Embed Size (px)
Citation preview

Neural Crest Cell Contribution to theDeveloping Circulatory SystemImplications for Vascular Morphology?
Maarten Bergwerff, Marlies E. Verberne, Marco C. DeRuiter,Robert E. Poelmann, Adriana C. Gittenberger-de Groot
Abstract—In this study, the distribution patterns of neural crest (NC) cells (NCCs) in the developing vascular system of thechick were thoroughly studied and examined for a correlation with smooth muscle cell differentiation and vascularmorphogenesis. For this purpose, we performed long-term lineage tracing using quail-chick chimera techniques andpremigratory NCC infection with a replication-incompetent retrovirus containing the LacZ reporter gene in combinationwith immunohistochemistry. Results indicate that NCC deposition around endothelial tubes is influenced by anteropos-terior positional information from the pharyngeal arterial system. NCCs were shown to be among the first cells todifferentiate into primary smooth muscle cells of the arch arteries. At later stages, NCCs eventually differentiated intoadventitial fibroblasts and smooth muscle cells and nonmuscular cells of the media and intima. NCCs were distributed inthe aortic arch and pulmonary arch arteries and in the brachiocephalic and carotid arteries. The coronary and pulmonaryarteries and the descending aorta, however, remained devoid of NCCs. A new finding was that the media of part of theanterior cardinal veins was also determined to be NC-derived. NC-derived elastic arteries differed from non-NC elasticvessels in their cellular constitution and elastic fiber organization, and the NC appeared not to be involved in designatinga muscular or elastic artery. Boundaries between NC-infested areas and mesodermal vessel structures were mostly verysharp and tended to coincide with marked changes in vascular morphology, with the exception of an intriguing area inthe aortic and pulmonary trunks. (Circ Res. 1998;82:221-231.)
Key Words: quail-chick chimera n pharyngeal arch artery n smooth muscle cell n great artery n neural crest migration
Neural crest cells have been studied extensively for theirmigratory behavior and potential to differentiate into a
wide variety of cell types. In cardiovascular research, the crestregion located between the midotic placode and somite 3, alsocalled the cardiac NC,1 has received much attention because ofits major role in cardiovascular development. Participation inoutflow tract septation,1,2 media formation of the pharyngealarch arteries,3–5 and contribution to parasympathetic cardiacinnervation3 have been documented using quail-chick chime-ras. The importance of the extensive contribution of NCCs tosuch a wide variety of cardiovascular structures is made clear bythe occurrence of congenital malformations involving the NCand presenting more or less severe cardiovascular anomalies.Syndromes associated with 22q11 deletions and vitamin Adeficiency are known NC-associated disorders, including car-diovascular malformations.6,7 Experimental removal of thecardiac crest in chick embryos,8,9 administration of excessretinoids,10–12 and deletion of retinoic acid receptor genes13 alsoresult in severe malformations like ventricular septal defects,common arterial trunk, and aortic arch interruptions. In thepresent study, the emphasis is not on malformations due to NC
disorders but on the actual role of the crest in normaldevelopment, focusing on the ectomesodermal lineage thatdifferentiates into vascular SMCs to form the media of thegreat thoracic arteries.
Vascular SMCs are reported to originate from either thesplanchnic mesoderm14,15 or the NC.3 Recently, a third possi-ble origin was ascribed to the endothelial cells, which appear totransdifferentiate into SMCs in the early avian dorsal aorta.16 LeLievre and Le Douarin3 have reported on the distribution ofNCCs to the great arteries in the thorax. Other studies havefurther elaborated on NC seeding of the pharyngeal archesboth in birds4,5,17–19 and in mammals.20,21 The tunica media ofthe great vessels derived from the pharyngeal arch arteries (eg,aortic arch, brachiocephalic arteries, and pulmonary archarteries) was shown to consist almost exclusively of NCCs,whereas other thoracic vessels (eg, pulmonary arteries, subcla-vian arteries, and descending aorta) appeared devoid of NCCsand thus entirely mesodermal in origin.3,5 This unequal distri-bution of cells of different embryonic origins may be ofinfluence on differential vascular patterning.
Recently, we reported on the differentiation of the thoracicarteries during chick embryonic development and suggested
Received May 12, 1997; accepted October 21, 1997.From the Department of Anatomy and Embryology, Leiden (the Netherlands) University Medical Centre.Correspondence to A.C. Gittenberger-de Groot, Department of Anatomy and Embryology, Leiden University Medical Centre, PO Box 9602, 2300 RC
Leiden, Netherlands.E-mail [email protected]© 1998 American Heart Association, Inc.
221
by guest on May 23, 2017
http://circres.ahajournals.org/D
ownloaded from

that morphogenesis of vessels described to be of NC origin wasdifferent from that of pure mesodermal derivatives. Thepresence of clearly distinguishable alternating SMC and non-muscular medial cell layers appeared to be confined to NCvessels. Furthermore, presumed NC vessels exhibited an elasticfiber organization different from that of mesodermal vessels.22
Moreover, several studies recently mentioned different in vitrocharacteristics of SMCs isolated from either mesodermal (ab-dominal aorta) or NC (thoracic aorta) vessels.23–25
In order to study both NCC distribution and NCC differ-entiation in the developing cardiovascular system at the sametime, we used a lineage tracing system with a replication-incompetent retrovirus containing the LacZ reporter gene.26
The particular retrovirus we used has been proven beneficial instudying the avian NC lineage27–29 as well as several other celllineage systems.30–32 Retroviral introduction of the LacZ geneis, in contrast to fluorescent dye injections, beneficial indetecting cell lineages over long time spans and is, at the sametime, much less invasive than chimera techniques, circumvent-ing major interference with normal development. In addition,clear whole-mount distribution patterns of NCCs can bevisualized using b-gal conversion. Using the retrovirus incombination with immunohistochemistry, we have been ableto map distribution patterns of NCCs at any desired develop-mental stage and to superimpose vascular differentiation pat-terns in an attempt to relate NC contribution and vascularmorphogenesis. For comparison, quail-chick chimeras werealso included in the present study. Chimeras have the advan-tage of labeling virtually all cells emanating from the cardiaccrest.
Materials and MethodsVirusThe replication-incompetent virus, designated CXL, that was used inthe present study was kindly provided by Dr T. Mikawa (CornellUniversity Medical College, New York, NY).26 The design of thisvector was based on spleen necrosis virus. It lacks the structural viralproteins necessary for replication and carries the LacZ reporter gene.Strong and stable expression of LacZ under control of the promoter inthe spleen necrosis virus long-terminal repeat has been established ina wide range of avian embryonic cell types.26,27,30,31 The virus wasproduced in canine packaging cells (D17.2G/CXL) cultured inIscove’s modified DMEM (IMDM, GIBCO) containing 5% FCS(GIBCO) and penicillin/streptomycin.
On the day of the experiments, CXL was harvested from confluentmonolayers 24 hours after changing the medium. Cells were scraped fromthe culture flasks and ground in their medium, followed by briefcentrifugation to remove cellular debris. CXL was subsequently pelletedby centrifugation of the medium at 17 000g and 22°C for 21⁄2 hours andresuspended in a small volume of medium containing 100 mg/mLpolybrene (Sigma Chemical Co) and indigo carmine blue. The harvestedvirus batch was used for both infection of embryos and titration on cells.Viral titers were determined in duplicate by infection of R2 rat fibroblasts
with dilutions of the virus batch on the day of the experiment. Titersvaried between 2 and 43106 transducing units/mL.
Infection of EmbryosFertilized specified pathogen-free white leghorn eggs (ID-DLO) wereincubated for 36 to 42 hours at 37°C, windowed, and stagedaccording to Hamburger and Hamilton.33 Embryos between stages 8and 10 were used for infection with freshly prepared CXL solutioncontaining polybrene. Using glass micropipettes and carefully exertedpressure by a Hamilton syringe connected to an oil-filled system, theneural groove/tube was filled with viral suspension from the somite-4to -5 region in an anterior direction. Spilling of virus over the neuralfolds or out of the anterior neuropore was minimized as much aspossible. After injection, the eggs were tightly sealed with Scotch tapeand returned to the incubator for further development.
Tissue Preparation and b-Gal StainingInfected embryos were allowed to develop until stages 19 to 40,collected, and further processed. The youngest embryos (stages 19 to33) were fixed in toto by immersion in 4% paraformaldehyde in PBS(4 hours, 4°C), whereas bigger embryos were first perfused with thefixative via the right atrium, followed by immersion-fixation. Afterextensive rinsing in PBS, the thoracic viscera were carefully removedfrom fixed older embryos. Next, embryos and heart-lung specimenswere stained by immersion in X-gal solution (PBS containing5 mmol/L potassium ferricyanide, 5 mmol/L potassium ferrocyanide,2 mmol/L MgCl2, and 0.1% [wt/vol] X-gal [C14H15BrClNO6, Boeh-
ringer]) while being shaken at 37°C for 2 to 4 hours. Again, thecardiovascular system of larger specimens was perfused with X-galsolution before immersion-incubation. Stained embryos were thor-oughly rinsed in PBS, evaluated macroscopically, and processed forimmunohistochemistry. The results are based on a total number of 56successfully labeled embryos.
