Embed Size (px)
Citation preview

Neurons in the Posterior Interposed Nucleus of the Cerebellum Relatedto Vergence and Accommodation. I. Steady-State Characteristics
HONGYU ZHANG AND PAUL D. R. GAMLINVision Science Research Center, University of Alabama at Birmingham, Birmingham, Alabama 35294
Zhang, Hongyu and Paul D. R. Gamlin. Neurons in the posterior VI and VII) and the underlying fastigial nuclei are involvedinterposed nucleus of the cerebellum related to vergence and ac- in the regulation of saccades and smooth pursuit (e.g., Fuchscommodation. I. Steady-state characteristics. J. Neurophysiol. 79: et al. 1993, 1994; Kase et al. 1979; Keller 1989; Noda et1255–1269, 1998. The present study used single-unit recording al. 1988; Optican and Robinson 1980; Suzuki and Kellerand electrical microstimulation techniques in alert, trained rhesus 1988a,b) . The dentate nucleus and adjacent y-group possessmonkeys to examine the involvement of the posterior interposed
neurons that discharge in relation to vertical pursuit eyenucleus (IP) of the cerebellum in vergence and accommodativemovements (Chubb and Fuchs 1982). The cerebellar floc-eye movements. Neurons related to vergence and ocular accommo-culus and paraflocculus are involved in the vestibulo-oculardation were encountered within a circumscribed region of the IPreflex, optokinetic nystagmus, and smooth pursuit (e.g.,and their activity during changes in viewing distance was character-
ized. The activity of these neurons increased with decreases in Buttner and Waespe 1984; Lisberger and Fuchs 1978; Milesvergence angle and accommodation (the far-response) but none et al. 1980). In general, however, these oculomotor studiesshowed changes in activity during changes in conjugate eye posi- have focused on the role of the cerebellum in controllingtion and we therefore term them ‘‘far-response neurons.’’ Far- conjugate eye movements and relatively little is known aboutresponse neurons were found within a restricted region of the IP the contribution of the cerebellum to the control of vergencethat extended Ç1 mm rostrocaudally and mediolaterally and 2 mm and ocular accommodation.dorsal to the fourth ventricle. Microstimulation of this far-response
Single-unit recording studies in alert, trained rhesus mon-region of the IP with low currents (õ30 mA) often elicited diver-keys have previously reported the presence of neurons re-gence and accommodation for far. The behavior of 37 IP far-lated to vergence and accommodation in the midbrain in theresponse neurons was examined during normal binocular viewing,vicinity of the oculomotor nucleus (Judge and Cummingduring monocular viewing (blur cue alone), and during binocular
viewing with accommodation open-loop (disparity cue alone). The 1986; Mays 1984; Mays et al. 1986; Zhang et al. 1992). Weactivity of all cells was modulated under all three conditions. How- have shown that neurons within the fastigial and interpositusever, the change in activity of some of these neurons was signifi- nuclei of the cerebellum have reciprocal connections withcantly different under these three viewing conditions. The behavior this midbrain region (May et al. 1992). We have also shownof 70 IP far-response neurons was compared during normal binocu- that some neurons within a medial zone of a precerebellarlar viewing and during viewing in which the accommodative re- nucleus, the nucleus reticularis tegmenti pontis (NRTP) aresponse was significantly dissociated from the vergence response.
related either to the near-response or to the far-responseThe data from these two conditions was pooled and multiple regres-(Gamlin and Clarke 1995). Furthermore, Westheimer andsion analyses for each neuron generated two coefficients expressingBlair (1973) reported transient vergence deficits in primatesthe activity of the neuron relative to the vergence and to accommo-after cerebellectomy. Thus there is some circumstantial evi-dative response respectively. On the basis of these coefficients,dence that the cerebellum plays a role in vergence and ocularthe overall activity of the neurons were classified as follows: 34
positively correlated with divergence, 11 positively correlated with accommodation in primates.far accommodation, 14 positively correlated with divergence and There is further evidence that the deep cerebellar nucleifar accommodation, 9 positively correlated with divergence and are involved in ocular accommodation. Recording studiesaccommodation, and 2 positively correlated with convergence and in paralyzed, anesthetized cats have reported cells in thefar accommodation. The results of this study demonstrate the interpositus nucleus that are related to accommodation, andinvolvement of a specific region of the posterior interposed nucleus electrical microstimulation of this region orthodromicallyof the cerebellum in vergence and accommodation. IP far-response
activated accommodation-related cells in the midbrainneurons are active for vergence and accommodation irrespective(Bando et al. 1979a,b) . In addition, stimulation of the poste-of whether or not these eye movements are elicited by blur orrior interposed nucleus bilaterally and the fastigial nucleusdisparity cues. The data in the present study strongly suggest thatcontralaterally is reported to elicit positive accommodationthis cerebellar region is a far-response region that is involved in(Hosoba et al. 1978).vergence as well as accommodative eye movements.
Lesions in humans also suggest a role for the cerebellumin vergence and accommodative eye movements. Some pa-
I N T R O D U C T I O N tients with cerebellar lesions have been reported to showconvergence excess or divergence failure (Leigh and Zee
Experimental and clinical studies of the cerebellum have 1991). More recently Kawasaki and coworkers reported ashown that it is involved in the dynamic and adaptive control case in which a patient with cerebellar signs manifested aof eye movements (see Lewis and Zee 1993 for a review). slow release of accommodation (Kawasaki et al. 1993).
Taken together, these studies indicate a role for the cerebel-It has been shown in primates that the dorsal vermis (lobules
12550022-3077/98 $5.00 Copyright q 1998 The American Physiological Society
J603-6/ 9k26$$mr05 02-06-98 08:58:49 neupa LP-Neurophys
by 10.220.33.4 on Septem
ber 27, 2016http://jn.physiology.org/
Dow
nloaded from

H. ZHANG AND P.D.R. GAMLIN1256
demand can be produced, with real targets denoted by positivelum in vergence and ocular accommodation. However, thevalues of accommodation and vergence demand. This modificationevidence described above is fragmentary. The present studyalso allows a stationary fixation target with a well-defined vergencewas therefore aimed at providing more detailed informationand accommodative demand (range 04 D to /12 D) to be pre-on cerebellar involvement in vergence and ocular accommo-sented on one pair of video displays at the same time as eitherdation. With the use of single-unit recording techniques, we static or moving visual probe stimuli with different vergence and
have identified neurons in a specific region of the posterior accommodative demands are presented on the other pair of dis-interposed nucleus (IP) of the rhesus monkey that are related plays. Two important aspects of this visual stimulator are that 1to divergence and ocular accommodation for far. Electrical cm of motion of lenses L1–4 produces a 4 D change in target focusmicrostimulation of this region produces divergence eye and that, because the optical arrangement closely approximates a
Badal lens system, changes in accommodative demand produce nomovements and accommodation for far. The activity of thesesignificant change in target size or luminance. To facilitate theIP far-response neurons is related to vergence and accommo-presentation and analysis of accommodation and vergence, we cal-dation regardless of whether or not these movements areculate them in the equivalent units of diopters (D) and meter angleselicited during normal viewing conditions, during blur-(MA) respectively. One meter angle corresponds to the reciprocaldriven monocular viewing (disparity open-loop), or duringof the target distance in meters. In this scheme, vergence angledisparity-driven viewing (accommodation open-loop). Also, measured in MA is related to the vergence angle measured in
with dissociation of vergence from accommodation we degrees by the interpupillary distance. For example, with an inter-found that, as is the case for midbrain neurons (e.g., Judge pupillary distance of 3.2 cm, the average for our animals, 1 MAand Cumming 1986; Zhang et al. 1992), the activity of is approximately equal to 1.837.some neurons is related predominately to vergence, somepredominantly to accommodation, and some to both
Standard behavioral tasksvergence and accommodation. In addition, the firing rate ofIP far-response cells is related not only to the steady-state TARGETS STEPPED IN DEPTH. In this trial type the animal wasfar-response, but is also correlated with the dynamics of the required to make a far- or a near-response to a visual target thatmovement. Some cells display an apparent sensory signal was stepped in depth. Except for accommodation open-loop trials,related to the appearance of the far target. These dynamic all of these trials and tracking in depth trials used a checkered
target subtending 1.87. Because of the independent video displays,and sensorimotor characteristics of IP far-response cells willthe initial target position could range from 04 D to /12 D andbe presented separately. Preliminary results of some of thesefrom 04 to /12 MA and the required target step could be pre-findings have been presented elsewhere (Gamlin 1991; Gam-sented over the same ranges. Also, because accommodative andlin et al. 1996; Zhang and Gamlin 1994).vergence demands could be varied independently in this opticalsystem, targets could be presented with either appropriate or con-
M E T H O D S flicting vergence and accommodative demands as describedbelow.
