontids The internarial bar and fourincisors are primitive features whereas the enlargement and pro-cumbency of the incisors are derived features shared with Ex-aeretodon and Scalenodontoides The postcranium shows severalsimilarities to that of Exaeretodon (apparent exclusion of theprocoracoid from the glenoid some features of the pelvis tallcaudal neural spines) but in general displays a rather conserva-tive morphology observed throughout Gomphodontia The mor-phology of the scapulocoracoids is closer akin to diademodontidsor trirachodontids than Exaeretodon and the expanded thoracicrib of Menadon has only slight differences from those of trirach-odontids or early traversodontids like Luangwa The atlas-axiscomplex of Menadon is similar to that of the Early Triassic cyn-odont Cynognathus in possessing a concave axial spine with along posterior process Although most gomphodonts have beenreconstructed with a convex axial spine as in early mammals thisspine is broken in most of the best described gomphodont post-crania (eg Diademodon Brink 1955) and the lsquosaddle-shapedrsquoaxial morphology may be more widely distributed among cyn-odonts than previously realized The new specimens of Menadondescribed in this paper demonstrate that a generalized gompho-dont postcranial morphology was retained deep within the tra-versodontid tree and that the derived morphologies of either ofthe well-known taxa Massetognathus or Exaeretodon should notbe taken as representative of traversodontids in general
ACKNOWLEDGMENTS
This work would not have been possible without the continu-ing support and encouragement of our collaborative programwith the Deacutepartement de Paleacuteontologie et drsquoAnthropologie Bi-ologique Universiteacute drsquoAntananarivo Madagascar under the di-rection of Armand Rasoamiaramanana (Department Chairman)and Gisegravele Ravololonarivo Randria (Head of the PaleontologyLaboratories) We are extremely grateful to the National Geo-graphic Society for their long-term support of the fieldwork as-sociated with this project Additional financial or logistical sup-port for fieldwork was provided by The Field Museum the Uni-versity of California-Santa Barbara and the WWF (WorldwideFund for Nature Madagascar) We acknowledge the Ministegraverede LrsquoEneacutergie et des Mines for authorizing field efforts and for
continuing to facilitate our joint US-Madagascar explorationresearch and education program Steve Goodman and AsminaGandhi generously assisted us in innumerable ways includinghospitality nonpareil We especially thank R Andriantompoha-vana L Razafimanantsoa A Toto Volahy JA Rabarison AGandie and P Vavisaro for assistance in the field during theexpedition in which these new specimens were discovered BillSimpson and Gina Wesley-Hunt for their patient excavation ofthis important material Allison Smith Ray Vodden Lisa Berg-wall and Deborah Wagner for excellent preparation of a chal-lenging specimen Jennifer Botha and Elize Butler (NationalMuseum Bloemfontein) Michael Raath and Fernando Abdala(Bernard Price Institute) Thomas Kemp (Oxford UniversityMuseum of Natural History) Ray Symonds (University Museumof Zoology Cambridge) Bernard Battail (Museacuteum nationaldrsquoHistoire naturelle Paris) Michael Maisch (Universitaumlt Tuumlbin-gen) and Charles Schaff and Farish Jenkins (Museum of Com-parative Zoology Harvard) for access to specimens and AgustiacutenMartinelli Fernando Abdala and James Hopson for helpful re-views of an earlier version of the manuscript
LITERATURE CITED
Abdala F M C Barbarena and J Dornelles 2002 A new species of thetraversodontid cynodont Exaeretodon from the Santa Maria Forma-tion (MiddleLate Triassic) of Southern Brazil Journal of Verte-brate Paleontology 22313ndash325
Abdala F and N P Giannini 2000 Gomphodont cynodonts of theChantildeares Formation the analysis of an ontogenetic sequence Jour-nal of Vertebrate Paleontology 20501ndash506
Abdala F J Neveling and J Welman 2006 A new trirachodontidcynodont from the lower levels of the Burgersdorp Formation(Lower Triassic) of the Beaufort Group South Africa and the cla-distic relationships of Gondwanan gomphodonts Zoological Jour-nal of the Linnean Society 147383ndash413
Abdala F A M Ribeiro and C L Schultz 2001 A rich cynodont faunaof Santa Cruz do Sul Santa Maria Formation (Middle-Late Trias-sic) southern Brazil Neues Jahrbuch fuumlr Geologie und Palaumlontolo-gie Monatshefte 2001669ndash687
Abdala F and A M Ribeiro 2002 Nuevos cinodontes traversodoacuten-tidos (Synapsida-Eucynodontia) de la Formacioacuten Santa Maria(Triaacutesico Medio-Superior) Rio Grande do Sul Brasil RevistaEspantildeola de Paleontologiacutea 17237ndash243
Abdala F and A M Ribeiro 2003 A new traversodontid cynodontfrom the Santa Maria Formation (Ladinian-Carnian) of southernBrazil with a phylogenetic analysis of Gondwanan traversodontidsZoological Journal of the Linnaean Society 139529ndash545
Abdala F and A M S Teixeira 2004 A traversodontid cynodont ofAfrican affinity in the South American Triassic Palaeontologia Af-ricana 4011ndash22
Bonaparte J F 1962 Descripcioacuten del craacuteneo y mandiacutebula de Ex-aeretodon frenguellii Cabrera y su comparacioacuten con Diademodon-tidae Tritylodontidae y los cinodontes sudamericanos Publica-ciones del Museo Municipal de Ciencias Naturales y Tradicional deMar del Plata 1135ndash202
Bonaparte J F 1963 Descripcioacuten del esqueleto postcraneano de Ex-aeretodon (CynodontiandashTraversodontidae) Acta Geoloacutegica Lil-loana 45ndash52
Bonaparte J F 1966 Una nueva ldquofaunardquo Triaacutesica de Argentina (Ther-apsida Cynodontia Dicynodontia) Consideracions filogeneacuteticas ypaleobiogeograacuteficas Ameghiniana 8243ndash296
Bradu D and F E Grine 1979 Multivariate analysis of diademodon-tine crania from South Africa and Zambia South African Journal ofScience 79441ndash448
Brink A S 1955 A study of the skeleton of Diademodon Palaeonto-logia Africana 33-39
Brink A S 1963 Two new cynodonts from the Ntawere Formation inthe Luangwa Valley of Northern Rhodesia Palaeontologia Africana877ndash96
Broom R 1905 On the use of the term Anomodontia Records of theAlbany Museum 1266ndash269
Cabrera A 1943 El primer hallazgo de teraacutepsidos en la ArgentinaNotas del Museo de La Plata Paleontologiacutea 55317ndash331
JOURNAL OF VERTEBRATE PALEONTOLOGY VOL 28 NO 2 2008460
Dow
nloa
ded
by [
UZ
H H
aupt
bibl
ioth
ek
Zen
tral
bibl
ioth
ek Z
uumlric
h] a
t 05
51 1
0 Se
ptem
ber
2013
Chatterjee S 1982 A new cynodont reptile from the Triassic of IndiaJournal of Paleontology 56203ndash214
Crompton A W 1955 On some Triassic cynodonts from TanganyikaProceedings of the Zoological Society of London 125617ndash669
Crompton A W 1972 Postcanine occlusion in cynodonts and tritylo-donts Bulletin of the British Museum (Natural History) Geology2129ndash71
Crompton A W and F Ellenberger 1957 On a new cynodont from theMolteno Beds and the origin of the tritylodontids Annals of theSouth African Museum 441ndash13
Flynn J J J M Parrish B Rakotosaminimanana W F Simpson R LWhatley and A R Wyss 1999 A Triassic fauna from Madagascarincluding early dinosaurs Science 286763ndash765
Flynn J J J M Parrish B Rakotosamimanana L RanivoharimananaW F Simpson and A R Wyss 2000 New traversodontids (Syn-apsida Eucynodontia) from the Triassic of Madagascar Journal ofVertebrate Paleontology 20422ndash427
Gow C E and P J Hancox 1993 First complete skull of the LateTriassic Scalenodontoides (Reptilia Cynodontia) from southern Af-rica In Lucas S G and M Morales (eds) The nonmarine TriassicNew Mexico Museum of Natural History amp Science Bulletin 3161ndash168
Grine F E B D Hahn and C E Gow 1978 Aspects of relativegrowth and variability in Diademodon (Reptilia Therapsida) SouthAfrican Journal of Science 7450ndash58
Hopson J A 1984 Late Triassic traversodont cynodonts from NovaScotia and southern Africa Palaeontologia Africana 25181ndash201
Hopson J A 1985 Morphology and relationships of Gomphodontosu-chus brasiliensis von Huene (Synapsida Cynodontia Tritylodontoi-dea) from the Triassic of Brazil Neues Jahrbuch fuumlr Geologie undPalaumlontologie Monatshefte 1985285ndash299
Hopson J A and J W Kitching 2001 A probainognathian cynodontfrom South Africa and the phylogeny of nonmammalian cynodontsBulletin of the Museum of Comparative Zoology 1565ndash35
Hopson J A and H-D Sues 2006 A traversodont cynodont from theMiddle Triassic (Ladinian) of Baden-Wuumlrttemberg (Germany)Palaumlontologische Zeitschrift 80124ndash129
Huene F von 1928 Ein Cynodontier aus der Trias Brasiliens Zentral-blatt fuumlr Mineralogie Geologie und Palaumlontologie 1928251ndash270
Huene F von 1936 Die fossilen Reptilien des suumldamerikanischenGondwanalandes Ergebnisse der Sauriergrabungen in Suumldbrasilien192829 Lieferung 2 (Cynodontia)93ndash159 Franz F Heine Tuumlbin-gen
Jenkins F A 1970 The Chantildeares (Argentina) Triassic reptile faunaVII The postcranial skeleton of the traversodontid Massetognathuspascuali (Therapsida Cynodontia) Breviora 3521ndash28
Jenkins F A 1971 The postcranial skeleton of African cynodonts prob-lems in the early evolution of the mammalian postcranial skeletonBulletin of the Peabody Museum of Natural History 361ndash216
Kemp T S 1969 The atlas-axis complex of mammal-like reptiles Jour-nal of Zoology 159223ndash248
Kemp T S 1980 Aspects of the structure and functional anatomy of theMiddle Triassic cynodont Luangwa Journal of Zoology 191193ndash239
Kemp T S 1982 Mammal-Like Reptiles and the Origin of MammalsAcademic Press New York
Lehman J-P 1961 Cynodontia pp 140ndash191 in Piveteau J (ed) Traiteacutede Paleacuteontologie Masson et Cie Paris
Osborn 1903 On the primary division of the Reptilia into two sub-classes Synapsida and Diapsida Science 17275ndash276
Owen R 1861 Palaeontology or a systematic summary of extinct ani-mals and their geological relations 2nd Ed Edinburgh
Parrington F R 1934 On the cynodont genus Galesaurus with a noteon the functional significance of the changes in the evolution of thetheriodont skull Annals and Magazine of Natural History 10(13)38ndash67
Parrington F R 1946 On the cranial anatomy of cynodonts Proceed-ings of the Zoological Society of London 116181ndash197
Razafimbelo E 1987 Le basin de Morondava (Madagascar) Synthegravesegeacuteologique et structurale PhD dissertation University Louis Pas-teur Strasbourg France 189 pp
Reisz R R and H-D Sues 2000 Herbivory in Late Paleozoic andTriassic terrestrial vertebrates pp 9ndash41 in Sues H-D (ed) Evo-lution of Herbivory in Terrestrial Vertebrates Cambridge Univer-sity Press New York
Romer A S 1967 The Chantildeares (Argentina) Triassic reptile fauna IIITwo new gomphodonts Massetognathus pascuali and M terugiiBreviora 2641ndash25
Romer A S 1972 The Chantildeares (Argentina) Triassic reptile faunaXVII The Chantildeares gomphodonts Breviora 3961ndash9
Rougier G W J R Wible and J A Hopson 1992 Reconstruction ofthe cranial vessels in the Early Cretaceous mammal Vincelestes neu-quenianus implications for the evolution of the mammalian cranialvascular system Journal of Vertebrate Paleontology 12188ndash216
Rowe T 1988 Definition diagnosis and origin of Mammalia Journal ofVertebrate Paleontology 8241ndash264
Rowe T 1993 Early mammal phylogenetic systematics pp 129ndash145 inSzalay F S M J Novacek and M C McKenna (eds) MammalPhylogeny Vol 1 Mesozoic Differentiation MultituberculatesMonotremes Early Therians and Marsupials Springer-Verlag NewYork
Seeley H G 1894 Researches on the structure organization and clas-sification of the fossil Reptilia Part IX Section 3 On DiademodonPhilosophical Transactions of the Royal Society of London 1851029ndash1041
Seeley H G 1895 Researches on the structure organization and clas-sification of the fossil Reptilia Part IX Section 4 On the Gompho-dontia Philosophical Transactions of the Royal Society of London1861ndash57
Seeley H G 1895 Researches on the structure organization and clas-sification of the fossil Reptilia Part IX Section 5 On the skeletonin new Cynodontia from the Karroo rocks Philosophical Transac-tions of the Royal Society of London 18659ndash148
Sues H-D J A Hopson and N H Shubin 1992 Affinities of Sca-lenodontoides plemmyridon Hopson 1984 (Synapsida Cynodontia)from the Upper Triassic of Nova Scotia Journal of Vertebrate Pa-leontology 12168ndash171
Sues H-D and P E Olsen 1990 Triassic vertebrates of Gondwananaspect from the Richmond basin of Virginia Science 2491020ndash1023
Swofford D L 2003 PAUP v 40b10 Sinauer Associates Inc Pub-lishers Sunderland Massachusetts
Watson D M S and A S Romer 1956 A classification of therapsidreptiles Bulletin of the Museum of Comparative Zoology 11435ndash89
Submitted September 20 2007 accepted November 3 2007
KAMMERER ET ALmdashCYNODONT FROM MADAGASCAR 461
Dow
nloa
ded
by [
UZ
H H
aupt
bibl
ioth
ek
Zen
tral
bibl
ioth
ek Z
uumlric
h] a
t 05
51 1
0 Se
ptem
ber
2013
APPENDIX Data matrix used in the phylogenetic analysesThis is a revised version of the matrix of Abdala et al (2006) with the following changes character 43 [Incisor procumbency (0) absent (1) present]added Menadon recoded for characters 0 (Number of upper incisors was 1 ldquothreerdquo changed to 0 ldquofourrdquo) 5 (Position of paracanine fossae in relationto the upper canine was changed to 1 ldquomedialrdquo) 16 (Overall morphology of lower postcanines in occlusal view was changed to 3 ldquoquadran-gularrdquo) 17 (Position of the transverse crest in lower postcanines was changed to 1 ldquoanteriorrdquo) 18 (Number of cusps in the transverse crest of thelower postcanines was changed to 0 ldquotwordquo) 20 (Anterior cingulum in the lower postcanines was changed to 2 ldquoabsentrdquo) 21 (Internarial barwas 1 ldquoabsentrdquo changed to 0 ldquopresentrdquo) 38 (Posterior cingulum in the lower postcanines was changed to 1 ldquoabsentrdquo) and 40 (Deep occlusalbasins in the postcanines was changed to 1 ldquopresentrdquo) Langbergia recoded for character 34 (Incisor cutting margins was 0 ldquoserratedrdquo changedto 1 ldquosmoothly ridgedrdquo) based on the description of the incisors in Abdala et al (2006) Gomphodontosuchus recoded for character 31 (Maxilla inthe margin of the subtemporal fenestra was 1 ldquoincludedrdquo changed to 0 ldquoexcludedrdquo) based on von Huene (1936) and Kammerer (pers obs of theholotype) contra Hopsonrsquos (1985) reconstruction lsquoScalenodonrsquo hirschsoni recoded for character 39 (Anterior cingulum in the upper postcanines was1 ldquoabsentrdquo changed to 0 ldquopresentrdquo) based on Abdala (pers comm 2006) Trirachodon recoded for character 30 (Ectopterygoid was changedto 0 ldquopresentrdquo) based on Abdala (pers comm 2006) and Traversodon recoded for character 15 (Number of cusps in the sectorial border of the upperpostcanines was 0 ldquomore than twordquo changed to 1 ldquotwordquo) based on Abdala (pers comm 2006) and Kammerer (pers obs)
