Embed Size (px)
Citation preview

Mycorrhizal networks: a review of their extent,function, and importance1
Suzanne W. Simard and Daniel M. Durall
Abstract: It is well known from laboratory studies that a single mycorrhizal fungal isolate can colonize different plantspecies, form interplant linkages, and provide a conduit for interplant transfer of isotopic carbon, nitrogen, phosphorus,or water. There is increasing laboratory and field evidence that the magnitude and direction of transfer is influenced byphysiological source–sink gradients between plants. There is also evidence that mycorrhizal fungi play a role in regu-lating transfer through their own source–sink patterns, frequency of links, and mycorrhizal dependency. Although it isplausible that connections are extensive in nature, field studies have been hampered by our inability to observe them insitu and by belowground complexity. In future, isotopic tracers, morphological observations, microsatellite techniques,and fluorescent dyes will be useful in the study of networks in nature. Mycorrhizal networks have the potential to in-fluence patterns of seedling establishment, interplant competition, plant diversity, and plant community dynamics, butstudies in this area are just beginning. Future plant community studies would benefit from concurrent experimental useof fungal network controls, isotopic labeling, direct observation of interplant linkages, and long-term observation in thefield. In this paper, we review recent literature on mycorrhizal networks and interplant carbon transfer, suggest futureresearch directions, and highlight promising scientific approaches.
Key words: common mycorrhizal network, carbon transfer, source–sink, establishment, competition, diversity.
Résumé : Les études conduites en laboratoire démontrent très bien qu’un seul isolat fongique mycorhizien peut coloni-ser différentes espèces de plantes, former des liens interplants et constituer un conduit pour le transfert d’eau ou decarbone isotopique, d’azote et de phosphore entre les plants. Il existe également des preuves, au laboratoire aussi bienque sur le terrain, que l’ordre de grandeur et la direction du transfert sont influencés par les gradients source–puit entreles plants. On a aussi démontré que les champignons mycorhiziens jouent un rôle dans la régulation du transfert vialeur propre patron source–puit, la fréquence des liens, et la dépendance mycorhizienne. Bien qu’il soit plausible que lesliens soient extensifs en nature, les études aux champs sont rendues difficiles par notre incapacité à les observer in situ,ainsi que par la complexité du milieu hypogé. L’étude des réseaux en nature devra faire appel à des techniques tellesque les traceurs isotopiques, les observations morphologiques, l’analyse par microsatellites, et les colorants fluorescents.Les réseaux mycéliens ont le potentiel d’influencer les patrons d’établissement des plantules, la compétition interplant,la diversité végétale, et la dynamique des communautés, mais les études sur ce sujet ne font que commencer. Les nou-velles études sur les communautés végétales gagneraient à utiliser expérimentalement et de façon concomitante le con-trôle des réseaux fongiques, le marquage isotopique, l’observation directe des connections interplants, et lesobservations à long terme sur le terrain. Cette revue de la littérature récente, sur les réseaux mycorhiziens et le trans-fert interplants de carbone, suggère de nouvelles avenues de recherches et des approches scientifiques prometteuses.
Mots clés : réseau mycorhizien commun, transfert de carbone, source–puit, établissement, compétition, diversité.
[Traduit par la Rédaction] Simard and Durall 1165
Introduction
The terms mycorrhizal network and common mycorrhizalnetwork (CMN) have recently been used by mycor-
rhizoligists. A CMN occurs where two or more root systemsare interconnected by mycorrhizal fungal hyphae. Mycor-rhizal networks have been shown to function by transferringcarbon (C) or nutrients from one plant to another (Finlayand Read 1986a, 1986b), but CMNs can also exist regard-less of whether they are involved with interplant elementaltransfer. For example, CMNs may allow movement of nutri-ents or C within the fungal mycelium, but without evertransferring elements into the tissue of plants interconnectedby the CMN. Thus, CMNs may or may not function withrespect to interplant elemental transfer.
The CMN can involve multiple fungal and plant specieswithin a community. One of the simplest mycorrhizal net-works occurs when the mycelium of one fungal individualconnects two plants of the same species. The complexity ofthe network increases with increasing numbers of fungalspecies, frequency of connections, number of plants within a
Can. J. Bot. 82: 1140–1165 (2004) doi: 10.1139/B04-116 © 2004 NRC Canada
1140
Received 22 September 2003. Published on the NRCResearch Press Web site at http://canjbot.nrc.ca on3 September 2004.
S.W. Simard.2 Forest Science Department, University ofBritish Columbia, 2424 Main Mall, Vancouver, BC V6T 1Z4,Canada.D.M. Durall. Biology Department, Okanagan UniversityCollege, 3333 College Way, Kelowna, BC V1V 1V7, Canada.
1This article is one of a selection of papers published in theSpecial Issue on Mycorrhizae and was presented at theFourth International Conference on Mycorrhizae.
2Corresponding author (e-mail: [email protected]).

species, and number of plant species. Complexity of CMNeffects also increases with interactions with other soil organ-isms, such as earthworms, nematodes, or bacteria.
The CMN has the potential to influence plant survival andgrowth by influencing C and nutrient status of individualplants. Research in this area has focused mainly on seedlingestablishment in the neighbourhood of larger trees sharingthe same fungal species. Growth of plants already estab-lished may also be affected to different degrees by the CMN,potentially altering intra- and inter-specific competitive in-teractions among plants within a plant community. Shifts incompetitive interactions could then affect dominance andevenness within a plant community, thereby affecting plantspecies diversity and structure.
In this paper, we review recent advances in the study ofmycorrhizal networks and C transfer, focusing primarily onliterature between 1997 and 2003. We build on earlier re-views by Newman (1988), Miller and Allen (1992), Fitter(2001), and Simard et al. (2002). The review focuses primar-ily on C transfer because it has been the focus of our ownresearch, as well as that of much of the recent literature, andwe refer readers to Simard et al. (2002) for our earlier re-view of nitrogen (N) and phosphorus (P) transfers. Wedevelop our discussion first by reviewing evidence for theexistence and size of CMNs, second by summarizing studiesthat have examined CMN-mediated C transfer betweenplants, and third by providing evidence for plant and fungalfactors that influence transfer. These sections provide back-ground for a more theoretical discussion of potential CMNand transfer influences on feedbacks among plants andmycorrhizal fungi. Finally, we discuss the potential influ-ences that CMNs may have on plant community dynamics(plant establishment, interplant competition, and plant diver-sity) using supporting evidence from the literature. In thecourse of our review, we identify areas in need of furtherstudy and suggest promising scientific approaches for exam-ining mycorrhizal networks.
Evidence for the existence and size of mycorrhizalnetworks
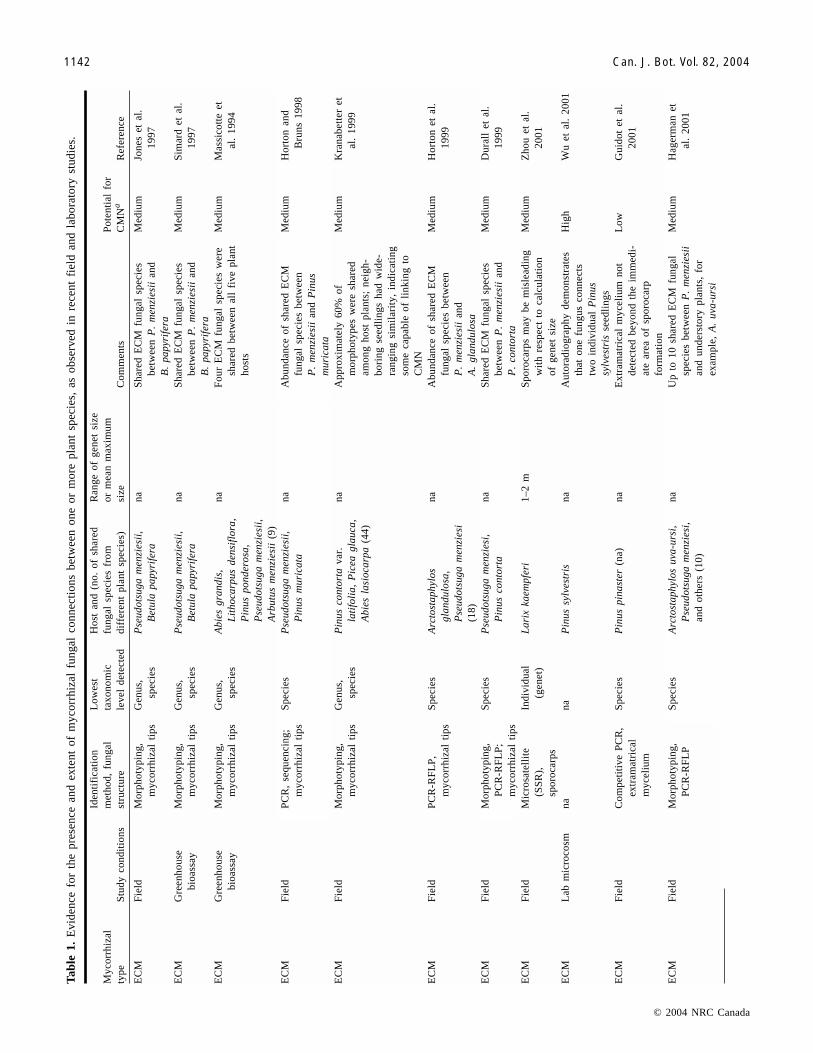
Newman (1988) suggested that many of the mycorrhizalplants growing together in forests or grasslands may be in-terconnected by a CMN and that this could have profoundimplications for ecosystem functioning. Only in laboratorystudies, however, have the concurrent existence of mycor-rhizal networks and transfer of elements from one root sys-tem to another been conclusively demonstrated (McKendricket al. 2000; Wu et al. 2001). Direct physical and functionalevidence of interplant hyphal linkages (listed in Table 1) hasalso been reported in several earlier studies using macro-scopic, microscopic, and autoradiographic tracings withintransparent laboratory microcosms for ectomycorrhizal(ECM) fungi (Reid and Woods 1969; Read et al. 1985; Fin-lay and Read 1986a, 1986b; Arnebrant et al. 1993; Wu et al.2001) and for arbuscular mycorrhizal (AM) fungi (Hirreland Gerdemann 1979; Francis and Read 1984). Some of themost conclusive evidence for ecologically significant levelsof C transfer occurs between conifers and myco-heterotrophic plants in the families Orchidaceae and Montro-paceae. In this relationship, the myco-heterotrophic plantreceives most or all of its C from an ECM fungal symbiont,
which links the myco-heterotrophic plant to a chloro-phyllous plant (Bjorkman 1960; McKendrick et al. 2000).Myco-heterotrophic plants can also be specialists with AMfungi, as shown recently by Bidartondo et al. (2002), butthere currently exists no direct evidence of C transfer facili-tated by AM fungi as it does with ECM fungi. Given that Ccan move from a chlorophyllous plant into a myco-heterotrophic plant via ECM hyphal links, it is our view thatC could also move into the shoot of a chlorophyllous plant,provided there is a strong enough source–sink gradient fororganic compounds. This topic is discussed further in a latersection.
In the field, direct observations of interplant hyphal link-ages are rare due to the small, hyaline, and interminglingnature of mycorrhizal hyphae (Newman 1988) and due todifficulties in restricting root and fungal growth to two di-mensions for enhanced visibility. Interplant C transfer be-tween plant species pairs sharing ECM (Simard et al. 1997a)and AM fungi (Lerat et al. 2002) has been measured in thefield, with small or negligible transfer to incompatiblemycorrhizal host plants, strongly suggesting that transferoccurred predominantly through a CMN pathway. In thesestudies, however, the mycorrhizal pathway was not visuallyquantified using autoradiography, leaving the identity of thetransfer pathway in question. Notably, all field studies on Ctransfer suffer from this shortcoming.
Additional evidence for mycorrhizal networks exists fromboth lab and field studies, showing low specificity (at leastat the species level) among mycorrhizal fungi and hostplants (van der Heidjden and Sanders 2002; Bruns et al.2002b). There is considerable evidence that many AM fungihave low host specificity (van der Heijden et al. 2003), eventhough a recent study suggests there is more specificityamong AM fungi than originally thought (Helgason et al.2002). ECM conifer and angiosperm hosts are also colo-nized more prevalently by generalist fungi (Horton andBruns 1998; Sakakibara et al. 2002) than by specialists. Oneexample of a specialist association occurs among myco-heterotrophic plant species, which associate with a narrowspecies group of ECM fungi (Bidartondo and Bruns 2002)and AM fungi (Bidartondo et al. 2002). More commonly,however, using morphotyping techniques, ECM fungal spe-cies or genera have been shown to colonize different individ-uals of the same plant species (Kranabetter et al. 1999)or different plant species (Jones et al. 1997; Simard et al.1997b). The use of molecular techniques, such as restrictionfragment length polymorphism (RFLP) and sequencing ofthe polymerase chain reaction (PCR)-amplified internal tran-scribed spacer region, confirm morphotyping studies show-ing that most ECM fungal species associate with multipleplant host species (Horton and Bruns 1998; Horton etal. 1999; Hagerman et al. 2001; Sakakibara et al. 2002;P.G. Kennedy, personal communication). These studies sug-gest there is a high potential for mycorrhizal networks inboth ECM and AM plant communities.
Specificity may also occur below the species level(Vrålstad et al. 2002). Although the same mycorrhizal fun-gal species may be found on different host plants, there maybe different fungal genetic variants specific to different hostplants. Genetic variability within a fungal species may re-duce the possibility of anastomoses between the mycelium
© 2004 NRC Canada
Simard and Durall 1141
© 2004 NRC Canada
1142 Can. J. Bot. Vol. 82, 2004
Myc
orrh
izal
type
Stu
dyco
ndit
ions
Iden
tifi
cati
onm
etho
d,fu
ngal
stru
ctur
e
Low
est
taxo
nom
icle
vel
dete
cted
Hos
tan
d(n
o.of
shar
edfu
ngal
spec
ies
from
diff
eren
tpl
ant
spec
ies)
Ran
geof
gene
tsi
zeor
mea
nm
axim
umsi
zeC
omm
ents
Pot
enti
alfo
rC
MN
aR
efer
ence
EC
MF
ield
Mor
phot
ypin
g,m
ycor
rhiz
alti
psG
enus
,sp
ecie
sP
seud
otsu
gam
enzi
esii
,B
etul
apa
pyri
fera
naS
hare
dE
CM
fung
alsp
ecie
sbe
twee
nP
.m
enzi
esii
and
B.
papy
rife
ra
Med
ium
Jone
set
al.
1997
EC
MG
reen
hous
ebi
oass
ayM
orph
otyp
ing,
myc
orrh
izal
tips
Gen
us,
spec
ies
Pse
udot
suga
men
zies
ii,
Bet
ula
papy
rife
rana
Sha
red
EC
Mfu
ngal
spec
ies
betw
een
P.
men
zies
iian
dB
.pa
pyri
fera
Med
ium
Sim
ard
etal
.19
97
EC
MG
reen
hous
ebi
oass
ayM
orph
otyp
ing,
myc
orrh
izal
tips
Gen
us,
spec
ies
Abi
esgr
andi
s,L
itho
carp
usde
nsif
lora
,P
inus
pond
eros
a,P
seud
otsu
gam
enzi
esii
,A
rbut
usm
enzi
esii
(9)
naF
our
EC
Mfu
ngal
spec
ies
wer
esh
ared
betw
een
all
five
plan
tho
sts
Med
ium
Mas
sico
tte
etal
.19
94
EC
MF
ield
PC
R,
sequ
enci
ng;
myc
orrh
izal
tips
Spe
cies
Pse
udot
suga
men
zies
ii,
Pin
usm
uric
ata
naA
bund
ance
ofsh
ared
EC
Mfu
ngal
spec
ies
betw
een
P.
men
zies
iian
dP
inus
mur
icat
a
Med
ium
Hor
ton
and
Bru
ns19
98
EC
MF
ield
Mor
phot
ypin
g,m
ycor
rhiz
alti
psG
enus
,sp
ecie
sP
inus
cont
orta
var.
lati
foli
a,P
icea
glau
ca,
Abi
esla
sioc
arpa
(44)
naA
ppro
xim
atel
y60
%of
mor
phot
ypes
wer
esh
ared
amon
gho
stpl
ants
;ne
igh-
bori
ngse
edli
ngs
had
wid
e-ra
ngin
gsi
mil
arit
y,in
dica
ting
som
eca
pabl
eof
link
ing
toC
MN
Med
ium
Kra
nabe
tter
etal
.19
99
EC
MF
ield
PC
R-R
FL
P,
myc
orrh
izal
tips
Spe
cies
Arc
tost
aphy
los
glan
dulo
sa,
Pse
udot
suga
men
zies
i(1
8)
naA
bund
ance
ofsh
ared
EC
Mfu
ngal
spec
ies
betw
een
P.
men
zies
iian
dA
.gl
andu
losa
Med
ium
Hor
ton
etal
.19
99
EC
MF
ield
Mor
phot
ypin
g,P
CR
-RF
LP
;m
ycor
rhiz
alti
ps
Spe
cies
Pse
udot
suga
men
zies
i,P
inus
cont
orta
naS
hare
dE
CM
fung
alsp
ecie
sbe
twee
nP
.m
enzi
esii
and
P.
cont
orta
Med
ium
Dur
all
etal
.19
99
EC
MF
ield
Mic
rosa
tell
ite
(SS
R),
spor
ocar
ps
Indi
vidu
al(g
enet
)L
arix
kaem
pfer
i1–
2m
Spo
roca
rps
may
bem
isle
adin
gw
ith
resp
ect
toca
lcul
atio
nof
gene
tsi
ze
Med
ium
Zho
uet
al.
2001
EC
ML
abm
icro
cosm
nana
Pin
ussy
lves
tris
naA
utor
adio
grap
hyde
mon
stra
tes
that
one
fung
usco
nnec
tstw
oin
divi
dual
Pin
ussy
lves
tris
seed
ling
s
Hig
hW
uet
al.
2001
EC
MF
ield
Com
peti
tive
PC
R,
extr
amat
rica
lm
ycel
ium
Spe
cies
Pin
uspi
nast
er(n
a)na
Ext
ram
atri
cal
myc
eliu
mno
tde
tect
edbe
yond
the
imm
edi-
ate
area
ofsp
oroc
arp
form
atio
n
Low
Gui
dot
etal
.20
01
EC
MF
ield
Mor
phot
ypin
g,P
CR
-RF
LP
Spe
cies
Arc
tost
aphy
los
uva-
ursi
,P
seud
otsu
gam
enzi
esi,
and
othe
rs(1
0)
naU
pto
10sh
ared
EC
Mfu
ngal
spec
ies
betw
een
P.
men
zies
iian
dun
ders
tory
plan
ts,
for
exam
ple,
A.
uva-
ursi
Med
ium
Hag
erm
anet
al.
2001
Tab
le1.
Evi
denc
efo
rth
epr
esen
cean
dex
tent
ofm
ycor
rhiz
alfu
ngal
conn
ecti
ons
betw
een
one
orm
ore
plan
tsp
ecie
s,as
obse
rved
inre
cent
fiel
dan
dla
bora
tory
stud
ies.
© 2004 NRC Canada
Simard and Durall 1143
Myc
orrh
izal
type
Stu
dyco
ndit
ions
Iden
tifi
cati
onm
etho
d,fu
ngal
stru
ctur
e
Low
est
taxo
nom
icle
vel
dete
cted
Hos
tan
d(n
o.of
shar
edfu
ngal
spec
ies
from
diff
eren
tpl
ant
spec
ies)
Ran
geof
gene
tsi
zeor
mea
nm
axim
umsi
zeC
omm
ents
Pot
enti
alfo
rC
MN
aR
efer
ence
EC
MF
ield
Mic
rosa
tell
ite
(IS
SR
)In
divi
dual
(gen
et)
Pin
usra
diat
a(n
a)1–
40m
Aw
ide
vari
ety
ofge
net
size
;ge
nets
may
repr
esen
ta
col-
lect
ion
ofra
met
s
Med
ium
Saw
yer
etal
.20
01
EC
MF
ield
Mic
rosa
tell
ite
Indi
vidu
al(g
enet
)P
icea
sitc
hens
is,
Pin
usco
ntor
ta(n
a)3–
18m
Mos
tge
nets
occu
pyin
gar
eas
of<
3m
Med
ium
Ber
gem
ann
and
Mil
ler
2002
EC
MF
ield
PC
R-R
FL
P,
sequ
enci
ngS
peci
esP
seud
otsu
gam
enzi
esi,
Lit
hoca
rpus
dens
iflo
ra(1
7)
naA
bund
ance
ofsh
ared
EC
Mfu
ngal
spec
ies
betw
een
P.
men
zies
iian
dL
.de
nsif
lora
Med
ium
Ken
nedy
etal
.20
03
EC
MF
ield
Mic
rosa
tell
ite
Indi
vidu
al(g
enet
)P
seud
otsu
gam
enzi
esi
1–13
m,
max
imum
<4
mG
enet
prop
agat
ion
inch
ante
rell
ege
nets
ism
ore
like
lyth
ere
sult
ofba
sidi
ospo
redi
sper
sal
than
vege
tati
vesp
read
Low
Dun
ham
etal
.20
03
AM
Fie
ldP
CR
,se
quen
cing
Spe
cies
Rub
usfr
utic
osus
,E
pilo
bium
angu
stif
oliu
m,
Ace
rps
eudo
plan
tanu
s,A
juga
rept
ans,
Gle
chom
ahe
dera
cea
naA
Mfu
ngi
can
colo
nize
diff
er-
ent
host
spec
ies,
but
host
sele
ctiv
ity
was
also
evid
ent
Med
ium
Hel
gaso
net
al.
2002
AM
Gro
wth
cham
ber,
mem
bran
esa
ndw
ich
naS
ubsp
ecie
sna
Des
crib
esa
mec
hani
smby
whi
chC
MN
can
form
betw
een
diff
eren
tA
Mpl
ants
ofth
esa
me
spec
ies
Med
ium
Gio
vane
tti
etal
.20
01
AM
Gre
enho
use,
pots
Roo
tle
ngth
infe
cted
Spe
cies
Bra
chyp
odiu
mpi
nnat
um,
Pru
nell
avu
lgar
is(1
–3)
naS
uppo
rts
the
com
mon
idea
that
inA
Mth
ere
isno
sele
ctiv
-it
yfo
rho
st
Med
ium
Van
der
Hei
jden
etal
.20
03O
rchi
d(m
yco-
hete
rotr
oph)
,E
CM
Lab
,P
etri
dish
esP
CR
-RF
LP
Sub
spec
ies,
anas
tam
ose
grou
ping
s
Pin
ussy
lves
tris
,G
oody
era
repe
nsna
Des
crib
esa
mec
hani
smby
whi
chC
MN
can
form
be-
twee
nor
chid
san
dE
CM
plan
tsp
ecie
s
Med
ium
Sen
etal
.19
99O
rchi
d(m
yco-
hete
rotr
oph)
,E
CM
Not
e:A
M,
arbu
scul
arm
ycor
rhiz
a;E
CM
,ec
tom
ycor
rhiz
a;SS
R,
sim
ple
sequ
ence
repe
at;
ISSR
,in
ters
impl
ese
quen
cere
peat
;na
,no
tap
plic
able
.a H
igh,
conc
lusi
veev
iden
cefo
rhy
phal
conn
ectio
nsbe
twee
ntw
oor
mor
epl
ants
and
the
CM
Nis
capa
ble
oftr
ansp
ortin
gel
emen
tsfr
omon
ero
otsy
stem
toan
othe
r;m
ediu
m,
high
prob
abili
tyth
atfu
nc-
tioni
ngC
MN
exis
ts,
but
conc
lusi
veev
iden
ceno
tre
port
ed;
low
,lit
tleev
iden
cefo
rth
eex
iste
nce
ofm
ycor
rhiz
alne
twor
ks.
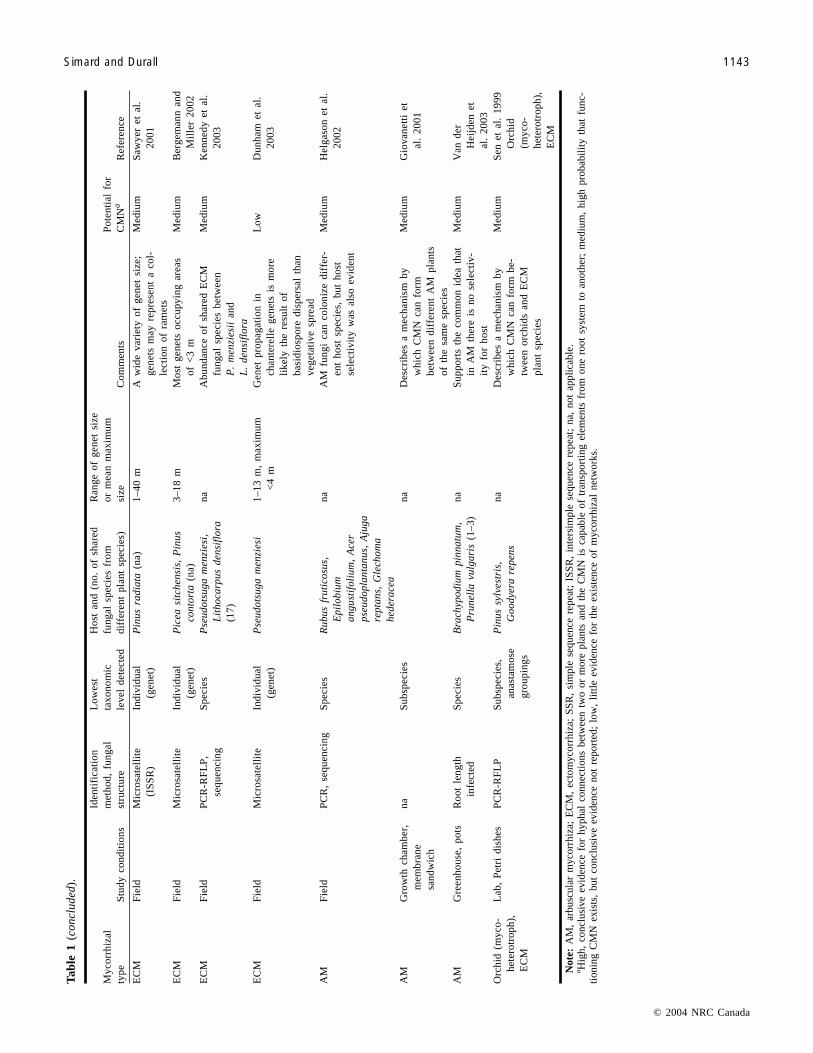
Tab
le1
(con
clud
ed).