Immunohistochemistryb-Gal–stained embryos and heart-lung specimens were swiftly dehy-drated in graded ethanol, followed by Paraclear (Earth Safe IndustriesInc) and subsequent embedding in paraffin. Xylene-free Paraclear wasused to prevent the blue precipitate from dissolving. Sections of 5 mmwere cut and distributed over several series of glass slides, allowing forvarious staining procedures of each embryo. Standard resorcin/fuchsinstaining was used to visualize elastic fibers, and monoclonal antibodiesagainst muscle-specific actin (HHF35,34 DAKO A/S) and HNK-135
enabled discernment of differential pathways of NC derivatives.Before antibody incubation, endogenous peroxidase activity wasquenched by treatment with 0.3% H2O2 in PBS. Routine immuno-histochemical staining was performed using overnight incubationswith the primary antibodies diluted in PBS with 0.05% Tween 20 and1% chicken egg albumin (HHF35, 1:1000; HNK-1 medium, 1:10).Sections were then thoroughly rinsed, and bound primary antibodieswere visualized with horseradish peroxidase–conjugated rabbit anti-mouse antibodies (1:300, DAKO A/S) and treatment with 0.04%diaminobenzidine tetrahydrochloride/0.06‰ H2O2 in 0.05 mol/LTRIS-maleic acid (pH 7.6) for 10 minutes at room temperature. Thenthe sections were counterstained with Mayer’s hematoxylin, dehy-drated in ethanol and Paraclear, and mounted in Entellan (Merck).
ChimerasFertilized eggs of White Leghorn chicken and Japanese quail (Coturnixcoturnix) were incubated for 40 to 42 and 36 to 38 hours, respectively,and windowed, and the embryos were staged after slight staining withNile blue sulfate. In stage-10 chicken and quail embryos, the cardiacNC (midotic to somite 3) was ablated by removing the dorsal part ofthe neural tube with a sharpened tungsten needle. Subsequently, quailNC tissue was homotopically transplanted onto the graft site of thechick embryo. After sealing the eggs, the chimeric embryos werereincubated and killed between stages 28 and 40. Whole embryos anddissected thorax segments were fixed in 2% acetic acid and 98%ethanol (overnight at 4°C), embedded in paraffin, and further pro-cessed according to the aforementioned immunohistochemical proce-
Selected Abbreviations and Acronyms
b-gal 5 b-galactosidaseDA 5 ductus arteriosusNC 5 neural crest
NCC 5 neural crest cellSMC 5 smooth muscle cellTGF 5 transforming growth factor
222 Neural Crest and Vascular Differentiation
by guest on May 23, 2017
http://circres.ahajournals.org/D
ownloaded from

dures. In addition to antibodies revealing differentiation antigens(HHF35, HNK-1, and smooth muscle a-actin [1A4, DAKO A/S]),the antibody QCPN (a quail nuclear marker, Developmental StudiesHybridoma Bank) was used to visualize distribution of quail NC-derived cells. Twenty-two successfully transplanted chimeras wereused in the present study.
ResultsStages 19 to 21NC-derived cells showing b-gal activity were shown to havemigrated into the entire pharyngeal arch region (Fig 1a). Thedense mesenchyme of the maxillary buds and branchial archesthat surrounds the arch arteries exhibited moderate to exten-sive blue staining. NCC distribution extended from the mostventrally located arch mesenchyme, alongside the pharynx, andgenerally up to the paired dorsal aortas and anterior cardinalveins. The NC distribution pattern along the pharyngealarterial system is depicted in Fig 2a. In contrast to theectomesenchyme of the pharyngeal arches, the mesenchymethat surrounds the aortic sac region is far less dense, and only afew labeled cells positioned both dorsally and ventrally of theaortic sac and in the endocardial cushion tissue of the outflowtract were shown. Ectomesenchymal contribution to the aorticsac therefore appeared limited at these stages. Endothelial cells
in the entire pharyngeal region never showed staining for b-galactivity. By use of a double staining against muscle-specificactins, part of the NCCs in the aortic sac revealed actinexpression indicative of early SMC differentiation (Fig 1b and1c). The pharyngeal arch arteries, which are also aligned byNC-derived ectomesenchyme, did not yet show signs of earlyactin expression at this stage in development. The paired dorsalaortas, on the other hand, showed a thin layer of actin-positivecells directly adjacent to the endothelium. These primarySMCs in the dorsal aorta were never shown to be of NC originin the region posterior to arch 4. However, the anterior dorsalaortas (ie, ductus caroticus and early carotids) showed coex-pression of actin and b-gal in a small number of cells,suggesting the onset of SMC differentiation of NC-derivedcells.
Stages 23 and 24Whole-mount–stained stage 23/24 embryos clearly demon-strated the disposition of the circumpharyngeal crest area (Fig1d). Sections revealed massive NC infestation of the arches,whereas further upstream toward the heart, the arch arteriesand aortic sac were surrounded by lower numbers of NCCs(Fig 1e and 1f). The lateral sides of the arch arteries clearly
Figure 1. a, Whole-mount b-gal–stainedstage-19 embryo showing NC-derivedcells in the head and pharyngeal region.Within the arches (I, II, III, and IV), individ-ual cells can be recognized as blue dots.E indicates eye; H, heart. Bar5100 mm. band c, Light micrographs of a sectionthrough the aortic sac region of thestage-19 embryo, depicted in panel a,showing presence of blue-stained NCCsin the aortic sac and near the outflowtract and double staining for muscle-specific actin (cells combining blue andbrown staining, arrow) indicative of pri-mary SMC differentiation. AoS indicatesaortic sac; OT, outflow tract. Bar550 mm(b) and 7,5 mm (c). d, Whole-mount–stained stage-24 embryo demonstratingmassive LacZ expression in the pharyn-geal arch region. The dashed line indi-cates the clear dorsal and posteriorboundary of the NC-infested circumpha-ryngeal crest area. Bar5250 mm. e and f,Micrographs of a stage-23 embryo show-ing periendothelial actin staining (brown)of the arch arteries (III, IV, and VI) andpreferential localization of NCCs (blue) atthe most lateral side of the pharyngealarches. Part of the NCC population hasbecome clearly associated with the arter-ies (arrows). AoS indicates aortic sac; PC,pharyngeal cleft; and T, trachea.Bar5150 mm (e and f).
Bergwerff et al 223
by guest on May 23, 2017
http://circres.ahajournals.org/D
ownloaded from
showed more seeding with NCCs than the median parts atthese stages. Moreover, no indication of a prospective aorti-copulmonary septum was found in any of the embryos, asNCC numbers were extremely low at the dorsal side of theaortic sac area. NCCs extended well into the outflow tractcushions, preferably in a dorsally located stretch of cells. In theaortic sac and arch arteries, part of the NCC population wasdouble-stained for LacZ and muscle actin.