Animal preparation and surgerySACCADIC TRIALS. Animals were required to make saccades to
Three juvenile rhesus monkeys (Macaca mulatta) were used visual targets over a range of {187 horizontally or vertically.in this study. All experimental procedures were approved by the
PURSUIT TRIALS. Animals were required to pursue a target thatUniversity of Alabama at Birmingham Institutional Animal Care moved in depth with either a ramp or a predictable sinusoidaland Use Committee and complied with the United States Public trajectory of different frequencies to elicit vergence and accommo-Health Service Policy on Human Care and Use of Laboratory dative tracking. These target could also move in the frontoparallelAnimals. All the described surgical procedures were performed plane so that smooth pursuit eye movements could be elicited.under sterile conditions using sodium pentobarbital anesthesia. In general, both saccadic and smooth pursuit eye movements inPostsurgically animals received analgesics (Numorphan, 0.15 the frontoparallel plane were used to test whether a putative far-mg/kg or Buprenex, 0.01 mg/kg) to minimize pain. Initially, ani- response neuron displayed activity that was influenced by conju-mals were implanted with four fixation plates attached to the skull gate eye position or velocity.by bone screws. One end of each plate was bent up Ç1 cm and aheadpost cemented between them with dental acrylic. The scalpwas sutured to tightly approximate the acrylic and the plates thus Further characterization of vergence and accommodationcovered completely by skin. With this system the head can be characteristicsheld stationary during training and subsequent recording sessions.Subsequently, scleral search coils for measuring eye movements As shown in Fig. 2, the vergence and accommodation systems(Fuchs and Robinson 1966) were implanted under the conjunctiva are cross-coupled, i.e., disparity-driven vergence signals influenceof each eye according to Judge et al. (1980). Finally, two recording accommodative output and, similarly, blur-driven accommodativecylinders, one on each side of the skull, were implanted stereotaxi- signals influence vergence output (e.g., Cumming and Judge 1986;cally over the cerebellum at a 157 angle to the sagittal plane. Fincham and Walton 1957; Hung and Semmlow 1980; Schor 1992;
Westheimer 1963). Thus to fully examine the relationship of thecell to its inputs and outputs, additional experiments are requiredVisual displaythat characterize the activity of the cell under viewing conditionsother than those imposed by normal viewing of a target that movesThe visual display system (Fig. 1) is similar to one used pre-
viously by us (Gamlin et al. 1989). It has a field of view of {187 in depth.As described below, we used the visual apparatus to put disparityand is based on the original design of Crane and Clark (1978). It
has however been modified to allow video displays to be used for into an open-loop condition by having the animal view a targetmonocularly. This apparatus was also used to put accommodationtarget presentation, with each having its own independent accom-
modative and vergence demand. By switching between two pairs of into an open-loop condition. Finally, the apparatus was used forconflict viewing experiments.video displays, step changes in both accommodative and vergence
J603-6/ 9k26$$mr05 02-06-98 08:58:49 neupa LP-Neurophys
by 10.220.33.4 on Septem
ber 27, 2016http://jn.physiology.org/
Dow
nloaded from

FAR-RESPONSE NEURONS IN THE POSTERIOR INTERPOSED NUCLEUS 1257
FIG. 1. Schematic view of visual display apparatus.L1–4 are 150-mm focal length moveable camera lenses.L5–8 are 50 mm focal length camera lenses. L9–12are achromatic doublets with an effective focal lengthof 106 mm. MM1 and MM2 are moveable mirror galva-nometers conjugate with the center of rotation of leftand right eye, respectively. M1–4 are front surface mir-rors. M5 and M6 are 50/50 beamsplitters. P1 and P2are pellicles. TV1–4 are super VGA monochrome mon-itors.
BLUR-DRIVEN ACCOMMODATION AND VERGENCE WITH DIS- CONFLICT VIEWING. This trial type was designed to better charac-terize the response characteristics of the far-response cells. ThisPARITY OPEN-LOOP. In this trial type, the animal views the targetapproach has been used in studies of the midbrain near-responsemonocularly placing disparity in an open-loop condition. The targetand far-response cells (Judge and Cumming 1986; Morley et al.is stepped or ramped from one accommodative demand to another1992; Zhang et al. 1992). Under normal viewing conditions, theand the animal is required to make an accurate accommodativevergence and accommodation demands of real targets are matchedmovement to the new target position. By measuring the accommo-in a predictable fashion determined by the interpupillary distance.dative response and vergence angle, one can calculate the responseBecause of this, vergence and accommodation responses are alsoAC/A [i.e., the ratio of accommodative convergence (AC) mea-closely matched. Therefore to determine more precisely how a cellsured in meter angles as a function of accommodation (A) mea-is related to each of these two components, the required vergencesured in diopters].and accommodation responses are put into conflict, with the animal
DISPARITY-DRIVEN VERGENCE AND ACCOMMODATION WITH required to increase its vergence response while increasing its ac-ACCOMMODATION OPEN-LOOP. In this trial type, the animal commodation by significantly less. So that animals could achieveviews binocularly with accommodation open-loop. The target is better dissociation of vergence from accommodation during conflictstepped or ramped from one disparity to another and the animal is viewing, most data were obtained during trials in which the targetrequired to make accurate vergence movements to the new target ramped in depth for 1–3 s and then maintained a steady vergence andposition. By measuring the accommodative response and vergence accommodative demand for the remainder of the trial. An example ofangle, one can calculate the response CA/C (i.e., the ratio of the behavior seen during this type of trial type is shown in Fig. 3.convergence accommodation (CA) measured in diopters as a func- The gain of the accommodative demand of the target was set to betion of convergence (C) measured in meter angles) . In our optical 0.25 of that appropriate for normal viewing. Thus for example, thesystem we placed accommodation into an open-loop condition by animal is required to converge 5 MA while only accommodatingpresenting a small (0.37) , circular image to the animal that was 1.25 D. Figure 3 shows that the animals were able to closely matchdefocused to such an extent (at least 04 D) that it was composedof spatial frequencies of õ0.5 cycles/deg. Because of the lowspatial frequency content of this stimulus it did not elicit accommo-dative eye movements (Charman and Tucker 1977; Tsuetaki andSchor 1987), but did elicit robust vergence responses. In confirma-tion of these previous reports, during monocular viewing of thisspecific stimulus in our optical system, we found that its movementin depth elicited no changes in accommodation.
FIG. 3. Typical behavior during normal binocular viewing (n) and con-flict viewing condition (m) . In conflict viewing condition, accommodativedemand was 25% of that in normal viewing condition. The dotted lineFIG. 2. Dual interaction model of accommodation and vergence control
(based on Hung and Semmlow 1980 and Schor 1992). AC, accommodative ( – – – ) represents vergence and accommodative demands under 2 condi-tions. Note that response closely matches demand under these 2 viewingconvergence; BA, blur-driven accommodation; CA, convergence accommo-
dation; DC, disparity-driven convergence. conditions.
J603-6/ 9k26$$mr05 02-06-98 08:58:49 neupa LP-Neurophys
by 10.220.33.4 on Septem
ber 27, 2016http://jn.physiology.org/
Dow
nloaded from

H. ZHANG AND P.D.R. GAMLIN1258
FR Å Ro / kdarAR / kdvrCR (1)their vergence and accommodative responses to these conflictingdemands and thus dissociate these normally tightly coupled re-
