Embed Size (px)
Citation preview

Neuritogenesis Induced by VasoactiveIntestinal Peptide, Pituitary AdenylateCyclase-Activating Polypeptide, and PeptideHistidine Methionine in SH-SY5Y Cells IsAssociated With Regulated Expression ofCytoskeleton mRNAs and Proteins
Celine Heraud,1 Sandrine Hilairet,2 Jean-Marc Muller,1 Jean-Francois Leterrier,1
and Corinne Chadeneau1*1Laboratoire des Biomembranes et Signalisation Cellulaire, Poitiers, France2Sanofi-SyntheLabo Recherche, Montpellier, France
Vasoactive intestinal peptide (VIP) and the related pep-tides pituitary adenylate cyclase-activating polypeptide(PACAP) and peptide histidine methionine (PHM) areknown to regulate proliferation and/or differentiation innormal and tumoral cells. In this study, neuritogenesis inhuman neuroblastoma SH-SY5Y cells cultured in serum-free medium was induced by VIP, PACAP, and PHM. Theestablishment of this process was followed by the quan-tification of neurite length and branching and the expres-sion of neurofilament mRNAs, neurofilament proteins,and other cytoskeletal protein markers of neuronal dif-ferentiation: neuron-specific MAPs and �-tubulin III. Neu-rite length and branching and the expression of mostmarkers tested were increased by VIP and PACAP in asimilar, although slightly different, fashion. In contrast,neuritic elongation induced by PHM was correlated withneither an increase in branching or neurofilament mRNAsnor a clear change in the expression of cytoskeletonproteins, with the exception of the stimulation by PHM ofdoublecortin, a microtubule-associated marker of mi-grating neuroblasts. These findings are the first evidencefrom a human neuron-like cell line for 1) a direct regula-tion of the metabolism of neurofilaments by VIP andPACAP and 2) the induction by PHM of neuritic pro-cesses of an apparent immature character.© 2003 Wiley-Liss, Inc.
Key words: VIP-related neuropeptides; SH-SY5Y; differ-entiation; neurofilaments; neuronal cytoskeleton
Vasoactive intestinal peptide (VIP), pituitary adenyl-ate cyclase-activating polypeptide (PACAP), and peptidehistidine isoleucine/methionine (PHI/PHM) belong tothe so-called VIP/secretin family of bioactive peptides.These neuropeptides are widely distributed in the body,particularly in the central and peripheral nervous systems.
VIP and PACAP modulate numerous functions and affectneural, circulatory, gastrointestinal, endocrine, and im-mune systems (Muller et al., 1995; Sherwood et al., 2000;Vaudry et al., 2000). They display neuroprotector andneurotrophic effects and induce neuronal survival (Gres-sens et al., 1993; Waschek, 1995; Gonzalez et al., 1997;Lindholm et al., 1998; Vaudry et al., 1999; DiCicco-Bloom et al., 2000). The functions of PHI/PHM are lesswell known. PHM modulates hormone secretion (Sasakiet al., 1987; Watanobe and Tamura 1994) and vesselsmooth muscle relaxation (Yao et al., 1996). In addition tothese functions, VIP, PACAP, and PHI/PHM are knownto regulate proliferation and/or differentiation in normaland tumoral cells. This regulation was notably investigatedin human and mouse neuroblastoma cells that were dem-onstrated to express high-affinity VIP, PACAP, or PHI/PHM binding sites (Muller et al., 1989, 1995; Vertongenet al., 1997; Waschek, 1997; Lelievre et al., 1998).
Neuroblastoma are embryonic tumors of neural crestorigin. These tumors are the most frequent extracranialsolid tumor of childhood. Some neuroblastoma undergo aspontaneous regression and/or differentiation into a be-nign form, the ganglioneuroma (Grosfeld, 2000). VIP andPACAP are frequently expressed in neuroblastoma, and anincreased VIP level is associated with the cell differentia-
Contract grant sponsor: Ligue Nationale Contre le Cancer.
*Correspondence to: Corinne Chadeneau, LBSC CNRS UMR 6558,Faculte des Sciences Fondamentales et Appliquees, 40 Avenue du RecteurPineau, 86022 Poitiers Cedex, France. E-mail: [email protected]
Received 10 May 2003; Revised 9 September 2003; Accepted 9 October2003
Journal of Neuroscience Research 75:320–329 (2004)
© 2003 Wiley-Liss, Inc.

tion grade of these tumors and a favorable prognosis(Qualman et al., 1995; Vertongen et al., 1997; Fruhwald etal., 1998; Grosfeld, 2000). VIP and/or PACAP induce thedecrease or arrest of cell proliferation in the human neu-roblastoma cell lines LA-N-5, SK-N-SH, IMR32, andNB-O-K1 (Pence and Shorter, 1990; O’Dorisio et al.,1992; Hoshino et al., 1993). Furthermore, in the NB-O-K1 cell line, VIP and PACAP27 induce neuritic out-growth (Hoshino et al., 1993). In contrast, in the humanneuroblastoma cell line NMB, VIP stimulates cell prolif-eration (Wollman et al., 2002). In the mouse neuroblas-toma cell line Neuro2A, VIP and PHI inhibit cell prolif-eration, whereas PACAP stimulates cell proliferation atsubnanomolar doses and inhibits cell proliferation athigher doses (Lelievre et al., 1998). Therefore, the effectsof neuropeptides on cellular proliferation or differentiationdepend on the type of NB cell lines, the neuropeptidedose, and perhaps the culture conditions, such as the serumlevel (Muller et al., 1995).
Distinct subclones are derived from the human neu-roblastoma cell line SK-N-SH. The SH-EP and SH-SY5Y subclones exhibit an epithelial and neuroblasticphenotype, respectively (Ross et al., 1983; Ross and Bie-dler, 1985; Biedler et al., 1985). SH-SY5Y cells are oftenused to study human neuronal differentiation and as amodel for reproducing and exploring the mechanisms ofneuronal diseases. In this study, we examined the effects ofVIP, PACAP, and PHM on these two cell lines culturedin serum-free medium. These neuropeptides inducedneurite-like process outgrowth only in SH-SY5Y cells.Because neuritogenesis associated with cell differentiationimplies up-regulation of specific cytoskeleton proteins, weexamined the expression of neurofilaments (NF), �-tubulinIII, and microtubule-associated proteins (MAPs) in SH-SY5Y cells treated or not with the neuropeptides.
MATERIALS AND METHODS
Cell Culture
SH-EP and SH-SY5Y cells were routinely seeded at den-sities of 2 and 3 � 106 cells/25-cm2 flasks, respectively, inhigh-glucose (4,500 mg/liter) Dulbecco’s modified Eagle’s me-dium (DMEM) with Glutamax I and sodium pyruvate (Invitro-gen, La Jolla, CA) supplemented with 10% fetal calf serum and100 U/ml penicillin and 100 �g/ml of streptomycin (Invitro-gen). Cells were incubated in a humidified 95% air/5% CO2controlled atmosphere at 37°C. Medium was changed every 3days. Passages were performed once per week, using 1�trypsin/EDTA (Invitrogen).
Cell Treatment
Cells were seeded and grown to subconfluence for 6 –7 days as described above. Serum-containing medium was thenreplaced with serum-free medium, and cells were cultured for6–7 days, with a renewal of serum-free medium on the fourthday. Before treatment, serum-free medium was changed forserum-free medium supplemented with 0.1% bovine serumalbumin (BSA) fraction V (Invitrogen). Neuropeptides wereprediluted to �1,000 or �100 in serum-free medium with 0.1%
BSA. Cells were treated with neuropeptides VIP, PACAP27, orPHM (Neosystem) by adding an appropriate volume of predi-luted neuropeptide to reach final concentrations ranging from10–10 M to 10–7 M. An equal volume of medium with 0.1%BSA without neuropeptide was added to control cells. Cellswere cultured for 1, 6, or 48 hr. For 48 hr of treatment, treatedand control cells received neuropeptides and medium with 0.1%BSA without neuropeptide, respectively, twice with a 24-hrinterval.
