Embed Size (px)
Citation preview

IntroductionPhyllopezus pollicaris (Spix, 1825) is a large-sized,
nocturnal and insectivorous gecko native to central South America (Rodrigues, 1986; Vanzolini, Costa and Vitt, 1980; Vitt, 1995). Currently, P. pollicaris is considered to represent a complex of cryptic species (Gamble et al., 2012) which is widespread along the South American great diagonal of open formations (Vanzolini, 1968; 1988) ranging from the Chaco, through the Brazilian Cerrado and Caatinga eastwards to the coastal Atlantic Forest of north-eastern Brazil (Vanzolini, 1968; Rodrigues, 1986; Cei, 1993). The presence of P. pollicaris has been frequently associated with rock outcrops (Rodrigues, 1986; Cei, 1993; Vitt, 1995; Werneck, Colli and Vitt, 2009).
Phyllopezus pollicaris is locally common but, despite its widespread distribution, ecological knowledge on the species is scarce. The available information is based on a limited number of studied populations (Rodrigues, 1986; Vitt, 1986; 1995; Dias and Lira-da-Silva, 1998; Werneck, Colli and Vitt, 2009) or on anecdotal data (Vanzolini 1974; Vanzolini, Costa and Vitt, 1980; Cei, 1993). Thus, this work aims to provide
information about aspects of the natural history (habitat use, morphology, diet, temperatures, reproductive condition and caudal autotomy) of a population of P. pollicaris from Central Brazil. We discuss local variation and compare our results with available data.
Materials and methods
Study areaField work was carried out in the Estação Ecológica Serra Geral
do Tocantins (EESGT), a large conservation unit in the Cerrado region of Central Brazil. The expedition was conducted with the main objective of making an inventory of the herpetofauna of the EESGT (results in Recoder et al., 2011; Valdujo et al., 2011). The area is characterized by typical Cerrado vegetation compo-sed of extensive grasslands and savannahs, interspersed by palm marshes and gallery forests. The EESGT region has three main levels of altimetric surfaces. The first is composed of a sandstone plateau with elevations ranging between 800 and 900 m, which occupies the eastern portion contiguous with Planalto dos Gerais. Isolated sandstone relicts are also present at the central portion of the Ecological Station. The second level composes an extensive sandy depression which resulted from erosion of the plateau, with elevations ranging between 400 and 550 m in the central, western and southern portions. The final level is an intermediary plateau crossing the station oriented NNW-SSE direction, with elevations ranging between 600 and 700 m.
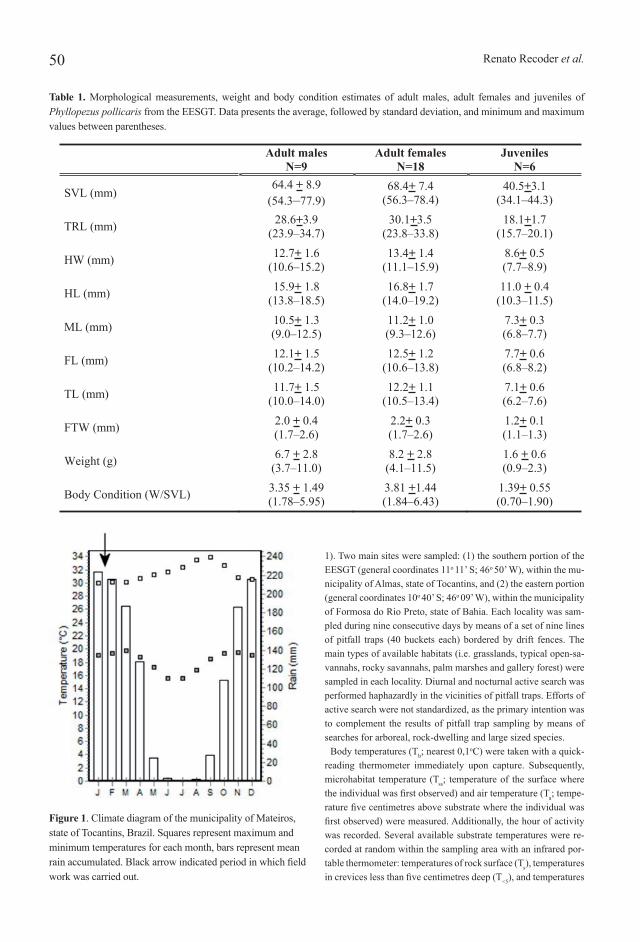
Field data collectionField work was conducted from the end of January through the
beginning of February 2008, at the end of the rainy season (Figure
Herpetology Notes, volume 5: 49-58 (2012) (published online on 18 March 2012)
Natural history of the tropical gecko Phyllopezus pollicaris (Squamata, Phyllodactylidae) from a sandstone outcrop
in Central Brazil.