where FR is the firing rate of the far-response cell, Ro is the pre-sponses. We therefore used this paradigm to compare the activitydicted firing rate for viewing a distant target ( i.e., AR Å 0,of a far-response cell during binocular viewing in which the vergenceCR Å 0), kda is the accommodation coefficient, kdv is the vergenceand accommodation responses were matched to the activity of thecoefficient, AR is the accommodation response, and CR is thecell when these responses were dissociated.vergence response. Averages of firing rate, vergence angle, andaccommodation were computed for sequential 200-ms periods dur-
Eye movement recording ing steady fixation (1–4 s in duration) for various levels ofvergence and accommodation. Data from the normal viewing andThe horizontal and vertical gains of each eye were calibratedconflict viewing conditions were combined and the coefficients forindependently at the beginning of each recording session. This wasvergence and accommodation were generated by using multipledone by requiring the animal to fixate targets that appeared atregression analyses. To determine whether or not the kdv and kdavarious horizontal and vertical positions using either eye alone.values were significantly different from zero, t-tests were per-Animals showed little variability in fixation from trial to trial andformed with a conservative a-level of P õ 0.001.saccades of õ0.27 could be reliably detected. Accommodation of
the left eye was measured by using a continuous recording, infraredUnit recording and electrical microstimulationoptometer, which was calibrated during normal binocular viewing
as described previously (Zhang et al. 1992). The positions of bothBy using a Kopf microdrive, a parylene-insulated tungsten micro-the right and left eyes, and accommodation were sampled at 1 KHz
electrode was advanced to the deep nuclei through a premeasuredand stored on computer disk for later analysis.26-gauge cannula that just penetrated the tentorium and a 21-gaugehypodermic needle used to puncture the dura. The electrode was
Data analysis advanced to the far-response region of the interposed nucleus (IP),which was often indicated by encounters with saccadic and blinkThe recorded data were analyzed off-line by using a Sun work-cells that are found nearby. Sometimes the drive was continued untilstation equipped with interactive graphics. Individual trials show-reaching the fourth ventricle and, as the electrode was subsequentlying vergence demand, accommodation demand, accommodationretracted, far-response cells were isolated. Unit activity was filteredresponse, the positions of both eyes, and the velocities of vergencesharply above 5 KHz and the occurrence of a spike was detectedand accommodation were displayed. Below these traces, concurrentwith a window discriminator and recorded to computer disk to thesingle-unit activity was also displayed.nearest 0.1 ms. When needed, the microelectrode could be switchedAverages of firing rate, vergence angle, and accommodationto electrical microstimulation, which was produced by using biphasicwere computed for sequential 200-ms periods during steady fixa-stimuli of 0.2 ms total duration generated by a Grass S88 stimulatortion (1–4 s in duration) at various vergence angles. Scatterplotsand two stimulation isolation units.of firing rate as a function of vergence angle during normal viewing
were produced and correlation coefficients and regression parame-ters were determined for each plot. The slopes of these regression Histology and verification of recording sitelines yielded a measure of the normal viewing coefficient (knv) for
Animals were used for several months and marking lesions couldeach cell. The firing rate for fixation of a distant target (0 vergencenot be made at all relevant sites. However, records were kept of theand accommodation demand; Ronv) was computed from the inter-location of familiar landmarks, the X-Y location of the microposi-cept of the regression line with the y-axis. The threshold (T ) fortioner, and the depth at which cells of interest were located. Tothe neuron was computed from the intercept of the regression lineverify the location of our electrodes, marking lesions were madewith the x-axis.during the last 2 wk of recording at the site of responsive neuronsThe activity of some far-response cells was examined duringby passing 30 mA anodal current through the microelectrode for 20blur-driven accommodation and vergence (disparity open-loop)s. Subsequently, monkeys were anesthetized deeply with sodiumand during disparity-driven vergence and accommodation (accom-pentobarbital and perfused transcardially with saline followed bymodation open-loop). To allow for comparisons between these4% paraformaldehyde in 0.1 M phosphate buffer. The brains wereconditions and normal viewing, firing rate was related to vergencecut coronally at 40 mm thickness and a Nissl series was prepared.angle for all cells and viewing conditions. Scatterplots of firingThe marking lesions were retrieved from these histological sections.rate as a function of vergence angle during blur-driven accommo-
dation and vergence (disparity open-loop) were produced and cor-relation coefficients and regression parameters were determined R E S U L T Sfor each plot. The slopes of these regression lines yielded a measureof the sensitivity of the cell to blur-driven movements (kb) . In Initial identification of far-response neuronsaddition, scatterplots of firing rate as a function of vergence angle
We examined the single-unit activity of all cells encoun-during disparity-driven vergence and accommodation (accommo-tered in the deep nuclei during static vergence and oculardation open-loop) were produced and correlation coefficients and
regression parameters were determined for each plot. The slopes accommodation. Neurons with activity specifically relatedof these regression lines yielded a measure of the sensitivity of the to the far-response were found in the posterior interposedcell to disparity-driven movements (kd) . To test for differences nucleus. Figure 4A shows the behavior of a cell (cell 32 inbetween the values for kb , kd , and knv F-tests were used with an Table 1) recorded in this nucleus during a decrease ina-level of P õ 0.001. This conservative a-level was chosen vergence angle and accommodation that occurred in re-throughout to minimize type I errors. sponse to a step change in target distance from 4 to 1 MA.To determine the degree to which the activity of a far-response
This cell shows a clear increase in its firing rate when lookingcell is associated with changes in vergence or accommodation, wefrom the near to the far target. Figure 4E summarizes theused the conflict viewing strategy as described above. To quantifyoverall relationship between vergence angle and firing ratethis procedure we used the same methods as reported by Zhang etof this cell during normal binocular viewing. The responseal. (1992). It assumes the following linear interaction between
vergence and accommodation: of the cell during sine-wave tracking of a target moving in
J603-6/ 9k26$$mr05 02-06-98 08:58:49 neupa LP-Neurophys
by 10.220.33.4 on Septem
ber 27, 2016http://jn.physiology.org/
Dow
nloaded from

FAR-RESPONSE NEURONS IN THE POSTERIOR INTERPOSED NUCLEUS 1259
depth from 1 to 4 MA was also examined (Fig. 4B) . Under Thus, on the basis of its responses during these standardtrials, the cell in Fig. 4 would satisfy our definition of athese conditions, the firing rate of the cell is modulated as
a function of viewing distance as would be expected from far-response cell. The conjugate sensitivity of all cells wasexamined on-line and none were modulated during saccadicchange in firing rate during the static trial shown in Fig. 4A .
In addition however, on the basis of the phase lead of the or smooth pursuit eye movements. To further document thisfinding, nine far-response neurons were examined systemati-cell during this trial and on the transient increase in firing
