Embed Size (px)
Citation preview

lable at ScienceDirect
Neurobiology of Aging 36 (2015) 3314e3320
Contents lists avai
Neurobiology of Aging
journal homepage: www.elsevier .com/locate/neuaging
Pitx3 deficiency produces decreased dopamine signaling andinduces motor deficits in Pitx3(�/�) mice
Weidong Le a, Lifen Zhang b, Wenjie Xie c, Song Li a, John A. Dani d,*aCenter for Translational Research on Neurological Diseases, the 1st Affiliated Hospital, Dalian Medical University, Dalian, Liaoning, ChinabDepartment of Neuroscience, Baylor College of Medicine, Houston, TX, USAcDepartment of Neurology, Baylor College of Medicine, Houston, TX, USAdDepartment of Neuroscience, Mahoney Institute for Neurosciences, Perelman School of Medicine, University of Pennsylvania, Philadelphia, PA, USA
a r t i c l e i n f o
Article history:Received 4 February 2015Received in revised form 10 August 2015Accepted 11 August 2015Available online 20 August 2015
Keywords:Transcription factorSubstantia nigraAnimal modelParkinson’s diseaseStriatum
* Corresponding author at: Department of NeuroscNeurosciences, Perelman School of Medicine, Universphia, PA 19104, USA. Tel.: 215 898 8498; fax: 215-573
E-mail address: [email protected] (J.A. Dani).
0197-4580/� 2015 The Authors. Published by Elsevierhttp://dx.doi.org/10.1016/j.neurobiolaging.2015.08.012
a b s t r a c t
Midbrain dopamine (DA) neurons are involved in cognition, control of motor activity, and emotion-related behaviors. Degeneration of DA neurons particularly in the substantia nigra is a hallmarkof Parkinson’s disease. The homeobox transcription factor, Pitx3, plays a critical role in thedevelopment, function, and maintenance of midbrain DA neurons. We found that in young adultPitx3-null mice, Pitx3(�/�), there was decreased tyrosine hydroxylase staining, indicating a loss of DAneurons particularly in the substantia nigra. In addition, fast-scan cyclic voltammetry and microdialysisassays of DA release indicated that the lack of Pitx3 caused a significant reduction of striatal DA release.Tonic DA release was impaired more significantly than the phasic DA release induced by burst firing ofDA neurons. Furthermore, behavioral tests revealed that Pitx3(�/�) mice displayed abnormal motoractivities, including impaired motor coordination and decreased locomotion. In summary, these dataprovide further evidence that Pitx3 is specifically required for DA-related function and, if impaired, Pitx3could contribute during the pathogenesis of Parkinson’s disease.� 2015 The Authors. Published by Elsevier Inc. This is an open access article under the CC BY-NC-ND
license (http://creativecommons.org/licenses/by-nc-nd/4.0/).
1. Introduction
Midbrain dopamine (DA) neurons play an essential role in thecontrol of motor activity and other behaviors related to emotionand cognition. Degeneration of these neurons, especially in thesubstantia nigra compacta (SNc), causes Parkinson’s disease (PD),which is a severe neurological disease primarily characterized byDA dysfunction in the nigrostriatal system (Braak et al., 2003;Jellinger, 2012; Li et al., 2009; Simon et al., 2003).
Several transcription factors regulate the differentiation ofmidbrain DA neuron precursors. Nurr1, a member of the Nur77/NGFI-B subfamily of the nuclear receptor superfamily of transcrip-tion factors, is strongly implicated in the growth, maintenance, andsurvival of DA neurons. Mice harboring null alleles of Nurr1 do notexpress tyrosine hydroxylase (TH) (Baffi et al.,1999; Eells et al., 2001;Kimet al., 2003; Le et al.,1999; Sakurada et al.,1999),which catalyzesthe initial step of DA neurotransmitter biosynthesis. Furthermore,mice deficient in Nurr1(þ/�) show a rapid onset of age-related
ience, Mahoney Institute fority of Pennsylvania, Philadel--0833.
Inc. This is an open access article u
decline in DA signaling that precedes the significant loss of DAneurons (Zhang et al., 2012). Another factor, LIM homeobox tran-scription factor 1 beta (LMX1B), contributes partially to the specifi-cation of DA neuronal progenitors beginning on embryonic day 12.5in the mouse, but LMX1B is not essential for TH gene expression(Smidt et al., 2000). A third transcription factor, the bicoid-relatedhomeodomain-containing transcription factor, Pitx3, plays animportant roles in the development, function, and maintenance ofmidbrain DA neurons (Andersson et al., 2006; Li et al., 2009;Wallenand Perlmann, 2003). Pitx3 is expressed in the SNc and the ventraltegmental area (VTA) DA neurons (Smidt et al., 1997; Zhao et al.,2004). However, the lack of Pitx3 results in the preferential loss ofthe SNc neuronal subpopulation whereas the VTA neurons remainrelatively intact, a phenotype that closely resembles that of PD(Hwang et al., 2003;Maxwell et al., 2005; Smidt et al., 2004; van denMunckhof et al., 2003). Moreover, Pitx3 directly regulates theexpression of a cascade of molecules in DA neurons, including TH,Vmat2, En1, brain-derived neurotrophic factor, and glial cell-derivedneurotrophic factor, indicating that Pitx3 expression is important forthe function of adultmidbrainDAneurons (Kimet al., 2014;Maxwellet al., 2005; Peng et al., 2007, 2011; Yang et al., 2008).
To further identify the biological functions of Pitx3 in themidbrain DA system, we used the Pitx3-null (Pitx3�/�) mice to
nder the CC BY-NC-ND license (http://creativecommons.org/licenses/by-nc-nd/4.0/).

W. Le et al. / Neurobiology of Aging 36 (2015) 3314e3320 3315
examine Pitx3-regulated DA release and the loss of DA neuronsassociated with the decline of motor activities. In the present study,we report that Pitx3-lacking mice (3e5 months in age) showed aloss of DA neurons mainly in the SNc, as revealed by the loss of TH-positive staining. In addition, fast-scan cyclic voltammetry andmicrodialysis assays of DA signaling indicated that the lack of Pitx3caused significant reduction of striatal DA release. Associated withthe loss of DA neurons and DA signaling there were motor-activitydeficits, including impaired motor coordination and decreasedlocomotion. In summary, these data provide direct evidence thatthe lack of Pitx3 caused DA neuron loss, decreased DA signaling, andcontributed to deficits in locomotor activities.
2. Methods
2.1. Pitx3-null mutant mice
Pitx3 heterozygous null mice (Pitx3þ/�) were kindly providedby Professor Beeler, Department of Neurobiology, The University ofChicago, Chicago, IL, USA. The mice were inbred and expanded atthe transgenic animal center of Baylor College of Medicine. Thesiblings of the inbred mice were analyzed for their genotypesthrough the restriction enzyme digestion of polymerase chain re-action product from mouse tail extracted DNA. We used 3- to 5-month-old Pitx3 homozygous null mice for our experiments andtheir wild-type littermates as controls with the same geneticbackground for the behavioral and anatomy experiments. Becausethe Pitx3(�/�) mice displayed a dramatic decrease in DA releasebeginning in the early ages (3e5 months) and there was not adramatic age-associated decline shortly beyond that age group, wegrouped data from mice at 3e5 months of age when examining DArelease with fast-scan cyclic voltammetry. Mice were housed andhandled in accordance with the guidelines approved by the animalcare committee at Baylor College of Medicine.