Measurement of Neurite Length and Branching
To measure neurite length and branching, video images ofcultured cells were captured using a digital camera connected toa phase-contrast microscope and to a computer (Photomat soft-ware; Microvision). For each treated or control cell culture,images of at least three fields were saved before treatment, and,to reduce variability, the same fields were captured after 48 hrwith or without peptide. Captured images were then analyzedwith VisioLab 2000 software (Biocom) for neurite length mea-surements. Only neurites as long as or longer than 14 �m, i.e.,the average length of cell bodies in control cells, were measured.For each field, the total length of neurites was measured anddivided by the number of emergences or connections of neuritesto or from the cell bodies. These measurement conditionsallowed application of the same rules for the different fields,even for those with complex networks of neurites betweenclusters of cells. However, this method led to an underestimatedaverage neurite length for networks in which branched neuriteslikely established several connections. Branching was measuredby counting the branch points, i.e., neurite bifurcations, man-ually on printed images of fields.
Western Immunoblotting
After treatments, cells were harvested with trypsin/EDTA, washed twice with cold phosphate-buffered saline(PBS), and suspended in 20 �l per 106 cells of ice-cold lysisbuffer [10 mM Tris, pH 7.5, 0.5 mM EDTA, pH 8.0, 0.5 mMdithiothreitol (DTT), 0.5% CHAPS, 10% glycerol, 0.1 mMphenylmethylsulfonyl fluoride (PMSF)]. After 30 min on ice,the samples were centrifuged at 4°C for 20 min at 10,000g. Thesupernatants were aliquoted, frozen in liquid nitrogen, andstored at –80°C. Protein concentration was determined by usingthe Bio-Rad DC Protein Assay (Bio-Rad, Hercules, CA). Pu-rified neurofilaments isolated from pig spinal cord preparedaccording to Leterrier et al. (1996) were used as positive controlsfor immunoanalysis. Proteins (20 �g) were resolved in 7.5%sodium dodecyl sulfate (SDS)-polyacrylamide gel as describedby Laemmli (1970) and electroblotted as described by Towbin etal. (1979) for 1 hr at 100 V onto Immobilon-P membranes(Millipore, Bedford, MA). Membranes were blocked overnightat room temperature by using 5% nonfat milk in Tris-bufferedsaline (TBS) containing 0.1% Tween 20 (TBST) and incubatedfor 2 hr at room temperature with primary antibodies in block-ing solution. The membranes were rinsed for 3 � 10 min atroom temperature in TBST, incubated for 1 hr with secondaryantibodies conjugated to horseradish peroxidase in 5% milk inTBST, and again rinsed with TBST. After a brief wash in TBS,bound antibodies were revealed by chemiluminescence with theECL� detection system (Amersham Biosciences, Arlington
VIP and Analogs in SH-SY5Y Human Cells 321

Heights, IL). The membranes were further stripped of boundantibodies as recommended in the manufacturer’s instructionsand saturated again (overnight with 5% milk in TBS) beforeincubation with other antibodies. After all immunodetections,total proteins on the membranes were detected by Coomassieblue staining. The ECL and Coomassie blue stainings werequantified by densitometry with the VisioLab 2000 image ana-lyzer (Biocom), with measures carried out independently by twoinvestigators. The densitometric measurements of ECL stainingwere normalized to the corresponding Coomassie blue stainingas loading control. The quantification procedure of the relativeamount of total proteins per lane was validated for membranestransferred with adjacent lanes loaded with 1� and 2� theamounts of total proteins of the same cell extract and stainedwith Coomassie blue after the blocking and stripping steps,reproducing the conditions used for immunoanalyses. Densito-metric measurements of each lane and of background on eachside of the lane were performed. The values obtained aftersubtraction of the background were in agreement with theexpected relative amount of total protein and showed a vari-ability not greater than 10%. Antibodies used were as follows:antiphosphorylated NFH (mouse RS18) 1:1,000 (Coleman andAnderton, 1990), antiphosphorylated NFM (rabbit serum 007)1:1,000 (Miehe et al., 1996), anti-NFL (rabbit NP70) 1:500(Hollenbeck and Bray, 1987), anti-MAP2 (rabbit serum 004)1:500 and antitau (rabbit serum 002) 1:500 (Leterrier and Eyer,1992), antidoublecortin (rabbit) 1:1,000 (Gleeson et al., 1999),monoclonal anti-�-tubulin III (mouse) 1:400 (Sigma, St. Louis,MO), anti-mouse IgG, H- and L-chain-specific (goat) peroxi-dase conjugate 1:20,000 (Calbiochem, La Jolla, CA), andperoxidase-conjugated goat anti-rabbit immunoglobulins (goat)1:2,000 (Dako, Carpinteria, CA).
cDNA Synthesis
After 1, 6, or 48 hr of treatment, total RNA was isolatedas described by Brown et al. (1999). Briefly, cells were lysed in400 �l per 106 cells of ice-cold lysis buffer [10 mM Tris-HCl,pH 7.5, 1 mM MgCl2, 0.15 mM NaCl, 0.5% NP40, and 20 URNaseOUT (Invitrogen)]. After 5 min on ice, samples werecentrifuged at 4°C for 30 min at 10,000g. The supernatant wasmixed to 50 �l 10% SDS and extracted with phenol/chloroform. RNA was precipitated with ethanol, resuspended inDNase/RNase-free water, and quantified using a GeneQuant IIspectrophotometer (Pharmacia, Piscataway, NJ). Aliquots oftotal RNA were treated with 1 U/1 �g RNA of DNase IAmplification Grade (Invitrogen) according to the manufactur-er’s instructions. After inactivation of DNase, RNA was reversetranscribed using an anchored oligo(dT)23 (Sigma) and M-MLVReverse Transcriptase RNase H Minus (Promega, Madison,WI) according to the manufacturer’s instructions. Because theprimer pairs used for the expression of NF mRNAs were able toamplify mRNA and DNA, DNase-treated RNA was subjectedto PCR amplification to ensure the absence of genomic DNA.
Real-Time PCR and Quantification
Real-time PCR was carried out with the LightCyclerSystem (Roche Molecular Biochemicals) by using theLightCycler-FastStart DNA Master SYBR Green I Kit accord-ing to the manufacturer’s instructions, except that the reaction
volume was 10 �l. Aliquots of cDNA (40 or 4 ng) and knownamounts of external standards (purified PCR products quanti-fied using a GeneQuant II spectrophotometer and diluted inDNase and Reverse Transcription mixes; 102–106 molecules)were amplified in parallel reactions. The optimal cycle programfor all amplified cDNA was determined to obtain a single,specific PCR product, verified by melting-curves analysis, fol-lowed by gel electrophoresis and DNA sequencing. These de-terminations showed that the same program could be optimalfor all amplified cDNA: 5 sec at 94°C, 5 sec at 60°C, and 10 secat 72°C. Quantification was performed by real-time detection offluorimetric emission of SYBR green I and by comparison ofthe fluorescence of PCR products of unknown concentrationwith the fluorescence of the external standards. Fluorescencevalues were analyzed with the LightCycler data-analysis software(version 3.5.3) by using the second derivative maximummethod. For each single determination, the PCRs were runtwice in duplicate. Initially, actin and GAPDH were used ascontrols of mRNA level. Because the actin/GAPDH mRNAratios between the untreated and the treated cells showed lessthan 10% variability, only GAPDH mRNA level was usedfurther to normalize the mRNA levels between samples. Prim-ers were designed with Primer3 software (http://www.genome.wi.mit.edu/cgi-bin/primer/primer3_www.cgi), except for ac-tin and GAPDH forward and reverse (Mayer et al., 2002) andwere as follows (5�-3�): NFH forward TGTCGAAAAGC-CCAAAGAAT, NFH reverse TTCTTGGCCACCTCTGT-CTT, NFM forward AAGGGATCCAGGAAGGAAGA, NFMreverse CTGCTGGGCTCAAGTCTAGG, NFL forward TA-CACCAGCCATGTCCAAGA, NFL reverse TCTTCAGCT-GCCTCCTCTTC.