Renato Recoder1*, Mauro Teixeira Junior1, Agustín Camacho1 and Miguel Trefaut Rodrigues1
1 Laboratório de Herpetologia, Departamento de Zoologia, Instituto de Biociências, Rua do Matão, no 101, CEP 05508-090, Universidade de São Paulo, São Paulo, Brazil;
*Corresponding author; e-mail: [email protected]
Abstract. Natural history aspects of the Neotropical gecko Phyllopezus pollicaris were studied at Estação Ecológica Serra Geral do Tocantins, in the Cerrado region of Central Brazil. Despite initial prospection at different types of habitats, all individuals were collected at sandstone outcrops within savannahs. Most individuals were observed at night, but several specimens were found active during daytime. Body temperatures were significantly higher in day-active individuals. We did not detect sexual dimorphism in size, shape, weight, or body condition. All adult males were reproductively mature, in contrast to just two adult females (11%), one of which contained two oviductal eggs. Dietary data indicates that P. pollicaris feeds upon a variety of arthropods. Dietary overlap between sexes and age classes was moderate to high. The rate of caudal autotomy varied between age classes but not between sexes. Our data, the first for a population of P. pollicaris from a savannah habitat, are in overall agreement with observations made in populations from Caatinga and Dry Forest, except for microhabitat use and reproductive cycle.
Keywords. Cerrado, lizard, local variation, niche breadth, thermal ecology, sexual dimorphism, tail autotomy.
Renato Recoder et al. 50
1). Two main sites were sampled: (1) the southern portion of the EESGT (general coordinates 11o 11’ S; 46o 50’ W), within the mu-nicipality of Almas, state of Tocantins, and (2) the eastern portion (general coordinates 10o 40’ S; 46o 09’ W), within the municipality of Formosa do Rio Preto, state of Bahia. Each locality was sam-pled during nine consecutive days by means of a set of nine lines of pitfall traps (40 buckets each) bordered by drift fences. The main types of available habitats (i.e. grasslands, typical open-sa-vannahs, rocky savannahs, palm marshes and gallery forest) were sampled in each locality. Diurnal and nocturnal active search was performed haphazardly in the vicinities of pitfall traps. Efforts of active search were not standardized, as the primary intention was to complement the results of pitfall trap sampling by means of searches for arboreal, rock-dwelling and large sized species.
Body temperatures (Tb; nearest 0,1oC) were taken with a quick-reading thermometer immediately upon capture. Subsequently, microhabitat temperature (Tss; temperature of the surface where the individual was first observed) and air temperature (Ta; tempe-rature five centimetres above substrate where the individual was first observed) were measured. Additionally, the hour of activity was recorded. Several available substrate temperatures were re-corded at random within the sampling area with an infrared por-table thermometer: temperatures of rock surface (Ts), temperatures in crevices less than five centimetres deep (T<5), and temperatures
Figure 1. Climate diagram of the municipality of Mateiros, state of Tocantins, Brazil. Squares represent maximum and minimum temperatures for each month, bars represent mean rain accumulated. Black arrow indicated period in which field work was carried out.
Adult males N=9
Adult females N=18
Juveniles N=6
SVL (mm) 64.4 + 8.9
(54.3–77.9) 68.4+ 7.4
(56.3–78.4) 40.5+3.1
(34.1–44.3)
TRL (mm) 28.6+3.9 (23.9–34.7)
30.1+3.5 (23.8–33.8)
18.1+1.7 (15.7–20.1)
HW (mm) 12.7+ 1.6 (10.6–15.2)
13.4+ 1.4 (11.1–15.9)
8.6+ 0.5 (7.7–8.9)
HL (mm) 15.9+ 1.8 (13.8–18.5)
16.8+ 1.7 (14.0–19.2)
11.0 + 0.4 (10.3–11.5)
ML (mm) 10.5+ 1.3 (9.0–12.5)
11.2+ 1.0 (9.3–12.6)
7.3+ 0.3 (6.8–7.7)
FL (mm) 12.1+ 1.5 (10.2–14.2)
12.5+ 1.2 (10.6–13.8)
7.7+ 0.6 (6.8–8.2)
TL (mm) 11.7+ 1.5 (10.0–14.0)
12.2+ 1.1 (10.5–13.4)
7.1+ 0.6 (6.2–7.6)
FTW (mm) 2.0 + 0.4 (1.7–2.6)
2.2+ 0.3 (1.7–2.6)
1.2+ 0.1 (1.1–1.3)
Weight (g) 6.7 + 2.8 (3.7–11.0)
8.2 + 2.8 (4.1–11.5)
1.6 + 0.6 (0.9–2.3)
Body Condition (W/SVL) 3.35 + 1.49 (1.78–5.95)
3.81 +1.44 (1.84–6.43)
1.39+ 0.55 (0.70–1.90)
Table 1. Morphological measurements, weight and body condition estimates of adult males, adult females and juveniles of Phyllopezus pollicaris from the EESGT. Data presents the average, followed by standard deviation, and minimum and maximum values between parentheses.

Natural history of the tropical gecko Phyllopezus pollicaris 51
Figure 2. Typical open savannah with sandstone outcrops where individuals were observed, at eastern portion of the Estação Ecológica Serra Geral do Tocantins (top); individual active by morning on rock surfaces (bottom).