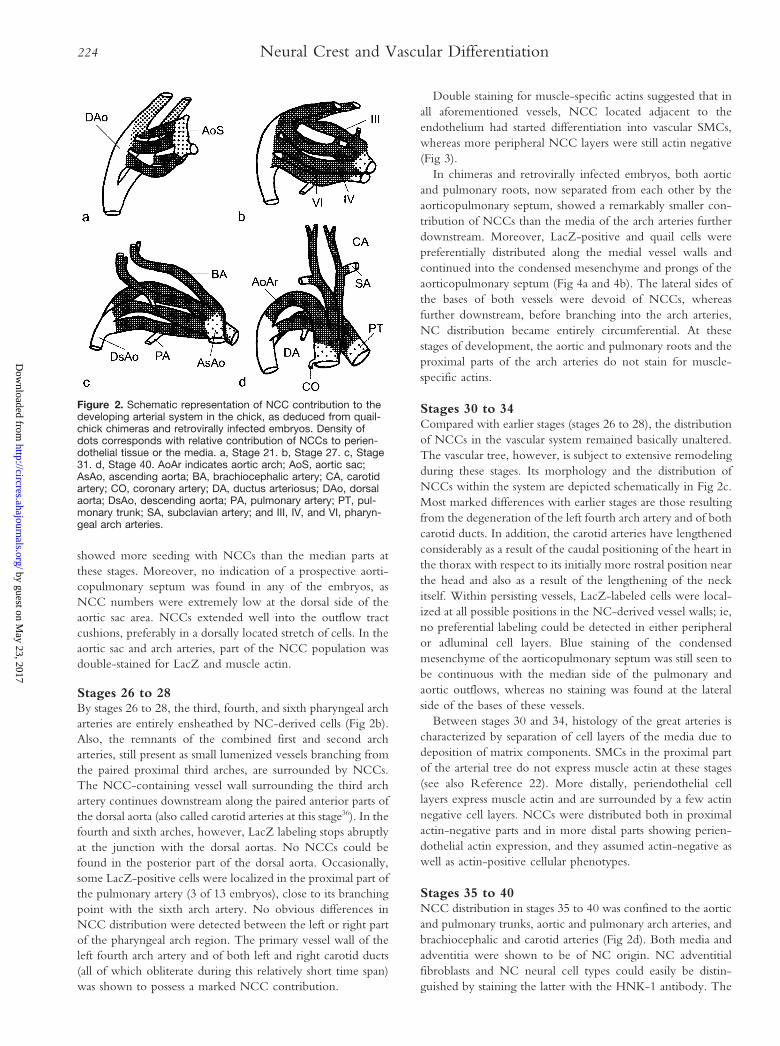
Stages 26 to 28By stages 26 to 28, the third, fourth, and sixth pharyngeal archarteries are entirely ensheathed by NC-derived cells (Fig 2b).Also, the remnants of the combined first and second archarteries, still present as small lumenized vessels branching fromthe paired proximal third arches, are surrounded by NCCs.The NCC-containing vessel wall surrounding the third archartery continues downstream along the paired anterior parts ofthe dorsal aorta (also called carotid arteries at this stage36). In thefourth and sixth arches, however, LacZ labeling stops abruptlyat the junction with the dorsal aortas. No NCCs could befound in the posterior part of the dorsal aorta. Occasionally,some LacZ-positive cells were localized in the proximal part ofthe pulmonary artery (3 of 13 embryos), close to its branchingpoint with the sixth arch artery. No obvious differences inNCC distribution were detected between the left or right partof the pharyngeal arch region. The primary vessel wall of theleft fourth arch artery and of both left and right carotid ducts(all of which obliterate during this relatively short time span)was shown to possess a marked NCC contribution.
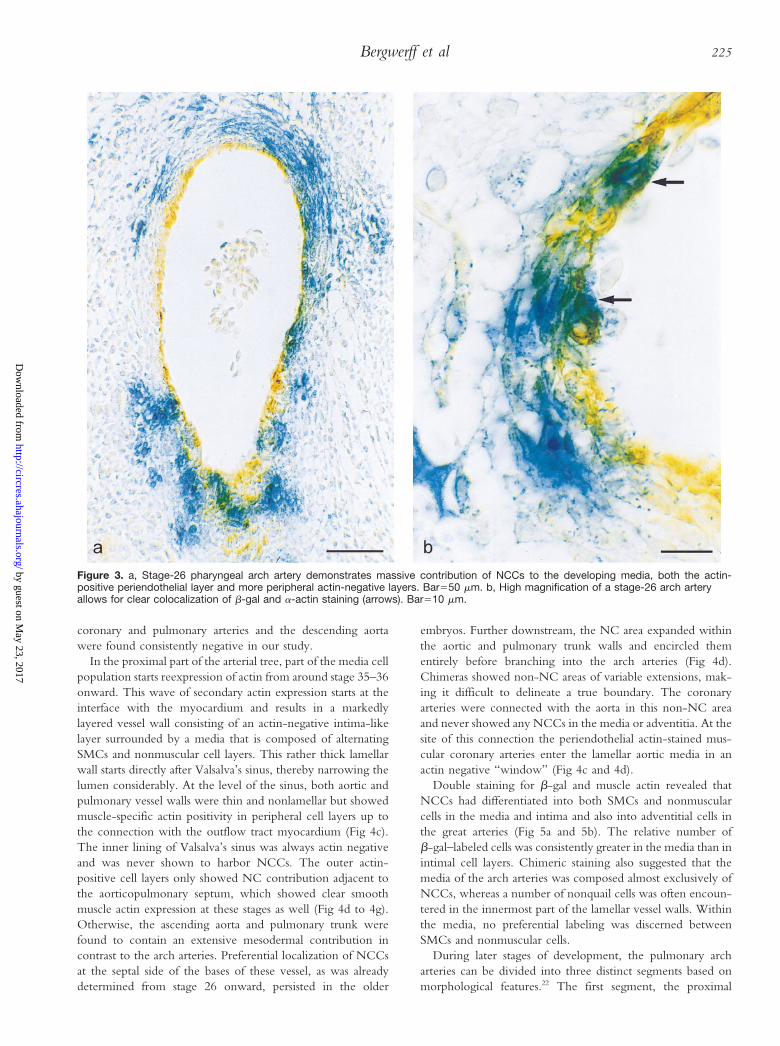
Double staining for muscle-specific actins suggested that inall aforementioned vessels, NCC located adjacent to theendothelium had started differentiation into vascular SMCs,whereas more peripheral NCC layers were still actin negative(Fig 3).
In chimeras and retrovirally infected embryos, both aorticand pulmonary roots, now separated from each other by theaorticopulmonary septum, showed a remarkably smaller con-tribution of NCCs than the media of the arch arteries furtherdownstream. Moreover, LacZ-positive and quail cells werepreferentially distributed along the medial vessel walls andcontinued into the condensed mesenchyme and prongs of theaorticopulmonary septum (Fig 4a and 4b). The lateral sides ofthe bases of both vessels were devoid of NCCs, whereasfurther downstream, before branching into the arch arteries,NC distribution became entirely circumferential. At thesestages of development, the aortic and pulmonary roots and theproximal parts of the arch arteries do not stain for muscle-specific actins.
Stages 30 to 34Compared with earlier stages (stages 26 to 28), the distributionof NCCs in the vascular system remained basically unaltered.The vascular tree, however, is subject to extensive remodelingduring these stages. Its morphology and the distribution ofNCCs within the system are depicted schematically in Fig 2c.Most marked differences with earlier stages are those resultingfrom the degeneration of the left fourth arch artery and of bothcarotid ducts. In addition, the carotid arteries have lengthenedconsiderably as a result of the caudal positioning of the heart inthe thorax with respect to its initially more rostral position nearthe head and also as a result of the lengthening of the neckitself. Within persisting vessels, LacZ-labeled cells were local-ized at all possible positions in the NC-derived vessel walls; ie,no preferential labeling could be detected in either peripheralor adluminal cell layers. Blue staining of the condensedmesenchyme of the aorticopulmonary septum was still seen tobe continuous with the median side of the pulmonary andaortic outflows, whereas no staining was found at the lateralside of the bases of these vessels.
Between stages 30 and 34, histology of the great arteries ischaracterized by separation of cell layers of the media due todeposition of matrix components. SMCs in the proximal partof the arterial tree do not express muscle actin at these stages(see also Reference 22). More distally, periendothelial celllayers express muscle actin and are surrounded by a few actinnegative cell layers. NCCs were distributed both in proximalactin-negative parts and in more distal parts showing perien-dothelial actin expression, and they assumed actin-negative aswell as actin-positive cellular phenotypes.
Stages 35 to 40NCC distribution in stages 35 to 40 was confined to the aorticand pulmonary trunks, aortic and pulmonary arch arteries, andbrachiocephalic and carotid arteries (Fig 2d). Both media andadventitia were shown to be of NC origin. NC adventitialfibroblasts and NC neural cell types could easily be distin-guished by staining the latter with the HNK-1 antibody. The
Figure 2. Schematic representation of NCC contribution to thedeveloping arterial system in the chick, as deduced from quail-chick chimeras and retrovirally infected embryos. Density ofdots corresponds with relative contribution of NCCs to perien-dothelial tissue or the media. a, Stage 21. b, Stage 27. c, Stage31. d, Stage 40. AoAr indicates aortic arch; AoS, aortic sac;AsAo, ascending aorta; BA, brachiocephalic artery; CA, carotidartery; CO, coronary artery; DA, ductus arteriosus; DAo, dorsalaorta; DsAo, descending aorta; PA, pulmonary artery; PT, pul-monary trunk; SA, subclavian artery; and III, IV, and VI, pharyn-geal arch arteries.
224 Neural Crest and Vascular Differentiation
by guest on May 23, 2017
http://circres.ahajournals.org/D
ownloaded from
coronary and pulmonary arteries and the descending aortawere found consistently negative in our study.