RESULTS
VIP, PACAP, and PHM Induce MorphologicalChanges in SH-SY5Y Cells
The effects of VIP, PACAP, and PHM on celldifferentiation were studied in the two cloned cell linesSH-SY5Y and SH-EP, derived from the human neuro-blastoma SK-N-SH cell line. SH-EP cells exhibit a non-neuroblastic phenotype, whereas the SH-SY5Y cell linehas a neuroblastic phenotype. In routine culture, small-cell-body SH-SY5Y cells grew as weakly adherent clustersextending relatively short neurite-like processes. In con-trast, the epithelial-like SH-EP cells grew as a monolayerand had a highly adherent flat shape without producingprocesses. To study the effects of neuropeptides in a de-fined medium, treatments with neuropeptides were per-formed on serum-starved cells for 6–7 days. The removalof serum led to the detachment of a fraction of cells,mainly in the neuroblastic cell line. However, numerouscells remained adherent. Only SH-SY5Y cells underwenta process outgrowth, a well-known effect of serum deple-tion on neuroblastoma cell lines (Gurwitz and Cunning-ham, 1988; Tsuneishi et al., 1993).
Serum-starved cells were treated or not with VIP,PACAP, or PHM as described in Materials and Methods atfinal concentrations ranging from 10–10 M to 10–7 M.After different periods of time, cells were examined for
322 Heraud et al.
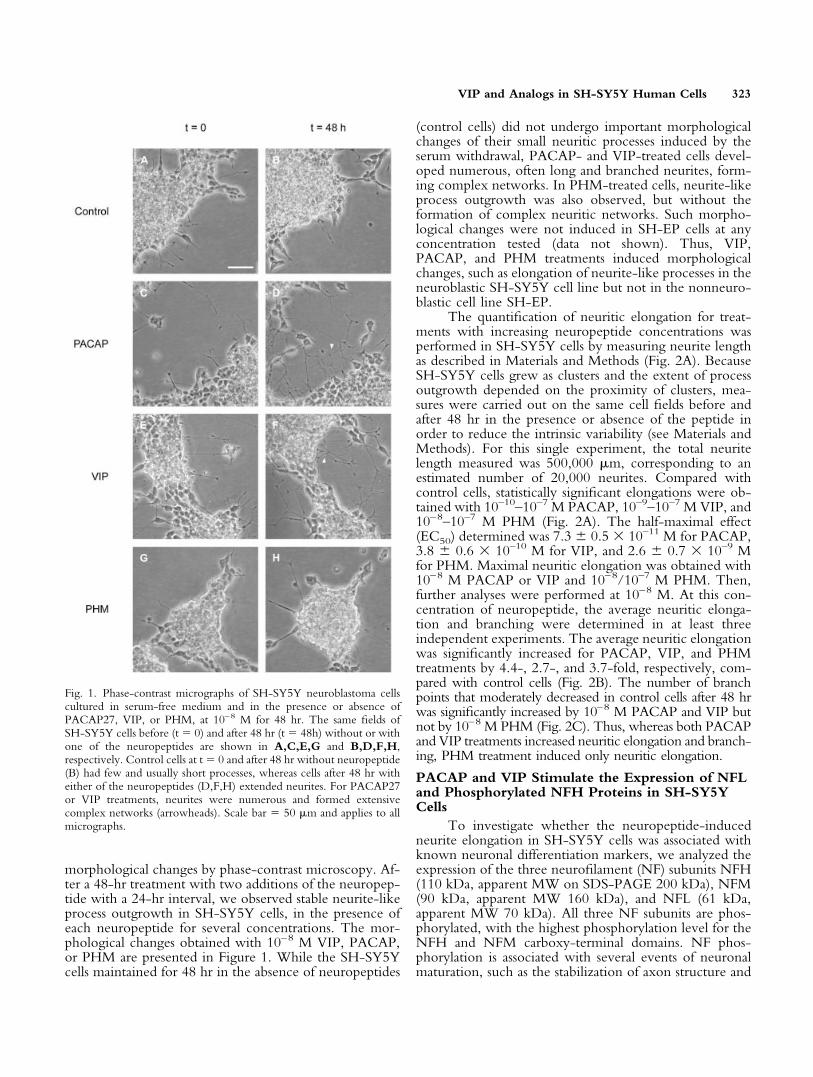
morphological changes by phase-contrast microscopy. Af-ter a 48-hr treatment with two additions of the neuropep-tide with a 24-hr interval, we observed stable neurite-likeprocess outgrowth in SH-SY5Y cells, in the presence ofeach neuropeptide for several concentrations. The mor-phological changes obtained with 10–8 M VIP, PACAP,or PHM are presented in Figure 1. While the SH-SY5Ycells maintained for 48 hr in the absence of neuropeptides
(control cells) did not undergo important morphologicalchanges of their small neuritic processes induced by theserum withdrawal, PACAP- and VIP-treated cells devel-oped numerous, often long and branched neurites, form-ing complex networks. In PHM-treated cells, neurite-likeprocess outgrowth was also observed, but without theformation of complex neuritic networks. Such morpho-logical changes were not induced in SH-EP cells at anyconcentration tested (data not shown). Thus, VIP,PACAP, and PHM treatments induced morphologicalchanges, such as elongation of neurite-like processes in theneuroblastic SH-SY5Y cell line but not in the nonneuro-blastic cell line SH-EP.
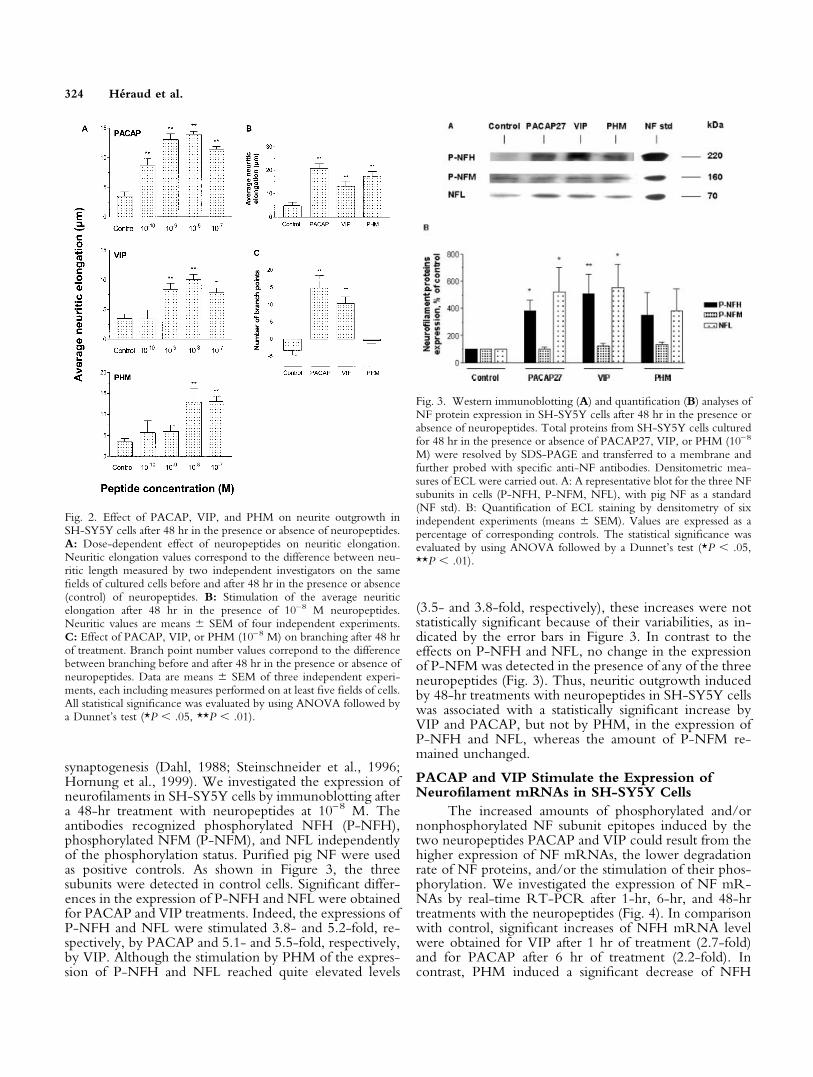
The quantification of neuritic elongation for treat-ments with increasing neuropeptide concentrations wasperformed in SH-SY5Y cells by measuring neurite lengthas described in Materials and Methods (Fig. 2A). BecauseSH-SY5Y cells grew as clusters and the extent of processoutgrowth depended on the proximity of clusters, mea-sures were carried out on the same cell fields before andafter 48 hr in the presence or absence of the peptide inorder to reduce the intrinsic variability (see Materials andMethods). For this single experiment, the total neuritelength measured was 500,000 �m, corresponding to anestimated number of 20,000 neurites. Compared withcontrol cells, statistically significant elongations were ob-tained with 10–10–10–7 M PACAP, 10–9–10–7 M VIP, and10–8–10–7 M PHM (Fig. 2A). The half-maximal effect(EC50) determined was 7.3 � 0.5 � 10–11 M for PACAP,3.8 � 0.6 � 10–10 M for VIP, and 2.6 � 0.7 � 10–9 Mfor PHM. Maximal neuritic elongation was obtained with10–8 M PACAP or VIP and 10–8/10–7 M PHM. Then,further analyses were performed at 10–8 M. At this con-centration of neuropeptide, the average neuritic elonga-tion and branching were determined in at least threeindependent experiments. The average neuritic elongationwas significantly increased for PACAP, VIP, and PHMtreatments by 4.4-, 2.7-, and 3.7-fold, respectively, com-pared with control cells (Fig. 2B). The number of branchpoints that moderately decreased in control cells after 48 hrwas significantly increased by 10–8 M PACAP and VIP butnot by 10–8 M PHM (Fig. 2C). Thus, whereas both PACAPand VIP treatments increased neuritic elongation and branch-ing, PHM treatment induced only neuritic elongation.