Renato Recoder et al. 52
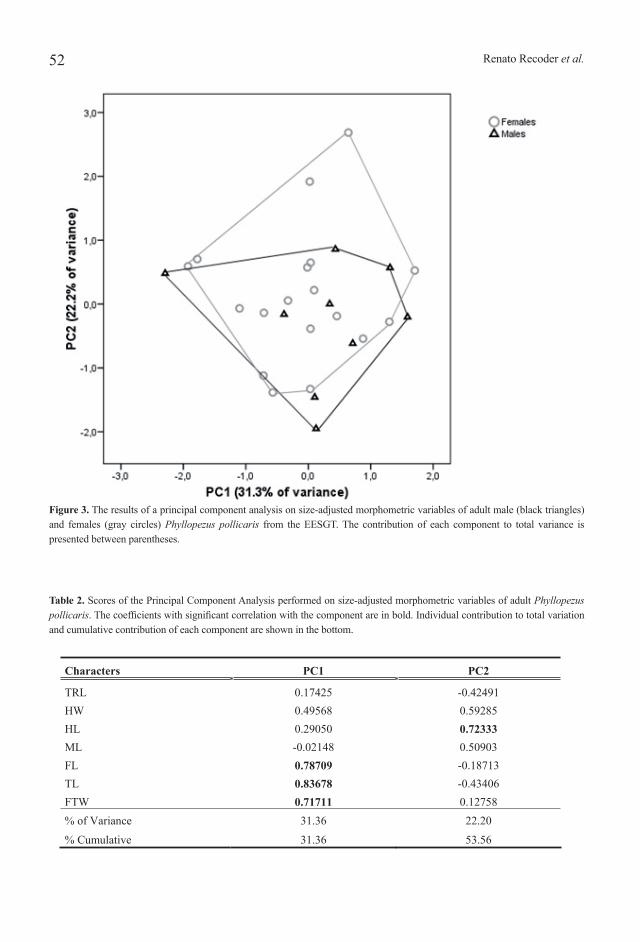
Table 2. Scores of the Principal Component Analysis performed on size-adjusted morphometric variables of adult Phyllopezus pollicaris. The coefficients with significant correlation with the component are in bold. Individual contribution to total variation and cumulative contribution of each component are shown in the bottom.
Characters PC1 PC2
TRL 0.17425 -0.42491 HW 0.49568 0.59285 HL 0.29050 0.72333 ML -0.02148 0.50903 FL 0.78709 -0.18713 TL 0.83678 -0.43406 FTW 0.71711 0.12758 % of Variance 31.36 22.20
% Cumulative 31.36 53.56
Figure 3. The results of a principal component analysis on size-adjusted morphometric variables of adult male (black triangles) and females (gray circles) Phyllopezus pollicaris from the EESGT. The contribution of each component to total variance is presented between parentheses.

in crevices deeper than five centimetres (T>5). Available substrate temperatures were taken at night (22:00 pm) and morning (11:00 am) periods. Variation in air temperature was recorded hourly with a HOBO data logger for eight consecutive days.
Laboratory data collectionSpecimens were euthanized by a lethal injection of anaesthetic,
weighed to the nearest 0.1 g using Pesola© string balances, fixed using a 10% formalin solution and preserved in 70% ethanol. Eight morphometric measures were taken with a digital calliper to the nearest 0.1 mm: snout-vent length (SVL), head length (HL), head width (HW), mandible length (ML), trunk length between members (TRL), femur length (FL), tibia length (TL) and fourth toe width (FTW). Individual body condition was estimated by di-viding the total body mass by SVL. Intact and regenerated porti-ons of tails were measured to estimate rates of tail autotomy.
Lizards were dissected to remove stomach contents for dietary analysis. Prey items were identified to the lowest taxonomic level. We calculated an index of importance (Ix; modified from Howard et al., 1999) to represent the relative importance of each prey item in relation to the entire diet:
where nx and fx are number and frequency of the prey category x, and N and F the equivalents for all prey items. We estimated the niche breadth for numerical data with the inverse of Simpson’s diversity index (Simpson, 1949):
where p is the proportion of utilization of prey item i, and n is the number of categories. B varies from 1 (utilization of a single prey category) to n (even utilization of all prey categories). We calculate dietary overlap between sexes and age groups with the Pianka’s niche overlap index (Pianka, 1974):
where pik is the proportion of utilization of prey item i by species k and pij is the proportion of utilization of prey item i by species j. Dietary overlap varies from 0 (no overlap) to 1 (complete over-lap).
Dissected lizards were sexed through examination of gonads. Females were considered reproductive if vitellogenic follicles, corpora lutea and/or oviductal eggs were present. Males were considered reproductive by presence of convoluted epididymides and enlarged testes. Voucher specimens are deposited in the her-petological collection of the Museu de Zoologia da Universidade de São Paulo, São Paulo, Brazil.
Statistical analysisIntraspecific sex variation in size, weight, body condition and
overall morphology was investigated with univariate and mul-
tivariate analysis of variance (ANOVA and MANOVA). Shape variation was summarized with a Principal Component Analysis (PCA) performed using the residuals of the linear regression of the morphometric data on the SVL. The effect of environmental temperatures over body temperatures was investigated with mul-tiple linear regression analysis. Differences in tail-loss frequen-cies between sexes and age groups were tested using chi-square test (χ2). We calculate estimations of complete tails for individuals with tail broken, with the coefficients of the linear regression of intact tails with SVL. The relative regeneration of tail and the frequency of tail-break in each quart of tail length (from proximal to distal portion of complete tails) were calculated based on the estimations of complete tails. Statistical analyses were performed with SPSS v.15.0 and in R (R Development Core Team, 2010), with a 0.05 significance level.
