Embed Size (px)
Citation preview

Neanderthals Revisited

Vertebrate Paleobiology and Paleoanthropology
Edited by
Eric Delson Vertebrate Paleontology, American Museum of Natural History,
New York, NY 10024, USA
Ross D. E. MacPheeVertebrate Zoology, American Museum of Natural History,
New York, NY 10024, USA
Focal topics for volumes in the series will include systematic paleontology of all vertebrates (from agnathans tohumans), phylogeny reconstruction, functional morphology, Paleolithic archaeology, taphonomy, geochronology,historical biogeography, and biostratigraphy. Other fields (e.g., paleoclimatology, paleoecology, ancient DNA,total organismal community structure) may be considered if the volume theme emphasizes paleobiology (orarchaeology). Fields such as modeling of physical processes, genetic methodology, nonvertebrates or neontologyare out of our scope.
Volumes in the series may either be monographic treatments (including unpublished but fully reviseddissertations) or edited collections, especially those focusing on problem- oriented issues, with multidisciplinarycoverage where possible.
Editorial Advisory Board
Nicholas Conard (University of Tübingen), John G. Fleagle (Stony Brook University), Jean-Jacques Hublin(Max Planck Institute for Evolutionary Anthropology), Sally McBrearty (University of Connecticut), Jin Meng(American Museum of Natural, History), Tom Plummer (Queens College/CUNY), Kristi Curry Rogers(Science Museum of Minnesota), Ken Rose (Johns Hopkins University).
Published and forthcoming titles in this series are listed at the end of this volume.

A Volume in
TheMax-Planck-InstituteSubseries in Human
EvolutionCoordinated by
Jean-Jacques HublinMax-Planck-Institute for Evolutionary Anthropology, Department of Human Evolution, Leipzig, Germany

Edited by
Max-Planck-Institute for Evolutionary Anthropology Leipzig, Germany
Department of Anthropology, New York UniversityNew York, USA
Neanderthals Revisited:
and PerspectivesNew Approaches
Katerina Harvati
Terry Harrison

A C.I.P. Catalogue record for this book is available from the Library of Congress.
ISBN-13 978-1-4020-5120-3 (HB)ISBN-13 978-1-4020-5121-0 (e-book)
Published by Springer,P.O. Box 17, 3300 AA Dordrecht, The Netherlands.
www.springer.com
Printed on acid-free paper
Cover image by Philipp Gunz, adapted from a CT scan image of La Ferrassie 1 by Jean-Jacques Hublin.
All Rights Reserved
No part of this work may be reproduced, stored in a retrieval system, or transmittedin any form or by any means, electronic, mechanical, photocopying, microfilming,
recording or otherwise, without written permission from the Publisher, with the exception of any material supplied specifically for the purpose of being entered
and executed on a computer system, for exclusive use by the purchaser of the work.
Reprinted with corrections 2008First edition published 2007
© 2008 Springer Science+Business Media B.V.

This volume is dedicated to the memory of W.W. Howells (1908–2005) for his remarkable and pioneering contributions to the study of human evolution, especially his role in the
greater understanding and appreciation of the Neanderthals. He was mentor and source of inspiration to generations of anthropologists, and his work continues to be a tremendous resource
for research in human variation and evolution.

Contents
Contributors xi
Preface xiii
1. Neanderthals revisited 1K. Harvati and T. Harrison
2. The distinctiveness and systematic context of Homo neanderthalensis 9I. Tattersall and J.H. Schwartz
3. Saccopastore 1: the earliest Neanderthal? A new look at an old cranium 23E. Bruner and G. Manzi
4. Inquiries into Neanderthal craniofacial development and evolution: “accretion” versus “organismic” models 37A. Rosas, M. Bastir, C. Martínez-Maza, A. García-Tabernero, and C. Lalueza-Fox
5. Neanderthals and modern humans – chimps and bonobos: similarities and differences in development and evolution 71M.S. Ponce de León and C.P.E. Zollikofer
6. Cranial growth models: heterochrony, heterotopy, and the kinematics of ontogeny 89C.P.E. Zollikofer and M.S. Ponce de León
7. Bioenergetic perspectives on Neanderthal thermoregulatory and activity budgets 113S.E. Churchill
8. How different were Neanderthals’ habitual activities? A comparative analysis with diverse groups of recent humans 135O.M. Pearson, R.M. Cordero, and A.M. Busby
9. Neanderthal hands in their proper perspective 157W.A. Niewoehner
10. Did Neanderthals make the Châtelperronian assemblage from La Grotte du Renne (Arcy-sur-Cure, France)? 191S.E. Bailey and J.-J. Hublin
11. The fate of European Neanderthals: results and perspectives from ancient DNA analyses 211D. Serre and S. Pääbo
12. Selection on mitochondrial DNA and the Neanderthal problem 221J. Hawks
13. Reliability of cranial morphology in reconstructing Neanderthal phylogeny 239K. Harvati and T.D. Weaver

14. Non-metric variation in recent humans as a model for understanding Neanderthal-early modern human differences: just how “unique” are Neanderthal unique traits 255J.C.M. Ahern
15. Earliest Upper Paleolithic crania from Mladec, Czech Republic, and the question of Neanderthal-modern continuity: metrical evidence from the fronto-facial region 269G. Bräuer, H. Broeg, and C.B. Stringer
16. Neanderthals and modern humans: an example of a mammalian syngameon? 281T.W. Holliday
17. Speciation by distance and temporal overlap: a new approach to understanding Neanderthal evolution 299J.-L. Voisin
18. The Neanderthal-H. sapiens interface in Eurasia 315C.B. Stringer
Index 325
CONTENTSx

J.C.M. AhernDepartment of AnthropologyUniversity of WyomingLaramie, Wyoming, [email protected]
S.E. BaileyCenter for the Study of Human OriginsDepartment of AnthropologyNew York University, New YorkNew York, [email protected]
M. BastirHull York Medical SchoolThe University of YorkHeslington, York, United [email protected]
G. BräuerAbteilung für HumanbiologieUniversität HamburgHamburg, [email protected]
H. BroegAbteilung für HumanbiologieUniversität HamburgHamburg, Germany
E. BrunerDipartimento di Biologia Animale e dell’UomoUniversità La SapienzaRome, [email protected]
A.M. BusbyDepartment of AnthropologyUniversity of New MexicoAlbuquerque, New Mexico, [email protected]
S.E. ChurchillDepartment of Biological Anthropology and AnatomyDuke UniversityDurham, North Carolina, [email protected]
R.M. CorderoDepartment of AnthropologyUniversity of New MexicoAlbuquerque, New Mexico, [email protected]
A. García-TaberneroDepartament de PaleobiologíaMuseo Nacional de Ciencias NaturalesMadrid, [email protected]
K. HarvatiAbteilung für HumanevolutionMax-Planck-Institut fürEvolutionäre AnthropologieLeipzig, [email protected]
T. HarrisonCenter for the Study of Human OriginsDepartment of AnthropologyNew York UniversityNew York, New York, [email protected]
J. HawksDepartment of AnthropologyUniversity of Wisconsin–MadisonMadison, Wisconsin, [email protected]
T.W. HollidayDepartment of AnthropologyTulane UniversityNew Orleans, Louisiana, [email protected]
J.-J. HublinAbteilung für-HumanevolutionMax-Planck-Institut fürEvolutionäre AnthropologieLeipzig, [email protected]
C. Lalueza-FoxDepartamento de Biologia AnimalUniversitat de BarcelonaBarcelona, [email protected]
G. ManziDipartimento di Biologia Animale e dell’UomoUniversità La SapienzaRome, [email protected]
Contributors

C. Martínez-MazaDepartament de PaleobiologíaMuseo Nacional de Ciencias NaturalesMadrid, [email protected]
W.A. NiewoehnerDepartment of Anthropology,California State University-San BernardinoSan Bernardino, California, [email protected]
S. PääboAbteilung für Evolutionare GenetikMax-Planck-Institut für Evolutionäre AnthropologieLeipzig, [email protected]
O.M. PearsonDepartment of AnthropologyUniversity of New MexicoAlbuquerque, New Mexico, [email protected]
M.S. Ponce de LeónAnthropologisches Institut und MuseumUniversität Zuerich-IrchelZurich, [email protected]
A. RosasDepartament de PaleobiologíaMuseo Nacional de Ciencias NaturalesMadrid, [email protected]
J.H. SchwartzDepartments of Anthropology and the History and Philosophy of ScienceUniversity of PittsburghPittsburgh, Pennsylvania, USAjhs�@pitt.edu
D. SerreMcGill University and Genome Quebec InnovationCenterMontreal, Quebec, [email protected]
C.B. StringerDepartment of PalaeontologyNatural History MuseumLondon, United Kingdomc.stringer@ nhm.ac.uk
I. TattersallDivision of AnthropologyAmerican Museum of Natural HistoryNew York, New York, [email protected]
J.-L. VoisinInstitut de Paléontologie HumaineParis, [email protected]
T.D. WeaverAbteilung für HumanevolutionMax-Planck-Institut für Evolutionäre AnthropologieLeipzig, [email protected]
C.P.E. ZollikoferAnthropologisches Institut und MuseumUniversität Zuerich-IrchelZurich, [email protected]
CONTRIBUTORSxii