1111111111 2222222222 3333333333 44440123456789 0123456789 0123456789 0123456789 0123
Cynognathus 00000000 000 000000000 00000000 000Diademodon 0000001000 0000100 000010A000 0000000100 0100Trirachodon 000000100B 1A002201 001A100010 0001001200 0110Langbergia 0000001000 10000201 0000101010 0001101200 0110Cricodon 0100001000 11000201 0010100 0001200 0110Andescynodon 0000002001 01013101 101120010 0112311 1100Massetognathus 00111121A2 1110003100 2011200011 1011212300 1100Exaeretodon 1110122112 010103101 2111110201 1111112311 1101Luangwa 0000001002 11010A3100 101010010 01002300 1100Scal angustifrons 000000100B 1111023100 10102101 00002301 1100Scal hirschsoni 11110202 1100113100 1010 1112300 1100Traversodon 0000002012 1101113100 201010 011230 1100Gomphodontosuchus 0011112112 0103101 2010 0112311 1100Pascualgnathus 1000001000 000231 1100001 0011230 1100Scalenodontoides 1112212 01003101 21111021 1212311 1101Menadon 0111112112 01012310 20121020 0102311 1101Dadadon 00002112 111112 013 11231 1100Santacruzodon 00002112 1110003101 201300 212300 1100
A(01) B(02)
JOURNAL OF VERTEBRATE PALEONTOLOGY VOL 28 NO 2 2008462
Dow
nloa
ded
by [
UZ
H H
aupt
bibl
ioth
ek
Zen
tral
bibl
ioth
ek Z
uumlric
h] a
t 05
51 1
0 Se
ptem
ber
2013
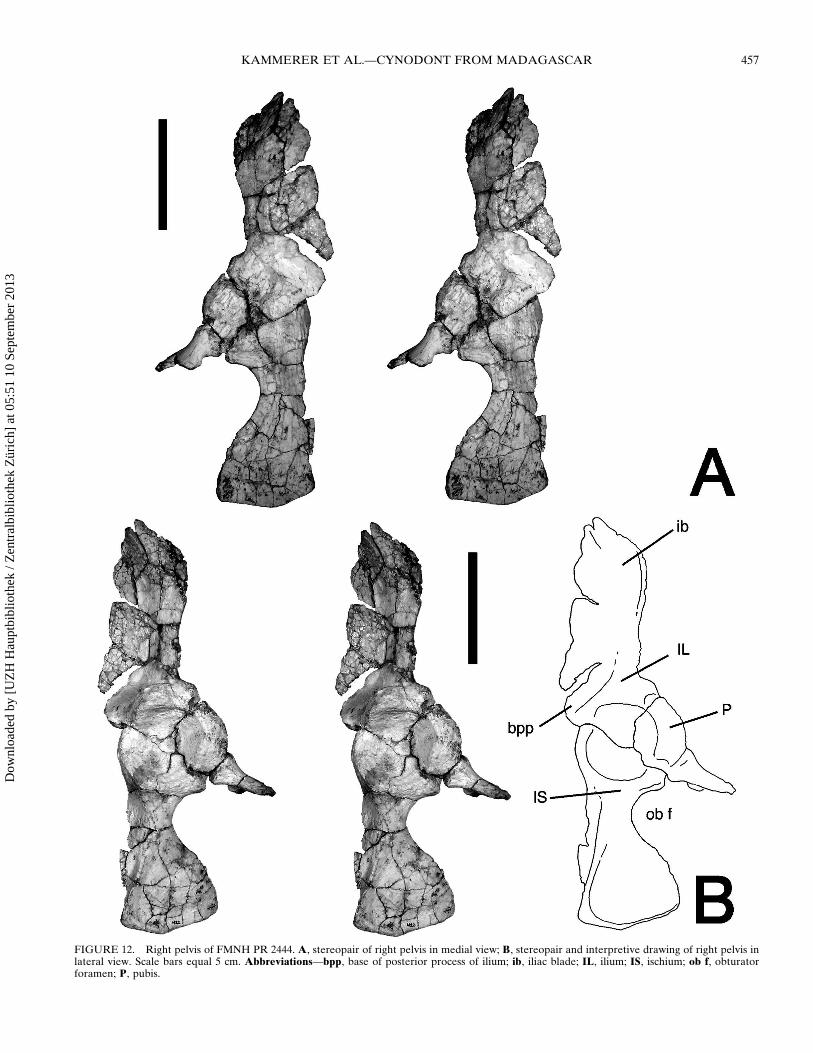
FIGURE 12 Right pelvis of FMNH PR 2444 A stereopair of right pelvis in medial view B stereopair and interpretive drawing of right pelvis inlateral view Scale bars equal 5 cm Abbreviationsmdashbpp base of posterior process of ilium ib iliac blade IL ilium IS ischium ob f obturatorforamen P pubis
KAMMERER ET ALmdashCYNODONT FROM MADAGASCAR 457
Dow
nloa
ded
by [
UZ
H H
aupt
bibl
ioth
ek
Zen
tral
bibl
ioth
ek Z
uumlric
h] a
t 05
51 1
0 Se
ptem
ber
2013
aeretodon (Abdala et al 2002) The orbit of FMNH PR 2444 isless upwardly directed than that of the holotype (being at anangle of sim65deg to a sagittal plane through the midline of the skullas opposed to sim45deg in the holotype) This difference in orbitalangle is probably attributable to postmortem deformation as theskull of FMNH PR 2444 is noticeably sheared to the front leftThe appearance of a wider squarer incisor arcade and broader
anterior palate in FMNH PR 2444 can also be attributed to thisshear Finally the occipital region of FMNH PR 2444 is alto-gether taller and narrower than that of the holotype For ex-ample the foramen magnum in FMNH PR 2444 approximates arounded isosceles triangle in shape while the foramen magnumof the holotype is more ovoid These differences can also beattributed to deformation Altogether the differences between
FIGURE 13 Stereopairs of field number 8-31-98-387 an isolated right pelvis in A medial and B lateral view Scale bars equal 5 cm
JOURNAL OF VERTEBRATE PALEONTOLOGY VOL 28 NO 2 2008458
Dow
nloa
ded
by [
UZ
H H
aupt
bibl
ioth
ek
Zen
tral
bibl
ioth
ek Z
uumlric
h] a
t 05
51 1
0 Se
ptem
ber
2013
FMNH PR 2444 and the holotype fall within the range of intra-specific variation recorded for other gomphodont cynodontssuch as the traversodontids Massetognathus pascuali (Abdalaand Giannini 2000 Kammerer pers obs) and Exaeretodon(Abdala et al 2002) and the diademodontid Diademodon tetra-gonus (Grine et al 1978 Bradu and Grine 1979)
Phylogenetic Analysis
Utilizing the new information afforded by FMNH PR 2444and a reexamination of the holotype (UA 10601) Menadon be-sairiei was recoded in the Gondwanan gomphodont data matrixof Abdala et al (2006) Abdala et alrsquos (2006) data matrix en-compassed 18 taxa and 43 characters For the present analysis anadditional character ldquoincisor procumbencyrdquo (states 0-absent1-present) was added to Abdala et alrsquos (2006) data set (see
Appendix for coding of this character) Several traversodontidtaxa (eg lsquoScalenodonrsquo hirschsoni) have relatively large incisorsbut only in Menadon Scalenodontoides and Exaeretodon are theincisors procumbent Three of Abdala et alrsquos (2006) originalcodings for Menadon besairiei characters 0 (ldquonumber of upperincisorsrdquo) 5 (ldquoposition of paracanine fossa in relation to theupper caninerdquo) and 21 (ldquointernarial barrdquo) were changed to in-corporate data from the new specimen (see Appendix) Addi-tionally further examination of the holotype (UA 10601) ofMenadon besairiei allowed us to code characters 16ndash18 20 38and 40 (morphology of the lower postcanines all marked as ldquordquoin Abdala et alrsquos analysis) Parsimony searches were performedin PAUP v40b10 using a branch-and-bound search Addition-ally a bootstrap analysis (also using branch-and-bound) was per-formed using 1000 replicates
A total of 34 most parsimonious trees of length 103 werefound with the scores CI0573 RI0725 RC0415 andHI0427 The strict consensus tree (Fig 15A) did not recovera polytomy between Menadon Exaeretodon and Scalenodont-oides as in Abdala and Ribeiro (2003) or a (Menadon + Ex-aeretodon) sister-group relationship as in Abdala et al (2006)Rather Menadon was recovered as the sister taxon of the (Ex-aeretodon + Scalenodontoides) clade supporting Flynn et alrsquos(2000) original placement of this taxon There is strong bootstrapsupport (88) for Traversodontidae but within traversodontidsthe only strongly supported subclade is (Gomphodontosuchus +(Menadon + (Scalenodontoides + Exaeretodon))) with a boot-strap value of 80 Within this clade the (Menadon + (Scalen-odontoides + Exaeretodon)) grouping is supported by a boot-strap value of 83 and the (Scalenodontoides + Exaeretodon)grouping is supported by a bootstrap value of 94 (Fig 15B) Inaddition to the positioning of Menadon the strict consensus tree(Fig 15A) differs from the topology of Abdala et al (2006) inrecovering a sister-group relationship between Massetognathusand Santacruzodon rather than having Massetognathus andSantacruzodon as successive outgroups to the clade containingGomphodontosuchus Menadon Scalenodontoides and Ex-aeretodon The (Massetognathus + Santacruzodon) clade is diag-nosed by the absence of an anterolingual cusp in the upper post-canines denticulated incisor cutting margins and the presence ofan anterior cingulum in the upper postcanines
FIGURE 14 Reconstruction of the pelvis of Menadon besairiei basedon information from FMNH PR 2444 and field number 8-31-98-387Dotted area not preserved in either specimen reconstruction based onPascualgnathus (Bonaparte 1966) and Exaeretodon (Bonaparte 1963)
FIGURE 15 Cladogram of traversodontid relationships based on revised data set of Abdala et al (2006) showing A strict consensus tree and B50 majority rule tree Numbers at nodes of majority rule tree represent bootstrap values
KAMMERER ET ALmdashCYNODONT FROM MADAGASCAR 459
Dow
nloa
ded
by [
UZ
H H
aupt
bibl
ioth
ek
Zen
tral
bibl
ioth
ek Z
uumlric
h] a
t 05
51 1
0 Se
ptem
ber
2013
Relationships of Menadon besairiei
Menadon besairiei is the sister taxon to (Exaeretodon + Sca-lenodontoides) the latter clade diagnosed by the possession ofthree upper incisors a posteromedial position of the paracaninefossae relative to the canines the absence of an internarial barlarge canines a projected zygomatic process of the jugal and amaxilla that contributes to the margin of the subtemporal fenes-tra (the last three character states represent reversals) The clade(Menadon + (Exaeretodon + Scalenodontoides)) is diagnosed bythe possession of large procumbent incisors The presence offour incisors a primitive retention distinguishes Menadon be-sairiei from Exaeretodon Scalenodontoides and an unnamed tra-versodontid (ldquoTraversodontid Type 1rdquo) from the Santa MariaFormation of Brazil (Abdala et al 2001 Abdala and Ribeiro2002) Gomphodontosuchus is recovered as the sister taxon to(Menadon (Exaeretodon + Scalenodontoides)) as has beenfound in previous studies (Flynn et al 2000 Abdala and Ribeiro2003) The taxon name Gomphodontosuchinae Watson andRomer 1956 (ex Gomphodontosuchidae Watson and Romer1956 first given subfamilial rank in Lehman 1961) is availablefor this clade As circumscribed here Gomphodontosuchinaeconsists of the most inclusive clade containing Gomphodontosu-chus brasiliensis von Huene 1928264 but not Massetognathuspascuali Romer 19672
Menadon besairiei displays a mixture of primitive and derivedcharacters within traversodontids The internarial bar and fourincisors are primitive features whereas the enlargement and pro-cumbency of the incisors are derived features shared with Ex-aeretodon and Scalenodontoides The postcranium shows severalsimilarities to that of Exaeretodon (apparent exclusion of theprocoracoid from the glenoid some features of the pelvis tallcaudal neural spines) but in general displays a rather conserva-tive morphology observed throughout Gomphodontia The mor-phology of the scapulocoracoids is closer akin to diademodontidsor trirachodontids than Exaeretodon and the expanded thoracicrib of Menadon has only slight differences from those of trirach-odontids or early traversodontids like Luangwa The atlas-axiscomplex of Menadon is similar to that of the Early Triassic cyn-odont Cynognathus in possessing a concave axial spine with along posterior process Although most gomphodonts have beenreconstructed with a convex axial spine as in early mammals thisspine is broken in most of the best described gomphodont post-crania (eg Diademodon Brink 1955) and the lsquosaddle-shapedrsquoaxial morphology may be more widely distributed among cyn-odonts than previously realized The new specimens of Menadondescribed in this paper demonstrate that a generalized gompho-dont postcranial morphology was retained deep within the tra-versodontid tree and that the derived morphologies of either ofthe well-known taxa Massetognathus or Exaeretodon should notbe taken as representative of traversodontids in general
ACKNOWLEDGMENTS
This work would not have been possible without the continu-ing support and encouragement of our collaborative programwith the Deacutepartement de Paleacuteontologie et drsquoAnthropologie Bi-ologique Universiteacute drsquoAntananarivo Madagascar under the di-rection of Armand Rasoamiaramanana (Department Chairman)and Gisegravele Ravololonarivo Randria (Head of the PaleontologyLaboratories) We are extremely grateful to the National Geo-graphic Society for their long-term support of the fieldwork as-sociated with this project Additional financial or logistical sup-port for fieldwork was provided by The Field Museum the Uni-versity of California-Santa Barbara and the WWF (WorldwideFund for Nature Madagascar) We acknowledge the Ministegraverede LrsquoEneacutergie et des Mines for authorizing field efforts and for
continuing to facilitate our joint US-Madagascar explorationresearch and education program Steve Goodman and AsminaGandhi generously assisted us in innumerable ways includinghospitality nonpareil We especially thank R Andriantompoha-vana L Razafimanantsoa A Toto Volahy JA Rabarison AGandie and P Vavisaro for assistance in the field during theexpedition in which these new specimens were discovered BillSimpson and Gina Wesley-Hunt for their patient excavation ofthis important material Allison Smith Ray Vodden Lisa Berg-wall and Deborah Wagner for excellent preparation of a chal-lenging specimen Jennifer Botha and Elize Butler (NationalMuseum Bloemfontein) Michael Raath and Fernando Abdala(Bernard Price Institute) Thomas Kemp (Oxford UniversityMuseum of Natural History) Ray Symonds (University Museumof Zoology Cambridge) Bernard Battail (Museacuteum nationaldrsquoHistoire naturelle Paris) Michael Maisch (Universitaumlt Tuumlbin-gen) and Charles Schaff and Farish Jenkins (Museum of Com-parative Zoology Harvard) for access to specimens and AgustiacutenMartinelli Fernando Abdala and James Hopson for helpful re-views of an earlier version of the manuscript
LITERATURE CITED
Abdala F M C Barbarena and J Dornelles 2002 A new species of thetraversodontid cynodont Exaeretodon from the Santa Maria Forma-tion (MiddleLate Triassic) of Southern Brazil Journal of Verte-brate Paleontology 22313ndash325
Abdala F and N P Giannini 2000 Gomphodont cynodonts of theChantildeares Formation the analysis of an ontogenetic sequence Jour-nal of Vertebrate Paleontology 20501ndash506
Abdala F J Neveling and J Welman 2006 A new trirachodontidcynodont from the lower levels of the Burgersdorp Formation(Lower Triassic) of the Beaufort Group South Africa and the cla-distic relationships of Gondwanan gomphodonts Zoological Jour-nal of the Linnean Society 147383ndash413
Abdala F A M Ribeiro and C L Schultz 2001 A rich cynodont faunaof Santa Cruz do Sul Santa Maria Formation (Middle-Late Trias-sic) southern Brazil Neues Jahrbuch fuumlr Geologie und Palaumlontolo-gie Monatshefte 2001669ndash687
Abdala F and A M Ribeiro 2002 Nuevos cinodontes traversodoacuten-tidos (Synapsida-Eucynodontia) de la Formacioacuten Santa Maria(Triaacutesico Medio-Superior) Rio Grande do Sul Brasil RevistaEspantildeola de Paleontologiacutea 17237ndash243
Abdala F and A M Ribeiro 2003 A new traversodontid cynodontfrom the Santa Maria Formation (Ladinian-Carnian) of southernBrazil with a phylogenetic analysis of Gondwanan traversodontidsZoological Journal of the Linnaean Society 139529ndash545
Abdala F and A M S Teixeira 2004 A traversodontid cynodont ofAfrican affinity in the South American Triassic Palaeontologia Af-ricana 4011ndash22
Bonaparte J F 1962 Descripcioacuten del craacuteneo y mandiacutebula de Ex-aeretodon frenguellii Cabrera y su comparacioacuten con Diademodon-tidae Tritylodontidae y los cinodontes sudamericanos Publica-ciones del Museo Municipal de Ciencias Naturales y Tradicional deMar del Plata 1135ndash202
