Embed Size (px)
Citation preview

Zeitschrift ffir Zellforschung 7 1 , 4 8 9 ~ 9 8 (1966)
NEURONAL LOCALIZATION OF DOPAMINE AND 5-HYDROXYTRYPTAMINE IN SOME MOLLUSCA*
ERIK DAItL, BENGT FALCK, CLAES VON MECKLENBURG, HARRY MYHRBERG,
and EVALD ROSENGREN Departments of Zoology, Histology, and Pharmacology, University of Lund
Sweden
Received November 18, 1965
Summary. The localization of biogenic monoamines in ganglionic tissues from Anodonta piscinalis, Helix pomatia, and Buccinum undatum has been studied by means of the histochemi- cal fluorescence method of FALCK and HILLARP.
In cerebral, visceral, and pedal ganglia (besides nonfluorescent nerve cells) neurons emitting a green or yellow fluorescence were found. No other cell systems exhibiting a specific fluorescence were observed. An abundance of monoaminergic terminals were found in the cen- tral parts of these ganglia. Speetrophotofluorimetric determinations showed that there are large quantities of dopamine and 5-hydroxytryptamine in the tissues investigated. The amounts of dopamine and 5-hydroxytryptamine agree well with the distribution of green and yellow fluorescence, respectively, in the ganglia.
There are many similarities between the vertebrate and the molluscan monoaminergic neurons. The morphology of the neurons is the same, the intraneuronal distribution of the monoamines is identical, depletion experiments with reserpine and denervation experiments give the same results, and the synaptic arrangement of monoaminergic fibres on non-adrenergic neurons has the same appearance. Apparently, however, dopamine and 5-hydroxytryptamine are the only monoamines acting as neuronal transmitters in the species investigated.
Introduction
By means of the his tochemical fluorescence me thod of FALCK and HILLARP
(FALCK, 1962a; FALCK et al., 1962; CORRODI and HILLARP, 1963, 1964), dopamine
(DA) and 5 -hydroxy t ryp tamine (5-HT) were shown to occur in neurons in molluscs
(FALCK, 1962a and b; DAHL et al., 1962; DAHL, 1963; DAHL et al., 1963a). I t was
demons t ra ted tha t DA - - probably as the only catecholamine - - occurs in large
amounts in mollusc gangha (cf. also SWEENEY, 1963). In this paper a more detai led
repor t on the localization of these amines is given.
Material and Methods Cerebral, pedal, and visceral ganglia from the fresh-water-bivalve Anodonta piscinalis as
well as cerebral and suboesophageal ganglia from the puhnonate snail Helix pomatia were used. To some extent tissues from the prosobranch Buccinum undatum were also examined. The ganglia from A. piscinalis were dissected out from living animals;the snails were dissected after decapitation. After removal the ganglia were immediately frozen in a propane-propylene mixture, cooled by liquid nitrogen, and thereafter freeze-dried and treated with formaldehyde gas, embedded and sectioned for fluorescence microscopy (see FALCK and 0WMA>r, 1965a). The catecholamines and 5-HT condense with formaldehyde to intensely fluorescent products. The fluorophores of the catecholamines emit a green or sometimes green to yellow-green light while the condensation product of 5-HT has a yellow fluorescence.
* The research reported in this document has been sponsored by the Air Force Office of Scientific Research under Grant AF EOAR 64-5 through the European Office of Aerospace Research (OAR), United States Air Force and by the Swedish Natural Science Research Council.
z. Zellforsch., Bd. 71 32

490 E. DAHL, B. FALCK, C. V. MECKLENBURG, H. MYHRBERG, and E. ROSENGREN:
The spectrophotofluorimetric determinations of the amines were carried out according to BERTLER et al. (1958) and BERTLER and ROSENGREN (1959).
Observations
Fluorescence Microscopy o~ Normal Tissues Anodonta piscinalis, Cerebral Ganglion. The formaldehyde t rea tment developed
a moderate yellow fluorescence in a large number of nerve cells with short pro- cesses, which were seen to fluoresce very weakly in comparison with the fluores- cence of the cell body. The cells were distributed th roughout the cortical layer, often lying closely together in groups consisting of up to ten cells. Within these groups the cells were often arranged in radial rows. These rows often extended from the connective neuri lemma throughout the cortex but never into the central neuropil.