of different host plants (Sen et al.1999; Giovannetti et al.2001), reducing the potential for formation of a CMN. Incontrast, finding a single genet on different plant specieswould provide strong evidence for the presence of a CMN,since the fungal hyphae would probably anastomose. Micro-satellite analysis of DNA allows mapping of ECM fungalgenets within a forest. Studies have shown that ECM fungalgenet size ranges from less than 1 m2 to 300 m2 (Bonello etal. 1998; Sawyer et al. 2001). Although microsatellite tech-niques have not been used specifically to demonstrate theexistence of a CMN, ECM fungal genets have been shownto extend over an area encompassing multiple plant hosts ofthe same or different plant species (Zhou et al. 2001; Sawyeret al. 2001; Bergemann and Miller 2002; Dunham et al.2003). Future research is necessary to determine whetherfungal genets occur on different host plants of the same ordifferent species.
Even though microsatellite techniques show promise,genet size may be problematic in estimating the size ofmycorrhizal networks. For example, genet size estimatesmay encompass a series of ramet clumps rather than onecontinuous genet and therefore may not represent a continu-ous mycorrhizal network (Sawyer et al. 2001; Dunham et al.2003). This is especially true when genet size is estimatedfrom sporocarps (Sawyer et al. 2001). In other ECM fungalspecies, however, whose hyphae are thought to extend forlarge distances, there is more chance that genet size wouldbe representative of a CMN (Bonello et al. 1998).
In this section, we reviewed recent evidence for the exis-tence of common mycorrhizal networks. Much more infor-mation is needed to confirm their existence, size,distribution, and function before we can make conclusivestatements about their ecological significance.
Pathways for interplant C transferThe molecular and morphological fungal specificity stud-
ies described above provide evidence that both AM andECM fungal species can associate with multiple plant hostsand that mycorrhizal hyphae emanating from one plant cancolonize the roots of neighboring plants, forming a CMN.Studies using isotopic tracers have suggested that interplanttransfer can occur along several different pathways. Inter-plant transfer can occur directly through an intact CMN(pathway 2a, Fig. 1a) (McKendrick et al. 2000; Wu et al.2001) or a CMN comminuted by soil organisms (pathway2b, Fig. 1a) (Tuffen et al. 2002). Alternatively, transfercan occur along a mycorrhizal–soil pathway (pathway 3,Fig. 1a), where C is exuded or leaked into the soil pool bythe mycorrhizal roots or mycelium of one plant and then ispicked up by the mycorrhizal roots or mycelium of a neigh-bouring plant (Newman et al. 1989), or along anonmycorrhizal–soil pathway (pathway 1, Fig. 1a), wherethere is no movement at any point within fungal hyphae. Inpathways 1, 2b, and 3, C or nutrients exuded into the soilpool are subject to biochemical transformations by soilorganisms, depletion or fixation by physical or chemical pro-cesses, movement as soluble organic compounds, or respira-tion, followed by anapleurotic uptake by neighbour roots.Individual ECM plants can have some root tips that associ-ate with host-specific fungi, others that associate with host-generalist fungi, and some that are nonmycorrhizal (e.g.,
Jones et al. 1997), raising the possibility that multiple path-ways can operate simultaneously within a single plant pair.Empirical evidence for simultaneous multiple pathways co-mes from Simard et al. 1997a, whose data support directtransfer between paper birch and Douglas-fir through theCMN as well as indirect transfer to AM western redcedarthrough the mycorrhizal–soil pathway.
© 2004 NRC Canada
1144 Can. J. Bot. Vol. 82, 2004
Fig. 1. (a) Model pathways for interplant carbon and nutrienttransfer. In pathway 1 (dotted arrow), plants A and B arenonmycorrhizal (NM), and transfer occurs through the soil poolwithout entering mycorrhizal hyphae at any point. In pathway 2a(solid arrows), transfer between plant A and plant B occurs en-tirely within the common mycorrhizal network (CMN) formedby generalist fungus Z. In pathway 2b (broken arrows), transferbetween plant A and plant B is facilitated by soil animals feed-ing on external hyphae produced by mycorrhizal fungi. In path-way 3 (broken–dotted arrow), plant A is colonized with host-specific fungus X and plant B with fungus Y (no CMN), andtransfer occurs partly through fungi X and Y and partly throughthe soil pool. (b) Simple model of interspecific competition be-tween plant A and plant B via the NM–soil pathway (generalinterplant competition for resources). The upper dotted line withend-circle denotes the competitive effect of plant A on plant Band vice versa.

C transfer between plantsAs outlined in the Introduction, our review primarily in-
cludes studies on C transfer published in the past 6 years,but with brief reference to key historical papers. Relevantstudies published between 1997 and 2003 are summarized inTable 2. The earliest evidence for C or nutrient transferthrough a CMN involved movement of 14C from large coni-fer trees in the field to neighbouring myco-heterotrophicplants of the species Monotropa hypopitys Crantz (Bjorkman1960). Since then, using PCR-RFLP techniques, severalmyco-heterotrophic plants have been shown to formendomycorrhizal associations with fungi that simultaneouslyform ECM with surrounding trees (Cullings et al. 1996; Tay-lor and Bruns 1997, 1999; McKendrick et al. 2000; Bidar-tondo and Bruns 2002; Bruns et al. 2002a; Bidartondo et al.2002). In a recent study, McKendrick et al. (2000) providedthe first experimental confirmation that growth of the myco-heterotrophic orchid Corallorhiza trifida Chatelain wassustained by C directly received from a neighbouringautotrophic tree (Betula pendula Roth. or Salix repens L.)through linked fungal mycelia of a shared symbiont. Theylabeled the two tree species with 14C and, using digitalautoradiography and tissue oxidation, found that one-waytransfer to the coralloid rhizome of Corallorhiza trifida wasequivalent to 0.31%–0.38% of label in the donor trees. Theyalso found that Corallorhiza trifida gained 6%–14% of masswhen linked to autotrophs, but lost 13% when not linked,probably resulting from uncompensated maintenance costs.These studies on myco-heterotrophic plants provide conclu-sive evidence that interplant C transport occurs throughhyphal networks.
Interplant transfer of C has also been studied extensivelyin AM and ECM chorophyllous plant systems sinceBjorkman’s original groundbreaking work. In early labora-tory studies, 14C was used to show C transfer from one ECMconifer seedling into the roots and shoots of another via aCMN (Reid and Woods 1969; Finlay and Read 1986a). Sim-ilar work was done in AM systems, but unlike the ECMstudies, transferred label did not move into receiver plant tis-sues (Hirrel and Gerdemann 1979; Francis and Read 1984).In the first field study with ECM plants, Read et al. (1985)showed transfer of 14C to multiple receiver plants, raisingquestions about the existence of bidirectional or net transferand about the ecological significance of the quantities of Ctransferred between plants (Newman 1988; Newman et al.1997).
C transfer between ectomycorrhizal plantsTo address the identified need to investigate bidirectional
and net transfer (Newman 1988), Simard et al. (1997a) andSimard et al. (1997d) used dual (13C–14C) pulse-labeling toexamine CMN-mediated transfer between ECM Pseudo-tsuga menziesii Mirb. (Franco) and Betula papyrifera Marsh.seedlings in the laboratory and field. Bidirectional C transferwas balanced between P. menziesii and B. papyrifera in thelaboratory and first-year field experiment, and in the secondfield year, P. menziesii received a net C gain fromB. papyrifera. Net gain by P. menziesii averaged 6% of Cisotope uptake through photosynthesis, with more in deepshade (10%) than in full or partial sun (3%–4%). Label wasdetected in receiver plant shoots in all transfer experiments,
indicating movement of transferred C out of fungal and intoplant tissues (Simard et al. 1997a, 1997d, 1997e). In thefield, equidistant AM Thuja plicata Donn ex D. Don seed-lings absorbed <1% and 18% of transferred isotope in thefirst and second years, respectively, suggesting that most Cwas transferred through the ECM hyphal pathway, but thatsome was also transferred through soil pathways. Based onthe field and lab results, Simard et al. (1997a) and Simard etal. (1997d) concluded that interplant C transfer was a com-plex and variable process that involved hyphal as well assoil pathways. One of the shortcomings of these experimentswas that the presence and functional status of hyphal con-nections were not unequivocally demonstrated (e.g., usingradiography); instead their presence was inferred from ECMshared-compatibility studies, using morphological and mo-lecular techniques (Jones et al. 1997; Simard et al. 1997b;Sakakibara et al. 2002), from observations of hyphal links inlaboratory rootboxes and from measurement of transfer be-tween EM versus AM seedlings.
The experiments of Simard et al. (1997a) and Simard etal. (1997d) were constructively criticized by Fitter et al.(1998), Robinson and Fitter (1999), and Fitter (2001), justi-fiably highlighting some of the limitations of the studies. Weare in agreement with Robinson and Fitter (1999) and Fitteret al. (1999) regarding several of these shortcomings. Theysuggested, instead of AM controls, use of physical barriersto prevent hyphal contact between ECM plants, full reci-procity of all dual-labeling treatments, quantification ofdonor root isotopic-specific activities, identification of trans-ferred C compounds, examination of both plant and fungalregulatory factors, and investigation of the significance oftransfer in a broader array of natural ecosystems and overlonger periods of time. We are addressing several of theselimitations in our current research programs, investigatingbidirectional transfer in mixed and single species forests(L.J. Philip, M.D Jones, and S.W. Simard, unpublished data;F. Teste, S.W. Simard, and D.M. Durall, unpublished data).
We felt that some of the criticisms of Robinson and Fitter(1999) and Fitter et al. (1999), however, were unfounded.We addressed most of these criticisms in the review bySimard et al. (2002), and we expand on them here. Robinsonand Fitter (1999) reiterated that a large amount of C (18%)was absorbed by AM T. plicata, suggesting the occurrenceof a soil-transfer pathway through release and capture ofexudates and possibly negating the primacy of a hyphal-transfer pathway. The significance of a soil-transfer pathwaywas never disputed by Simard et al. (1997a), and the com-mon occurrence of host-general and host-specific fungi on asingle ECM root system (Molina et al. 1992; Simard et al.1997b; Jones et al. 1997) suggests that multiple transferpathways operating simultaneously in a given plant pair areto be expected (see model pathways above). Even so, theoccurrence of pathway 2a (Fig. 1a) is suggested by the ex-periment of Simard et al. (1997a) as well as others (Francisand Read 1984), which show greater transfer when mycor-rhizal links are present. Secondly, Robinson and Fitter(1999) argued that our studies showed one-way rather thanbidirectional transfer, based on the nearly 10-fold larger iso-tope transfer from B. papyrifera to P. menziesii than viceversa in the second-year field experiment and the possibilitythat received isotope lies within the error calculation limits.
© 2004 NRC Canada
Simard and Durall 1145