Bonaparte J F 1963 Descripcioacuten del esqueleto postcraneano de Ex-aeretodon (CynodontiandashTraversodontidae) Acta Geoloacutegica Lil-loana 45ndash52
Bonaparte J F 1966 Una nueva ldquofaunardquo Triaacutesica de Argentina (Ther-apsida Cynodontia Dicynodontia) Consideracions filogeneacuteticas ypaleobiogeograacuteficas Ameghiniana 8243ndash296
Bradu D and F E Grine 1979 Multivariate analysis of diademodon-tine crania from South Africa and Zambia South African Journal ofScience 79441ndash448
Brink A S 1955 A study of the skeleton of Diademodon Palaeonto-logia Africana 33-39
Brink A S 1963 Two new cynodonts from the Ntawere Formation inthe Luangwa Valley of Northern Rhodesia Palaeontologia Africana877ndash96
Broom R 1905 On the use of the term Anomodontia Records of theAlbany Museum 1266ndash269
Cabrera A 1943 El primer hallazgo de teraacutepsidos en la ArgentinaNotas del Museo de La Plata Paleontologiacutea 55317ndash331
JOURNAL OF VERTEBRATE PALEONTOLOGY VOL 28 NO 2 2008460
Dow
nloa
ded
by [
UZ
H H
aupt
bibl
ioth
ek
Zen
tral
bibl
ioth
ek Z
uumlric
h] a
t 05
51 1
0 Se
ptem
ber
2013
Chatterjee S 1982 A new cynodont reptile from the Triassic of IndiaJournal of Paleontology 56203ndash214
Crompton A W 1955 On some Triassic cynodonts from TanganyikaProceedings of the Zoological Society of London 125617ndash669
Crompton A W 1972 Postcanine occlusion in cynodonts and tritylo-donts Bulletin of the British Museum (Natural History) Geology2129ndash71
Crompton A W and F Ellenberger 1957 On a new cynodont from theMolteno Beds and the origin of the tritylodontids Annals of theSouth African Museum 441ndash13
Flynn J J J M Parrish B Rakotosaminimanana W F Simpson R LWhatley and A R Wyss 1999 A Triassic fauna from Madagascarincluding early dinosaurs Science 286763ndash765
Flynn J J J M Parrish B Rakotosamimanana L RanivoharimananaW F Simpson and A R Wyss 2000 New traversodontids (Syn-apsida Eucynodontia) from the Triassic of Madagascar Journal ofVertebrate Paleontology 20422ndash427
Gow C E and P J Hancox 1993 First complete skull of the LateTriassic Scalenodontoides (Reptilia Cynodontia) from southern Af-rica In Lucas S G and M Morales (eds) The nonmarine TriassicNew Mexico Museum of Natural History amp Science Bulletin 3161ndash168
Grine F E B D Hahn and C E Gow 1978 Aspects of relativegrowth and variability in Diademodon (Reptilia Therapsida) SouthAfrican Journal of Science 7450ndash58
Hopson J A 1984 Late Triassic traversodont cynodonts from NovaScotia and southern Africa Palaeontologia Africana 25181ndash201
Hopson J A 1985 Morphology and relationships of Gomphodontosu-chus brasiliensis von Huene (Synapsida Cynodontia Tritylodontoi-dea) from the Triassic of Brazil Neues Jahrbuch fuumlr Geologie undPalaumlontologie Monatshefte 1985285ndash299
Hopson J A and J W Kitching 2001 A probainognathian cynodontfrom South Africa and the phylogeny of nonmammalian cynodontsBulletin of the Museum of Comparative Zoology 1565ndash35
Hopson J A and H-D Sues 2006 A traversodont cynodont from theMiddle Triassic (Ladinian) of Baden-Wuumlrttemberg (Germany)Palaumlontologische Zeitschrift 80124ndash129
Huene F von 1928 Ein Cynodontier aus der Trias Brasiliens Zentral-blatt fuumlr Mineralogie Geologie und Palaumlontologie 1928251ndash270
Huene F von 1936 Die fossilen Reptilien des suumldamerikanischenGondwanalandes Ergebnisse der Sauriergrabungen in Suumldbrasilien192829 Lieferung 2 (Cynodontia)93ndash159 Franz F Heine Tuumlbin-gen
Jenkins F A 1970 The Chantildeares (Argentina) Triassic reptile faunaVII The postcranial skeleton of the traversodontid Massetognathuspascuali (Therapsida Cynodontia) Breviora 3521ndash28
Jenkins F A 1971 The postcranial skeleton of African cynodonts prob-lems in the early evolution of the mammalian postcranial skeletonBulletin of the Peabody Museum of Natural History 361ndash216
Kemp T S 1969 The atlas-axis complex of mammal-like reptiles Jour-nal of Zoology 159223ndash248
Kemp T S 1980 Aspects of the structure and functional anatomy of theMiddle Triassic cynodont Luangwa Journal of Zoology 191193ndash239
Kemp T S 1982 Mammal-Like Reptiles and the Origin of MammalsAcademic Press New York
Lehman J-P 1961 Cynodontia pp 140ndash191 in Piveteau J (ed) Traiteacutede Paleacuteontologie Masson et Cie Paris
Osborn 1903 On the primary division of the Reptilia into two sub-classes Synapsida and Diapsida Science 17275ndash276
Owen R 1861 Palaeontology or a systematic summary of extinct ani-mals and their geological relations 2nd Ed Edinburgh
Parrington F R 1934 On the cynodont genus Galesaurus with a noteon the functional significance of the changes in the evolution of thetheriodont skull Annals and Magazine of Natural History 10(13)38ndash67
Parrington F R 1946 On the cranial anatomy of cynodonts Proceed-ings of the Zoological Society of London 116181ndash197
Razafimbelo E 1987 Le basin de Morondava (Madagascar) Synthegravesegeacuteologique et structurale PhD dissertation University Louis Pas-teur Strasbourg France 189 pp
Reisz R R and H-D Sues 2000 Herbivory in Late Paleozoic andTriassic terrestrial vertebrates pp 9ndash41 in Sues H-D (ed) Evo-lution of Herbivory in Terrestrial Vertebrates Cambridge Univer-sity Press New York
Romer A S 1967 The Chantildeares (Argentina) Triassic reptile fauna IIITwo new gomphodonts Massetognathus pascuali and M terugiiBreviora 2641ndash25
Romer A S 1972 The Chantildeares (Argentina) Triassic reptile faunaXVII The Chantildeares gomphodonts Breviora 3961ndash9
Rougier G W J R Wible and J A Hopson 1992 Reconstruction ofthe cranial vessels in the Early Cretaceous mammal Vincelestes neu-quenianus implications for the evolution of the mammalian cranialvascular system Journal of Vertebrate Paleontology 12188ndash216
Rowe T 1988 Definition diagnosis and origin of Mammalia Journal ofVertebrate Paleontology 8241ndash264
Rowe T 1993 Early mammal phylogenetic systematics pp 129ndash145 inSzalay F S M J Novacek and M C McKenna (eds) MammalPhylogeny Vol 1 Mesozoic Differentiation MultituberculatesMonotremes Early Therians and Marsupials Springer-Verlag NewYork
Seeley H G 1894 Researches on the structure organization and clas-sification of the fossil Reptilia Part IX Section 3 On DiademodonPhilosophical Transactions of the Royal Society of London 1851029ndash1041
Seeley H G 1895 Researches on the structure organization and clas-sification of the fossil Reptilia Part IX Section 4 On the Gompho-dontia Philosophical Transactions of the Royal Society of London1861ndash57
Seeley H G 1895 Researches on the structure organization and clas-sification of the fossil Reptilia Part IX Section 5 On the skeletonin new Cynodontia from the Karroo rocks Philosophical Transac-tions of the Royal Society of London 18659ndash148
Sues H-D J A Hopson and N H Shubin 1992 Affinities of Sca-lenodontoides plemmyridon Hopson 1984 (Synapsida Cynodontia)from the Upper Triassic of Nova Scotia Journal of Vertebrate Pa-leontology 12168ndash171
Sues H-D and P E Olsen 1990 Triassic vertebrates of Gondwananaspect from the Richmond basin of Virginia Science 2491020ndash1023
Swofford D L 2003 PAUP v 40b10 Sinauer Associates Inc Pub-lishers Sunderland Massachusetts
Watson D M S and A S Romer 1956 A classification of therapsidreptiles Bulletin of the Museum of Comparative Zoology 11435ndash89
Submitted September 20 2007 accepted November 3 2007
KAMMERER ET ALmdashCYNODONT FROM MADAGASCAR 461
Dow
nloa
ded
by [
UZ
H H
aupt
bibl
ioth
ek
Zen
tral
bibl
ioth
ek Z
uumlric
h] a
t 05
51 1
0 Se
ptem
ber
2013
APPENDIX Data matrix used in the phylogenetic analysesThis is a revised version of the matrix of Abdala et al (2006) with the following changes character 43 [Incisor procumbency (0) absent (1) present]added Menadon recoded for characters 0 (Number of upper incisors was 1 ldquothreerdquo changed to 0 ldquofourrdquo) 5 (Position of paracanine fossae in relationto the upper canine was changed to 1 ldquomedialrdquo) 16 (Overall morphology of lower postcanines in occlusal view was changed to 3 ldquoquadran-gularrdquo) 17 (Position of the transverse crest in lower postcanines was changed to 1 ldquoanteriorrdquo) 18 (Number of cusps in the transverse crest of thelower postcanines was changed to 0 ldquotwordquo) 20 (Anterior cingulum in the lower postcanines was changed to 2 ldquoabsentrdquo) 21 (Internarial barwas 1 ldquoabsentrdquo changed to 0 ldquopresentrdquo) 38 (Posterior cingulum in the lower postcanines was changed to 1 ldquoabsentrdquo) and 40 (Deep occlusalbasins in the postcanines was changed to 1 ldquopresentrdquo) Langbergia recoded for character 34 (Incisor cutting margins was 0 ldquoserratedrdquo changedto 1 ldquosmoothly ridgedrdquo) based on the description of the incisors in Abdala et al (2006) Gomphodontosuchus recoded for character 31 (Maxilla inthe margin of the subtemporal fenestra was 1 ldquoincludedrdquo changed to 0 ldquoexcludedrdquo) based on von Huene (1936) and Kammerer (pers obs of theholotype) contra Hopsonrsquos (1985) reconstruction lsquoScalenodonrsquo hirschsoni recoded for character 39 (Anterior cingulum in the upper postcanines was1 ldquoabsentrdquo changed to 0 ldquopresentrdquo) based on Abdala (pers comm 2006) Trirachodon recoded for character 30 (Ectopterygoid was changedto 0 ldquopresentrdquo) based on Abdala (pers comm 2006) and Traversodon recoded for character 15 (Number of cusps in the sectorial border of the upperpostcanines was 0 ldquomore than twordquo changed to 1 ldquotwordquo) based on Abdala (pers comm 2006) and Kammerer (pers obs)
1111111111 2222222222 3333333333 44440123456789 0123456789 0123456789 0123456789 0123
Cynognathus 00000000 000 000000000 00000000 000Diademodon 0000001000 0000100 000010A000 0000000100 0100Trirachodon 000000100B 1A002201 001A100010 0001001200 0110Langbergia 0000001000 10000201 0000101010 0001101200 0110Cricodon 0100001000 11000201 0010100 0001200 0110Andescynodon 0000002001 01013101 101120010 0112311 1100Massetognathus 00111121A2 1110003100 2011200011 1011212300 1100Exaeretodon 1110122112 010103101 2111110201 1111112311 1101Luangwa 0000001002 11010A3100 101010010 01002300 1100Scal angustifrons 000000100B 1111023100 10102101 00002301 1100Scal hirschsoni 11110202 1100113100 1010 1112300 1100Traversodon 0000002012 1101113100 201010 011230 1100Gomphodontosuchus 0011112112 0103101 2010 0112311 1100Pascualgnathus 1000001000 000231 1100001 0011230 1100Scalenodontoides 1112212 01003101 21111021 1212311 1101Menadon 0111112112 01012310 20121020 0102311 1101Dadadon 00002112 111112 013 11231 1100Santacruzodon 00002112 1110003101 201300 212300 1100
A(01) B(02)
JOURNAL OF VERTEBRATE PALEONTOLOGY VOL 28 NO 2 2008462
Dow
nloa
ded
by [
UZ
H H
aupt
bibl
ioth
ek
Zen
tral
bibl
ioth
ek Z
uumlric
h] a
t 05
51 1
0 Se
ptem
ber
2013
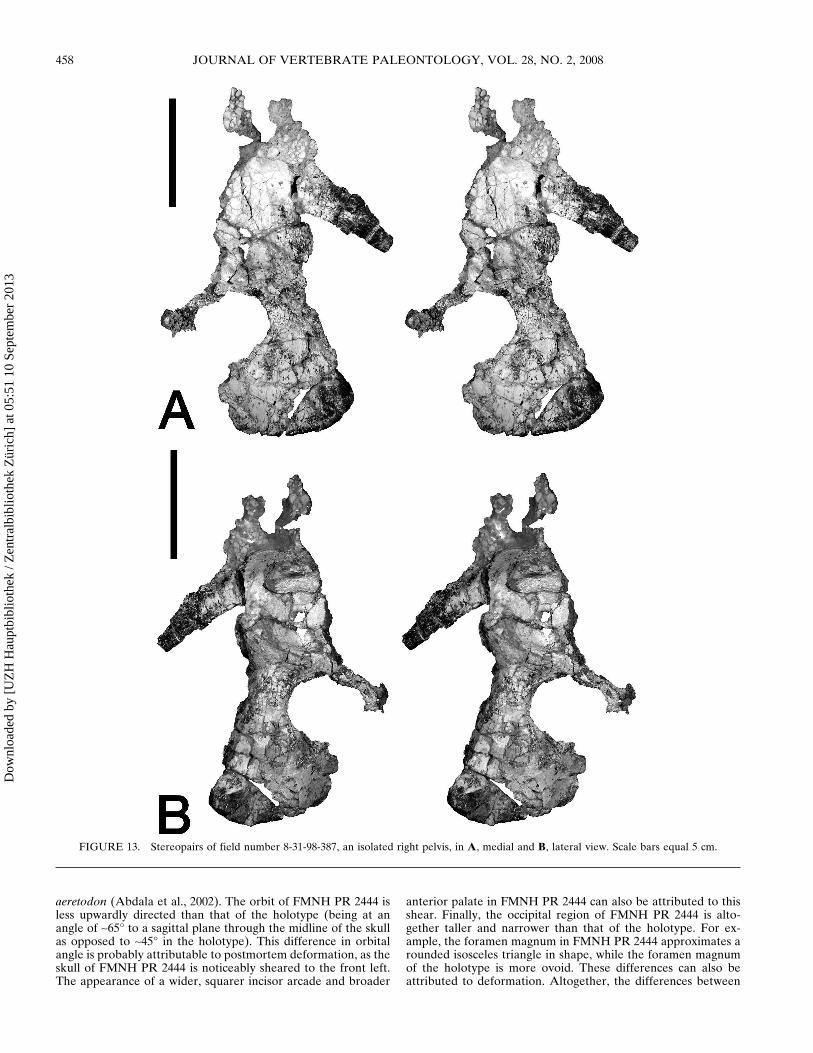
aeretodon (Abdala et al 2002) The orbit of FMNH PR 2444 isless upwardly directed than that of the holotype (being at anangle of sim65deg to a sagittal plane through the midline of the skullas opposed to sim45deg in the holotype) This difference in orbitalangle is probably attributable to postmortem deformation as theskull of FMNH PR 2444 is noticeably sheared to the front leftThe appearance of a wider squarer incisor arcade and broader
anterior palate in FMNH PR 2444 can also be attributed to thisshear Finally the occipital region of FMNH PR 2444 is alto-gether taller and narrower than that of the holotype For ex-ample the foramen magnum in FMNH PR 2444 approximates arounded isosceles triangle in shape while the foramen magnumof the holotype is more ovoid These differences can also beattributed to deformation Altogether the differences between
FIGURE 13 Stereopairs of field number 8-31-98-387 an isolated right pelvis in A medial and B lateral view Scale bars equal 5 cm
JOURNAL OF VERTEBRATE PALEONTOLOGY VOL 28 NO 2 2008458
Dow
nloa
ded
by [
UZ
H H
aupt
bibl
ioth
ek
Zen
tral
bibl
ioth
ek Z
uumlric
h] a
t 05
51 1
0 Se
ptem
ber
2013
FMNH PR 2444 and the holotype fall within the range of intra-specific variation recorded for other gomphodont cynodontssuch as the traversodontids Massetognathus pascuali (Abdalaand Giannini 2000 Kammerer pers obs) and Exaeretodon(Abdala et al 2002) and the diademodontid Diademodon tetra-gonus (Grine et al 1978 Bradu and Grine 1979)
Phylogenetic Analysis
Utilizing the new information afforded by FMNH PR 2444and a reexamination of the holotype (UA 10601) Menadon be-sairiei was recoded in the Gondwanan gomphodont data matrixof Abdala et al (2006) Abdala et alrsquos (2006) data matrix en-compassed 18 taxa and 43 characters For the present analysis anadditional character ldquoincisor procumbencyrdquo (states 0-absent1-present) was added to Abdala et alrsquos (2006) data set (see
Appendix for coding of this character) Several traversodontidtaxa (eg lsquoScalenodonrsquo hirschsoni) have relatively large incisorsbut only in Menadon Scalenodontoides and Exaeretodon are theincisors procumbent Three of Abdala et alrsquos (2006) originalcodings for Menadon besairiei characters 0 (ldquonumber of upperincisorsrdquo) 5 (ldquoposition of paracanine fossa in relation to theupper caninerdquo) and 21 (ldquointernarial barrdquo) were changed to in-corporate data from the new specimen (see Appendix) Addi-tionally further examination of the holotype (UA 10601) ofMenadon besairiei allowed us to code characters 16ndash18 20 38and 40 (morphology of the lower postcanines all marked as ldquordquoin Abdala et alrsquos analysis) Parsimony searches were performedin PAUP v40b10 using a branch-and-bound search Addition-ally a bootstrap analysis (also using branch-and-bound) was per-formed using 1000 replicates
