Embed Size (px)
Citation preview

J. Exp. Biol. (1966), 45, 343-356 2 4 3With 1 plate and 9 text-figures
Printed in Great Britain
NEUROMUSCULAR MECHANISMSOF BURROW IRRIGATION IN THE ECHIUROID WORM
URECHIS CAUPO FISHER & MACGINITIE
I. ANATOMY OF THE NEUROMUSCULAR SYSTEM ANDACTIVITY OF INTACT ANIMALS
BY JAMES V. LAWRY, JR.*
Department of Biological Sciences, Stanford University,Stanford, California
(Received 8 June 1966)
INTRODUCTION
Animals exposed to the direct influences of the sun, moon and seasons may exhibitrhythmic behaviour displaying a solar, lunar or annual periodicity, whereas organismsliving where periodic stimuli are reduced or absent often rely upon endogenousrhythms for initiating behaviour. Among the latter are some of the inhabitants ofmudflats and estuaries which may occupy permanent burrows or tubes that provideanchorage in the shifting substrate. Such a mode of life poses problems of foodacquisition and waste disposal, and tube dwellers may rely on neuromuscular activitygoverned by intrinsic rhythms to produce a current of water which irrigates the burrow.
Urechis caupo, described by Fisher & MacGinitie (1928a), inhabits a 'U'-shapedburrow in California sloughs and estuaries. The vertical arms of a burrow extenddownward about 40 cm and are united by a horizontal portion of about 90 cm.Throughout its adult life an individual remains in this burrow, enlarging it to keeppace with growth. Water is circulated by peristaltic contractions of the body-wallinitiated behind the proboscis, which force water backward between the worm and thewall of the burrow. The flow of water brings in food and oxygen and expels wastesand gametes (Text-fig. 1). Fisher & MacGinitie report that, when feeding, Urechismoves toward one entrance of the burrow, and a ring of glands at the base of theproboscis secretes a band of mucus which adheres to the wall. The worm then movesslowly downward, forming a funnel of mucus attached to the rim of the burrow at itsupper end and to the ring of glands at its lower. The water that is subsequentlypumped through the burrow must pass through the mucus funnel, which strainsparticles from the current. After an interval of several minutes to an hour of pumping,the worm detaches the mucus tube from its body and consumes the tube and its con-tained particles, feeding while it moves upward to the mouth of the burrow where thecycle is begun again.
It appears that oxygen exchange occurs through the walls of the spacious thin-walled hind gut. Urechis has an open circulatory system and lacks a heart and bloodvessels. The coelomic fluid, containing cellular haemoglobin, is circulated by peri-
• Present address: Gatty Marine Laboratory, St Andrews, Scotland.

344 JAMES V. LAWRY
stalsis of the hind gut and body-wall. Peristaltic waves originating from an anal pace-maker draw water into the hind gut through the anus by a series of one to twenty-fivesmall inhalations. These movements are terminated by a contraction expelling waterfrom the hind gut. Fisher & MacGinitie (19286) recorded periods of inhalation lastingfrom 25 to 90 sec. and exhalations of 10-50 sec. Redfield & Florkin (1931) collectedsamples of hind-gut water at the moment of expiration and found the oxygen contentof the expired water to be two-thirds that of the sea water in the burrow.
Text-fig. 1. UreMs caupo in its burrow. MN, mucus net; P, proboscis; PW, peristalticwave; A, anus.
Two mechanisms might give rise to spontaneous cycles of patterned behaviour: theactivity could be triggered or paced by environmental stimuli acting as cues, or theorganism could instead rely upon intrinsic pacemakers to initiate behaviouralsequences. In Aremcola feeding movements and longer cycles are regulated by spon-taneously active elements in the oesophagus and nerve cord. The rhythmic burstsbegin without apparent stimulus or biological need, and they subside without apparentexternal influence; their rhythm plays a vital role in patterning the life of the animal(Wells, 1950). Urechis appears to show a similar dependence upon autogenic behavioursequences.
The neuromuscular system of Urechis produces a current of water which irrigatesthe burrow, thus providing an opportunity for quantitative study of the relationshipbetween muscular activity and the mechanism of exchange with the environment. Thispaper reports the findings resulting from a study of the neuromuscular system.Anatomical description provided a structural basis for subsequent behavioural andphysiological investigations. Two pacemaker systems apparently initiate the pumpingand respiratory movements of this echiuroid. The activity of these pacemakers wasstudied, and the frequency of peristalsis and the volume of water pumped per daywere determined for worms kept under constant conditions in the laboratory.