rate that is apparent during the divergence movement shown cally during vertical and horizontal saccades and during hori-zontal smooth pursuit. Summary data plots from these cellsin Fig. 4A , it is evident that this cell also shows a strongare shown in Fig. 5. Figure 5A shows the vergence sensitivitycorrelation between firing rate and vergence velocity and aof these cells and demonstrates that their firing rates decreasescatterplot of this relationship is shown for this particularsignificantly during convergence eye movements. In con-cell in Fig. 4F . Such a vergence velocity sensitivity is atrast, Fig. 5, B– D shows that the firing rate of these cellscommon feature of IP far-response neurons and will be ad-remains essentially constant for vertical and horizontal sac-dressed at length in a future communication.cades and for horizontal smooth pursuit eye movements overEach far-response neuron was also examined for its sensi-an even greater oculomotor range than that shown for thetivity to changes in conjugate eye position. The activity ofvergence eye movements.each cell was tested during trials in which the animal made
smooth pursuit eye movements and other trials in which itmade saccadic eye movements. Figure 4, C and D show the Location of far-response neuronsbehavior of the same far-response cell as is shown in Fig.4, A and B during these eye movements while viewing a A total of 185 cells with the characteristics describedtarget with 1 MA of vergence demand. Note that the activity above were recorded from the posterior interposed nucleusof this cell is not altered during smooth pursuit eye move- of three animals. Sufficient data were collected from 70 of
these cells for the steady-state analyses described below.ments (Fig. 4C) or saccadic eye movements (Fig. 4D) .
FIG. 4. A : behavior of an IP far-response cellfor a symmetrical far-response movement in re-sponse to a step change in accommodative andvergence demand (normal binocular viewing). Inthis and subsequent figures, convergence, accom-modation for near, and rightward eye movementsare represented as upward on traces. B : responseof this cell during sine-wave tracking of a targetmoving in depth. C : activity of this cell is unaf-fected by smooth pursuit of a horizontally movingtarget at optical infinity. D : activity of this cellis unaffected by saccadic eye movements madebetween targets at optical infinity. E : summaryplot of firing rate as a function of vergence anglefor this cell. F : summary plot of firing rate as afunction of vergence velocity for this cell. ACC,accommodation; HL, horizontal left eye position;HR, horizontal right eye position; VA, vergenceangle; VR, vertical right eye position. Scalebars Å 4 meter angle (MA) in A and B ; 47 in Cand D .
J603-6/ 9k26$$mr05 02-06-98 08:58:49 neupa LP-Neurophys
by 10.220.33.4 on Septem
ber 27, 2016http://jn.physiology.org/
Dow
nloaded from
H. ZHANG AND P.D.R. GAMLIN1260
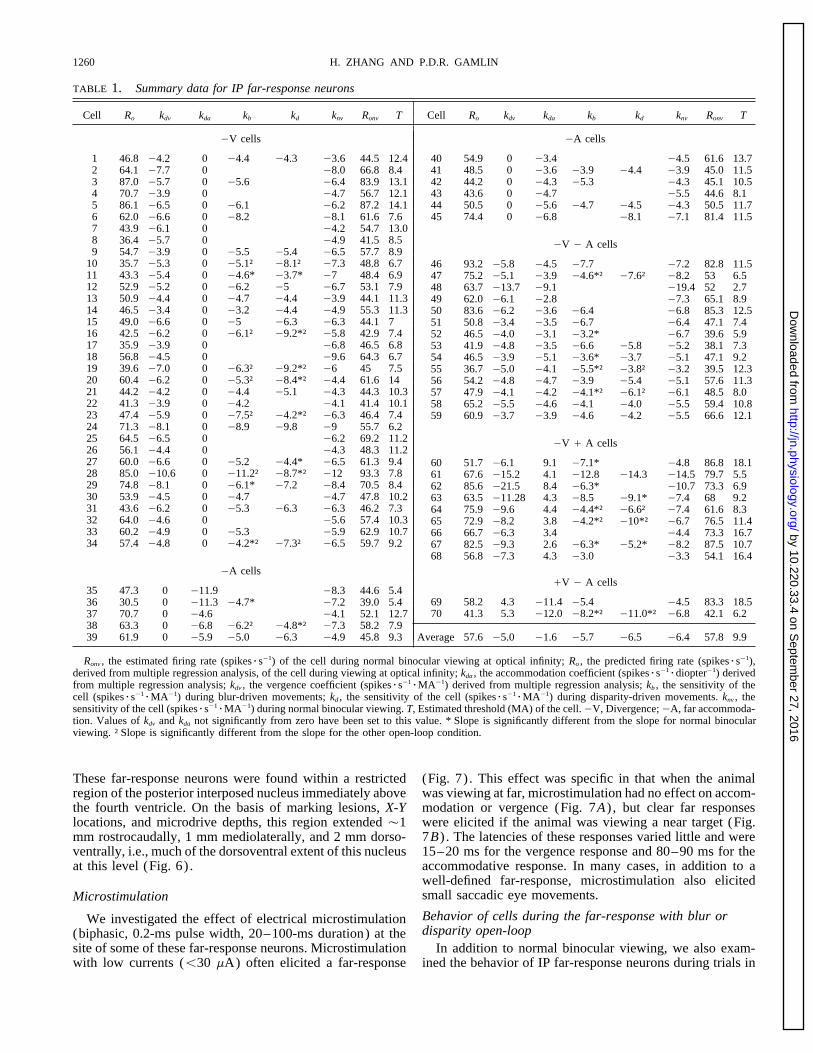
TABLE 1. Summary data for IP far-response neurons
Cell Ro kdv kda kb kd knv Ronv T Cell Ro kdv kda kb kd knv Ronv T
0V cells
1 46.8 04.2 0 04.4 04.3 03.6 44.5 12.42 64.1 07.7 0 08.0 66.8 8.43 87.0 05.7 0 05.6 06.4 83.9 13.14 70.7 03.9 0 04.7 56.7 12.15 86.1 06.5 0 06.1 06.2 87.2 14.16 62.0 06.6 0 08.2 08.1 61.6 7.67 43.9 06.1 0 04.2 54.7 13.08 36.4 05.7 0 04.9 41.5 8.59 54.7 03.9 0 05.5 05.4 06.5 57.7 8.9
10 35.7 05.3 0 05.1† 08.1† 07.3 48.8 6.711 43.3 05.4 0 04.6* 03.7* 07 48.4 6.912 52.9 05.2 0 06.2 05 06.7 53.1 7.913 50.9 04.4 0 04.7 04.4 03.9 44.1 11.314 46.5 03.4 0 03.2 04.4 04.9 55.3 11.315 49.0 06.6 0 05 06.3 06.3 44.1 716 42.5 06.2 0 06.1† 09.2*† 05.8 42.9 7.417 35.9 03.9 0 06.8 46.5 6.818 56.8 04.5 0 09.6 64.3 6.719 39.6 07.0 0 06.3† 09.2*† 06 45 7.520 60.4 06.2 0 05.3† 08.4*† 04.4 61.6 1421 44.2 04.2 0 04.4 05.1 04.3 44.3 10.322 41.3 03.9 0 04.2 04.1 41.4 10.123 47.4 05.9 0 07.5† 04.2*† 06.3 46.4 7.424 71.3 08.1 0 08.9 09.8 09 55.7 6.225 64.5 06.5 0 06.2 69.2 11.226 56.1 04.4 0 04.3 48.3 11.227 60.0 06.6 0 05.2 04.4* 06.5 61.3 9.428 85.0 010.6 0 011.2† 08.7*† 012 93.3 7.829 74.8 08.1 0 06.1* 07.2 08.4 70.5 8.430 53.9 04.5 0 04.7 04.7 47.8 10.231 43.6 06.2 0 05.3 06.3 06.3 46.2 7.332 64.0 04.6 0 05.6 57.4 10.333 60.2 04.9 0 05.3 05.9 62.9 10.734 57.4 04.8 0 04.2*† 07.3† 06.5 59.7 9.2
0A cells
35 47.3 0 011.9 08.3 44.6 5.436 30.5 0 011.3 04.7* 07.2 39.0 5.437 70.7 0 04.6 04.1 52.1 12.738 63.3 0 06.8 06.2† 04.8*† 07.3 58.2 7.939 61.9 0 05.9 05.0 06.3 04.9 45.8 9.3
0A cells
40 54.9 0 03.4 04.5 61.6 13.741 48.5 0 03.6 03.9 04.4 03.9 45.0 11.542 44.2 0 04.3 05.3 04.3 45.1 10.543 43.6 0 04.7 05.5 44.6 8.144 50.5 0 05.6 04.7 04.5 04.3 50.5 11.745 74.4 0 06.8 08.1 07.1 81.4 11.5
0V 0 A cells
46 93.2 05.8 04.5 07.7 07.2 82.8 11.547 75.2 05.1 03.9 04.6*† 07.6† 08.2 53 6.548 63.7 013.7 09.1 019.4 52 2.749 62.0 06.1 02.8 07.3 65.1 8.950 83.6 06.2 03.6 06.4 06.8 85.3 12.551 50.8 03.4 03.5 06.7 06.4 47.1 7.452 46.5 04.0 03.1 03.2* 06.7 39.6 5.953 41.9 04.8 03.5 06.6 05.8 05.2 38.1 7.354 46.5 03.9 05.1 03.6* 03.7 05.1 47.1 9.255 36.7 05.0 04.1 05.5*† 03.8† 03.2 39.5 12.356 54.2 04.8 04.7 03.9 05.4 05.1 57.6 11.357 47.9 04.1 04.2 04.1*† 06.1† 06.1 48.5 8.058 65.2 05.5 04.6 04.1 04.0 05.5 59.4 10.859 60.9 03.7 03.9 04.6 04.2 05.5 66.6 12.1
0V / A cells
60 51.7 06.1 9.1 07.1* 04.8 86.8 18.161 67.6 015.2 4.1 012.8 014.3 014.5 79.7 5.562 85.6 021.5 8.4 06.3* 010.7 73.3 6.963 63.5 011.28 4.3 08.5 09.1* 07.4 68 9.264 75.9 09.6 4.4 04.4*† 06.6† 07.4 61.6 8.365 72.9 08.2 3.8 04.2*† 010*† 06.7 76.5 11.466 66.7 06.3 3.4 04.4 73.3 16.767 82.5 09.3 2.6 06.3* 05.2* 08.2 87.5 10.768 56.8 07.3 4.3 03.0 03.3 54.1 16.4
/V 0 A cells
69 58.2 4.3 011.4 05.4 04.5 83.3 18.570 41.3 5.3 012.0 08.2*† 011.0*† 06.8 42.1 6.2
Average 57.6 05.0 01.6 05.7 06.5 06.4 57.8 9.9
Ronv , the estimated firing rate (spikesrs01) of the cell during normal binocular viewing at optical infinity; Ro , the predicted firing rate (spikesrs01),derived from multiple regression analysis, of the cell during viewing at optical infinity; kda , the accommodation coefficient (spikesrs01
rdiopter01) derivedfrom multiple regression analysis; kdv , the vergence coefficient (spikesrs01
rMA01) derived from multiple regression analysis; kb , the sensitivity of thecell (spikesrs01
rMA01) during blur-driven movements; kd , the sensitivity of the cell (spikesrs01rMA01) during disparity-driven movements. knv , the
sensitivity of the cell (spikesrs01rMA01) during normal binocular viewing. T, Estimated threshold (MA) of the cell.0V, Divergence; 0A, far accommoda-
tion. Values of kdv and kda not significantly from zero have been set to this value. * Slope is significantly different from the slope for normal binocularviewing. † Slope is significantly different from the slope for the other open-loop condition.
These far-response neurons were found within a restricted (Fig. 7) . This effect was specific in that when the animalwas viewing at far, microstimulation had no effect on accom-region of the posterior interposed nucleus immediately above
the fourth ventricle. On the basis of marking lesions, X-Y modation or vergence (Fig. 7A) , but clear far responseswere elicited if the animal was viewing a near target (Fig.locations, and microdrive depths, this region extended Ç1
mm rostrocaudally, 1 mm mediolaterally, and 2 mm dorso- 7B) . The latencies of these responses varied little and were15–20 ms for the vergence response and 80–90 ms for theventrally, i.e., much of the dorsoventral extent of this nucleus
at this level (Fig. 6) . accommodative response. In many cases, in addition to awell-defined far-response, microstimulation also elicitedsmall saccadic eye movements.Microstimulation
Behavior of cells during the far-response with blur orWe investigated the effect of electrical microstimulationdisparity open-loop(biphasic, 0.2-ms pulse width, 20–100-ms duration) at the
site of some of these far-response neurons. Microstimulation In addition to normal binocular viewing, we also exam-ined the behavior of IP far-response neurons during trials inwith low currents (õ30 mA) often elicited a far-response
J603-6/ 9k26$$mr05 02-06-98 08:58:49 neupa LP-Neurophys
by 10.220.33.4 on Septem
ber 27, 2016http://jn.physiology.org/
Dow
nloaded from

FAR-RESPONSE NEURONS IN THE POSTERIOR INTERPOSED NUCLEUS 1261
FIG. 5. Firing rate of IP far-response neuronsincreases as a function of increases in vergenceangle (A) . No such change in firing rate is seenfor horizontal saccades (B) , vertical saccades (C) ,or horizontal smooth pursuit (D) . Horizontalscale: A, meter angles; B–D, degrees.