A total of 34 most parsimonious trees of length 103 werefound with the scores CI0573 RI0725 RC0415 andHI0427 The strict consensus tree (Fig 15A) did not recovera polytomy between Menadon Exaeretodon and Scalenodont-oides as in Abdala and Ribeiro (2003) or a (Menadon + Ex-aeretodon) sister-group relationship as in Abdala et al (2006)Rather Menadon was recovered as the sister taxon of the (Ex-aeretodon + Scalenodontoides) clade supporting Flynn et alrsquos(2000) original placement of this taxon There is strong bootstrapsupport (88) for Traversodontidae but within traversodontidsthe only strongly supported subclade is (Gomphodontosuchus +(Menadon + (Scalenodontoides + Exaeretodon))) with a boot-strap value of 80 Within this clade the (Menadon + (Scalen-odontoides + Exaeretodon)) grouping is supported by a boot-strap value of 83 and the (Scalenodontoides + Exaeretodon)grouping is supported by a bootstrap value of 94 (Fig 15B) Inaddition to the positioning of Menadon the strict consensus tree(Fig 15A) differs from the topology of Abdala et al (2006) inrecovering a sister-group relationship between Massetognathusand Santacruzodon rather than having Massetognathus andSantacruzodon as successive outgroups to the clade containingGomphodontosuchus Menadon Scalenodontoides and Ex-aeretodon The (Massetognathus + Santacruzodon) clade is diag-nosed by the absence of an anterolingual cusp in the upper post-canines denticulated incisor cutting margins and the presence ofan anterior cingulum in the upper postcanines
FIGURE 14 Reconstruction of the pelvis of Menadon besairiei basedon information from FMNH PR 2444 and field number 8-31-98-387Dotted area not preserved in either specimen reconstruction based onPascualgnathus (Bonaparte 1966) and Exaeretodon (Bonaparte 1963)
FIGURE 15 Cladogram of traversodontid relationships based on revised data set of Abdala et al (2006) showing A strict consensus tree and B50 majority rule tree Numbers at nodes of majority rule tree represent bootstrap values
KAMMERER ET ALmdashCYNODONT FROM MADAGASCAR 459
Dow
nloa
ded
by [
UZ
H H
aupt
bibl
ioth
ek
Zen
tral
bibl
ioth
ek Z
uumlric
h] a
t 05
51 1
0 Se
ptem
ber
2013
Relationships of Menadon besairiei
Menadon besairiei is the sister taxon to (Exaeretodon + Sca-lenodontoides) the latter clade diagnosed by the possession ofthree upper incisors a posteromedial position of the paracaninefossae relative to the canines the absence of an internarial barlarge canines a projected zygomatic process of the jugal and amaxilla that contributes to the margin of the subtemporal fenes-tra (the last three character states represent reversals) The clade(Menadon + (Exaeretodon + Scalenodontoides)) is diagnosed bythe possession of large procumbent incisors The presence offour incisors a primitive retention distinguishes Menadon be-sairiei from Exaeretodon Scalenodontoides and an unnamed tra-versodontid (ldquoTraversodontid Type 1rdquo) from the Santa MariaFormation of Brazil (Abdala et al 2001 Abdala and Ribeiro2002) Gomphodontosuchus is recovered as the sister taxon to(Menadon (Exaeretodon + Scalenodontoides)) as has beenfound in previous studies (Flynn et al 2000 Abdala and Ribeiro2003) The taxon name Gomphodontosuchinae Watson andRomer 1956 (ex Gomphodontosuchidae Watson and Romer1956 first given subfamilial rank in Lehman 1961) is availablefor this clade As circumscribed here Gomphodontosuchinaeconsists of the most inclusive clade containing Gomphodontosu-chus brasiliensis von Huene 1928264 but not Massetognathuspascuali Romer 19672
Menadon besairiei displays a mixture of primitive and derivedcharacters within traversodontids The internarial bar and fourincisors are primitive features whereas the enlargement and pro-cumbency of the incisors are derived features shared with Ex-aeretodon and Scalenodontoides The postcranium shows severalsimilarities to that of Exaeretodon (apparent exclusion of theprocoracoid from the glenoid some features of the pelvis tallcaudal neural spines) but in general displays a rather conserva-tive morphology observed throughout Gomphodontia The mor-phology of the scapulocoracoids is closer akin to diademodontidsor trirachodontids than Exaeretodon and the expanded thoracicrib of Menadon has only slight differences from those of trirach-odontids or early traversodontids like Luangwa The atlas-axiscomplex of Menadon is similar to that of the Early Triassic cyn-odont Cynognathus in possessing a concave axial spine with along posterior process Although most gomphodonts have beenreconstructed with a convex axial spine as in early mammals thisspine is broken in most of the best described gomphodont post-crania (eg Diademodon Brink 1955) and the lsquosaddle-shapedrsquoaxial morphology may be more widely distributed among cyn-odonts than previously realized The new specimens of Menadondescribed in this paper demonstrate that a generalized gompho-dont postcranial morphology was retained deep within the tra-versodontid tree and that the derived morphologies of either ofthe well-known taxa Massetognathus or Exaeretodon should notbe taken as representative of traversodontids in general
ACKNOWLEDGMENTS
This work would not have been possible without the continu-ing support and encouragement of our collaborative programwith the Deacutepartement de Paleacuteontologie et drsquoAnthropologie Bi-ologique Universiteacute drsquoAntananarivo Madagascar under the di-rection of Armand Rasoamiaramanana (Department Chairman)and Gisegravele Ravololonarivo Randria (Head of the PaleontologyLaboratories) We are extremely grateful to the National Geo-graphic Society for their long-term support of the fieldwork as-sociated with this project Additional financial or logistical sup-port for fieldwork was provided by The Field Museum the Uni-versity of California-Santa Barbara and the WWF (WorldwideFund for Nature Madagascar) We acknowledge the Ministegraverede LrsquoEneacutergie et des Mines for authorizing field efforts and for
continuing to facilitate our joint US-Madagascar explorationresearch and education program Steve Goodman and AsminaGandhi generously assisted us in innumerable ways includinghospitality nonpareil We especially thank R Andriantompoha-vana L Razafimanantsoa A Toto Volahy JA Rabarison AGandie and P Vavisaro for assistance in the field during theexpedition in which these new specimens were discovered BillSimpson and Gina Wesley-Hunt for their patient excavation ofthis important material Allison Smith Ray Vodden Lisa Berg-wall and Deborah Wagner for excellent preparation of a chal-lenging specimen Jennifer Botha and Elize Butler (NationalMuseum Bloemfontein) Michael Raath and Fernando Abdala(Bernard Price Institute) Thomas Kemp (Oxford UniversityMuseum of Natural History) Ray Symonds (University Museumof Zoology Cambridge) Bernard Battail (Museacuteum nationaldrsquoHistoire naturelle Paris) Michael Maisch (Universitaumlt Tuumlbin-gen) and Charles Schaff and Farish Jenkins (Museum of Com-parative Zoology Harvard) for access to specimens and AgustiacutenMartinelli Fernando Abdala and James Hopson for helpful re-views of an earlier version of the manuscript
LITERATURE CITED
Abdala F M C Barbarena and J Dornelles 2002 A new species of thetraversodontid cynodont Exaeretodon from the Santa Maria Forma-tion (MiddleLate Triassic) of Southern Brazil Journal of Verte-brate Paleontology 22313ndash325
Abdala F and N P Giannini 2000 Gomphodont cynodonts of theChantildeares Formation the analysis of an ontogenetic sequence Jour-nal of Vertebrate Paleontology 20501ndash506
Abdala F J Neveling and J Welman 2006 A new trirachodontidcynodont from the lower levels of the Burgersdorp Formation(Lower Triassic) of the Beaufort Group South Africa and the cla-distic relationships of Gondwanan gomphodonts Zoological Jour-nal of the Linnean Society 147383ndash413
Abdala F A M Ribeiro and C L Schultz 2001 A rich cynodont faunaof Santa Cruz do Sul Santa Maria Formation (Middle-Late Trias-sic) southern Brazil Neues Jahrbuch fuumlr Geologie und Palaumlontolo-gie Monatshefte 2001669ndash687
Abdala F and A M Ribeiro 2002 Nuevos cinodontes traversodoacuten-tidos (Synapsida-Eucynodontia) de la Formacioacuten Santa Maria(Triaacutesico Medio-Superior) Rio Grande do Sul Brasil RevistaEspantildeola de Paleontologiacutea 17237ndash243
Abdala F and A M Ribeiro 2003 A new traversodontid cynodontfrom the Santa Maria Formation (Ladinian-Carnian) of southernBrazil with a phylogenetic analysis of Gondwanan traversodontidsZoological Journal of the Linnaean Society 139529ndash545
Abdala F and A M S Teixeira 2004 A traversodontid cynodont ofAfrican affinity in the South American Triassic Palaeontologia Af-ricana 4011ndash22
Bonaparte J F 1962 Descripcioacuten del craacuteneo y mandiacutebula de Ex-aeretodon frenguellii Cabrera y su comparacioacuten con Diademodon-tidae Tritylodontidae y los cinodontes sudamericanos Publica-ciones del Museo Municipal de Ciencias Naturales y Tradicional deMar del Plata 1135ndash202
Bonaparte J F 1963 Descripcioacuten del esqueleto postcraneano de Ex-aeretodon (CynodontiandashTraversodontidae) Acta Geoloacutegica Lil-loana 45ndash52
Bonaparte J F 1966 Una nueva ldquofaunardquo Triaacutesica de Argentina (Ther-apsida Cynodontia Dicynodontia) Consideracions filogeneacuteticas ypaleobiogeograacuteficas Ameghiniana 8243ndash296
Bradu D and F E Grine 1979 Multivariate analysis of diademodon-tine crania from South Africa and Zambia South African Journal ofScience 79441ndash448
Brink A S 1955 A study of the skeleton of Diademodon Palaeonto-logia Africana 33-39
Brink A S 1963 Two new cynodonts from the Ntawere Formation inthe Luangwa Valley of Northern Rhodesia Palaeontologia Africana877ndash96
Broom R 1905 On the use of the term Anomodontia Records of theAlbany Museum 1266ndash269
Cabrera A 1943 El primer hallazgo de teraacutepsidos en la ArgentinaNotas del Museo de La Plata Paleontologiacutea 55317ndash331
JOURNAL OF VERTEBRATE PALEONTOLOGY VOL 28 NO 2 2008460
Dow
nloa
ded
by [
UZ
H H
aupt
bibl
ioth
ek
Zen
tral
bibl
ioth
ek Z
uumlric
h] a
t 05
51 1
0 Se
ptem
ber
2013
Chatterjee S 1982 A new cynodont reptile from the Triassic of IndiaJournal of Paleontology 56203ndash214
Crompton A W 1955 On some Triassic cynodonts from TanganyikaProceedings of the Zoological Society of London 125617ndash669
Crompton A W 1972 Postcanine occlusion in cynodonts and tritylo-donts Bulletin of the British Museum (Natural History) Geology2129ndash71
Crompton A W and F Ellenberger 1957 On a new cynodont from theMolteno Beds and the origin of the tritylodontids Annals of theSouth African Museum 441ndash13
Flynn J J J M Parrish B Rakotosaminimanana W F Simpson R LWhatley and A R Wyss 1999 A Triassic fauna from Madagascarincluding early dinosaurs Science 286763ndash765
Flynn J J J M Parrish B Rakotosamimanana L RanivoharimananaW F Simpson and A R Wyss 2000 New traversodontids (Syn-apsida Eucynodontia) from the Triassic of Madagascar Journal ofVertebrate Paleontology 20422ndash427
Gow C E and P J Hancox 1993 First complete skull of the LateTriassic Scalenodontoides (Reptilia Cynodontia) from southern Af-rica In Lucas S G and M Morales (eds) The nonmarine TriassicNew Mexico Museum of Natural History amp Science Bulletin 3161ndash168
Grine F E B D Hahn and C E Gow 1978 Aspects of relativegrowth and variability in Diademodon (Reptilia Therapsida) SouthAfrican Journal of Science 7450ndash58
Hopson J A 1984 Late Triassic traversodont cynodonts from NovaScotia and southern Africa Palaeontologia Africana 25181ndash201
Hopson J A 1985 Morphology and relationships of Gomphodontosu-chus brasiliensis von Huene (Synapsida Cynodontia Tritylodontoi-dea) from the Triassic of Brazil Neues Jahrbuch fuumlr Geologie undPalaumlontologie Monatshefte 1985285ndash299
Hopson J A and J W Kitching 2001 A probainognathian cynodontfrom South Africa and the phylogeny of nonmammalian cynodontsBulletin of the Museum of Comparative Zoology 1565ndash35
Hopson J A and H-D Sues 2006 A traversodont cynodont from theMiddle Triassic (Ladinian) of Baden-Wuumlrttemberg (Germany)Palaumlontologische Zeitschrift 80124ndash129
Huene F von 1928 Ein Cynodontier aus der Trias Brasiliens Zentral-blatt fuumlr Mineralogie Geologie und Palaumlontologie 1928251ndash270
Huene F von 1936 Die fossilen Reptilien des suumldamerikanischenGondwanalandes Ergebnisse der Sauriergrabungen in Suumldbrasilien192829 Lieferung 2 (Cynodontia)93ndash159 Franz F Heine Tuumlbin-gen
Jenkins F A 1970 The Chantildeares (Argentina) Triassic reptile faunaVII The postcranial skeleton of the traversodontid Massetognathuspascuali (Therapsida Cynodontia) Breviora 3521ndash28
Jenkins F A 1971 The postcranial skeleton of African cynodonts prob-lems in the early evolution of the mammalian postcranial skeletonBulletin of the Peabody Museum of Natural History 361ndash216
Kemp T S 1969 The atlas-axis complex of mammal-like reptiles Jour-nal of Zoology 159223ndash248
Kemp T S 1980 Aspects of the structure and functional anatomy of theMiddle Triassic cynodont Luangwa Journal of Zoology 191193ndash239
Kemp T S 1982 Mammal-Like Reptiles and the Origin of MammalsAcademic Press New York
Lehman J-P 1961 Cynodontia pp 140ndash191 in Piveteau J (ed) Traiteacutede Paleacuteontologie Masson et Cie Paris
Osborn 1903 On the primary division of the Reptilia into two sub-classes Synapsida and Diapsida Science 17275ndash276
Owen R 1861 Palaeontology or a systematic summary of extinct ani-mals and their geological relations 2nd Ed Edinburgh
Parrington F R 1934 On the cynodont genus Galesaurus with a noteon the functional significance of the changes in the evolution of thetheriodont skull Annals and Magazine of Natural History 10(13)38ndash67
Parrington F R 1946 On the cranial anatomy of cynodonts Proceed-ings of the Zoological Society of London 116181ndash197
Razafimbelo E 1987 Le basin de Morondava (Madagascar) Synthegravesegeacuteologique et structurale PhD dissertation University Louis Pas-teur Strasbourg France 189 pp
Reisz R R and H-D Sues 2000 Herbivory in Late Paleozoic andTriassic terrestrial vertebrates pp 9ndash41 in Sues H-D (ed) Evo-lution of Herbivory in Terrestrial Vertebrates Cambridge Univer-sity Press New York
Romer A S 1967 The Chantildeares (Argentina) Triassic reptile fauna IIITwo new gomphodonts Massetognathus pascuali and M terugiiBreviora 2641ndash25
Romer A S 1972 The Chantildeares (Argentina) Triassic reptile faunaXVII The Chantildeares gomphodonts Breviora 3961ndash9
Rougier G W J R Wible and J A Hopson 1992 Reconstruction ofthe cranial vessels in the Early Cretaceous mammal Vincelestes neu-quenianus implications for the evolution of the mammalian cranialvascular system Journal of Vertebrate Paleontology 12188ndash216
Rowe T 1988 Definition diagnosis and origin of Mammalia Journal ofVertebrate Paleontology 8241ndash264