Neuromuscular mechanisms of burrow irrigation. I 345
METHODS
Urechis caupo was collected from its type locality, Elkhorn Slough, California, andwas maintained in sea-water aquaria in the laboratory. Some worms were kept in glassor Lucite tubes (technique of Fisher & MacGinitie, 19286), and others were allowedto burrow in sand. Animals prepared for histological study were fixed in Bouin's orGilson's fixatives, and were mounted in paraffin after a double-embedding procedure,in which they remained for 24 hr. in a 1 % solution of celloidin in methyl benzoate.Sections were cut at 10 /i and stained using the silver method of Rowell (1963) andMasson's trichrome. The nervous systems of living preparations were stained withmethylene blue in sea water.
Text-fig. 2. Diagram of the apparatus for measuring initiation times and conduction rates ofperistaltic waves in intact Urechis. Numbers indicate channels of strip-chart recorder (SCR)sequentially activated as the expanding body-wall in the wake of a ring of propagating circularmuscle contraction pushes pistons (P) flush with the wall of the burrow. The pool of mercury(Hg) on the top of each piston (P) closes the circuit each time it contacts the two electrodes.A voltage source (EMF) is supplied by a 6 V battery in series with the recorder. A section ofchart is shown moving in the direction of the arrow. Channels 4 and 5 are momentarilyactivated by the expansion beneath the corresponding pistons. The vertical bars on the chartindicate the times when circuits were closed. {W) is Urechis with the proboscis to the leftWaves of contraction propagate to the right.
Observations were made of the behaviour of Urechis, either kept free in pans of coldsea water or confined in artificial burrows which were blocked to prevent anterior orposterior movements of the worms. Water currents were made visible by the additionof methylene blue or suspensions of Urechis eggs. Analyses of the actions of specificmuscle layers were made by viewing worms placed on flat surfaces in air, followed bypartial dissections of the several muscle layers to determine whether their contractionswere involved in specific actions. Irrigation behaviour was analysed cinematographic-ally, and measurements of the rate of peristaltis were made from prints using the film

346 JAMES V. LAWRY
transport as a time base and from images projected on a grid. An apparatus was de-signed for determining the frequency of initiation and rates of conduction of peri-staltic waves behind the proboscis of Urecfns over a long period (Text-fig. 2). Theseexperiments were conducted at 130 C , and the water in the tube was aerated con-tinuously.
An apparatus for recording periods of activity and the volume of water pumped byUreckis is diagrammed in Text-fig. 3. A single animal was maintained in a Luciteburrow at 13° C. for several weeks. Aerated sea water entered both ends of the tubethrough syringes, opened by solenoids and closed by springs. The solenoid circuitswere made and broken by the movements of glass floats activating microswitches.Each time the worm initiated a pulse which moved water from one end of the apparatus
Text-fig. 3. Apparatus for recording periods of activity and the volume of water pumped byUrecttu. For explanation of apparatus, see text. SCR, Strip-chart recorder; arrow indicatesthe direction of chart movement; EMF, 6 V battery supplying current to activate solenoidsand channels of SCR; M, microswitch; F, float; SO, solenoid; SP, spring; S, syringe; I, inlet;O, overflow; U, union through which Ureckit was introduced into the apparatus. The arrowabove Urecfris, W, shows direction of water moved by the worm. The microswitch on the rightside is closed and the solenoid is activated. The microswitch on the left is open and the springhas closed the syringe preventing water entry through this inlet.
to the other, the float on the end from which the pulse initiated dropped, thus closingthe solenoid at that end. Upon activation of the solenoid, a volume of water equal tothat moved by the worm entered the apparatus through the inlet and restored the floatto its original position. When the circuit was broken by the rising float, the springclosed the syringe, stopping the inflow of water. On the other end of the apparatus thevolume of water pumped by the worm was recorded as overflow. Since the worm wasable to turn around in the burrow, the solenoid and inlet were duplicated on theopposite end of the apparatus. The two overflows were measured separately, and each