Scattered cells in the cortical layer also showed a green fluorescence of moder- ate to strong intensity. These cells had the same general appearance as the yellow cells, except t ha t the beginning of their processes were seen to fluoresce more clearly for a short distance from the cell body. In good preparations such processes could sometimes be followed a rather long distance in the tissue - - this par t of the axon then had only a barely visible greenish fluorescence and no varicosities. The green fluorescence and the reaction conditions under which it was developed suggest the presence of a pr imary catecholaminc (cf. FALCK and OWMAN, 1965a).
I n the cortical layer there were also numerous nerve cells, which did not show any specific fluorescence but often contained more or less coarse granules showing a yellow-orange autofluorescence. Such autofluorescent granules were also loca- ted elsewhere in the ganglia but could readily be distinguished from the specific fluorescence. These granules which occur also in cells exhibiting a specific fluorescence are paraldehyde-fuchsin-positive.
I n the central par t of the ganglion, which in Anodonta is filled with nerve fibres, a ra ther strong yellow fluorescence developed in numerous small granules which in m a n y instances were arranged in rows. The whole picture suggested the existence of rather closely packed varicose nerve fibres of the same morphology as the axons running in the mammal ian autonomic ground plexus. This has been confirmed in other experiments (cf. depletion and dcnervat ion experiments below).
However, the dominat ing fluorescent structure in the central par t consisted of fine varicose fibres showing a green fluorescence. Their varicosities showed an intense fluorescence whereas between the varicosities the axon had only a weak fluorescence, sometimes hardly visible. I n m a n y cases connections between presumed varicosities could not be seen. The green fibres were often arranged in distinct tracts. Varicose green and yellow fibres were sometimes seen to enter the cortical layer.
Smooth, fine fibres with a weak g r e e n - - but never ye l l ow- - fluorescence occurred in the big nerves, the proximal parts of which were dissected out together with the ganglia. Sometimes these fibres were seen to pass over into the varicose fibre type.
I n some instances green smooth axons from green cells in the cortical layer could be seen entering smaller nerves and traced for some distance within them. Smaller nerves leaving the cerebral ganglion sometimes contained numerous green and yellow varicose fibres.
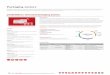
Dopamine and 5-Hydroxytryptamine in some Mollusca 491
Fig. 1. Densely packed varicose, monoaminergie fibres in the central neuropil in the visceral ganglion of Anodonta piscinalis. 400 •
A. piscinalis, Visceral Ganglion. The majority of the ganglion cells contained only autofiuorescent granules. A specific green fluorescence developed in nerve cells resembling those in the cerebral ganglion, but more numerous. The central part, which is more voluminous than that of the cerebral ganglion, contained an enormous number of varicose green fibres and numerous yellowish granules arranged in the same manner as in the cerebral ganglion (Fig. 1). These were more scattered than in the cerebral ganglion giving the impression that their number did not exceed or was even less than that of the yellow granules in the cerebral ganglion. As in the cerebral ganglion, varicose fibres were found in the cortical layer.
A. piscinalis, Pedal Ganglion. The neuropil consisted of a dense tangle of green fluorescent fibres even more numerous than in the visceral ganglion. Besides non- fluorescent nerve cells, the cortical layer contained only yellow fluorescent cells whose processes could be followed for a very short distance but were lost in the green centraI fibre mass.
Helix pomatia. The whole ganglionic ring consisting of the cerebral, visceral, pleural, and pedal ganglia was dissected out. A specific green or yellow fluorescence developed in such a rich variety of cells and fibre arrangements that only a summary of the more important observations will be presented here.