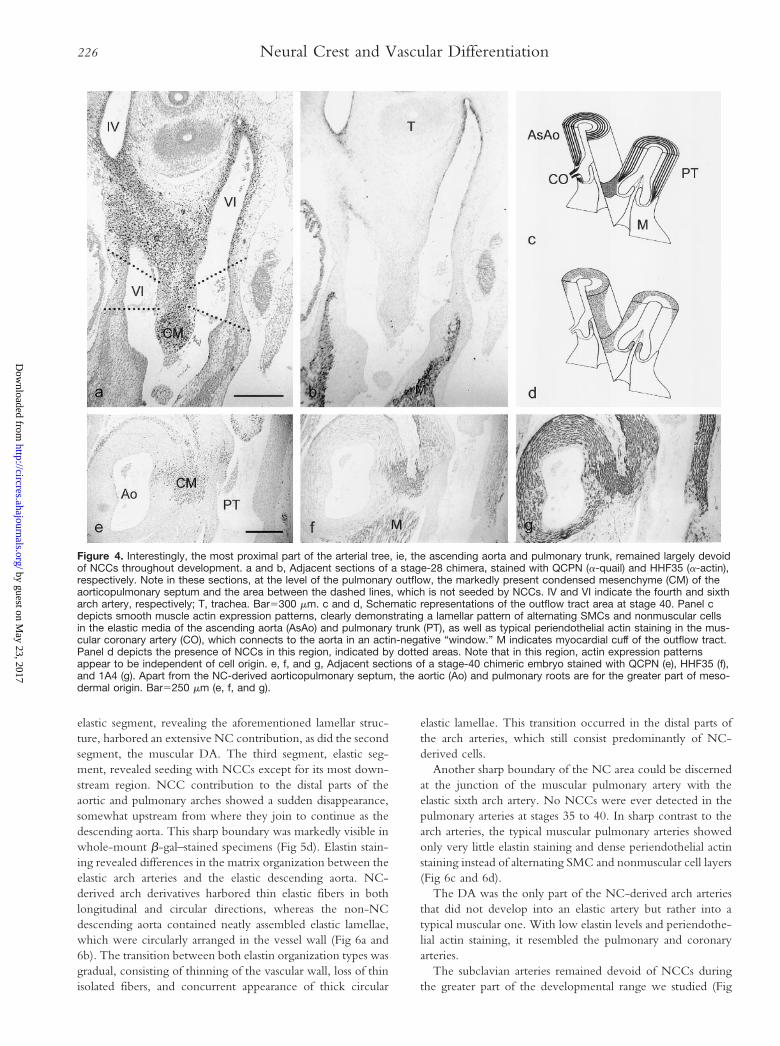
In the proximal part of the arterial tree, part of the media cellpopulation starts reexpression of actin from around stage 35–36onward. This wave of secondary actin expression starts at theinterface with the myocardium and results in a markedlylayered vessel wall consisting of an actin-negative intima-likelayer surrounded by a media that is composed of alternatingSMCs and nonmuscular cell layers. This rather thick lamellarwall starts directly after Valsalva’s sinus, thereby narrowing thelumen considerably. At the level of the sinus, both aortic andpulmonary vessel walls were thin and nonlamellar but showedmuscle-specific actin positivity in peripheral cell layers up tothe connection with the outflow tract myocardium (Fig 4c).The inner lining of Valsalva’s sinus was always actin negativeand was never shown to harbor NCCs. The outer actin-positive cell layers only showed NC contribution adjacent tothe aorticopulmonary septum, which showed clear smoothmuscle actin expression at these stages as well (Fig 4d to 4g).Otherwise, the ascending aorta and pulmonary trunk werefound to contain an extensive mesodermal contribution incontrast to the arch arteries. Preferential localization of NCCsat the septal side of the bases of these vessel, as was alreadydetermined from stage 26 onward, persisted in the older
embryos. Further downstream, the NC area expanded withinthe aortic and pulmonary trunk walls and encircled thementirely before branching into the arch arteries (Fig 4d).Chimeras showed non-NC areas of variable extensions, mak-ing it difficult to delineate a true boundary. The coronaryarteries were connected with the aorta in this non-NC areaand never showed any NCCs in the media or adventitia. At thesite of this connection the periendothelial actin-stained mus-cular coronary arteries enter the lamellar aortic media in anactin negative “window” (Fig 4c and 4d).
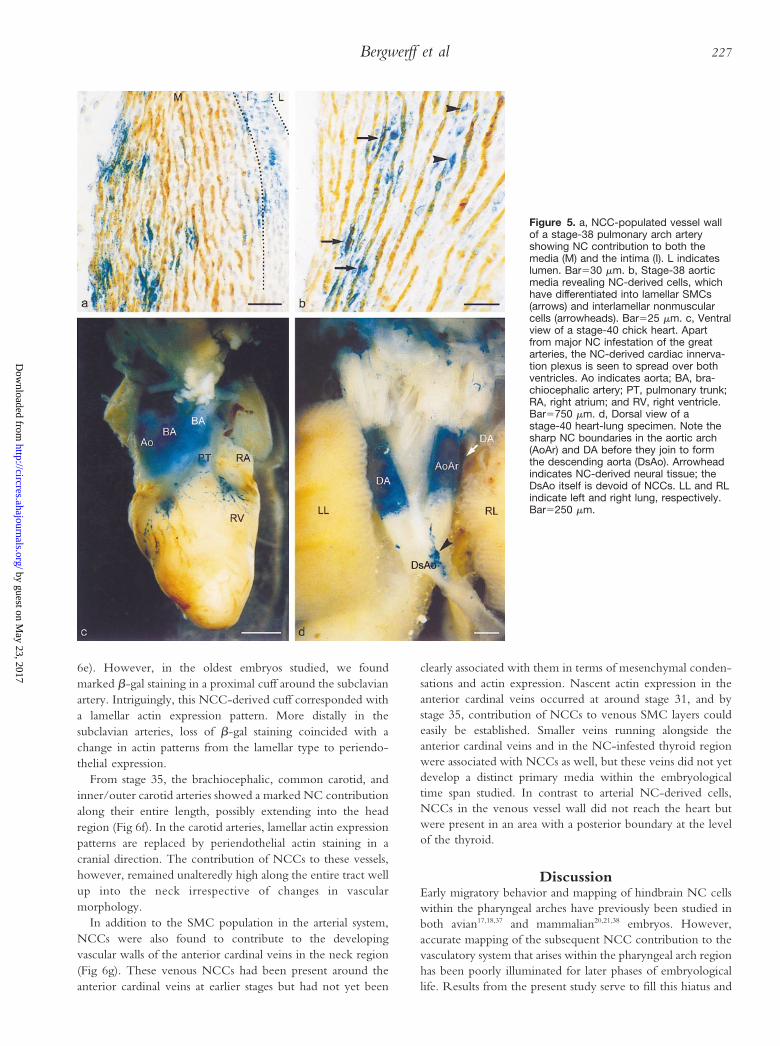
Double staining for b-gal and muscle actin revealed thatNCCs had differentiated into both SMCs and nonmuscularcells in the media and intima and also into adventitial cells inthe great arteries (Fig 5a and 5b). The relative number ofb-gal–labeled cells was consistently greater in the media than inintimal cell layers. Chimeric staining also suggested that themedia of the arch arteries was composed almost exclusively ofNCCs, whereas a number of nonquail cells was often encoun-tered in the innermost part of the lamellar vessel walls. Withinthe media, no preferential labeling was discerned betweenSMCs and nonmuscular cells.
During later stages of development, the pulmonary archarteries can be divided into three distinct segments based onmorphological features.22 The first segment, the proximal
Figure 3. a, Stage-26 pharyngeal arch artery demonstrates massive contribution of NCCs to the developing media, both the actin-positive periendothelial layer and more peripheral actin-negative layers. Bar550 mm. b, High magnification of a stage-26 arch arteryallows for clear colocalization of b-gal and a-actin staining (arrows). Bar510 mm.