© 2004 NRC Canada
1146 Can. J. Bot. Vol. 82, 2004
Myc
orrh
izal
type
;st
udy
cond
itio
nsIs
otop
esu
ppli
ed;
anal
ysis
met
hod
Pla
ntsp
ecie
spa
irs;
fung
alin
ocul
ant
Met
hod
used
tose
para
tefu
ngal
from
soil
path
way
Tra
nsfe
rpa
thw
ayfo
und
Fact
orfo
und
toin
flue
nce
tran
sfer
Type
oftr
ansf
erfo
und
and
othe
rre
leva
ntre
sult
sM
easu
reof
amou
nttr
ansf
erre
dR
ecei
ver
tiss
ueR
efer
ence
EC
M;
fiel
dP
ulse
d14
Can
d13
C;
bioa
ssay
Pse
udot
suga
men
zies
ii,
Bet
ula
papy
rife
ra;
fiel
dso
il
Thu
japl
icat
a(A
Mco
ntro
l)1°
:E
CM
;2°
:so
ilS
hadi
ngof
P.m
enzi
esii
;pr
esen
ceof
hyph
alli
nks
(i)
Two-
way
tran
s-fe
rbe
twee
nE
Mpl
ants
;(i
i)ne
ttr
ansf
erto
P.m
enzi
esii
;(i
ii)
twof
old
mor
etr
ansf
erin
shad
eth
anin
full
ligh
t,co
nsis
-te
ntw
ith
Cso
urce
–sin
kpa
tter
n
Net
tran
sfer
was
3%–1
0%of
tota
lis
otop
ein
dono
rs
Roo
tan
dsh
oot
Sim
ard
etal
.19
97a
EC
M;
root
box
inla
b
Pul
sed
14C
and
13C
;bi
oass
ayP.
men
zies
ii,
B.
papy
rife
ra;
fiel
dso
il
28- µ
mm
esh
barr
ier
EC
Man
dso
ilpa
thw
ayno
tdi
stin
guis
habl
e
Pre
senc
eof
hyph
alli
nks
test
edbu
tno
effe
ctfo
und
(i)
Two-
way
tran
s-fe
r;(i
i)no
net
tran
sfer
Two-
way
tran
sfer
was
5%of
tota
lis
otop
ein
dono
rs
Roo
tan
dsh
oot
Sim
ard
etal
.19
97b
EC
M;
root
box
inla
b
Pul
sed
13C
;bi
oass
ayP.
men
zies
ii,
B.
papy
rife
ra;
fiel
dso
il
Non
eN
otkn
own
Non
ete
sted
(i)
One
-way
tran
s-fe
rto
P.m
enzi
esii
(tra
nsfe
rin
reve
rse
dire
ctio
nno
tte
sted
)
One
-way
tran
sfer
was
4.7%
ofis
otop
ein
dono
r
Roo
tan
dsh
oot
Sim
ard
etal
.19
97c
EC
M;
root
box
inla
b
Pul
sed
14C
;au
tora
diog
raph
yP
inus
dens
iflo
rapa
irs;
Pis
olit
hus
tinc
tori
us,
T01
Non
myc
orrh
izal
cont
rol
1°:
EC
MP
rese
nce
ofhy
phal
link
s(i
)O
ne-w
ayan
dtw
o-w
aytr
ans-
fer;
(ii)
isot
ope
mov
emen
tfr
omba
seto
apex
ofm
ycel
ium
mor
era
pid
than
apex
toba
se
Not
dete
rmin
edF
unga
lW
uet
al.
2001
AM
;ro
otbo
xin
lab
Nat
ural
abun
danc
e13
C;
bioa
ssay
Pla
ntag
ola
nceo
lata
,C
ynod
onda
ctyl
on;
nativ
ero
otfr
agm
ents
Dif
fere
nt13
Cdi
scri
min
atio
nbe
twee
nC
3an
dC
4pl
ants
;28
- µm
and
0.45
- µm
mes
hba
rrie
rs
1°:
AM
;2°
:so
ilP
rese
nce
ofhy
phal
link
s(i
)O
ne-w
aytr
ans-
fer
toC
ynod
onda
ctyl
on(t
rans
-fe
rin
reve
rse
dire
ctio
nno
tde
term
ined
);(i
i)on
e-w
aytr
ansf
erto
P.la
nceo
lata
Typi
call
y0%
–10%
(up
to41
%)
ofC
inre
ceiv
erro
ots
deri
ved
from
dono
r
Roo
tan
dfu
ngal
ofC
.da
ctyl
on;
root
and
shoo
tof
P.la
nceo
lata
Wat
kins
etal
.19
96
AM
;ro
otbo
xin
lab
Pul
sed-
depl
eted
13C
;bi
oass
ayFe
stuc
aov
ina
turf
pair
s;fo
urG
lom
ussp
.
Non
myc
orrh
izal
cont
rol
1°:
AM
Pre
senc
eof
hyph
alli
nks
One
-way
and
two-
way
tran
sfer
One
-way
tran
sfer
40%
ofis
otop
ein
dono
r
Roo
tan
dfu
ngal
Gra
ves
etal
.19
97
Tab
le2.
Cha
ract
eris
tics
ofca
rbon
tran
sfer
inE
CM
and
AM
syst
ems
mea
sure
din
rece
ntfi
eld
and
labo
rato
ryst
udie
s.

© 2004 NRC Canada
Simard and Durall 1147
Myc
orrh
izal
type
;st
udy
cond
itio
nsIs
otop
esu
ppli
ed;
anal
ysis
met
hod
Pla
ntsp
ecie
spa
irs;
fung
alin
ocul
ant
Met
hod
used
tose
para
tefu
ngal
from
soil
path
way
Tra
nsfe
rpa
thw
ayfo
und
Fact
orfo
und
toin
flue
nce
tran
sfer
Type
oftr
ansf
erfo
und
and
othe
rre
leva
ntre
sult
sM
easu
reof
amou
nttr
ansf
erre
dR
ecei
ver
tiss
ueR
efer
ence
AM
;ro
otbo
xin
lab
Nat
ural
-ab
unda
nce
13C
;bi
oass
ayP
lant
ago
lanc
eola
ta,
Cyn
odon
dact
ylon
;G
lom
usm
osse
ae,
UY
21
Dif
fere
nt13
Cdi
scri
min
atio
nbe
twee
nC
3an
dC
4pl
ants
;28
- µm
and
0.45
- µm
mes
hba
rrie
rs
1°:
AM
;2°
:so
ilS
ink
and
sour
cest
reng
thal
tere
dby
clip
ping
rece
iver
shoo
tsan
del
evat
ing
CO
2,bu
tno
effe
ct
One
-way
and
two-
way
tran
sfer
App
rox.
5%–1
5%of
root
Cfr
omtr
ansf
er,
wit
hm
ore
inP
lant
ago
lanc
eola
ta
Roo
tan
dfu
ngal
Fit
ter
etal
.19
98
AM
;m
icro
-co
sms
inla
b
Pul
sed
14C
bioa
ssay
;au
tora
diog
raph
y
Cor
rall
orhi
zatr
ifid
a,B
etul
ape
ndul
a;Sa
lix
repe
ns,
Pin
ussy
lves
tris
;fi
eld
soil
Mes
hba
gsan
dno
nmyc
orrh
izal
cont
rols
1°:
AM
link
s(E
CM
onau
totr
ophs
)
Pre
senc
eof
hyph
alli
nks
One
-way
tran
sfer
toC
orra
llor
hiza
trif
ida
from
both
B.
pend
ula
and
S.re
pens
,bu
tno
tP
inus
sylv
estr
is
One
-way
tran
sfer
toC
orra
llor
hiza
trif
ida
was
0.31
%–0
.38%
ofis
otop
ein
dono
rs;
itga
ined
6%–1
4%m
ass
ofw
hen
link
edto
auto
trop
hsan
dlo
st13
%w
hen
not
Cor
allo
idrh
izom
eM
cKen
dric
ket
al.
2000
AM
;fi
bre
pots
infi
eld
Pul
sed
14C
;bi
oass
ayA
cer
sacc
haru
m,
Ery
thro
nium
amer
ican
um;
fiel
dso
il
Bet
ula
alle
ghan
iens
is(E
CM
cont
rol)
1°:
AM
;2°
:so
ilP
heno
logy
ofdo
nor
and
rece
iver
plan
ts
(i)
One
-way
tran
s-fe
r;(i
i)tr
ansf
erre
vers
edbe
twee
nsp
ring
and
fall
,co
rres
pond
ing
wit
hC
sour
ce–
sink
patt
erns
info
liat
ion
One
-way
tran
sfer
toA
.sa
ccha
rum
was
1.6%
ofle
afC
dem
and
Roo
tan
dsh
oot
Ler
atet
al.
2002
AM
;sp
lit
Pet
ridi
shin
lab
13C
and
15N
NM
R;
gas
chro
mat
ogra
ph,
mas
ssp
ectr
os-
copy
anal
ysis
ofpr
otei
nex
trac
t
Dau
cus
caro
ta;
Glo
mus
intr
arad
ices
Roo
t-fr
eeco
mpa
rtm
ent
AM
Non
eT
rans
ferr
ed13
Cre
-m
aine
din
rece
iver
fung
alpo
ols
(tre
halo
se,
lipi
ds),
whi
le15
Nfo
und
inre
-ce
iver
root
prot
eins
naF
unga
lP.
E.
Pfe
ffer
,pe
rson
alco
mm
unic
atio
n
Not
e:A
M,
arbu
scul
arm
ycor
rhiz
a;E
CM
,ec
tom
ycor
rhiz
a;na
,no
tap
plic
able
.
Tab
le2
(con
clud
ed).

However, Simard et al. (1997a) showed that isotope re-ceived by both tree species was well above detection andcalculation error limits in all experiments, and Simard et al.(1997a, 1997d, 1997e) showed that large amounts of isotopewere transferred in both directions in both the first-year fieldand laboratory studies, exceeding one-way transfer consid-ered significant in other systems (e.g., Read et al. 1985;Finlay and Read 1986b). Thirdly, Robinson and Fitter(1999) suggested that [13CO2] far exceeded [14CO2] in ourfield labeling chambers, possibly altering C assimilation, al-location, and interplant flux patterns. However, out-plantedB. papyrifera and P. menziesii were pulsed with similar12 + 13CO2 (0.05%) and 12 + 14CO2 (0.03%) concentrations(Simard et al. 1997a), resulting in the same 13C and 14C tis-sue allocation patterns (Simard et al. 1997a). This findingwas supported by another study by Simard et al. (1997e),who found no effect of increasing 12 + 13CO2 concentrationsfrom 0.04% to 0.05% on 13C allocation patterns. Fourthly,Robinson and Fitter (1999) and Fitter (2001) argued thatSimard et al. (1997a) inferred a mutualistic plant–fungus–plant relationship from their results and that they had specu-lated that competition interactions be replaced by mutualismas the dominant interaction shaping plant communities.While these speculations may have been made in otherpapers, Simard et al. (1997a) carefully stated that the netcompetitive effect of one species on another should not bepredicted without an understanding of interplant transferthrough hyphal and soil pathways (i.e., model pathways 1, 2,and 3) and in so doing recognized the primacy of competi-tive interactions in shaping plant community dynamics. Ourstudies were insufficient, however, to determine the impor-tance of C transfer relative to other competitive interactionsin plant community dynamics. Nevertheless, the magnitudeof net transfer we measured was in the range considered suf-ficient to increase survival and growth of interconnectedramets in clonal plants (Alpert et al. 1991). This suggests tous that C transfer has the potential to influence plant interac-tions some time during community development, but longerterm, broader scope field research is needed before we canquantify this effect.
Following the ECM studies of Simard and associates, Wuet al. (2001) carefully examined the pathway, movement,and fate of 14C transferred between ECM-connected Pinusdensiflora Sieb. & Zucc. seedling pairs using time-courseautoradiography in laboratory microcosms. They demon-strated unequivocally that 14C was transferred from ECMdonor plants to the mycelia and ECM of receiver seedlingswithin 3 d. They also found that 14C moved bidirectionallywithin the mycelia, with the greatest transfer through densemycelial fans of one fungal species and hyphal strands ofanother. Hyphal strands of ECM had earlier been hypothe-sized as the most important conduits of transferred C(Cairney and Smith 1992), but this study confirms thathyphae not associated with rhizomorphs can also be an im-portant pathway.
In contrast to many of the earlier C-transfer studies in-volving ectomycorrhizae (Bjorkman 1960; Brownlee et al.1983; Francis and Read 1984; Read et al. 1985; Finlay andRead 1986b; Arnebrant et al. 1993; Simard et al. 1997a;Simard et al. 1997d; Simard et al. 1997e), Wu et al. (2001)found no transfer into root or shoot tissues of receiver
plants. Reasons for the inconsistency in root-to-shoot Ctransfer are not understood, but Wu et al. (2001) differedfrom the earlier studies in that shoots of receiver plants werewrapped in aluminum foil during the chase period to preventrefixation of respired 14CO2, whereas most of the other stud-ies used equidistant control plants to quantify and subtractrefixed 14CO2, leaving receiver plants in an unaltered physi-ological condition. By preventing fixation and respiration ofreceiver shoots, Wu et al. (2001) may have weakened thesink strength of plant tissues for organic compounds. Addi-tionally, the use of radiography to detect C transfer mayhave underestimated C transfer into pine shoots because it isless sensitive than the tissue oxidation method commonlyused by others (e.g., Simard et al. 1997a; Lerat et al. 2002).
Some have argued that C movement to shoots in earlierECM studies may have actually followed transfer betweennonmycorrhizal rather than between mycorrhizal roots (Wuet al. 2001; Fitter et al. 2001). These authors also recog-nized, however, that previous research demonstrated move-ment of isotopically labeled organic N compounds from themycorrhizal fungus to plant tissue in ectomycorrhizal plants(Arnebrant et al. 1993; Cliquet and Stewart 1993; Finlay etal. 1989; Chalot and Brun 1998; Näsholm et al. 1998) andmyco-heterotrophic plants (McKendrick et al. 2000). Amechanism for C transfer into receiver tissue has been pro-posed, where sugar or organic N is transported from the do-nor plant to the fungus, low molecular weight organic Nmoves from the donor to receiver plant through the intercon-necting fungus along a hydrostatic pressure gradient fromhigh to low assimilate and N concentrations, and the organiccompound is then actively transported across the plant–fungal membrane into plant tissues (Smith and Smith 1990;Martin et al. 1997; Chalot and Brun 1998). Whether trans-ferred C moves into receiver plant tissues or remains in fun-gal tissues is still under debate, and this has been proposedas the key piece of evidence that would underscore the func-tional significance of interplant transfer in ecological inter-actions (Fitter et al. 1999; Robinson and Fitter 1999). Weand others argue, however, that C transferred to the ectomy-corrhizal fungi of the receiver plant, even without transferinto plant tissues, is still a subsidy to the nutrient-gatheringsystem of the plant (Perry 1998; Simard et al. 2002) that po-tentially can affect plant community dynamics (Bever 2003).
C transfer between arbuscular mycorrhizal plantsRecent studies in AM systems are providing increasing
evidence that the pattern of interplant C transfer contrastswith that in ECM systems. These differences in part have fu-elled the aforementioned debates. Several laboratory studieshave demonstrated C movement from one AM plant to an-other (Francis and Read 1984; Martins 1993; Watkins et al.1996), but none have demonstrated net transfer (Perry 1999),and most have found little (Francis and Read 1984; Read etal. 1985) or no movement of transferred C from roots toshoots (Waters and Borowicz 1994; Watkins et al. 1996;Graves et al. 1997; Fitter et al. 1998), because it appearedto remain associated with fungal tissue (Fitter et al. 1999;P.E. Pfeffer, personal communication). Watkins et al. (1996)used differences in natural abundance of 13C to quantify Ctransfer between laboratory-grown C3 (Plantago lanceo-lata L.) and C4 (Cynodon dactylon (L.) Pers.) plants, as well
© 2004 NRC Canada
1148 Can. J. Bot. Vol. 82, 2004

as different-sized mesh barriers, to distinguish AM fromsoil-transfer pathways and showed that one-way transfer toCynodon dactylon was generally <10% of donor isotopecontent, ranging between 0% and 41%. They also found thattransfer between potentially linked plants far exceeded thatof unlinked plants and most transferred C remained in themycorrhizal tissues of Cynodon dactylon, but that mosttransferred C moved into both roots and shoots of Plantagolanceolata. Fitter et al. (1998) further tested the fate oftransferred C in Cynodon dactylon and Plantago lanceolataby examining the regrowth of clipped receiver plants, con-firming that all transferred C remained in mycorrhizal struc-tures. They also showed that the extent of transfer waspositively correlated with fungal vesicle abundance, nothyphal abundance (Fitter 2001), underscoring differenceswith ECM transfer patterns shown by Wu et al. (2001).These results were corroborated by Graves et al. (1997),who fumigated sections of mycorrhizal and nonmycorrhizalC4 Festuca ovina S. Wats. turf with 13C-depleted air andshowed that 41% of C was transferred from donor roots tolinked neighbours, most transferred C remained in donormycorrhizal structures, and no C was transferred to unlinkedneighbours. The lack of transferred C movement from AMfungal tissues into associated plant tissues is supported bythe recent work of P.E. Pfeffer (personal communication),who showed that 15N substrates supplied in vitro toextraradical mycelium (ERM) of AM carrot-root cultures re-sulted in substantial labeling of root proteins, but that 13Csubstrates supplied to the ERM entered only fungal com-pounds (trehalose and storage lipids) and not host root me-tabolites. Similar to Wu et al. (2001), however, sink strengthof receiver plants may have been substantially altered by ex-cising the carrot shoots and placing the roots in a Petri dish.
A recent field study that investigated interplant C transferin an AM Acer saccharum Marsh. forest in Quebec providesdivergent results to the AM laboratory, greenhouse, and invitro studies described previously. Lerat et al. (2002) grewA. saccharum, Erythronium americanum Ker-Gawl., and, asa control, ECM Betula alleghaniesis Britt. individuals to-gether in fibre pots in the forest understory. In spring, theylabeled fully expanded E. americanum leaves with 14C whileA. saccharum overstory and sapling leaves were still leaf-less, but were bursting bud, and followed this with a falllabeling of fully expanded A. saccharum foliage, afterE. americanum leaves had been shed. In the spring, 0.064%of the fixed label in donor E. americanum was transferredinto A. saccharum, with 13 times less transferred into Betulacontrol plants. Although the amount transferred was small, itwas equivalent to 1.6% of leaf expansion costs for the maplesaplings, which the authors suggest is a conservative esti-mate, considering that maple saplings would be connectedto several E. americanum plants in the undisturbed forest.Autoradiographs clearly showed that 14C had moved intoroots, stems, and leaves of receiver maple seedlings. In thefall labeling, there was evidence that the direction of transferhad reversed, with 14C moving from A. saccharum saplingsto 7 of 22 E. americanum mycorrhizal root systems andnone moving to Betula controls. The lack of radioactivity inbirch indicated that transfer through soil pool pathways wasinsignificant. The pattern of C transfer was consistent withsource–sink relationships between donor and receiver plants
during spring and fall seasons. Fully developed E. ameri-canum were strong sources and newly developing mapleleaves were strong sinks for carbohydrates in spring,whereas defoliated E. americanum roots became sinks fornew maple-leaf assimilate in the fall. This study provides thefirst evidence that the direction and amount of C transfer canchange with season, emphasizing that one-time measure-ments are inadequate for evaluating the significance oftransfer to plant fitness where receivers and donors havenonsynchronous phenologies.
Reasons why transfer patterns in AM systems differ fromthose in EM systems have yet to be fully explored. Severalmorphological, physiological, and genetic factors may be in-volved, including AM and EM differences at the fungus, in-terface, root, plant-species, and ecosystem levels. Some ofthese differences may be controlled by examining transferamong plant species forming both ECM and AM associa-tions within the same ecosystem. A striking difference isalso emerging in some field versus laboratory or greenhousestudies for both ECM (e.g., Simard et al. (1997a) vs. Wu etal.(2001)) and AM systems (e.g., Lerat et al. (2002) vs. Fit-ter et al. (1998)). There are numerous conditions that differbetween the field and laboratory or greenhouse, including al-terations in source–sink patterns in plant combinations andsimplification of the CMN. These factors should be exam-ined to improve our understanding of the mechanisms andsignificance of C transfer in nature.
Factors regulating C transferStudies that have investigated factors influencing the
transfer of C among interconnected plants have focused onthree areas: source–sink relationships between donor and re-ceiver plants, sink patterns of fungi making up the CMN,and perturbations to the CMN. The original pioneering labo-ratory studies by Read and coworkers showed that theamount of C transferred among interconnected AM or ECMplants increased with shading of the receiver, suggesting thatC moved between plants along a source–sink gradient of acurrent C assimilate (Francis and Read 1984; Read et al.1985; Finlay and Read 1986a). These studies were sup-ported by the field study of Simard et al. (1997a), whoshowed that net transfer from ECM B. papyrifera to P. men-ziesii increased twofold when P. menziesii was deeplyshaded. New corroborating evidence for plant-drivensource–sink regulation of transfer comes from Lerat et al.(2002), who showed that seasonal changes in source–sinkgradients driven by shifting plant phenologies affected thedirection of C transfer between AM plants. In another study,Waters and Borowicz (1994) altered sink strength of AM-linked plants by clipping one of the partners and found thatC flowed away from clipped plants toward unclipped plants,the opposite of that expected. They reasoned, however, thatclipping the plants increased labile C concentration in theroots (i.e., turned them into source plants), thereby increas-ing the diffusion gradient for C out of roots into mycorrhizalfungi and into connected, neighbouring, unclipped plants. Incontrast to sink-strength manipulations, Fitter et al. (1998)found that increasing source strength by growing plants inenriched CO2 environments had no effect on the occurrenceor extent of C transfer. Other experiments examined transferbetween plants that naturally differed in C physiology or
© 2004 NRC Canada
Simard and Durall 1149