Preface
xiii
The question of modern human origins andthe role of Neanderthals in human evolutionhave fascinated paleoanthropologists and thepublic alike for more than a century. Recentyears have witnessed important scientificbreakthroughs in these areas of investigationas a result of ancient DNA studies, the appli-cation of new imaging and analytical tools inthe study of morphology, and novel theoreticaland conceptual approaches in evolutionarybiology. These exciting and important devel-opments have transformed our understandingand appreciation of the paleobiology, environ-mental adaptations, evolutionary relation-ships, and extinction of the Neanderthals.Although a number of influential edited vol-umes have previously dealt with these ques-tions, there have been no recent compendia onthe subject that have allowed leading interna-tional scholars specializing in Neanderthalresearch to present their latest ideas and toexplore new methods and approaches to theselong-standing problems.
With the 150th anniversary of the discov-ery of the first Neanderthal specimen drawingnear, and while Katerina Harvati was still anAssistant Professor at the AnthropologyDepartment at New York University (NYU),we conceived of a conference that would pro-vide an opportunity for those scientists active-ly involved in all aspects of Neanderthalresearch to present their latest findings and todiscuss the implications of these advances forunderstanding the evolutionary history ofNeanderthals. The “Neanderthals Revisited:New Approaches and Perspectives” confer-ence was held at NYU on January 27–29,2005. An important aim was to uniteresearchers in the early stages of their careerswith more established authorities, therebyencouraging a fresh look at some enduringproblems using innovative new methods andperspectives. We invited some 35 scholars
from the USA, Europe and the Near East topresent their research or to act as discussants,most of who were able to attend. This editedvolume is the outcome of that conference, andit presents, in more detailed fashion, the cut-ting-edge research that was showcased in NewYork. From the close of the conference to sub-mitting the final manuscript to the Press hastaken just slightly more than one year, so weare confident that the collection of papersincluded in this volume present up-to-dateresearch and current ideas on Neanderthals.
The conference program included twodays of presentations and discussions open tothe academic community and to the public,followed by a half-day workshop restricted toconference participants. Contributionsranged from the re-evaluation of Neanderthaland modern human anatomy, inferredNeanderthal adaptations and habitual activi-ties, developmental patterns, phylogeneticrelationships, and Neanderthal extinction.Participants also applied new methods,including computer tomography, 3D geomet-ric morphometrics, experimental growthstudies, genetic and paleogenetic analyses,and presented new perspectives andapproaches, including dental analysis, cladis-tic methodologies, bioenergetics, and broadercomparative analyses. One of the mostremarkable aspects of this gathering was thatthe venue provided an opportunity forresearchers to present contrasting views onmodern human origins without the rancourthat has characterized much of this debate inpast years. At the close of the conferencethere was general agreement that we wereclose to reaching a consensus on the criticalissue of the Neanderthal-modern humanrelationship, while other key questions, suchas the relationship of development withmorphological change, the relationshipbetween genetics and morphology, and the

re-evaluation of presumed Neanderthal adap-tations, were given closer critical scrutiny.
We would like to thank all the contributorsto this volume for participating in the“Neanderthals Revisited” conference and forproviding us with uniformly outstandingpapers. We are also grateful to the invited dis-cussants at the conference: Susan Antón,Jean-Jacques Hublin, Clifford Jolly, GiorgioManzi and Milford Wolpoff, as well as AlisonBrooks and Randy White. They all providedinsightful comments and initiated lively dis-cussion during the meeting. Bob Franciscus,Yoel Rak, and Mark Stoneking participated inthe conference, but were unable to contributeto this volume due to other pressing profes-sional commitments. Special thanks go to JenLeClair, Jennie Tichenor, Myriam Haas, andAllison Cleveland for their invaluable help inorganizing the conference, creating the con-ference poster, and formatting the manu-scripts. Several NYU students graciouslyvolunteered their time to help with the smoothrunning of the conference, thereby ensuring aminimum of technical and organizational hic-cups. Among these, Tom Rein, Maja Seselj,Connie Fellmann, Tim Cavaretta, Joe Califf,Andres Link, Gisselle Garcia, Laura Gaydosh,Deena Emera, and Ilana Soloman deservespecial mention.
Finally, we are grateful to all the col-leagues, students, members of the press, andpublic at large who attended the conference inlarge numbers and showed their remarkableenthusiasm for all things Neanderthal.
Financial support for the conference wasprovided by the Center for the Study ofHuman Origins at NYU, the Department ofHuman Evolution of the Max-Planck-Institutefor Evolutionary Anthropology, and the Deanfor Social Sciences at NYU. Fred Myers,Richard Foley, Jean-Jacques Hublin and EricDelson were especially supportive and instru-mental in the realization of both the confer-ence and the volume. We are very grateful toSpringer and to the editors of the VertebratePaleobiology and Paleoanthropology series,Eric Delson and Ross MacPhee, for agreeingto publish this volume, and to all the col-leagues who devoted their time to reviewingthe manuscripts and providing helpful input.Finally, we owe the greatest gratitude to ourfamilies, and particularly to our spouses, Eliasand Terri, for their support and encouragementin this and all endeavors.
Katerina HarvatiTerry Harrison
March 2006
PREFACExiv

1. Neanderthals revisited
K. HARVATIMax-Planck-Institute for Evolutionary AnthropologyLeipzig [email protected]
T. HARRISONCenter for the Study of Human Origins,Department of AnthropologyNew York UniversityNew York, NY 10003, [email protected]
Keywords: Neanderthals, phylogeny, evolution, taxonomy, paleobiology, species
Abstract
Neanderthals are the best represented and most studied group in the fossil human record. The relatively large numberof Neanderthal fossils and their good preservation offers the possibility of robust inferences about their evolutionand paleobiology. Nevertheless, debate still continues on important issues, and this suggests that deeper theoreticaland methodological differences lie at the root of the lack of consensus. Such disagreements are not likely to beresolved by additional fossil findings, but rather require critical re-evaluation of the evidence at hand and theapplication of novel techniques and perspectives. This is the premise and main goal of this volume. The majordebates in Neanderthal research are re-examined with the use of innovative state-of-the art methods and excitingnew theoretical and conceptual approaches. The diverse contributions presented here offer fresh insights andadvances that move us closer to reaching a consensus.
As the contributions to this volume illustrate,the Neanderthals are the best represented,most comprehensively studied, and most thor-oughly understood group of fossil hominins.The wealth of specimens currently available tothe scientific community, including dozens ofrelatively complete crania and partial skele-tons from across a broad geographic range,affords scholars the opportunity to developwell-informed and robust inferences about
the anatomy, phylogenetic relationships, tax-onomy, and paleobiology of the Neanderthals.Equally importantly, we know a great dealabout their archaeology, paleoecology, paleo-environment, and zoogeography, all of whichoffer key evidence for interpreting theirpaleobiology in a broader environmen-tal, behavioral, and phylogenetic context.Paleoanthropologists studying earlier parts ofthe human fossil record are less fortunate,
1
K. Harvati and T. Harrison (eds.), Neanderthals Revisited: New Approaches and Perspectives, 1–8.© 2008 Springer.