PACAP and VIP Stimulate the Expression of NFLand Phosphorylated NFH Proteins in SH-SY5YCells
To investigate whether the neuropeptide-inducedneurite elongation in SH-SY5Y cells was associated withknown neuronal differentiation markers, we analyzed theexpression of the three neurofilament (NF) subunits NFH(110 kDa, apparent MW on SDS-PAGE 200 kDa), NFM(90 kDa, apparent MW 160 kDa), and NFL (61 kDa,apparent MW 70 kDa). All three NF subunits are phos-phorylated, with the highest phosphorylation level for theNFH and NFM carboxy-terminal domains. NF phos-phorylation is associated with several events of neuronalmaturation, such as the stabilization of axon structure and
Fig. 1. Phase-contrast micrographs of SH-SY5Y neuroblastoma cellscultured in serum-free medium and in the presence or absence ofPACAP27, VIP, or PHM, at 10–8 M for 48 hr. The same fields ofSH-SY5Y cells before (t � 0) and after 48 hr (t � 48h) without or withone of the neuropeptides are shown in A,C,E,G and B,D,F,H,respectively. Control cells at t � 0 and after 48 hr without neuropeptide(B) had few and usually short processes, whereas cells after 48 hr witheither of the neuropeptides (D,F,H) extended neurites. For PACAP27or VIP treatments, neurites were numerous and formed extensivecomplex networks (arrowheads). Scale bar � 50 �m and applies to allmicrographs.
VIP and Analogs in SH-SY5Y Human Cells 323
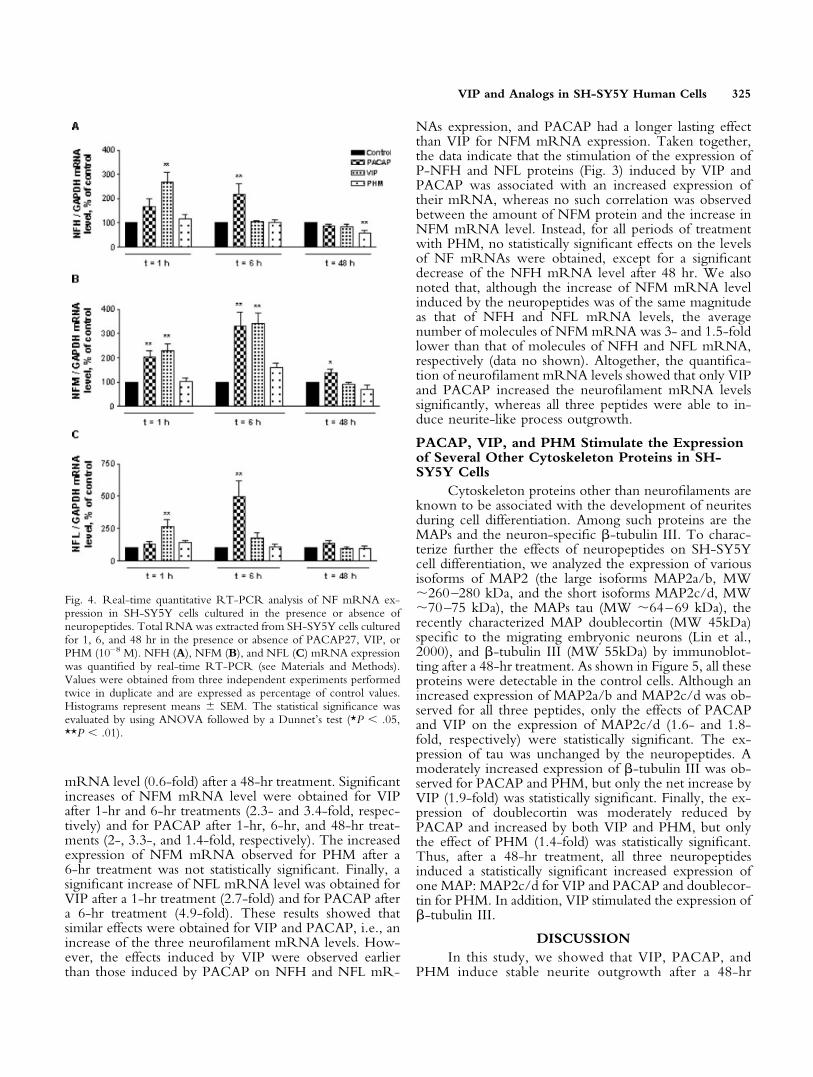
synaptogenesis (Dahl, 1988; Steinschneider et al., 1996;Hornung et al., 1999). We investigated the expression ofneurofilaments in SH-SY5Y cells by immunoblotting aftera 48-hr treatment with neuropeptides at 10–8 M. Theantibodies recognized phosphorylated NFH (P-NFH),phosphorylated NFM (P-NFM), and NFL independentlyof the phosphorylation status. Purified pig NF were usedas positive controls. As shown in Figure 3, the threesubunits were detected in control cells. Significant differ-ences in the expression of P-NFH and NFL were obtainedfor PACAP and VIP treatments. Indeed, the expressions ofP-NFH and NFL were stimulated 3.8- and 5.2-fold, re-spectively, by PACAP and 5.1- and 5.5-fold, respectively,by VIP. Although the stimulation by PHM of the expres-sion of P-NFH and NFL reached quite elevated levels
(3.5- and 3.8-fold, respectively), these increases were notstatistically significant because of their variabilities, as in-dicated by the error bars in Figure 3. In contrast to theeffects on P-NFH and NFL, no change in the expressionof P-NFM was detected in the presence of any of the threeneuropeptides (Fig. 3). Thus, neuritic outgrowth inducedby 48-hr treatments with neuropeptides in SH-SY5Y cellswas associated with a statistically significant increase byVIP and PACAP, but not by PHM, in the expression ofP-NFH and NFL, whereas the amount of P-NFM re-mained unchanged.
PACAP and VIP Stimulate the Expression ofNeurofilament mRNAs in SH-SY5Y Cells
The increased amounts of phosphorylated and/ornonphosphorylated NF subunit epitopes induced by thetwo neuropeptides PACAP and VIP could result from thehigher expression of NF mRNAs, the lower degradationrate of NF proteins, and/or the stimulation of their phos-phorylation. We investigated the expression of NF mR-NAs by real-time RT-PCR after 1-hr, 6-hr, and 48-hrtreatments with the neuropeptides (Fig. 4). In comparisonwith control, significant increases of NFH mRNA levelwere obtained for VIP after 1 hr of treatment (2.7-fold)and for PACAP after 6 hr of treatment (2.2-fold). Incontrast, PHM induced a significant decrease of NFH
Fig. 2. Effect of PACAP, VIP, and PHM on neurite outgrowth inSH-SY5Y cells after 48 hr in the presence or absence of neuropeptides.A: Dose-dependent effect of neuropeptides on neuritic elongation.Neuritic elongation values correspond to the difference between neu-ritic length measured by two independent investigators on the samefields of cultured cells before and after 48 hr in the presence or absence(control) of neuropeptides. B: Stimulation of the average neuriticelongation after 48 hr in the presence of 10–8 M neuropeptides.Neuritic values are means � SEM of four independent experiments.C: Effect of PACAP, VIP, or PHM (10–8 M) on branching after 48 hrof treatment. Branch point number values correpond to the differencebetween branching before and after 48 hr in the presence or absence ofneuropeptides. Data are means � SEM of three independent experi-ments, each including measures performed on at least five fields of cells.All statistical significance was evaluated by using ANOVA followed bya Dunnet’s test (*P � .05, **P � .01).