Bergwerff et al 225
by guest on May 23, 2017
http://circres.ahajournals.org/D
ownloaded from
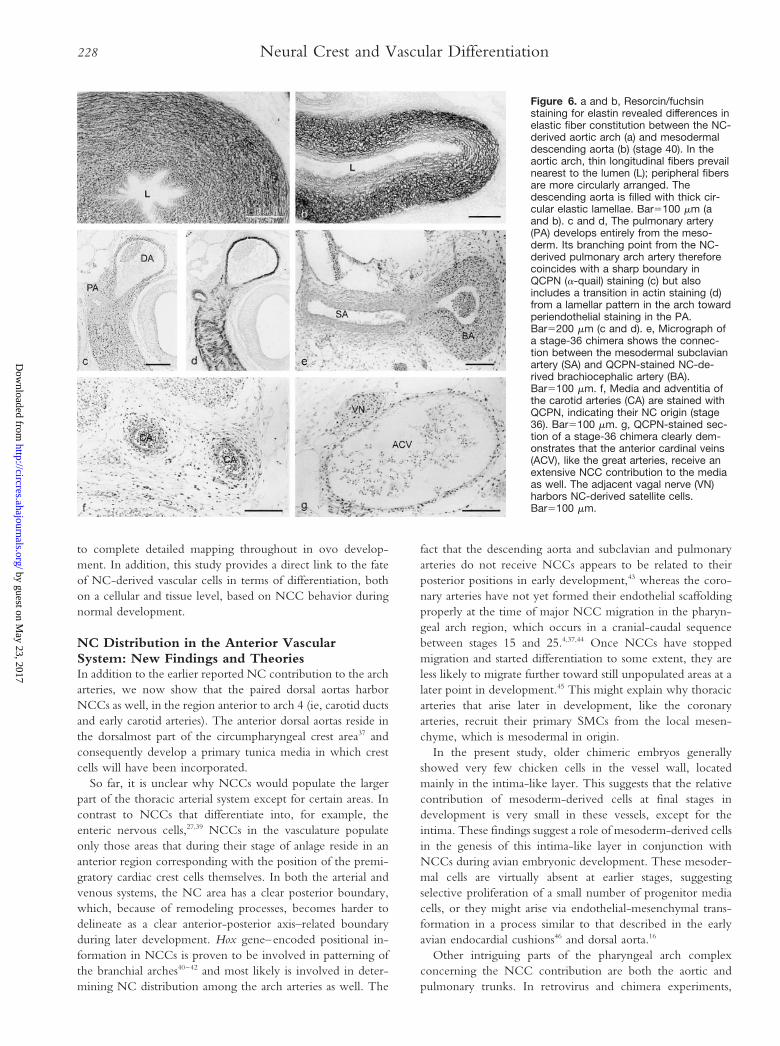
elastic segment, revealing the aforementioned lamellar struc-ture, harbored an extensive NC contribution, as did the secondsegment, the muscular DA. The third segment, elastic seg-ment, revealed seeding with NCCs except for its most down-stream region. NCC contribution to the distal parts of theaortic and pulmonary arches showed a sudden disappearance,somewhat upstream from where they join to continue as thedescending aorta. This sharp boundary was markedly visible inwhole-mount b-gal–stained specimens (Fig 5d). Elastin stain-ing revealed differences in the matrix organization between theelastic arch arteries and the elastic descending aorta. NC-derived arch derivatives harbored thin elastic fibers in bothlongitudinal and circular directions, whereas the non-NCdescending aorta contained neatly assembled elastic lamellae,which were circularly arranged in the vessel wall (Fig 6a and6b). The transition between both elastin organization types wasgradual, consisting of thinning of the vascular wall, loss of thinisolated fibers, and concurrent appearance of thick circular
elastic lamellae. This transition occurred in the distal parts ofthe arch arteries, which still consist predominantly of NC-derived cells.
Another sharp boundary of the NC area could be discernedat the junction of the muscular pulmonary artery with theelastic sixth arch artery. No NCCs were ever detected in thepulmonary arteries at stages 35 to 40. In sharp contrast to thearch arteries, the typical muscular pulmonary arteries showedonly very little elastin staining and dense periendothelial actinstaining instead of alternating SMC and nonmuscular cell layers(Fig 6c and 6d).
The DA was the only part of the NC-derived arch arteriesthat did not develop into an elastic artery but rather into atypical muscular one. With low elastin levels and periendothe-lial actin staining, it resembled the pulmonary and coronaryarteries.
The subclavian arteries remained devoid of NCCs duringthe greater part of the developmental range we studied (Fig
Figure 4. Interestingly, the most proximal part of the arterial tree, ie, the ascending aorta and pulmonary trunk, remained largely devoidof NCCs throughout development. a and b, Adjacent sections of a stage-28 chimera, stained with QCPN (a-quail) and HHF35 (a-actin),respectively. Note in these sections, at the level of the pulmonary outflow, the markedly present condensed mesenchyme (CM) of theaorticopulmonary septum and the area between the dashed lines, which is not seeded by NCCs. IV and VI indicate the fourth and sixtharch artery, respectively; T, trachea. Bar5300 mm. c and d, Schematic representations of the outflow tract area at stage 40. Panel cdepicts smooth muscle actin expression patterns, clearly demonstrating a lamellar pattern of alternating SMCs and nonmuscular cellsin the elastic media of the ascending aorta (AsAo) and pulmonary trunk (PT), as well as typical periendothelial actin staining in the mus-cular coronary artery (CO), which connects to the aorta in an actin-negative “window.” M indicates myocardial cuff of the outflow tract.Panel d depicts the presence of NCCs in this region, indicated by dotted areas. Note that in this region, actin expression patternsappear to be independent of cell origin. e, f, and g, Adjacent sections of a stage-40 chimeric embryo stained with QCPN (e), HHF35 (f),and 1A4 (g). Apart from the NC-derived aorticopulmonary septum, the aortic (Ao) and pulmonary roots are for the greater part of meso-dermal origin. Bar5250 mm (e, f, and g).
226 Neural Crest and Vascular Differentiation
by guest on May 23, 2017
http://circres.ahajournals.org/D
ownloaded from
6e). However, in the oldest embryos studied, we foundmarked b-gal staining in a proximal cuff around the subclavianartery. Intriguingly, this NCC-derived cuff corresponded witha lamellar actin expression pattern. More distally in thesubclavian arteries, loss of b-gal staining coincided with achange in actin patterns from the lamellar type to periendo-thelial expression.
From stage 35, the brachiocephalic, common carotid, andinner/outer carotid arteries showed a marked NC contributionalong their entire length, possibly extending into the headregion (Fig 6f). In the carotid arteries, lamellar actin expressionpatterns are replaced by periendothelial actin staining in acranial direction. The contribution of NCCs to these vessels,however, remained unalteredly high along the entire tract wellup into the neck irrespective of changes in vascularmorphology.
In addition to the SMC population in the arterial system,NCCs were also found to contribute to the developingvascular walls of the anterior cardinal veins in the neck region(Fig 6g). These venous NCCs had been present around theanterior cardinal veins at earlier stages but had not yet been
clearly associated with them in terms of mesenchymal conden-sations and actin expression. Nascent actin expression in theanterior cardinal veins occurred at around stage 31, and bystage 35, contribution of NCCs to venous SMC layers couldeasily be established. Smaller veins running alongside theanterior cardinal veins and in the NC-infested thyroid regionwere associated with NCCs as well, but these veins did not yetdevelop a distinct primary media within the embryologicaltime span studied. In contrast to arterial NC-derived cells,NCCs in the venous vessel wall did not reach the heart butwere present in an area with a posterior boundary at the levelof the thyroid.
DiscussionEarly migratory behavior and mapping of hindbrain NC cellswithin the pharyngeal arches have previously been studied inboth avian17,18,37 and mammalian20,21,38 embryos. However,accurate mapping of the subsequent NCC contribution to thevasculatory system that arises within the pharyngeal arch regionhas been poorly illuminated for later phases of embryologicallife. Results from the present study serve to fill this hiatus and
Figure 5. a, NCC-populated vessel wallof a stage-38 pulmonary arch arteryshowing NC contribution to both themedia (M) and the intima (I). L indicateslumen. Bar530 mm. b, Stage-38 aorticmedia revealing NC-derived cells, whichhave differentiated into lamellar SMCs(arrows) and interlamellar nonmuscularcells (arrowheads). Bar525 mm. c, Ventralview of a stage-40 chick heart. Apartfrom major NC infestation of the greatarteries, the NC-derived cardiac innerva-tion plexus is seen to spread over bothventricles. Ao indicates aorta; BA, bra-chiocephalic artery; PT, pulmonary trunk;RA, right atrium; and RV, right ventricle.Bar5750 mm. d, Dorsal view of astage-40 heart-lung specimen. Note thesharp NC boundaries in the aortic arch(AoAr) and DA before they join to formthe descending aorta (DsAo). Arrowheadindicates NC-derived neural tissue; theDsAo itself is devoid of NCCs. LL and RLindicate left and right lung, respectively.Bar5250 mm.
Bergwerff et al 227
by guest on May 23, 2017
http://circres.ahajournals.org/D
ownloaded from
to complete detailed mapping throughout in ovo develop-ment. In addition, this study provides a direct link to the fateof NC-derived vascular cells in terms of differentiation, bothon a cellular and tissue level, based on NCC behavior duringnormal development.
NC Distribution in the Anterior VascularSystem: New Findings and TheoriesIn addition to the earlier reported NC contribution to the archarteries, we now show that the paired dorsal aortas harborNCCs as well, in the region anterior to arch 4 (ie, carotid ductsand early carotid arteries). The anterior dorsal aortas reside inthe dorsalmost part of the circumpharyngeal crest area37 andconsequently develop a primary tunica media in which crestcells will have been incorporated.