2.2. Behavioral assessment of Pitx3(�/�) mice
The behavioral performance of Pitx3(�/�) or their wild-typelittermates was evaluated using the rotarod test and the locomo-tor activity test. Motor coordination was determined with anaccelerating rotarod treadmill (Columbus Instruments, Columbus,OH, USA). Initially, each mouse was required to perch on the sta-tionary rod for 30 seconds to accustom itself to the environment.Then the animals were trained at a constant speed of 5 rpm for90 seconds. After this training session, the mice were tested 3 timesat 1-hour intervals on 3 consecutive days for a total of 9 tests.During each test, the rotarod was set at a starting speed of 5 rpm for30 seconds, and the speed was increased by 0.1 revolution persecond. All animals were tested 3 times for each experiment, andthe means of the test results underwent statistical analysis. Loco-motive activities were carried out from9 AM to 11 AM in a relativelydark room, and the behavioral performance was monitored by theAccuScan Digiscan system (AccuScan Instruments, Inc, Columbus,OH, USA). Data collected by computer included total distancetraveled (centimeter per 60minutes) andmoving time (seconds per60 minutes). Each mouse was placed in the testing chamber for30 minutes for adaptation, followed by a 60-minute recording bythe computer-generated automatic analysis system.
2.3. Tyrosine hydroxylase immunohistochemistry
Mice were transcardially perfused with ice-cold phosphate-buffered saline (PBS) (pH 7.4), and the central part of the striatumand the midbrain were dissected for DA biochemistry and TH im-munostaining. The midbrains were fixed, rapidly frozen, and
sectioned into 30-mm slices. The slices were systematically chosenat 150-mm intervals. Free-floating sections were incubated succes-sively for 15 minutes with 0.05% H2O2 in 0.1-M PBS to removeendogenous peroxidase activity for 1 hour with 2% goat serum per0.1% Triton X-100 in 0.1-M PBS to block nonspecific binding sitesand for 24 hours at 4 �C with the primary anti-TH antibody (rabbit,1:4000; Protos Biotech, New York, NY, USA) to detect DA neurons.After washing, sections were then incubated for 2 hours at a routinetime with the appropriate biotinylated secondary antibody (anti-rabbit, 1:200; Vector Laboratories Inc, Burlingame, CA, USA). Theavidin-biotin method was used to amplify the signal (ABC Kit;Vector Laboratories Inc) and 3,3’-diaminobenzidine tetrachloridewas used to visualize bound antibodies.
We used stereological methods to evaluate the number ofTH-positive DA neurons in the SNc using a stereological system(Axioskop 2; Carl Zeiss Inc, Thornwood, NY, USA). Counting wasperformed with the computer-assisted stereological toolbox soft-ware program, Stereo Investigator 7.0 (MicroBrightField, Inc,Willston, VT, USA). The substantia nigra was outlined under2.5� magnification, and 50% of the outlined region was analyzedusing a sampling design generated with the following stereologicparameters: grid size, 200 � 200 mm; counting frame size,150 � 150 mm; and dissector height, 14 mm.
2.4. Determination of striatal DA content
The concentration of DA in the striatal tissue was quantified byhigh-performance liquid chromatography (HPLC). We made coro-nal sections. Although avoiding the ventral striatum that containsthe nucleus accumbens, we selected themiddle part of the striatumby dissection. Then that tissue was homogenized (10% wt/vol) bysonication in ice-cold 0.1-M perchloric acid. Homogenates werecentrifuged at 10,000 g for 10 minutes at 4 �C, and the supernatantwas collected and filtered through acro-disc filters (0.25 mm; FisherScientific, Pittsburgh, PA, USA) and subjected to HPLC (HTEC-500;Eicom, Kyoto, Japan) with the SC-3ODS column (EICOMPAK; Eicom,Kyoto, Japan) and detected by an electrochemical detector (AD In-struments Pty Ltd, Castle Hill, New South Wales, Australia). Themobile phase consisted of 0.1-mM citric acid, 0.1-M sodium acetate,220-mg/L octane sulfate sodium, 5-mg/L EDTA, and 20% methanol.The pH was adjusted to 3.5.
2.5. Fast-scan cyclic voltammetric measurement of DA signals fromstriatal brain slices
A combination of ketamine, xylazine, and acepromazine wasused to achieve deep anesthesia before mice were decapitated andthe brains were rapidly removed. Horizontal slices (350 mm) werecut on a vibratome, and they were incubated at 32 � 0.5 �C for30 minutes (Zhou et al., 2001). Then the slices were held at roomtemperature for>30 minutes, and they were studied at 34� 1 �C inartificial cerebrospinal fluid: 125-mM NaCl, 2.5-mM KCl, 1.3-mMMgCl2, 2.5-mM CaCl2, 26-mM NaHCO3, 1.25-mM NaH2PO4, and10-mM glucose, saturated with 95% O2 and 5% CO2.
Carbon-fiber microelectrodes (10-mm diameter and approxi-mately 100 mm exposed length; P55s, Amoco Polymers, Greenville,SC, USA) were used to perform fast-scan cyclic voltammetry withinthemiddle portion of the dorsolateral striatum (Zhang et al., 2009a;Zhou et al., 2001). The carbon-fiber microelectrode potential waslinearly scanned at 10 Hz from 0 to �400 to 1000 to �400 to 0 mVagainst a silver-silver chloride reference electrode at a rate of300 mV/ms. The data were acquired and analyzed using an Axo-patch 200B amplifier, a Digidata1320 interface, and a pClamp 8system (Axon Instruments Inc, Union City, CA, USA). The back-ground current was subtracted digitally from the voltammograms,
W. Le et al. / Neurobiology of Aging 36 (2015) 3314e33203316
which were sampled at 50 kHz. The peak amplitude oxidationcurrents was at roughly 600 mV for DA, and each voltammogramwas converted into a DA concentration based on a post-experimental calibration of the carbon-fiber electrode againstfresh solutions of 0.5e10 mM DA.
A bipolar tungsten electrode was used to deliver intrastriatalstimuli. The poles of the stimulating electrode were about 150 mmaway from each other, and the tip of the carbon-fiber recordingmicroelectrode was about 200 mm away from each of the 2 poles ofthe stimulating electrode. Different stimulation protocols wereused, and phasic burst stimulations were applied at an intraburstfrequency of 20 Hz based on our previous in vivo unit recordings ofDA neuron bursts from freely moving rats (Zhang et al., 2009b).Each stimulus pulse was 1 ms in duration and about 0.6 mA atconstant current. Unless trains of stimuli were applied, isolatedstimuli were separated by 2 minutes to allow recovery of the DArelease.
The electrically stimulated DA response was measured as thepeak amplitude (mM) or was measured as the area under the curveof DA concentration (mM-s). The phasic and/or tonic relative DAsignal was calculated by comparing the burst evoked DA signal tothe single-pulseeevoked DA signal. The results are presented as themeans � standard error of the mean. Two-way analysis of variancewas used to judge the changes in frequency-dependent DA releasebetween genotypes.