Fig. 3. Western immunoblotting (A) and quantification (B) analyses ofNF protein expression in SH-SY5Y cells after 48 hr in the presence orabsence of neuropeptides. Total proteins from SH-SY5Y cells culturedfor 48 hr in the presence or absence of PACAP27, VIP, or PHM (10–8
M) were resolved by SDS-PAGE and transferred to a membrane andfurther probed with specific anti-NF antibodies. Densitometric mea-sures of ECL were carried out. A: A representative blot for the three NFsubunits in cells (P-NFH, P-NFM, NFL), with pig NF as a standard(NF std). B: Quantification of ECL staining by densitometry of sixindependent experiments (means � SEM). Values are expressed as apercentage of corresponding controls. The statistical significance wasevaluated by using ANOVA followed by a Dunnet’s test (*P � .05,**P � .01).
324 Heraud et al.
mRNA level (0.6-fold) after a 48-hr treatment. Significantincreases of NFM mRNA level were obtained for VIPafter 1-hr and 6-hr treatments (2.3- and 3.4-fold, respec-tively) and for PACAP after 1-hr, 6-hr, and 48-hr treat-ments (2-, 3.3-, and 1.4-fold, respectively). The increasedexpression of NFM mRNA observed for PHM after a6-hr treatment was not statistically significant. Finally, asignificant increase of NFL mRNA level was obtained forVIP after a 1-hr treatment (2.7-fold) and for PACAP aftera 6-hr treatment (4.9-fold). These results showed thatsimilar effects were obtained for VIP and PACAP, i.e., anincrease of the three neurofilament mRNA levels. How-ever, the effects induced by VIP were observed earlierthan those induced by PACAP on NFH and NFL mR-
NAs expression, and PACAP had a longer lasting effectthan VIP for NFM mRNA expression. Taken together,the data indicate that the stimulation of the expression ofP-NFH and NFL proteins (Fig. 3) induced by VIP andPACAP was associated with an increased expression oftheir mRNA, whereas no such correlation was observedbetween the amount of NFM protein and the increase inNFM mRNA level. Instead, for all periods of treatmentwith PHM, no statistically significant effects on the levelsof NF mRNAs were obtained, except for a significantdecrease of the NFH mRNA level after 48 hr. We alsonoted that, although the increase of NFM mRNA levelinduced by the neuropeptides was of the same magnitudeas that of NFH and NFL mRNA levels, the averagenumber of molecules of NFM mRNA was 3- and 1.5-foldlower than that of molecules of NFH and NFL mRNA,respectively (data no shown). Altogether, the quantifica-tion of neurofilament mRNA levels showed that only VIPand PACAP increased the neurofilament mRNA levelssignificantly, whereas all three peptides were able to in-duce neurite-like process outgrowth.
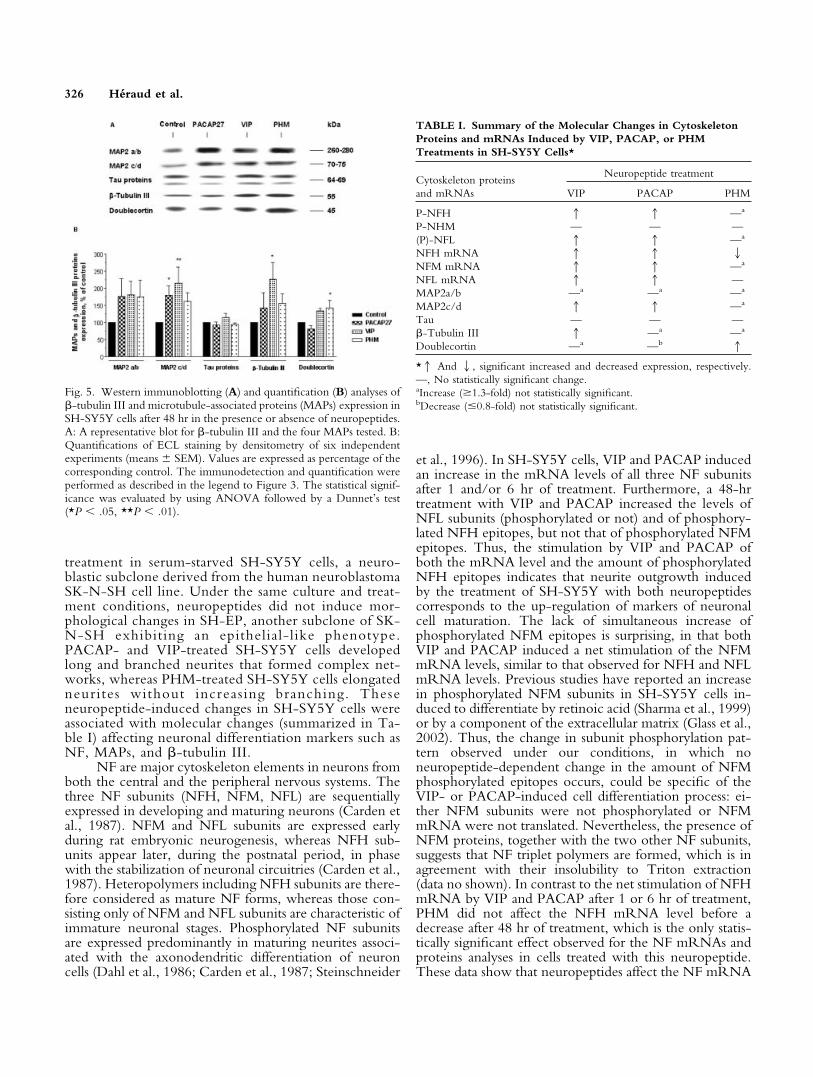
PACAP, VIP, and PHM Stimulate the Expressionof Several Other Cytoskeleton Proteins in SH-SY5Y Cells
Cytoskeleton proteins other than neurofilaments areknown to be associated with the development of neuritesduring cell differentiation. Among such proteins are theMAPs and the neuron-specific �-tubulin III. To charac-terize further the effects of neuropeptides on SH-SY5Ycell differentiation, we analyzed the expression of variousisoforms of MAP2 (the large isoforms MAP2a/b, MW260–280 kDa, and the short isoforms MAP2c/d, MW70–75 kDa), the MAPs tau (MW 64–69 kDa), therecently characterized MAP doublecortin (MW 45kDa)specific to the migrating embryonic neurons (Lin et al.,2000), and �-tubulin III (MW 55kDa) by immunoblot-ting after a 48-hr treatment. As shown in Figure 5, all theseproteins were detectable in the control cells. Although anincreased expression of MAP2a/b and MAP2c/d was ob-served for all three peptides, only the effects of PACAPand VIP on the expression of MAP2c/d (1.6- and 1.8-fold, respectively) were statistically significant. The ex-pression of tau was unchanged by the neuropeptides. Amoderately increased expression of �-tubulin III was ob-served for PACAP and PHM, but only the net increase byVIP (1.9-fold) was statistically significant. Finally, the ex-pression of doublecortin was moderately reduced byPACAP and increased by both VIP and PHM, but onlythe effect of PHM (1.4-fold) was statistically significant.Thus, after a 48-hr treatment, all three neuropeptidesinduced a statistically significant increased expression ofone MAP: MAP2c/d for VIP and PACAP and doublecor-tin for PHM. In addition, VIP stimulated the expression of�-tubulin III.