So far, it is unclear why NCCs would populate the largerpart of the thoracic arterial system except for certain areas. Incontrast to NCCs that differentiate into, for example, theenteric nervous cells,27,39 NCCs in the vasculature populateonly those areas that during their stage of anlage reside in ananterior region corresponding with the position of the premi-gratory cardiac crest cells themselves. In both the arterial andvenous systems, the NC area has a clear posterior boundary,which, because of remodeling processes, becomes harder todelineate as a clear anterior-posterior axis–related boundaryduring later development. Hox gene–encoded positional in-formation in NCCs is proven to be involved in patterning ofthe branchial arches40–42 and most likely is involved in deter-mining NC distribution among the arch arteries as well. The
fact that the descending aorta and subclavian and pulmonaryarteries do not receive NCCs appears to be related to theirposterior positions in early development,43 whereas the coro-nary arteries have not yet formed their endothelial scaffoldingproperly at the time of major NCC migration in the pharyn-geal arch region, which occurs in a cranial-caudal sequencebetween stages 15 and 25.4,37,44 Once NCCs have stoppedmigration and started differentiation to some extent, they areless likely to migrate further toward still unpopulated areas at alater point in development.45 This might explain why thoracicarteries that arise later in development, like the coronaryarteries, recruit their primary SMCs from the local mesen-chyme, which is mesodermal in origin.
In the present study, older chimeric embryos generallyshowed very few chicken cells in the vessel wall, locatedmainly in the intima-like layer. This suggests that the relativecontribution of mesoderm-derived cells at final stages indevelopment is very small in these vessels, except for theintima. These findings suggest a role of mesoderm-derived cellsin the genesis of this intima-like layer in conjunction withNCCs during avian embryonic development. These mesoder-mal cells are virtually absent at earlier stages, suggestingselective proliferation of a small number of progenitor mediacells, or they might arise via endothelial-mesenchymal trans-formation in a process similar to that described in the earlyavian endocardial cushions46 and dorsal aorta.16
Other intriguing parts of the pharyngeal arch complexconcerning the NCC contribution are both the aortic andpulmonary trunks. In retrovirus and chimera experiments,
Figure 6. a and b, Resorcin/fuchsinstaining for elastin revealed differences inelastic fiber constitution between the NC-derived aortic arch (a) and mesodermaldescending aorta (b) (stage 40). In theaortic arch, thin longitudinal fibers prevailnearest to the lumen (L); peripheral fibersare more circularly arranged. Thedescending aorta is filled with thick cir-cular elastic lamellae. Bar5100 mm (aand b). c and d, The pulmonary artery(PA) develops entirely from the meso-derm. Its branching point from the NC-derived pulmonary arch artery thereforecoincides with a sharp boundary inQCPN (a-quail) staining (c) but alsoincludes a transition in actin staining (d)from a lamellar pattern in the arch towardperiendothelial staining in the PA.Bar5200 mm (c and d). e, Micrograph ofa stage-36 chimera shows the connec-tion between the mesodermal subclavianartery (SA) and QCPN-stained NC-de-rived brachiocephalic artery (BA).Bar5100 mm. f, Media and adventitia ofthe carotid arteries (CA) are stained withQCPN, indicating their NC origin (stage36). Bar5100 mm. g, QCPN-stained sec-tion of a stage-36 chimera clearly dem-onstrates that the anterior cardinal veins(ACV), like the great arteries, receive anextensive NCC contribution to the mediaas well. The adjacent vagal nerve (VN)harbors NC-derived satellite cells.Bar5100 mm.
228 Neural Crest and Vascular Differentiation
by guest on May 23, 2017
http://circres.ahajournals.org/D
ownloaded from

both vascular roots appeared to have a dual origin in whichmesodermal cells clearly outnumbered NCCs. Moreover,NCCs generally turned out to be spatially organized towardthe median (septal) side at the very bases, gradually substitutingmesodermal cells in a downstream direction; a somewhatsimilar phenomenon was described by Takamura et al.2 Thechick ascending aorta and pulmonary trunk, or at least theirvery bases, are generally hypothesized to originate from theseptated aortic sac. In contrast to designated derivatives of thearch arteries (aortic arch, brachiocephalic arteries, and pulmo-nary arch arteries), the aortic sac appears to develop mainlyfrom the mesoderm, incorporating only a minor NC contri-bution. Actual translation of these chick NCC patterns into thefully remodeled mammalian arterial tree remains difficult. Theaortic sac in mammals is considered to develop into the entireascending aorta, brachiocephalic artery, and pulmonarytrunk.9,47 However, the extension of the proximal cuff, whichwe found largely devoid of NCCs, is unknown in these majormammalian vessels. Unfortunately, long-term tracing studies,similar to those in avian models, cannot be performed inintrauterine mammalian development, and specific markers formesenchymal cells and SMCs derived from the NC, which areexpressed for a prolonged period of time, have not beenidentified yet.
Our present finding of the contribution of NCCs to themedia of the anterior cardinal veins is the first so far. Thisfinding was accomplished by use of both retrovirus andchimera experiments and strengthens the hypothesis that vas-cular NCC distribution is dictated by the presence of endo-thelial tubes at the early hindbrain level in both the arterial andvenous system. Unlike the outflow tract of the heart and thegreat arteries, however, the venous system was never describedto be affected after NC ablation.48 To date, however, thevenous system has received relatively little attention in vascularresearch, and our results suggest that it should be given moreconsideration in NC-related experimental and clinicalresearch.
Implications for Cellular Phenotype andVascular Morphology?Using retroviral NC tracing in combination with the HHF35-antibody, we have been able to show that NCCs are amongthe first cells to differentiate into primary SMCs in thedeveloping pharyngeal arterial system. This finding is incontradiction with earlier reports by Rosenquist and Beall.49
Using an anti–smooth muscle actin antibody (1A4) on chi-meric embryos and without the benefit of the QCPN anti-body, they concluded that primary actin expression in thearterial system occurred in mesodermal cells. This primarymesodermal tunica media was to be replaced at a later stage byactin-negative NCCs, which in turn expressed actin fromaround day 12 onward, resulting in a wave of secondary actinexpression. We have recently reported on similar actin expres-sion patterns22 but now reveal that NCCs are involved in bothprimary and secondary expression waves.
Adventitial fibroblasts as well as SMCs and nonmuscularcells of the media, which arise after the secondary actinexpression wave within the lamellar vessel wall,22,49 were in thepresent study shown to be NC-derived. NCCs did not show
preferential differentiation into any of these cell types. Al-though true analogs of avian nonmuscular cells do not appearto be present in mammalian vessels, the latter do harbor distinctSMC phenotypes, which are usually referred to as contractileand synthetic cells.50,51 Yablonka-Reuveni et al52 reported thatin chicks NCCs have the potential to differentiate into typicalSMCs, which express desmin and a-smooth muscle actin andare surrounded by a laminin and collagen IV basementmembrane, and into cells lacking those characteristics (non-muscular cells). Cellular phenotype in the pharyngeal archderivatives, therefore, appears not to be lineage-related.
On the basis of (immuno)histological examination only, werecently suggested that NC-derived thoracic arteries differfrom mesodermal arteries.22 In the present study, we confirmthis hypothesis. In general, it turned out that the typicallamellar vessel wall structure only developed in vessels thathave an NC contribution. Interestingly, the boundaries of NCareas tend to coincide with marked changes in vascularmorphology. At least three good examples of this phenomenonwere seen: (1) the junction between pulmonary arch artery(NC-derived) and pulmonary artery (mesoderm), (2) theproximal subclavian artery (NC)–distal subclavian artery (me-soderm), and (3) the junction of the fourth and sixth archarteries (NC) with the descending aorta (mesoderm).
Although these coinciding spatial boundaries are very sug-gestive of lineage-related morphogenesis, other findings partlycontradict this hypothesis. In both aortic and pulmonary roots,for instance, we demonstrated a spatially organized dual originof the vessel wall. However, the areas in these vessels that wereshown to be of mesodermal origin did not differ morpholog-ically from the NC areas nearby. An NC boundary, the exactposition of which could not be established by us, is certainlynot reflected in the histology of this part of the arterial tree,suggesting the major influence of other factors, such ashemodynamics. This area nonetheless remains very interestingbecause of matrix-related pathologies, which predominantlyaffect this region. Fibrillin-1–associated Marfan’s syndrome53,54
often presents dilatation or dissection of the ascending aorta,whereas the elastin-related Williams syndrome involves supra-valvular aortic stenosis.55
In addition, the sixth arch derivative, the DA, was shown toreceive an extensive NC contribution but did not join theother arch derivatives in their elastogenic differentiation.NC-derived arteries therefore can differentiate into both elasticand muscular (DA) vessels. Although the DA shares the sameorigin and presumably comparable hemodynamic and envi-ronmental factors with the other arch derivatives, its differen-tiation deviates rather early in development. Since NCCs carryHox gene–encoded positional information into the pharyngealarches,40,41 one could speculate on a further refinement oflineage-dependent morphogenesis, as cardiac crest cells differin Hox gene expression according to their original position onthe anteroposterior axis. However, cardiac NCCs originatingfrom different rhombomeric origins were shown to mix ratherextensively within the circumpharyngeal crest before finishingmigration into the forming pharyngeal arches.4,18 In this way,the sixth arch receives NCCs emanating from the somite 1 tosomite 3 level and therefore largely shares its cellular compo-sition with that of arch 4. Yet regardless of these similarities,
Bergwerff et al 229
by guest on May 23, 2017
http://circres.ahajournals.org/D
ownloaded from

only the sixth arch was shown to retain HOX 2.1 (HOXB5)expression,56 thereby emphasizing its unique identity.