The fluorescent cells varied considerably in size and shape. The most common and most intensely fluorescent types were formed of very small green and yellow cells in closely packed groups of either type. The processes of the yellow cells were more or less inconspicuous, whereas the axons originating from the green cells
32*
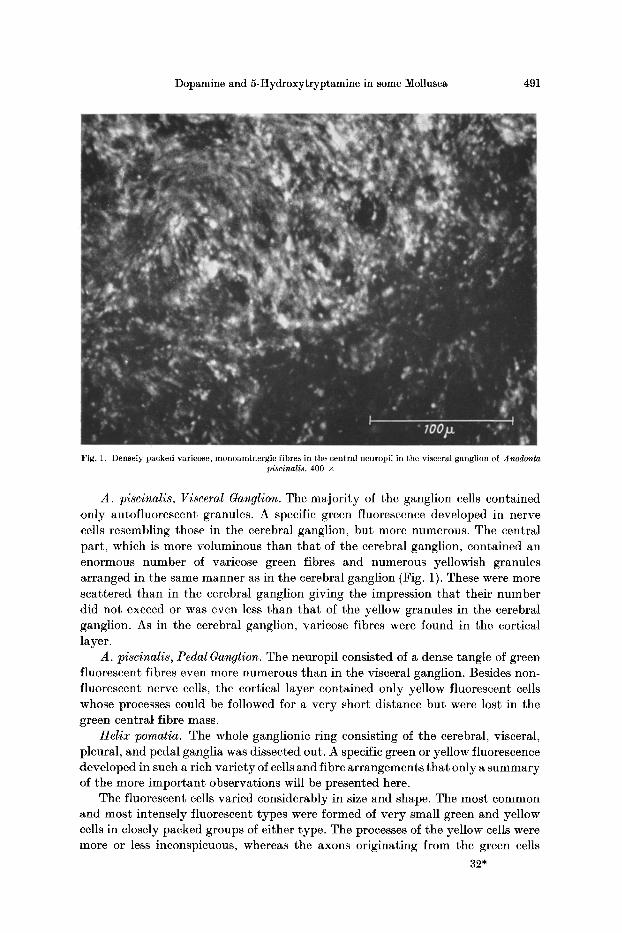
492 E. DAHL, B. FALCK, C. V. MECKLENBURG, H. MYttRBERG, and E. ROSE•OREN:
Fig. 2. A group of small dopamine-containing neurons from the cerebral ganglion of Helix pomatia. The pretermi- nals form a short bundle with comparatively weak fluorescence but are rapidly transformed into a dense mass of
intensely fluorescent varicose terminals. 360 •
could often be followed for a certain distance. These axons consistently showed a weak fluorescence compared with the cell body. Such a cell group in the cerebral ganglion exhibited an interesting distribution of the fluorescent material in the neurons (Fig. 2). Here the axons left the cell mass in a dense bundle which fluores- ced weakly. After a short distance a sudden change was seen. The fibre bundle then assumed an intense fluorescence and dissolved into more spaciously distributed varicose fibres. The significance of this observation will be discussed below. The green and yellow groups of small cells were located in the vicinity of rather large areas of extremely tightly packed fibres which fluoresced in a green of very high intensity making it impossible to recognize the morphology of the individual fibres. No doubt a great many fibres, perhaps all, were beaded for after partial depletion of the fluorescence with low doses of reserpine all fibres in this area appeared to be of the varicose type. Yellow varicose fibres were observed in the immediate neighbourhood of the green fibre masses. No nerve cells, whether fluorescent or non-fluorescent, could be detected within the fibre masses, which, on the other hand, seemed to be mixed with a considerable number of non- fluorescent fibres. The picture suggests that the fibre arrangement belongs to the neuropil (HANSTROM, 1928).