2.6. In vivo microdialysis and HPLC for DA quantification
Microdialysis was performed as described previously (Donget al., 2010; Zhang et al., 2012). Guide cannulae (CMA/7) (CMA/Microdialysis, Solna, Sweden) were aimed at the dorsal striatum.That is, we avoided the ventral striatum containing the nucleusaccumbens while placing the probe in the middle of the striatum.The probe was not in the extreme dorsal area but rather morecentral in location. The stereotaxic coordinates (relative to bregma)were 0.38-mm anteroposterior, 2.0-mm lateral, and �2.5-mmdorsal-ventral, and the post-microdialysis anatomy found all theprobes within the striatum indicated in Fig. 3C (Paxinos andFranklin, 2001). We allowed 3e5 days of recovery from surgery.At least 14 hours before the experiment, the probe was positionedand microdialysate buffer (149-mM NaCl, 2.8-mM KCl, 1.2-mMCaCl2, 1.2-mM MgCl2, and 0.25-mM ascorbic acid, 5.4-mM D-glucose) was perfused at 0.5 mL/min overnight and, then, 2.0-mL/min beginning at least 1 hour before baseline sampling.
The DA content of microdialysates was determined by HPLC:pump (Model 582; ESA, Inc, Chelmsford, MA, USA), autosampler(Model 542; ESA, Inc), and an HR-80 � 3.2 mm column (3-mmparticle size; ESA Inc). A coulometric cell (5014B; ESA, Inc) was
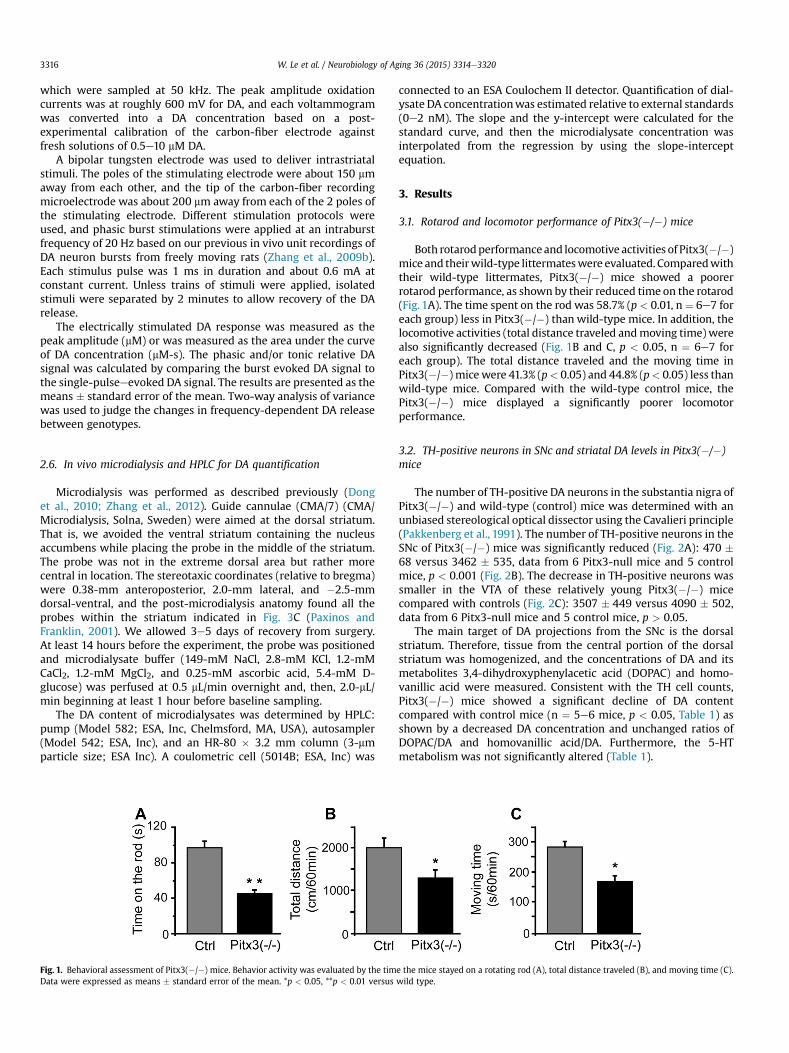
Fig. 1. Behavioral assessment of Pitx3(�/�) mice. Behavior activity was evaluated by the timData were expressed as means � standard error of the mean. *p < 0.05, **p < 0.01 versus
connected to an ESA Coulochem II detector. Quantification of dial-ysate DA concentrationwas estimated relative to external standards(0e2 nM). The slope and the y-intercept were calculated for thestandard curve, and then the microdialysate concentration wasinterpolated from the regression by using the slope-interceptequation.
3. Results
3.1. Rotarod and locomotor performance of Pitx3(�/�) mice
Both rotarodperformanceand locomotive activities of Pitx3(�/�)mice and theirwild-type littermateswere evaluated. Comparedwiththeir wild-type littermates, Pitx3(�/�) mice showed a poorerrotarod performance, as shown by their reduced time on the rotarod(Fig. 1A). The time spent on the rod was 58.7% (p < 0.01, n¼ 6e7 foreach group) less in Pitx3(�/�) than wild-type mice. In addition, thelocomotive activities (total distance traveled andmoving time) werealso significantly decreased (Fig. 1B and C, p < 0.05, n ¼ 6e7 foreach group). The total distance traveled and the moving time inPitx3(�/�) micewere 41.3% (p< 0.05) and 44.8% (p< 0.05) less thanwild-type mice. Compared with the wild-type control mice, thePitx3(�/�) mice displayed a significantly poorer locomotorperformance.
3.2. TH-positive neurons in SNc and striatal DA levels in Pitx3(�/�)mice
The number of TH-positive DA neurons in the substantia nigra ofPitx3(�/�) and wild-type (control) mice was determined with anunbiased stereological optical dissector using the Cavalieri principle(Pakkenberg et al., 1991). The number of TH-positive neurons in theSNc of Pitx3(�/�) mice was significantly reduced (Fig. 2A): 470 �68 versus 3462 � 535, data from 6 Pitx3-null mice and 5 controlmice, p < 0.001 (Fig. 2B). The decrease in TH-positive neurons wassmaller in the VTA of these relatively young Pitx3(�/�) micecompared with controls (Fig. 2C): 3507 � 449 versus 4090 � 502,data from 6 Pitx3-null mice and 5 control mice, p > 0.05.
The main target of DA projections from the SNc is the dorsalstriatum. Therefore, tissue from the central portion of the dorsalstriatum was homogenized, and the concentrations of DA and itsmetabolites 3,4-dihydroxyphenylacetic acid (DOPAC) and homo-vanillic acid were measured. Consistent with the TH cell counts,Pitx3(�/�) mice showed a significant decline of DA contentcompared with control mice (n ¼ 5e6 mice, p < 0.05, Table 1) asshown by a decreased DA concentration and unchanged ratios ofDOPAC/DA and homovanillic acid/DA. Furthermore, the 5-HTmetabolism was not significantly altered (Table 1).
e the mice stayed on a rotating rod (A), total distance traveled (B), and moving time (C).wild type.