Rowe T 1993 Early mammal phylogenetic systematics pp 129ndash145 inSzalay F S M J Novacek and M C McKenna (eds) MammalPhylogeny Vol 1 Mesozoic Differentiation MultituberculatesMonotremes Early Therians and Marsupials Springer-Verlag NewYork
Seeley H G 1894 Researches on the structure organization and clas-sification of the fossil Reptilia Part IX Section 3 On DiademodonPhilosophical Transactions of the Royal Society of London 1851029ndash1041
Seeley H G 1895 Researches on the structure organization and clas-sification of the fossil Reptilia Part IX Section 4 On the Gompho-dontia Philosophical Transactions of the Royal Society of London1861ndash57
Seeley H G 1895 Researches on the structure organization and clas-sification of the fossil Reptilia Part IX Section 5 On the skeletonin new Cynodontia from the Karroo rocks Philosophical Transac-tions of the Royal Society of London 18659ndash148
Sues H-D J A Hopson and N H Shubin 1992 Affinities of Sca-lenodontoides plemmyridon Hopson 1984 (Synapsida Cynodontia)from the Upper Triassic of Nova Scotia Journal of Vertebrate Pa-leontology 12168ndash171
Sues H-D and P E Olsen 1990 Triassic vertebrates of Gondwananaspect from the Richmond basin of Virginia Science 2491020ndash1023
Swofford D L 2003 PAUP v 40b10 Sinauer Associates Inc Pub-lishers Sunderland Massachusetts
Watson D M S and A S Romer 1956 A classification of therapsidreptiles Bulletin of the Museum of Comparative Zoology 11435ndash89
Submitted September 20 2007 accepted November 3 2007
KAMMERER ET ALmdashCYNODONT FROM MADAGASCAR 461
Dow
nloa
ded
by [
UZ
H H
aupt
bibl
ioth
ek
Zen
tral
bibl
ioth
ek Z
uumlric
h] a
t 05
51 1
0 Se
ptem
ber
2013
APPENDIX Data matrix used in the phylogenetic analysesThis is a revised version of the matrix of Abdala et al (2006) with the following changes character 43 [Incisor procumbency (0) absent (1) present]added Menadon recoded for characters 0 (Number of upper incisors was 1 ldquothreerdquo changed to 0 ldquofourrdquo) 5 (Position of paracanine fossae in relationto the upper canine was changed to 1 ldquomedialrdquo) 16 (Overall morphology of lower postcanines in occlusal view was changed to 3 ldquoquadran-gularrdquo) 17 (Position of the transverse crest in lower postcanines was changed to 1 ldquoanteriorrdquo) 18 (Number of cusps in the transverse crest of thelower postcanines was changed to 0 ldquotwordquo) 20 (Anterior cingulum in the lower postcanines was changed to 2 ldquoabsentrdquo) 21 (Internarial barwas 1 ldquoabsentrdquo changed to 0 ldquopresentrdquo) 38 (Posterior cingulum in the lower postcanines was changed to 1 ldquoabsentrdquo) and 40 (Deep occlusalbasins in the postcanines was changed to 1 ldquopresentrdquo) Langbergia recoded for character 34 (Incisor cutting margins was 0 ldquoserratedrdquo changedto 1 ldquosmoothly ridgedrdquo) based on the description of the incisors in Abdala et al (2006) Gomphodontosuchus recoded for character 31 (Maxilla inthe margin of the subtemporal fenestra was 1 ldquoincludedrdquo changed to 0 ldquoexcludedrdquo) based on von Huene (1936) and Kammerer (pers obs of theholotype) contra Hopsonrsquos (1985) reconstruction lsquoScalenodonrsquo hirschsoni recoded for character 39 (Anterior cingulum in the upper postcanines was1 ldquoabsentrdquo changed to 0 ldquopresentrdquo) based on Abdala (pers comm 2006) Trirachodon recoded for character 30 (Ectopterygoid was changedto 0 ldquopresentrdquo) based on Abdala (pers comm 2006) and Traversodon recoded for character 15 (Number of cusps in the sectorial border of the upperpostcanines was 0 ldquomore than twordquo changed to 1 ldquotwordquo) based on Abdala (pers comm 2006) and Kammerer (pers obs)
1111111111 2222222222 3333333333 44440123456789 0123456789 0123456789 0123456789 0123
Cynognathus 00000000 000 000000000 00000000 000Diademodon 0000001000 0000100 000010A000 0000000100 0100Trirachodon 000000100B 1A002201 001A100010 0001001200 0110Langbergia 0000001000 10000201 0000101010 0001101200 0110Cricodon 0100001000 11000201 0010100 0001200 0110Andescynodon 0000002001 01013101 101120010 0112311 1100Massetognathus 00111121A2 1110003100 2011200011 1011212300 1100Exaeretodon 1110122112 010103101 2111110201 1111112311 1101Luangwa 0000001002 11010A3100 101010010 01002300 1100Scal angustifrons 000000100B 1111023100 10102101 00002301 1100Scal hirschsoni 11110202 1100113100 1010 1112300 1100Traversodon 0000002012 1101113100 201010 011230 1100Gomphodontosuchus 0011112112 0103101 2010 0112311 1100Pascualgnathus 1000001000 000231 1100001 0011230 1100Scalenodontoides 1112212 01003101 21111021 1212311 1101Menadon 0111112112 01012310 20121020 0102311 1101Dadadon 00002112 111112 013 11231 1100Santacruzodon 00002112 1110003101 201300 212300 1100
A(01) B(02)
JOURNAL OF VERTEBRATE PALEONTOLOGY VOL 28 NO 2 2008462
Dow
nloa
ded
by [
UZ
H H
aupt
bibl
ioth
ek
Zen
tral
bibl
ioth
ek Z
uumlric
h] a
t 05
51 1
0 Se
ptem
ber
2013

FMNH PR 2444 and the holotype fall within the range of intra-specific variation recorded for other gomphodont cynodontssuch as the traversodontids Massetognathus pascuali (Abdalaand Giannini 2000 Kammerer pers obs) and Exaeretodon(Abdala et al 2002) and the diademodontid Diademodon tetra-gonus (Grine et al 1978 Bradu and Grine 1979)
Phylogenetic Analysis
Utilizing the new information afforded by FMNH PR 2444and a reexamination of the holotype (UA 10601) Menadon be-sairiei was recoded in the Gondwanan gomphodont data matrixof Abdala et al (2006) Abdala et alrsquos (2006) data matrix en-compassed 18 taxa and 43 characters For the present analysis anadditional character ldquoincisor procumbencyrdquo (states 0-absent1-present) was added to Abdala et alrsquos (2006) data set (see
Appendix for coding of this character) Several traversodontidtaxa (eg lsquoScalenodonrsquo hirschsoni) have relatively large incisorsbut only in Menadon Scalenodontoides and Exaeretodon are theincisors procumbent Three of Abdala et alrsquos (2006) originalcodings for Menadon besairiei characters 0 (ldquonumber of upperincisorsrdquo) 5 (ldquoposition of paracanine fossa in relation to theupper caninerdquo) and 21 (ldquointernarial barrdquo) were changed to in-corporate data from the new specimen (see Appendix) Addi-tionally further examination of the holotype (UA 10601) ofMenadon besairiei allowed us to code characters 16ndash18 20 38and 40 (morphology of the lower postcanines all marked as ldquordquoin Abdala et alrsquos analysis) Parsimony searches were performedin PAUP v40b10 using a branch-and-bound search Addition-ally a bootstrap analysis (also using branch-and-bound) was per-formed using 1000 replicates
A total of 34 most parsimonious trees of length 103 werefound with the scores CI0573 RI0725 RC0415 andHI0427 The strict consensus tree (Fig 15A) did not recovera polytomy between Menadon Exaeretodon and Scalenodont-oides as in Abdala and Ribeiro (2003) or a (Menadon + Ex-aeretodon) sister-group relationship as in Abdala et al (2006)Rather Menadon was recovered as the sister taxon of the (Ex-aeretodon + Scalenodontoides) clade supporting Flynn et alrsquos(2000) original placement of this taxon There is strong bootstrapsupport (88) for Traversodontidae but within traversodontidsthe only strongly supported subclade is (Gomphodontosuchus +(Menadon + (Scalenodontoides + Exaeretodon))) with a boot-strap value of 80 Within this clade the (Menadon + (Scalen-odontoides + Exaeretodon)) grouping is supported by a boot-strap value of 83 and the (Scalenodontoides + Exaeretodon)grouping is supported by a bootstrap value of 94 (Fig 15B) Inaddition to the positioning of Menadon the strict consensus tree(Fig 15A) differs from the topology of Abdala et al (2006) inrecovering a sister-group relationship between Massetognathusand Santacruzodon rather than having Massetognathus andSantacruzodon as successive outgroups to the clade containingGomphodontosuchus Menadon Scalenodontoides and Ex-aeretodon The (Massetognathus + Santacruzodon) clade is diag-nosed by the absence of an anterolingual cusp in the upper post-canines denticulated incisor cutting margins and the presence ofan anterior cingulum in the upper postcanines
FIGURE 14 Reconstruction of the pelvis of Menadon besairiei basedon information from FMNH PR 2444 and field number 8-31-98-387Dotted area not preserved in either specimen reconstruction based onPascualgnathus (Bonaparte 1966) and Exaeretodon (Bonaparte 1963)
FIGURE 15 Cladogram of traversodontid relationships based on revised data set of Abdala et al (2006) showing A strict consensus tree and B50 majority rule tree Numbers at nodes of majority rule tree represent bootstrap values
KAMMERER ET ALmdashCYNODONT FROM MADAGASCAR 459
Dow
nloa
ded
by [
UZ
H H
aupt
bibl
ioth
ek
Zen
tral
bibl
ioth
ek Z
uumlric
h] a
t 05
51 1
0 Se
ptem
ber
2013
Relationships of Menadon besairiei
Menadon besairiei is the sister taxon to (Exaeretodon + Sca-lenodontoides) the latter clade diagnosed by the possession ofthree upper incisors a posteromedial position of the paracaninefossae relative to the canines the absence of an internarial barlarge canines a projected zygomatic process of the jugal and amaxilla that contributes to the margin of the subtemporal fenes-tra (the last three character states represent reversals) The clade(Menadon + (Exaeretodon + Scalenodontoides)) is diagnosed bythe possession of large procumbent incisors The presence offour incisors a primitive retention distinguishes Menadon be-sairiei from Exaeretodon Scalenodontoides and an unnamed tra-versodontid (ldquoTraversodontid Type 1rdquo) from the Santa MariaFormation of Brazil (Abdala et al 2001 Abdala and Ribeiro2002) Gomphodontosuchus is recovered as the sister taxon to(Menadon (Exaeretodon + Scalenodontoides)) as has beenfound in previous studies (Flynn et al 2000 Abdala and Ribeiro2003) The taxon name Gomphodontosuchinae Watson andRomer 1956 (ex Gomphodontosuchidae Watson and Romer1956 first given subfamilial rank in Lehman 1961) is availablefor this clade As circumscribed here Gomphodontosuchinaeconsists of the most inclusive clade containing Gomphodontosu-chus brasiliensis von Huene 1928264 but not Massetognathuspascuali Romer 19672
Menadon besairiei displays a mixture of primitive and derivedcharacters within traversodontids The internarial bar and fourincisors are primitive features whereas the enlargement and pro-cumbency of the incisors are derived features shared with Ex-aeretodon and Scalenodontoides The postcranium shows severalsimilarities to that of Exaeretodon (apparent exclusion of theprocoracoid from the glenoid some features of the pelvis tallcaudal neural spines) but in general displays a rather conserva-tive morphology observed throughout Gomphodontia The mor-phology of the scapulocoracoids is closer akin to diademodontidsor trirachodontids than Exaeretodon and the expanded thoracicrib of Menadon has only slight differences from those of trirach-odontids or early traversodontids like Luangwa The atlas-axiscomplex of Menadon is similar to that of the Early Triassic cyn-odont Cynognathus in possessing a concave axial spine with along posterior process Although most gomphodonts have beenreconstructed with a convex axial spine as in early mammals thisspine is broken in most of the best described gomphodont post-crania (eg Diademodon Brink 1955) and the lsquosaddle-shapedrsquoaxial morphology may be more widely distributed among cyn-odonts than previously realized The new specimens of Menadondescribed in this paper demonstrate that a generalized gompho-dont postcranial morphology was retained deep within the tra-versodontid tree and that the derived morphologies of either ofthe well-known taxa Massetognathus or Exaeretodon should notbe taken as representative of traversodontids in general
ACKNOWLEDGMENTS
This work would not have been possible without the continu-ing support and encouragement of our collaborative programwith the Deacutepartement de Paleacuteontologie et drsquoAnthropologie Bi-ologique Universiteacute drsquoAntananarivo Madagascar under the di-rection of Armand Rasoamiaramanana (Department Chairman)and Gisegravele Ravololonarivo Randria (Head of the PaleontologyLaboratories) We are extremely grateful to the National Geo-graphic Society for their long-term support of the fieldwork as-sociated with this project Additional financial or logistical sup-port for fieldwork was provided by The Field Museum the Uni-versity of California-Santa Barbara and the WWF (WorldwideFund for Nature Madagascar) We acknowledge the Ministegraverede LrsquoEneacutergie et des Mines for authorizing field efforts and for
continuing to facilitate our joint US-Madagascar explorationresearch and education program Steve Goodman and AsminaGandhi generously assisted us in innumerable ways includinghospitality nonpareil We especially thank R Andriantompoha-vana L Razafimanantsoa A Toto Volahy JA Rabarison AGandie and P Vavisaro for assistance in the field during theexpedition in which these new specimens were discovered BillSimpson and Gina Wesley-Hunt for their patient excavation ofthis important material Allison Smith Ray Vodden Lisa Berg-wall and Deborah Wagner for excellent preparation of a chal-lenging specimen Jennifer Botha and Elize Butler (NationalMuseum Bloemfontein) Michael Raath and Fernando Abdala(Bernard Price Institute) Thomas Kemp (Oxford UniversityMuseum of Natural History) Ray Symonds (University Museumof Zoology Cambridge) Bernard Battail (Museacuteum nationaldrsquoHistoire naturelle Paris) Michael Maisch (Universitaumlt Tuumlbin-gen) and Charles Schaff and Farish Jenkins (Museum of Com-parative Zoology Harvard) for access to specimens and AgustiacutenMartinelli Fernando Abdala and James Hopson for helpful re-views of an earlier version of the manuscript
LITERATURE CITED
Abdala F M C Barbarena and J Dornelles 2002 A new species of thetraversodontid cynodont Exaeretodon from the Santa Maria Forma-tion (MiddleLate Triassic) of Southern Brazil Journal of Verte-brate Paleontology 22313ndash325
Abdala F and N P Giannini 2000 Gomphodont cynodonts of theChantildeares Formation the analysis of an ontogenetic sequence Jour-nal of Vertebrate Paleontology 20501ndash506