To date, an increasing number of in vitro experiments favorthe hypothesis of lineage-related SMC differentiation. SMCsisolated from normally developed NC vessels were shown todiffer in vitro from “surrogate” SMCs that infest the arteriesafter experimental NC ablation. SMCs (of undeterminedorigin) that replaced the ablated NCCs showed much higherlevels of a-actin, tropoelastin, and expression of c-jun during invitro assays.57 In addition, in vitro experiments also elaboratedon differential responses to TGF-b1 by either thoracic orabdominal aortic SMCs. TGF-b was shown to be growthinhibitory when administered to abdominal (mesodermal)SMCs, whereas it increased DNA synthesis in SMCs isolatedfrom the thoracic (NC) aorta.24,58 Topouzis and Majesky24
suggested that this differential TGF-b responsiveness mightresult from lineage-dependent differences in glycosylation ofthe type II TGF-b receptor. However, the implications oftheir findings for vessel differentiation in vivo remain to beelucidated, since cellular morphology and protein expressionlevels of cultured cells, oddly enough, were similar in bothgroups. Actin expression in vivo, however, was shown to berestricted to only a subpopulation of media cells in theNC-derived aortic arch. All other cells were of a nonmuscularactin-negative phenotype.22,49,52 Nonetheless, differences inpotency unraveled by in vitro studies will aid evaluation ofmorphological and physiological heterogeneity among vesselsthat so far are considered to be similar. If SMC origin andtopographical localization indeed are of major importance indefining the characteristics and performance of the cell, thenthis should be taken into account in design and evaluation of invitro experiments as well as in surgical vessel grafting.
Last, origin-related SMC features may well be of signifi-cance in vascular pathogenesis. Intriguingly, spontaneous aorticplaques in the chicken were only found in the descending partof this vessel.59 The proximal part, showing the intricatecellular lamellar organization, which we now know is predom-inantly of NC origin, does not develop such arterioscleroticpathology in the adult chicken. Because a proper correlation ofhuman and chick patterning is still lacking and because thisphenomenon cannot yet be translated to human arteriosclero-sis-prone areas, more research into the NC in mammals will beneeded to elucidate its role in vascular biology.
AcknowledgmentsThis study was financially supported by grant 93.111 from theNetherlands Heart Foundation. The excellent technical assistance ofBenno Vrolijk and Marie-Joze Verdonk and graphic support by JanLens and Bas Blankevoort are greatly appreciated.
References1. Kirby ML, Gale TF, Stewart DE. Neural crest cells contribute to normal
aorticopulmonary septation. Science. 1983;220:1059–1061.2. Takamura K, Okishima T, Ohdo S, Hayakawa K, Okamoto N. Sequential
observation of cardiac neural crest cell distribution in the developing heart:effects of transplantation regions. In: Clark EB, Takao A, eds. DevelopmentalCardiology: Morphogenesis and Function. Mt Kisco, NY: Futura Press; 1990:159–173.
3. Le Lievre CS, Le Douarin NM. Mesenchymal derivatives of the neuralcrest: analysis of chimaeric quail and chick embryos. J Embryol Exp Morphol.1975;34:125–154.
4. Miyagawa-Tomita S, Waldo K, Tomita H, Kirby ML. Temporospatialstudy of the migration and distribution of cardiac neural crest in quail-chickchimeras. Am J Anat. 1991;192:79–88.
5. Waldo KL, Kirby ML. Cardiac neural crest contribution to the pulmonaryartery and sixth aortic arch artery complex in chick embryos aged 6 to 18days. Anat Rec. 1993;237:385–399.
6. Driscoll DA, Goldmuntz E, Emanuel BS. Detection of 22q11 deletions inpatients with conotruncal cardiac malformations, DiGeorge, velocardio-facial, and conotruncal anomaly face syndromes. In: Clark EB, MarkwaldRR, Takao A, eds. Developmental Mechanisms of Heart Disease. Armonk,NY: Futura Press; 1995:569–575.
7. Wilson JG, Roth CB, Warkany J. An analysis of the syndrome of malfor-mations induced by maternal vitamin A deficiency: effects of restoration ofvitamin A at various times during gestation. Am J Anat. 1953;92:189–217.
8. Bockman DE, Redmond ME, Kirby ML. Alteration of early vasculardevelopment after ablation of cranial neural crest. Anat Rec. 1989;225:209–217.
9. Kirby ML, Waldo KL. Neural crest and cardiovascular patterning. Circ Res.1995;77:211–215.
10. Hart RC, Ragland WL, Winn KJ, Unger ER. Avian model for 13-cis-retinoic acid embryopathy demonstration of neural crest related defects.Teratology. 1990;41:463–472.
11. Broekhuizen MLA, Wladimiroff JW, Tibboel D, Poelmann RE, WeninkACG, Gittenberger-de Groot AC. Induction of cardiac anomalies withall-trans retinoic acid in the chick embryo. Cardiol Young. 1992;2:311–317.
12. Pexieder T, Blanc O, Pelouch V, Ostadalova I, Milerova M, Ostadal B.Late fetal development of retinoic acid-induced transposition of greatarteries: morphology, physiology and biochemistry. In: Clark EB,Markwald RR, Takao A, eds. Developmental Mechanisms of Heart Disease.Armonk, NY: Futura Press; 1995:297–307.
13. Mendelsohn C, Lohnes D, Decimo D, Lufkin T, LeMeur M, Chambon P,Mark M. Function of the retinoic acid receptors (RARs) during devel-opment (II) Multiple abnormalities at various stages of organogenesis inRAR double mutants. Development. 1994;120:2749–2771.
14. Saint-Jeannet JP, Levi G, Girault JM, Koteliansky V, Thiery JP. Ventro-lateral regionalization of Xenopus laevis mesoderm is characterized by theexpression of a-smooth muscle actin. Development. 1992;115:1165–1173.
15. Hungerford JE, Owens GK, Argraves WS, Little CD. Development of theaortic vessel wall as defined by vascular smooth muscle and extracellularmatrix markers. Dev Biol. 1996;178:375–392.
16. DeRuiter MC, Poelmann RE, VanMunsteren JC, Mironov V, MarkwaldRR, Gittenberger-de Groot AC. Embryonic endothelial cells transdiffer-entiate into mesenchymal cells expressing smooth muscle actins in vivo andin vitro. Circ Res. 1997;80:444–451.
17. Lumsden A, Sprawson N, Graham A. Segmental origin and migration ofneural crest cells in the hindbrain region of the chick embryo. Development.1991;113:1281–1291.
18. Shigetani Y, Aizawa S, Kuratani S. Overlapping origins of pharyngeal archcrest cells on the postotic hind-brain. Dev Growth Differ. 1995;37:733–746.
19. Waldo KL, Kumiski D, Kirby ML. Cardiac neural crest is essential for thepersistence rather than the formation of an arch artery. Dev Dyn. 1996;205:281–292.
20. Fukiishi Y, Morriss-Kay GM. Migration of cranial neural crest cells to thepharyngeal arches and heart in rat embryos. Cell Tissue Res. 1992;268:1–8.
21. Osumi-Yamashita N, Ninomiya Y, Doi H, Eto K. Rhombomere for-mation and hind-brain crest cell migration from prorhombomeric originsin mouse embryos. Dev Growth Differ. 1996;38:107–118.
22. Bergwerff M, DeRuiter MC, Poelmann RE, Gittenberger-de Groot AC.Onset of elastogenesis and downregulation of smooth muscle actin asdistinguishing phenomena in artery differentiation in the chick embryo.Anat Embryol. 1996;194:545–557.
23. Thieszen SL, Dalton M, Gadson PF, Patterson E, Rosenquist TH.Embryonic lineage of vascular smooth muscle cells determines responses tocollagen matrices and integrin receptor expression. Exp Cell Res. 1996;227:135–145.
24. Topouzis S, Majesky MW. Smooth muscle lineage diversity in the chickembryo: two types of aortic smooth muscle cell differ in growth andreceptor-mediated transcriptional responses to transforming growthfactor-b. Dev Biol. 1996;178:430–445.
25. Gadson PF, Dalton M, Patterson E, Svoboda DD, Hutchinson L, SchramD, Rosenquist TH. Differential response of mesoderm- and neural crest-derived smooth muscle to TGFb1: regulation of c-myb and a1 (I) pro-collagen genes. Exp Cell Res. 1997;230:169–180.
230 Neural Crest and Vascular Differentiation
by guest on May 23, 2017
http://circres.ahajournals.org/D
ownloaded from

26. Mikawa T, Fischman DA, Dougherty JP, Brown AMC. In vivo analysis ofa new lacZ retrovirus vector suitable for cell lineage marking in avian andother species. Exp Cell Res. 1991;195:516–523.