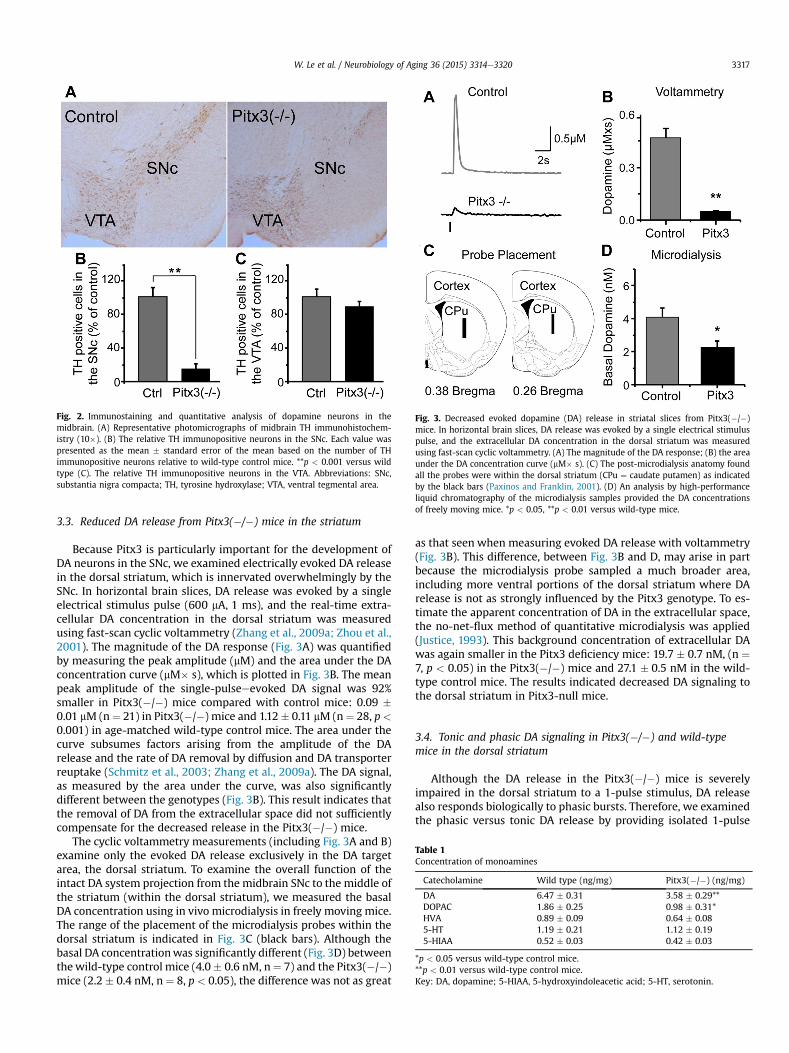
Fig. 3. Decreased evoked dopamine (DA) release in striatal slices from Pitx3(�/�)mice. In horizontal brain slices, DA release was evoked by a single electrical stimuluspulse, and the extracellular DA concentration in the dorsal striatum was measuredusing fast-scan cyclic voltammetry. (A) The magnitude of the DA response; (B) the areaunder the DA concentration curve (mM� s). (C) The post-microdialysis anatomy foundall the probes were within the dorsal striatum (CPu ¼ caudate putamen) as indicatedby the black bars (Paxinos and Franklin, 2001). (D) An analysis by high-performanceliquid chromatography of the microdialysis samples provided the DA concentrationsof freely moving mice. *p < 0.05, **p < 0.01 versus wild-type mice.
Fig. 2. Immunostaining and quantitative analysis of dopamine neurons in themidbrain. (A) Representative photomicrographs of midbrain TH immunohistochem-istry (10�). (B) The relative TH immunopositive neurons in the SNc. Each value waspresented as the mean � standard error of the mean based on the number of THimmunopositive neurons relative to wild-type control mice. **p < 0.001 versus wildtype (C). The relative TH immunopositive neurons in the VTA. Abbreviations: SNc,substantia nigra compacta; TH, tyrosine hydroxylase; VTA, ventral tegmental area.
Table 1Concentration of monoamines
Catecholamine Wild type (ng/mg) Pitx3(�/�) (ng/mg)
DA 6.47 � 0.31 3.58 � 0.29**DOPAC 1.86 � 0.25 0.98 � 0.31*HVA 0.89 � 0.09 0.64 � 0.085-HT 1.19 � 0.21 1.12 � 0.195-HIAA 0.52 � 0.03 0.42 � 0.03
*p < 0.05 versus wild-type control mice.**p < 0.01 versus wild-type control mice.Key: DA, dopamine; 5-HIAA, 5-hydroxyindoleacetic acid; 5-HT, serotonin.
W. Le et al. / Neurobiology of Aging 36 (2015) 3314e3320 3317
3.3. Reduced DA release from Pitx3(�/�) mice in the striatum
Because Pitx3 is particularly important for the development ofDA neurons in the SNc, we examined electrically evoked DA releasein the dorsal striatum, which is innervated overwhelmingly by theSNc. In horizontal brain slices, DA release was evoked by a singleelectrical stimulus pulse (600 mA, 1 ms), and the real-time extra-cellular DA concentration in the dorsal striatum was measuredusing fast-scan cyclic voltammetry (Zhang et al., 2009a; Zhou et al.,2001). The magnitude of the DA response (Fig. 3A) was quantifiedby measuring the peak amplitude (mM) and the area under the DAconcentration curve (mM� s), which is plotted in Fig. 3B. The meanpeak amplitude of the single-pulseeevoked DA signal was 92%smaller in Pitx3(�/�) mice compared with control mice: 0.09 �0.01 mM (n¼ 21) in Pitx3(�/�) mice and 1.12� 0.11 mM (n¼ 28, p<
0.001) in age-matched wild-type control mice. The area under thecurve subsumes factors arising from the amplitude of the DArelease and the rate of DA removal by diffusion and DA transporterreuptake (Schmitz et al., 2003; Zhang et al., 2009a). The DA signal,as measured by the area under the curve, was also significantlydifferent between the genotypes (Fig. 3B). This result indicates thatthe removal of DA from the extracellular space did not sufficientlycompensate for the decreased release in the Pitx3(�/�) mice.
The cyclic voltammetry measurements (including Fig. 3A and B)examine only the evoked DA release exclusively in the DA targetarea, the dorsal striatum. To examine the overall function of theintact DA system projection from the midbrain SNc to the middle ofthe striatum (within the dorsal striatum), we measured the basalDA concentration using in vivo microdialysis in freely moving mice.The range of the placement of the microdialysis probes within thedorsal striatum is indicated in Fig. 3C (black bars). Although thebasal DA concentrationwas significantly different (Fig. 3D) betweenthe wild-type control mice (4.0� 0.6 nM, n¼ 7) and the Pitx3(�/�)mice (2.2 � 0.4 nM, n ¼ 8, p < 0.05), the difference was not as great
as that seen when measuring evoked DA release with voltammetry(Fig. 3B). This difference, between Fig. 3B and D, may arise in partbecause the microdialysis probe sampled a much broader area,including more ventral portions of the dorsal striatum where DArelease is not as strongly influenced by the Pitx3 genotype. To es-timate the apparent concentration of DA in the extracellular space,the no-net-flux method of quantitative microdialysis was applied(Justice, 1993). This background concentration of extracellular DAwas again smaller in the Pitx3 deficiency mice: 19.7 � 0.7 nM, (n ¼7, p < 0.05) in the Pitx3(�/�) mice and 27.1 � 0.5 nM in the wild-type control mice. The results indicated decreased DA signaling tothe dorsal striatum in Pitx3-null mice.