Neuromuscular mechanisms of burrow irrigation. I 347
solenoid activated a separate channel of an Esterline Angus strip-chart recorder—a penstroke registering each pumping event on the channel corresponding to the end of theapparatus from which the pulse originated.
RESULTS
(1) Anatomy of the neuromuscidar system
Average specimens of UrecMs caupo are 15-18 cm. long. The body of the worm iscigar-shaped, and the anterior end narrows, tapering into a short prostomial proboscis.The anus is terminal and is surrounded by an incomplete ring of ten or eleven analsetae. A single pair of setae is located on the ventral surface of the proboscis, justposterior to the mouth. Seitz (1907) describes the anatomy of U. cMlensis, and Fisher& MacGinitie (1928 a) use Seitz's description as a basis for their description of
Text-fig. 4. Stereogram of the neuromuscular system of Urechis in the anal region. Distensiblehind gut is in expanded state. NC, Nerve cord; GC, gut connective; HG, hind gut; AS, analsac; ICM, inner circular muscle layer; LM, longitudinal muscle layer; OCM, outer circularmuscle layer; / , integument; PN, peripheral nerve.
U. caupo which has been extended to the microscopic level in the present investiga-tion. The body-wall (Text-fig. 4 and PL I A) consists of an outer rugose, papillateintegument, an underlying areolar dermis, an outer layer of muscle fibres 1-3 fi indiameter arranged circularly, an inner layer of longitudinal muscle fibres 5 fi indiameter, an inner layer of circular muscle fibres 3 and 5 fi in diameter, and a peri-toneum. The nervous system is composed of a central nervous system consisting ofa ventral, solid nerve cord bifurcating rostrally to form two circumpharyngeal con-nectives that unite dorsally in the tissue of the proboscis, and a peripheral nervoussystem of several hundred pairs of segmentally arranged peripheral nerves which

348 JAMES V. LAWRY
innervate the body wall, the setiferous apparatus and the coelomic viscera. Thecircumpharyngeal connectives run around the edge of the hood of the proboscis,forming a ring from which roots leave at intervals to innervate the proboscis (Text-fig. 5 and PI. IB). The ventral nerve cord hangs freely in the coelom and is connected
NC
wText-fig. 5. Diagram of the circumpharyngeal connectives in the proboscis of Ureckis. Thebody-wall was cut along the mid-ventral line anteriorly to the bifurcation of the connectives;the cut was extended along both sides of the buccal funnel, and the two flaps thus formed wereremoved. The body-wall has been retracted laterally in the region of the bifurcation. Peripheralnerves and the anterior setiferous apparatus have been deleted. The dotted line divides theanterior segmentation cavity of the proboscis from the coelom and trunk. GC, gut connective;CC, circumpharyngeal connective; SC, segmentation cavity; CO, coelom; M, mouth; NC,nerve cord.
to the body-wall solely by the peripheral nerves, as the ventral mesentery of otherechiuroids is absent in Ureckis (Text-fig. 6). The cord extends from the anteriorbifurcation of the circumpharyngeal connectives to the posterior bifurcation of thecord where it surrounds the hind gut and innervates both it and the body wall of theanal region. (PI. IC) The cord is invested with a multilayered sheath and sometimesappears convoluted, sometimes straight (Text-fig. 6). When the longitudinal musclesof the body-wall contract, the cord is thrown into a series of tight bends which dis-appear on relaxation. The sheath is covered with a layer of peritoneum externally,below which are found spindle-shaped cells resembling the smooth muscle cells of