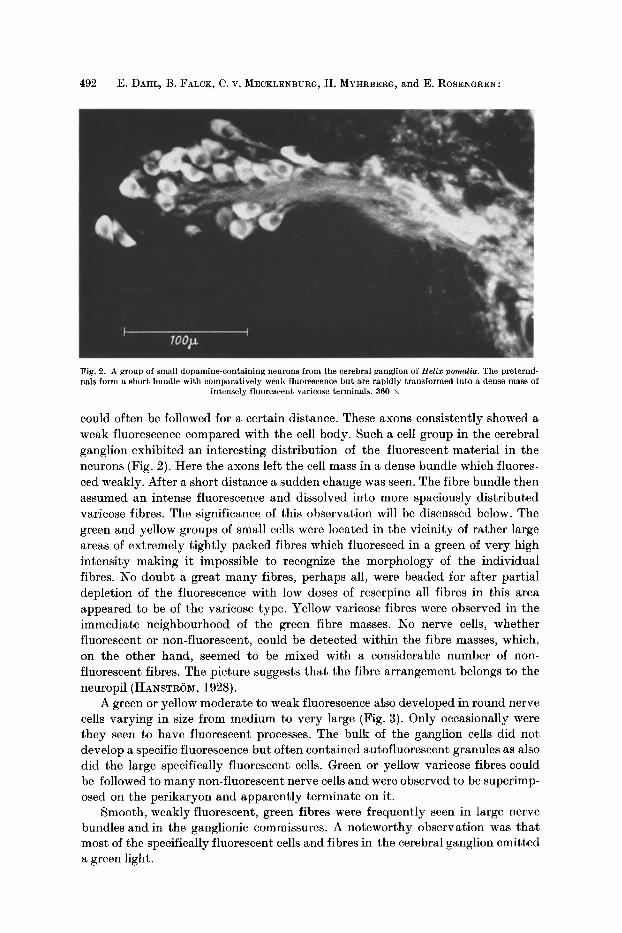
A green or yellow moderate to weak fluorescence also developed in round nerve cells varying in size from medium to very large (Fig. 3). Only occasionally were they seen to have fluorescent processes. The bulk of the ganglion cells did not develop a specific fluorescence but often contained autofluorescent granules as also did the large specifically fluorescent cells. Green or yellow varicose fibres could be followed to many non-fluorescent nerve cells and were observed to be superimp- osed on the perikaryon and apparently terminate on it.
Smooth, weakly fluorescent, green fibres were frequently seen in large nerve bundles and in the ganglionic commissures. A noteworthy observation was that most of the specifically fluorescent cells and fibres in the cerebral ganglion emitted a green light.
Dopamine and 5-Hydroxytryptamine in some Mollusca 493
Fig. 3. A large dopamine-conta in ing per ikaryon in the cerebral gangl ion of Helix pomatia. The small bodies are auto-f luorescent s t ruc tures and varicose terminals out of focus. 1135 •
Buccinum undatum. In a current investigation of the cerebral ganglion from the prosobranch marine snail Buccinum undatum, fluorescence microscopic fin- dings similar to the above-mentioned were made. An interesting finding was that numerous fibres in the optical nerve exhibited a relatively strong green fluores- cence. These fibres were smooth except in the vicinity of the eye bulb where some of the fibres had transformed into the varicose type. They spread out to form a net-work of fibres in the subepithelial layer around the eye bulb but did not pene- trate into the retina. Sparsely distributed in the tentacular tissue appeared also fine varicose green fibres. Also green fluorescence was seen in varicose elongated cells located under certain epithelial areas. These cells were provided with two delicate processes, one penetrating into the epithelial lining and the other directed towards the central part of the tentacle. Thus, they appear to belong to the adrenergic sensory cell system previously described (DAHL et al., 1963a and b). Their connections with the central nervous system are not yet clear.
Spectrophoto/luorimetric Determinations
The characteristics of the reaction and the colour of the emitted light suggest that the observed fluorescence in the tissues is due to the presence of amines be- longing to both the catecholamine and tryptamine groups. The conditions under which the green fluorescence developed seem to exclude the occurrence of secon- dary catecholamines. To further evaluate the above-mentioned observations, spectrophotofluorimetric determinations were carried out (partly performed in collaboration with M. LINDQUIST, Department of Pharmacology, Gothenburg; cf. DAHL et al., 1962). In the catecholamine analyses little or no adrenaline and noradrenaline was found. However, a compound was detected which behaved as DA on the ion exchange column and in spectrophotofluorimetric determinations.