3.4. Tonic and phasic DA signaling in Pitx3(�/�) and wild-typemice in the dorsal striatum
Although the DA release in the Pitx3(�/�) mice is severelyimpaired in the dorsal striatum to a 1-pulse stimulus, DA releasealso responds biologically to phasic bursts. Therefore, we examinedthe phasic versus tonic DA release by providing isolated 1-pulse
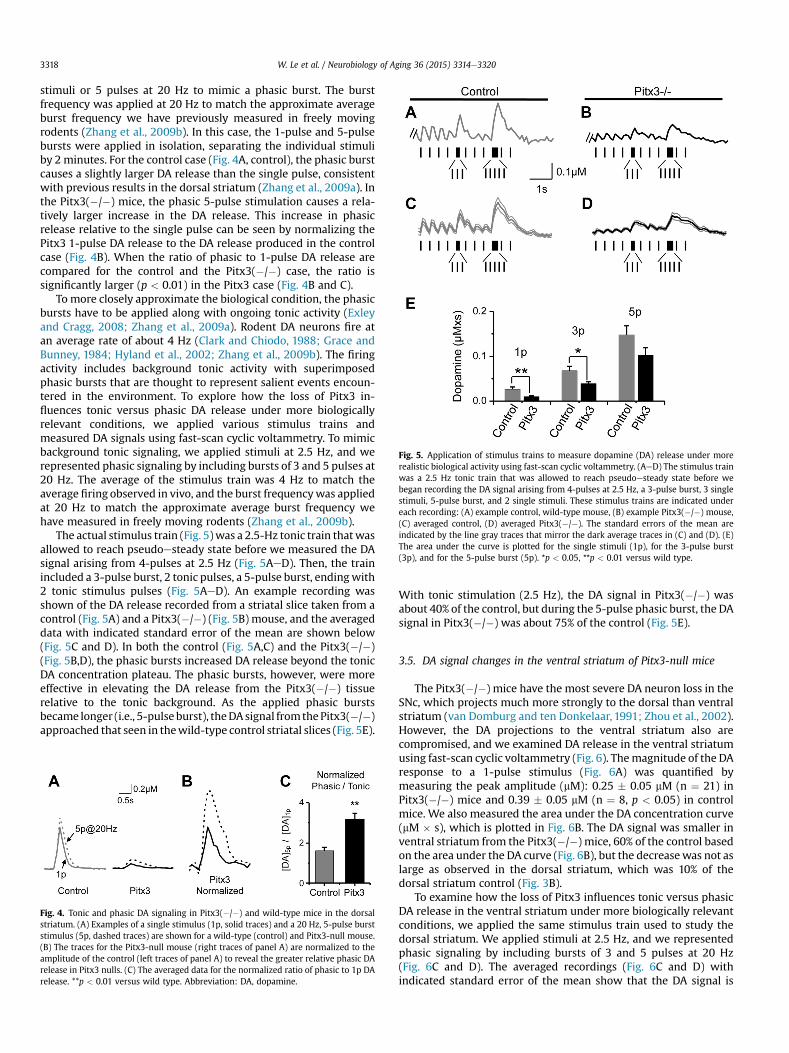
Fig. 5. Application of stimulus trains to measure dopamine (DA) release under morerealistic biological activity using fast-scan cyclic voltammetry. (AeD) The stimulus trainwas a 2.5 Hz tonic train that was allowed to reach pseudoesteady state before webegan recording the DA signal arising from 4-pulses at 2.5 Hz, a 3-pulse burst, 3 singlestimuli, 5-pulse burst, and 2 single stimuli. These stimulus trains are indicated undereach recording: (A) example control, wild-type mouse, (B) example Pitx3(�/�) mouse,(C) averaged control, (D) averaged Pitx3(�/�). The standard errors of the mean areindicated by the line gray traces that mirror the dark average traces in (C) and (D). (E)The area under the curve is plotted for the single stimuli (1p), for the 3-pulse burst(3p), and for the 5-pulse burst (5p). *p < 0.05, **p < 0.01 versus wild type.
W. Le et al. / Neurobiology of Aging 36 (2015) 3314e33203318
stimuli or 5 pulses at 20 Hz to mimic a phasic burst. The burstfrequency was applied at 20 Hz to match the approximate averageburst frequency we have previously measured in freely movingrodents (Zhang et al., 2009b). In this case, the 1-pulse and 5-pulsebursts were applied in isolation, separating the individual stimuliby 2 minutes. For the control case (Fig. 4A, control), the phasic burstcauses a slightly larger DA release than the single pulse, consistentwith previous results in the dorsal striatum (Zhang et al., 2009a). Inthe Pitx3(�/�) mice, the phasic 5-pulse stimulation causes a rela-tively larger increase in the DA release. This increase in phasicrelease relative to the single pulse can be seen by normalizing thePitx3 1-pulse DA release to the DA release produced in the controlcase (Fig. 4B). When the ratio of phasic to 1-pulse DA release arecompared for the control and the Pitx3(�/�) case, the ratio issignificantly larger (p < 0.01) in the Pitx3 case (Fig. 4B and C).
To more closely approximate the biological condition, the phasicbursts have to be applied along with ongoing tonic activity (Exleyand Cragg, 2008; Zhang et al., 2009a). Rodent DA neurons fire atan average rate of about 4 Hz (Clark and Chiodo, 1988; Grace andBunney, 1984; Hyland et al., 2002; Zhang et al., 2009b). The firingactivity includes background tonic activity with superimposedphasic bursts that are thought to represent salient events encoun-tered in the environment. To explore how the loss of Pitx3 in-fluences tonic versus phasic DA release under more biologicallyrelevant conditions, we applied various stimulus trains andmeasured DA signals using fast-scan cyclic voltammetry. To mimicbackground tonic signaling, we applied stimuli at 2.5 Hz, and werepresented phasic signaling by including bursts of 3 and 5 pulses at20 Hz. The average of the stimulus train was 4 Hz to match theaverage firing observed in vivo, and the burst frequencywas appliedat 20 Hz to match the approximate average burst frequency wehave measured in freely moving rodents (Zhang et al., 2009b).
The actual stimulus train (Fig. 5)was a 2.5-Hz tonic train thatwasallowed to reach pseudoesteady state before we measured the DAsignal arising from 4-pulses at 2.5 Hz (Fig. 5AeD). Then, the trainincluded a 3-pulse burst, 2 tonic pulses, a 5-pulse burst, endingwith2 tonic stimulus pulses (Fig. 5AeD). An example recording wasshown of the DA release recorded from a striatal slice taken from acontrol (Fig. 5A) and a Pitx3(�/�) (Fig. 5B) mouse, and the averageddata with indicated standard error of the mean are shown below(Fig. 5C and D). In both the control (Fig. 5A,C) and the Pitx3(�/�)(Fig. 5B,D), the phasic bursts increased DA release beyond the tonicDA concentration plateau. The phasic bursts, however, were moreeffective in elevating the DA release from the Pitx3(�/�) tissuerelative to the tonic background. As the applied phasic burstsbecame longer (i.e., 5-pulse burst), theDAsignal fromthePitx3(�/�)approached that seen in thewild-type control striatal slices (Fig. 5E).