Neuromuscular mechanisms of burrow irrigation. I 349
vertebrates (PL ID). Whether these cells function in movements of the cord is un-known; the cord does not appear to contract when stimulated electrically. Inter-digitations of connective tissue from the sheath divide up the rind of neuron somatabelow it into indistinct masses of neurons (PL IE). These send fibres centrally intothe neuropile, and in intersegmental areas fascicles of fibres from ipsi- and contra-lateral somata form peripheral nerve trunks.
Text-fig. 6. External features of the nerve cord of UreMs. PNBW, Peripheral nerve enteringbody-wall; PN, peripheral nerve; PNR, peripheral nerve root; S, sheath. The distance froma to b is 7 mm.
A multicellular 'giant fibre' 18-20 fi in diameter is located medio-dorsally justbelow the rind of somata. It is invested with a multilayered sheath and conductsimpulses at 1-5 m/sec. (Lawry, 1966). The cells composing this compound structureappear linked by characteristic tight junctions and send processes into the circum-pharyngeal connectives (Lawry, unpublished).
Several hundred peripheral nerves emerge from the cord of Urechis on the rightand left sides. These are sometimes opposite, sometimes alternate and sometimesirregular in position. Frequently a peripheral nerve emerges as a number of rootletswhich then fuse; or it may in other cases emerge as a single trunk (Text-fig. 6). Theperipheral nerves run in the coelom before entering the body-wall where they ramifybetween muscle layers, usually innervating the layers on either side of them (Text-fig. 7). Nerve fibres destined for the dermis and integument emerge from peripheralnerves running in the longitudinal muscle layer and penetrate the outer circular layerof muscle before entering these structures. Peripheral nerves run around the circum-ference of the worm in annular fashion, but since they leave the cord alternately they

350 JAMES V. LA WRY
do not meet in the dorsal mid-line. The wall of the gut is innervated along its lengthby nerve fibres that enter connectives from nerve trunks in the body-wall. A connectivewith its contained nerve fibres is shown (PI. i C).
(2) Activity of intact animals
The sequence of events during the conduction of a peristaltic wave along a worm isas follows:
(a) Cinematographic analysis shows that before a wave begins in the proboscis theprostomium is expanded and extended by 5-10 mm. through contraction of thecircular muscle layers. If the forward thrust of the proboscis is prevented by blocksplaced in the tube to keep the worm at resting length, no subsequent propagated waves
GC
Text-fig. 7. The branching patterns of peripheral nerves in the body wall of Urechis. A. Diagramof peripheral nerve running at junction of the outer circular muscle layer and the longitudinalmuscle layer, sending branches through the outer circular muscle layer to the integument. B.Diagram of peripheral nerve running at junction between inner circular muscle layer andlongitudinal muscle layer, sending branches to other layers and fibres to the gut connectives.C. Diagram of peripheral nerve with branches running at junction of inner circular musclelayer and longitudinal muscle layer and outer circular muscle layer. NC, Nerve cord; ICM,inner layer of circular muscle; LM, longitudinal muscle layer; OCM, outer circular layer ofmuscle; /, integument; PN, peripheral nerve; GC, gut connective.