mycorrhizal dependency (Grime et al. 1987; van der Heijden2002).
The form of C transferred between interconnected plantsis important for understanding the mechanism of interplanttransfer and how C compounds are used by receiver plants.Trehalose, mannitol, and arabitol are major carbohydratesassociated with mycelium interconnecting ECM plants(Söderström et al. 1988), and trehalose is important in myce-lium of AM plants (Lammers et al. 2001; P.E. Pfeffer, per-sonal communication). The occurrence and concentration ofthese compounds can differ between extramatrical mycelium(EMM) and fungal tissue of ECM. For example, mannitolis found in free-living mycelium of Pisolithus tinctorius(Mich.: Pers.) Coker & Couch, whereas trehalose tends toaccumulate in Pisolithus tinctorius ECM (Martin et al.1998). Furthermore, trehalose predominates in ECMbasidiomycetes, whereas mannitol is commonly found as asoluble form of C in ECM ascomycete tissue (Martin 1985;Martin et al. 1988; Ceccaroli et al. 2003). Source–sink gradi-ents of these compounds likely play a role in movement ofcarbohydrates within the CMN, but it is unknown whetherthese gradients are important in moving C via the commonmycorrhizal network from one plant to another. Free aminoacids also represent an important sink of assimilated C inectomycorrhizal fungi (Martin et al. 1988; Ceccaroli et al.2003). N source–sink gradients may be as much or more im-portant than C assimilate gradients in moving C from oneplant to another, because there is greater evidence that or-ganic N compounds transfer more readily from fungal toplant tissues than do carbohydrates. Amino acids or low-weight nitrogenous compounds have been shown to passfrom mycorrhizal fungi into host-plant tissues (Finlay 1989;Cliquet and Stewart 1993; Arnebrant et al. 1993; Näsholm etal. 1998). Although slow movement of carbohydrates fromthe fungal mycobiont to the plant symbiont has been demon-strated in excised roots (Lewis and Harley 1965), other stud-ies have failed to demonstrate this (Smith and Smith 1990).In addition to carbohydrates and amino acids, lipids, such astriacylglycerides, may play an important role in the move-ment of C in the ERM of AM plants (Pfeffer et al. 1999).There is a great need to identify compounds that are trans-ferred through the CMN and into mycorrhizal and host-planttissues and to trace their movement using biochemical andmolecular markers. This would help identify the mechanismfor simultaneous C and nutrient exchange between intercon-nected plants and fungi.
There is increasing evidence that mycorrhizal fungi alsoplay a role in controlling the direction, magnitude, and rateof C or nutrient transfer between linked plants, particularlyin AM systems. Although the ability of the fungi to seques-ter and provide nutrients and C to linked plants at differentrates and amounts has largely been unexplored (Miller andAllen 1992), different characteristics of the CMN have re-cently been shown to influence interplant transfer. For exam-ple, Martensson et al. (1998) found great variation in Ntransfer from bean to chicory among different linking iso-lates of Glomus spp., which they suggested reflected differ-ences in the isolates’ capacity to initiate a plant trigger for Ntransfer. In other studies, hyphal links have been shown toenhance herbicide injury to weeds, possibly because AMfungi adjusted their own source–sink relations in favour of
sink-driven fluxes (Bethlenfalvay et al. 1996b; Rejon et al.1997). Several studies provide evidence that magnitudeof transfer is affected by the degree of physical root–rhizosphere overlap or frequency of mycorrhizal links(Johansen and Jensen 1996; Walter et al. 1996; Simard et al.1997a). By contrast, Watkins et al. (1996) and Fitter et al.(1998) found little relationship between C transfer and thedegree or direction of AM colonization and little evidencethat colonization was related to number or functioning oflinks. Additional fungal factors that appear to play a rolein transfer include identity of the fungal species, seasonalor phenological variation in mycorrhizal activity (Frey andSchüepp 1992; Hamel et al. 1992), as well as differentialmycorrhizal dependency of plant species in mixtures (Zhu etal. 2000; van der Heijden 2002).
Factors that may disrupt the CMN and transferSoil animals, such as collembola and earthworms, feed on
the hyphae and spores of mycorrhizal fungi (Finlay 1985;Klironomos and Moutoglis 1999; Tuffen et al. 2002).Collembola can reduce the number of AM spores found insoil and in some cases reduce external hyphal length of AMfungi (Larsen et al. 1996). In cases where collembola havehad no effect or resulted in an increase in AM hyphal length(Klironomos and Kendrick 1995), interconnections betweenAM fungi and plants may still have occurred. For example,collembola have been found to sever hyphal branches with-out shortening hyphal length (Klironomos and Ursic 1998).These studies suggest that soil animals may have a profoundeffect on the presence, size, and function of the CMN.Tuffen et al. (2002), using AM-linked leek plants in a labo-ratory microcosm experiment, showed that increased earth-worm grazing of the AM mycelium led to enhancedavailability of 32P to a receiver plant. Thus, transfer of 32Pfrom one plant to another was greater when earthworms hadcomminuted the CMN (pathway 2b in Fig. 1a) than when anintact CMN was connecting the two plants (pathway 2a inFig. 1a). This lab study demonstrates that soil animals areable to influence the functioning of a CMN, but we awaitfield results to see if similar effects occur in nature.
Few studies have investigated the influence of soil animalson EMM of ECM fungi. Bonkowski et al. (2001) usedPaxillus involutus (Batsch: Fr.) Fr. in a laboratory micro-cosm study to show that ECM hyphal length was reducedwhen protozoa were present compared with when they wereabsent. This study confirms the findings from AM studiesthat soil animals can disrupt the mycelial networks; however,much more evidence under natural conditions is needed. Inaddition to soil animals, other factors that may limit the sizeand function of CMNs include drought, natural disturbances(e.g., fire, flooding), and anthropogenic disturbances (e.g.,ploughing, site preparation, harvesting). These perturbationsform gaps in the plant community and can directly or indi-rectly disrupt the CMN (Jones et al. 2003).
CMN influences on feedbacks among plants and fungiWe have presented evidence that C transferring from one
plant to another may enter the host root or shoot tissue(Watkins et al. 1996; Simard et al. 1997a; Lerat et al. 2002)or, alternatively, remain within the fungal tissues of theirmycorrhizal associations (Graves et al. 1997; Fitter et al.
© 2004 NRC Canada
1150 Can. J. Bot. Vol. 82, 2004

1998; Wu et al. 2001; P.E. Pfeffer, personal communica-tion). Even where transferred C remains in mycorrhizal fun-gal tissues and is not transferred into receiver plant tissues,we suggest there may be a net benefit to the plant, becausethe CMN is subsidized, reducing C demand by the plant’sfungal symbionts (Wilkinson 1998; Amaranthus et al. 1999;Fitter 2001; Zabinski et al. 2002; Simard et al. 2002). Withor without C transfer to plant tissues, a CMN is likely tohave important influences on feedback among and betweenplants and mycorrhizal fungi (Bever 2003). Based on theconcept that growth rates and fitness of plants and soil mi-crobes are interdependent, Bever et al. (1997) used a model-ing approach to predict the structure and diversity of plantand microbial communities, later extending the models tofocus specifically on relationships between mycorrhizalfungi and host plants (Bever 2003). These models stronglysuggest that the CMN has the potential to influence structureand diversity of the whole plant–microbe community,whether or not C transfer occurs, or whether transferred Cmoves into receiver plant tissues.
We can use the models developed by Bever (2003) toillustrate potential feedbacks among plants and mycorrhizalfungi resulting from the transfer pathways shown in Fig. 1a.Understanding these feedbacks may help us predict the ef-fects of CMNs and C transfer on plant community dynam-ics, a field of research that we discuss later in this paper. Inour nonmycorrhizal soil pathway (pathway 1), the simplestmodel is where plants compete for resources in the soil pool(Tilman and Pacala 1993). In that case, interspecific differ-ences in nutrient or water demand and uptake, as well as in-trinsic growth rates, would determine which plant specieswere favored in the plant community (Fig. 1b). Even in thesimplest plant systems, however, the additional importanceof facilitative between-plant interactions (e.g., frost protec-tion, sunscald protection, reduced herbivory) in structuringplant communities is being increasingly recognized (Perryet al. 1989; Burton et al. 1992; Callaway 1995; Howe etal. 2002). Facilitative relationships between plants and mi-crobes, such as the mycorrhizal association, are also increas-ingly being recognized as important determinants of plantcommunity dynamics (van der Heijden et al. 1998; Bever etal. 2002; DeLong et al. 2002).
The mycorrhizal–soil-transfer pathway (pathway 3) shownin Fig. 1a, which involves specific mycorrhizal relationshipswith host plants, can have complex effects on plant dynam-ics (see mutualistic models in Bever (2003)). In symmetricrelationships between two plants and their mycorrhizalfungi, in which the host-specific fungus delivers the greatestbenefit to the host plant, there is a positive feedback dy-namic (Fig. 2a). One plant–fungal combination is reinforcedmore than the other in the community, leading to eventuallocal extinction of other types. Dominance of grasslands byexotic invading plants, such as Centaurea L. species that arefavored by native AM fungi, is one example of a positivefeedback (Marler et al. 1999; Klironomos 2002). Asymmet-ric relationships can also occur, however, where an AMfungi that has the greatest benefit to a particular plant mayalso result in a greater growth rate of a second plant species(i.e., cosmopolitan specificity). This results in a sustaineddynamic between the plants and AMF community, where nosingle plant species is able to replace the other, leading to in-
creased plant community diversity. Examples of negativefeedback (Fig. 2b) are numerous (e.g., increasing density-dependent mortality in conspecific patches in tropical forests(Harms et al. 2000)) and are considered more common thanpositive-feedback relationships in nature (Bever 2003). Be-cause mycorrhizal associations range from predominantlylow specificity (e.g., AM) to, less commonly, high specific-ity (e.g., orchid mycorrhizae), with most fungi able to colo-nize and thrive on a number of host-plant species (Molina etal. 1992), there exists a mechanism for negative feedbackwithin mycorrhizal plant communities.
The potential coexistence of the direct CMN transfer path-way (pathway 2a, Fig. 1a) with pathways 1, 2b, and 3 canlead to increasing complexity of potential feedbacks amongplants and fungi and less predictable effects on plant com-munity dynamics. This complexity will increase as the num-ber of fungi and plants involved in the CMN increase and asthe interaction strengths between the plants and shared fungichange as a result of successive feedbacks. Taking a reduc-tionist approach to pathway 2a, however, we can see thatseveral positive- and negative-feedback models are possiblein a simple system containing two plants and one sharedfungus (Fig. 3). In the simplest case (Fig. 3a), there is a con-stant host C / fungal nutrient-exchange ratio between the twoplants, where both plants supply the fungus with the sameamount of C in return for equal benefits from the fungus(e.g., equal amounts of organic nutrients). Feedback betweeneach host and fungus is positive, but both plants coexist be-cause there is no net benefit or net loss (e.g., no net transferbetween plants) to either (i.e., the shared fungus does not al-ter the interaction between the two species). One example ofthis is provided by Simard et al. (1997d), where same-sizedEM paper birch and Douglas-fir grown in laboratory root-boxes exchanged C below ground in equivalent amounts, re-sulting in no net C gain by either species. If we alter themagnitude of host C and fungal nutrient exchange betweenhost plants A and B, but don’t alter the exchange ratio be-tween linked plants, no net effect on the plant communityis also expected (Fig. 3b). This could occur in a self-
© 2004 NRC Canada
Simard and Durall 1151
Fig. 2. Two potential consequences of the mutual interdepen-dence of plant and mycorrhiza fungal growth rates. The directionof benefit between the two plant species, A and B, and the twofungal species, X and Y, is depicted with the direction of the ar-rows. Thickness of the arrows represents the degree of the bene-fit received, with a thick line meaning greater benefit than a thinone. (a) The symmetric fitness relation between the plants andfungi results in a positive feedback dynamic. (b) The highlyasymmetric fitness relations between the plants and fungi wouldresult in a negative-feedback dynamic. See text for more detailedexplanation. (From Bever et al. 1997., reproduced with permis-sion of J. Ecol., Vol. 85, p. 568, © 1997 Blackwell ScientificPublications Ltd.)

regenerating stand of conspecific trees, where trees are ofdifferent sizes but their C physiologies and nutrient demandsare proportionally the same (Jonsson et al. 2001). A net ben-efit to a small tree is still possible, even though there is no Ccost to the adult or C gain by the small tree (i.e., no nettransfer), because it has established within a CMN accessinga large soil-nutrient pool (sensu Zabinski et al. 2002).
However, host C / fungal nutrient-exchange ratios, andhence interactions between host plants and a shared fungus,are not always homogenous. Therefore, feedback from theCMN can be expected to affect growth rates of species Aand B differently. Plant A may have a higher ratio of plantC / fungal organic nutrient exchange than plant B, for exam-ple, resulting in net C transfer from plant A to B (or net“benefit” to B if we consider the more general case of inor-ganic nutrients) (Fig. 3c). When Douglas-fir and paper birch
were grown together for 2 years in the field, for example,there was net C transfer from the more photosyntheticallyactive paper birch to shaded Douglas-fir (Simard et al.1997a). Transfer occurred along a source–sink gradient forC that was driven by plant-species differences in C assimila-tion rates and organic nutrient demands. These source–sinkor interplant C – fungal nutrient relationships can changedramatically over time depending on the phenology of thelinked plants (Fig. 3d). Lerat et al. (2002), for example,found that foliated trout lily was a C source for maple seed-lings in an emerging maple canopy in spring, but that defoli-ated trout-lily corms were a C sink in the fall, when themaple canopy was fully expanded (Lerat et al. 2002). Themost extreme case of direct C transfer occurs from auto-trophic trees to myco-heterotrophic plants through specificshared mycorrhizal symbionts (McKendrick et al. 2000)
© 2004 NRC Canada
1152 Can. J. Bot. Vol. 82, 2004
Fig. 3. Feedbacks and direct transfer between plants A and B through the ECM hyphal pathway, where fungus Z represents a CMN.Arrows pointing to the plant represent flow of organic nutrients, whereas arrows pointing to the fungus represent carbon (C) flow. Dif-ferences in size of arrows are explained in the caption for Fig. 3. (a) Plant A and plant B have similar plant C / fungal nutrient-exchange ratios. (b) C / fungal nutrient-exchange ratios of plants A and B are the same, but the magnitude is different, resulting inbenefit to both but no net transfer. In (c) and (d), plant C / fungal nutrient-exchange ratios differ between A and B, resulting in net Ctransfer. (e) The C / fungal nutrient-exchange ratios differ between plants A and B, but C transfer to the myco-heterotrophic plant(plant B) is negligible to plant A.