In order to further establish the identity of this catecholamine two extracts from pooled cerebral and visceral ganglia of Anodonta piscinalis were provided
494 E. DAHL, B. FALGK, C. V. MECKLENBURG, H. Mrm~BERO, and E. ROSENGREN:
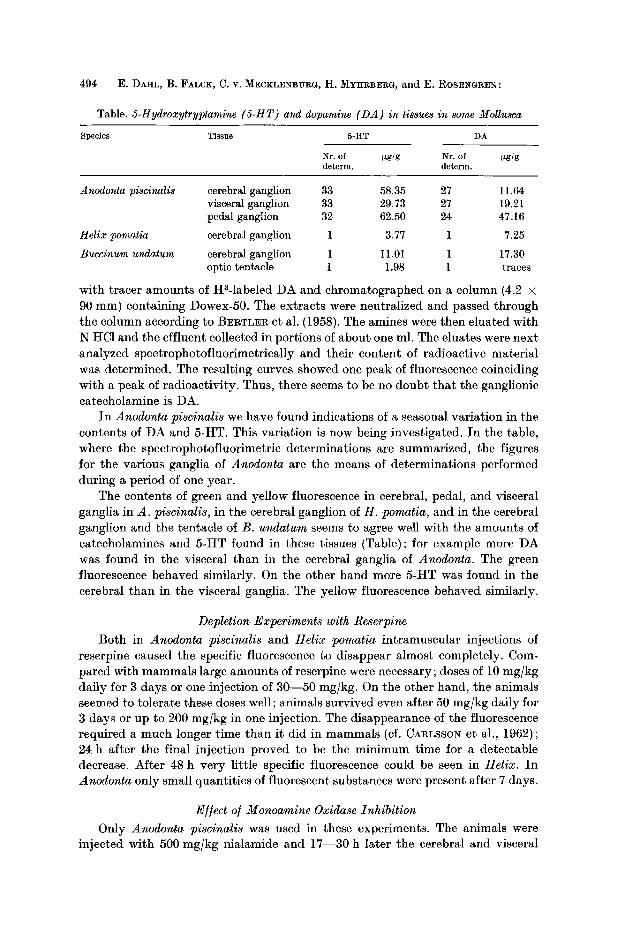
Table. 5-Hydroxytryptamine (5-HT) and dopamine (DA) in tissues in some Mollusca
Species Tissue 5-HT DA
Nr. of ~zg/g Nr. of ~g/g determ, determ.
Anodonta piscinalis cerebral ganglion 33 58.35 27 11.64 visceral ganglion 33 29.73 27 19.21 pedal ganglion 32 62.50 24 47.16
Helix pomatia cerebral ganglion 1 3.77 1 7.25
Buccinum undatum cerebral ganglion 1 I 1.01 1 17.30 optic tentacle 1 1.98 1 traces
with tracer amounts of Ha-labeled DA and chromatographed on a column (4.2 • 90 mm) containing Dowex-50. The extracts were neutralized and passed through the column according to BERTLER et al. (1958). The amines were then eluated with N HC1 and the effluent collected in portions of about one ml. The eluates were next analyzed spectrophotofluorimetrically and their content of radioactive material was determined. The resulting curves showed one peak of fluorescence coinciding with a peak of radioactivity. Thus, there seems to be no doubt that the ganglionic catecholamine is DA.
In Anodonta piscinalis we have found indications of a seasonal variation in the contents of DA and 5-HT. This variation is now being investigated. In the table, where the spectrophotofluorimetric determinations are summarized, the figures for the various ganglia of Anodonta are the means of determinations performed during a period of one year.
The contents of green and yellow fluorescence in cerebral, pedal, and visceral ganglia in A. piscinalis, in the cerebral ganglion of H. pomatia, and in the cerebral ganglion and the tentacle of B. undatum seems to agree well with the amounts of catecholamines and 5-HT found in these tissues (Table); for example more DA was found in the visceral than in the cerebral ganglia of Anodonta. The green fluorescence behaved similarly. On the other hand more 5-HT was found in the cerebral than in the visceral ganglia. The yellow fluorescence behaved similarly.
Depletion Experiments with Reserpine
Both in Anodonta piscinalis and Helix pomatia intramuscular injections of reserpine caused the specific fluorescence to disappear almost completely. Com- pared with mammals large amounts of reserpine were necessary ; doses of 10 mg/kg daily for 3 days or one injection of 30---50 mg/kg. On the other hand, the animals seemed to tolerate these doses well ; animals survived even after 50 mg/kg daily for 3 days or up to 200 mg/kg in one injection. The disappearance of the fluorescence required a much longer time than it did in mammals (cf. CARLSSON et al., 1962); 24 h after the final injection proved to be the minimum time for a detectable decrease. After 48 h very little specific fluorescence could be seen in Helix. In Anodonta only small quantities of fluorescent substances were present after 7 days.