Fig. 4. Tonic and phasic DA signaling in Pitx3(�/�) and wild-type mice in the dorsalstriatum. (A) Examples of a single stimulus (1p, solid traces) and a 20 Hz, 5-pulse burststimulus (5p, dashed traces) are shown for a wild-type (control) and Pitx3-null mouse.(B) The traces for the Pitx3-null mouse (right traces of panel A) are normalized to theamplitude of the control (left traces of panel A) to reveal the greater relative phasic DArelease in Pitx3 nulls. (C) The averaged data for the normalized ratio of phasic to 1p DArelease. **p < 0.01 versus wild type. Abbreviation: DA, dopamine.
With tonic stimulation (2.5 Hz), the DA signal in Pitx3(�/�) wasabout 40% of the control, but during the 5-pulse phasic burst, the DAsignal in Pitx3(�/�) was about 75% of the control (Fig. 5E).
3.5. DA signal changes in the ventral striatum of Pitx3-null mice
The Pitx3(�/�) mice have the most severe DA neuron loss in theSNc, which projects much more strongly to the dorsal than ventralstriatum (van Domburg and ten Donkelaar, 1991; Zhou et al., 2002).However, the DA projections to the ventral striatum also arecompromised, and we examined DA release in the ventral striatumusing fast-scan cyclic voltammetry (Fig. 6). Themagnitude of the DAresponse to a 1-pulse stimulus (Fig. 6A) was quantified bymeasuring the peak amplitude (mM): 0.25 � 0.05 mM (n ¼ 21) inPitx3(�/�) mice and 0.39 � 0.05 mM (n ¼ 8, p < 0.05) in controlmice. We also measured the area under the DA concentration curve(mM � s), which is plotted in Fig. 6B. The DA signal was smaller inventral striatum from the Pitx3(�/�) mice, 60% of the control basedon the area under the DA curve (Fig. 6B), but the decreasewas not aslarge as observed in the dorsal striatum, which was 10% of thedorsal striatum control (Fig. 3B).
To examine how the loss of Pitx3 influences tonic versus phasicDA release in the ventral striatum under more biologically relevantconditions, we applied the same stimulus train used to study thedorsal striatum. We applied stimuli at 2.5 Hz, and we representedphasic signaling by including bursts of 3 and 5 pulses at 20 Hz(Fig. 6C and D). The averaged recordings (Fig. 6C and D) withindicated standard error of the mean show that the DA signal is
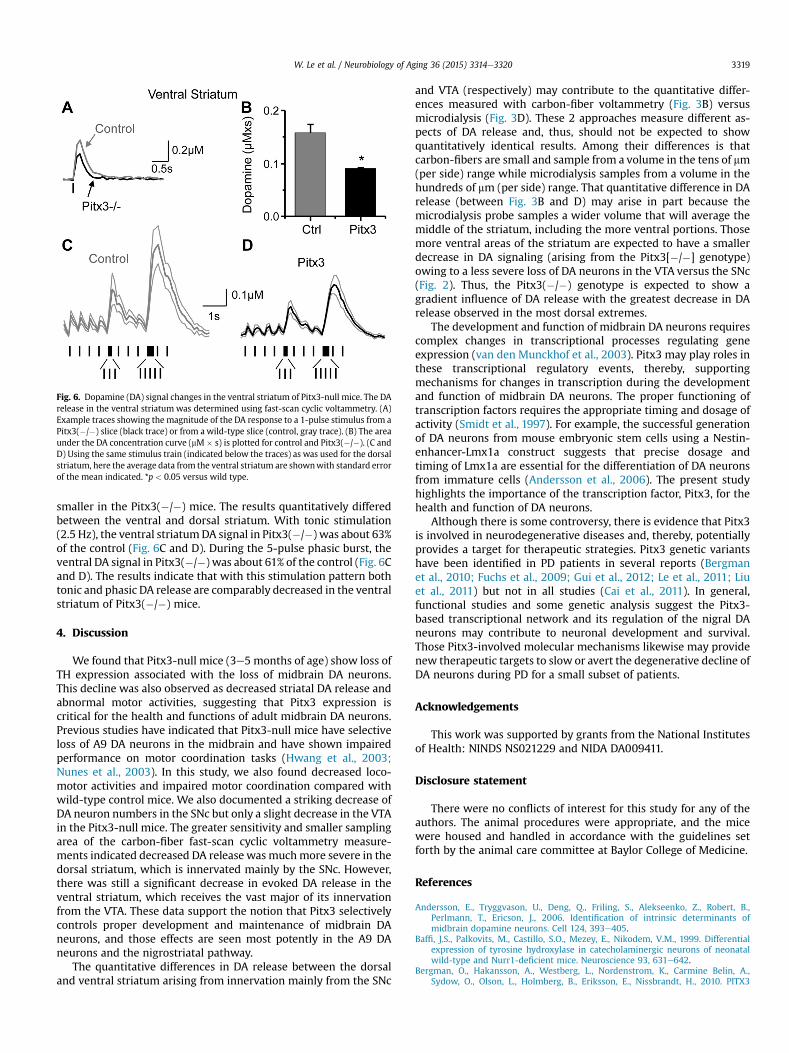
Fig. 6. Dopamine (DA) signal changes in the ventral striatum of Pitx3-null mice. The DArelease in the ventral striatum was determined using fast-scan cyclic voltammetry. (A)Example traces showing the magnitude of the DA response to a 1-pulse stimulus from aPitx3(�/�) slice (black trace) or from a wild-type slice (control, gray trace). (B) The areaunder the DA concentration curve (mM � s) is plotted for control and Pitx3(�/�). (C andD) Using the same stimulus train (indicated below the traces) as was used for the dorsalstriatum, here the average data from the ventral striatum are shownwith standard errorof the mean indicated. *p < 0.05 versus wild type.
W. Le et al. / Neurobiology of Aging 36 (2015) 3314e3320 3319
smaller in the Pitx3(�/�) mice. The results quantitatively differedbetween the ventral and dorsal striatum. With tonic stimulation(2.5 Hz), the ventral striatumDA signal in Pitx3(�/�) was about 63%of the control (Fig. 6C and D). During the 5-pulse phasic burst, theventral DA signal in Pitx3(�/�) was about 61% of the control (Fig. 6Cand D). The results indicate that with this stimulation pattern bothtonic and phasic DA release are comparably decreased in the ventralstriatum of Pitx3(�/�) mice.
4. Discussion
We found that Pitx3-null mice (3e5 months of age) show loss ofTH expression associated with the loss of midbrain DA neurons.This decline was also observed as decreased striatal DA release andabnormal motor activities, suggesting that Pitx3 expression iscritical for the health and functions of adult midbrain DA neurons.Previous studies have indicated that Pitx3-null mice have selectiveloss of A9 DA neurons in the midbrain and have shown impairedperformance on motor coordination tasks (Hwang et al., 2003;Nunes et al., 2003). In this study, we also found decreased loco-motor activities and impaired motor coordination compared withwild-type control mice. We also documented a striking decrease ofDA neuron numbers in the SNc but only a slight decrease in the VTAin the Pitx3-null mice. The greater sensitivity and smaller samplingarea of the carbon-fiber fast-scan cyclic voltammetry measure-ments indicated decreased DA release was muchmore severe in thedorsal striatum, which is innervated mainly by the SNc. However,there was still a significant decrease in evoked DA release in theventral striatum, which receives the vast major of its innervationfrom the VTA. These data support the notion that Pitx3 selectivelycontrols proper development and maintenance of midbrain DAneurons, and those effects are seen most potently in the A9 DAneurons and the nigrostriatal pathway.