(Fig. 3e). Although one-way C transfer from mature trees tothe relatively tiny orchid is an example of positive feedback,theoretically leading to community domination by the re-ceiver plant (i.e., orchids), this does not appear to happen,because of the negligible net cost to the tree (Leake 1994).
Adding more plant and fungal species to the CMN createsa greater array of possible feedbacks and potential effects onplant and fungal community structure and diversity (Bruns etal. 2002b). At the same time, the network can still existwithout all plants being tapped into a common mycorrhizalfungal species and without any direct transfer of C betweenplants. In one relatively simple scenario (Fig. 4), three plants(A, B, C) may be directly and indirectly interconnected bytwo fungi (W, Z). In this example, fungus Z connects plantsA and B; fungus W connects plants B and C. Thus, eventhough plants A and C are not associated with the samemycelium, they still can be considered part of the largermycorrhizal network. At a functional level, the interactionbetween plant C and fungus W may be indirectly affectingplant A through the mutual association with plant B. For ex-ample, plant C could be allocating large amounts of C belowground to fungus W, reducing plant B’s need to allocate C tofungus W and indirectly allowing plant B to contribute tothe C demand imposed by fungus Z, thereby reducing the re-quirement of plant A to provide C to fungus Z (Fig. 4). Thisscenario would occur without any interplant transfer. Al-though there is no evidence supporting the occurrence of thisscenario in nature, it demonstrates the complexity of interac-tions that could exist between different plant species andtheir CMN. It is important to remember that this is a rela-tively simple scenario. In nature, feedbacks among plantsand fungi are much more complex and community effectsmuch less predictable because there exists an enormous po-tential for many plants to be interlinked by many differentshared fungi, all with contrasting and shifting C / nutrient-exchange ratios. These are further affected by additionalfeedbacks generated in pathways 1 and 2.
CMN influences on plant community dynamicsMany studies have discussed the implications of plants
tapping into an existing CMN, and increasing their access toa larger pool of host-derived C or soil nutrients, on plantpopulation or community dynamics. However, almost allstudies suffer from inadequate measurement of C transfer orinclude inadequate controls that separate CMN from soilpathways, leaving unanswered questions about the relativeimportance of CMNs, or of C transfer through alternativepathways, on plant community development. As a result, wefeel the field has great potential for further study. Futurestudies should include (i) demonstration that hyphal linksactually exist in the field, (ii) establishment of adequate con-trols, so that CMN effects can be separated from non-CMNeffects, (iii) measurement of bidirectional or net transfer ofC, nutrients, or water among mycorrhizal plants that arelinked by the CMN, (iv) quantification of changes in themagnitude and duration of CMN effects or transfer overextended periods of time and over critical periods of plantdevelopment, and (v) measurements of CMN and (or) trans-fer effects on plant survival, growth, and fitness. Until theseresearch needs are met, the importance of CMNs and C
transfer to plant population and community dynamics willremain unknown.
The potential influences of the CMN on plant communitydynamics have been discussed in several earlier review pa-pers (Newman 1988; Miller and Allen 1992; Perry et al.1992; Amaranthus and Perry 1994; Zobel and Moora 1997;Fitter 2001; Simard et al. 2002). In the most recent review,Simard et al. (2002) summarized six possible effects onplant communities and ecosystems that had previously beenproposed in the literature, citing several papers that sup-ported or contradicted the hypotheses. These were that theCMN may (i) assist with seedling establishment near matureplants by allowing seedlings to become colonized more rap-idly or tap into an established CMN supported by otherplants; (ii) assist with recovery of species following distur-bance, which is particularly important where disturbancebehavior is unpredictable; (iii) reduce competition intensityand promote species diversity by allowing C or nutrients todirectly flow from sufficient to deficient plants, resulting ina more even distribution of C or nutrients; (iv) affect fungalnutrient demands, source–sink relationships, foraging anduptake, and therefore fungal ecology; (v) reduce nutrientlosses from ecosystems by keeping more nutrients in livingbiomass; and (vi) increase productivity, stability, and sustain-ability of ecosystems. Most recent research has examinedCMN effects on seedling establishment, competition amongplants, and plant community diversity, and we therefore fo-cus our review of plant community dynamics on those threeareas. In addition, most studies have examined CMN effectson plant communities rather than populations, providing acommunity focus to our review; however, many of the CMNimplications are similar for populations. Recent relevantstudies are summarized in Table 3.
CMN assistance in seedling establishmentIn forests, subalpine parklands, or shrublands, it has been
recognized that seedlings can preferentially establish nearexisting plants of the same or different species, forming is-lands or clumps of regeneration (Perry et al. 1992). Among anumber of hypotheses suggested to explain this phenomenonis one where mycorrhizae of an existing plant communityinfects new seedlings, facilitating their establishment. Theseedlings may benefit in a number of ways: they may be in-
© 2004 NRC Canada
Simard and Durall 1153
Fig. 4. Feedbacks between plants A, B, and C through the ECMhyphal pathway, where fungi Z and W represent a CMN. Thereis no direct interplant C transfer between A and C, but benefit ofplant C to plants A and B still occurs because of benefits to theCMN. Thickness of the arrows represents the degree of the bene-fit received, with a thick line meaning greater benefit than a thinone. See text for detailed explanation.
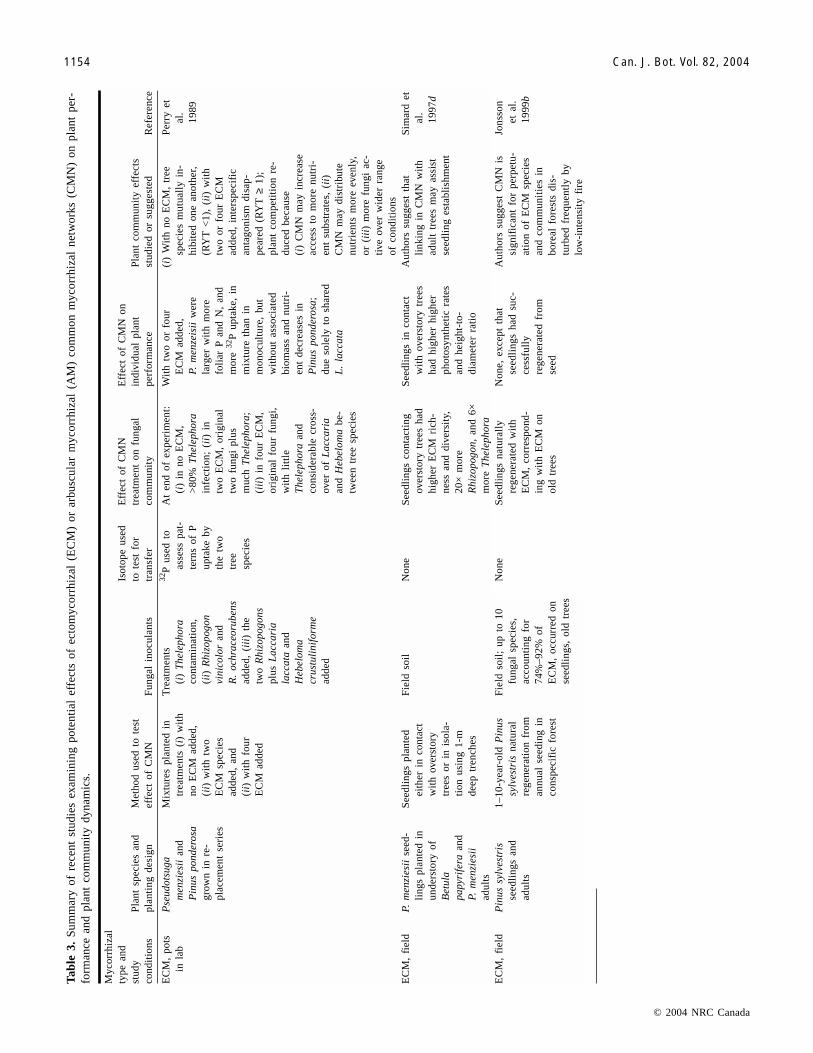
© 2004 NRC Canada
1154 Can. J. Bot. Vol. 82, 2004
Myc
orrh
izal
type
and
stud
yco
ndit
ions
Pla
ntsp
ecie
san
dpl
anti
ngde
sign
Met
hod
used
tote
stef
fect
ofC
MN
Fun
gal
inoc
ulan
ts
Isot
ope
used
tote
stfo
rtr
ansf
er
Eff
ect
ofC
MN
trea
tmen
ton
fung
alco
mm
unit
y
Eff
ect
ofC
MN
onin
divi
dual
plan
tpe
rfor
man
ceP
lant
com
mun
ity
effe
cts
stud
ied
orsu
gges
ted
Ref
eren
ce
EC
M,
pots
inla
bP
seud
otsu
gam
enzi
esii
and
Pin
uspo
nder
osa
grow
nin
re-
plac
emen
tse
ries
Mix
ture
spl
ante
din
trea
tmen
ts(i
)w
ith
noE
CM
adde
d,(i
i)w
ith
two
EC
Msp
ecie
sad
ded,
and
(ii)
wit
hfo
urE
CM
adde
d
Tre
atm
ents
(i)
The
leph
ora
cont
amin
atio
n,(i
i)R
hizo
pogo
nvi
nico
lor
and
R.
ochr
aceo
rube
nsad
ded,
(iii
)th
etw
oR
hizo
pogo
nspl
usL
acca
ria
lacc
ata
and
Heb
elom
acr
ustu
lini
form
ead
ded
32P
used
toas
sess
pat-
tern
sof
Pup
take
byth
etw
otr
eesp
ecie
s
At
end
ofex
peri
men
t:(i
)in
noE
CM
,>
80%
The
leph
ora
infe
ctio
n;(i
i)in
two
EC
M,
orig
inal
two
fung
ipl
usm
uch
The
leph
ora;
(iii
)in
four
EC
M,
orig
inal
four
fung
i,w
ith
litt
leT
hele
phor
aan
dco
nsid
erab
lecr
oss-
over
ofL
acca
ria
and
Heb
elom
abe
-tw
een
tree
spec
ies
Wit
htw
oor
four
EC
Mad
ded,
P.m
enze
isii
wer
ela
rger
wit
hm
ore
foli
arP
and
N,
and
mor
e32
Pup
take
,in
mix
ture
than
inm
onoc
ultu
re,
but
wit
hout
asso
ciat
edbi
omas
san
dnu
tri-
ent
decr
ease
sin
Pin
uspo
nder
osa;
due
sole
lyto
shar
edL
.la
ccat
a
(i)
Wit
hno
EC
M,
tree
spec
ies
mut
uall
yin
-hi
bite
don
ean
othe
r,(R
YT
<1)
,(i
i)w
ith
two
orfo
urE
CM
adde
d,in
ters
peci
fic
anta
goni
smdi
sap-
pear
ed(R
YT
≥1)
;pl
ant
com
peti
tion
re-
duce
dbe
caus
e(i
)C
MN
may
incr
ease
acce
ssto
mor
enu
tri-
ent
subs
trat
es,
(ii)
CM
Nm
aydi
stri
bute
nutr
ient
sm
ore
even
ly,
or(i
ii)
mor
efu
ngi
ac-
tive
over
wid
erra
nge
ofco
ndit
ions
Per
ryet
al.
1989
EC
M,
fiel
dP.
men
zies
iise
ed-
ling
spl
ante
din
unde
rsto
ryof
Bet
ula
papy
rife
raan
dP.
men
zies
iiad
ults
See
dlin
gspl
ante
dei
ther
inco
ntac
tw
ith
over
stor
ytr
ees
orin
isol
a-ti
onus
ing
1-m
deep
tren
ches
Fie
ldso
ilN
one
See
dlin
gsco
ntac
ting
over
stor
ytr
ees
had
high
erE
CM
rich
-ne
ssan
ddi
vers
ity,
20×
mor
eR
hizo
pogo
n,an
d6×
mor
eT
hele
phor
a
See
dlin
gsin
cont
act
wit
hov
erst
ory
tree
sha
dhi
gher
high
erph
otos
ynth
etic
rate
san
dhe
ight
-to-
diam
eter
rati
o
Aut
hors
sugg
est
that
link
ing
inC
MN
wit
had
ult
tree
sm
ayas
sist
seed
ling
esta
blis
hmen
t
Sim
ard
etal
.19
97d
EC
M,
fiel
dP
inus
sylv
estr
isse
edli
ngs
and
adul
ts
1–10
-yea
r-ol
dP
inus
sylv
estr
isna
tura
lre
gene
rati
onfr
oman
nual
seed
ing
inco
nspe
cifi
cfo
rest
Fie
ldso
il;
upto
10fu
ngal
spec
ies,
acco
unti
ngfo
r74
%–9
2%of
EC
M,
occu
rred
onse
edli
ngs,
old
tree
s
Non
eS
eedl
ings
natu
rall
yre
gene
rate
dw
ith
EC
M,
corr
espo
nd-
ing
wit
hE
CM
onol
dtr
ees
Non
e,ex
cept
that
seed
ling
sha
dsu
c-ce
ssfu
lly
rege
nera
ted
from
seed
Aut
hors
sugg
est
CM
Nis
sign
ific
ant
for
perp
etu-
atio
nof
EC
Msp
ecie
san
dco
mm
unit
ies
inbo
real
fore
sts
dis-
turb
edfr
eque
ntly
bylo
w-i
nten
sity
fire
Jons
son
etal
.19
99b
Tab
le3.
Sum
mar
yof
rece
ntst
udie
sex
amin
ing
pote
ntia
lef
fect
sof
ecto
myc
orrh
izal
(EC
M)
orar
busc
ular
myc
orrh
izal
(AM
)co
mm
onm
ycor
rhiz
alne
twor
ks(C
MN
)on
plan
tpe
r-fo
rman
cean
dpl
ant
com
mun
ity
dyna
mic
s.

© 2004 NRC Canada
Simard and Durall 1155
Myc
orrh
izal
type
and
stud
yco
ndit
ions
Pla
ntsp
ecie
san
dpl
anti
ngde
sign
Met
hod
used
tote
stef
fect
ofC
MN
Fun
gal
inoc
ulan
ts
Isot
ope
used
tote
stfo
rtr
ansf
er
Eff
ect
ofC
MN
trea
tmen
ton
fung
alco
mm
unit
y
Eff
ect
ofC
MN
onin
divi
dual
plan
tpe
rfor
man
ceP
lant
com
mun
ity
effe
cts
stud
ied
orsu
gges
ted
Ref
eren
ce
EC
M,
fiel
dP.
men
zies
iise
ed-
ling
spl
ante
din
shru
bpa
tche
sdo
min
ated
byar
buto
idA
rcto
stap
hylo
sgl
andu
losa
and
AM
Ade
nost
oma
fasc
icul
atum
P.m
enzi
esii
seed
-li
ngs
grow
n4–
10m
onth
sfr
omse
edin
patc
hes
ofA
deno
stom
aan
dA
rcto
stap
hylo
s;li
ght,
soil
,nu
tri-
ents
,an
dal
lelo
path
yw
ere
not
impo
rtan
tto
seed
ling
resp
onse
Fie
ldso
il.
Aft
er10
mon
ths,
17E
CM
spec
ies
colo
-ni
zed
both
P.m
enzi
esii
and
A.
glan
dulo
sain
patc
hes
ofA
rcto
stap
hylo
s;in
sam
eco
re,
85%
ofP.
men
zies
iian
d49
%of
thei
rE
CM
biom
ass
colo
nize
dby
fung
iof
Arc
tost
aphy
los
Non
eA
fter
4m
onth
s,P.
men
zies
iise
ed-
ling
sin
four
offi
veA
rcto
stap
hylo
spa
tche
sw
ere
colo
-ni
zed
bysi
xE
Msp
ecie
sw
here
asse
edli
ngs
intw
oof
five
Ade
nost
oma
patc
hes
wer
eco
lo-
nize
dby
two
EC
Msp
ecie
sof
The
leph
ora
P.m
enzi
esii
seed
ling
mor
tali
tyhi
gher
inA
deno
stom
ath
anA
rcto
stap
hylo
spa
tche
s;no
seed
-li
ngs
surv
ived
inpa
tche
sof
Ade
nost
oma,
whe
reas
20%
sur-
vive
din
the
Arc
tost
aphy
los,
mai
nly
due
toE
CM
inoc
ulum
diff
eren
ces
Sur
viva
lof
esta
blis
hing
seed
ling
sw
asgr
eatl
yin
crea
sed
whe
nas
soci
-at
edw
ith
the
exis
ting
CM
Nin
Arc
tost
aphy
los
patc
hes.
Aut
hors
sugg
est
that
EC
Msp
ecie
ssh
ared
byP.
men
zies
iian
dA
rcto
stap
hylo
sco
n-ne
cted
the
two
host
plan
ts,
crea
ting
cond
i-ti
ons
cond
uciv
efo
rC
tran
sfer
tow
ard
P.m
enzi
esii
Hor
ton
etal
.19
99
EC
M,
fiel
dM
icro
berl
inia
bisu
lcat
a,Te
trab
erli
nia
bifo
liat
a,an
dTe
trab
erli
nia
mor
elia
nase
ed-
ling
spl
ante
din
unde
rsto
ry
Con
spec
ific
seed
-li
ngs
plan
ted
into
fore
sts
wit
hhi
ghan
dlo
wba
sal
area
ofE
CM
adul
ttr
ees
Fie
ldso
ilN
one
Hig
her
myc
orrh
izat
ion
inhi
gh-E
CM
than
inlo
w-E
CM
fore
st;
noef
fect
ofm
ycor
rhiz
atio
non
seed
ling
biom
ass,
but
thos
ew
ith
high
EC
Mha
dhe
avie
rta
proo
ts
Onl
yon
esp
ecie
sin
crea
sed
insu
r-vi
val
orbi
omas
sin
the
high
-EC
Mfo
rest
;su
rviv
ors
oftw
otr
eesp
ecie
sw
ere
heav
ier
inhi
gh-E
CM
fore
sts
Hig
hba
sal
area
ofE
CM
adul
ts,
less
effe
ctiv
ene
twor
kco
nnec
tion
s,or
NM
tran
spla
nts
may
have
mas
ked
the
impo
rtan
ceof
the
CM
Nto
esta
blis
hing
rege
nera
tion
prov
ided
byco
nspe
cifi
cad
ults
New
berr
yet
al.
2000
EC
M,
fiel
dPa
rabe
rlin
iabi
foli
ata
seed
-li
ngs
plan
ted
unde
rA
fzel
iabi
pind
ensi
s,B
rach
yste
gia
cyno
met
roid
es,
Para
berl
inia
bifo
liat
a,an
dT.
bifo
liat
aad
ults
See
dlin
gspl
ante
d5–
30m
away
from
adul
tsei
ther
inco
ntac
tw
ith
adul
tro
ots
oris
olat
edin
side
aP
VC
tube
Fie
ldso
ilN
one
Num
ber
ofse
edli
ngs
wit
hm
ycor
rhiz
aean
dde
gree
ofin
fect
ion
high
erw
here
seed
ling
sin
cont
act
wit
had
ult
root
sth
anis
olat
ed;
myc
orrh
izat
ion
decl
ined
wit
hdi
s-ta
nce
from
adul
ts
See
dlin
gsu
rviv
alan
dbi
omas
shi
gher
whe
reco
ntac
tw
ith
adul
tro
ots
than
whe
reis
olat
ed;
per-
cent
age
ofE
CM
seed
ling
sde
clin
edw
ith
dist
ance
,bu
tno
tsu
rviv
al,
soE
CM
and
com
peti
-ti
onim
port
ant
CM
Nw
asim
port
ant
inth
ere
gene
rati
onof
EC
Mtr
ees
inan
AM
-do
min
ated
trop
ical
rain
fore
stin
Cam
er-
oon;
diff
eren
tad
ult
tree
spec
ies
had
diff
er-
ent
inoc
ulum
pote
ntia
lsfo
rpl
ante
dse
edli
ngs;
rege
nera
tion
shou
ldbe
carr
ied
out
unde
rco
nspe
cifi
cpa
rent
tree
s
Ong
uene
and
Kuy
per
2002
Tab
le3.
(con
tinu
ed).

© 2004 NRC Canada
1156 Can. J. Bot. Vol. 82, 2004
Myc
orrh
izal
type
and
stud
yco
ndit
ions
Pla
ntsp
ecie
san
dpl
anti
ngde
sign
Met
hod
used
tote
stef
fect
ofC
MN
Fun
gal
inoc
ulan
ts
Isot
ope
used
tote
stfo
rtr
ansf
er
Eff
ect
ofC
MN
trea
tmen
ton
fung
alco
mm
unit
y
Eff
ect
ofC
MN
onin
divi
dual
plan
tpe
rfor
man
ceP
lant
com
mun
ity
effe
cts
stud
ied
orsu
gges
ted
Ref
eren
ce
EC
M,
fiel
dQ
uerc
usru
bra
acor
nspl
ante
dad
jace
ntto
live
EM
Que
rcus
mon
tana
(nea
r-Q
uerc
us),
dead
Q.
mon
tana
(nea
r-de
ad-
Que
rcus
),an
dA
MA
cer
rubr
um(n
ear-
Ace
r)st
ump
spro
uts
EC
Mvs
.A
Mst
ump
spro
uts
test
edE
CM
infl
uenc
ew
itho
utso
ildi
s-tu
rban
ce;
Que
rcus
mon
tana
redu
ced
root
graf
ting
wit
hQ
.ru
brum
;liv
evs
.de
adte
sted
site
alte
rati
onvs
.in
dica
tion
Fie
ldso
ilN
one
Nea
r-Q
uerc
usse
ed-
ling
sin
fect
edby
EC
Mm
ore
rapi
dly
and
toa
grea
ter
exte
nt(4
×)
wit
ha
diff
eren
tan
dm
ore
dive
rse
EM
com
-m
unit
yth
anne
ar-
Ace
ror
near
-dea
d-Q
uerc
usse
edli
ng.
Nea
r-Q
uerc
usse
ed-
ling
sha
dgr
eate
rN
and
Pco
nten
tsth
anin
the
othe
rtr
eat-
men
tsan
dgr
eate
rgr
owth
inth
anth
ene
ar-A
cer
trea
tmen
t
Aut
hors
sugg
est
grea
ter
infe
ctio
nof
seed
ling
sN
ear-
Que
rcus
occu
rred
via
hyph
alli
nks
wit
hliv
ero
ots
ofst
ump
spro
uts;
data
supp
ort
mic
rosi
teal
tera
tion
hypo
thes
isth
atliv
ero
ots
ofQ
.m
onta
nain
crea
sed
qual
ity
and
quan
tity
ofE
CM
,le
adin
gto
incr
ease
dse
edli
nggr
owth
and
nutr
itio
n;in
crea
sing
dens
ity
ofQ
.m
onta
naco
uld
mas
kbe
nefi
ts
Dic
kie
etal
.20
02
AM
,m
icro
-co
sms
inla
b
20he
rbsp
ecie
sin
trod
uced
toea
chm
icro
cosm
whe
regr
azin
g,m
ycor
rhiz
atio
n,an
dhe
tero
geno
ussp
atia
ltr
eat-
men
tsap
plie
d
NM
cont
rols
Glo
mus
cons
tric
tum
used
toin
ocul
ate
seed
ling
sin
myc
orrh
izal
trea
tmen
t
14C
Pla
ntse
edli
ngs
wer
ein
fect
edw
ith
AM
Pla
ntdi
vers
ity
incr
ease
d,w
hile
yiel
dof
dom
inan
tFe
stuc
aov
ina
decr
ease
dan
dA
Msu
bord
inat
esp
ecie
sin
crea
sed;
14C
tran
sfer
red
toA
Mbu
tno
tN
Msu
bord
inat
es
Aut
hors
sugg
est
that
Cm
ovem
ent
from
sour
ceto
sink
plan
tsvi
aa
CM
Nm
aybe
impo
r-ta
ntin
mai
ntai
ning
spec
ies-
rich
com
mun
i-ti
esin
infe
rtil
eso
ils
Gri
me
etal
.19
87
AM
,fi
eld
Tall
gras
spr
airi
edo
min
ated
byA
ndro
pogo
nge
rard
ii,
And
ropo
gon
scop
ariu
s,So
rgha
stru
mnu
tans
,Pa
nicu
mvi
rgat
umpl
usm
ixof
war
m-,
cool
-sea
son
spec
ies
MC
was
com
pare
dw
ith
NM
trea
t-m
ent;
inN
M,
AM
fung
iw
ere
sup-
pres
sed
wit
hre
peat
edap
plic
a-ti
ons
ofbe
nom
yl
InM
C,
fiel
dso
ilin
ocul
ated
plan
tsw
ith
nativ
eG
lom
ussp
ecie
s;in
NM
trea
tmen
t,be
nom
yltr
eat-
men
tsw
ere
appl
ied
ever
y2
wee
ksth
roug
h-ou
tth
egr
owin
gse
ason
for
4ye
ars
Non
eB
enom
yltr
eatm
ent
redu
ced
AM
colo
ni-
zati
onof
indi
vidu
alpl
ant
spec
ies
to<
5%(a
ppro
x.25
%of
MC
)
Sup
pres
sion
ofm
ycor
rhiz
aere
duce
dab
unda
nce
ofdo
min
ant
obli
-ga
teC
4ta
llgr
asse
s,an
din
crea
sed
sub-
ordi
nate
facu
ltat
ive
C3
plan
ts;
plan
tbi
omas
sun
chan
ged,
dive
rsit
yin
crea
sed
inN
M
Two
poss
ible
mec
ha-
nism
sof
myc
orrh
izal
-m
edia
ted
chan
ges
inpl
ant
dive
rsit
yan
dco
mpo
siti
on:
(i)
alte
r-at
ions
inre
sour
cedi
stri
buti
onam
ong
neig
hbor
svi
aC
MN
or,
(ii)
diff
eren
tho
stre
spon
ses
toA
Mco
lo-
niza
tion
inw
hich
plan
tdo
min
ants
diff
erin
myc
orrh
izal
depe
nd-
ency
than
neig
hbor
s
Har
tnet
tan
dW
ilso
n19
99
Tab
le3.
(con
tinu
ed).