which blur or disparity were open-loop. The behavior of value that was not significantly different from zero (Table1; Cells 1–34) . During normal binocular viewing, Fig.53 IP far-response neurons were studied during monocular
viewing. Of these neurons, 37 were also studied during bin- 10A shows the decrease in activity of one of these cells(cell 5 in Table 1) in response to an increase in conver-ocular viewing with accommodation open-loop. A summary
of the results obtained during these trials is presented in gence of 6 MA and in accommodation of 6 D. Figure 10Bshows the behavior of the same cell during conflict viewingTable 1. The activity of all tested cells was modulated under
both of these viewing conditions. The response of one neuron in which there is a convergence movement of 6 MA (asimilar amount to that seen in Fig. 10A ) but an increase of(Cell 58 in Table 1) is shown in Fig. 8 during normal binocu-
lar viewing (Fig. 8A) , viewing with disparity open-loop accommodation of only 2 D. This reduced accommodativeresponse can be appreciated by referring to Fig. 10B(Fig. 8B) , and viewing with blur open-loop (Fig. 8C) of
a target that stepped from an apparent distance of 20 cm to ( – – – ) , indicating the approximate level of the accom-modative response in Fig. 10A . Under both viewing condi-an apparent distance of 1 m. Overall, the relationship be-
tween the activity of this particular neuron and vergence tions, the change in firing rate is the same and is closelyrelated to the vergence angle. This is confirmed by theangle is not significantly different under these three condi-
tions (Fig. 8D) . This is also true for the population averages scatterplots in Fig. 10C of the relationship between thevergence angle and the cell’s firing rate under the twoas is indicated in Table 1. However, as can also be seen in
Table 1, the activity of some cells is significantly different viewing conditions. Clearly neither the slopes nor inter-cepts of the regression lines are significantly different frombetween these two open-loop conditions. In some cases, it
is also different between normal viewing and these open- one another. Figure 10D shows scatterplots of the cell’sfiring rate as a function of accommodation for the sameloop conditions. Summary plots in Fig. 9 illustrate these
differences. two viewing conditions. In contrast to 10C , there is a pro-nounced difference in the relationship between firing rate
Determination of far-response cell type and accommodation under these two conditions. This cellwould therefore be characterized as a divergence cell (-VAfter the initial identification of a far-response cell, wecell) on the basis of the classification scheme previouslydetermined its relationship to vergence and to ocular ac-introduced by us (Zhang et al. 1992) .commodation under conflict viewing conditions. These
cells were classified into different groups using a classifi-cation scheme similar to that developed by us previously Far accommodation cells (0A cells)(Zhang et al. 1992) . Table 1 summarizes the response prop-erties based on multiple regression analyses for these far- Eleven far-response cells had a negative kda coefficientresponse cell during normal binocular and conflict viewing that was significantly different from zero and a kdv value thatconditions. was not significantly different from zero (Table 1; cells 35–
45). Figure 11A shows the decrease in activity of one ofDivergence cells (0V cells) these cells (cell 38 in Table 1) in response to convergence
of 6 MA and accommodation of 6 D during normal binocularThirty-four far-response cells had a negative kdv coeffi-cient that was significantly different from zero and a kda viewing. Figure 11B shows the behavior of the same cell
J603-6/ 9k26$$mr05 02-06-98 08:58:49 neupa LP-Neurophys
by 10.220.33.4 on Septem
ber 27, 2016http://jn.physiology.org/
Dow
nloaded from

H. ZHANG AND P.D.R. GAMLIN1262
FIG. 6. A : schematic illustration of a coronal sec-tion through cerebellum. Enclosed area indicates re-gion of deep nuclei from which photomicrograph wastaken. B : photomicrograph of a coronal section at levelof deep cerebellar nuclei showing site of a markingelectrolytic lesion in posterior interposed nucleus. F,fastigial nucleus; IA, anterior interposed nucleus; IP,posterior interposed nucleus; D, dentate nucleus. ScaleBar in A and B Å 2 mm.
during conflict viewing in which there is a convergence ted lines above the frequency (FREQ) traces in Figs. 11, Aand B , indicate that the change in firing rate is very differentmovement of 6 MA (a similar amount to that seen in Fig.
11A) but an increase of accommodation of only 2 D. This under these two viewing conditions and that it is closelyrelated to the accommodative response. This is confirmedreduced accommodative response can be appreciated by re-
ferring to the dashed line above the accommodative trace by the scatterplots in Fig. 11D of the relationship betweenaccommodation and the cell’s firing rate under the two view-(ACC) in Fig. 11B , indicating the approximate level of the
accommodative response in Fig. 11A . Reference to the dot- ing conditions. Neither the slopes nor intercepts of the re-
FIG. 7. A : electrical microstimulationwith low current (30 mA; 100 ms; 500 Hz)at location of a far-response cell elicits novergence or accommodation response wheneyes are diverged. B : microstimulationwith same parameters elicits well-defineddecreases in convergence and accommoda-tion when eyes are converged and accom-modating for near. Scale bar Å 47 and 2diopters.
J603-6/ 9k26$$mr05 02-06-98 08:58:49 neupa LP-Neurophys
by 10.220.33.4 on Septem
ber 27, 2016http://jn.physiology.org/
Dow
nloaded from

FAR-RESPONSE NEURONS IN THE POSTERIOR INTERPOSED NUCLEUS 1263
FIG. 8. A : behavior of an IP far-re-sponse cell (cell 58 in Table 1) during nor-mal binocular viewing. B : behavior of thiscell during monocular accommodativevergence. C : behavior of this cell duringbinocular viewing with accommodationopen-loop. D : scatter plot of firing rate asa function of vergence angle for 3 viewingconditions. ( ) , normal viewing;(rrr) , monocular viewing; ( – – – ), bin-ocular viewing condition with accommoda-tion open-loop. Scale bar Å 4 MA and 4diopters.
gression lines are significantly different from one another. the basis of the classification scheme previously introducedby us (Zhang et al. 1992).Figure 11C shows scatterplots of the cell’s firing rate as a
function of vergence angle for the same two viewing condi-tions. In contrast to 11D , there is a pronounced difference Divergence plus far accommodation cells (0V 0 A cells)in the relationship between firing rate and vergence angleunder the two viewing conditions. This cell would therefore Fourteen far-response cells had negative kdv and kda coef-be characterized as a far-accommodation cell (-A cell) on ficients, both of which were significantly different from zero
(Table 1, cells 46–59). During normal binocular viewingand conflict viewing the change in firing rate of these cellswas not related entirely either to vergence angle or to theaccommodative response. This is shown for one cell (cell 49in Table 1) in the scatterplots in Fig. 12A of the relationshipbetween vergence angle and firing rate under the two view-ing conditions. The slope of the regression line during con-flict viewing is significantly less steep from that seen duringnormal viewing. Thus the firing rate of the cell is not entirelyrelated to vergence angle. Figure 12B shows scatterplots ofthe cell’s firing rate as a function of accommodation for thesame two viewing conditions. As in 12A , the relationshipbetween firing rate and accommodation is significantly dif-ferent under the two viewing conditions. This cell wouldtherefore be characterized as a divergence plus far-accom-modation cell (0V0 A cell) on the basis of the classificationscheme previously introduced by us (Zhang et al. 1992).
Divergence minus far accommodation cells (0V / A cells)
Nine far-response cells had negative kdv and positive kda
coefficients that were significantly different from zero (Table1; cells 60–68). During normal binocular and conflict view-ing the change in firing rate of these cells was not related
FIG. 9. A : plot of 37 far-response cells comparing their response during entirely either to vergence angle or to the accommodativenormal viewing (knv) and disparity open-loop (kb) . B : plot of same far- response and their firing rates decreased more with conflictresponse cells comparing their response during normal viewing (knv) and viewing than with normal viewing. This observation is con-accommodation open-loop (kd) . C : plot of same far-response cells compar-
firmed for one such cell (cell 63 in Table 1) by the scat-ing their response during disparity open-loop (kb) and accommodation open-loop (kd) . terplots in Fig. 12C of the relationship between the vergence
J603-6/ 9k26$$mr05 02-06-98 08:58:49 neupa LP-Neurophys
by 10.220.33.4 on Septem
ber 27, 2016http://jn.physiology.org/
Dow
nloaded from

H. ZHANG AND P.D.R. GAMLIN1264
FIG. 10. Response properties of a di-vergence (0V) IP far-response cell (cell 5in Table 1) during normal binocular view-ing and partial dissociation of vergencefrom accommodation. Initial VA and ACCof 1 MA and 1 D respectively in A and B.A : during normal binocular viewing activ-ity of neuron decreases as a function ofnear response. B : during conflict viewingcondition, in which accommodative de-mand and responses are less than normal,both vergence amplitude and activity ofneuron is similar to that in A . – – – : ac-commodative response seen in A . C : scat-terplot and linear regression of firing rateas a function of vergence angle for normalviewing (m) , and conflict viewing (n) . D :scatterplot and linear regression of firingrate as a function of accommodation fornormal viewing (m) and conflict viewingcondition (n) . Scale bar Å 4 MA and 4diopters.
angle and firing rate under the two viewing conditions. The during conflict viewing is more than would be expected fora far-response cell that was solely related to vergence angleslope of the regression line during conflict viewing is sig-
nificantly steeper from that seen during normal viewing. ( i.e., a 0V cell) . Indeed, as described above, this was con-firmed by the multiple regression analysis, which yielded aFigure 12D shows scatterplots of the firing rate of the cell
as a function of accommodation for the same two viewing negative kdv and a positive kda for this cell. Thus this cellwould be characterized as a divergence minus far-accomoda-conditions. As in 12C , the relationship between firing rate
and accommodation is significantly different under the two tion cell (0V / A cell) on the basis of the classificationscheme previously introduced by us (Zhang et al. 1992).viewing conditions and the decrease in firing rate of this cell
FIG. 11. Response properties of a faraccommodation (0A) IP far-response cell(cell 38 in Table 1) during normal binocu-lar viewing and partial dissociation ofvergence from accommodation during con-flict viewing. Initial VA and ACC of 1 MAand 1 D respectively in A and B. A : duringnormal binocular viewing, activity of thisneuron decreases as a function of near re-sponse. B : response of this cell during aconvergence movement similar in ampli-tude to that in A and an accommodativeresponse substantially lower than that inA . This difference is best appreciated byreference to upper of 2 dashed lines thatindicates accommodative response in A .For reference, lower of 2 dashed line islocated at same relative level as it is in A .Note that decrease in firing rate is muchless than in A . C : scatterplot and linearregression of firing rate as a function ofvergence angle for normal viewing (m) andconflict viewing (n) . D : scatterplot andlinear regression of firing rate as a functionof accommodation for normal viewing (m)and conflict viewing condition (n) . Scalebar Å 4 MA and 4 diopters.