Results
Sample distribution We obtained 33 individuals of Phyllopezus pollicarus
in the EESGT. All lizards were hand-collected during active search. Most individuals (31 specimens; 93.9%) were obtained within the eastern portion of the station. Two individuals (6.1%) were obtained at the central portion (“Morro do Fumo” hill). No observations of P. pollicarus were made in the southern portion of the station. All individuals were collected on sandstone outcrops in typical savannah habitat (Fig. 2).
Morphological variation and sexual dimorphismThe largest female measured 78.4 mm SVL and
weighted 11.5 g, the largest male 77.9 mm SVL and weighted 11.0 g (see Table 1 for descriptive statistics). The smallest individual measured 34.1 mm and weighted 0.9 g. Adult females and males did not differ in size (ANOVA; F1,25 = 1.538, P = 0.226), weight (F1,25 = 1.685, P = 0.206) or body condition (F1,25 = 1.794, P = 0.192). The sexes did not differ in any other morphometric character (MANOVA; F7,19 = 1.779, P = 0.150). Overall shape variation was summarized by PCA with size-adjusted data, which did not reveal any conspicuous morphological differentiation between sexes (Fig. 3, Table 2).
Reproductive conditions The sample consisted of six immature juveniles and
twenty seven adult individuals. The smallest juvenile could not be accurately sexed. Of the eighteen adult females analysed, two were reproductive (11.0%), one presenting a single vitellogenic follicle, the other containing two enlarged oviductal eggs. Corpora lutea was not observed.
Of the thirteen males, eight presented convoluted epididymides and enlarged testes (mean ratio of testicle
Natural history of the tropical gecko Phyllopezus pollicaris 53

Renato Recoder et al. 54
Body temperatures Microhabitat temperatures Environmental temperatures
Tb Tss Ta Ts T<5 T>5
Night 24.4 + 1.0 (23.0–26.6)
22.9 + 1.0 (21.0–25.2)
21.9 + 1.0 (20.0–24.4)
22.7 + 0.7 (21.1–23.7)
23.2 + 0.7 (21.8–25.4)
23.5 + 0.6 (22.6–25.1)
Morning 28.5 + 2.2 (25.0–31.2)
26.5 + 1.8 (25.0–31.2)
28.1 + 2.5 (24.9–30.8)
27.5 + 2.4 (24.2–32.0)
26.3 + 2.4 (23.0–32.0)
23.7 + 1.9 (22.0–29.0)
Table 3. Selected temperatures at microhabitat (body Tb, substrate Tss and air Ta) and available environmental temperatures (rock surfaces Ts, crevices less than five centimetres deep T<5, and crevices deeper than five centimetres T>5). Data show averages followed by standard deviation, and minimum and maximum values in parentheses.
Prey Type N %N F %F Ix
INSECTA Orthoptera 8 8.3 7 29.2 0.19 Blattodea 5 6.0 4 16.7 0.11 Coleoptera 3 3.6 2 8.3 0.06 Coleoptera larvae 2 2.4 1 4.2 0.03 Hemiptera
Heteroptera 3 3.6 3 12.5 0.08 Homoptera 3 3.6 2 8.3 0.06
Hymenoptera Formicidae 14 16.7 5 20.8 0.19 Formicidae alate 1 1.2 1 4.2 0.03
Isoptera 17 20.2 4 16.7 0.18 Apterygota 11 13.1 8 33.3 0.23 Diptera 1 1.2 1 4.2 0.03 Mantodea 1 1.2 1 4.2 0.03 Lepidoptera larvae 2 2.4 2 8.3 0.05
ARACHNIDA Araneae 9 10.7 8 33.3 0.22Scorpionida 1 1.2 1 4.2 0.03
MIRIAPODA Diplopoda 1 1.2 1 4.2 0.03
OTHERSLizard skin 2 2.4 2 8.3 0.05 Egg shell pieces 1 1.2 1 4.2 0.03
Total 85 24Niche Breadth 8.80
Table 4. Number (N), percentage (%N), frequency (F), frequency in percentage (%F) and importance index (Ix) of prey categories in the diet of Phyllopezus pollicaris in the EESGT. In the bottom is presented the dietary niche breadth.
diameter/SVL = 0.079 + 0.006 mm) and were considered to be reproductive (69.2%). One sub adult had slightly convoluted epididymides and moderately enlarged testes (ratio of testicle diameter/SVL = 0.055). Four juvenile males did not have developed testicles (ratio of testicle diameter/SVL < 0.015) thus were considered immature.
Activity and temperaturesSix individuals were captured during the morning
(between 11:00 and 11:30 am), of which four were directly exposed to sunlight (Fig. 2). Four individuals were captured between 17:00 and 17:30 pm hidden under rocks and crevices. The remaining 23 individuals
were captured at night (between 21:10 and 23:35 pm) from rock surfaces and large crevices.