Neuromuscular mechanisms of burrow irrigation. I 351
of peristalsis occur. Cuts in the circular muscle layers perpendicular to the long axisof muscle fibres gape during proboscis extension, indicating contraction of the musclefibres in these layers. Cuts in the longitudinal muscle layer perpendicular to the direc-tion of these fibres either close or remain unchanged, indicating that they are notinvolved in proboscis extension.
(b) The pulse appears as a sequential contraction of the circular muscle layers pro-pagating down the length of the worm. When a worm with a hind gut emptied ofwater is placed on a flat surface in air, the contractions of the outer circular muscle layerare seen as transverse furrows in the integument. In a dissected preparation the pulseappears as a contraction of the inner circular muscle layer. Cuts placed perpendicularto the long axes of muscle fibres in the circular layers gape as the pulse passes.
(c) The hydrostatic pressure of the coelomic fluid and contraction of the longtiu-dinal muscle layer in the wake of the propagating contraction help to restore thecontour of the worm. Animals from which the coelomic fluid has been drained remainlimp, and following the passage of a wave of circular muscle contraction longitudinalfurrows appear in the integument. That these are the result of contraction of long-tiudinal muscles is indicated by the fact that cuts placed perpendicular to the fibres ofthe longitudinal layer gape.
(d) The velocity of propagation of a peristaltic wave from the proboscis decreasesin the anal region, and in normal specimens the wave is usually obliterated severalcentimetres anterior to the anus by peristalsis propagating rostrally from a posteriorpacemaker.
(e) Some water is expelled through the anus every time a wave from the proboscisenters the anal region.
(/) While a peristaltic wave from the proboscis propagates to the anus, two or threepulsations of the posterior end of the worm draw water into the hind gut through theanus, causing a swelling of the hind gut and posterior end of the worm. Inhalantpulsations proceed rostrally from the anal regions for several centimetres as waves ofperistalsis in the circular muscle layers of the body wall before they are obliterated bya pulse from the proboscis. The hind-gut swelling is reduced periodically by a forcefulexhalant contraction of the musculature of the hind gut and body wall in the analregion. This exhalation may or may not be coincident with the arrival of a wave fromthe proboscis at the anus. If the proboscis pacemaker is inactive, peristaltic wavesoriginating from the anus may propagate rostrally to the anterior end of the worm.
(3) Measurements of rates of conduction of peristaltic waves
Measurements of initiation times and rates of conduction of a series of peristalticwaves initiated behind the proboscis of Urechis are presented in Table 1. The datafrom the strip-chart recorder were reduced by measuring and tabulating chart dis-tances which were converted into times. In some instances it was impossible to recordeach pulse at every piston; consequently, n (the number of measurements) differs foreach piston and interval. The means of these measurements are presented graphicallyin Text-fig. 8. The broken sections in the horizontal line for each piston represent theduration of the annulus of contraction of circular muscle at the piston. The shortsegments, of which Dx is an example, are measurements of the time for an annulusto travel the 20 mm. between pistons. The larger segments, Dit represent the duration,

352 JAMES V. LAWRY
Table i . Averaged measurements of the distances indicated in Text-fig. 8for each of two experiments
Distances(mm.)
I*o
2 0
4 06 080
1 0 0
1 2 0
140
lit 02 0
406080
D, (sec.)
1 2 5
i7-517-51 6 8
17-51 7 11 7 11 2 8
1 2 58-8
11-2
7-27 2
S.E. Y ̂ S.D./Wjfor D,
2 - 0
3 63-63-54'42 - 26 l2-4
I'O
1-21-5i -o
4'4
n = no. ofobservations
for/),
3626
2525172 0
8z
62626258
3
/(sec.)
3-33 14-63-6S'35'35-4
3-24 06 4
I I - 2
s.B. V(s.D./n)for/
o-83-81-2I T
1-42 30
°-sOSo-86-4
n — no. ofobservations
for/
26
25251619
81
626258
3
= S3 sec; S.E. = 0-5, n = 16.•\IWI = 127 sec., S.E. = 2-6, n = 101.
Seconds10 20 30 40
20
Text-fig. 8. Chart of averaged intervals calculated for components of the peristaltic waves ofUreckis recorded with the apparatus diagrammed in Text-fig. 2. Abscissa: time in seconds;ordinate: distances travelled in millimetres by peristaltic waves along body of worm in each oftwo experiments, I and II. Longitudinal bars represent durations of the components of thepropagated wave at each piston. For further explanation, see text.
at a piston, of a wave of expansion of the body-wall in the wake of an annulus. Theinterval, /, is a measurement of the time for a wave of expansion to traverse the distancebetween pistons. IWIis the time between the initiations of pulses behind the proboscis.
A conducted wave of contraction takes about 2$ sec. to progress from proboscis toanus in a specimen 140 mm. long. Slowing of pulses from the proboscis posteriorlymay result from interaction of the posterior pacemaker with the conducting elements.