DISCUSSIONIn this study, we showed that VIP, PACAP, and
PHM induce stable neurite outgrowth after a 48-hr
Fig. 4. Real-time quantitative RT-PCR analysis of NF mRNA ex-pression in SH-SY5Y cells cultured in the presence or absence ofneuropeptides. Total RNA was extracted from SH-SY5Y cells culturedfor 1, 6, and 48 hr in the presence or absence of PACAP27, VIP, orPHM (10–8 M). NFH (A), NFM (B), and NFL (C) mRNA expressionwas quantified by real-time RT-PCR (see Materials and Methods).Values were obtained from three independent experiments performedtwice in duplicate and are expressed as percentage of control values.Histograms represent means � SEM. The statistical significance wasevaluated by using ANOVA followed by a Dunnet’s test (*P � .05,**P � .01).
VIP and Analogs in SH-SY5Y Human Cells 325
treatment in serum-starved SH-SY5Y cells, a neuro-blastic subclone derived from the human neuroblastomaSK-N-SH cell line. Under the same culture and treat-ment conditions, neuropeptides did not induce mor-phological changes in SH-EP, another subclone of SK-N-SH exhibiting an epithelial-like phenotype.PACAP- and VIP-treated SH-SY5Y cells developedlong and branched neurites that formed complex net-works, whereas PHM-treated SH-SY5Y cells elongatedneurites without increasing branching. Theseneuropeptide-induced changes in SH-SY5Y cells wereassociated with molecular changes (summarized in Ta-ble I) affecting neuronal differentiation markers such asNF, MAPs, and �-tubulin III.
NF are major cytoskeleton elements in neurons fromboth the central and the peripheral nervous systems. Thethree NF subunits (NFH, NFM, NFL) are sequentiallyexpressed in developing and maturing neurons (Carden etal., 1987). NFM and NFL subunits are expressed earlyduring rat embryonic neurogenesis, whereas NFH sub-units appear later, during the postnatal period, in phasewith the stabilization of neuronal circuitries (Carden et al.,1987). Heteropolymers including NFH subunits are there-fore considered as mature NF forms, whereas those con-sisting only of NFM and NFL subunits are characteristic ofimmature neuronal stages. Phosphorylated NF subunitsare expressed predominantly in maturing neurites associ-ated with the axonodendritic differentiation of neuroncells (Dahl et al., 1986; Carden et al., 1987; Steinschneider
et al., 1996). In SH-SY5Y cells, VIP and PACAP inducedan increase in the mRNA levels of all three NF subunitsafter 1 and/or 6 hr of treatment. Furthermore, a 48-hrtreatment with VIP and PACAP increased the levels ofNFL subunits (phosphorylated or not) and of phosphory-lated NFH epitopes, but not that of phosphorylated NFMepitopes. Thus, the stimulation by VIP and PACAP ofboth the mRNA level and the amount of phosphorylatedNFH epitopes indicates that neurite outgrowth inducedby the treatment of SH-SY5Y with both neuropeptidescorresponds to the up-regulation of markers of neuronalcell maturation. The lack of simultaneous increase ofphosphorylated NFM epitopes is surprising, in that bothVIP and PACAP induced a net stimulation of the NFMmRNA levels, similar to that observed for NFH and NFLmRNA levels. Previous studies have reported an increasein phosphorylated NFM subunits in SH-SY5Y cells in-duced to differentiate by retinoic acid (Sharma et al., 1999)or by a component of the extracellular matrix (Glass et al.,2002). Thus, the change in subunit phosphorylation pat-tern observed under our conditions, in which noneuropeptide-dependent change in the amount of NFMphosphorylated epitopes occurs, could be specific of theVIP- or PACAP-induced cell differentiation process: ei-ther NFM subunits were not phosphorylated or NFMmRNA were not translated. Nevertheless, the presence ofNFM proteins, together with the two other NF subunits,suggests that NF triplet polymers are formed, which is inagreement with their insolubility to Triton extraction(data no shown). In contrast to the net stimulation of NFHmRNA by VIP and PACAP after 1 or 6 hr of treatment,PHM did not affect the NFH mRNA level before adecrease after 48 hr of treatment, which is the only statis-tically significant effect observed for the NF mRNAs andproteins analyses in cells treated with this neuropeptide.These data show that neuropeptides affect the NF mRNA
Fig. 5. Western immunoblotting (A) and quantification (B) analyses of�-tubulin III and microtubule-associated proteins (MAPs) expression inSH-SY5Y cells after 48 hr in the presence or absence of neuropeptides.A: A representative blot for �-tubulin III and the four MAPs tested. B:Quantifications of ECL staining by densitometry of six independentexperiments (means � SEM). Values are expressed as percentage of thecorresponding control. The immunodetection and quantification wereperformed as described in the legend to Figure 3. The statistical signif-icance was evaluated by using ANOVA followed by a Dunnet’s test(*P � .05, **P � .01).
TABLE I. Summary of the Molecular Changes in CytoskeletonProteins and mRNAs Induced by VIP, PACAP, or PHMTreatments in SH-SY5Y Cells*
Cytoskeleton proteinsand mRNAs
Neuropeptide treatment
VIP PACAP PHM
P-NFH 1 1 —a
P-NHM — — —(P)-NFL 1 1 —a
NFH mRNA 1 1 2NFM mRNA 1 1 —a
NFL mRNA 1 1 —MAP2a/b —a —a —a
MAP2c/d 1 1 —a
Tau — — —�-Tubulin III 1 —a —a
Doublecortin —a —b 1
*1 And 2, significant increased and decreased expression, respectively.—, No statistically significant change.aIncrease (�1.3-fold) not statistically significant.bDecrease (�0.8-fold) not statistically significant.
326 Heraud et al.

levels and the NF protein levels in distinct manners andwith net differences in effects among the three neuropep-tides tested. Indeed, VIP and PACAP stimulate similarly atransient expression of all three NF mRNAs, in contrast toPHM, but with slightly different time courses betweenVIP and PACAP (Fig. 4). Furthermore, the increasedexpression of the two NFH and NFM mRNAs by VIPand PACAP and the increased phosphorylated epitopeslevel only for NFH show that these two neuropeptidesregulate independently the synthesis or life span of NFmRNAs and the phosphorylation or life span of NFsubunits. A related phenomenon was observed in murineneocortex, where prenatal blockade of VIP induced anincreased expression of NF 160 kDa, whereas the expres-sion of NFH and NFL was unaffected (Zupan et al., 2000).Taken together, the present data suggest that the stimula-tion of neuritogenesis in SH-SY5Y cells by two of theneuropeptides tested (VIP and PACAP), correlated with atransient increase in NF mRNAs and a longer lastingincrease in expressed NF proteins, reflects an early step ofgenuine neuritic differentiation. The third neuropeptidetested, PHM, likely induces NF-poor neurites that do notrequire a stimulation of the metabolism of NF as importantas that observed for VIP and PACAP.
MAPs and �-tubulin III are other markers of neu-ronal cell differentiation. Shea et al. (1992) have shownthat, in neuroblastoma NB2a/d1 cells, the preexistingcytoplasmic pools of the axon-specific tau proteins canmediate initial neuritogenesis. In our study, we found thattau isoforms were expressed but not affected by any neu-ropeptide treatment in SH-SY5Y cells. Preexisting poolsof tau proteins seem sufficient to allow the neurite elon-gation observed. The superfamily of MAP2 proteins playsan important role in the development of neuronal cells.Indeed, MAP2c is expressed in early developmental stages,whereas MAP2a is expressed mostly in the adult brain.MAP2b and MAP2d are present throughout nervous sys-tem development (Sanchez et al., 2000). MAP2a/b wereexpressed and only a nonstatistically significant increasewas observed in SH-SY5Y cells following neuropeptidetreatments. In contrast, a statistically significant increase inMAP2c/d proteins was induced by VIP and PACAP aftera 48-hr treatment. Therefore, VIP and PACAP inducedthe expression of MAP2 isoforms characteristic of earlydevelopmental stages. Doublecortin, a novel MAP, is animmature neuronal marker highly expressed in the distalparts of developing processes (Francis et al., 1999; Gleesonet al., 1999). Only PHM induced a statistically significantincrease of the expression of doublecortin. This increase,associated with the decrease of NFH mRNA level, whichis not expressed during early development, indicates thatPHM-treated cells were at a very early differentiationstage. The �-tubulin III isoform is a neuronal-specifictubulin expressed early in neuronal differentiation. Its ex-pression is differentiation dependent in rat peripheral neu-rons (Jiang and Oblinger, 1992) and human neuroblastictumours such as neuroblastoma (Katsetos et al., 1994).Only VIP induced a clear increased expression of
�-tubulin III, suggesting that VIP induces a more maturestate of differentiation than PACAP and PHM. Takentogether, analyses of cytoskeleton proteins indicate that,after a 48-hr treatment, 1) PHM, for which the onlystatistically significant increase in cytoskeleton proteinsaffected doublecortin, a marker of migrating neuroblasts,induces a very early differentiation state; 2) VIP andPACAP, which increase phosphorylated NFH character-istic of mature NF predominantly in axons and expressionof MAP2c/d associated with development, induce a moreadvanced differentiation state; and 3) VIP, which increasesa differentiation-dependent marker, such as �-tubulin III,induces a more mature state than PACAP.