© 2004 NRC Canada
Simard and Durall 1157
Myc
orrh
izal
type
and
stud
yco
ndit
ions
Pla
ntsp
ecie
san
dpl
anti
ngde
sign
Met
hod
used
tote
stef
fect
ofC
MN
Fun
gal
inoc
ulan
ts
Isot
ope
used
tote
stfo
rtr
ansf
er
Eff
ect
ofC
MN
trea
tmen
ton
fung
alco
mm
unit
y
Eff
ect
ofC
MN
onin
divi
dual
plan
tpe
rfor
man
ceP
lant
com
mun
ity
effe
cts
stud
ied
orsu
gges
ted
Ref
eren
ce
AM
,po
tsin
lab
Cen
taur
eam
acul
osa,
Fest
uca
idah
oens
is
C.
mac
ulos
aan
dF.
idah
oens
isgr
own
alon
ean
din
mix
ture
,w
ith
and
wit
hout
AM
fung
i;te
sted
effe
cts
ofsm
all,
larg
eFe
stuc
a
AM
trea
tmen
tsgr
own
wit
hna
tive
fiel
dso
il.
NM
trea
t-m
ents
had
auto
clav
edfi
eld
soil
wit
hm
icro
bial
was
h;fo
rla
rge
Fest
uca,
beno
myl
was
used
tocr
eate
NM
cont
rols
Non
eG
ood
AM
colo
niza
-ti
onin
myc
orrh
izal
trea
tmen
ts,
and
0%an
dre
duce
din
cula
tion
inN
Mco
ntro
lsfo
rsm
all
and
larg
eFe
stuc
a,re
spec
tivel
y
AM
incr
ease
dth
eco
mpe
titiv
eef
fect
ofC
.m
acul
osa
onsm
all
orla
rge
F.id
ahoe
nsis
AM
cont
ribu
ted
toco
m-
peti
tive
dom
inan
ceby
C.
mac
ulos
a.A
utho
rssu
gges
tth
atA
Mm
ayha
vem
edia
ted
apa
ra-
siti
cC
ornu
trie
ntdr
ain
byC
.m
acul
osa
onF.
idah
oens
isvi
aa
CM
Nor
that
com
peti
-ti
onfo
rre
sour
ces
was
impr
oved
Mar
ler
etal
.19
99
AM
,po
tsin
lab
C.
mac
ulos
a,pa
ired
wit
hA
chil
lea
mil
lefo
lium
,G
aill
ardi
aar
ista
t,F.
idah
oens
is,
Pse
udor
oegn
eria
spic
ata,
orK
oele
ria
cris
tata
28- µ
mm
esh
barr
ier
and
0.45
- µm
mem
bran
eba
rrie
r
Fie
ldso
il13
CC
olon
izat
ion
good
for
all
spec
ies
C.
mac
ulos
ala
rger
wit
hgr
eate
rti
ssue
[P]
whe
rem
esh
barr
ier
occu
rred
,pa
rtic
ular
lyw
hen
pair
edw
ith
gras
ses,
beca
use
the
CM
Nin
crea
sed
its
acce
ssto
Pon
both
side
ofth
epo
t;no
Ctr
ansf
er
AM
enha
nced
C.
mac
ulos
aby
incr
easi
ngP
upta
ke;
CM
Ndi
dno
tfa
cili
-ta
ted
Ctr
ansf
er,
but
may
have
incr
ease
dnu
trie
ntac
cess
toin
terc
onne
cted
plan
ts
Zab
insk
iet
al.
2002
AM
,po
tsin
lab
Cen
taur
eam
elit
ensi
s,N
asse
lla
pulc
hra,
Ave
naba
rbat
a
C.
mel
iten
sis,
N.
pulc
hra,
and
A.
barb
ata
grow
nal
one
and
inm
ixtu
re,
wit
han
dw
itho
utA
Mfu
ngi
AM
trea
tmen
tsgr
own
wit
hna
tive
fiel
dso
ilas
inoc
ulum
,an
dN
Mtr
eatm
ents
crea
ted
wit
hbe
nom
yl
Non
eB
enom
ylsi
gnif
ican
tly
redu
ced
AM
fung
iac
ross
all
thre
epl
ant
spec
ies.
Bio
mas
sof
lone
C.
mel
iten
sis
50%
the
size
wit
hA
Mth
anN
M,
but
larg
erw
ith
AM
whe
npa
ired
wit
hN
.pu
lchr
a;bi
omas
sun
affe
cted
bybe
nom
ylw
hen
wit
hA
vena
barb
ata
Aut
hors
sugg
est
Cm
ayha
vetr
ansf
erre
dfr
omN
.pu
lchr
ato
C.
mel
iten
sis
via
CM
N,
but
not
from
Ave
naba
rbat
a;al
ter-
nativ
ely,
N.
pulc
hra
inhi
bite
dby
soil
path
ogen
sin
mix
ture
,or
posi
tivel
yaf
fect
sso
ilfu
ngi
bene
fici
alto
C.
mel
iten
sis
Cal
law
ayet
al.
2003
AM
,po
tsin
lab
Sibb
aldi
apr
ocum
bens
adul
t,se
edli
ngpa
ired
wit
har
c-ti
che
rbs
NM
cont
rol
Glo
mus
hoi,
Glo
mus
sp.,
Glo
mus
clar
oide
um
Non
eC
ontr
ols
wer
eN
M;
G.
hoi
and
G.
clar
oide
umhi
ghly
infe
ctiv
e;ad
ults
infe
cted
seed
ling
svi
aC
MN
See
dlin
ggr
owth
im-
prov
edw
ith
AM
,bu
tno
tw
hen
inC
MN
wit
hes
tab-
lish
edSi
bbal
dia
be-
caus
eof
com
peti
tive
effe
cts
CM
Nm
ayve
ctor
com
pe-
titi
onbe
twee
nes
tab-
lish
edpl
ants
and
seed
ling
s,an
dlo
nese
edli
ngs
bene
fit
mor
efr
omA
Mth
anse
ed-
ling
sne
arad
ult
plan
ts;
pote
ntia
lpo
sitiv
eef
-fe
cts
ofC
MN
mas
ked
byco
mpe
titi
on
Kyt
õvii
taet
al.
2003
Not
e:M
C,
myc
orrh
izal
cont
rol;
NM
,no
nmyc
orrh
izal
.
Tab
le3.
(con
clud
ed).

fected more rapidly with a greater diversity of mycorrhizalfungi; they may tap into an established CMN supported by Cinputs from other plants; they may access a larger pool ofsoil nutrients and water through the CMN; they may receiveC, nutrients, or water transferred directly from other plants;or they may have greater access to available nutrientsthrough CMN interactions with other soil organisms. Thesebenefits may be of life-or-death significance to a plant ifthey come, no matter how briefly, at a particular critical pe-riod during the plant-establishment phase. Once tapped intothe CMN, seedlings may be more likely to survive the criti-cal regeneration period in stressed or harsh environments,such as in deep shade (Simard et al. 1997a; Onguene andKuyper 2002), drought (Borchers and Perry 1990; Marler etal. 1999; Horton et al. 1999), at high elevations (Perry etal. 1989; Jumpponen and Trappe 1998), or on severely orfrequently disturbed sites. The CMN may be particularlysignificant for natural regeneration of temperate and borealforests that are frequently disturbed by wildfire, where itprovides critical linkages through time between the old andnew forests (Perry et al. 1992; Jonsson et al. 1999a).
Potential mycorrhizal network influences on plant estab-lishment have been studied fairly extensively in the field.Studies have used several techniques to separate CMN fromnon-CMN effects, including soil transfers (Perry et al. 1989;Perry et al. 1992), soil trenching (Simard et al. 1997c;Onguene and Kuyper 2002), mycorrhizal versus nonmy-corrhizal or incompatible control plants (Simard et al.1997a; Lerat et al. 2002; Kytoviita et al. 2003), plantingseedlings in mycorrhizal-compatible versus noncompatibleplant communities (Horton et al. 1999), or by seeding orplanting into an existing plant community followed byexamination of mycorrhizae and regeneration patterns(Jumpponen and Trappe 1998; Jonsson et al. 1999b;Newberry et al. 2000; Bidartondo et al. 2000; Dickie et al.2002). None of these studies investigated C or nutrient trans-fers, but rather have quantified general responses of seed-lings and mycorrhizae following association or linkage intothe CMN of established plants. In most cases, seedling bene-fits appear to have resulted from increased mycorrhizal di-versity by association with mature plants. Several of thesestudies are described next.
A number of field studies that support the hypothesis thata CMN can enhance plant establishment and performancehave been conducted in the temperate coniferous forests ofwestern North America. In a series of elegant experimentsin high-elevation Oregon mountains, Perry and associatesfound that regenerating P. menziesii seedlings benefitedgreatly from growing in soils transferred from beneath hard-woods compared with unamended, disturbed soils (Perry etal. 1992). Their work suggested that direct transfer ofmycorrhizal inocula from hardwoods to Douglas-fir was oneof several important factors that helped facilitate establish-ment of Douglas-fir. It corroborated earlier evidence fromDeacon et al. (1983) that seedlings planted close to birchparents formed shared, “late-stage” fungi while those at dis-tance formed different “early-stage” fungi. Since then,Simard et al. (1997c) found that P. menziesii seedlingsplanted in the understory of mature P. menziesii andB. papyrifera forests in British Columbia had higher netphotosynthetic rates, higher ECM diversity and richness, and
a different community of ECM morphotypes (more late-stage and strand-forming ECM) in untrenched (in contactwith root systems of mature trees) than in trenched plots(isolated from mature trees). Access to the higher mycelialinoculum density near the mature trees in the untrenchedtreatment likely accounted for the more diverse and robustECM community on understory P. menziesii seedlings, pos-sibly imposing a greater C drain on the seedlings and there-fore stimulating higher net photosynthetic rates than in thetrenched treatment. In another study that sought to explainpreferential establishment of P. menziesii in Arctostaphylosshrub patches within the California chaparral vegetationtype, Horton et al. (1999) found that P. menziesii seedlingswere colonized more frequently by a greater richness ofECM fungi in Arctostaphylos patches than in Adenostomapatches, corresponding with greater survival rates in theArctostaphylos patches. The authors suggested ECM ino-culum differences between shrub species largely accountedfor the P. menziesii survival differences and that ECM spe-cies shared by P. menziesii and Arctostaphylos connectedthe two host plants, creating conditions conducive for Ctransfer toward P. menziesii. Both the studies of Simard etal. (1997c) and Horton et al. (1999) were able to discountmost environmental factors (e.g., light, available nutrients,or soil water) that could provide an alternative explanationfor seedling performance differences in their treatments.
Similar patterns of tree-seedling establishment have alsobeen found in temperate and tropical forests dominated byhardwood tree species (Newberry et al. 2000; Dickie et al.2002; Onguene and Kuyper 2002). These studies make thefollowing two important contributions: (i) mycorrhizal facili-tation is likely important in secondary succession wheresymbiont populations are potentially limiting, contrastingwith more traditional concepts of facilitation involving pri-mary succession or harsh environmental conditions, and(ii) benefits of mycorrhizal facilitation may be masked bycompetition for resources when tree densities increasebeyond some threshold level. The benefits of mycorrhizalinfection probably happens at relatively low tree densities(Dickie et al. 2002), a suggestion that is corroborated byother studies (Kranabetter 1999; Hagerman et al. 2001).Dickie et al. (2002) found that establishment of ECMQuercus rubrum L. seedlings in a burned and salvage-loggedforest was facilitated by planting the acorns near sproutingstumps of Q. montana Willd. (congeneric was used to elimi-nate possibility of root grafting) but was relatively sup-pressed when planted near sprouting stumps of AM Acerrubrum L. Seedlings planted near Q. montana had a morediverse and different community of ECM fungi, greater Nand P contents, and greater growth relative to seedlingsplanted near A. rubrum, indicating indirect facilitationthrough increased seedling ECM formation. The authorssuggest that with increasing overstory density, mycorrhizalinfection benefits would be overwhelmed by competition forresources, resulting in decreasing Q. rubrum growth. In asimilar study, Onguene and Kuyper (2002) found that 1-month-old seedlings planted into a tropical rainforest hadgreater survival, biomass, and ectomycorrhizal formationwhen they were in contact with conspecific adult roots thanwhen they were isolated. They also found that mycor-rhization declined with distance from parents, while survival
© 2004 NRC Canada
1158 Can. J. Bot. Vol. 82, 2004

did not, suggesting that seedling survival was probably alsopositively influenced by declining competition with distancefrom parents. Corroborating results have been found inclearcut, mixed hardwood – conifer sites in British Colum-bia. Pseudotsuga menziesii planted in a low-density mixturewith B. papyrifera had faster leader growth and higher netphotosynthetic rates in untrenched than in trenched plotsduring the first 3 years after establishment, but this was re-versed in subsequent years after B. papyrifera had outgrownP. menziesii (S.W. Simard, unpublished data). The crossoverfrom positive to negative effects of B. papyrifera neighborson P. menziesii performance happened more quickly in high-than in low-density mixtures.
In some ecosystems, the competitive effects of neighborsmay completely mask facilitative effects of the CMN, evenat low densities and very early during plant establishment. Ina pot study using subarctic herbs, Kytoviita et al. (2003)found that biomass growth of solitary seedlings improvedwith AM inoculation, but these benefits were completelylost when the AM seedlings were linked to adult Sibbaldiaprocumbens L. plants via the CMN. In the absence ofmycorrhizal fungi, by contrast, nonmycorrhizal seedlingsgrew equally well alone or in the presence of a non-mycorrhizal Sibbaldia adult. The authors suggested that theCMN vectored competition between established plants andseedlings, masking any benefits that may have been con-ferred by the CMN. These studies show that the effectsof mycorrhizal symbioses and the CMN on seedling-establishment patterns can vary considerably, depending onthe plant species, fungal species, growing environment, andlife stage of the different plants within a community.
In summary, studies examining CMN effects on plant es-tablishment have found that association with a mature plantresults in seedlings infected with a greater richness or diver-sity of mycorrhizal fungi, a greater proportion of fungishared in common with the mature plant, or a greater pro-portion of strand-, cord-, or rhizomorph-forming fungi. Inmany cases, this has been associated with greater plant per-formance, as measured by increased survival, growth, photo-synthetic rate, or tissue nutrient contents. In none of thesestudies, however, is there convincing evidence that these ef-fects have resulted from C or nutrient transfer from matureplants, even though it is commonly suggested as one of sev-eral potential facilitative mechanisms. Conversely, there ismore evidence that the benefits arise simply from increasedmycorrhization, which may or may not require CMN forma-tion. The beneficial effects of mature plants have beenshown to decrease with increasing neighbour density, prox-imity, or size, suggesting there may be threshold levels atwhich competition for resources outweighs the beneficial ef-fects of CMN inoculation. Future studies in this area need toseparate the effects of C or nutrient transfer from simplemycorrhization and to identify plant community factors thatregulate seedling facilitation by the CMN.
CMN effects on plant competition and diversitySeveral studies have examined the potential effects of a
CMN on inter- and intra-specific competition among plants,even though all suffered from inadequacies in clearly dem-onstrating the existence of a functional CMN. These studiessuggest, however, that the CMN has the potential to decrease
(Grime et al. 1987; Perry et al. 1989; Gange et al. 1993;Moora and Zobel 1996) or increase interplant competition(Eissenstat and Newman 1990; Eason et al. 1991; Hartnett etal. 1993; Bethlenfalvay et al. 1996a; Rejon et al. 1997).Grime et al. (1987) suggested that competitive intensity maybe reduced by the direct flow of C or nutrients from suffi-cient to deficient plants, resulting in greater growth of subor-dinate relative to dominant plants, and greater plantcommunity diversity. On the other hand, there is some evi-dence that competitive dominance may increase, becauselarger plants acquire greater resources from the CMN be-cause of their higher nutrient demand (Zabinski et al. 2002).In some cases, competitive intensity within plant communi-ties may simply be affected by greater, faster, more diverse,or different mycorrhization of plants tapping into the CMN,resulting in increased nutrient access for some plant species.
In one of the first studies that found evidence for CMN ef-fects on interspecific competition, Perry et al. (1989) exam-ined interactions between P. menziesii and Pinus ponderosaDougl. ex P. Laws. & C. Laws. in a replacement-series de-sign with none, two, or four common ECM species added.They found that the two tree species mutually inhibited eachother with no ECM added (but with a Thelphora contami-nant), but that this antagonism disappeared with added ECMspecies, particularly with added generalist Laccaria laccata(Scop.: Fr.) Cooke, resulting in larger P. menziesii seedlingswith more foliar N and P content. The benefits to P. men-ziesii came at no expense to growth or nutrient content ofPinus ponderosa. The authors suggested that competitionwas reduced because a CMN increased access to nutrients, aCMN distributed nutrients more evenly, or more simply thatthe greater diversity of fungi was active over a broader rangeof environmental conditions. Perry et al. (1989) were veryeffective in separating inter- and intra-specific competitiveeffects as well as eliminating the confounding effects ofcompetitor density through the use of a replacement-seriesdesign (Harper 1977). In their experiment, however, contam-ination of the “no-ECM” control by Thelephora may havemasked some of the CMN effects. In addition, the designmade it difficult to separate CMN effects from fungal diver-sity effects on competitive interactions. Some of the prob-lems with this experimental design may be avoided in futureexperiments by use of growth chambers to reduce contami-nation, mesh barrier controls that prevent formation ofhyphal connections, and isotopic tracers to confirm the pres-ence of links and interplant nutrient transfer.
A similar replacement-series approach was also used inthe field by Jones et al. (1997) to examine the effects of treespecies composition and density on the ECM communitiesof P. menziesii and B. papyrifera. Evenness of the ECMcommunity on P. menziesii was higher when grown in mix-ture with B. papyrifera than when grown alone because ofincreases in minor types, and this may have resulted from ei-ther coinoculation from neighboring birch hosts (Massicotteet al. 1994; Simard et al. 1997b) or modification of soilsby birch favoring different mycorrhizal fungi. Jones et al.(1997) found that 91% of B. papyrifera and 56% of P. men-ziesii mycorrhizae were types common to the two tree spe-cies, and they hypothesized that the resulting CMN mayreduce the negative effects of competition between the twotree species. This hypothesis was supported by Simard et al.
© 2004 NRC Canada
Simard and Durall 1159