Neitromuscular mechanisms of burrow irrigation. I 353
Large broken sections appear in the horizontal bars representing the activity of thefirst and last pistons (o and 140 in Text-fig. 8). These spaces are in part explained bythe following:
(a) The anterior and posterior ends of a worm taper. Pulses in these regions arenot as large as those in the central regions of the body and, thus, do not activate thepistons for as long a period.
(b) The space under piston eight is in part due to the marked slowing of a posteriormoving pulse in the anal region. At this piston pulses were often cancelled prematurelyby breathing movements of the anus.
Piston five in Expt. II was activated by anal pulsations and was rarely reached bydescending pulses from the proboscis, which were obliterated in the interval betweenpistons four and five by pulses propagating anteriorly from the anus. The anal pulseswere also extinguished in the process.
Days
Text-fig. 9. Histogram of the number of pumping events per hour over an 8-day period and per10 min. interval for a 24 hr. period day 5 (inset). Shaded bars indicate events that occurred afterthe worm reversed in the burrow. For further explanation, see text.
(4) Periods of activity and volume of water pumped per day
Experiments were undertaken to ascertain whether the behaviour of Urechis inartificial burrows under constant temperature conditions displayed any recurrentpatterns, since various sorts of endogenous rhythms in phase with tidal cycles maypersist unaltered for a time under constant conditions, whereas behaviour triggereddirectly by environmental stimuli ceases or changes upon removal of the stimulus.Observations while digging Urechis indicate that when the burrow is uncovered at lowtide the worm ceases pumping and remains expanded in the horizontal portion of theburrow.
Some representative data from the apparatus diagrammed in Text-fig. 3 are graphedin Text-fig. 9. They were obtained from a large female Urechis taken from the mudflatand maintained for 192 hr. in semi-darkness at 130 C. from 9 to 17 August 1964. Eachhour of strip-chart record was divided into 10 min. intervals and the number ofpumping events in each was plotted; twelve events per channel per interval was the

354 JAMES V. LAWRY
maximum number discernible. Since in some intervals fifteen or twenty events weredistributed between the two channels, more than twelve strokes could occur on onechannel in an interval and not be recorded.
A sample histogram of daily activity appears in the inset of Text-fig. 9. The wormwas active continuously throughout the 24 hr. period, and it pumped at least twicein every 10 min. interval. This rinding contradicts that of Hall (1931) who reportedresting periods from 20 min. to 1 hr. in length. In only 1 hr. of the 8-day period wasno pumping recorded, and at all other times there were at least nine events per hour.Periods of most rapid activity were obscured by lack of resolution in the apparatus.In the 8-day period many bars contain saturated values which could be a major sourceof error in interpreting this data were it not for the agreement between the number oflitres of water moved and the number of pumping events per day. Table 2 correlatesthe number of litres circulated and the number of pumping events recorded on thestrip-chart recorder. The arithmetic mean of ten measurements of the volume of watermoved per wave of contraction was 20 ml. The variability among the averages inHall's experiments was considerable; he collected water at 5 min. intervals and did notrecord the number of pumping movements accomplished by his animals during theperiod of observation.
Table 2. Correlation between the number of litres of water circulated and the number ofpumping events per day for the 8 days in Text-fig. 9
(For explanation of table, see text.)
f n o .
1
2
34S678
Litrespumped
28-1426-0823-3839'OS28603°-3726-7728-33
Pumpingevents
9991244128515191328136913141186
Litres/event
0-0280-0210-0180-026O-O2I0-022O-O2O0024
No. of saturatedintervals (12 events
in 10 min.)
38374 1
5536474 2
34
In the present apparatus worms displayed a marked preference for one particularanterior-posterior direction for pumping water. The worms reversed their directiononce every few hours, but the volume subsequently pumped was small compared withthe volume of water moved in the preferred direction. Shaded bars indicate the volumeof water pumped after such reversals by the worm in the experiment described. Othersbehaved similarly. The causes and significance of such turning behaviour are unknown.
Fifty millilitres of a concentrated suspension of cultured Nitzschia were added tothe inlet reservoir on several occasions to see if the increased food content of the wateraffected the pumping rate. One might assume the rate of production of the mucus netswould be related to the particle content of the water, and Hall noted an increase inpumping rate during periods of feeding. One of his worms averaged 36-5 c.c./min.while feeding and 7-2 c.c./min. when not feeding. Hall gives the rate of production ofmucus nets as 0-7 per hour for one worm and 1-5 per hour for a second. The worms inthe present experiment fed continuously, as is evidenced by the large volumes of