J603-6/ 9k26$$mr05 02-06-98 08:58:49 neupa LP-Neurophys
by 10.220.33.4 on Septem
ber 27, 2016http://jn.physiology.org/
Dow
nloaded from

FAR-RESPONSE NEURONS IN THE POSTERIOR INTERPOSED NUCLEUS 1265
FIG. 12. A and B : divergence plus faraccommodation (0V 0 A) IP far-responsecell (cell 49 in Table 1). A : scatterplot andlinear regression of firing rate as a functionof vergence angle for normal viewing (m)and conflict viewing (n) . B : scatterplot andlinear regression of firing rate as a functionof accommodation for normal viewing (m)and conflict viewing condition (n) . C andD : divergence minus far accommodation(0V / A) IP far-response cell (cell 63 inTable 1). C : scatterplot and linear regres-sion of firing rate as a function of vergenceangle for normal viewing (m) and conflictviewing (n) . D : scatterplot and linear re-gression of firing rate as a function ofaccommodation for normal viewing (m)and conflict viewing condition (n) . Scalebar Å 4 MA and 4 diopters.
Far accommodation minus divergence cells (/V 0 A) Threshold for IP far-response neurons
Figure 14 presents the threshold data for all 70 IP far-Only two far-response cells had positive kdv and negativeresponse neurons. The curve represents a logistic fit to thekda coefficients that were significantly different from zerosample of thresholds for this population of neurons. Thisand these were classified as far accommodation minus diver-curve shows clearly that with as little convergence as 5 MAgence (/V 0 A) cells (Table 1, cells 69 and 70).some IP far-response neurons become inactive and that with
Relationship of kda and kdv to knv
During normal viewing the accommodative and vergenceresponses are matched and therefore Eq. 1 , which was usedin our multiple regression analysis linking the activity ofthe cell independently to the accommodative and vergenceresponses by their respective gain factors of kda and kdv ,reduces to:
FR Å Ro / (kda / kdv)rCR (2)
Equation 2 is directly comparable to a single linear regres-sion of firing rate as a function of vergence angle in which
FR Å Ronv / knvrCR (3)
Thus for normal viewing, knv should be more closely matchedto the sum of the kda and kdv coefficients than to either ofthese two coefficients alone. As can be seen from Fig. 13, A–C , this is the case. The similarity is confirmed by statisticalanalysis showing that the correlation between knv and kdv islow (r Å 0.55) and that the correlation between knv and kda
is insignificant, while that between knv and (kda / kdv) isrobust (r Å 0.74). Comparison of Eqs. 2 and 3 also impliesthat the multiple regression analysis will yield an Ro value
FIG. 13. A : comparison of kda with knv for a sample of 70 cells. B :that approximates the Ronv value generated by single linear comparison of kdv with knv for same group of cells. C : comparison ofregression. As illustrated in Fig. 13D , the correlation be- kdv / kda with knv for same group of cells. D : comparison of Ronv with Ro
for same group of cells.tween these two values is strong (r Å 0.84).
J603-6/ 9k26$$mr05 02-06-98 08:58:49 neupa LP-Neurophys
by 10.220.33.4 on Septem
ber 27, 2016http://jn.physiology.org/
Dow
nloaded from

H. ZHANG AND P.D.R. GAMLIN1266
are summarized in Fig. 15. More specifically, the afferentconnections of the IP are arranged so that the NRTP is in aposition to relay a signal related to vergence and accommo-dation to this nucleus. Because the NRTP is reported toreceive input from the prearcuate cortex (Leichnetz et al.1984), it is possible that a far-response signal reaches NRTPneurons from the far-response neurons that have recentlybeen reported in this region (Gamlin and Yoon 1995; Gamlinet al. 1996). The connections of the IP with neurons in themidbrain near-response region are such that these midbrainneurons could relay a signal related to motor output (corol-lary discharge) to the IP. The efferent projections from theIP to the midbrain near-response neurons are in a positionto modulate the activity of these midbrain neurons on thebasis of mismatches between the desired and actual motoroutput signals impinging on the IP. Furthermore, these sig-nals, if supplemented by signals related to visual errors ( i.e.,
FIG. 14. Plot of number of IP far-response neurons that would reach disparity, blur) are precisely those that are required for thethreshold (T) as viewing distance is decreased. Data points were fitted with
roles that have generally been ascribed to the cerebelluma logistic function of formsuch as the continuous adjustment of movement parameters,predictive control, and long-term adaptation (see Stein andy Å (a 0 b)
1 / (x /c) d/ b
Glickstein 1992 for a review).where a is maximum y value, b is minimum y value, c is inflection point,and d is a slope function. Function shown here had following parameters: Latency of vergence and accommodation responses toa Å 73, b Å 04.0, c Å 9.6, d Å 4.4. electrical microstimulation of IP
The latency to elicit vergence changes from the IP of9.5 MA of convergence half the population of neurons is 15–20 ms is consistent with the shortest latency of 13 msinactive. The entire population is inactive by 18 MA. As required to elicit convergence from electrical microstimula-discussed below, this decrease in number of active far-re- tion of the midbrain near-response region (Judge and Cum-sponse neurons with increases in vergence angle may have ming 1986). Similarly, the latency to elicit an accommoda-important consequences for vergence control. tive response of 80–90 ms from IP stimulation is consistentwith the average latency of 75 ms reported for stimulation
D I S C U S S I O N of the Edinger-Westphal nucleus (Gamlin et al. 1994) andthe shortest latency of 75 ms reported for stimulation of the
To our knowledge, this is the first detailed report of the midbrain near-response region (Judge and Cumming 1986).steady-state activity of neurons in the posterior interposednucleus of the cerebellum in the alert, behaving primate Relationship to single-unit studies of primate midbrainduring vergence and ocular accommodation. Previous re- neuronsports linking cerebellar activity to these oculomotor func-
Previous studies have reported that the activity of Ç75%tions have been conducted in paralyzed, anesthetized catsof the neurons in the midbrain near-response region is corre-(e.g., Bando et al. 1980; Hosoba et al. 1978). Our results
raise a number of important questions regarding the role ofthe posterior interposed nucleus in the eye movements
Location and connections of IP far-response neurons
The region of the IP from which we have recorded far-response neurons coincides closely with the region of thenucleus that we have previously shown, using anterogradeand retrograde techniques, to be reciprocally connected withthe midbrain near-response region (May et al. 1992). Thisregion of the interposed nucleus is also reported to receiveinputs from the nucleus reticularis tegmenti pontis (NRTP)(Mihailoff 1993; Noda et al. 1990) and from the intermedi-ate cerebellar cortex (e.g., Bishop et al. 1979; Haines andRubertone 1979). Neurons within this region of the inter-posed nucleus project to the NRTP (e.g., Noda et al. 1990),to the midbrain near-response region (May et al. 1992) and
FIG. 15. Summary diagram of known efferent and afferent connectionspresumably to the ventrolateral thalamic nucleus (e.g., of posterior interposed nucleus potentially involved in vergence and accom-McCrea et al. 1978; Tolbert et al. 1977). The afferent and modation. EW, Edinger-Westphal nucleus; MRMN, medial rectus motoneu-
rons; SOA, Supraoculomotor area (midbrain near-response region).efferent connections of IP that are relevant to this discussion
J603-6/ 9k26$$mr05 02-06-98 08:58:49 neupa LP-Neurophys
by 10.220.33.4 on Septem
ber 27, 2016http://jn.physiology.org/
Dow
nloaded from

FAR-RESPONSE NEURONS IN THE POSTERIOR INTERPOSED NUCLEUS 1267
lated with the near-response, whereas the activity of Ç25% that, in the present study, the majority of vergence-only neu-rons are related to the vergence output pathway and theis related to the far-response (Judge and Cumming 1986;
Mays 1984). Just as is the case for the IP far-response neu- majority of accommodation-only neurons are related to theaccommodative output pathway.rons, neither the midbrain near-response or far-response neu-
rons display a conjugate signal. They are active for both Another feature of midbrain near-response neurons thatmay be explained by our data is the tendency for someblur-driven and disparity-driven movements as well as for
normal binocular movements. Figure 16 compares the kda of these neurons to show ‘‘soft-saturation’’ with increasingvergence angles (Morley et al 1992). As discussed below,and kdv values from the IP far-response neurons with those
reported for midbrain near-response neurons (Zhang et al. these near-response cells may be operating in a push-pullmode with far-response cells. If the activity of the population1992). The distributions form mirror images of one another
that only differ in the absolute values of these two coeffi- of far-response cells increasingly decreases above 5 MA (asmight be suggested by Fig. 14), then near-response cellscients. This quantitative difference is to be expected because
the sensitivity of IP far-response neurons to accommodation would have to decreasingly increase their activity in orderto produce the same degree of change in vergence angle.and vergence is, on average, approximately three times less
than that of midbrain near-response neurons i.e., on average This would appear as soft-saturation in the activity of thenear-response neurons with increasing vergence angles.kdv and kda values are three times higher for midbrain near-
response neurons than for IP far-response neurons. Also, therelative numbers of neurons related to vergence as opposed Potential roles for the cerebellum in the control ofto accommodation are approximately the same between the vergence and accommodationtwo populations of cells. As pointed out by Zhang and col-