Abdala F J Neveling and J Welman 2006 A new trirachodontidcynodont from the lower levels of the Burgersdorp Formation(Lower Triassic) of the Beaufort Group South Africa and the cla-distic relationships of Gondwanan gomphodonts Zoological Jour-nal of the Linnean Society 147383ndash413
Abdala F A M Ribeiro and C L Schultz 2001 A rich cynodont faunaof Santa Cruz do Sul Santa Maria Formation (Middle-Late Trias-sic) southern Brazil Neues Jahrbuch fuumlr Geologie und Palaumlontolo-gie Monatshefte 2001669ndash687
Abdala F and A M Ribeiro 2002 Nuevos cinodontes traversodoacuten-tidos (Synapsida-Eucynodontia) de la Formacioacuten Santa Maria(Triaacutesico Medio-Superior) Rio Grande do Sul Brasil RevistaEspantildeola de Paleontologiacutea 17237ndash243
Abdala F and A M Ribeiro 2003 A new traversodontid cynodontfrom the Santa Maria Formation (Ladinian-Carnian) of southernBrazil with a phylogenetic analysis of Gondwanan traversodontidsZoological Journal of the Linnaean Society 139529ndash545
Abdala F and A M S Teixeira 2004 A traversodontid cynodont ofAfrican affinity in the South American Triassic Palaeontologia Af-ricana 4011ndash22
Bonaparte J F 1962 Descripcioacuten del craacuteneo y mandiacutebula de Ex-aeretodon frenguellii Cabrera y su comparacioacuten con Diademodon-tidae Tritylodontidae y los cinodontes sudamericanos Publica-ciones del Museo Municipal de Ciencias Naturales y Tradicional deMar del Plata 1135ndash202
Bonaparte J F 1963 Descripcioacuten del esqueleto postcraneano de Ex-aeretodon (CynodontiandashTraversodontidae) Acta Geoloacutegica Lil-loana 45ndash52
Bonaparte J F 1966 Una nueva ldquofaunardquo Triaacutesica de Argentina (Ther-apsida Cynodontia Dicynodontia) Consideracions filogeneacuteticas ypaleobiogeograacuteficas Ameghiniana 8243ndash296
Bradu D and F E Grine 1979 Multivariate analysis of diademodon-tine crania from South Africa and Zambia South African Journal ofScience 79441ndash448
Brink A S 1955 A study of the skeleton of Diademodon Palaeonto-logia Africana 33-39
Brink A S 1963 Two new cynodonts from the Ntawere Formation inthe Luangwa Valley of Northern Rhodesia Palaeontologia Africana877ndash96
Broom R 1905 On the use of the term Anomodontia Records of theAlbany Museum 1266ndash269
Cabrera A 1943 El primer hallazgo de teraacutepsidos en la ArgentinaNotas del Museo de La Plata Paleontologiacutea 55317ndash331
JOURNAL OF VERTEBRATE PALEONTOLOGY VOL 28 NO 2 2008460
Dow
nloa
ded
by [
UZ
H H
aupt
bibl
ioth
ek
Zen
tral
bibl
ioth
ek Z
uumlric
h] a
t 05
51 1
0 Se
ptem
ber
2013
Chatterjee S 1982 A new cynodont reptile from the Triassic of IndiaJournal of Paleontology 56203ndash214
Crompton A W 1955 On some Triassic cynodonts from TanganyikaProceedings of the Zoological Society of London 125617ndash669
Crompton A W 1972 Postcanine occlusion in cynodonts and tritylo-donts Bulletin of the British Museum (Natural History) Geology2129ndash71
Crompton A W and F Ellenberger 1957 On a new cynodont from theMolteno Beds and the origin of the tritylodontids Annals of theSouth African Museum 441ndash13
Flynn J J J M Parrish B Rakotosaminimanana W F Simpson R LWhatley and A R Wyss 1999 A Triassic fauna from Madagascarincluding early dinosaurs Science 286763ndash765
Flynn J J J M Parrish B Rakotosamimanana L RanivoharimananaW F Simpson and A R Wyss 2000 New traversodontids (Syn-apsida Eucynodontia) from the Triassic of Madagascar Journal ofVertebrate Paleontology 20422ndash427
Gow C E and P J Hancox 1993 First complete skull of the LateTriassic Scalenodontoides (Reptilia Cynodontia) from southern Af-rica In Lucas S G and M Morales (eds) The nonmarine TriassicNew Mexico Museum of Natural History amp Science Bulletin 3161ndash168
Grine F E B D Hahn and C E Gow 1978 Aspects of relativegrowth and variability in Diademodon (Reptilia Therapsida) SouthAfrican Journal of Science 7450ndash58
Hopson J A 1984 Late Triassic traversodont cynodonts from NovaScotia and southern Africa Palaeontologia Africana 25181ndash201
Hopson J A 1985 Morphology and relationships of Gomphodontosu-chus brasiliensis von Huene (Synapsida Cynodontia Tritylodontoi-dea) from the Triassic of Brazil Neues Jahrbuch fuumlr Geologie undPalaumlontologie Monatshefte 1985285ndash299
Hopson J A and J W Kitching 2001 A probainognathian cynodontfrom South Africa and the phylogeny of nonmammalian cynodontsBulletin of the Museum of Comparative Zoology 1565ndash35
Hopson J A and H-D Sues 2006 A traversodont cynodont from theMiddle Triassic (Ladinian) of Baden-Wuumlrttemberg (Germany)Palaumlontologische Zeitschrift 80124ndash129
Huene F von 1928 Ein Cynodontier aus der Trias Brasiliens Zentral-blatt fuumlr Mineralogie Geologie und Palaumlontologie 1928251ndash270
Huene F von 1936 Die fossilen Reptilien des suumldamerikanischenGondwanalandes Ergebnisse der Sauriergrabungen in Suumldbrasilien192829 Lieferung 2 (Cynodontia)93ndash159 Franz F Heine Tuumlbin-gen
Jenkins F A 1970 The Chantildeares (Argentina) Triassic reptile faunaVII The postcranial skeleton of the traversodontid Massetognathuspascuali (Therapsida Cynodontia) Breviora 3521ndash28
Jenkins F A 1971 The postcranial skeleton of African cynodonts prob-lems in the early evolution of the mammalian postcranial skeletonBulletin of the Peabody Museum of Natural History 361ndash216
Kemp T S 1969 The atlas-axis complex of mammal-like reptiles Jour-nal of Zoology 159223ndash248
Kemp T S 1980 Aspects of the structure and functional anatomy of theMiddle Triassic cynodont Luangwa Journal of Zoology 191193ndash239
Kemp T S 1982 Mammal-Like Reptiles and the Origin of MammalsAcademic Press New York
Lehman J-P 1961 Cynodontia pp 140ndash191 in Piveteau J (ed) Traiteacutede Paleacuteontologie Masson et Cie Paris
Osborn 1903 On the primary division of the Reptilia into two sub-classes Synapsida and Diapsida Science 17275ndash276
Owen R 1861 Palaeontology or a systematic summary of extinct ani-mals and their geological relations 2nd Ed Edinburgh
Parrington F R 1934 On the cynodont genus Galesaurus with a noteon the functional significance of the changes in the evolution of thetheriodont skull Annals and Magazine of Natural History 10(13)38ndash67
Parrington F R 1946 On the cranial anatomy of cynodonts Proceed-ings of the Zoological Society of London 116181ndash197
Razafimbelo E 1987 Le basin de Morondava (Madagascar) Synthegravesegeacuteologique et structurale PhD dissertation University Louis Pas-teur Strasbourg France 189 pp
Reisz R R and H-D Sues 2000 Herbivory in Late Paleozoic andTriassic terrestrial vertebrates pp 9ndash41 in Sues H-D (ed) Evo-lution of Herbivory in Terrestrial Vertebrates Cambridge Univer-sity Press New York
Romer A S 1967 The Chantildeares (Argentina) Triassic reptile fauna IIITwo new gomphodonts Massetognathus pascuali and M terugiiBreviora 2641ndash25
Romer A S 1972 The Chantildeares (Argentina) Triassic reptile faunaXVII The Chantildeares gomphodonts Breviora 3961ndash9
Rougier G W J R Wible and J A Hopson 1992 Reconstruction ofthe cranial vessels in the Early Cretaceous mammal Vincelestes neu-quenianus implications for the evolution of the mammalian cranialvascular system Journal of Vertebrate Paleontology 12188ndash216
Rowe T 1988 Definition diagnosis and origin of Mammalia Journal ofVertebrate Paleontology 8241ndash264
Rowe T 1993 Early mammal phylogenetic systematics pp 129ndash145 inSzalay F S M J Novacek and M C McKenna (eds) MammalPhylogeny Vol 1 Mesozoic Differentiation MultituberculatesMonotremes Early Therians and Marsupials Springer-Verlag NewYork
Seeley H G 1894 Researches on the structure organization and clas-sification of the fossil Reptilia Part IX Section 3 On DiademodonPhilosophical Transactions of the Royal Society of London 1851029ndash1041
Seeley H G 1895 Researches on the structure organization and clas-sification of the fossil Reptilia Part IX Section 4 On the Gompho-dontia Philosophical Transactions of the Royal Society of London1861ndash57
Seeley H G 1895 Researches on the structure organization and clas-sification of the fossil Reptilia Part IX Section 5 On the skeletonin new Cynodontia from the Karroo rocks Philosophical Transac-tions of the Royal Society of London 18659ndash148
Sues H-D J A Hopson and N H Shubin 1992 Affinities of Sca-lenodontoides plemmyridon Hopson 1984 (Synapsida Cynodontia)from the Upper Triassic of Nova Scotia Journal of Vertebrate Pa-leontology 12168ndash171
Sues H-D and P E Olsen 1990 Triassic vertebrates of Gondwananaspect from the Richmond basin of Virginia Science 2491020ndash1023
Swofford D L 2003 PAUP v 40b10 Sinauer Associates Inc Pub-lishers Sunderland Massachusetts
Watson D M S and A S Romer 1956 A classification of therapsidreptiles Bulletin of the Museum of Comparative Zoology 11435ndash89
Submitted September 20 2007 accepted November 3 2007
KAMMERER ET ALmdashCYNODONT FROM MADAGASCAR 461
Dow
nloa
ded
by [
UZ
H H
aupt
bibl
ioth
ek
Zen
tral
bibl
ioth
ek Z
uumlric
h] a
t 05
51 1
0 Se
ptem
ber
2013
APPENDIX Data matrix used in the phylogenetic analysesThis is a revised version of the matrix of Abdala et al (2006) with the following changes character 43 [Incisor procumbency (0) absent (1) present]added Menadon recoded for characters 0 (Number of upper incisors was 1 ldquothreerdquo changed to 0 ldquofourrdquo) 5 (Position of paracanine fossae in relationto the upper canine was changed to 1 ldquomedialrdquo) 16 (Overall morphology of lower postcanines in occlusal view was changed to 3 ldquoquadran-gularrdquo) 17 (Position of the transverse crest in lower postcanines was changed to 1 ldquoanteriorrdquo) 18 (Number of cusps in the transverse crest of thelower postcanines was changed to 0 ldquotwordquo) 20 (Anterior cingulum in the lower postcanines was changed to 2 ldquoabsentrdquo) 21 (Internarial barwas 1 ldquoabsentrdquo changed to 0 ldquopresentrdquo) 38 (Posterior cingulum in the lower postcanines was changed to 1 ldquoabsentrdquo) and 40 (Deep occlusalbasins in the postcanines was changed to 1 ldquopresentrdquo) Langbergia recoded for character 34 (Incisor cutting margins was 0 ldquoserratedrdquo changedto 1 ldquosmoothly ridgedrdquo) based on the description of the incisors in Abdala et al (2006) Gomphodontosuchus recoded for character 31 (Maxilla inthe margin of the subtemporal fenestra was 1 ldquoincludedrdquo changed to 0 ldquoexcludedrdquo) based on von Huene (1936) and Kammerer (pers obs of theholotype) contra Hopsonrsquos (1985) reconstruction lsquoScalenodonrsquo hirschsoni recoded for character 39 (Anterior cingulum in the upper postcanines was1 ldquoabsentrdquo changed to 0 ldquopresentrdquo) based on Abdala (pers comm 2006) Trirachodon recoded for character 30 (Ectopterygoid was changedto 0 ldquopresentrdquo) based on Abdala (pers comm 2006) and Traversodon recoded for character 15 (Number of cusps in the sectorial border of the upperpostcanines was 0 ldquomore than twordquo changed to 1 ldquotwordquo) based on Abdala (pers comm 2006) and Kammerer (pers obs)
1111111111 2222222222 3333333333 44440123456789 0123456789 0123456789 0123456789 0123
Cynognathus 00000000 000 000000000 00000000 000Diademodon 0000001000 0000100 000010A000 0000000100 0100Trirachodon 000000100B 1A002201 001A100010 0001001200 0110Langbergia 0000001000 10000201 0000101010 0001101200 0110Cricodon 0100001000 11000201 0010100 0001200 0110Andescynodon 0000002001 01013101 101120010 0112311 1100Massetognathus 00111121A2 1110003100 2011200011 1011212300 1100Exaeretodon 1110122112 010103101 2111110201 1111112311 1101Luangwa 0000001002 11010A3100 101010010 01002300 1100Scal angustifrons 000000100B 1111023100 10102101 00002301 1100Scal hirschsoni 11110202 1100113100 1010 1112300 1100Traversodon 0000002012 1101113100 201010 011230 1100Gomphodontosuchus 0011112112 0103101 2010 0112311 1100Pascualgnathus 1000001000 000231 1100001 0011230 1100Scalenodontoides 1112212 01003101 21111021 1212311 1101Menadon 0111112112 01012310 20121020 0102311 1101Dadadon 00002112 111112 013 11231 1100Santacruzodon 00002112 1110003101 201300 212300 1100
A(01) B(02)
JOURNAL OF VERTEBRATE PALEONTOLOGY VOL 28 NO 2 2008462
Dow
nloa
ded
by [
UZ
H H
aupt
bibl
ioth
ek
Zen
tral
bibl
ioth
ek Z
uumlric
h] a
t 05
51 1
0 Se
ptem
ber
2013

Relationships of Menadon besairiei
Menadon besairiei is the sister taxon to (Exaeretodon + Sca-lenodontoides) the latter clade diagnosed by the possession ofthree upper incisors a posteromedial position of the paracaninefossae relative to the canines the absence of an internarial barlarge canines a projected zygomatic process of the jugal and amaxilla that contributes to the margin of the subtemporal fenes-tra (the last three character states represent reversals) The clade(Menadon + (Exaeretodon + Scalenodontoides)) is diagnosed bythe possession of large procumbent incisors The presence offour incisors a primitive retention distinguishes Menadon be-sairiei from Exaeretodon Scalenodontoides and an unnamed tra-versodontid (ldquoTraversodontid Type 1rdquo) from the Santa MariaFormation of Brazil (Abdala et al 2001 Abdala and Ribeiro2002) Gomphodontosuchus is recovered as the sister taxon to(Menadon (Exaeretodon + Scalenodontoides)) as has beenfound in previous studies (Flynn et al 2000 Abdala and Ribeiro2003) The taxon name Gomphodontosuchinae Watson andRomer 1956 (ex Gomphodontosuchidae Watson and Romer1956 first given subfamilial rank in Lehman 1961) is availablefor this clade As circumscribed here Gomphodontosuchinaeconsists of the most inclusive clade containing Gomphodontosu-chus brasiliensis von Huene 1928264 but not Massetognathuspascuali Romer 19672