having to work with taxa that are much morepoorly represented, and in some cases knownonly by a few fragmentary specimens. It iscertainly an enviable position to be in, one inwhich most vertebrate paleontologists, whouniversally lament the shortcomings of thefossil record as an impediment to resolvingkey problems, would be most content to findthemselves.
Nevertheless, despite the quality and weightof the evidence, there continue to be majordebates (that have lasted for 150 years) about anumber of contentious issues, especiallywhether or not Neanderthals should be includ-ed in the same species as anatomically modernhumans, and what is the precise phylogeneticrelationship between these two forms. Ourinability to agree on these fundamental ques-tions is a matter of serious concern for paleo-anthropologists: it leads to the inevitableconclusion that if we are unable to come to adecision about the nature of the relationshipbetween Neanderthals and modern humans,how can we have confidence in our ability toresolve relationships in the earlier, much morescanty, fossil human record. However, the lackof unanimity is unrelated to the quality of thematerial. It is more a consequence of deepertheoretical and conceptual issues that relate tohow different researchers analyze and interpretthe anatomical and genetic evidence, and to themanner in which these are ultimately situatedin the broader context of how biological sys-tems operate in the natural world. If this is thecase, then it will take some time before a con-sensus can be reached, regardless of theamount of fossil material available for study.One way forward is to explore new methodsand theoretical approaches in order to betterunderstand the paleobiology and phylogeneticrelationships of Neanderthals.
The main theme of this volume is to revisitthe major debates concerning the place ofNeanderthals in human evolution. How mor-phologically distinct are the Neanderthals frommodern humans, and what do these distinctions
mean in terms of their paleobiology and phy-logeny? How genetically distinct areNeanderthals from modern humans, and whatdoes this mean for interpreting the populationdynamics, taxonomy and phylogenetic struc-ture of Late Pleistocene hominins in Europe?Were Neanderthals and modern humans capa-ble of interbreeding, and can they be consideredthe same or different species? What were thepaleoenvironmental and paleoecological con-texts of Neanderthals, and how did this impacton their paleobiology, evolution, and extinc-tion? All of these issues are tackled head-on inthis volume. By presenting new evidence, usinginnovative and state-of-the art techniques andmethods, and exploring exciting new theoreti-cal and conceptual approaches, the contributorsgain fresh insights into these issues, and ulti-mately succeed in edging the debate closer to aconsensus. However, we leave it up to the rea-der to decide just how far we still have to go inorder to attain a satisfactory solution to some ofthese long-term problems.
As editors of this volume, our aim was toassemble a collection of papers written by lead-ing international researchers who have tackledmany of these important questions using a vari-etyofnovelapproaches.Equally importantly,ascan be discerned from the chapter titles and thecontent of this volume, we have also tried toaccommodate a diversity of opinions and per-spectives that reflect the plurality of viewpointsamong contemporary scholars. The range oftopics covered include phylogeny, taxonomy,speciation, development, lifeways and adapta-tion, population genetics, extinction, paleo-ecology and archaeology, while the methodsadopted include morphological analyses (i.e.,traditional comparative morphology, dentalanthropology, developmental biology, unilin-ear measurements, and three-dimensional geo-metric morphometrics), genetics (i.e., mtDNA,microsatellite data), experimental modeling,and computer imaging.
This volume is not organized in formal sec-tions, but rather it follows the logic of the
K. HARVATI & T. HARRISON2

general themes addressed by the contributors.It starts with the Middle-Late Pleistocenehuman fossil record and the evolution of theNeanderthal morphotype; continues with anexamination of Neanderthal and modernhuman ontogeny, bioenergetics, and paleobiol-ogy, and their implications for inferring behav-ior; followed by genetic perspectives onNeanderthals and the utility of mtDNA andcranial morphological data in reconstructingphylogeny, the possibility of Neanderthal-modern human interbreeding and its taxonom-ic implications; and concludes with a review ofthe factors that may have contributed to theextinction of the Neanderthals.
In the opening chapter, Tattersall andSchwartz (Chapter 2) review the abundantmorphological and genetic evidence support-ing the distinctiveness of Neanderthals frommodern humans, but also from earlier MiddlePleistocene hominins. To them, this evidenceclearly confirms the status of Neanderthals asa separate clade. Does this mean that they are adifferent species? This is a difficult, perhapseven impossible, question to settle, because asTattersall and Schwartz highlight, nature doesnot come neatly packaged and there are noabsolute criteria by which to recognize species,especially in the fossil record. Nevertheless,Tattersall and Schwartz make the crucialobservation (echoed by other authors in thevolume) that Neanderthals as a group consti-tute a clear-cut morphologically and historica-lly individuated entity, evidently equivalent tothose commonly recognized today as species.They further point out that the morphologicalvariability in the European Middle Pleistoceneis little understood, and provocatively proposethat several hominin clades might have beencontemporaries in Europe during this period.This chapter sets the stage for the subsequentdiscussion of both the taxonomic position ofNeanderthals and the tempo and mode of theirevolution.
The next two chapters address the appearanceof the Neanderthal morphotype and its evolution.
Bruner and Manzi (Chapter 3) reassess theSaccopastore 1 cranium, which is correlated withoxygen isotope stage (OIS) 5, and is commonlyconsidered to be an “early Neanderthal”. Eventhough the specimen has been known since 1929,Bruner and Manzi are able to gain new insightsinto the endocranial morphology of the specimenusing computer tomography. Their observationson cranial capacity, degree of pneumatization,and inner ear morphology support previousconclusions that this specimen exhibits aNeanderthal-like morphology despite its smallsize, thereby pinpointing the appearance of thismorphotype to at least 130–100 ka. The authorssuggest that the demographic impact of OIS 6was probably catalytic in the evolution of full-blown Neanderthal features through geneticdrift. Rosas, Bastir, Martínez-Maza, García-Tabernero and Lalueza-Fox (Chapter 4) proposethe “organismic model” for Neanderthal evolu-tion as an alternative hypothesis to the widelyaccepted “accretion model,” drawing insightfrom their work on the extensive Spanish Middleand Late Pleistocene material. The authors pos-tulate a two-phase evolutionary process in theEuropean Middle-Late Pleistocene fossil record.The first phase is proposed to involve an increasein body size, greater postcranial robusticity, andincreased midfacial prognathism. The secondphase in turn would represent a true speciationevent at about 300–250 ka, corresponding with amajor re-organization of cranial architecture inNeanderthals relative to their Middle Pleistoceneprecursors. Their hypotheses point to a promis-ing direction of research in the study of humanevolution in Europe in the Middle Pleistocene.
Chapters 5 and 6 compare Neanderthal andmodern human ontogeny from several differ-ent viewpoints. Ponce de León and Zollikofer(Chapter 5) obtain three-dimensional datafrom computer tomography scans and analyzethem using geometric morphometric methods,in order to compare the ontogenetic trajecto-ries of two sets of sister taxa: Neanderthalsand modern humans, as opposed to chim-panzees and bonobos. Their analysis indicates
NEANDERTHALS REVISITED 3

that the two human taxa share a common onto-genetic trajectory, but have different perinatalmorphologies resulting from differences inprenatal growth. The two species of Pan,although overall more similar in shape to eachother than the human groups, differ not onlyin the length of their ontogeny, but also in thedirection. As Ponce de León and Zollikoferobserve in their concluding comments, one ofthe most important findings of their study isthat “spatial and temporal differences ingrowth and development not only generatedistinct adult morphologies, but also give riseto taxon-specific life histories”. This willsurely be a very fruitful avenue of futureresearch that will dramatically improve ourunderstanding of the phylogenetic relation-ships and paleobiology of fossil hominins. Inthe following chapter, Zollikofer and Ponce deLeón (Chapter 6) use computer modeling oftheir 3-D data to simulate cranial growthunder diverse conditions. This approachallows them to explore the ways in which asimple developmental system can be modifiedto produce different outcomes. Their resultsdemonstrate the complexity of developmentalprocesses, with intricate patterns potentiallyarising from simple changes and vice versa.The differences in the developmental patternbetween Neanderthals and modern humanssuggest that a change in the initial conditionsmay result in subsequent differences in theirdevelopmental trajectories.
Various aspects of Neanderthal anatomy,and their implications for understandingNeanderthal behavior, are explored in the nextfour chapters. Churchill (Chapter 7) applies abioenergetics approach, coupled with experi-mental modeling of the Neanderthal bodyform. His innovative analysis indicates that thecapacious Neanderthal ribcage may have beenrelated to heat production, rather than to heatretention, as is commonly postulated underBergmann’s rule. Churchill’s results also sug-gest a very high-caloric diet, with importantimplications for Neanderthal hunting abilities,
ranging behavior, and demographics. Eventhough Neanderthals appear to have had bodiesbetter adapted to generate and conserve heatthan early modern Europeans and modern-daycold-adapted populations, the finding thatNeanderthals occupied sites with warmer win-ter temperatures than early modern humans,suggests that they were less able to tolerateextreme glacial conditions. This may reflect agreater capability by early modern humans tocapture sufficient calories for sustaining ade-quate heat generation or the use of clothing orshelters with higher insulative values. Pearson,Cordero and Busby (Chapter 8) re-assessNeanderthal habitual activities, commonlythought (based on anatomical differences) todiffer markedly from those of modern humansin their extreme activity levels and foraginginefficiency. They compare Neanderthal upperand lower limb robusticity to those from seve-ral recent human foraging groups, and con-clude that Neanderthals do not appear unique,but instead are quite similar to modern forag-ing peoples that exploit limited territories. Theauthors conclude that these results, far fromindicating foraging inefficiency, may insteadimply a more intensive form of foraging.
Niewoehner (Chapter 9) uses three-dimensional geometric morphometric methodsto evaluate Neanderthal hand morphologycompared to that of Early and Late UpperPaleolithic modern humans. He relates theobserved shape differences to differences ininferred habitual grip positions, possibly sug-gesting differences in hafting technology andpreference for wood as a raw material. In con-trast, Upper Paleolithic human hand morphol-ogy is consistent with the archaeologicallyobserved expansion of the technological reper-toire that would have required increasedemphasis on precision handling and shifts inmanipulative postures. His results suggest agradual transition in hand morphology fromthe Middle to the Late Paleolithic. Baileyand Hublin (Chapter 10) re-examine the iso-lated dental remains associated with the
K. HARVATI & T. HARRISON4