(1997a), who showed on the same sites that net C transferfrom B. papyrifera to P. menziesii increased with increasingshading of P. menziesii.
In studies seeking to explain why AM fungi increased thecompetitive dominance of the exotic invasive Centaureamaculosa auct. non Lam over native Festuca idahoensisElmer in Montana grasslands, Marler et al. (1999) andZabinski et al. (2002) used 13C to test whether the CMN hadmediated a parasitic drain by Centaurea maculosa fromF. idahoensis or whether it simply enhanced the competitiveability of Centaurea maculosa for soil resources. Zabinski etal. (2002) found that the CMN did not facilitate C transferfrom F. idahoensis to Centaurea maculosa, but rather thatit increased P uptake by Centaurea maculosa, particularlywhen paired with grasses rather than forbs. Forming a CMNwith grass increased access of AM Centaurea maculosa to Pover a wider soil pool covering the neighbor’s rooting zone,resulting in greater plant biomass and luxury consumption ofP. While the CMN in this study did not appear to serve as aconduit of resource transfer, it may have benefited plants inthe community by increasing hyphal development with mul-tiple hosts over a larger soil pool of resources. Either mecha-nism can result in changes in the plant community.
Changes in competitive interactions among plants result-ing from a CMN have been suggested to affect plant com-munity diversity. Reduced interspecific competition, forexample, can reduce the abundance of canopy dominants,increase abundance of subordinates, and result in a moreevenly distributed and diverse plant community. Grime et al.(1987) observed apparent one-way 14C transfer from canopy-dominant F. ovina to understory herbs and grasses, whichthey suggested occurred through a CMN along a naturalsource–sink gradient for assimilate and may have accountedfor increased diversity and more even biomass distributionamong plants. This work was later criticized, because nettransfer and the transfer pathway were not examined (Fitteret al. 1999), and interplant variation in C physiology andAM dependency was not adequately demonstrated. van derHeijden (2002) subsequently reanalysed the data of Grime etal. (1987) and found that subordinate plant species with thehighest mycorrhizal dependency obtained the most 14C fromF. ovina, thus benefiting the most from the CMN. He sug-gested that the number and relative abundance ofmycorrhizal-dependent plant species may help explain thedifferences in how AM fungi, the CMN, and interplant Ctransfer affect different plant communities. It may explain,for example, why Hartnett and Wilson (1999) found thatplant diversity increased when AM mycorrhization declinedfollowing benomyl treatments in a tallgrass prairie, the op-posite result of Grime et al. (1987). Suppression of AMmycorrhizae decreased abundance of the dominant, obli-gately mycotrophic C4 tallgrasses and increased abundanceof subordinate facultatively mycotrophic C3 plants. However,Walter et al. (1996) earlier showed that more 32P was trans-ferred to facultatively mycorrhizal plants than to obligatelymycorrhizal plants in a similar prairie, leading Hartnett andWilson (1999) to conclude that changes in plant diversity intheir experiment resulted more from differences in host re-sponses to AM than to alteration in resource distributionamong neighbors via the CMN.
Evidence for CMN or C transfer effects on competitive
interactions and plant community diversity is inadequate.Several studies above have used isotopes to trace transferthrough a CMN, but most suffer from lack of adequate non-CMN controls (e.g., Grime et al. 1987; Perry et al. 1989;Walter et al. 1996), and few have been conducted underfield conditions. Most studies suggest that non-CMN mecha-nisms, such as changes in soil chemistry or mycorrhizalinfection patterns, are just as likely to explain plant commu-nity competition or diversity changes as CMN or transfermechanisms. Although laboratory and greenhouse studiesprovide greater opportunity for precisely examining theCMN, they provide an unrealistic picture of how the CMNaffects plant communities in nature (Read 2002). More re-search is needed to improve our understanding of whether,where, and how the CMN affects competition and commu-nity diversity in nature. We also need to research those fac-tors that may alter CMN effects on plant communities,including different attributes of site quality (nutrient, water,and light limitations), plant species composition (source–sink patterns, mycorrhizal dependency), fungal species com-position, and stage of plant community development.
Conclusions
Recent field studies have increased our knowledge of theextent, function, and implications of CMNs, but only labstudies have unequivocally demonstrated the existence ofCMNs and their specific role in transferring C from one rootsystem to another. Field evidence for CMNs is supported byboth functional- and taxonomic-level data. The general lackof specificity, at least at the species level, of both AM andECM fungi to their host suggests that CMNs within grass-lands and forests are abundant and extensive. Whether speci-ficity to different host species occurs at a level lower thanspecies is still not well understood. Further research usingmolecular and microsatellite techniques will help to deter-mine whether this specificity occurs and whether it affectsCMN potential. C transfer between plants has been observedin both ECM and AM field studies (Simard et al. 1997a;Lerat et al. 2002), but the transfer mechanism has yet to befully investigated.
Newman (1988) emphasized the need to know whether(i) interplant C transfer in the field is one-way or bidirec-tional, (ii) transfer occurs through a CMN, (iii) transferred Centers the receiver shoot, and (iv) receiver plants benefit sig-nificantly from the CMN. Since this pivotal review, there isfield evidence of bidirectional and net transfer (Simard et al.1997a) and transfer to the receiver shoot in both ECM andAM plants (Simard et al. 1997a; Lerat et al. 2002). Thesestudies did not conclusively show whether the CMN acted asa conduit for C transfer, but they did clearly demonstratethat mycorrhizae facilitated transfer. These studies also pro-vided evidence that the direction and magnitude of C trans-fer was highly dependent on source–sink relations betweenlinked plants. Further field research is needed on plant andfungal source–sink relationships, how they affect magnitudeand direction of transfer, and the mechanisms by which theyinfluence transfer in nature.
Several studies have suggested that the CMN may play arole in improving seedling establishment, reducing or in-creasing plant competition, and reducing or increasing plant
© 2004 NRC Canada
1160 Can. J. Bot. Vol. 82, 2004

community diversity. The strongest evidence for CMN ef-fects on plant communities exists for seedling establishment.There is a need to examine CMN effects on plant competi-tion and diversity in nature with adequate controls for theCMN, adequate designs for addressing the confoundingeffects of density, composition, and diversity of plant andfungal communities, direct observation of the functionalCMN, and concurrent isotopic labeling to quantify bidi-rectional and net transfer. Once we have this information,we will be better able to understand the ecological signifi-cance of mycorrhizal networks.
Acknowledgements
We are grateful to the Organizing Committee for the invi-tation to present this review paper at the Fourth InternationalConference on Mycorrhizae in Montréal, Canada, in August2003. We thank Melanie D. Jones for providing valuablecomments on the manuscript.
References
Alpert, P., Warembourg, F.R., and Roy, J. 1991. Transport of car-bon among connected ramets of Eichhornia crassipes (Ponte-deriaceae) at normal and high levels of CO2. Am. J. Bot. 78:1459–1466.
Amaranthus, M.P., and Perry, D.A. 1994. The functioning ofectomycorrhizal fungi in the field: linkages in space and time.Plant Soil, 159: 133–140.
Arnebrant, K., Ek, H., Finlay, R.D., and Söderström, B. 1993. Ni-trogen translocation between Alnus glutinosa (L.) Gaertn. seed-lings inoculated with Frankia sp., and Pinus contorta Doug. exLoud seedlings connected by a common ectomycorrhizal myce-lium. New Phytol. 124: 231–242.
Amaranthus, M.P., Cázares, E., and Perry, D.A. 1999. The role ofsoil organisms in restoration. In Proceeding of the Pacific North-west Forest and Rangeland Soil Organism Symposium. Editedby R.T. Meurisse, W.G. Ypsilantis, and C. Seybold. USDA For.Serv. Pac. Northwest Res. Stn. Gen. Tech. Rep. PNW-GTR-461.pp. 179–189.
Bergemann, S.E., and Miller, S.L. 2002. Size, distribution, and per-sistence of genets in local populations of the late-stage ectomy-corrhizal basidiolycete, Russula brevipes. New Phytol. 156:313–320.
Bethlenfalvay, G.J., Mihara, K.L., Schreiner, R.P., and McDaniel,H. 1996a. Mycorrhizae, biocides, and biocontrol. 1. Herbicide-mycorrhiza interactions in soybean and cocklebur treated withbentazon. Appl. Soil Ecol. 3: 197–204.
Bethlenfalvay, G.J., Schreiner, R.P., Mihara, K.L., and McDaniel,H. 1996b. Mycorrhizae, biocides, and biocontrol. 2. Mycorrhizalfungi enhance weed control and crop growth in a soybean-cocklebur association treated with the herbicide bentazon. Appl.Soil Ecol. 3: 205–214.
Bergemann, S.E., and Miller, S.L. 2002. Size, distribution, and per-sistence of genets in local populations of the late-stage ectomy-corrhizal basidiomycete, Russula brevipes. New Phytol. 156:313–320.
Bever, J.D. 2003. Soil community feedback and the coexistence ofcompetitiors: conceptual frameworks and empirical tests. NewPhytol. 157: 465–473.
Bever, J.D., Westover, K.M., and Antonovics, J. 1997. Incorpo-rating the soil community into plant population dynamics: theutility of the feedback approach. J. Ecol. 85: 561–573.
Bever, J.D., Pringle, A., and Schultz, P.A. 2002. Dynamics withinth plant – arbuscular mycorrhizal fungal mutualism: testing thenature of community feedback. In Mycorrhizal ecology. Editedby M.G.A. van der Heijden and I. Sanders. Springer-Verlag,Berlin. pp. 267–292.
Bidartondo, M.I., and Bruns, T.D. 2002. Fine-level mycorrhizalspecificity in the Monotropoideae (Ericaceae): specificity forfungal species groups. Mol. Ecol. 11: 557–569.
Bidartondo, M.I., Kretzer, A.M., Pine, E.M., and Bruns, T.D. 2000.High root concentration and uneven ecto-mycorrhizal diversitynear Sarcodes sanguinea (Ericaceae). Am. J. Bot. 87: 1783–1788.
Bidartondo, M.I., Redecker, D., Hijri, I., Wiemken, A., Bruns,T.D., Dominguez, L., Sersic, A., Leake, J.R., and Read, D.J.2002. Epiparasitic plants specialized on arbuscular mycorrhizalfungi. Nature (Lond.), 419: 389–392.
Bjorkman, E. 1960. Monotropa hypopitys L. — an epiparasite ontree roots. Physiol. Plant. 13: 308–327.
Bonello, P., Bruns, T.D., and Gardes, M. 1998. Genetic structureof a natural population of the ectomycorrhizal fungus Suilluspungens. New Phytol. 138: 533–542.
Bonkowski, M., Jentschke, G., and Scheu, S. 2001. Contrastingeffects of microbial partners in the rhizosphere: interactionsbetween Norway Spruce seedlings (Picea abies Karst.), mycor-rhiza (Paxillus involutus (Batsch) Fr.) and naked amoebae (pro-tozoa). Appl. Soil Ecol. 18: 193–204.
Borchers, S.L., and Perry, D.A. 1990. Growth and ectomycorrhizaformation of Douglas-fir seedlings grown in soils collected atdifferent distances from pioneering hardwoods in southwest Or-egon clear-cuts. Can. J. For. Res. 20: 712–721.
Brownlee, C., Duddridge, J.A., Malibari, A., and Read, D.J. 1983.The structure and function of mycelial systems of ectomy-corrhizal roots with special reference to their role in forminginter-plant connections and providing pathways for assimilateand water transport. Plant Soil, 71: 433–443.
Bruns, T., Tan, J., Bidartondo, M., Szaro, T., and Redecker, D.2002a. Survival of Suillus pungens and Amanita franchetiectomycorrhizal genets was rare or absent after a stand-replacing wildfire. New Phytol. 155: 517–523.
Bruns, T.D., Bidartondo, M.I., and Taylor, D.L. 2002b. Host speci-ficity in ectomycorrhizal communities: What do the exceptionstell us? Integ. Comp. Biol. 42: 352–359.
Burton, P.J., Balisky, A.C., Coward, L.P., Cumming, S.G., andKneeshaw, D.D. 1992. The value of managing for biodiversity.For. Chron. 68: 225–237.
Callaway, R.M. 1995. Positive interactions among plants. Bot. Rev.61: 306–349.
Callaway, R.M., Marshall, B.E., Wicks, C., Pankey, J., and Zabin-ski, C. 2003. Soil fungi and the effects of an invasive forb ongrasses: neighbor identity matters. Ecology, 84: 129–135.
Cairney, J.W.G., and Smith, S.E. 1992. Influence of intracellularphosphorus concentration on phosphate absorption by theectomycorrhizal basidiomycete Pisolithus tinctorius. Mycol.Res. 96: 673–676.
Ceccaroli, P., Saltarelli, R., Cesari, P., Pierleoni, R., Sacconi, C.,Vallorani, L., Rubini, P., Stocchi, V., and Martin, F. 2003. Car-bohydrate and amino acid metabolism in Tuber borchii myce-lium during glucose utilization: a 13C NMR study. FungalGenet. Biol. 39: 168–175.
Chalot, M., and Brun, A. 1998. Physiology of organic nitrogen ac-quisition by ectomycorrhizal fungi and ectomycorrhizas. FEMSMicrobiol. Rev. 22: 21–44.
Cliquet, J.B., and Stewart, G.R. 1993. Ammonia assimilationin Zea Mays L. infected with a vesicular–arbuscular fungusGlomus fasciculatum. Plant Physiol. 101: 685–671.
© 2004 NRC Canada
Simard and Durall 1161

Cullings, K.W., Szaro, T.M., and Bruns, T.D. 1996. Evolution ofextreme specialization within a lineage of ectomycorrhizal epi-parasites. Nature (Lond.), 379: 63–66.
Deacon, J.W., Donaldson, S.J., and Last, F.T. 1983. Sequences andinteractions of mycorrhizal fungi on birch. Plant Soil, 71: 257–262.
DeLong, R.L., Lewis, K.J., Simard, S.W., Gibson, S. 2002. Fluo-rescent pseudomonad population sizes baited from soils underpure birch, pure Douglas-fir, and mixed forest stands and theirantagonism toward Armillaria ostoyae in vitro. Can. J. For. Res.32: 2146–2159.
Dickie, I.A., Koide, R.T., and Steiner, K.C. 2002. Influences of es-tablished trees on mycorrhizas, nutrition, and growth of Quercusrubra seedlings. Ecol. Monogr. 72: 505–521.
Dunham, S.M., Kretzer, A., and Pfrender, M.E. 2003. Character-ization of Pacific golden chanterelle (Cantharellus formosus)genet size using co-dominant microsatellite markers. Mol. Ecol.12: 1607–1618.
Durall, D.M., Jones, M.D., Kroeger, P., and Coates, K.D. 1999.Ectomycorrhizae and ectomycorrhizal sporocarps in cutblocksof different sizes in the interior cedar–hemlock forests of north-western British Columbia. Can. J. For. Res. 29: 1322–1332.
Eason, W.R., Newman, E.I., and Chuba, P.N. 1991. Specificity ofinterplant cycling of phosphorus: the role of mycorrhizas. PlantSoil, 137: 267–274.
Eissenstat, D.M., and Newman, E.I. 1990. Seedling establishmentnear large plants: effects of vesicular–arbuscular mycorrhizas onthe intensity of plant competition. Funct. Ecol. 4: 95–99.
Finlay, R.D. 1989. Functional aspects of phosphorus uptake andcarbon translocation in incompatible ectomycorrhizal associa-tions between Pinus sylvestris and Suillus grevillei and Bole-tinus cavipes. New Phytol. 112: 185–192.
Finlay, R.D., and Read, D.J. 1986a. The structure and function ofthe vegetative mycelium of ectomycorrhizal plants. I. Trans-location of 14C- labelled carbon between plants interconnectedby a common mycelium. New Phytol. 103: 143–156.
Finlay, R.D., and Read, D.J. 1986b. The structure and function ofthe vegetative mycelium of ectomycorrhizal plants. II. The up-take and distribution of phosphorus by mycelial strands inter-connecting host plants. New Phytol. 103: 157–165.
Finlay, R.D., Ek, H., and Odham, G. 1989. Uptake, translocation andassimilation of nitrogen from 15N-labelled ammonium and nitratesources by intact ecto-mycorrhizal systems of Fagus sylvatica in-fected with Paxillus involutus. New Phytol. 113: 47–55.
Fitter, A.H. 2001. Specificity, links and networks in the control ofdiversity in plant and microbial communities. In Ecology:achievement and challenge. 41st Symposium of the British Eco-logical Society. Edited by M.C. Press, N.J. Huntly, and S. Levin.Blackwell Scientific, Oxford, UK. pp. 95–114.
Fitter, A.H., Graves, J.D., Watkins, N.K., Robinson, D., andScrimgeour, C. 1998. Carbon transfer between plants and itscontrol in networks of arbuscular mycorrhizas. Funct. Ecol. 12:406–412.
Fitter, A.H., Hodge, A., Daniell, T.J., and Robinson, D. 1999. Re-source sharing in plant–fungus communities: Did the carbonmove for you? Trends Ecol. Evol. 14: 70–71.
Francis, R., and Read, D.J. 1984. Direct transfer of carbon betweenplants connected by vesicular–arbuscular mycorrhizal myce-lium. Nature (Lond.), 307: 53–56.
Frey, B., and Schüepp, H. 1992. Transfer of symbiotically fixed ni-trogen from berseem (Trifolium alexandrinum L.) to maize viavesicular–arbuscular mycorrhizal hyphae. New Phytol. 122:447–454.
Gange, A.C., Brown, V.K., and Sinclair, G.S. 1993. Vesicular–arbuscular mycorrhizal fungi: a determinant of plant communitystructure in early succession. Funct. Ecol. 7: 616–622.
Giovannetti, M., Fortuna, P., Citernesi, A.S., Morini, S., and Nuti,S.P. 2001. The occurence of anastomosis formation and nuclearexchange in intact arbuscular mycorrhizal networks. NewPhytol. 151: 717–724.
Graves, J.D., Watkins, N.K., Fitter, A.H., Robinson, D., andScrimgeour, C. 1997. Intraspecific transfer of carbon betweenplants linked by a common mycorrhizal network. Plant Soil,192: 153–159.
Grime, J.P., Mackey, J.M.L., Hillier, S.H., and Read, D.J. 1987.Floristic diversity in a model system using experimental micro-cosms. Nature (Lond.), 328: 420–422.
Guidot, A., Debaud, J.C., and Marmeisse, R. 2001. Correspon-dence between genetic diversity and spatial distribution ofabove- and below-ground populations of the ectomycorrhizalfungus Hebeloma cylindrosporum. Mol. Ecol. 10: 1121–1131.
Hagerman, S.M., Sakakibara, S.M., and Durall, D.M. 2001. Thepotential for woody understory plants to provide refuge forectomycorrhizal inoculum at an interior Douglas-fir forest afterclear-cut logging. Can. J. For. Res. 31: 711–721.
Hamel, C., Furlan, V., and Smith, D.L. 1992. Mycorrhizal effectson interspecific plant competition and nitrogen transfer inlegume–grass mixtures. Crop Sci. 138: 991–996.
Harms, K.E., Wright, S.J., Calderon, O., Hernandez, A., and Herre,E.A. 2000. Pervasive density-dependent recruitment enhances seed-ling diversity in a tropical forest. Nature (Lond.), 404: 493–495.
Harper, J.L. 1977. Population biology of plants. London, AcademicPress.
Hartnett, D.C., and Wilson, G.W.T. 1999. Mycorrhizae influenceon plant community structure and diversity in tallgrass prairie.Ecology, 80: 1187–1195.
Hartnett, D.C., Hetrick, B.A.D., Wilson, G.W.T., and Gibson, D.J.1993. Mycorrhizal influence on intra- and interspecific neigh-bour interactions among co-occurring prairie grasses. J. Ecol.81: 787–795.
Helgason, T., Merryweather, J.W., Denison, J., Wilson, P., Young,J.P.W., and Fitter, A.H. 2002. Selectivity and functional diver-sity in arbuscular mycorrhizas of co-occuring fungi and plantsfrom a temperate deciduous woodland. J. Ecol. 90: 371–384.
Hirrel, M.C., and Gerdemann, J.W. 1979. Enhanced carbon transferbetween onions infected with a vesicular–arbuscular mycor-rhizal fungus. New Phytol. 83: 731–738.
Horton, T.R., and Bruns, T.D. 1998. Multiple-host fungi are themost frequent and abundant ectomycorrhizal types in a mixedstand of Douglas-fir (Pseudotsuga menziesii) and bishop pine(Pinus muricata). New Phytol. 139: 331–339.
Horton, T.R., Bruns, T.D., and Parker, V.T. 1999. Ectomycorrhizalfungi associated with Arctostaphylos contribute to Pseudotsugamenziesii establishment. Can. J. Bot. 77: 93–102.
Howe, H., Brown, J., and Zorn-Arnold, B. 2002. A rodent plagueon prairie diversity. Ecol. Lett. 5: 30–36.
Johansen, A., and Jensen, E.S. 1996. Transfer of N and P from in-tact or decomposing roots of pea to barley interconnected by anarbuscular mycorrhizal fungus. Soil Biol. Biochem. 28: 73–81.
Jones, M.D., Durall, D.M., Harniman, S.M.K., Classen, D.C., andSimard, S.W. 1997. Ectomycorrhizal diversity on Betulapapyrifera and Pseudotsuga menziesii seedlings grown in thegreenhouse or outplanted in single-species and mixed plots insouthern British Columbia. Can. J. For. Res. 27: 1872–1889.
Jones, M.D., Durall, D.M., and Cairney, J.W.G. 2003. Ectomy-corrhizal fungal communities in young forest stands regenerat-ing after clearcut logging. New Phytol. 157: 399–422.
© 2004 NRC Canada
1162 Can. J. Bot. Vol. 82, 2004