E/feet o/Monoamine Oxidase Inhibition
Only Anodonta piseinalis was used in these experiments. The animals were injected with 500 mg/kg nialamide and 17--30 h later the cerebral and visceral

Dopamine and 5-Hydroxytryptamine in some Mollusca 495
ganglia were excised. This t rea tment in most specimens produced a distinct in- crease of the fluorescence in the green fibres but not in the green cell bodies. In some cases the yellow fibres in the cerebral and visceral ganglia showed a considera- bly enhanced fluorescence intensity, which also held true in normal animals for the thin and often invisible parts of the axons between the varicosities. This finding confirmed tha t the yellow granules and sometimes also the green ones occurring in normal ganglia and often arranged in rows, represent true varicosities of nerve fibres. For unknown reason this enhancement of fluorescence could not be repro- duced consistently.
Denervation Experiments As yet only the effects of severing of the cerebro-visceral connective (i. e. the
nerve trunk connecting the cerebral and visceral ganglia) in the vicinity of the cerebral ganglion have been investigated. All specimens submitted to bilateral sectioning of the connectives died within few days, but the survival rate after unilateral operation was well above 50 %.
The cerebral and visceral ganglia on the normal and the operated sides were excised 12 days after the operation. In the cerebral ganglion, whose connective had been cut, most of the green cells and fibres had disappeared. The yellow neu- rons in some specimens appeared unchanged, in others a considerable accumulation of fluorescent material was observed in the fibre parts situated proximal to the lesion. In a few cases this increase also appeared in the yellow cell bodies in the ganglion cortex. In the visceral ganglion on the same side all the yellow fluores- cence and a large part of the green varicose fibres had disappeared.
Discussion
I t has previously been shown tha t some invertebrate nervous systems contain high amounts of 5-HT (FLogEY and FLOREY, 1954; WELSh, 1954), especially those of the molluscs (WELSH and MOORHEAD, 1960). These observations were recently confirmed by using ganglionic tissues from Anodonta piscinalis, Helix pomatia, a n d Buecinum undatum, and for the first t ime the presence of high amounts of DA, but probably no other catecholamine, was simultaneously demonstrated; both amines were shown to be localized in neurons (FALCK, 1962a and b; DAHL et al., 1962; DA~IL, 1963; DAHL et al., 1963a). The presence of DA in molluscan nervous tissue was independently demonstrated by SWE~N]~Y (1963) and CARDOT (1963) with fluorimetric and paper chromatographic methods.
The fluorescence method for cellular localization of monoamines has a high specificity for certain catecholamines and tryptamines. This and the fact tha t of amines belonging to these groups only 5-HT and DA are present in significant amounts in the investigated tissues strongly indicates tha t the fluorescent material demonstrates the localization of these amines in nerve cells and nerve fibres. Furthermore, the distribution of green and yellow fluorescent material agrees well with the content of DA and 5-HT in the different tissues. The disappearance of the specific fluorescence in animals treated with reserpine also shows that the fluores- cent material is due to the presence of such amines.
While these facts conclusively prove tha t 5-HT and DA are localized in mollus- can neurons, they do not prove tha t these amines act as transmitters. Various observations, however, indicate that this might be the case:

496 E. D~L, B. FALCK, C. V. MECKLENBURG, H. MYHRBERG, and E. ROSENGREN:
1. I t is clearly established (cf. v. EULER, 1961) that noradrenaline is a sympa- thetic transmitter. That this is not an exclusive property of this biogenic mono- amine was demonstrated by FALCK et al. (1963) who found adrenaline-adrenergic nerves in the frog. The mammalian brain contains extensive system of neurons carrying DA and 5-I tT (CARLSSON et al., 1962; ANDI~N et al., 1964; BI~RTLER et al., 1964; CARLSSON et al., 1964; DAHLSTRSM and FuxE, 1964). I f the molluscs are supplied with adrenergic nervous systems, the t ransmit ter cannot be noradrena- line or adrenaline since no significant amounts of these amines are present in the ganglia. Neuronal localization of 5-HT is not only found in molluscs but also in numerous other groups, e.g. oligochaetes (MYHI~BERO and ROSENGREN, unpublis- hed observations), cyclostomes, teleosts, and amphibians (DAHL, FALCK, V. MECK- LENBVRO, MYHRBERG, and I~OSENG~N, unpublished observations).