Châtelperronian levels of the Grotte du Renne(Arcy-sur-Cure) site in France. These were pre-viously considered taxonomically unidentifi-able, but using a new dental scoring methoddeveloped by Bailey, the authors are able toestablish the Neanderthal identity of the dentalassemblage from this site. Their findings sub-stantiate earlier inferences that Neanderthalsare associated with the Châtelperronian indus-try in Western Europe, and that they were mostlikely the makers of these archaeologicalassemblages. Equally importantly, the recogni-tion that isolated teeth from Late Pleistocenesites can be identified taxonomically opens upthe possibility of investigating the mode andtempo of human evolution in Europe with muchbetter sampling and a finer-grained temporalresolution than was previously possible.
Chapters eleven and twelve consider aspectsof Neanderthal and modern human genetics.Serre and Pääbo (Chapter 11) present a newmethod for ancient DNA recovery. Thisresolves the problem of contamination by mod-ern human DNA, which leads to the inability todetect modern-human like genetic materialfrom fossil humans. Among the specimensexamined under this protocol, all Neanderthalsyielded Neanderthal-like mtDNA sequences,while all early modern Europeans yielded onlymodern human like mtDNA. The authors inter-pret their findings as indicating a minimaldegree of possible Neanderthal contribution tothe modern human gene pool. They also showthat major demographic changes occurred inLate Pleistocene mammal species that coincidetemporally with the extinction of Neanderthals.Such analyses highlight the importance of thestudy of population history for understandingNeanderthal evolution, and for providingimportant clues as to the timing and causes oftheir extinction. In the next chapter, Hawks(Chapter 12) critically re-examines the conclu-sions derived from mtDNA evidence aboutthe phylogenetic relationships betweenNeanderthals and modern humans. In particu-lar, he questions whether previous models
predicated on the assumption of selective neu-trality are valid, and proposes an alternativehypothesis that human mtDNA may have recent-ly undergone a “selective sweep,” possibly relat-ed to climate adaptation. Hawks suggests that itwas positive selection rather than populationreplacement that explains the disappearanceof archaic mtDNA variants. In this case, theobserved differences between the mtDNA inNeanderthals and modern humans would berendered phylogenetically uninformative.
The relationship between genetics and mor-phology in modern humans is explored furtherin chapter thirteen. Harvati and Weaver(Chapter 13) evaluate the usefulness of differ-ent cranial regions (i.e., face, vault, and tempo-ral bone) in reconstructing the phylogeneticplacement of Neanderthals. They assess thedegree to which morphological differences(represented by three-dimensional geometricmorphometric data) among recent human pop-ulations correspond to known neutral geneticdifferences (as represented by microsatellitedata) and/or to climatic differences. Althoughfacial morphology alone shows a relationshipwith climate, both vault and temporal bonemorphology track neutral genetics, with thetemporal bone tracking older events more suc-cessfully. The authors conclude that temporalbone morphology may be most appropriate forreconstructing the phylogeny of Neanderthalsand early modern humans. Their analysis doesnot support a unique phylogenetic linkbetween Neanderthals and early modernEuropeans.
The issue of Neanderthal-modern humanrelationships, and the possibility of inter-breeding between these populations, is takenup in greater detail in chapters fourteen andfifteen, with conflicting opinions expressed.Ahern (Chapter 14) addresses the question ofwhether Neanderthals and Upper PaleolithicEuropeans differ in a significantly greaternumber of distinct morphological traits thando two modern human populations: a “replac-ing” (European Americans) and “replaced”
NEANDERTHALS REVISITED 5

(Native Americans) group. The author isunable to reject the hypothesis thatNeanderthals and modern humans were con-specific for most of the features used, althoughhe does add a note of caution by acknowledg-ing that additional traits or combinations offeatures might eventually falsify the singlemorphospecies hypothesis. Bräuer, Broeg andStringer (Chapter 15) address the same ques-tion by re-examining the most complete craniafrom Mladec in the Czech Republic. These areamong the earliest modern European speci-mens known and they have often been sug-gested to exhibit Neanderthal-like features.However, the univariate and multivariate sta-tistical analyses of frontal bone metric datapresented by Bräuer and his colleagues offerno support for the claim that the Mladec indi-viduals might represent hybrids.
Further discussion of the concepts of specia-tion and interbreeding is explored by Hollidayand Voisin in the next two chapters. These con-tributions draw on studies of other vertebrates toreframe the species question in a broader com-parative perspective. Holliday (Chapter 16)reviews the literature on hybridization amongmammalspecies,andapplies it to theNeanderthal-early modern human case. He demonstrates thatamong interbreeding mammal species, thosethat have diverged as recently as these twohuman taxa are still able to produce fertilehybrids. He concludes from this evidence thatNeanderthal-modern human hybridization waspossible in all likelihood, even though there is noevidence from the genetic data or the fossilrecord to confirm that it actually took place.Building on ideas and concepts developed inprevious work by Clifford Jolly, Holliday arguesthat Neanderthals and modern humans areperhaps best considered “allotaxa,” good mor-phological species that may still have been ableto interbreed. He invokes the concept of syn-gameon – that closely related interbreedingspecies can be grouped into larger taxa – as auseful model for interpreting Late PleistoceneEuropean hominins. As a flip side to this study,
it would be interesting to examine how manynon-reproducing pairs of large mammal specieshave diverged in the past 700 ka. We suspect thatit would be very few; a finding that would serveto underscore just how rapidly Neanderthals andmodern humans diverged. Neanderthals are evi-dently autapomorphic, but it needs to berecalled that much of the distinction betweenNeanderthals and modern humans stem fromthe highly autapomorphic nature of modernhumans.
Voisin (Chapter 17) extends this approachby adding a broader comparative dimensionusing recent models of speciation in birds. Inparticular, he focuses on the ring speciesPhylloscopus trochiloides (greenish warbler)from central Asia as an analogy for Europeanhominin migration and evolution. His model ofspeciation by distance and temporal overlap isoffered to explain the East-West morphologi-cal cline observed in Neanderthals, as well asthe possible intermediate morphology of someCentral European Upper Paleolithic modernhumans. Voisin suggests that modern humansarriving in Eurasia during the Late Pleistocenewere able to interbreed to some degree with theless derived Neanderthals in Western Asia andCentral Europe, but hybridization was not pos-sible or was extremely limited with the morehighly derived Neanderthal populations at thefurthest extreme of the morphological cline inWestern Europe. In other words, Neanderthalsand modern humans behaved as two distinctbiological species in Western Europe, but notelsewhere.
Finally, Stringer (Chapter 18) presents newdata on the chronology of the appearance ofmodern humans in Europe and the extinction ofthe Neanderthals. Current paleoclimatic evi-dence indicates a much harsher and increas-ingly unstable climate during the period ofNeanderthal extinction (OIS 3). Stringer showsthat the greatest climatic stress would have beenexperienced around 30 ka, and he argues that thisstress probably played a key role in the demiseof the Neanderthals. Stringer’s contribution
K. HARVATI & T. HARRISON6

highlights the challenge of integrating thediverse kinds of data that are becoming incr-easingly available to fully test hypotheses ofNeanderthal and early modern humanpopulation histories.
The contributors to this volume provideimportant new insights that help us to betterappreciate and understand the evolution andpaleobiology of the Neanderthals. Theseadvances have been brought about not throughthe discovery of startling new fossil finds, butthrough the application of exciting new methods
and technologies, and a critical questioning ofestablished theoretical and conceptual para-digms.This is certainly the way forward, and weecho Chris Stringer’s concluding remarks thatwe undoubtedly have some exciting years aheadof us in Neanderthal and modern human originsresearch. Finally, although this volume is devo-ted entirely to the study of Neanderthals, itshould not be overlooked that it is through com-parisons with them that we are able to recognizeand reflect on the uniqueness and remarkablepeculiarities of our own evolutionary history.
NEANDERTHALS REVISITED 7