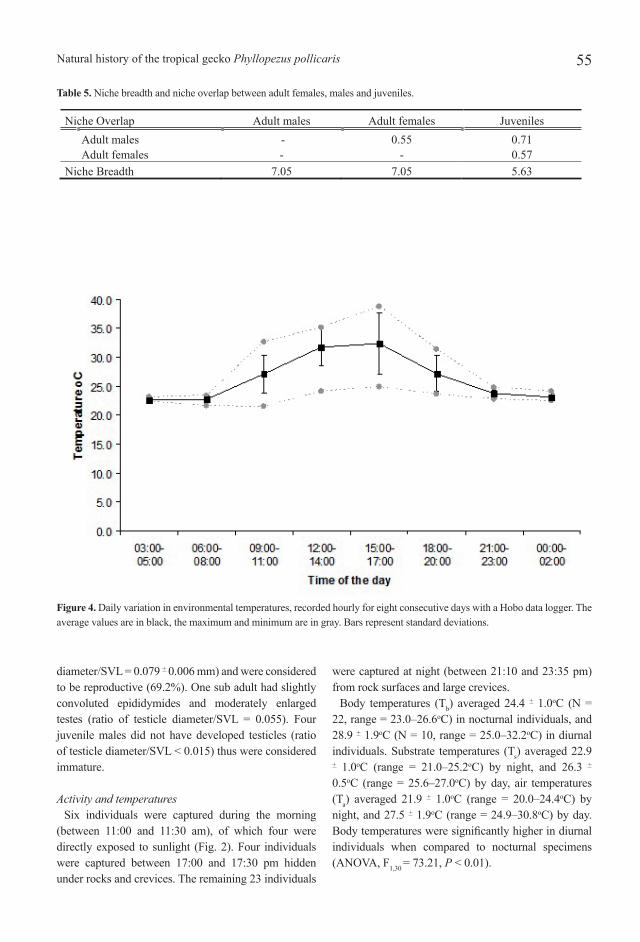
Body temperatures (Tb) averaged 24.4 + 1.0oC (N = 22, range = 23.0–26.6oC) in nocturnal individuals, and 28.9 + 1.9oC (N = 10, range = 25.0–32.2oC) in diurnal individuals. Substrate temperatures (Ts) averaged 22.9 + 1.0oC (range = 21.0–25.2oC) by night, and 26.3 + 0.5oC (range = 25.6–27.0oC) by day, air temperatures (Ta) averaged 21.9 + 1.0oC (range = 20.0–24.4oC) by night, and 27.5 + 1.9oC (range = 24.9–30.8oC) by day. Body temperatures were significantly higher in diurnal individuals when compared to nocturnal specimens (ANOVA, F1,30 = 73.21, P < 0.01).
Natural history of the tropical gecko Phyllopezus pollicaris 55
Table 5. Niche breadth and niche overlap between adult females, males and juveniles.
Niche Overlap Adult males Adult females Juveniles Adult males - 0.55 0.71 Adult females - - 0.57
Niche Breadth 7.05 7.05 5.63
Figure 4. Daily variation in environmental temperatures, recorded hourly for eight consecutive days with a Hobo data logger. The average values are in black, the maximum and minimum are in gray. Bars represent standard deviations.

The multiple regression model indicated that body temperature at night is weakly associated with environmental temperatures (r2 = 0.292; F = 2.290; P = 0.147). Body temperatures of lizards active between 11.00-11.30 am is significantly associated with air temperature (r2 = 0.864; F = 9.515; P = 0.05).
Daily variation in air temperatures has been summarized in Figure 4. Temperatures of selected microhabitats (Tss) did not differ from random substrate temperatures at night (ANOVA; F1,44 = 2.138, P = 0.150) and morning (F1,28 = 2.116, P = 0.156). However, temperatures of selected microhabitats by active lizards at morning are significantly lower than temperatures of adjacent rock surfaces exposed to sun (F1,28 = 8.360, P < 0.01). Table 3 summarizes the selected and available substrate temperatures.
DietWe analysed 33 stomach contents and identified 85
items in 18 prey categories. Nine stomachs were empty (27.3%). The prey items with the highest importance index were Apterygota (0.23), Araneae (0.22), Orthoptera (0.19), Formicidae (0.19) and Isoptera (0.18) respectively (Table 4). Niche breadth based on numerical data was 8.80.
Overlap in dietary niches between adult males and juveniles was high (0.712), but moderate between adult females and males (0.556), or adult females and juveniles (0.579) (Table 5). Niche breadth was similar for adults of both sexes (7.05) but lower for juveniles (5.63). We found no evidence for intra-population niche partitioning between sexes or between adults and juveniles in diet.
Tail autotomy and regenerationSix specimens had intact tails (18.2%), five individuals
had broken tails (15.1%) and the remaining twenty two lizards (66.7%) possessed regenerated tails. Excluding individuals with broken tails (in which case tail loss cannot be readily attributed to natural causes), we found a frequency of regenerated tails of 78.6% in the sample. Tail loss did not differ between sexes (χ2 = 0.330; P = 0.565), but the frequency was significantly higher in adults when compared to juveniles (χ2 = 10.889; P < 0.001).
A linear regression of intact tails with SVL as independent variable shows that size is a strong factor in explaining tail length (r2= 0.981; F4,1 = 202.87; P < 0.001). The relative regeneration varied from 47.5% to 100% (mean = 82.6 + 0.14%) of the
estimated complete tail. Tail-break occurred more frequently in the first quart of the tail (59.3%, N=16); less frequently in the second (37.0%, N=10) and on a single occasion in the third quarter (3.7%).