2. In adrenergic systems working with noradrenaline and adrenaline as trans- mitters, these, like DA and 5-HT in molluscan neurons, are located within the neu- rons (v. EULER, 1961; FALCK, 1962a; FALCK and ToRe, 1962; FALCK et al., 1963) and not in some extraneuronal stores such as chromaffin cells. Noradrenaline in adrenergic nerves is present in the whole neuron but accumulates mostly in its terminal par t (v. EULER, 1956, 1961; FALCK, 1962a; FALCK and TORP, 1962). The same intraneuronal distribution has been observed in this investigation for both 5-HT and DA. The axons could usually only be observed - - if a t all - - for a very short distance from the cell body and the green fibres in larger nerves showed at most a weak fluorescence; yellow fibres in such nerves were not seen at all. On the other hand, the fibres which apparently represent the varicose terminal parts of the neurons exhibit a strong fluorescence intensity. In Hel ix small cell groups were actually observed giving off weakly fluorescent smooth fibres which after a short distance changed into varicose, strongly fluorescent fibres (Fig. 2) terminat- ing in an area probably belonging to the neuropfl (which is known to contain numerous synapses between nerve fibres).
Thus a comparison with vertebrate adrenergic neurons reveals many similari- ties:
1. The morphology of the neurons is the same in the respect tha t the axons consist of a smooth preterminal part and a varicose terminal part (FALoK, 1962a; NORBERG and HAMBEROER, 1964). Exceptions from this general morphology exist, however, both among invertebrates and vertebrates; in some instances the axons of adrenergic cell bodies are entirely varicosed, as has been observed in some adren- ergic sensory systems.
2. The intraneuronal distribution of the monoamines is identical (see above), the terminal varicose parts always show a high fluorescence intensity.
3. As in vertebrates a depletion of the neuronal monoamines can be produced by means of t reatment with reserpine.
4. In the molluscan ganglia, varicose adrenergic fibres superimpose on non- adrenergic cell bodies in an apparently synaptic arrangement of the same appear- ance as found in the mammalian brain (CARLSSON et al., 1962) and autonomic ganglia (for ref. see F~LCK and OWMA~, 1965b).
Thus there are reasons/or believing that among the /our monoamines which act as neurohumoral agents, 5 - H T and D A are those active in molluscs.

Dopamine and 5-Hydroxytryptamine in some Mollusca 497
Refe rences
ANDI~N, N.-E., A. CARLSSON, A. DAHLSTROM, K. FUXE, N.-A. HILLARP, and K. L~RSSON: Demonstration and mapping out of nigro-neostriatal dopamine neurons. Life Sci. 3, 523--530 (1964).
BERTL~R, A., A. CARLSSON, E. ROS~NGR~N, and B. WALDECK: A method for the fluorimetrie determination of adrenaline, noradrenaline, and dopamine in tissues. Kgl. Fysiogr. S~illsk. Lund, FSrh. 28, Nr. 12, 121--123 (1958).
- - , B. FALCK, C. G. GOTTFRIES, L. LJUNGGREN, and E. ROSENGREN: Some observations on adrenergic connections between the cerebral hemispheres and the mesencephalon. Acta pharmacol. (Kbh.) 21, 283--289 (1964).
- - , and E. ROS~.NORE~: Occurrence and distribution of catecholamines in brain. Acta physiol. scand. 47, 350--361 (1959).
CARDOT, J. : Sur la pr6sence de dopamine dans le syst~me nerveux et ses relations avec la d6earboxylation de la dioxyph6nylalanine chez le Mollusque Helix pomatia. C. R. Acad. Sci. (Paris) 257, 1364--1366 (1963).
CARLSSON, A., B. FALCK, K. FUXE, and N.-A. HILLARP: Cellular localization of monoamines in the spinal cord. Acta physiol scand. 60, 112--119 (1964).
- - - - and N.-A. HILLARP: Cellular localization of brain monoamines. Acta physiol, scand. 56, Suppl. 196, 1--28 (1962).
CORRODI, H., u. N.-A. HILLARP: Fluoreszenzmethoden zur histochemisehen Sichtbarmachung von Monoaminen. 1. Identifizierung der fluoreszierenden Produkte aus Modellversuchen mit 6,7-Dimethoxyisochinolinderivaten und Formaldehyd. Helv. chim. Acta 46, 2425 bis 2430 (1963).