Neuromuscular mechanisms of burrow irrigation. I 355
water pumped and the fact that mucus nets were observed at all hours. The additionof diatoms to the inlet reservoirs had no effect upon the pumping rate.
DISCUSSION
Anatomy of the neuromuscular system
The present account is in keeping with most of the early work on the anatomy ofUrechis. Spengel (1880) thought that the 'neural canal' of EchiuruspaUacii might bea giant axon, and this appears to be the case in U. caupo. The peripheral nerves, how-ever, do not fuse mid-dorsally (Grasse, fig. 717, p. 901) but ramify within the body-wall, as they approach the dorsal mid-line.
Activity of intact animals
Muscular contraction against the fluid within the coelomic and hind gut cavitiesprovides Urechis with a flexible hydrostatic skeleton. The animal can control internalhydrostatic pressure and body form by adjusting the volume of water in the hind gut.The internal hydrostatic pressure of Urechis is increased by (a) contractions of body-wall musculature during the propagation of a pulse from proboscis to anus, and (b)inhalant breathing movements of the anus which fill the hind gut. The pressurecreated by (a) is released when the anus opens upon arrival of a pulse from the pro-boscis. This pressure may be very small compared with pressure created by (b) whichcauses noticeable swelling of the posterior portions of animals observed outside then-tubes. The pressure created by (b) is relieved by the forceful exhalant movements ofthe hind gut. The jet of water could also serve to clear the posterior part of the burrowof organic wastes and particulate debris.
The extension of the proboscis before the commencement of a pulse suggests thatthis movement could serve to initiate the pulse reflexly, since pulses fail to occur whenelongation is inhibited.
Cinematographic studies indicate that peristaltic waves decelerate upon leaving theproboscis and reaching the trunk. The reasons for this slowing are not known, but itmay be related to the increased volume of musculature that must be activated whena pulse enters the trunk.
Measurements of rates of conduction of peristaltic reaves
In the chart of averaged intervals for components of peristaltic waves (Text-fig. 8)the time needed for a pulse to propagate from the first piston to succeeding pistonsshould be some multiple of the time for a pulse to propagate between the first andsecond piston, if the pulse width (the length of body-wall activated) and the velocityof propagation are constant. However, the time required for a wave to progress frompiston one to successive pistons tends to increase as a wave moves caudally. The com-ponents producing this delay must be ascertained and may be indicative of the under-lying mechanism.
Periods of activity and volume of water pumped per day
The behaviour of the worms in this apparatus did not appear cyclic and could not becorrelated with tidal or diurnal rhythms. Populations of Urechis in the field might