2. The distinctiveness and systematic context of Homo neanderthalensis
I. TATTERSALLDivision of AnthropologyAmerican Museum of Natural HistoryNew York, NY 10024, [email protected]
J.H. SCHWARTZDepartments of Anthropology and the History and Philosophy of ScienceUniversity of PittsburghPittsburgh, PA 15260, USAjhs�@pitt.edu
Keywords: Homo neanderthalensis, Neanderthals, Homo heidelbergensis, Mauer, Arago, Atapuerca, Sima de losHuesos, reconstructed skeleton
Abstract
The “packaging” of the diverse living world is untidy, with the result that there are no absolute criteria for recog-nizing in all contexts the bounded historical entities we call species. However, there is no doubt whatsoever thatHomo neanderthalensis is as clear-cut a morphological entity as any in the hominid fossil record: one that is char-acterized by a whole host of cranial apomorphies. Further, a recent full-skeleton reconstruction further emphasizesjust how different Neanderthal body structure was from that of Homo sapiens, not simply in numerous anatomi-cal details, but in the proportions of the thorax and its relation to the pelvic region. These bodily proportions wouldhave given these extinct hominids a very distinctive appearance on the landscape, and enhance the likelihood thatwe are dealing here with a reproductively differentiated entity. Still, Homo neanderthalensis is not unique in allthose features that distinguish it from Homo sapiens. Many “Neanderthal” cranial features are shared with vari-ous middle Pleistocene European hominids, notably the Steinheim specimen and, to a lesser extent, the Sima delos Huesos hominids from Atapuerca. Indeed, it appears that, far from being an isolated phenomenon, Homo nean-derthalensis formed part of a larger endemic European hominid clade. This clade seems to have existed contem-poraneously in Europe with at least one other hominid lineage or clade, exemplified by the Homo heidelbergensisfossils from Mauer, Arago and Petralona.
Introduction
Over the century and a half since the firstdescription of a Neanderthal fossil, an impres-sive record of these extinct hominids hasaccumulated. Indeed, not only has the mor-
phological distinctiveness of Homo nean-derthalensis for long been vastly better docu-mented than that of any other of our fossilrelatives (Tattersall, 1986), but we are also in abetter position than in the case of any otherextinct hominid to appreciate the morphological
9
K. Harvati and T. Harrison (eds.), Neanderthals Revisited: New Approaches and Perspectives, 9–22.© 2008 Springer.

variations (around a very distinctive mean) thatare shown by the various populations of thisform over time and space (Tattersall andSchwartz, 2000). Yet many paleoanthropolo-gists continue to equivocate over the questionof whether the Neanderthals actually constitutea bounded historical entity (Ghiselin, 1974) ofthe kind that warrants recognition as a species.Since this problem appears to be related, at leastpartly, to more general difficulties of speciesdefinition and recognition, it seems appropri-ate to begin our discussion of the status of theNeanderthals with a brief consideration ofthe nature of the boundaries that exist in theliving world.
Species as Bounded Historical Entities
It must be very clear to anyone concerned withthe luxuriant variety of living organisms that atsome level Nature is “packaged.” The biosphereis composed of a mass of discrete (but nested)units. At higher taxonomic levels there is noproblem distinguishing these units: all horsesare distinct from all whales by any definition.But as we approach finer degrees of distinction,particularly at intrageneric levels, difficultiesproliferate. These difficulties are reflected inthe extraordinary plethora of definitions of thespecies, by practice and by common consent thebasic systematic unit, that is currently on offer.As Hey (2001) observed, literally dozens of newsuch definitions have been proposed in recentdecades. This is not the place to trawl yet againthrough this lengthening list, but perhaps it isappropriate to point out that it is vanishinglyunlikely that any single definition of the term“species” will ever fit all cases. This is not onlybecause any universal definition would have tofit both living and extinct species, which offer usdifferent information sets; it is also because spe-ciation, the process by which individuated, non-reticulating units come about, is not a unitarymechanism. It is not, for example, simply an
inevitable, passive, consequence of themorphological differentiation of populationsover time (though this routine if poorly under-stood process certainly furnishes the basis forthe morphological differences by which speciesmay often be distinguished). Instead, speciationis a result (individuation, expressed most essen-tially among living populations as reproductiveindependence, but always seen a posteriori:Tattersall, 1994), which may eventuate fromshifts in developmental regulation at many dif-
Human beings are instinctively reductionistcreatures; but for all these reasons, and more,we may be unrealistic in expecting Nature to beneatly packaged. Ultimately, the boundariesdefining historically (evolutionarily) independ-ent units must lie in their (effective or absolute)reproductive isolation. But even reproductivebehaviors may not provide us with a goldenbullet. The studies of Clifford Jolly and his col-leagues (e.g. Jolly, 2001) have shown that evi-dence of quite extensive hybridization betweenadjacent populations of baboons that are some-times well differentiated to the eye may often bereadily observed; yet evidence is still lackingthat such behaviors are necessarily associatedwith the progressive integration of what evi-dently continue to be distinctive gene pools.Jolly (2001: 17) has, indeed, penetratinglyobserved that baboon allotaxa may at one andthe same time be “ ‘phylogenetic’ species, but‘biological’ subspecies.” And if this is truly thecase, from a historical (evolutionary) perspec-tive morphological differentiation becomesmuch the most significant factor to consider.Still, this hardly simplifies matters much. For,from the phenotypic standpoint, remarkableamounts of geographically (or artificially)maintained morphological variety may accu-mulate within a species without the disruptionof reproductive continuity – although, at thesame time, the latter can occur in the absence ofreadily detectable phenotypic change. Indeed,Schwartz (1999) has emphasized that there are
I. TATTERSALL & J.H. SCHWARTZ10
ferent levels (Schwartz, 2005).

no biological reasons whatever for expectingthat morphological change, deriving ultimatelyfrom changes in communication at the molecu-lar level, should be associated with reproduc-tive incompatibility unless it specificallyaffects such factors as protein recognitionbetween sperm and ova, gametic or zygotic via-bility, or reproductive organ morphology.Nonetheless, in the fossil record morphologicalcharacteristics are almost invariably all we haveto go on.
Given these awkward realities, we have toaccept the unfortunate fact that even amongsympatric or parapatric living forms, whosenatural behaviors we can directly observe, itwill sometimes never be certain whether twoclose relatives are “biologically” specificallydistinct or not. And the difficulties only mul-tiply when fossils are involved, for no specifi-able or quantifiable degree of morphologicaldifferentiation can be associated with specia-tion; and in almost all cases fossils can offerus only morphology as a basis for makingspecies judgments. It might be argued that theNeanderthals are actually a partial exceptionto this, given that fragments of their (distinc-tive) mitochondrial genome are now available(Krings et al., 1997, 2000; Ovchinnikov et al.,2000; Schmitz et al., 2002); but while thelarge mtDNA differences between availableNeanderthal samples and all living Homosapiens populations tested are indeed stronglysuggestive, it nonetheless remains true thatDNA differences suffer from analogous limi-tations to morphology in the context of assess-ing “species” status.
How, then, do we determine whether twoclearly related forms represent independent(individual) entities? It has recently becomefashionable to argue that species are mostusefully or practically equated with “basal(smallest) clusters of diagnosably distinct pop-ulations” (Cracraft, 2002: 130). But while thisapproach may have its possibly Siren attrac-tions for the working systematist, it still leaves
paleontologists in a quandary. Because whilesystematists studying living populations willoften already know from direct evidence whatthe geographical boundaries (and the morpho-logical variation occurring within those bound-aries) of their populations happen to be, thepaleontologist has to work backwards, inferen-tially, from morphology alone. Moreover, dis-tinctive morphologies frequently characterizegeographic variants that are not reproductively/historically individuated. Given this constraint,it would appear appropriate for paleontologiststo exercise caution, and to err on the side ofinclusivity when using morphological differ-entiation as a basis for inferring the existenceof reproductively and historically independententities. Where, then, do the Neanderthals fitwithin this perspective?
Homo neanderthalensis as aMorphologically and Historically Individuated Entity
There has been remarkably little debate overwhich hominid fossils are Neanderthal or notNeanderthal (as witnessed, for example, by theremarkable unanimity on this question, at least,among the very heterogeneous assortment ofcontributors to the classic volume edited bySmith and Spencer [1984]). Only at the fringeshas there been any significant discussion, butthis has usually focused upon whether or not aparticular fossil was somehow related to theNeanderthals, as a precursor or otherwise, ratherthan on the matter of its inclusion within thegroup itself (see historical account inTattersall,1995). This is largely because, among allextinct hominid entities, the Neanderthalgroup appears by far the most clearly delimit-ed morphologically. Instructively, of all otherfossil hominids only Homo erectus is compara-bly well represented in the record, yet no end isin sight to the debate over which specimensshould or should not be attributed to this
SYSTEMATIC OF NEANDERTHALS 11