The region of the IP from which we have recorded projectsleagues (1992), the behavior of the near-response cells dur-to the midbrain near-response region. As stated above,ing conflict viewing depends not only on whether or notÇ25% of neurons in the midbrain region are far-responsethey are in the vergence or accommodation output pathwaysneurons. If the projections from IP far-response neuronsbut also on their relative blur and disparity inputs. Thesewere to the midbrain far-response neurons then these connec-authors demonstrated that a cell identified as a vergence-tions could be directly excitatory. Alternatively, the IP far-only cell by conflict viewing might in fact be in the accom-response neurons could have an inhibitory influence on near-modative output pathway if the blur and disparity controllerresponse neurons in this midbrain region. We have recentlyinputs to it were sufficiently mismatched. However, Morleyreported that a region of the posterior fastigial nucleus con-and colleagues (1992) pointed out that, for midbrain near-tains near-response neurons (Zhang and Gamlin 1994). Be-response neurons to be ascribed to the wrong output path-cause this region of the fastigial nucleus also projects to theway, large and specific mismatches between the blur andmidbrain near-response region (May et al. 1992), it is possi-disparity controller inputs would be required, such that thisble that the fastigial near-response region and the interposedwould be a rare occurrence. Although our study of IP far-far-response region represent a push-pull system for modu-response neurons found more cells with mismatches betweenlating vergence and accommodation. Importantly, increasestheir kb and kd values than were reported for midbrain neu-in the tonic activity of the vergence-related cells of the cere-rons, these mismatches were not sufficiently large or specificbellar near-response region combined with decreases in thethat many IP far-response neurons would have been ascribedtonic activity of vergence-related cells of the cerebellar far-to the wrong output pathway. It is therefore safe to assumeresponse region would result in an overall increase in theconvergence signal, but not the accommodation signal, im-pinging on the midbrain near-response region. A signal ofthis nature could underlie the ability of individuals to showphoria adaptation and would be consistent with one reportthat indicated that cerebellar damage compromises this abil-ity (Milder and Reinecke 1983), although this was not con-firmed in a later study (Hain and Luebke 1990)
Relationship to studies in cats
Previous studies in cats have reported that both the fastig-ial and interposed nucleus are related to positive accommo-dation (e.g., Bando et al. 1979a,b; Hosoba et al. 1978).Because the animals in these studies were paralyzed andanesthetized, the influence of these cerebellar nuclei onvergence eye movements could not be evaluated. It is possi-ble that slight positive accommodation was obtained withmicrostimulation of the interposed nucleus because of cur-rent spread to the fastigial nucleus or activation of fibersfrom this nucleus. Alternatively, the small amount (Ç0.2
FIG. 16. Scatterplot comparing kda versus kdv values for sample of 70D) of positive accommodation that was obtained with micro-IP far-response neurons in this study (n) with those from a sample of
midbrain near-response neurons (s) (from Zhang et al. 1992). stimulation of the interposed nucleus in cats can be interpre-
J603-6/ 9k26$$mr05 02-06-98 08:58:49 neupa LP-Neurophys
by 10.220.33.4 on Septem
ber 27, 2016http://jn.physiology.org/
Dow
nloaded from

H. ZHANG AND P.D.R. GAMLIN1268
This research was supported by National Eye Institute Grant R01 EY-ted in light of our results in the primate. We found that the07558 to P.D.R. Gamlin and by P30 EY-03039.eyes must be converged and accommodating for electrical
Address for reprint requests: P.D.R. Gamlin, Vision Science Researchmicrostimulation of the interposed nucleus to elicit clear Center, University of Alabama at Birmingham, Birmingham, AL 35294.divergence and accommodation for far. Given the physiolog-
Received 30 July 1996; accepted in final form November 4, 1997.ical state of the cats in the studies described above, it isunlikely that they were accommodating or that their eyes
REFERENCESwere converged. In our studies, electrical microstimulationBANDO, T., ISHIHARA, A., AND TSUKAHARA, N. Interpositus neurons control-of the IP under these conditions had no clear effect and the
ling lens accommodation. Proc. Japan Acad. 55: 153–156, 1979a.very small positive accommodative responses obtained inBANDO, T., ISHIHARA, A., AND TSUKAHARA, N. The mode of cerebellar
cats under comparable conditions may therefore not have control of lens accommodation. In: Integrative Control Functions ofbeen functionally relevant. For these same reasons, the spon- Brain, edited by M. Ito, N. Tsukahara, K. Kubota, and K. Yogi. 2: 149–
150, 1979b.taneous activity of IP neurons in cats that apparently corre-BISHOP, G. A., MCCREA, R. A., LIGHTHALL, J. W., AND KITAI, S. T. Anlated with positive accommodation (Bando et al. 1979a) is
HRP and autoradiographic study of the projection from the cerebellarhard to interpret. cortex to the nucleus interpositus anterior and nucleus interpositus poste-rior of the cat. J. Comp. Neurol. 185: 735–756, 1979.
Relationship to previous studies BUTTNER, U. AND WAESPE, W. Purkinje cell activity in the primate flocculusduring optokinetic stimulation, smooth pursuit eye movements and VOR-
The region of the IP from which we have recorded far- suppression. Exp. Brain Res. 55: 97–104, 1984.response neurons has recently been reported by Van Kan BUTTNER-ENNEVER, J. A. AND BUTTNER, U. The reticular formation. In:
Neuroanatomy of the Oculomotor System, edited by J. A. Buttner-En-and colleagues (1993) to contain neurons related to eyenever. Amsterdam: Elsevier, p. 119–202, 1988.movements, but the specific eye movement to which these
CHARMAN, W. N. AND TUCKER, J. Dependence of accommodation responsecells were related was not investigated. Apart from the far- on the spatial frequency spectrum of the observed object. Vision Res. 17:response neurons reported here, we have not examined in 129–139, 1977.
CHUBB, M. C. AND FUCHS, A. F. Contribution of y group of vestibular nucleiany detail the activity of other eye movement related neuronsand dentate nucleus of cerebellum to generation of vertical smooth eyein IP, but we have encountered some saccade-related andmovements. J. Neurophysiol. 48: 75–99, 1982.blink-related neurons within this region, and have found that CRANE, H. D. AND CLARK, M. R. Three-dimensional visual stimulus deflec-
electrical microstimulation of it often elicits small saccades. tor. Appl. Opt. 17: 706–714, 1978.Previous studies of oculomotor control by the deep cere- CUMMING, B. G. AND JUDGE, S. J. Disparity-induced and Blur-induced con-
vergence eye movement and accommodation in the monkey. J. Neuro-bellar nuclei have concentrated on the role of the oculomotorphysiol. 55: 896–914, 1986.region of the fastigial nucleus (e.g., Fuchs et al. 1993, 1994;
FINCHAM, E. F. AND WALTON, J. The reciprocal actions of accommodationNoda et al. 1988; Ohtsuka and Noda 1991). Single-unit and vergence. J. Physiol. (Lond.) 137: 488–508, 1957.recording studies of the role of the fastigial nucleus in oculo- FUCHS, A. F. AND ROBINSON, D. A. A method for measuring horizontal and
vertical eye movement chronically in the monkey. J. Applied Physiol.motor control have reported that the tonic activity of sac-21: 1068–1070, 1966.cade-related fastigial neurons is very weakly correlated with
FUCHS, A. F., ROBINSON, F. R., AND STRAUBE, A. Participation of the caudaleye position and that these neurons, at best, show a relation- fastigial nucleus in smooth-pursuit eye movements. I. Neuronal activity.ship of approximately 0.4 spikes/second per degree of J. Neurophysiol. 72: 2714–2728, 1994.change in eye position (Fuchs et al. 1993). Furthermore, FUCHS, A. F., ROBINSON, F. R., AND STRAUBE, A. Role of the caudal fastigial
nucleus in saccade generation. I. Neuronal discharge pattern. J. Neuro-those fastigial neurons related to smooth pursuit eye move-physiol. 70: 1723–1740, 1993.ments are not reported to have any eye position sensitivity
GAMLIN, P.D.R. Neural control of vergence and accommodation: a cerebel-(Fuchs et al. 1994). In contrast, as shown by our regression lar ‘‘far response’’ region. Assoc. Res. Vis. Ophthalmol. 32, Suppl. : 1125,analyses, the tonic activity of many far-response neurons 1991.
GAMLIN, P.D.R. AND CLARKE, R. J. Single-unit activity in the nucleus reticu-recorded from the IP correlates well with vergence anglelaris tegmenti pontis related to vergence and ocular accommodation. J.and accommodation. Although this finding is different fromNeurophysiol. 73: 2115–2119, 1995.the findings for fastigial neurons during conjugate eye move- GAMLIN, P.D.R., GNADT, J. W., AND MAYS, L. E. Lidocaine-induced unilat-
ments, it is not inconsistent with a report by Thach (1978) eral internuclear ophthalmoplegia: effects on convergence and conjugateon interpositus neuron activity during wrist flexion/exten- eye movements. J. Neurophysiol. 62: 82–95, 1989.
GAMLIN, P.D.R. AND YOON, K. Single-unit activity related to the near-sion. He found that the tonic activity of these neurons in-response in area 8 of the primate frontal cortex. Soc. Neurosci. Abstr.creased as a function of the force required to overcome21: 1918, 1995.either an extensor or flexor load. The force generated by the GAMLIN, P.D.R., K. YOON, AND ZHANG, H. The role of cerebro-ponto-
extraocular muscles and the ciliary muscle must change for cerebellar pathways in the control of vergence eye movements. Eye 10:167–171, 1996.divergence and accommodation for far, and the changes in
GAMLIN, P.D.R. AND ZHANG, H. Y. Effects of muscimol blockade of theactivity of interpositus neurons during the far-response mayposterior fastigial nucleus on vergence and ocular accommodation in thewell be related to these required changes in muscle force. primate. Soc. Neurosci. Abstr. 22: 1034, 1996.