Discussion
In agreement with several other South American Phyllodactylids (sensu Gamble et al., 2008; 2011) which occur in similar habitats, the savannah population of P. pollicaris studied here occurred in close association with rocks (e.g. Vanzolini, 1982; Marquet et al., 1990; Cei, 1993; Colli et al., 2003; Koch, Venegas and Böhme, 2006; Cacciali, Ávila and Bauer, 2007; Blanco, Villavicencio and Acosta, 2009; Werneck, Colli and Vitt, 2009). Nevertheless, P. pollicaris can be found within a wide range of microhabitats (e.g. tree trunks, fallen logs, cactuses, palm leaves, rock crevices, bushes, bark, fences) in the Caatinga and Atlantic Forest domains (Rodrigues, 1986; 2003; pers. obs.). Such environmental plasticity has also been observed in geckos of the genus Gymnodactylus. While G. amarali seems to be a microhabitat specialist within the Cerrado (Colli et al., 2003; Vitt et al., 2007), G. geckoides utilizes a wide range of microhabitats in the Caatinga (Vitt, 1995; Rodrigues, 2003). Microhabitat choice by lizards may be constrained in habitats with high temperature variation to sites that offer sufficient opportunities for thermoregulation (Marquet et al., 1990; Kearney, 2002; Hitchcock and McBreyer, 2006; Aguilar and Cruz, 2010). Thus, rocks may constitute a thermally suitable microhabitat for nocturnal lizards in the Cerrado, by providing refuges with a gradient of temperatures (Kearney and Predavec, 2000; Kearney, 2002; Vitt et al., 2007). Although we took temperatures of several day and night-active individuals of P. pollicaris, by lack of evidence and lack of an adequate experimental design, we can not address whether the studied specimens were thermoconformers in nocturnal activity and if individuals thermoregulate during the day as our empirical observation suggests, and as do other New World nocturnal geckos (e.g. Vitt 1995; Vitt and Zani, 1997; Colli et al., 2003).
Our results show that P. pollicaris is an opportunistic predator that feeds upon a variety of active arthropods. Upon comparison of our data with other three populations for which dietary data are available, we found that a similar variety of prey items were ingested. In all cases, crickets, ants, termites, spiders, insect larvae and beetles represent the most abundant prey items (Rodrigues, 1986; Vitt, 1995; Dias and Lira-da-
Renato Recoder et al. 56

Silva, 1998; Werneck, Colli and Vitt, 2009). Also, P. pollicaris has been commonly placed among the species with widest dietary niche in lizard communities they occur, with estimated niche breadths varying between 4.90 and 9.09 (Vitt, 1995; Dias and Lira-da-Silva, 1998; Werneck, Colli and Vitt, 2009).
The results presented herein regarding female reproductive state are in agreement with those of Rodrigues (1986), who found no signs of reproductive activity in a sample of ten adult P. pollicaris obtained during February in the Caatinga. In contrast, Vitt (1986, 1995) studied a lizard community in the Caatinga for over a year, and found that four sympatric geckos, including P. pollicaris, show nearly continuous reproduction. Seasonal reproduction can however be expected for P. pollicaris populations within the Cerrado, as this pattern has been observed in most lizards from the domain, due to its highly marked seasonality in precipitation and/or food resources (Colli, 1991; Van Sluys, 1993; Wiederhecker, Pinto and Colli, 2002; Colli et al., 2003; Mesquita and Colli, 2003).
In consensus with our results, Vitt (1986) did not recover sexual dimorphism in a populations of P. pollicaris from the Caatinga. Lack of the same phenomenon was found in two other nocturnal Geckonidae, while sexual dimorphism was present in two diurnal species (Vitt, 1986). The latter author stated that sexually dimorphic characters may differ in visually-oriented compared to auditory-oriented tropical geckos, what may explain the lack of sexual dimorphism in the nocturnal species (Vitt, 1986).
The high frequencies of tail-break were similar to those reported for other tropical geckos, which varies between 32.3% in Hemidactylus palaichthus (Zani, 1996), 65.6% in Thecadactylus rapicauda (Vitt and Zani, 1997), to 72.5% in Gymnodactylus amarali (Vitt et al., 2007). High incidence of tail-break in P. pollicaris may be a phylogenetically conserved propensity for caudal autotomy in geckos (Arnold, 1984; Van Sluys, Vrcibradic and Rocha, 2002; Bateman and Fleming, 2009). We also found a high relative tail regeneration in the population, reinforcing the importance of tail autotomy as a defensive strategy for this species (Vitt, Congdon and Dickson, 1977). Although males P. pollicaris occasionally engage in aggressive encounters (Vitt, 1983), no differences in tail-break frequencies was observed between sexes. Nevertheless, frequencies are significantly higher in adults than juveniles. This pattern has also been observed in other tropical geckos (e.g. Vitt, 1983; Vitt and Zani, 1997). Thus, variation in frequency
of regenerated tails more likely reflects escape success from predation attempts than intra-specific aggression (Vitt, 1983; Medel, 1988; Vitt and Zani, 1997; Cooper Jr., Pérez-Mellado and Vitt, 2004).