The quantitative differences in DA release between the dorsaland ventral striatum arising from innervation mainly from the SNc
and VTA (respectively) may contribute to the quantitative differ-ences measured with carbon-fiber voltammetry (Fig. 3B) versusmicrodialysis (Fig. 3D). These 2 approaches measure different as-pects of DA release and, thus, should not be expected to showquantitatively identical results. Among their differences is thatcarbon-fibers are small and sample from a volume in the tens of mm(per side) range while microdialysis samples from a volume in thehundreds of mm (per side) range. That quantitative difference in DArelease (between Fig. 3B and D) may arise in part because themicrodialysis probe samples a wider volume that will average themiddle of the striatum, including the more ventral portions. Thosemore ventral areas of the striatum are expected to have a smallerdecrease in DA signaling (arising from the Pitx3[�/�] genotype)owing to a less severe loss of DA neurons in the VTA versus the SNc(Fig. 2). Thus, the Pitx3(�/�) genotype is expected to show agradient influence of DA release with the greatest decrease in DArelease observed in the most dorsal extremes.
The development and function of midbrain DA neurons requirescomplex changes in transcriptional processes regulating geneexpression (van den Munckhof et al., 2003). Pitx3 may play roles inthese transcriptional regulatory events, thereby, supportingmechanisms for changes in transcription during the developmentand function of midbrain DA neurons. The proper functioning oftranscription factors requires the appropriate timing and dosage ofactivity (Smidt et al., 1997). For example, the successful generationof DA neurons from mouse embryonic stem cells using a Nestin-enhancer-Lmx1a construct suggests that precise dosage andtiming of Lmx1a are essential for the differentiation of DA neuronsfrom immature cells (Andersson et al., 2006). The present studyhighlights the importance of the transcription factor, Pitx3, for thehealth and function of DA neurons.
Although there is some controversy, there is evidence that Pitx3is involved in neurodegenerative diseases and, thereby, potentiallyprovides a target for therapeutic strategies. Pitx3 genetic variantshave been identified in PD patients in several reports (Bergmanet al., 2010; Fuchs et al., 2009; Gui et al., 2012; Le et al., 2011; Liuet al., 2011) but not in all studies (Cai et al., 2011). In general,functional studies and some genetic analysis suggest the Pitx3-based transcriptional network and its regulation of the nigral DAneurons may contribute to neuronal development and survival.Those Pitx3-involved molecular mechanisms likewise may providenew therapeutic targets to slow or avert the degenerative decline ofDA neurons during PD for a small subset of patients.
Acknowledgements
This work was supported by grants from the National Institutesof Health: NINDS NS021229 and NIDA DA009411.
Disclosure statement
There were no conflicts of interest for this study for any of theauthors. The animal procedures were appropriate, and the micewere housed and handled in accordance with the guidelines setforth by the animal care committee at Baylor College of Medicine.
References
Andersson, E., Tryggvason, U., Deng, Q., Friling, S., Alekseenko, Z., Robert, B.,Perlmann, T., Ericson, J., 2006. Identification of intrinsic determinants ofmidbrain dopamine neurons. Cell 124, 393e405.
Baffi, J.S., Palkovits, M., Castillo, S.O., Mezey, E., Nikodem, V.M., 1999. Differentialexpression of tyrosine hydroxylase in catecholaminergic neurons of neonatalwild-type and Nurr1-deficient mice. Neuroscience 93, 631e642.
Bergman, O., Hakansson, A., Westberg, L., Nordenstrom, K., Carmine Belin, A.,Sydow, O., Olson, L., Holmberg, B., Eriksson, E., Nissbrandt, H., 2010. PITX3

W. Le et al. / Neurobiology of Aging 36 (2015) 3314e33203320
polymorphism is associated with early onset Parkinson’s disease. Neurobiol.Aging 31, 114e117.
Braak, H., Del Tredici, K., Rub, U., de Vos, R.A., Jansen Steur, E.N., Braak, E., 2003.Staging of brain pathology related to sporadic Parkinson’s disease. Neurobiol.Aging 24, 197e211.
Cai, Y., Ding, H., Gu, Z., Baskys, A., Ma, J., Chan, P., 2011. PITX3 polymorphism is notassociated with Parkinson’s disease in a Chinese population. Neurosci. Lett. 505,260e262.
Clark, D., Chiodo, L.A., 1988. Electrophysiological and pharmacological character-ization of identified nigrostriatal and mesoaccumbens dopamine neurons in therat. Synapse 2, 474e485.
Dong, Y., Zhang, T., Li, W., Doyon, W.M., Dani, J.A., 2010. Route of nicotine admin-istration influences in vivo dopamine neuron activity: habituation, needle in-jection, and cannula infusion. J. Mol. Neurosci. 40, 164e171.
Eells, J.B., Rives, J.E., Yeung, S.K., Nikodem, V.M., 2001. In vitro regulated expressionof tyrosine hydroxylase in ventral midbrain neurons from Nurr1-null mousepups. J. Neurosci. Res. 64, 322e330.
Exley, R., Cragg, S.J., 2008. Presynaptic nicotinic receptors: a dynamic and diversecholinergic filter of striatal dopamine neurotransmission. Br. J. Pharmacol. 153(Suppl 1), S283eS297.
Fuchs, J., Mueller, J.C., Lichtner, P., Schulte, C., Munz, M., Berg, D., Wullner, U., Illig, T.,Sharma, M., Gasser, T., 2009. The transcription factor PITX3 is associated withsporadic Parkinson’s disease. Neurobiol. Aging 30, 731e738.
Grace, A.A., Bunney, B.S., 1984. The control of firing pattern in nigral dopamineneurons: single spike firing. J. Neurosci. 4, 2866e2876.
Gui, Y., Zhao, Y., Liu, H., Fu, J., Xu, Z., Hu, X., 2012. A novel synonymous SNP in PITX3is associated with Parkinson’s disease in Chinese population. Swiss Med. Wkly142, w13521.
Hwang, D.Y., Ardayfio, P., Kang, U.J., Semina, E.V., Kim, K.S., 2003. Selective loss ofdopaminergic neurons in the substantia nigra of Pitx3-deficient aphakia mice.Brain Res. Mol. Brain Res. 114, 123e131.
Hyland, B.I., Reynolds, J.N., Hay, J., Perk, C.G., Miller, R., 2002. Firing modes ofmidbrain dopamine cells in the freely moving rat. Neuroscience 114, 475e492.
Jellinger, K.A., 2012. Neuropathology of sporadic Parkinson’s disease: evaluation andchanges of concepts. Mov. Disord. 27, 8e30.
Justice Jr., J.B., 1993. Quantitative microdialysis of neurotransmitters. J. Neurosci.Methods 48, 263e276.
Kim, H., Quan, X., Seong, Y., Kim, J., 2014. Impaired motor coordination in Pitx3overexpression mice. Biochem. Biophys. Res. Commun. 446, 1211e1218.
Kim, K.S., Kim, C.H., Hwang, D.Y., Seo, H., Chung, S., Hong, S.J., Lim, J.K., Anderson, T.,Isacson, O., 2003. Orphan nuclear receptor Nurr1 directly transactivates thepromoter activity of the tyrosine hydroxylase gene in a cell-specific manner.J. Neurochem. 85, 622e634.