Therefore, our results can be considered analogous to those GAMLIN, P.D.R., ZHANG, Y., CLENDANIEL, R. A., AND MAYS, L. E. Behaviorof Thach and they strongly suggest that for both types of of identified Edinger-Westphal neurons during ocular accommodation.
J. Neurophysiol. 72: 2368–2382, 1994.movements the tonic activity of interpositus neurons directlyHAIN, T. C. AND LUEBKE, A. E. Phoria adaptation in patients with cerebellaror indirectly contributes to the change in steady-state activity
dysfunction. Invest. Ophthalmol. Vis. Sci. 31: 1394–1397, 1990.of motoneurons that is required to match force generation HAINES, D. E. AND RUBERTONE, J. A. Cerebellar corticonuclear fibers of theto specific loads. dorsal culminate lobule (anterior lobe–lobule V) in a prosimian primate,
Galago senegalensis. J. Comp. Neurol. 186: 321–341, 1979.HOSOBA M., BANDO T., TSUKAHARA N. The cerebellar control of accommo-We thank S. Mason and K. Mitchell for technical assistance and S.
Hayley for computer programming. dation of the eye in the cat. Brain Res. 153: 495–505, 1978.
J603-6/ 9k26$$mr05 02-06-98 08:58:49 neupa LP-Neurophys
by 10.220.33.4 on Septem
ber 27, 2016http://jn.physiology.org/
Dow
nloaded from

FAR-RESPONSE NEURONS IN THE POSTERIOR INTERPOSED NUCLEUS 1269
HUNG, G. K. AND SEMMLOW, J. L. Static behavior of accommodation and MORLEY, J. W., JUDGE, S. J., AND LINDSEY, J. W. Role of monkey midbrainnear-response neurons in phoria adaptation. J. Neurophysiol. 67: 1475–vergence: computer simulation of an interactive dual-feedback system
IEEE Trans. Biomed. Eng. 29: 439–447, 1980. 1492, 1992.NODA, H., MURAKAMI, S., YAMADA, J., TAMADA, J., TAMAKI, Y., AND ASO,JUDGE S. J. AND CUMMING B. G. Neurons in the monkey midbrain with
T. Saccadic eye movements evoked by microstimulation of the fastigialactivity related to vergence eye movement and accommodation. J. Neuro-nucleus of macaque monkeys. J. Neurophysiol. 60: 1036–1052, 1988.physiol. 55: 915–930, 1986.
NODA, H., SUGITA, S., AND IKEDA, Y. Afferent and efferent connections ofJUDGE, S. J., RICHMOND, B. S., AND CHU, F. C. Implantation of magneticthe oculomotor region of the fastigial nucleus in the macaque monkey.search coils for measurement of eye position: an improved method. VisionJ. Comp. Neurol. 302: 330–348, 1990.Res. 20: 535–538, 1980.
OHTSUKA, K. AND NODA, H. Saccadic burst neurons in the oculomotorKASE, M., NODA, H., SUZUKI, D. A., AND MILLER, D. C. Target velocityregion of the fastigial nucleus of macaque monkeys. J. Neurophysiol.signals of visual tracking in vermal Purkinje cells of the monkey. Science65: 1422–1434, 1991.205: 717–720, 1979.
OPTICAN, L. M. AND ROBINSON, D. A. Cerebellar-dependent adaptive con-KATO, L., HARADA, K. NAKAMURA, T., SATO, Y., AND KAWASAKI, T. Roletrol of primate saccadic system. J. Neurophysiol. 44: 1058–1076, 1980.of the nucleus reticularis tegmenti pontis on visually induced eye move-
SCHOR, C. M. A dynamic model of cross-coupling between accommodationments. Exp. Neurol. 78: 503–516, 1982.and convergence: simulations of step and frequency responses. Optom.KAWASAKI, T., KIYOSAWA, M., FUJINO, T., AND TOKORO, T. Slow accommo-Vis. Sci. 69: 258–269, 1992.dation release with a cerebellar lesion. British J. Ophthalmol. 77: 678,
STEIN, J. F. AND GLICKSTEIN, M. Role of the cerebellum in visual guidance1993.of movement. Physiol. Rev. 967–1017: 1992.KELLER, E. L. The cerebellum. Rev. of Oculomotor Res. 3: 391–411, 1989.
SUZUKI, D. A. AND KELLER, E. L. The role of the posterior vermis of monkeyLEICHNETZ, G. R., SMITH, D. J., AND SPENCER R. F. Cortical projections tocerebellum in smooth-pursuit eye movement control. I. Eye and headthe paramedian tegmental and basilar pons in the monkey. J. Comp.movement-related activity. J. Neurophysiol. 59: 1–18, 1988.Neurol. 228: 388–408, 1984.
SUZUKI, D. A. AND KELLER, E. L. The role of the posterior vermis of monkeyLEIGH, R. J. AND ZEE, D. S. The Neurology of Eye Movements. Philadelphia,cerebellum in smooth-pursuit eye movement control. II. Target velocity-PA: F. A. Davis, 1991.related Purkinje cell activity. J. Neurophysiol. 59: 19–40, 1988.LEWIS, R. F. AND ZEE, D. S. Ocular motor disorders associated with cerebel-
THACH, W. T. Correlation of neural discharge with pattern and force oflar lesions: pathophysiology and topical localization. Rev. Neurol. (Paris)muscular activity, joint position, and direction of intended next movement149: 665–677, 1993.in motor cortex and cerebellum. J. Neurophysiol. 41: 654–676, 1978.LISBERGER, S. G. AND FUCHS, A. F. Role of primate flocculus during rapid
TOLBERT, D. L., BANTLI, H., AND BLOEDEL, J. R. The intracerebellarbehavioral modification of vestibuloocular reflex. I. Purkinje cell activitynucleocortical projection in a primate. Exp. Brain Res. 30: 425–434,during visually guided horizontal smooth-pursuit eye movements and1977.passive head rotation. J. Neurophysiol. 41: 733–763, 1978.
TSUETAKI, T. K. AND SCHOR, C. M. Clinical method for measuring adapta-MAY, P. J., PORTER, J. D., AND GAMLIN, P.D.R. Interconnections betweention of tonic accommodation and vergence accommodation. Am. J. Op-the cerebellum and midbrain near response regions. J. Comp. Neurol. tom. Physiol. Opt. 64: 437–449, 1987.
315: 98–116, 1992. VAN KAN, P. L., HOUK, J. C., AND GIBSON, A. R. Output organization ofMAYS, L. E. Neural control of vergence eye movements: convergence and intermediate cerebellum of the monkey. J. Neurophysiol. 69: 57–73,
divergence neurons in the midbrain. J. Neurophysiol. 51: 1091–1108, 1993.1984. WESTHEIMER, G. Amphetamine, barbiturates, and accommodative conver-
MAYS, L. E., PORTER, J. D., GAMLIN, P.D.R., AND TELLO, C. A. Neural gence. Arch. Ophthalmol. 70: 830–836, 1963.control of vergence eye movements: neurons encoding vergence velocity. WESTHEIMER, G. AND BLAIR, S. M. Oculomotor defects in cerebellectom-J. Neurophysiol. 56: 1007–1021, 1986. ized monkeys. Invest. Opthalmol. 12: 618–620, 1973.
MCCREA, R. A., BISHOP, G. A., AND KITAI, S. T. Morphological and electro- YAMADA, J. AND NODA, H. Afferent and efferent connections of the oculo-physiological characteristics of projection neurons in the nucleus inter- motor cerebellar vermis in the macaque monkey. J. Comp. Neurol. 265:positus of the cat cerebellum. J. Comp. Neurol. 181: 397–419, 1978. 224–241, 1987.
MIHAILOFF, G. A. Cerebellar nuclear projections from the basilar pontine ZHANG, H. Y. AND GAMLIN, P.D.R. Sensorimotor characteristics of far re-nuclei and nucleus reticularis tegmenti pontis as demonstrated with PHA- sponse neurons in the cerebellum of the rhesus monkey. Assoc. Res. Vis.L tracing in the rat. J. Comp. Neurol. 330: 130–146, 1993. Ophthalmol. 35, Suppl. : 1282, 1994.
MILDER, D. G. AND REINECKE, R. D. Phoria adaptation to prisms: a cerebel- ZHANG, H. Y. AND GAMLIN, P.D.R. Single-unit activity within the posteriorlar-dependent response. Arch. Neurol. 40: 339–342, 1983. fastigial nucleus during vergence and accommodation in the alert primate.
MILES, F. A., FULLER, J. H., BRAITMAN, D. J., AND DOW, B. M. Long-term Soc. Neurosci. Abstr. 22: 2034, 1996.adaptive changes in primate vestibuloocular reflex. III. Electrophysiologi- ZHANG, Y., MAYS, L. E., AND GAMLIN, P.D.R. Characteristics of near re-cal observations in flocculus of normal monkeys. J. Neurophysiol. 43: sponse cells projecting to the oculomotor nucleus. J. Neurophysiol. 67:
944–960, 1992.1437–1476, 1980.
J603-6/ 9k26$$mr05 02-06-98 08:58:49 neupa LP-Neurophys
by 10.220.33.4 on Septem
ber 27, 2016http://jn.physiology.org/
Dow
nloaded from