Menadon besairiei displays a mixture of primitive and derivedcharacters within traversodontids The internarial bar and fourincisors are primitive features whereas the enlargement and pro-cumbency of the incisors are derived features shared with Ex-aeretodon and Scalenodontoides The postcranium shows severalsimilarities to that of Exaeretodon (apparent exclusion of theprocoracoid from the glenoid some features of the pelvis tallcaudal neural spines) but in general displays a rather conserva-tive morphology observed throughout Gomphodontia The mor-phology of the scapulocoracoids is closer akin to diademodontidsor trirachodontids than Exaeretodon and the expanded thoracicrib of Menadon has only slight differences from those of trirach-odontids or early traversodontids like Luangwa The atlas-axiscomplex of Menadon is similar to that of the Early Triassic cyn-odont Cynognathus in possessing a concave axial spine with along posterior process Although most gomphodonts have beenreconstructed with a convex axial spine as in early mammals thisspine is broken in most of the best described gomphodont post-crania (eg Diademodon Brink 1955) and the lsquosaddle-shapedrsquoaxial morphology may be more widely distributed among cyn-odonts than previously realized The new specimens of Menadondescribed in this paper demonstrate that a generalized gompho-dont postcranial morphology was retained deep within the tra-versodontid tree and that the derived morphologies of either ofthe well-known taxa Massetognathus or Exaeretodon should notbe taken as representative of traversodontids in general
ACKNOWLEDGMENTS
This work would not have been possible without the continu-ing support and encouragement of our collaborative programwith the Deacutepartement de Paleacuteontologie et drsquoAnthropologie Bi-ologique Universiteacute drsquoAntananarivo Madagascar under the di-rection of Armand Rasoamiaramanana (Department Chairman)and Gisegravele Ravololonarivo Randria (Head of the PaleontologyLaboratories) We are extremely grateful to the National Geo-graphic Society for their long-term support of the fieldwork as-sociated with this project Additional financial or logistical sup-port for fieldwork was provided by The Field Museum the Uni-versity of California-Santa Barbara and the WWF (WorldwideFund for Nature Madagascar) We acknowledge the Ministegraverede LrsquoEneacutergie et des Mines for authorizing field efforts and for
continuing to facilitate our joint US-Madagascar explorationresearch and education program Steve Goodman and AsminaGandhi generously assisted us in innumerable ways includinghospitality nonpareil We especially thank R Andriantompoha-vana L Razafimanantsoa A Toto Volahy JA Rabarison AGandie and P Vavisaro for assistance in the field during theexpedition in which these new specimens were discovered BillSimpson and Gina Wesley-Hunt for their patient excavation ofthis important material Allison Smith Ray Vodden Lisa Berg-wall and Deborah Wagner for excellent preparation of a chal-lenging specimen Jennifer Botha and Elize Butler (NationalMuseum Bloemfontein) Michael Raath and Fernando Abdala(Bernard Price Institute) Thomas Kemp (Oxford UniversityMuseum of Natural History) Ray Symonds (University Museumof Zoology Cambridge) Bernard Battail (Museacuteum nationaldrsquoHistoire naturelle Paris) Michael Maisch (Universitaumlt Tuumlbin-gen) and Charles Schaff and Farish Jenkins (Museum of Com-parative Zoology Harvard) for access to specimens and AgustiacutenMartinelli Fernando Abdala and James Hopson for helpful re-views of an earlier version of the manuscript
LITERATURE CITED
Abdala F M C Barbarena and J Dornelles 2002 A new species of thetraversodontid cynodont Exaeretodon from the Santa Maria Forma-tion (MiddleLate Triassic) of Southern Brazil Journal of Verte-brate Paleontology 22313ndash325
Abdala F and N P Giannini 2000 Gomphodont cynodonts of theChantildeares Formation the analysis of an ontogenetic sequence Jour-nal of Vertebrate Paleontology 20501ndash506
Abdala F J Neveling and J Welman 2006 A new trirachodontidcynodont from the lower levels of the Burgersdorp Formation(Lower Triassic) of the Beaufort Group South Africa and the cla-distic relationships of Gondwanan gomphodonts Zoological Jour-nal of the Linnean Society 147383ndash413
Abdala F A M Ribeiro and C L Schultz 2001 A rich cynodont faunaof Santa Cruz do Sul Santa Maria Formation (Middle-Late Trias-sic) southern Brazil Neues Jahrbuch fuumlr Geologie und Palaumlontolo-gie Monatshefte 2001669ndash687
Abdala F and A M Ribeiro 2002 Nuevos cinodontes traversodoacuten-tidos (Synapsida-Eucynodontia) de la Formacioacuten Santa Maria(Triaacutesico Medio-Superior) Rio Grande do Sul Brasil RevistaEspantildeola de Paleontologiacutea 17237ndash243
Abdala F and A M Ribeiro 2003 A new traversodontid cynodontfrom the Santa Maria Formation (Ladinian-Carnian) of southernBrazil with a phylogenetic analysis of Gondwanan traversodontidsZoological Journal of the Linnaean Society 139529ndash545
Abdala F and A M S Teixeira 2004 A traversodontid cynodont ofAfrican affinity in the South American Triassic Palaeontologia Af-ricana 4011ndash22
Bonaparte J F 1962 Descripcioacuten del craacuteneo y mandiacutebula de Ex-aeretodon frenguellii Cabrera y su comparacioacuten con Diademodon-tidae Tritylodontidae y los cinodontes sudamericanos Publica-ciones del Museo Municipal de Ciencias Naturales y Tradicional deMar del Plata 1135ndash202
Bonaparte J F 1963 Descripcioacuten del esqueleto postcraneano de Ex-aeretodon (CynodontiandashTraversodontidae) Acta Geoloacutegica Lil-loana 45ndash52
Bonaparte J F 1966 Una nueva ldquofaunardquo Triaacutesica de Argentina (Ther-apsida Cynodontia Dicynodontia) Consideracions filogeneacuteticas ypaleobiogeograacuteficas Ameghiniana 8243ndash296
Bradu D and F E Grine 1979 Multivariate analysis of diademodon-tine crania from South Africa and Zambia South African Journal ofScience 79441ndash448
Brink A S 1955 A study of the skeleton of Diademodon Palaeonto-logia Africana 33-39
Brink A S 1963 Two new cynodonts from the Ntawere Formation inthe Luangwa Valley of Northern Rhodesia Palaeontologia Africana877ndash96
Broom R 1905 On the use of the term Anomodontia Records of theAlbany Museum 1266ndash269
Cabrera A 1943 El primer hallazgo de teraacutepsidos en la ArgentinaNotas del Museo de La Plata Paleontologiacutea 55317ndash331
JOURNAL OF VERTEBRATE PALEONTOLOGY VOL 28 NO 2 2008460
Dow
nloa
ded
by [
UZ
H H
aupt
bibl
ioth
ek
Zen
tral
bibl
ioth
ek Z
uumlric
h] a
t 05
51 1
0 Se
ptem
ber
2013
Chatterjee S 1982 A new cynodont reptile from the Triassic of IndiaJournal of Paleontology 56203ndash214
Crompton A W 1955 On some Triassic cynodonts from TanganyikaProceedings of the Zoological Society of London 125617ndash669
Crompton A W 1972 Postcanine occlusion in cynodonts and tritylo-donts Bulletin of the British Museum (Natural History) Geology2129ndash71
Crompton A W and F Ellenberger 1957 On a new cynodont from theMolteno Beds and the origin of the tritylodontids Annals of theSouth African Museum 441ndash13
Flynn J J J M Parrish B Rakotosaminimanana W F Simpson R LWhatley and A R Wyss 1999 A Triassic fauna from Madagascarincluding early dinosaurs Science 286763ndash765
Flynn J J J M Parrish B Rakotosamimanana L RanivoharimananaW F Simpson and A R Wyss 2000 New traversodontids (Syn-apsida Eucynodontia) from the Triassic of Madagascar Journal ofVertebrate Paleontology 20422ndash427
Gow C E and P J Hancox 1993 First complete skull of the LateTriassic Scalenodontoides (Reptilia Cynodontia) from southern Af-rica In Lucas S G and M Morales (eds) The nonmarine TriassicNew Mexico Museum of Natural History amp Science Bulletin 3161ndash168
Grine F E B D Hahn and C E Gow 1978 Aspects of relativegrowth and variability in Diademodon (Reptilia Therapsida) SouthAfrican Journal of Science 7450ndash58
Hopson J A 1984 Late Triassic traversodont cynodonts from NovaScotia and southern Africa Palaeontologia Africana 25181ndash201
Hopson J A 1985 Morphology and relationships of Gomphodontosu-chus brasiliensis von Huene (Synapsida Cynodontia Tritylodontoi-dea) from the Triassic of Brazil Neues Jahrbuch fuumlr Geologie undPalaumlontologie Monatshefte 1985285ndash299
Hopson J A and J W Kitching 2001 A probainognathian cynodontfrom South Africa and the phylogeny of nonmammalian cynodontsBulletin of the Museum of Comparative Zoology 1565ndash35
Hopson J A and H-D Sues 2006 A traversodont cynodont from theMiddle Triassic (Ladinian) of Baden-Wuumlrttemberg (Germany)Palaumlontologische Zeitschrift 80124ndash129
Huene F von 1928 Ein Cynodontier aus der Trias Brasiliens Zentral-blatt fuumlr Mineralogie Geologie und Palaumlontologie 1928251ndash270
Huene F von 1936 Die fossilen Reptilien des suumldamerikanischenGondwanalandes Ergebnisse der Sauriergrabungen in Suumldbrasilien192829 Lieferung 2 (Cynodontia)93ndash159 Franz F Heine Tuumlbin-gen
Jenkins F A 1970 The Chantildeares (Argentina) Triassic reptile faunaVII The postcranial skeleton of the traversodontid Massetognathuspascuali (Therapsida Cynodontia) Breviora 3521ndash28
Jenkins F A 1971 The postcranial skeleton of African cynodonts prob-lems in the early evolution of the mammalian postcranial skeletonBulletin of the Peabody Museum of Natural History 361ndash216
Kemp T S 1969 The atlas-axis complex of mammal-like reptiles Jour-nal of Zoology 159223ndash248
Kemp T S 1980 Aspects of the structure and functional anatomy of theMiddle Triassic cynodont Luangwa Journal of Zoology 191193ndash239
Kemp T S 1982 Mammal-Like Reptiles and the Origin of MammalsAcademic Press New York
Lehman J-P 1961 Cynodontia pp 140ndash191 in Piveteau J (ed) Traiteacutede Paleacuteontologie Masson et Cie Paris
Osborn 1903 On the primary division of the Reptilia into two sub-classes Synapsida and Diapsida Science 17275ndash276
Owen R 1861 Palaeontology or a systematic summary of extinct ani-mals and their geological relations 2nd Ed Edinburgh
Parrington F R 1934 On the cynodont genus Galesaurus with a noteon the functional significance of the changes in the evolution of thetheriodont skull Annals and Magazine of Natural History 10(13)38ndash67
Parrington F R 1946 On the cranial anatomy of cynodonts Proceed-ings of the Zoological Society of London 116181ndash197
Razafimbelo E 1987 Le basin de Morondava (Madagascar) Synthegravesegeacuteologique et structurale PhD dissertation University Louis Pas-teur Strasbourg France 189 pp
Reisz R R and H-D Sues 2000 Herbivory in Late Paleozoic andTriassic terrestrial vertebrates pp 9ndash41 in Sues H-D (ed) Evo-lution of Herbivory in Terrestrial Vertebrates Cambridge Univer-sity Press New York
Romer A S 1967 The Chantildeares (Argentina) Triassic reptile fauna IIITwo new gomphodonts Massetognathus pascuali and M terugiiBreviora 2641ndash25
Romer A S 1972 The Chantildeares (Argentina) Triassic reptile faunaXVII The Chantildeares gomphodonts Breviora 3961ndash9
Rougier G W J R Wible and J A Hopson 1992 Reconstruction ofthe cranial vessels in the Early Cretaceous mammal Vincelestes neu-quenianus implications for the evolution of the mammalian cranialvascular system Journal of Vertebrate Paleontology 12188ndash216
Rowe T 1988 Definition diagnosis and origin of Mammalia Journal ofVertebrate Paleontology 8241ndash264
Rowe T 1993 Early mammal phylogenetic systematics pp 129ndash145 inSzalay F S M J Novacek and M C McKenna (eds) MammalPhylogeny Vol 1 Mesozoic Differentiation MultituberculatesMonotremes Early Therians and Marsupials Springer-Verlag NewYork
Seeley H G 1894 Researches on the structure organization and clas-sification of the fossil Reptilia Part IX Section 3 On DiademodonPhilosophical Transactions of the Royal Society of London 1851029ndash1041
Seeley H G 1895 Researches on the structure organization and clas-sification of the fossil Reptilia Part IX Section 4 On the Gompho-dontia Philosophical Transactions of the Royal Society of London1861ndash57
Seeley H G 1895 Researches on the structure organization and clas-sification of the fossil Reptilia Part IX Section 5 On the skeletonin new Cynodontia from the Karroo rocks Philosophical Transac-tions of the Royal Society of London 18659ndash148
Sues H-D J A Hopson and N H Shubin 1992 Affinities of Sca-lenodontoides plemmyridon Hopson 1984 (Synapsida Cynodontia)from the Upper Triassic of Nova Scotia Journal of Vertebrate Pa-leontology 12168ndash171
Sues H-D and P E Olsen 1990 Triassic vertebrates of Gondwananaspect from the Richmond basin of Virginia Science 2491020ndash1023
Swofford D L 2003 PAUP v 40b10 Sinauer Associates Inc Pub-lishers Sunderland Massachusetts
Watson D M S and A S Romer 1956 A classification of therapsidreptiles Bulletin of the Museum of Comparative Zoology 11435ndash89
Submitted September 20 2007 accepted November 3 2007
KAMMERER ET ALmdashCYNODONT FROM MADAGASCAR 461
Dow
nloa
ded
by [
UZ
H H
aupt
bibl
ioth
ek
Zen
tral
bibl
ioth
ek Z
uumlric
h] a
t 05
51 1
0 Se
ptem
ber
2013
APPENDIX Data matrix used in the phylogenetic analysesThis is a revised version of the matrix of Abdala et al (2006) with the following changes character 43 [Incisor procumbency (0) absent (1) present]added Menadon recoded for characters 0 (Number of upper incisors was 1 ldquothreerdquo changed to 0 ldquofourrdquo) 5 (Position of paracanine fossae in relationto the upper canine was changed to 1 ldquomedialrdquo) 16 (Overall morphology of lower postcanines in occlusal view was changed to 3 ldquoquadran-gularrdquo) 17 (Position of the transverse crest in lower postcanines was changed to 1 ldquoanteriorrdquo) 18 (Number of cusps in the transverse crest of thelower postcanines was changed to 0 ldquotwordquo) 20 (Anterior cingulum in the lower postcanines was changed to 2 ldquoabsentrdquo) 21 (Internarial barwas 1 ldquoabsentrdquo changed to 0 ldquopresentrdquo) 38 (Posterior cingulum in the lower postcanines was changed to 1 ldquoabsentrdquo) and 40 (Deep occlusalbasins in the postcanines was changed to 1 ldquopresentrdquo) Langbergia recoded for character 34 (Incisor cutting margins was 0 ldquoserratedrdquo changedto 1 ldquosmoothly ridgedrdquo) based on the description of the incisors in Abdala et al (2006) Gomphodontosuchus recoded for character 31 (Maxilla inthe margin of the subtemporal fenestra was 1 ldquoincludedrdquo changed to 0 ldquoexcludedrdquo) based on von Huene (1936) and Kammerer (pers obs of theholotype) contra Hopsonrsquos (1985) reconstruction lsquoScalenodonrsquo hirschsoni recoded for character 39 (Anterior cingulum in the upper postcanines was1 ldquoabsentrdquo changed to 0 ldquopresentrdquo) based on Abdala (pers comm 2006) Trirachodon recoded for character 30 (Ectopterygoid was changedto 0 ldquopresentrdquo) based on Abdala (pers comm 2006) and Traversodon recoded for character 15 (Number of cusps in the sectorial border of the upperpostcanines was 0 ldquomore than twordquo changed to 1 ldquotwordquo) based on Abdala (pers comm 2006) and Kammerer (pers obs)