356 JAMES V. LAWRY
display cycles of behaviour which could give way to aperiodic patterns if the environ-mental phasing stimulus were absent in the laboratory.
SUMMARY
1. The anatomy of the muscular and nervous systems of Ureckis is described.2. An apparatus is described whereby it is possible to record the parameters of the
peristaltic waves used in irrigating the tube.3. Urechis is continuously active and shows no periodicity under conditions of
constant temperature. About 1280 peristaltic waves are initiated, and about 29 L ofwater are moved, in 24 hr.
The work described in this paper constitutes part of a dissertation submitted to theDepartment of Biological Sciences at Stanford University in partial fulfilment of therequirements for the degree of Doctor of Philosophy and was supported in part bya Public Health Service Fellowship I-F1-GM-20, 539-01A2 from the NationalInstitute of General Medical Sciences. The author is indebted to Prof. DonaldKennedy for his advice, support and encouragement during the course of theseexperiments.
REFERENCES
FISHER, W. K. & MAcGrNrnB, E. (1928a). A new echiuroid worm from California. Aim. Mag. Nat. Hist.(Ser. 10), 1, 199-204.
FISHER, W. K. & MAcGiNrnB, E. (19286). The natural history of an echiuroid worm. Ann. Mag. Nat.Hist. (Ser. io), I, 204-14.
HALL, V. E. (1931). The muscular activity and oxygen consumption of Urechis caupo. Biol. Bull. Mar.Biol. Lab., Woods Hole 61, 400-416.
LAWRY, J. V. (1966). Neuromuscular mechanisms of burrow irrigation in the echiuroid worm Urechiscaupo Fisher & MacGinitie. II Neuromuscular activity of dissected preparations. .7. Exp. Biol. 357-68,
REDFTKLD, A, C. & FLORKIN, M. (193 I ) . The respiratory function of the blood of Urechis caupo. Biol.Bull. Mar. BioL Lab., Woods Holt 61, 185-210.
ROWELL, C. H. FRASHR (1963). A general method for silvering invertebrate central nervous systems.Q. Jl Microsc. Sri. 104, 81-97.
SETTZ, P. (1907). Der Bau von Eckutrus ckilensis (Ureckis n.g. ckilensis). Zool. Jb. (Abt. Anat,), 24,323-56.
SPBNGEL, J. W. (1880). Beitrfige zum Zenntnis der Gephyreen II. Die Organisation des Echiuruspallasii. Z. wist Zool. 34. 460-538.
WELLS, G. P. (1950). Spontaneous activity cycles in polychaete worms. Symp. Soc. Exp. Biol. 4, 127-42.
EXPLANATION OF PLATE
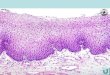
A. Transverse section of the body-wall of a small Urechis. (Silver method of Rowell.) CU, Cuticle;E, epithelium; D, dermis, OCM, outer circular muscle; LM, longitudinal muscle, ICM, inner circularmuscle; PN, peripheral nerve; CO, coelom.B. Transverse section of a portion of the proboscis of a small Urechis, (Silver method of Rowell.) PN,Peripheral nerve; SC, segmentation cavity; CC, circumpharyngeal connective; BCE, buccal ciliatedepithelium.C. Transverse section of the nerve cord in the region of the hind gut. (Silver method of Rowell.) DCB,Dorsal cell bodies in nerve cord; PN, transverse section of peripheral nerve; GC, gut connective withcontained nerve fibres; ICM, inner layer of circular muscle; HE, epithelium and hind-gut wall;C. coelom.D. Parasagittal section of Urechis nerve cord. (Silver method of Rowell.) S, Spindle-shaped cells of theconnective tissue sheath intermingling with cell-bodies which send processes centrally into neuropile,NP. Section cut perpendicularly in region 1 of E.E. Transverse intersegmental section of the nerve cord of Ureckis. (Silver method of Rowell.) S, Sheathwith spindle-shaped cells; DCB, dorsal rind of cell-bodies; PN, peripheral nerve trunk; GF, giant fibre.

Journal of Experimental Biology, Vol. 45, No. 2 Plate 1
• 1 mm.
8C£
-0 8 mm.
JAMES V. LAWRY {Facing p. 356)