Le, W., Conneely, O.M., Zou, L., He, Y., Saucedo-Cardenas, O., Jankovic, J., Mosier, D.R.,Appel, S.H., 1999. Selective agenesis of mesencephalic dopaminergic neurons inNurr1-deficient mice. Exp. Neurol. 159, 451e458.
Le, W., Nguyen, D., Lin, X.W., Rawal, P., Huang, M., Ding, Y., Xie, W., Deng, H.,Jankovic, J., 2011. Transcription factor PITX3 gene in Parkinson’s disease. Neu-robiol. Aging 32, 750e753.
Li, J., Dani, J.A., Le,W., 2009. The role of transcription factor Pitx3 in dopamine neurondevelopment and Parkinson’s disease. Curr. Top Med. Chem. 9, 855e859.
Liu, J., Sun, Q.Y., Tang, B.S., Hu, L., Yu, R.H., Wang, L., Shi, C.H., Yan, X.X., Pan, Q.,Xia, K., Guo, J.F., 2011. PITX3 gene polymorphism is associated with Parkinson’sdisease in Chinese population. Brain Res. 1392, 116e120.
Maxwell, S.L., Ho, H.Y., Kuehner, E., Zhao, S., Li, M., 2005. Pitx3 regulates tyrosinehydroxylase expression in the substantia nigra and identifies a subgroup ofmesencephalic dopaminergic progenitor neurons during mouse development.Dev. Biol. 282, 467e479.
Nunes, I., Tovmasian, L.T., Silva, R.M., Burke, R.E., Goff, S.P., 2003. Pitx3 is required fordevelopment of substantia nigra dopaminergic neurons. Proc. Natl. Acad. Sci.U. S. A. 100, 4245e4250.
Pakkenberg, B., Moller, A., Gundersen, H.J., Mouritzen Dam, A., Pakkenberg, H., 1991.The absolute number of nerve cells in substantia nigra in normal subjects and inpatients with Parkinson’s disease estimated with an unbiased stereologicalmethod. J. Neurol. Neurosurg. Psychiatry 54, 30e33.
Paxinos, G., Franklin, K.B.J., 2001. The Mouse Brain in Stereotaxic Coordinates, sec-ond ed. Academic Press, Elsevier Science, USA.
Peng, C., Aron, L., Klein, R., Li, M., Wurst, W., Prakash, N., Le, W., 2011. Pitx3 is acritical mediator of GDNF-induced BDNF expression in nigrostriatal dopami-nergic neurons. J. Neurosci. 31, 12802e12815.
Peng, C., Fan, S., Li, X., Fan, X., Ming, M., Sun, Z., Le, W., 2007. Overexpression of pitx3upregulates expression of BDNF and GDNF in SH-SY5Y cells and primary ventralmesencephalic cultures. FEBS Lett. 581, 1357e1361.
Sakurada, K., Ohshima-Sakurada, M., Palmer, T.D., Gage, F.H., 1999. Nurr1, an orphannuclear receptor, is a transcriptional activator of endogenous tyrosine hydrox-ylase in neural progenitor cells derived from the adult brain. Development 126,4017e4026.
Schmitz, Y., Benoit-Marand, M., Gonon, F., Sulzer, D., 2003. Presynaptic regulation ofdopaminergic neurotransmission. J. Neurochem. 87, 273e289.
Simon, H.H., Bhatt, L., Gherbassi, D., Sgado, P., Alberi, L., 2003. Midbrain dopami-nergic neurons: determination of their developmental fate by transcriptionfactors. Ann. N. Y. Acad. Sci. 991, 36e47.
Smidt, M.P., Asbreuk, C.H., Cox, J.J., Chen, H., Johnson, R.L., Burbach, J.P., 2000.A second independent pathway for development of mesencephalic dopami-nergic neurons requires Lmx1b. Nat. Neurosci. 3, 337e341.
Smidt, M.P., Smits, S.M., Bouwmeester, H., Hamers, F.P., van der Linden, A.J.,Hellemons, A.J., Graw, J., Burbach, J.P., 2004. Early developmental failure ofsubstantia nigra dopamine neurons in mice lacking the homeodomain genePitx3. Development 131, 1145e1155.
Smidt, M.P., van Schaick, H.S., Lanctot, C., Tremblay, J.J., Cox, J.J., van der Kleij, A.A.,Wolterink, G., Drouin, J., Burbach, J.P., 1997. A homeodomain gene Ptx3 hashighly restricted brain expression in mesencephalic dopaminergic neurons.Proc. Natl. Acad. Sci. U. S. A. 94, 13305e13310.
van den Munckhof, P., Luk, K.C., Ste-Marie, L., Montgomery, J., Blanchet, P.J.,Sadikot, A.F., Drouin, J., 2003. Pitx3 is required for motor activity and for survivalof a subset of midbrain dopaminergic neurons. Development 130, 2535e2542.
van Domburg, P.H., ten Donkelaar, H.J., 1991. The human substantia nigra andventral tegmental area. A neuroanatomical study with notes on aging and agingdiseases. Adv. Anat. Embryol. Cell Biol. 121, 1e132.
Wallen, A., Perlmann, T., 2003. Transcriptional control of dopamine neuron devel-opment. Ann. N. Y. Acad. Sci. 991, 48e60.
Yang, D., Peng, C., Li, X., Fan, X., Li, L., Ming, M., Chen, S., Le, W., 2008. Pitx3-transfected astrocytes secrete brain-derived neurotrophic factor and glial cellline-derived neurotrophic factor and protect dopamine neurons in mesen-cephalon cultures. J. Neurosci. Res. 86, 3393e3400.
Zhang, L., Doyon, W.M., Clark, J.J., Phillips, P.E., Dani, J.A., 2009a. Controls of tonicand phasic dopamine transmission in the dorsal and ventral striatum. Mol.Pharmacol. 76, 396e404.
Zhang, L., Le, W., Xie, W., Dani, J.A., 2012. Age-related changes in dopamine signalingin Nurr1 deficient mice as a model of Parkinson’s disease. Neurobiol. Aging 33,1001.e7e1001.e16.
Zhang, T., Zhang, L., Liang, Y., Siapas, A.G., Zhou, F.M., Dani, J.A., 2009b. Dopaminesignaling differences in the nucleus accumbens and dorsal striatum exploitedby nicotine. J. Neurosci. 29, 4035e4043.
Zhao, S., Maxwell, S., Jimenez-Beristain, A., Vives, J., Kuehner, E., Zhao, J., O’Brien, C.,de Felipe, C., Semina, E., Li, M., 2004. Generation of embryonic stem cells andtransgenic mice expressing green fluorescence protein in midbrain dopami-nergic neurons. Eur. J. Neurosci. 19, 1133e1140.
Zhou, F.M., Liang, Y., Dani, J.A., 2001. Endogenous nicotinic cholinergic activityregulates dopamine release in the striatum. Nat. Neurosci. 4, 1224e1229.
Zhou, F.M., Wilson, C.J., Dani, J.A., 2002. Cholinergic interneuron characteristics andnicotinic properties in the striatum. J. Neurobiol. 53, 590e605.























![Treadmill 93t.service[1]](https://img.pdfslide.us/doc/110x75/55cf9032550346703ba3c7b9/treadmill-93tservice1.jpg)