1111111111 2222222222 3333333333 44440123456789 0123456789 0123456789 0123456789 0123
Cynognathus 00000000 000 000000000 00000000 000Diademodon 0000001000 0000100 000010A000 0000000100 0100Trirachodon 000000100B 1A002201 001A100010 0001001200 0110Langbergia 0000001000 10000201 0000101010 0001101200 0110Cricodon 0100001000 11000201 0010100 0001200 0110Andescynodon 0000002001 01013101 101120010 0112311 1100Massetognathus 00111121A2 1110003100 2011200011 1011212300 1100Exaeretodon 1110122112 010103101 2111110201 1111112311 1101Luangwa 0000001002 11010A3100 101010010 01002300 1100Scal angustifrons 000000100B 1111023100 10102101 00002301 1100Scal hirschsoni 11110202 1100113100 1010 1112300 1100Traversodon 0000002012 1101113100 201010 011230 1100Gomphodontosuchus 0011112112 0103101 2010 0112311 1100Pascualgnathus 1000001000 000231 1100001 0011230 1100Scalenodontoides 1112212 01003101 21111021 1212311 1101Menadon 0111112112 01012310 20121020 0102311 1101Dadadon 00002112 111112 013 11231 1100Santacruzodon 00002112 1110003101 201300 212300 1100
A(01) B(02)
JOURNAL OF VERTEBRATE PALEONTOLOGY VOL 28 NO 2 2008462
Dow
nloa
ded
by [
UZ
H H
aupt
bibl
ioth
ek
Zen
tral
bibl
ioth
ek Z
uumlric
h] a
t 05
51 1
0 Se
ptem
ber
2013

Chatterjee S 1982 A new cynodont reptile from the Triassic of IndiaJournal of Paleontology 56203ndash214
Crompton A W 1955 On some Triassic cynodonts from TanganyikaProceedings of the Zoological Society of London 125617ndash669
Crompton A W 1972 Postcanine occlusion in cynodonts and tritylo-donts Bulletin of the British Museum (Natural History) Geology2129ndash71
Crompton A W and F Ellenberger 1957 On a new cynodont from theMolteno Beds and the origin of the tritylodontids Annals of theSouth African Museum 441ndash13
Flynn J J J M Parrish B Rakotosaminimanana W F Simpson R LWhatley and A R Wyss 1999 A Triassic fauna from Madagascarincluding early dinosaurs Science 286763ndash765
Flynn J J J M Parrish B Rakotosamimanana L RanivoharimananaW F Simpson and A R Wyss 2000 New traversodontids (Syn-apsida Eucynodontia) from the Triassic of Madagascar Journal ofVertebrate Paleontology 20422ndash427
Gow C E and P J Hancox 1993 First complete skull of the LateTriassic Scalenodontoides (Reptilia Cynodontia) from southern Af-rica In Lucas S G and M Morales (eds) The nonmarine TriassicNew Mexico Museum of Natural History amp Science Bulletin 3161ndash168
Grine F E B D Hahn and C E Gow 1978 Aspects of relativegrowth and variability in Diademodon (Reptilia Therapsida) SouthAfrican Journal of Science 7450ndash58
Hopson J A 1984 Late Triassic traversodont cynodonts from NovaScotia and southern Africa Palaeontologia Africana 25181ndash201
Hopson J A 1985 Morphology and relationships of Gomphodontosu-chus brasiliensis von Huene (Synapsida Cynodontia Tritylodontoi-dea) from the Triassic of Brazil Neues Jahrbuch fuumlr Geologie undPalaumlontologie Monatshefte 1985285ndash299
Hopson J A and J W Kitching 2001 A probainognathian cynodontfrom South Africa and the phylogeny of nonmammalian cynodontsBulletin of the Museum of Comparative Zoology 1565ndash35
Hopson J A and H-D Sues 2006 A traversodont cynodont from theMiddle Triassic (Ladinian) of Baden-Wuumlrttemberg (Germany)Palaumlontologische Zeitschrift 80124ndash129
Huene F von 1928 Ein Cynodontier aus der Trias Brasiliens Zentral-blatt fuumlr Mineralogie Geologie und Palaumlontologie 1928251ndash270
Huene F von 1936 Die fossilen Reptilien des suumldamerikanischenGondwanalandes Ergebnisse der Sauriergrabungen in Suumldbrasilien192829 Lieferung 2 (Cynodontia)93ndash159 Franz F Heine Tuumlbin-gen
Jenkins F A 1970 The Chantildeares (Argentina) Triassic reptile faunaVII The postcranial skeleton of the traversodontid Massetognathuspascuali (Therapsida Cynodontia) Breviora 3521ndash28
Jenkins F A 1971 The postcranial skeleton of African cynodonts prob-lems in the early evolution of the mammalian postcranial skeletonBulletin of the Peabody Museum of Natural History 361ndash216
Kemp T S 1969 The atlas-axis complex of mammal-like reptiles Jour-nal of Zoology 159223ndash248
Kemp T S 1980 Aspects of the structure and functional anatomy of theMiddle Triassic cynodont Luangwa Journal of Zoology 191193ndash239
Kemp T S 1982 Mammal-Like Reptiles and the Origin of MammalsAcademic Press New York
Lehman J-P 1961 Cynodontia pp 140ndash191 in Piveteau J (ed) Traiteacutede Paleacuteontologie Masson et Cie Paris
Osborn 1903 On the primary division of the Reptilia into two sub-classes Synapsida and Diapsida Science 17275ndash276
Owen R 1861 Palaeontology or a systematic summary of extinct ani-mals and their geological relations 2nd Ed Edinburgh
Parrington F R 1934 On the cynodont genus Galesaurus with a noteon the functional significance of the changes in the evolution of thetheriodont skull Annals and Magazine of Natural History 10(13)38ndash67
Parrington F R 1946 On the cranial anatomy of cynodonts Proceed-ings of the Zoological Society of London 116181ndash197
Razafimbelo E 1987 Le basin de Morondava (Madagascar) Synthegravesegeacuteologique et structurale PhD dissertation University Louis Pas-teur Strasbourg France 189 pp
Reisz R R and H-D Sues 2000 Herbivory in Late Paleozoic andTriassic terrestrial vertebrates pp 9ndash41 in Sues H-D (ed) Evo-lution of Herbivory in Terrestrial Vertebrates Cambridge Univer-sity Press New York
Romer A S 1967 The Chantildeares (Argentina) Triassic reptile fauna IIITwo new gomphodonts Massetognathus pascuali and M terugiiBreviora 2641ndash25
Romer A S 1972 The Chantildeares (Argentina) Triassic reptile faunaXVII The Chantildeares gomphodonts Breviora 3961ndash9
Rougier G W J R Wible and J A Hopson 1992 Reconstruction ofthe cranial vessels in the Early Cretaceous mammal Vincelestes neu-quenianus implications for the evolution of the mammalian cranialvascular system Journal of Vertebrate Paleontology 12188ndash216
Rowe T 1988 Definition diagnosis and origin of Mammalia Journal ofVertebrate Paleontology 8241ndash264
Rowe T 1993 Early mammal phylogenetic systematics pp 129ndash145 inSzalay F S M J Novacek and M C McKenna (eds) MammalPhylogeny Vol 1 Mesozoic Differentiation MultituberculatesMonotremes Early Therians and Marsupials Springer-Verlag NewYork
Seeley H G 1894 Researches on the structure organization and clas-sification of the fossil Reptilia Part IX Section 3 On DiademodonPhilosophical Transactions of the Royal Society of London 1851029ndash1041
Seeley H G 1895 Researches on the structure organization and clas-sification of the fossil Reptilia Part IX Section 4 On the Gompho-dontia Philosophical Transactions of the Royal Society of London1861ndash57
Seeley H G 1895 Researches on the structure organization and clas-sification of the fossil Reptilia Part IX Section 5 On the skeletonin new Cynodontia from the Karroo rocks Philosophical Transac-tions of the Royal Society of London 18659ndash148
Sues H-D J A Hopson and N H Shubin 1992 Affinities of Sca-lenodontoides plemmyridon Hopson 1984 (Synapsida Cynodontia)from the Upper Triassic of Nova Scotia Journal of Vertebrate Pa-leontology 12168ndash171
Sues H-D and P E Olsen 1990 Triassic vertebrates of Gondwananaspect from the Richmond basin of Virginia Science 2491020ndash1023
Swofford D L 2003 PAUP v 40b10 Sinauer Associates Inc Pub-lishers Sunderland Massachusetts
Watson D M S and A S Romer 1956 A classification of therapsidreptiles Bulletin of the Museum of Comparative Zoology 11435ndash89
Submitted September 20 2007 accepted November 3 2007
KAMMERER ET ALmdashCYNODONT FROM MADAGASCAR 461
Dow
nloa
ded
by [
UZ
H H
aupt
bibl
ioth
ek
Zen
tral
bibl
ioth
ek Z
uumlric
h] a
t 05
51 1
0 Se
ptem
ber
2013
APPENDIX Data matrix used in the phylogenetic analysesThis is a revised version of the matrix of Abdala et al (2006) with the following changes character 43 [Incisor procumbency (0) absent (1) present]added Menadon recoded for characters 0 (Number of upper incisors was 1 ldquothreerdquo changed to 0 ldquofourrdquo) 5 (Position of paracanine fossae in relationto the upper canine was changed to 1 ldquomedialrdquo) 16 (Overall morphology of lower postcanines in occlusal view was changed to 3 ldquoquadran-gularrdquo) 17 (Position of the transverse crest in lower postcanines was changed to 1 ldquoanteriorrdquo) 18 (Number of cusps in the transverse crest of thelower postcanines was changed to 0 ldquotwordquo) 20 (Anterior cingulum in the lower postcanines was changed to 2 ldquoabsentrdquo) 21 (Internarial barwas 1 ldquoabsentrdquo changed to 0 ldquopresentrdquo) 38 (Posterior cingulum in the lower postcanines was changed to 1 ldquoabsentrdquo) and 40 (Deep occlusalbasins in the postcanines was changed to 1 ldquopresentrdquo) Langbergia recoded for character 34 (Incisor cutting margins was 0 ldquoserratedrdquo changedto 1 ldquosmoothly ridgedrdquo) based on the description of the incisors in Abdala et al (2006) Gomphodontosuchus recoded for character 31 (Maxilla inthe margin of the subtemporal fenestra was 1 ldquoincludedrdquo changed to 0 ldquoexcludedrdquo) based on von Huene (1936) and Kammerer (pers obs of theholotype) contra Hopsonrsquos (1985) reconstruction lsquoScalenodonrsquo hirschsoni recoded for character 39 (Anterior cingulum in the upper postcanines was1 ldquoabsentrdquo changed to 0 ldquopresentrdquo) based on Abdala (pers comm 2006) Trirachodon recoded for character 30 (Ectopterygoid was changedto 0 ldquopresentrdquo) based on Abdala (pers comm 2006) and Traversodon recoded for character 15 (Number of cusps in the sectorial border of the upperpostcanines was 0 ldquomore than twordquo changed to 1 ldquotwordquo) based on Abdala (pers comm 2006) and Kammerer (pers obs)
1111111111 2222222222 3333333333 44440123456789 0123456789 0123456789 0123456789 0123
Cynognathus 00000000 000 000000000 00000000 000Diademodon 0000001000 0000100 000010A000 0000000100 0100Trirachodon 000000100B 1A002201 001A100010 0001001200 0110Langbergia 0000001000 10000201 0000101010 0001101200 0110Cricodon 0100001000 11000201 0010100 0001200 0110Andescynodon 0000002001 01013101 101120010 0112311 1100Massetognathus 00111121A2 1110003100 2011200011 1011212300 1100Exaeretodon 1110122112 010103101 2111110201 1111112311 1101Luangwa 0000001002 11010A3100 101010010 01002300 1100Scal angustifrons 000000100B 1111023100 10102101 00002301 1100Scal hirschsoni 11110202 1100113100 1010 1112300 1100Traversodon 0000002012 1101113100 201010 011230 1100Gomphodontosuchus 0011112112 0103101 2010 0112311 1100Pascualgnathus 1000001000 000231 1100001 0011230 1100Scalenodontoides 1112212 01003101 21111021 1212311 1101Menadon 0111112112 01012310 20121020 0102311 1101Dadadon 00002112 111112 013 11231 1100Santacruzodon 00002112 1110003101 201300 212300 1100
A(01) B(02)
JOURNAL OF VERTEBRATE PALEONTOLOGY VOL 28 NO 2 2008462
Dow
nloa
ded
by [
UZ
H H
aupt
bibl
ioth
ek
Zen
tral
bibl
ioth
ek Z
uumlric
h] a
t 05
51 1
0 Se
ptem
ber
2013
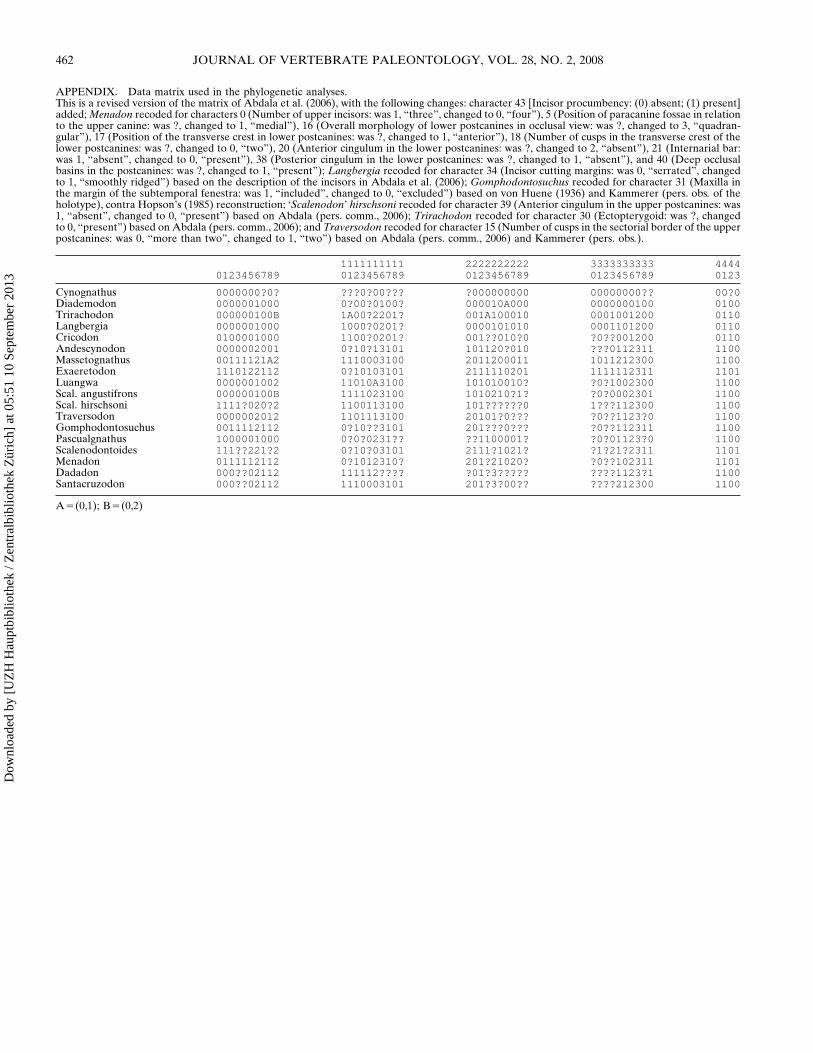
APPENDIX Data matrix used in the phylogenetic analysesThis is a revised version of the matrix of Abdala et al (2006) with the following changes character 43 [Incisor procumbency (0) absent (1) present]added Menadon recoded for characters 0 (Number of upper incisors was 1 ldquothreerdquo changed to 0 ldquofourrdquo) 5 (Position of paracanine fossae in relationto the upper canine was changed to 1 ldquomedialrdquo) 16 (Overall morphology of lower postcanines in occlusal view was changed to 3 ldquoquadran-gularrdquo) 17 (Position of the transverse crest in lower postcanines was changed to 1 ldquoanteriorrdquo) 18 (Number of cusps in the transverse crest of thelower postcanines was changed to 0 ldquotwordquo) 20 (Anterior cingulum in the lower postcanines was changed to 2 ldquoabsentrdquo) 21 (Internarial barwas 1 ldquoabsentrdquo changed to 0 ldquopresentrdquo) 38 (Posterior cingulum in the lower postcanines was changed to 1 ldquoabsentrdquo) and 40 (Deep occlusalbasins in the postcanines was changed to 1 ldquopresentrdquo) Langbergia recoded for character 34 (Incisor cutting margins was 0 ldquoserratedrdquo changedto 1 ldquosmoothly ridgedrdquo) based on the description of the incisors in Abdala et al (2006) Gomphodontosuchus recoded for character 31 (Maxilla inthe margin of the subtemporal fenestra was 1 ldquoincludedrdquo changed to 0 ldquoexcludedrdquo) based on von Huene (1936) and Kammerer (pers obs of theholotype) contra Hopsonrsquos (1985) reconstruction lsquoScalenodonrsquo hirschsoni recoded for character 39 (Anterior cingulum in the upper postcanines was1 ldquoabsentrdquo changed to 0 ldquopresentrdquo) based on Abdala (pers comm 2006) Trirachodon recoded for character 30 (Ectopterygoid was changedto 0 ldquopresentrdquo) based on Abdala (pers comm 2006) and Traversodon recoded for character 15 (Number of cusps in the sectorial border of the upperpostcanines was 0 ldquomore than twordquo changed to 1 ldquotwordquo) based on Abdala (pers comm 2006) and Kammerer (pers obs)
1111111111 2222222222 3333333333 44440123456789 0123456789 0123456789 0123456789 0123
Cynognathus 00000000 000 000000000 00000000 000Diademodon 0000001000 0000100 000010A000 0000000100 0100Trirachodon 000000100B 1A002201 001A100010 0001001200 0110Langbergia 0000001000 10000201 0000101010 0001101200 0110Cricodon 0100001000 11000201 0010100 0001200 0110Andescynodon 0000002001 01013101 101120010 0112311 1100Massetognathus 00111121A2 1110003100 2011200011 1011212300 1100Exaeretodon 1110122112 010103101 2111110201 1111112311 1101Luangwa 0000001002 11010A3100 101010010 01002300 1100Scal angustifrons 000000100B 1111023100 10102101 00002301 1100Scal hirschsoni 11110202 1100113100 1010 1112300 1100Traversodon 0000002012 1101113100 201010 011230 1100Gomphodontosuchus 0011112112 0103101 2010 0112311 1100Pascualgnathus 1000001000 000231 1100001 0011230 1100Scalenodontoides 1112212 01003101 21111021 1212311 1101Menadon 0111112112 01012310 20121020 0102311 1101Dadadon 00002112 111112 013 11231 1100Santacruzodon 00002112 1110003101 201300 212300 1100
A(01) B(02)
JOURNAL OF VERTEBRATE PALEONTOLOGY VOL 28 NO 2 2008462
Dow
nloa
ded
by [
UZ
H H
aupt
bibl
ioth
ek
Zen
tral
bibl
ioth
ek Z
uumlric
h] a
t 05
51 1
0 Se
ptem
ber
2013