27. Epstein ML, Mikawa T, Brown AMC, McFarlin DR. Mapping, the originof the avian enteric nervous system with a retroviral marker. Dev Dyn.1994;201:236–244.
28. Noden DM, Poelmann RE, Gittenberger-de Groot AC. Cell origins andtissue boundaries during outflow tract development. Trends Cardiovasc Med.1995;5:69–75.
29. Gittenberger-de Groot AC, Slomp J, DeRuiter MC, Poelmann RE.Smooth muscle cell differentiation during early development and duringintimal thickening formation in the ductus arteriosus. In: Schwartz SM,Mecham R, eds. The Vascular Smooth Muscle Cell. San Diego, Calif:Academic Press; 1995:17–36.
30. Mikawa T, Borisov A, Brown AMC, Fischman DA. Clonal analysis ofcardiac morphogenesis in the chicken embryo using a replication-defectiveretrovirus, I: formation of the ventricular myocardium. Dev Dyn. 1992;193:11–23.
31. Fekete DM, Perez-Miguelsanz J, Ryder EF, Cepko CL. Clonal analysis inthe chicken retina reveals tangential dispersion of clonally related cells. DevBiol. 1994;166:666–682.
32. Mikawa T, Gourdie RG. Pericardial mesoderm generates a population ofcoronary smooth muscle cells migrating into the heart along with ingrowthof the epicardial organ. Dev Biol. 1996;174:221–232.
33. Hamburger V, Hamilton HL. A series of normal stages in the developmentof the chick embryo. J Morphol. 1951;88:49–92.
34. Tsukada T, Tippens D, Gordon D, Ross R, Gown AM. HHF35, amuscle-actin-specific monoclonal antibody, I: immunocytochemical andbiochemical characterization. Am J Pathol. 1987;126:51–60.
35. Abo T, Balch CM. A differentiation antigen of human NK and K cellsidentified by a monoclonal antibody (HNK-1). J Immunol. 1981;127:1024–1029.
36. Romanoff AL. The Avian Embryo: Structural and Functional Development.New York, NY: Macmillan Publishing Co Inc; 1960.
37. Kuratani SC, Kirby ML. Initial migration and distribution of the cardiaccrest in the avian embryo: an introduction to the concept of the circum-pharyngeal crest. Am J Anat. 1991;191:215–227.
38. Serbedzija GN, Bronner-Fraser M, Fraser SE. Vital dye analysis of cranialneural crest cell migration in the mouse embryo. Development. 1992;116:297–307.
39. Le Douarin NM, Teillet MA. The migration of neural crest cells to the wallof the digestive tract in avian embryo. J Embryol Exp Morphol. 1973;30:31–48.
40. Hunt P, Gulisano M, Cook M, Sham M, Faiella A, Wilkinson D,Boncinelli E, Krumlauf R. A distinct Hox code for the branchial region ofthe head. Nature. 1991;353:861–864.
41. Krumlauf R. Hox genes and pattern formation in the branchial region ofthe vertebrate head. Trends Genet. 1993;9:106–112.
42. Kirby ML, Hunt P, Wallis K, Thorogood P. Abnormal patterning of theaortic arch arteries does not evoke cardiac malformations. Dev Dyn. 1997;208:34–47.
43. DeRuiter MC, Gittenberger-de Groot AC, Poelmann RE, van Iperen L,Mentink MMT. Development of the pharyngeal arch system related to the
pulmonary and bronchial vessels in the avian embryo. Circulation. 1993;87:1306–1319.
44. Kuratani SC, Kirby ML. Migration and distribution of circumpharyngealcrest cells in the chick embryo: formation of the circumpharyngeal ridgeand E/C81 crest cells in the vertebrate head region. Anat Rec. 1992;234:263–280.
45. Newgreen DF, Tan SS. Adhesion molecules in neural crest development.Pharmacol Ther. 1993;60:517–537.
46. Krug EL, Rezaee M, Isokawa K, Turner DK, Litke LL, Wunsch AM, BainJL, Riley DA, Capehart AA, Markwald RR. Transformation of cardiacendothelium into cushion mesenchyme is dependent on es/130: temporal,spatial, and functional studies in the early chick embryo. Cell Mol Biol Res.1995;41:263–277.
47. Netter FH; Yonkman FF, ed. The CIBA Collection of Medical Illustrations.Summit, NJ: CIBA; 1969.
48. Kirby ML, Waldo KL. Role of neural crest in congenital heart disease.Circulation. 1990;82:332–340.
49. Rosenquist TH, Beall AC. Elastogenic cells in the developing cardiovas-cular system: smooth muscle, nonmuscle, and cardiac neural crest. Ann N YAcad Sci. 1990;558:106–119.
50. Thyberg J, Hedin U, Sjolund M, Palmberg L, Bottger BA. Regulation ofdifferentiated properties and proliferation of arterial smooth muscle cells.Arteriosclerosis. 1990;10:966–990.
51. Campbell GR, Campbell JH. The phenotypes of smooth muscle expressedin human atheroma. Ann N Y Acad Sci. 1990;598:143–158.
52. Yablonka-Reuveni Z, Schwartz SM, Christ B. Development of chickenaortic smooth muscle: expression of cytoskeletal and basement membraneproteins defines two distinct cell phenotypes emerging from a commonlineage. Cell Mol Biol Res. 1995;41:241–249.
53. Dietz HC, Pyeritz RE. Mutations in the human gene for fibrillin-1 (FBN1)in the Marfan syndrome and related disorders. Hum Mol Genet. 1995;4:1799–1809.
54. Roberts WC, Honig HS. The spectrum of cardiovascular disease in theMarfan syndrome: a clinico-morphologic study of 18 necropsy patients andcomparison to 151 previously reported necropsy patients. Am Heart J.1982;104:115–134.
55. Curran ME, Atkinson DL, Ewart AK, Morris CA, Leppert MF, KeatingMT. The elastin gene is disrupted by a translocation associated withsupravalvular aortic stenosis. Cell. 1993;73:159–168.
56. Kuratani SC, Wall NA. Expression of Hox 2.1 protein in restricted pop-ulations of neural crest cells and pharyngeal ectoderm. Dev Dyn. 1992;195:15–28.
57. Gadson PF, Rossignol C, McCoy J, Rosenquist TH. Expression of elastin,smooth muscle alpha-actin, and c-Jun as a function of the embryoniclineage of vascular smooth muscle cells. In Vitro Cell Dev Biol. 1993;29A:773–781.
58. Wrenn RW, Raeuber CL, Herman LE, Walton WJ, Rosenquist TH.Transforming growth factor-beta: signal transduction via protein kinase Cin cultured embryonic vascular smooth muscle cells. In Vitro Cell Dev Biol.1993;29A:73–78.
59. Moss NS, Benditt EP. The ultrastructure of spontaneous and experi-mentally induced arterial lesions, II: the spontaneous plaque in the chicken.Lab Invest. 1970;23:231–245.
Bergwerff et al 231
by guest on May 23, 2017
http://circres.ahajournals.org/D
ownloaded from

C. Gittenberger-de-GrootMaarten Bergwerff, Marlies E. Verberne, Marco C. DeRuiter, Robert E. Poelmann and Adriana
Vascular Morphology?Neural Crest Cell Contribution to the Developing Circulatory System: Implications for
Print ISSN: 0009-7330. Online ISSN: 1524-4571 Copyright © 1998 American Heart Association, Inc. All rights reserved.is published by the American Heart Association, 7272 Greenville Avenue, Dallas, TX 75231Circulation Research
doi: 10.1161/01.RES.82.2.2211998;82:221-231Circ Res.
http://circres.ahajournals.org/content/82/2/221World Wide Web at:
The online version of this article, along with updated information and services, is located on the
http://circres.ahajournals.org//subscriptions/
is online at: Circulation Research Information about subscribing to Subscriptions:
http://www.lww.com/reprints Information about reprints can be found online at: Reprints:
document. Permissions and Rights Question and Answer about this process is available in the
located, click Request Permissions in the middle column of the Web page under Services. Further informationEditorial Office. Once the online version of the published article for which permission is being requested is
can be obtained via RightsLink, a service of the Copyright Clearance Center, not theCirculation Researchin Requests for permissions to reproduce figures, tables, or portions of articles originally publishedPermissions:
by guest on May 23, 2017
http://circres.ahajournals.org/D
ownloaded from













![A Stable Cranial Neural Crest Cell Line from Mouse · Neural crest cell culture Cranial neural crest cells labeled with Wnt1-Cre; R26R-GFP [7,11,12] were obtained from E8.5 mouse](https://img.pdfslide.us/doc/110x75/5f42417ff2821645233c9c4f/a-stable-cranial-neural-crest-cell-line-from-mouse-neural-crest-cell-culture-cranial.jpg)