species (compare, e.g., Wolpoff et al. [1994] toSchwartz and Tattersall [2000] and Antón[2003]).
Traditional cranial characters that are com-monly cited as typical of Neanderthals (see e.g.,Hublin, 1978, 1998; Santa Luca, 1978;Vandermeersch, 1981; Stringer et al., 1984;Schwartz and Tattersall, 1996a, b, 2005; Rakand Hylander, 2003) are numerous and include:double-arched supraorbital ridges whose sur-faces roll smoothly upward from the orbitalroofs and onto the frontal squama; orbits thatare obliquely truncated inferomedially; a nar-row lower face and a sharply retreatingmidface; medial projections emerging above aspinoturbinal crest that delineates a prenasalfossa lying just within the very large nasal aper-ture; very long and typically thin zygomaticarches; a “puffy” midface that reflects the pres-ence internally of expanded maxillary sinusesthat swell out the infraorbital and medial orbitalregions; an angulation along the anteriorsquamosal suture, delineating distinct anteriorand posterior temporal fossae; a smoothlyrounded (“en bombe”) cranial profile in rearview; a pitted suprainiac fossa that lies above asuperior nuchal line that is undercut by thenuchal plane but poorly delineated above; along and more or less straight parietomastoidsuture that flows directly behind into an anteri-or lambdoid suture; widespread pneumatizationwithin the petrosal; incomplete ossification ofthe ectotympanic tube laterally; and a long, nar-row, ovoid foramen magnum. A useful tablepublished by Hublin (1998, Table 1) lists sever-al others in addition. A particularly interestingnew source of cranial information has been pro-vided by CT studies (e.g., Hublin et al., 1996;Spoor et al., 2003) showing that the bonylabyrinth of the inner ear in Neanderthals is dis-tinguished in numerous derived features fromthat of Homo sapiens (and of Homo erectus). Inthe most comprehensive such study to date,Spoor et al. (2003: 141) suggested that suchdifferences reflected a distinctive pattern of
head movements possibly related to “aspects oflocomotor behaviour and the kinematic proper-ties of their head and neck.”
In the Neanderthal mandible are seen retro-molar spaces; sigmoid notches that are deep-est posteriorly, in front of low-set condyles,and sigmoid notch crests that terminate med-ial to the lateral extremities of the condyles;obliquely truncated gonial angles; symphysealbone that, when viewed from below, is thinnerfrom side to side than the bone distal to it. Inthe dentition the molars have relatively com-plex occlusal surfaces, with centroconids andcentrocones present on the lower and uppermolars respectively, and distinct talonid andtrigonid basins in the relatively long and nar-row lower molars. The molar occlusal surfacesare well defined peripherally by blunt crests,and are constricted in area by their inwardlysloping sides. Additional apomorphies of theNeanderthal upper and lower cheek dentitionshave recently been cited by Bailey (2002,2004) and Bailey and Lynch (2005).
Unsurprisingly, not all these and other cran-iodental characters typical of Neanderthals areequally strongly expressed in all unarguablyNeanderthal specimens that preserve them;and indeed it is evident that, in any widely dis-tributed group that is as close-knit phylogenet-ically as the genus Homo, there is bound to besome overlap among differentiated popula-tions in the frequency and extent of expression,as well as in the presence/absence of particulartraits. Thus, among the large assemblage ofunquestionably Neanderthal fossils individu-als vary, sometimes substantially, in featuressuch as the degree of bunning of the occiput,the size of the suprainiac fossa, the length ofthe retromolar space, the presence or absenceof a distinct postorbital plane behind thebrow ridges, the prominence of the medialprojections in the nasal fossa, the extent ofcheektooth taurodontism, the depth of thezygomatic arches, and so forth. But it isabundantly clear – and almost universally
I. TATTERSALL & J.H. SCHWARTZ12

acknowledged – that, despite their manifestindividual and geographic variation, theNeanderthals represent an unusually coherentand readily recognizable group. Certainauthors (e.g., Wolpoff, 1980; Frayer, 1984)have suggested that the relatively restrainedexpression of some of these characters in cer-tain Neanderthals of late date indicates adegree of intermediacy with Homo sapiens,but there are numerous reasons for rejectingthis notion (Tattersall and Schwartz, 2000).
Still, as impressive as the list of craniodentalapomorphies of the Neanderthals undoubtedly
is, the extent to which these hominids are dis-tinguished morphologically from Homo sapi-ens is made particularly evident by the full-skeleton reconstruction (Figures 1, 2) recentlyreported by Sawyer and Maley (2005).Reconstituted from the remains of a half-dozenpartial skeletons from four countries, this post-cranial reconstruction draws attention to theclassically Neanderthal characteristics of thepostcranial skeleton that have already beenexhaustively documented by numerous authors(e.g., Boule, 1911–1913; McCown and Keith,1939; Straus and Cave, 1957; Trinkaus, 1983).
SYSTEMATIC OF NEANDERTHALS 13
Figure 1. Front and lateral views of a complete Neanderthal skeleton as reconstructed, using elements from five partial skeletons (principally La Ferrassie 1 and Kebara 2), by
G. J. Sawyer and B. Maley. Photo courtesy of Ken Mowbray.
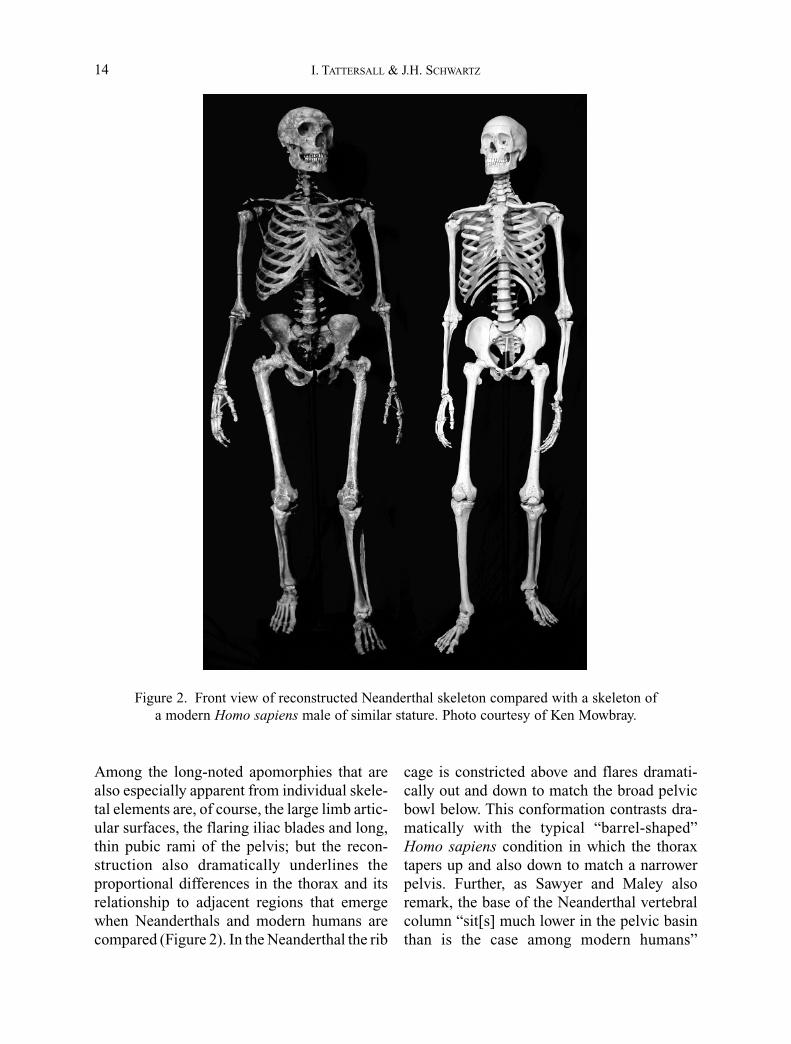
Among the long-noted apomorphies that arealso especially apparent from individual skele-tal elements are, of course, the large limb artic-ular surfaces, the flaring iliac blades and long,thin pubic rami of the pelvis; but the recon-struction also dramatically underlines theproportional differences in the thorax and itsrelationship to adjacent regions that emergewhen Neanderthals and modern humans arecompared (Figure 2). In the Neanderthal the rib
cage is constricted above and flares dramati-cally out and down to match the broad pelvicbowl below. This conformation contrasts dra-matically with the typical “barrel-shaped”Homo sapiens condition in which the thoraxtapers up and also down to match a narrowerpelvis. Further, as Sawyer and Maley alsoremark, the base of the Neanderthal vertebralcolumn “sit[s] much lower in the pelvic basinthan is the case among modern humans”
I. TATTERSALL & J.H. SCHWARTZ14
Figure 2. Front view of reconstructed Neanderthal skeleton compared with a skeleton of a modern Homo sapiens male of similar stature. Photo courtesy of Ken Mowbray.