- - - Fluoreszenzmethoden zur histochemischen Sichtbarmachung von Monoaminen. 2. Identifizierung des fluoreszierenden Produktes aus Dopamin und Formaldehyd. Helv. chim. Aeta 47, 911--918 (1964).
DAHL, E.: Cellular localization of monoamines in mollusc nervous systems. Proceedings of the XVI Internat. Congr. of Zoology, Washington, D. C. August 20--27, 1963, 2, 105 (1963).
- - B. FALCK, ]~r LINDQVIST, and C. v. MECKLENBURG: Monoamines in mollusc neurons. Kgl. Fysiogr. S~llsk. Lund, FSrh. 32, Nr. 8, 89--92 {1962).
- - - - C. V. ~r and H. MYHRBERG: Adrenergic sensory neurons in invertebrates. Gen. comp. Endoer. 3, 693 (1963a).
An adrenergic nervous system in sea anemones. Quart. J. micr. Sei 104, 531--534 (1963b).
DAHLSTROM, A., and K. FUXE: Evidence for the existence of monoamine-containing neurons in the central nervous system. I. Demonstration of monoamines in the cell bodies of brain stem neurons. Acta physiol, scand. 62, Suppl. 232, 1--55 (1964).
EULER, U. S. v.: Noradrenaline. Springfield (Ill.): Ch. C. Thomas 1956. - - Neurotransmission in the adrenergic nervous system. Harvey Lect. 55, 43--65 (1961).
FALCK, B.: Observations on the possibilities of the cellular localization of monoamines by a fluorescence method. Acta physiol, scand. 56, Suppl. 197, 1--25 (1962a).
- - Histochemical localization of monoamines in tissues. In: Reports from the third Scand. Conf. on Cell Research (Ed. K. J. PEDERSEN), Copenhagen 1962b.
- - J. H)iOCENDAL, and CH. OWMA~: The localization of adrenaline in adrenergic nerves in the frog. Quart. J. exp. Physiol. 48, 253--257 (1963).
- - N.-A. HILLARP, G. TtIIEME, and A. TORt: Fluorescence of catechol amines and related compounds condensed with formaldehyde. J. Histochem. Cytochem. 10, 348--354 (1962).
- - , and CH. OWMAN: A detailed methodological description of the fluorescence method for the cellular demonstration of biogenic monoamines. Acta Univ. Lund., Sect. II , No 7, 1--23 (1965a).
- - - - Histochemistry of monoaminergic mechanisms in peripheral neurons. Symposium on mechanisms of release of biogenic amines, Stockholm 1965. Oxford: Pergamon Press (in press) (1965b).
- - , and A. TORP: New evidence for the localization of noradrenaline in the adrenergic nerve terminals. IVied. exp. (Basel) 6, 169--172 (1962).

498 E. DAHr, et al. : Dopamine and 5-tIydroxytryptamine in some Mollusea
FLOREu E., u. E. FLOttEY: ~ber die m6gliche Bedeutung yon Enteramin (5-Oxy-Tryptamin) als nerv6se Aktionssubstanz bei Cephalopoden und dekapoden Crustaeeen. Z. Naturforseh. 9b, 58--68 (1954).
HANSTR6•, B.: Vergleiehende Anatomie des Nervensystems der wirbellosen Tiere. Berlin: Springer 1928.
NORBERO, K.-A., and B. HAMBEROER: The sympathetic adrenergic neuron. Some characteris- tics revealed by histoehemical studies on the intraneuronal distribution of the transmitter. Acta physiol, scand. 6~1, Suppl. 238, 1--42 (1964).
SWEENEH, D.: Dopamine: Its occurrence in molluscan ganglia. Science 189, 1051 (1963). WELSH, J. H.: Hydroxytryptamine: a neurohormone in the invertebrates. Fed. Proc. 18,
162--163 (1954). - - , and M. MOORttEAD: The quantitative distribution of 5-hydroxytryptamine in the inverte-
brates, especially in their nervous systems. J. Neurochem. 6, 146--169 (1960).
Prof. Dr. EttIK DAHL Department of Zoology, University of Lund Lund, Sweden