Increased but not statiscally significant expressionswere obtained for several markers, mainly with PHM(Table I), after a 48-hr treatment. Preliminary experimentsperformed after longer treatments seem to indicate that theexpression of NF continues to increase in PHM-treatedcells. Therefore, such treatments could allow a determi-nation of whether the nonstatistically significant increasesobserved for several markers after a 48-hr treatment cor-respond to effects requiring longer than 48 hr to be fullyestablished.
Three known receptors mediate most effects of theseneuropeptides in various tissues. PAC1 is the PACAP-specific receptor, whereas VPAC1 and VPAC2 are poly-valent receptors for VIP and PACAP, which also bindPHI/PHM but with a lower affinity (Harmar et al., 1998).This study emphasizes common effects of PACAP andVIP on SH-SY5Y cell differentiation, suggesting thatthese neuropeptides could act via a polyvalent receptor.However, the dose-dependent effect of neuropeptides onneuritic elongation indicated that the relative potencies(EC50 PACAP/EC50 peptide) of PACAP, VIP, and PHMwere 1, 0.2, and 0.03, respectively. This potency orderdoes not clearly correspond to that observed at knownreceptors. Selective antagonists and agonists could allowidentification of the receptor(s) involved in theneuropeptide-induced neurite outgrowth in SH-SY5Ycells.
In this study, we have shown that VIP, PACAP, andPHM induce a neuroblastic cell differentiation in serum-starved SH-SY5Y cells after a 48-hr treatment, as shownby the early stage of neurite outgrowth indicated by thechanges in neuronal markers. VIP increased the expressionof more markers than PACAP, which itself stimulatedmore markers than PHM. The present differentiationmodel using VIP, PACAP, and PHM treatments of hu-man SH-SY5Y cells could allow the identification oftranscriptional factors implicated in the early differentia-tion stages. To our knowledge, this report is the firstshowing an effect of PHM on neurite outgrowth and celldifferentiation, and the direct stimulation of the NF me-tabolism by the neuropeptides VIP and PACAP, in ahuman cell line.
ACKNOWLEDGMENTSWe are very grateful to B.H. Anderton (Institute of
Psychiatry, London) for the kind gift of the monoclonal
VIP and Analogs in SH-SY5Y Human Cells 327

antibody RS18 and to J.G. Gleeson (Department of Neu-rosciences, University of California, San Diego) for thepolyclonal antibody against doublecortin. We thank D.Guyonnet for his technical assistance with DNA sequenc-ing. C.H. was the recipient of PhD fellowships from theLigue Nationale Contre le Cancer and from the Fondationpour la Recherche Medicale.
REFERENCESBiedler JL, Rozen MG, el-Badry O, Meyers MB, Melera PW, Ross RA,
Spengler BA. 1985. Growth stage-related synthesis and secretion ofproteins by human neuroblastoma cells and their variants. Prog Clin BiolRes 175:202–221.
Brown AJ, Hutchings C, Burke JF, Mayne LV. 1999. Application of a rapidmethod (targeted display) for the identification of differentially expressedmRNAs following NGF-induced neuronal differentiation in PC12 cells.Mol Cell Neurosci 13:119–130.
Carden MJ, Trojanowski JQ, Schlaepfer WW, Lee VM. 1987. Two-stageexpression of neurofilament polypeptides during rat neurogenesis withearly establishment of adult phosphorylation patterns. J Neurosci 7:3489–3504.
Coleman MP, Anderton BH. 1990. Phosphate-dependent monoclonalantibodies to neurofilaments and Alzheimer neurofibrillary tangles recog-nize a synthetic phosphopeptide. J Neurochem 54:1548–1555.
Dahl D. 1988. Early and late appearance of neurofilament phosphorylatedepitopes in rat nervous system development: in vivo and in vitro studywith monoclonal antibodies. J Neurosci Res 20:431–441.
Dahl D, Bignami A. 1986. Neurofilament phosphorylation in development.A sign of axonal maturation? Exp Cell Res 162:220–230.
DiCicco-Bloom E, Deutsch PJ, Maltzman J, Zhang J, Pintar JE, Zheng J,Friedman WF, Zhou X, Zaremba T. 2000. Autocrine expression andontogenic functions of the PACAP ligand/receptor system during sym-pathetic development. Dev Biol 219:197–213.
Francis F, Koulakoff A, Boucher D, Chafey P, Schaar B, Vinet MC,Friocourt G, McDonnell N, Reiner O, Kahn A, McConnell SK,Berwald-Netter, Denoulet P, Chelly J. 1999. Doublecortin is a develop-mentally regulated, microtubule-associated protein expressed in migratingand differentiating neurons. Neuron 23:247–256.
Fruhwald MC, O’Dorisio MS, Cottingham SL, Qualman SJ, O’DorisioTM. 1998. Neuropeptides in developmental tumors of the central andperipheral nervous system. Ann N Y Acad Sci 865:420–426.
Glass TL, Raabe TD, Garcia DM, Koke JR. 2002. Phosphorylated neu-rofilaments and SNAP-25 in cultured SH-SY5Y neuroblastoma cells.Brain Res 26:43–48.
Gleeson JG, Lin PT, Flanagan LA, Walsh CA. 1999. Doublecortin is amicrotubule-associated protein and is expressed widely by migratingneurons. Neuron 23:257–271.
Gonzalez BJ, Basille M, Vaudry D, Fournier A, Vaudry H. 1997. Pituitaryadenylate cyclase-activating polypeptide promotes cell survival and neu-rite outgrowth in rat cerebellar neuroblasts. Neuroscience 78:419–430.
Gressens P, Hill JM, Gozes I, Fridkin M, Brenneman DE. 1993. Growthfactor function of vasoactive intestinal peptide in whole cultured mouseembryos. Nature 362:155–158.
Grosfeld JL. 2000. Risk-based management of solid tumors in children.Am J Surg 180:322–325.
Gurwitz D, Cunningham DD. 1988. Thrombin modulates and reversesneuroblastoma neurite outgrowth. Proc Natl Acad Sci USA 85:3440–3444.
Harmar AJ, Arimura A, Gozes I, Journot L, Laburthe M, Pisegna JR,Rawlings SR, Robberecht P, Said SI, Sreedharan SP, Wank SA, WaschekJA. 1998. International Union of Pharmacology. XVIII. Nomenclature ofreceptors for vasoactive intestinal peptide and pituitary adenylate cyclase-activating polypeptide. Pharmacol Rev 50:265–270.
Hollenbeck PJ, Bray D. 1987. Rapidly transported organelles containing
membrane and cytoskeletal components: their relation to axonal growth.J Cell Biol 105:2827–2835.
Hornung JP, Riederer BM. 1999. Medium-sized neurofilament proteinrelated to maturation of a subset of cortical neurons. J Comp Neurol414:348–360.
Hoshino M, Li M, Zheng LQ, Suzuki M, Mochizuki T, Yanaihara N.1993. Pituitary adenylate cyclase activating peptide and vasoactive intes-tinal polypeptide: differentiation effects on human neuroblastoma NB-O-K1 cells. Neurosci Lett 159:35–38.
Jiang YQ, Oblinger MM. 1992. Differential regulation of betaIII and othertubulin genes during peripheral and central neuron development. J CellSci 103:643–651.
Katsetos CD, Karkavelas G, Frankfurter A, Vlachos IN, Vogeley K,Schober R, Wechsler W, Urich H. 1994. The stromal Schwann cellduring maturation of peripheral neuroblastomas: immunohistochemicalobservations with antibodies to the neuronal class III beta-tubulin isotype(betaIII) and S-100 protein. Clin Neuropathol 13:171–180.