(Sawyer and Maley, 2005: 30), contributing toan extreme shortness of the waist in the formerthat, in limiting thoracic torsion, would havehad significant consequences for gait, as wellas for appearance. Altogether, this reconstruc-tion makes it abundantly evident that in life theNeanderthals would have cut a very differentfigure on the landscape from Homo sapiens.
This major Gestalt difference adds, ofcourse, to the existing morphological basis forsurmising that the two kinds of hominid weresignificantly differentiated reproductivelyas well as morphologically. And, while notdemonstrating this conclusively, from the per-spective of “inclusive,” rather than “exclusive,”species concepts it also adds weight to theinference that specific mate recognitionsystems would have differed significantlybetween the two. This does not, of course,eliminate the possibility that occasionalinstances of coupling might have occurredwhen the two kinds of hominids came intocontact; but it does very strongly buttress thealready substantial reasons furnished by thefossil record for inferring that no significantintegration of the two populations ever tookplace. Overwhemingly, then, the probabilitymust be that the two kinds of hominid,Neanderthal and modern, were/are independ-ent historical as well as morphological entities.In which case, we are fully justified, undervirtually any set of criteria, in regarding theformer as constituting the species Homo nean-derthalensis (see also Harvati et al., 2004).
Neanderthals in Wider Systematic Context
The morphological distinctiveness of Homoneanderthalensis is hardly surprising whenone considers that both molecular (Kringset al., 1997, 2000) and paleontological (e.g.,Stringer and Gamble, 1993; Hublin, 1998;Tattersall and Schwartz, 2000) studies suggestthat this species last shared a common ancestor
with Homo sapiens at least a half-million yearsago. For close to a century now paleoanthro-pologists have sought the roots of theNeanderthals far back in time, and in one guiseor another many have discerned evidence inEurope for what Hublin (1998: 297) hasrecently termed a “Neandertalization Process,”linking forms from the early MiddlePleistocene (Mauer, Tautavel, etc.) to the latest(stages 5–3) “classical” Neanderthals, via“Holstein-Hoxnian” (Bilzingsleben, Sima delos Huesos, etc.) and “Saalian” (Biache-Saint-Vaast, Ehringsdorf, etc.) intermediates. Still, ifHomo neanderthalensis really was the endproduct of a steady course of phyletic modifi-cation, as this notion of a process implies, whatis perhaps most surprising is how clearly themorphological boundaries of the speciesHomo neanderthalensis actually appear to bedrawn.
Hublin himself finds it difficult as a matterof logic to exclude any of the “specimensinvolved in the Neandertalization process”from Homo neanderthalensis, “even if theydisplay only a few derived Neandertal fea-tures” (Hublin 1998: 302). But from our ownexamination of the fossils concerned, whatseems most remarkable in the longer-estab-lished record is that in strictly morphologicalterms there is really only one potentially ques-tionable case of attribution to Homo nean-derthalensis, apart from such apparentlypermanently inscrutable specimens as theFontéchevade fragments (see Vallois, 1949;Trinkaus, 1973). This is the rather fragmen-tary assemblage from Ehringsdorf which,despite its limited size, shows unusual varia-tion in morphology (Vlcek, 1993; Schwartzand Tattersall, 2005). Thus, one occipital (EhrH9 1032/69) lacks a suprainiac fossa, andshows instead twinned depressions bothbelow and above the occipital “torus,” whilethe Ehr H3 1026/69 parietal is also atypicalfor a Neanderthal in showing a “tent-shaped”coronal profile in rear view. And the temporalEhr H3 1026/69 shows the straight and long
SYSTEMATIC OF NEANDERTHALS 15

parietomastoid suture of a Neanderthal, butuncharacteristically for this group the posteri-or root of its zygomatic arch diverges stronglyfrom the cranial wall. Still, if we provisionallyregard this material as early Neanderthal, itserves to emphasize the relative homogeneityof the rest of the Neanderthal hypodigm.
None of this is to say, however, that all“Neanderthal” characteristics of the kind welisted in the last section are confined to thelarge group of fossils that we may comfortablyregard as belonging to Homo neanderthalen-sis.This is because in the middle Pleistocene ofEurope we can indeed, like Hublin and mostothers, identify a variety of hominid fossils dis-playing some, but not all, of the features thattypify Neanderthals (Hublin, 1998; Schwartzand Tattersall, 2005). Perhaps most notableamong these is the Steinheim cranium (proba-bly stage 7, around 225 ka), which possesses afairly standard Neanderthal-like morphologyof the upper face, with separately arching andsmoothly rolled supraciliary ridges over orbitsthat have truncated inferomedial margins. Thelarge nasal fossa and the presence of a prenasalfossa between well defined lateral and spino-turbinal crests are characteristics shared withNeanderthals, as are the angulation apparentalong the anterior squamous suture, the long,straightish parietomastoid and anterior lamb-doid sutures, the (rather faint) suprainiacdepression, the horizontal occipital “torus”that is only fully defined below, and the ratherrounded posterior profile of the braincase.There are even the rudiments of a verticallyoriented medial projection faintly evidentwithin the nasal cavity. On the other hand, thepuffy midface of the Neanderthals is absentfrom the Steinheim specimen, as are thesharply retreating zygomas, with their laterallyrising anterior roots, that give Neanderthalfaces their highly characteristic allure. Thepoorly inflated Steinheim braincase departsfrom the Neanderthal condition in having fair-ly vertical side walls in coronal section and inshowing a smoothly rounded lateral profile at
the rear. As a result, despite the manyNeanderthal resemblances of this specimen, ithas been more or less universally regarded (seereviews by Day, 1986; Schwartz and Tattersall,2002) as an example of “archaic Homo sapi-ens” rather than as of Neanderthal affinity. Fewhave ever called Steinheim a Neanderthal, andit would certainly be inaccurate to do so.However, based on the constellation of charac-ters it exhibits, it would seem entirely reason-able to consider the Steinheim specimen asrepresentative of the sister taxon to Homoneanderthalensis. Conceivably this taxon wasdirectly ancestral to the Neanderthals; but tomake that claim would involve a variety ofassumptions that we would prefer to avoid here(see discussion by Tattersall and Eldredge,1977).
Another German specimen with many of thecharacteristic Neanderthal cranial traits is thefrustratingly incomplete partial calvaria fromReilingen, which is possibly penecontempora-neous with that from Steinheim (Ziegler andDean, 1998). Early analyses resulted in attribu-tions to Homo erectus (Czarnetzki, 1989) or to“archaic Homo sapiens” (Adam, 1989, Schott,1990), but more recent contributions havefocused on the Neanderthal affinities of thisspecimen (e.g., Condemi, 1996; Dean et al.,1998). Like Neanderthals, this fossil possessesexpanded petrosal pneumatization; an occipital“torus” that is fully delineated only below, witha suprainiac depression above; an en bombecoronal profile of the expansive braincase; andincomplete lateral ossification of the ectotym-panic tube. As in Steinheim, though, the sagit-tal profile of the occiput is quite rounded, andthe weakly undercut occiput is fairly narrow,though the cranial vault itself appears consider-ably flatter and more Neanderthal-like than thatof Steinheim. This specimen exemplifies thedifficulty of categorizing members of thiswider group strictly on characters of the cranialrear, though it is abundantly clear thatReilingen is either Homo neanderthalensis or aclose relative.
I. TATTERSALL & J.H. SCHWARTZ16

The impressive assemblage of hominid fos-sils from the Sima de los Huesos at Atapuercain Spain, now thought to be around 400 ka orpossibly more (Bischoff et al., 2003), appears tobe relatively homogeneous. All of this materialhas been referred by its describers (e.g.,Arsuaga et al., 1997) to the species Homo hei-delbergensis.This species is, however, based onthe Mauer jaw, a specimen to which none of themandibles known from the Sima appears tobear notable similarities. At the same time,Arsuaga and colleagues have noted that variousfeatures of the Sima fossils are “transitional” toNeanderthal morphology, and have concludedthat these hominids are early members of theNeanderthal lineage, as well as simultaneouslylinked to other European middle Pleistocenefossils (Arsuaga et al., 1997). And certainly,while the Sima hominids do show fewercomponents of the Neanderthal character con-stellation than Steinheim does, the number ofapparent Neanderthal synapomorphies thatthey possess is nonetheless quite extensive.Cranially, such resemblances to Homo nean-derthalensis include: bilaterally arced supraor-bital tori with tall and evenly rounded anteriorsurfaces; orbits with obliquely truncated infer-omedial corners; a large nasal aperture showinga distinct prenasal fossa with a continuousinternal margin; some projection of the frontalprocesses around the nasal aperture; an angula-tion along the anterior squamous suture; a long,straight parietomastoid suture; incompletelylaterally ossified ectotympanic tubes; and a pit-ted suprainiac depression. Like those ofNeanderthals, the Sima mandibles displaymedial pterygoid tubercles on the inner surfaceof the ramus, and have sigmoid notch crests thatterminate just lateral to the midline of thecondyle.
At the same time, however, the Simahominids are cranially less derived and quitedistinct from Homo neanderthalensis. In thestructure of the face, differences from the latterinclude: an uninflated infraorbital region; hor-izontal conchal crests just within the aperture
of the nasal cavity in lieu of vertical medialprojections; no sharply retreating and inferi-orly tapering midface; and anterior zygomaticroots that angle out more sharply laterally.Farther posteriorly, the Sima hominid showsdeep zygomatic arches, a very short anteriorlambdoid suture, and there is no clearly under-cut occipital “torus.” In sagittal profile thebraincase is smoothly rounded, and in coronalprofile it has parallel sides and a sagittal peak.Postcranially the robust Sima pelvis showsgreatly flaring iliac blades, long pubic rami anda capacious pelvic canal. These pelvic featuresrecall the Neanderthals; but they are likely torepresent the primitive condition for the genusHomo, or at least for the subclade that containsboth the Neanderthals and the Sima hominids.
Based purely on the material discussed upto this point, it might (just) be possible toargue that this “Neanderthal clade,” fairly welldefined in terms of synapomorphies, includeda chronological succession of taxa of whichHomo neanderthalensis was the terminal out-come. But from the evidence presented below,it is evident that the larger story of middlePleistocene hominid evolution in Europe wasmore complex than the notion of a singleevolving lineage can accommodate.
For the conclusion that hominid evolutionin Europe prior to the abrupt arrival of Homosapiens was not an essentially unilineal affairis dramatically reinforced by a survey of theentire variety of hominid fossils known fromEurope in the period centering on 400 ka.Such fossils include the specimens fromSwanscombe, in England (about 400 ka:Stringer and Hublin, 1999), the Aragohominids from southern France (perhaps 450ka: Iacumin et al., 1996); the German Mauerjaw (around 500 ka: Cook et al., 1982); theBilzingsleben hominids, also from Germany(300–400 ka: Schwarcz et al., 1988), and pos-sibly the Vérteszöllös occipital from Hungarywhich might be as old as 350–250ka (Cherdyntsev, 1971) though Schwarczand Latham (1990) consider it younger.
SYSTEMATIC OF NEANDERTHALS 17