Jonsson, L., Dahlberg, A., Nilsson, M.-C., Zackrisson, O., andKaren, O. 1999a. Ectomycorrhizal fungal communities in late-successional Swedish boreal forests, and their composition fol-lowing wildfire. Mol. Ecol. 8: 205–215.
Jonsson, L., Dahlbergh, A., Nilsson, M.-C., Karen, O., andZackrisson, O. 1999b. Continuity of ectomycorrhizal fungi inself-regenerating boreal Pinus sylvestris forests studied by com-paring mycobiont diversity on seedlings and mature trees. NewPhytol. 142: 151–162.
Jonsson, L.M., Nilsson, M.-C., Wardle, D.A., and Zackrisson, O.2001. Context dependent effects of ectomycorrhizal speciesrichness on tree seedling productivity. Oikos, 93: 353–364.
Jumpponen, A., and Trappe, J.M. 1998. Dark septate endophytes: areview of facultative biotrophic root-colonizing fungi. NewPhytol. 140: 295–310.
Kennedy, P.G., Izzo, A.D., and Bruns, T.D. 2003. There is high po-tential for formation of common mycorrhizal networks betweenunderstorey and canopy trees in a mixed evergreen forest. J.Ecol. 91: 1071–1080.
Klironomos, J.N. 2002. Diversity of arbuscular mycorrhizal fungiand ecosystem functioning. In Mycorrhizal ecology. Edited byM.G.A. van der Heijden and I. Sanders. Springer-Verlag, Berlin.
Klironomos, J.N., and Moutoglis, P. 1999. Colonization of nonmy-corrhizal plants by mycorrhizal neighbours as influenced by thecollembolan, Folsomia candida. Biol. Fertil. Soils, 29: 277–281.
Klironomos, J.N., and Ursic, M. 1998. Density-dependent grazingon the extraradical hyphal network of the arbuscular mycor-rhizal fungus, Glomus intraradices, by the collembolan,Folsomia candida. Biol. Fertil. Soils, 26: 250–253.
Klironomos, J., and Kendrick, W.B. 1995. Stimulative effects ofarthropods on endomycorrhizas of sugar maple in the presenceof decaying litter. Funct. Ecol. 9: 528–536.
Kranabetter, J.M. 1999. The effect of refuge trees on a paper birchectomycorrhiza community. Can. J. Bot. 77: 1523–1528.
Kranabetter, J.M., Hayden, S., and Wright, E.F. 1999. A compari-son of ectomycorrhiza communities from three conifer speciesplanted on forest gap edges. Can. J. Bot. 77: 1193–1198.
Kytoviita, M.M., Vestberg, M., and Tuomi, J. 2003. A test of mu-tual aid in common mycorrhizal networks: established vegeta-tion negates benefit in seedlings. Ecology, 84: 898–906.
Lammers, P.J., Jun, J., Abubaker, J., Arreola, R., Gopalan, A.,Bago, B., Sebastia, C.-H., Allen, J.W., Douds, D.D., Pfeffer,P.E., and Shachar-Hill, Y. 2001. The glyoxylate cycle in anarbuscular mycorrhizal fungus. Carbon flux and gene expres-sion. Plant Physiol. 127: 1287–1298.
Larsen, J., Thingstrup, I., Jakobsen, I., and Rosendahl, S. 1996.Benomyl inhibits phosphorus transport but not fungal alkalinephosphatase activity in a Glomus–cucumber symbiosis. NewPhytol. 132: 127–133.
Leake, J.R. 1994. The biology of myco-heterotrophic (saprophytic)plants. New Phytol. 127: 171–216.
Lerat, S., Gauci, R., Catford, J.G., Vierheilig, H., Piché, Y., andLapointe, L. 2002. 14C transfer between the spring ephemeralErythronium americanum and sugar maple saplings via arbus-cular mycorrhizal fungi in natural stands. Oecologia, 132: 181–187.
Lewis, D.H., and Harley, J.L. 1965. Carbohydrate physiology ofmycorrhizal roots of beech. II. Utilization of exogenous sugarsby uninfected and mycorrhizal roots. New Phytol. 64: 238–256.
Marler, M.J., Zabinski, C.A., and Callaway, R.M. 1999. Mycor-rhizae indirectly enhance competitive effects of an invasive forbon a native bunchgrass. Ecology, 80: 1180–1186.
Martensson, A.M., Rydberg, I., and Vestberg, M. 1998. Potential to
improve transfer of N in intercropped systems by optimizinghost–endophyte combinations. Plant Soil, 205: 57–66.
Martin, F. 1985. 15N-NMR studies of nitrogen assimilation andamino acid biosynthesis in the ectomycorrhizal fungus Ceno-coccum graniforme. FEBS Letters, 180: 350–354
Martin, F., Ramstedt, M., Soderhall, K., and Canet, D. 1988. Car-bohydrate and amino acid metabolism in the ectomycorrhizalascomycete Sphaerosporella brunnea during glucose utilization.A 13C NMR study. Plant Physiol. 86: 935–940.
Martin, F., Lapeyrie, F., and Tagu, D. 1997. Altered gene expres-sion during ectomycorrhizal development. In The Mycota. Vol.V. Springer-Verlag, Berlin. pp 223–242.
Martin, F., Boiffin, V., and Pfeffer, P.E. 1998. Carbohydrate andamino acid metabolism in the Eucalyptus globules – Pisolithustinctorius ectomycorrhiza during glucose utilization. PlantPhysiol. 118: 627–635.
Martins, M.A. 1993. The role of the external mycelium of arbus-cular mycorrhizal fungi in the carbon transfer process betweenplants. Mycol. Res. 97: 807–810.
Massicotte, H.B., Molina, R., Luoma, D.L., and Smith, J.E. 1994.Biology of the ecto-mycorrhizal genus, rhizopogon. II. Patternsof host–fungus specificity following spore inoculation of diversehosts grown in monoculture and dual culture. New Phytol. 126:677–690.
McKendrick, S.L., Leake, J.R., and Read, D.J. 2000. Symbioticgermination and development of myco-heterotrohic plants in na-ture: transfer of carbon from ectomycorrhizal Salix repens andBetula pendula to the orchid Corallorhiza trifida through sharedhyphal connections. New Phytol. 145: 539–548.
Miller, S.L., and Allen, E.B. 1992. Mycorrhizae, nutrient trans-location, and interactions between plants. In Mycorrhizal func-tioning: an integrative plant–fungal process. Edited by M.F.Allen. Chapman and Hall, New York. pp. 301–325.
Molina, R., Massicotte, H., and Trappe, J.M. 1992. Specificity phe-nomena in mycorrhizal symbiosis: community–ecological con-sequences and practical implications. In Mycorrhizalfunctioning: an integrative plant–fungal process. Edited by M.F.Allen. Chapman and Hall, New York. pp. 357–423.
Moora, M., and Zobel, M. 1996. Effect of arbuscular mycorrhizaon inter- and intraspecific competition of two grassland species.Oecologia, 108: 79–84.
Näsholm, T., Ekblad, A., Nordin, A., Giesler, R., Högberg, M., andHögberg, P. 1998. Boreal forest plants take up organic nitrogen.Nature (Lond.), 392: 914–916.
Newberry, D.M., Alexander, I.J., and Rother, J.A. 2000. Doesproximity to conspecific adults influence the establishment ofectomycorrhizal trees in rain forest? New Phytol. 147: 401–409.
Newman, E.I. 1988. Mycorrhizal links between plants: their func-tioning and ecological significance. Adv. Ecol. Res. 18: 243–270.
Newman, E.I., Ritz, K., and Jupp, A.P. 1989. The functioning ofroots in the grassland ecosystem. Aspects Appl. Biol. 22: 263–269.
Onguene, N.A., and Kuyper, T.W. 2002. Importance of the ectomy-corrhizal network for seedling survival and ectomycorrhizaformation in rain forests of south Cameroon. Mycorrhiza, 12:13–17.
Perry, D.A. 1998. A moveable feast: the evolution of resource shar-ing in plant–fungus communities. Trends Ecol. Evol. 13: 432–434.
Perry, D.A. 1999. Reply from D.A. Perry. Trends Ecol. Evol. 14:70–71.
Perry, D.A., Margolis, H., Choquette, C., Molina, R., and Trappe,J.M. 1989. Ectomycorrhizal mediation of competition betweenconiferous tree species. New Phytol. 112: 501–511.
© 2004 NRC Canada
Simard and Durall 1163

Perry, D.A., Bell, T., and Amaranthus, M.P. 1992. Mycorrhizalfungi in mixed-species forests and tales of positive feedback, re-dundancy and stability. In The ecology of mixed-species standsof trees. Edited by M.G.R. Cannell, D.C. Malcolm, and P.A.Robertson. Blackwell, Oxford. pp. 151–179.
Pfeffer, P.E., Douds, D.D., Becard, G., and Shachar-Hill, Y. 1999.Carbon uptake and the metabolism and transport of lipids in andarbuscular mycorrhiza. Plant Physiol. 120: 587–598.
Read, D.J. 2002. Towards ecological relevance: progress and pit-falls in the path towards an understanding of mycorrhizal func-tions in nature. In Mycorrhizal ecology. Edited by M.G.A. vander Heijden and I. Sanders. Springer-Verlag, Berlin.
Read, D.J., Francis, R., and Finlay, R.D. 1985. Mycorrhizal myce-lia and nutrient cycling in plant communities. In Ecological in-teractions in soil. Edited by A.H., Fitter, D. Atkinson, D.J. Read,and M.B. Usher. Blackwell Scientific, Oxford. pp. 193–217.
Reid, C.P.P., and Woods, F.W. 1969. Translocation of 14C-labeledcompounds in mycorrhizae and its implications in interplant nu-trient cycling. Ecology, 50: 179–187.
Rejon, A., Garcia-Romera, I., Ocampo, J.A., and Bethlenfalvay,G.J. 1997. Mycorrhizal fungi influence competition in a wheat–ryegrass association treated with the herbicide diclofop. Appl.Soil Ecol. 7: 51–57.
Robinson, D., and Fitter, A. 1999. The magnitude and control ofcarbon transfer between plants linked by a common mycorrhizalnetwork. J. Exp. Bot. 50: 9–13.
Sakakibara, S.M., Jones, M.D., Gillespie, M., Hagvermann, S.M.,Forrest, M.E., Simard, S.W., and Durall, D.M. 2002. A compari-son of ectomycorrhiza identification based on morphotyping andPCR-RFLP analysis. Mycol. Res. 106: 868–878.
Sawyer, N.A., Chambers, S.M., and Cairney, J.W.G. 2001. Distri-bution and persistence of Amanita muscaria genotypes in Aus-tralian Pinus radiata plantations. Mycol. Res. 105: 966–970.
Sen, R., Hietala, A.M., Zelmer, C.D. 1999. Common anastomosisand internal transcribed spacer RFLP groupings in binucleateRhizoctonia isolates representing root endophytes of Pinussylvestris, Ceratorhiza spp. from orchid mycorrhizas and aphytopathogenic anastomosis group. New Phytol. 144: 331–341.
Simard, S.W., Perry, D.A., Jones, M.D., Myrold, D.D., Durall,D.M., and Molina, R. 1997a. Net transfer of carbon betweenectomycorrhizal tree species in the field. Nature (Lond.), 388:579–582.
Simard, S.W., Molina, R., Smith, J.E., Perry, D.A., and Jones,M.D. 1997b. Shared compatibility of ectomycorrhizae onPseudotsuga menziesii and Betula papyrifera seedlings grown inmixture in soils from southern British Columbia. Can. J. For.Res. 27: 331–342.
Simard, S.W., Perry, D.A., Smith, J.E., and Molina, R. 1997c. Ef-fects of soil trenching on occurrence of ectomycorrhizas onPseudotsuga menziesii seedlings grown in mature forests ofBetula papyrifera and Pseudotsuga menziesii. New Phytol. 136:327–340.
Simard, S.W., Jones, M.D., Durall, D.M., Perry, D.A., Myrold,D.D., and Molina, R. 1997d. Reciprocal transfer of carbon iso-topes between ectomycorrhizal Betula papyrifera and Pseudo-tsuga menziesii. New Phytol. 137: 529–542.
Simard, S.W., Durall, D.M., and Jones, M.D. 1997e. Carbon allo-cation and carbon transfer between Betula papyrifera andPseudotsuga menziesii seedlings using a 13C pulse-labelingmethod. Plant Soil, 191: 41–55.
Simard, S.W., Jones, M.D., and Durall, D.M. 2002. Carbon and nu-trient fluxes within and between mycorrhizal plants. In Mycor-rhizal ecology. Edited by M.G.A. van der Heijden and I.Sanders. Springer-Verlag, Berlin. pp. 33–74.
Smith, S.E., and Smith F.A. 1990. Structure and function of the in-terface in biotrophic symbioses as they relate to nutrient trans-port. New Phytol. 114: 1–38.
Söderström, B., Finlay, R.D., and Read, D.J. 1988. The structureand function of the vegetative mycelium of ectomycorrhizalplants. IV. Qualitative analysis of carbohydrate contents of my-celium interconnecting host plants. New Phytol. 109: 163–166.
Taylor, D.L., and Bruns, T.D. 1997. Independent, specialized inva-sions of ectomycorrhizal mutualism by two nonphotosyntheticorchids. Proc. Natl. Acad. Sci. U.S.A. 94: 4510–4515.
Taylor, D.L., and Bruns, T.D. 1999. Population, habitat and geneticcorrelates of mycorrhizal specialization in the “cheating” or-chids Corallorhiza maculata and C. mertensiana. Mol. Ecol. 8:1719–1732.
Tilman, D., and Pacala, S. 1993. The maintenance of speciesrechness in plant communities. In Species diversity in ecologicalcommunities. Edited by R.E. Ricklefs and D. Schulter. The Uni-versity of Chicago Press, Chicago, Ill. pp. 13–25.
Tuffen, F., Eason, W.R., and Scullion, J. 2002. The effect of earth-worms and arbuscular mycorrhizal fungi on growth of and 32Ptransfer between Allium porrum plants. Soil Biol. Biochem. 34:1027–1036.
van der Heijden, M.G.A. 2002. Arbuscular mycorrhizal fungi as adeterminant of plant diversity: in search for underlying mecha-nisms and general principles. In Mycorrhizal ecology. Edited byM.G.A. van der Heijden and I. Sanders. Springer-Verlag, Berlin.
van der Heidjden, M.G.A., and Sanders, I.R. 2002. Mycorrhizalecology: synthesis and perspectives. In Mycorrhizal ecology.Edited by M.G.A. van der Heijden and I. Sanders. Springer-Verlag, Berlin.
van der Heijden, M.G.A., Boller, T., Wiemken, A., and Sanders,I.R. 1998. Different arbuscular mycorrhizal fungal species arepotential determinants of plant community structure. Ecology,76: 2082–2091.
van der Heijden, M.G.A., Wiemken, A., and Sanders, I.R. 2003.Different arbuscular mycorrhizal fungi alter coexistence andresource distribution between co-occurring plant. New Phytol.157: 569–578.
Vrålstad, T., Schumacher, T., and Taylor, A.F.S. 2002. Mycorrhizalsynthesis between fungal strains of the Hymenoscyphus ericaeaggregate and potential ectomycorrhizal and ericoid hosts. NewPhytol. 153: 143–152.
Walter, L.E.F., Hartnett, D.C., Hetrick, A.D., and Schwab, A.P.1996. Interspecific nutrient transfer in a tallgrass prairie plantcommunity. Am. J. Bot. 83: 180–184.
Waters, J.R., and Borowicz, V.A. 1994. Effect of clipping,benomyl, and genet on 14C transfer between mycorrhizal plants.Oikos, 71: 246–252.
Watkins, N.K., Fitter, A.H., Graves, J.D., and Robinson, D. 1996.Carbon transfer between C3 and C4 plants linked by a commonmycorrhizal network quantified using stable isotopes. Soil Biol.Biochem. 28: 471–477.
Wilkinson, D.A. 1998. The evolutionary ecology of mycorrhizalnetworks. Oikos, 82: 407–410.
Wu, B., Nara, K., and Hogetsu, T. 2001. Can 14C-labelled photo-synthetic products move between Pinus densiflora seedlingslinked by ectomycorrhizal mycelia? New Phytol. 149: 137–146.
Zabinski, C.A., Quinn, L., and Callaway, R.M. 2002. Phosphorusuptake, not carbon transfer, explains arbuscular mycorrhizal en-hancement of Centaurea maculosa in the presence of nativegrassland species. Funct. Ecol. 16: 758–765.
Zhou, Z., Miwa, M., Matsuda, Y., and Hogetsu, T. 2001. Spatialdistribution of the subterranean mycelia and ectomycorrhizae ofSuillus grevillei genets. J. Plant Res. 114: 179–185.
© 2004 NRC Canada
1164 Can. J. Bot. Vol. 82, 2004

Zhu, Y.-G., Laidlaw, A.S., Christie, P., and Hammond, M.E.R.2000. The specificity of arbuscular mycorrhizal fungi in a peren-nial ryegrass-white clover pasture. Agric. Ecosyst. & Environ.77: 211–218.
Zobel, M., and Moora, M. 1997. Plant coexistence in the interac-tive environment: arbuscular mycorrhiza should not be out ofmind. Oikos, 78: 202–208.
© 2004 NRC Canada
Simard and Durall 1165





