Acknowledgements. We thank Cristiano Nogueira, Paula Valdujo, Pedro Nunes, Tami Mott and José Mario Ghellere for field assistance and logistical support, and Rodrigo Dios and Silvio Nihei whom kindly helped with identification of prey items. We thank Wouter Beukema for his critical review and valuable suggestions that improved the manuscript. We also thank the “Instituto Chico Mendes de Conservação da Biodiversidade” (ICMBIO) for allowing the faunal survey in EESGT and providing collection permit (12187-1). The Fundação Boticário de Conservação da Natureza, “Conselho Nacional de Desenvolvimento Científico e Tecnológico” (CNPq) and “Fundação de Apoio à Pesquisa do Estado de São Paulo” (FAPESP) provided financial support.
References
Aguilar, R., Cruz, F.B. (2010): Refuge use in a patagonian noc-turnal lizard, Homonota darwini: the role of temperature. J. Herpetol. 44: 236-241.
Arnold, E.N. (1984): Evolutionary aspects of tail shedding in li-zards and their relatives. J. Nat. Hist. 18: 127-169.
Bateman, P.W., Fleming, P.A. (2009): To cut a long tail short: a review of lizard caudal autotomy studies carried out over the last 20 years. J. Zool. 277: 1-14.
Blanco, G., Villavicencio, H.J., Acosta, J.C. (2009): Field body temperature, diet, and reproduction of Homonota andicola (Gekkonidae) in Catamarca, Argentina. Herp. Rev. 40: 156-158.
Cacciali, P., Ávila, I., Bauer, F. (2007): A new species of Homo-nota (Squamata, Gekkonidae) from Paraguay, with a key to the genus. Phyllomedusa 6: 137-146.
Cei, J.M. (1993): Reptiles del noroeste, nordeste y este de la Ar-gentina. Museo Regionale di Scienze Naturali, Torino. Mono-grafie 14: 1-949.
Colli, G.R. (1991): Reproductive ecology of Ameiva ameiva (Sauria: Teiidae) in the cerrado of central Brazil. Copeia 1991: 1002-1012.
Colli, G.R., Mesquita, D.O., Rodrigues, P.V.V., Kitayama, K. (2003): Ecology of the Gecko Gymnodactylus geckoides ama-rali in a Neotropical Savanna. J. Herpetol. 37: 694-706.
Cooper Jr., W.E., Pérez-Mellado, V., Vitt, L.J. (2004): Ease and effectiveness of costly autotomy vary with predation intensity among lizard populations. J. Zool. 262: 243-255.
Dias, E.J.R., Lira–da–Silva, R. (1998): Utilização dos recursos alimentares por quatro espécies de lagartos (Phyllopezus polli-caris, Tropidurus hispidus, Mabuya machrorhyncha e Vanzos-saura rubricauda) da Caatinga UHE-Xingó. Braz. J. Ecol. 1998: 97-101.
Gamble, T., Bauer, A.M., Greenbaum, E., Jackman, T.R. (2008): Out of the blue: a novel, trans-Atlantic clade of geckos (Gekko-ta, Squamata). Zool. Scripta 37: 355-366.
Gamble, T., Bauer, A.M., Colli, G.R. Greenbaum, E. Jackman, T.R. Vitt, L.J., Simons, A.M. (2011): Coming to America: mul-
Natural history of the tropical gecko Phyllopezus pollicaris 57

tiple origins of New World geckos. J. Evol. Biol. 24: 231-244.Gamble, T., Colli, G.R., Rodrigues, M.T., Werneck F. P., Simons,
A.M. (2012): Phylogeny and cryptic diversity in geckos (Phyl-lopezus; Phyllodactylidae; Gekkota) from South America’s open biomes. Mol. Phylogenet. Evol. 62: 943-953.
Hitchcock, M.A., McBrayer, L.D. (2006): Thermoregulation in nocturnal ecthotherms: seasonal and intraspecific variation in the Mediterranean Gecko (Hemidactylus turcicus). J. Herpetol. 40: 185-195.
Howard, A.K., Forester, J.D., Ruder, J.M., Pamerlee, Jr., J.S., Powell, R. (1999): Natural history of a terrestrial Hispaniolan Anole: Anolis barbouri. J. Herpetol. 33: 702-706.
Kearney, M. (2002): Hot rocks and much-too-hot rocks: seasonal patterns of retreat-site selection by a nocturnal ectotherm. J. Therm. Biol. 27: 205-218.
Kearney, M., Pradavec, M. (2000): Do nocturnal ectotherms ther-moregulate? A study of the temperate gecko Christinus mar-moratus. Ecology 81: 2984-2996.
Koch, C., Venegas, P.J., Böhme, W. (2006): A remarkable dis-covery: description of a big-growing new gecko (Squamata: Gekkonidae: Phyllopezus) from northwestern Peru. Salamand-ra 42: 145-150.
Marquet, P.A., Bozinović, F., Medel, R.G., Werner, Y.L., Jaksić F.M. (1990): Ecology of Garthia gaudichaudi, a gecko en-demic to the Semiarid Region of Chile. J. Herpetol. 24: 431-434.
Medel, R.G., Jiménez, J.E., Fox, S.F., Jaksic, F.M. (1988): Experi-mental evidence that high population frequencies of lizard tail autotomy indicate inefficient predation. Oikos 53: 321-324.