Laemmli UK. 1970. Cleavage of structural proteins during the assembly ofthe head of bacteriophage T4. Nature 227:680–685.
Lelievre V, Pineau N, Du J, Wen CH, Nguyen T, Janet T, Muller JM,Waschek JA. 1998. Differential effects of peptide histidine isoleucine(PHI) and related peptides on stimulation and suppression of neuroblas-toma cell proliferation. J Biol Chem 273:19685–19690.
Leterrier JF, Eyer J. 1992. Age-dependent changes in the ultrastructure andin the molecular composition of rat brain microtubules. J Neurochem59:1126–1137.
Leterrier JF, Kas J, Hartwig J, Vegners R, Janmey PA. 1996. Mechanicaleffects of neurofilament cross-bridges. Modulation by phosphorylation,lipids and interaction with F-actin. J Biol Chem 271:15687–15694.
Lin PT, Gleeson JG, Corbo JC, Flanagan L, Walsh CA. 2000. DCAMKL1encodes a protein kinase with homology to doublecortin that regulatesmicrotubule polymerization. J Neurosci 20:9152–9161.
Lindholm D, Skoglosa Y, Takei N. 1998. Developmental regulation ofpituitary adenylate cyclase-activating polypeptide (PACAP) and its recep-tor 1 in rat brain: function of PACAP as a neurotrophic factor. Ann N YAcad Sci 865:189–196.
Mayer C, Popanda O, Zelezny O, von Brevern MC, Bach A, Bartsch H,Schmezer P. 2002. DNA repair capacity after gamma-irradiation andexpression profiles of DNA repair genes in resting and proliferatinghuman peripheral blood lymphocytes. DNA Repair 1:237–250.
Miehe M, Leterrier JF, Deloulme JC, Gensburger C, Knoetgen MF,Sensenbrenner M. 1996. Influence of basic fibroblast growth factor andastroglial cells on the ultrastructure of developing rat brain neuronalprecursors in vitro. Dev Neurosci 18:210–223.
Muller JM, Lolait SJ, Yu VC, Sadee W, Waschek JA. 1989. Functionalvasoactive intestinal polypeptide (VIP) receptors in human neuroblastomasubclones that contain VIP precursor mRNA and release VIP-like sub-stances. J Biol Chem 264:3647–3650.
Muller JM, Lelievre V, Becq-Giraudon L, Meunier AC. 1995. VIP as acell-growth and differentiation neuromodulator role in neurodevelop-ment. Mol Neurobiol 10:115–134.
O’Dorisio MS, Fleshman DJ, Qualman SJ, O’Dorisio TM. 1992. Vasoac-tive intestinal peptide: autocrine growth factor in neuroblastoma. RegulPept 37:213–226.
Pence JC, Shorter NA. 1990. In vitro differentiation of human neuroblas-toma cells caused by vasoactive intestinal peptide. Cancer Res 50:5177–5183.
Qualman SJ, O’Dorisio MS, Fleshman DJ, Shimada H, O’Dorisio TM.1992. Neuroblastoma. Correlation of neuropeptide expression in tumortissue with other prognostic factors. Cancer 70:2005–2012.
Ross RA, Biedler JL. 1985. Presence and regulation of tyrosinase activity inhuman neuroblastoma cell variants in vitro. Cancer Res 45:1628–1632.
328 Heraud et al.

Ross RA, Spengler BA, Biedler JL. 1983. Coordinate morphological andbiochemical interconversion of human neuroblastoma cells. JNCI 71:741–747.
Sanchez C, Dıaz-Nido J, Avila J. 2000. Phosphorylation of microtubule-associated protein 2 (MAP2) and its relevance for the regulation of theneuronal cytoskeleton function. Prog Neurobiol 61:133–168.
Sasaki A, Sato S, Go M, Shimizu Y, Murakami O, Hanew K, Yumita S,Andoh N, Sasano N, Yoshinaga K. 1987. Distribution, plasma concen-tration, and in vivo prolactin-releasing activity of peptide histidine me-thionine in humans. J Clin Endocrinol Metab 65:683–688.
Sharma M, Sharma P, Pant HC. 1999. CDK-5-mediated neurofilamentphosphorylation in SH-SY5Y human neuroblastoma cells. J Neurochem73:79–86.
Shea TB, Beermann ML. 1994. Respective roles of neurofilaments, micro-tubules, MAP1B, and tau in neurite outgrowth and stabilization. Mol BiolCell 5:863–875.
Sherwood NM, Krueckl SL, McRory JE. 2000. The origin and function ofthe pituitary adenylate cyclase-activating polypeptide (PACAP)/glucagonsuperfamily. Endocrine Rev 21:619–670.
Steinschneider R, Delmas P, Nedelec J, Gola M, Bernard D, Boucraut J.1996. Appearance of neurofilament subunit epitopes correlates with elec-trophysiological maturation in cortical embryonic neurons coculturedwith mature astrocytes. Brain Res Dev Brain Res 20:15–27.
Towbin H, Staehelin T, Gordon J. 1979. Electrophoretic transfer of pro-teins from polyacrylamide gels to nitrocellulose sheets: procedure andsome applications. Proc Natl Acad Sci USA 76:4350–4354.
Tsuneishi S, Sano K, Nakamura H. 1993. Serum depletion increases theneurofilament protein mRNA levels in a neuroblastoma cell line, GOTO.Brain Res Mol Brain Res 17:119–128.
Vaudry D, Gonzalez BJ, Basille M, Fournier A, Vaudry H. 1999. Neuro-trophic activity of pituitary adenylate cyclase-activating polypeptide on rat
cerebellar cortex during development. Proc Natl Acad Sci USA 96:9415–9420.
Vaudry D, Gonzalez BJ, Basille M, Yon L, Fournier A, Vaudry H. 2000.Pituitary adenylate cyclase-activating polypeptide and its receptors: fromstructure to functions. Pharmacol Rev 52:269–324.
Vertongen P, De Clerck P, Fournet JC, Martelli H, Helardot P, Devalck C,Peeters T, Sariban E, Robberecht P. 1997. Comparison between vaso-active intestinal peptide and pituitary adenylate cyclase-activatingpolypeptide levels in neuroblastoma tumour tissues. Neuropeptides 31:409–413.
Waschek JA. 1995. Vasoactive intestinal peptide: an important trophicfactor and developmental regulator? Dev Neurosci 17:1–7.
Waschek JA, Lelievre V, Bravo DT, Nguyen T, Muller JM. 1997. Retinoicacid regulation of the VIP and PACAP autocrine ligand and receptorsystem in human neuroblastoma cell lines. Peptides 18:835–841.
Watanobe H, Tamura T. 1994. Stimulation by peptide histidine methio-nine (PHM) of adrenocorticotropin secretion in patients with Cushing’sdisease: a comparison with the effect of vasoactive intestinal peptide (VIP)and a study on the effect of combined administration of corticotrophin-releasing hormone with PHM or VIP. J Clin Endocrinol Metab 78:1372–1377.
Wollman Y, Blumberg S, Spungin A, Brenneman DE, Fridkin M, Woll-man J, Iaina A, Gozes I. 2002. The increased proliferation of culturedneuroblastoma cells treated with vasoactive intestinal peptide is enhancedby simultaneous inhibition of neutral endopeptidase. Regul Pept 108:175–177.
Yao W, Sheikh SP, Ottesen B, Jorgensen JC. 1996. Vascular effects andcyclic AMP production produced by VIP, PHM, PHV, PACAP-27,PACAP-38, and NPY on rabbit ovarian artery. Peptides 17:809–815.
Zupan V, Nehlig A, Evrard P, Gressens P. 2000. Prenatal blockade ofvasoactive intestinal peptide alters cell death and synaptic equipment inthe murine neocortex. Pediatr Res 47:53–63.
VIP and Analogs in SH-SY5Y Human Cells 329





















![Terminally Differentiated SH-SY5Y Cells Provide a Model [887035]](https://img.pdfslide.us/doc/110x75/577c832e1a28abe054b3f104/terminally-differentiated-sh-sy5y-cells-provide-a-model-887035.jpg)