Morphologically there is strong justificationfor associating the Greek Petralona craniumwith the Arago hominids (Schwartz andTattersall, 2005), although Grün (1996) con-cluded that this exceptionally poorly datedfossil most likely derives from significantlylater in time, around 250–150 ka.
The committee that was originally con-vened to evaluate the two first-found elementsof the Swanscombe cranial rear emphasizedmetrical comparisons to both Homo sapiensand Steinheim (Morant, 1938), and influentiallater contributions (e.g., Howell, 1960;Stringer et al., 1984) continued an essentially“presapiens” assignment. However, followingthe lead of Santa Luca (1978), most workershave moved toward comparisons with Homoneanderthalensis (see Stringer and Gamble,1993; Stringer and Hublin, 1999). And,indeed, Swanscombe clearly does sort into theNeanderthal clade, although it does not seemto represent a typical Neanderthal, and insome ways it more closely resembles its coun-terparts from the Sima de los Huesos, forinstance in having a relatively narrow andweakly undercut occipital “torus” and a poo-rly defined suprainiac fossa. Still, while hav-ing more vertical cranial walls than is usualfor Neanderthals, it does also show a morerounded coronal cranial contour than is seenin the Spanish material and it does possess afairly large, ovoid foramen magnum.
The Vérteszöllös occipital was initiallyannounced as a representative of Homo erec-tus (Vértes, 1965) – in hindsight a virtuallymeaningless attribution in the European con-text – and was subsequently moved to “arch-aic Homo sapiens” by Stringer et al. (1979) ata time when the Petralona cranium was simi-larly classified. The two hominids share along and very horizontal occipital “torus,” butotherwise the Vérteszöllös specimen remainsfairly enigmatic. Also rather inscrutable arethe Bilzingsleben hominids, which have beenascribed to Homo erectus by their finders(e.g., Mania, 1983; Vlcek and Mania, 1987).
Stringer et al. (1984) found these fossils tobe the “most erectus-like and the leastNeanderthal or modern-like” of the “archaicHomo sapiens” group, a conclusion later sus-tained by Stringer (1989) through compar-isons with the Saldanha calotte. The twoBilzingsleben crania are both fragmentary, butawkwardly they appear to display differencesfrom one another (Schwartz and Tattersall,2005) that are substantial enough at least toraise the question of whether they belonged toa single population. The affinities of neitherare clearly evident, but there is little reasonto associate all of the material with theNeanderthal clade and the net effect is to addto the growing impression of hominid divers-ity in the European middle Pleistocene.
The Arago and Petralona crania are nowa-days widely accepted as classic Europeanexemplars of the species Homo heidelbergen-sis, originally founded solely on the basis ofthe Mauer mandible. It is fortunate that theexistence of both mandibles and crania in theArago collection allows this attribution to besubstantiated (Schwartz and Tattersall, 2005).There is a substantial Gestalt differencebetween the Mauer jaw and the two better pre-served Arago mandibles because the latter aremore gracile than the former, and lack itsremarkable ramal length. However, both theMauer and Arago 13 mandibles show a com-mon configuration of the anteroinferior mar-gins of the jaw. They also share excavated androunded gonial regions, anteroposteriorly longcoronoid bases, posteriorly decreasing corpo-ral height, and tall but shallow infracondylarsulci that lie along the posterior margins of therami. More striking, though, are the dentalsimilarities. Notably, the anterior teeth in bothmandibles were large, and the molars long andovoid; the P1s are obliquely truncatedmesiodistally along their lingual surfaces, andare hence more mesiodistally tapering andelongate than the short, buccolingually wideand more ovoid P2s; the protoconids on boththe P1s and P2s are centrally placed; on P1 the
I. TATTERSALL & J.H. SCHWARTZ18

low lingual swelling lies opposite the proto-conid, while on P2 the metaconid is mesiallysituated relative to the protoconid, and on bothpremolars the lingual swelling or cusp isbounded by a very small fovea mesially and amuch deeper fovea distally. In the molars M2
is larger than both M1 and M3; the protoconidsand metaconids are situated very mesially onthe crowns, and in the same mutual relation-ship; M1 shows evidence of a tiny trigonidbasin, while this basin is more pronounced onM2–3. On all molars the hypoconulid lies justbuccal to the crown midline, and the talonidbasin is or was quite long mesiodistally andtruncated buccolingually, with some evidenceof enamel wrinkling.
If we can allocate the Mauer andArago lowerjaws to the same species, then it appears permis-sible to regard the well-preserved Arago 21 fos-sil as the classic exemplar of the face of Homoheidelbergensis. In turn, we can associate withthis species the more complete and potentiallymuch more recent Petralona cranium, whichshares withArago 21 a massive and broad lowerface that lies below hugely developed and super-oinferiorly tall supraorbital margins that attaintheir maximum thickness around mid-orbit. Inboth fossils, the superior margin of the orbitbears a blunt edge that demarcates the anterosu-periorly twisting front surface of the torus fromthe shallow posttoral sulcus behind. The samefeatures unite these European specimens withothers from Africa (Bodo, Kabwe, Saldanha)and Asia (Dali, Jinniushan). Interestingly, whilethere is also a fair amount of variation in otherfeatures of the cranium among this cosmopoli-tan group (see discussion by Schwartz andTattersall, 2005), that variation does not appearto be geographically organized in any clear-cutmanner. In the present context, however, it isimportant to note that the upper facial featuresof Homo heidelbergensis noted above distin-guish this species absolutely from Homoneanderthalensis as well as from broadly con-temporaneous members of the latter’s cladesuch as the Sima hominids.
Conclusion
Homo neanderthalensis is an unambiguouslydemarcated morphospecies, recognizable on ahost of cranial and postcranial characters, thatwas also, as definitely as such things can beknown in the fossil record, a fully individuat-ed historical entity. Its identity did not shadeinto that of any other known hominid, and cer-tainly not into that of the species Homo sapi-ens that entered its European and westernAsian zone of distribution some 40 kyr agoand entirely displaced it within about a dozenmillennia. Further, it clearly emerges from theevidence just adumbrated that the largerNeanderthal clade, if not the species Homoneanderthalensis itself, was already in exis-tence in the period centering on 400 ka.
Inadequate specimens or poor dating make ithard to be sure whether or not multiple kinds ofhominid belonging to this “Neanderthal clade”coexisted at this juncture, although the proba-bility may lie in that direction. On the otherhand, the new datings make it evident that ataround this same time point there also existed inEurope at least one other hominid lineage, rep-resented by Homo heidelbergensis (as definedby the Mauer/Arago fossils). This species isclearly demarcated from all members of theequally derived Neanderthal/Steinheim/ Simaclade in which this cranial region is known bythe apparently derived structure of its upperface (among other features). It is beyond thescope of our discussion here to broach the manyproblems of apparently geography-independ-ent morphological variation in the larger groupto which the species name Homo heidelbergen-sis has been applied in recent years (e.g.,Rightmire, 1990; Stringer and McKie, 1996;Schwartz and Tattersall, 2005). But it is alreadyevident that this species, as broadly defined,somehow shared the European habitat with theNeanderthals and/or their relatives, potentiallyover a period of several hundred thousand yearsif the late dating of Petralona turns out to beaccurate.
SYSTEMATIC OF NEANDERTHALS 19





