Mesquita, D.O., Colli, G.R. (2003): Geographical variation in the ecology of populations of some Brazilian species of Cnemido-phorus (Squamata, Teiidae). Copeia 2003: 285-298.
Pianka, E.R. (1974): Niche overlap and diffuse competition. Proc. Natl. Acad. Sci. USA 71: 2141-2145.
R Development Core Team (2010): R: A language and environ-ment for statistical computing. R Foundation for Statistical Computing, Vienna, Austria. URL http://www.R-project.org.
Recoder, R.S., Teixeira Jr., M., Camacho, A.G., Nunes, P.M.S., Mott, T., Valdujo, P.H., Ghellere, J.M., Nogueira, C., Rodri-gues, M.T. (2011): Répteis da Estação Ecológica Serra Geral do Tocantins, Brasil Central. Biota Neotropica 11: 1-19.
Rodrigues, M.T. (1986): Uma nova espécie do gênero Phyllo-pezus de Cabaceiras: Paraíba: Brasil; com comentários sobre a fauna de lagartos da área (Sauria Gekkonidae). Pap. Avul. Zool., São Paulo 36: 237-250.
Rodrigues, M.T. (2003): Herpetofauna da Caatinga. In: Ecologia e Conservação da Caatinga, p. 181-236. Leal, I.R., Tabarelli, M., da Silva, J.M.C., Eds., Universidade Federal de Pernam-buco, Recife.
Simpson, E.H. (1949): Measurement of diversity. Nature 163: 688.
Valdujo, P.H., Camacho, A.G., Recoder, R.S., Teixeira Jr., M., Ghellere, J.M.B., Mott, T., Nunes, P.M.S., Nogueira, C., Rodri-gues, M.T. (2011): Anfíbios da Estação Ecológica Serra Geral do Tocantins, região do Jalapão, Estados do Tocantins e Bahia. Biota Neotropica 11: 251-261.
Van Sluys, M. (1993): The reproductive cycle of Tropidurus itam-bere (Sauria: Tropiduridae) in southeastern Brazil. J. Herpetol. 27: 28-32.
Van Sluys, M., Vrcibradic, D., Rocha, C.F.D. (2002): Tail loss in the syntopic lizards Tropidurus itambere (Tropiduridae) and Mabuya frenata (Scincidae) in southeastern Brazil. Stud. Neo-trop. Fauna Environ. 37: 227-231.
Vanzolini, P.E. (1968): Geography of the South American Gekko-nidae (Sauria). Arqu. Zool. 17: 85-112.
Vanzolini, P.E. (1974): Ecological and geographical distribution of lizards in Pernambuco, Northeastern Brasil (Sauria). Pap. Avul. Zool. 28: 61-90.
Vanzolini, P.E. (1982): A new Gymnodactylus from Minas Gerais, Brazil, with remarks on the genus, on the area and on montane endemisms in Brazil (Sauria, Gekkonidae). Pap. Avul. Zool. 34: 403-413.
Vanzolini, P.E. (1988): Distributional patterns of South American lizards. In: Proceedings of a workshop on Neotropical distribu-tional patterns, p. 317-342. Vanzolini, P.E., Heyer, W.R, Eds., Academia Brasileira de Ciências. Rio de Janeiro.
Vanzolini, P.E., Costa A.M., Vitt, L.J. (1980): Répteis das Caa-tingas. Academia Brasileira de Ciências. Rio de Janeiro.
Vitt, L.J. (1983): Tail loss in lizards: the significance of foraging and predator escape modes. Herpetologica 39: 151-162.
Vitt, L.J. (1986): Reproductive Tactics of Sympatric Gekkonid Lizards with a Comment on the Evolutionary and Ecological Consequences of Invariant Clutch Size. Copeia 1986: 773-786.
Vitt, L.J. (1995): The ecology of tropical lizards in the Caatinga of northeast Brazil. Occ. Pap. Oklahoma Mus. Nat. Hist. 1: 1-29.
Vitt, L.J., Zani, P.A. (1997): Ecology of the Nocturnal Lizard Thecadactylus rapicauda (Sauria: Gekkonidae) in the Amazon Region. Herpetologica 53: 165-179.
Vitt, L.J., Congdon, J.D., Dickson, N.A. (1977): Adaptive stra-tegies and energetics of tail autotomy in lizards. Ecology 58: 326-337.
Vitt, L.J., Shepard, D.B., Caldwell, J.P., Vieira, G.H.C., França, F.G.R., Colli, G.R. (2007): Living with your food: geckos in termitaria of Cantão. J. Zool. 272: 1-8.
Zani, P.A. (1996): Patterns of caudal autotomy evolution in li-zards. J. Zool. 240: 210-220.
Werneck, F.R, Colli, G.R., Vitt, L.J. (2009): Determinants of as-semblage structure in Neotropical dry forest lizards. Austral Ecol. 34: 97-115.
Wiederhecker, H.C., Pinto, A.C.S., Colli, G.R. (2002): Reproduc-tive Ecology of Tropidurus torquatus (Squamata: Tropiduri-dae) in the Highly Seasonal Cerrado Biome of Central Brazil. J. Herpetol. 36: 82-91.
Renato Recoder et al. 58
Accepted by Wouter Beukema





























