Embed Size (px)
Citation preview

BioOne sees sustainable scholarly publishing as an inherently collaborative enterprise connecting authors, nonprofit publishers, academic institutions, researchlibraries, and research funders in the common goal of maximizing access to critical research.
Mycoplasma gallisepticum Hemagglutinin VlhA, Pyruvate DehydrogenasePdhA, Lactate Dehydrogenase, and Elongation Factor Tu Share Epitopes withMycoplasma imitans HomologuesAuthor(s): Miha Lavrič, Dušan Benčina, Mojca NaratSource: Avian Diseases, 49(4):507-513. 2005.Published By: American Association of Avian PathologistsDOI: http://dx.doi.org/10.1637/7377-051105R.1URL: http://www.bioone.org/doi/full/10.1637/7377-051105R.1
BioOne (www.bioone.org) is a nonprofit, online aggregation of core research in the biological, ecological, andenvironmental sciences. BioOne provides a sustainable online platform for over 170 journals and books publishedby nonprofit societies, associations, museums, institutions, and presses.
Your use of this PDF, the BioOne Web site, and all posted and associated content indicates your acceptance ofBioOne’s Terms of Use, available at www.bioone.org/page/terms_of_use.
Usage of BioOne content is strictly limited to personal, educational, and non-commercial use. Commercial inquiriesor rights and permissions requests should be directed to the individual publisher as copyright holder.

Mycoplasma gallisepticum Hemagglutinin VlhA, Pyruvate DehydrogenasePdhA, Lactate Dehydrogenase, and Elongation Factor Tu Share
Epitopes with Mycoplasma imitans Homologues
Miha Lavric, Dusan Bencina, and Mojca NaratA
Department of Animal Sciences, Biotechnical Faculty, University of Ljubljana, 1230 Domzale, Slovenia
Received 11 May 2005; Accepted 30 June 2005
SUMMARY. Mycoplasma gallisepticum is a major pathogen of poultry. Mycoplasma imitans is genetically and antigenically closelyrelated to M. gallisepticum, but so far, only a few proteins of M. imitans have been identified as sharing epitopes withM. gallisepticum. In this study, we identified three proteins of M. gallisepticum that share with M. imitans epitopes defined bymonoclonal antibodies (MAbs). MAb 9D4 reacted with the 67-kD hemagglutinin VlhA (previously termed pMGA) of M.gallisepticum and with its continuously expressed 40-kD protein. This MAb also reacted with a 40-kD protein of M. imitans, butnot with its putative VlhA. Two-dimensional (2D) immunoblots of M. gallisepticum strains showed that their 40-kD proteinsreacting with MAb 9D4 are expressed as major forms with isoelectric points (pI) around 6, and also as less-abundant formsdiffering in pI. In M. imitans, major forms of 40-kD proteins recognized by MAb 9D4 had pI around 6, whereas minor forms hadpI between 5.5 and 5.8. The N-terminal sequence of the M. gallisepticum 40-kD protein recognized by MAb 9D4 strongly indicatesthat this protein is pyruvate dehydrogenase E1, subunit a (PdhA protein, also termed AcoA). The position of elongation factor Tu(EF-Tu), detected by the reference MAb GB8, was very similar in the 2D proteome maps of M. gallisepticum and M. imitans (MWof about 45 kD; pI ; 5.6). In both M. gallisepticum and M. imitans, MAb 7G1 reacted with proteins of about 36 kD with similarcharges (major forms with pI of about 8). The position of this protein in the proteome map of M. gallisepticum and its N-terminalsequence strongly suggest that MAb 7G1 recognizes lactate (malate) dehydrogenase (Ldh or Mdh). Comparison of 2D proteomesof 10 M. gallisepticum strains indicated that positions of EF-Tu, PdhA, and Ldh proteins are rather consistent and can be used asreference points in further analyses of the M. gallisepticum proteome.
RESUMEN. Las proteınas V1hA, piruvato deshidrogenasa PdhA, lactato deshidrogenasa y factor de elongacion Tu delMycoplasma gallisepticum comparten epıtopes con sus homologas en el Mycoplasma imitans.
El Mycoplasma gallisepticum es un patogeno de gran importancia para la avicultura. El Mycoplasma imitans esta genetica yantigenicamente relacionado con el M. gallisepticum, pero hasta ahora, solo contadas proteınas del M. imitans han sido identificadascomo compartiendo epıtopes con el M. gallisepticum. En el presente estudio fueron determinadas tres proteınas del M. gallisepticumque comparten epıtopes con el M. imitans, mediante anticuerpos monoclonales. El anticuerpo monoclonal 9D4 reacciono con lahemoaglutinina V1hA de 67-kDs del M. gallisepticum (previamente denominada pMGA) y con su proteına de expresion continuade 40-kD. Este anticuerpo monoclonal tambien reacciono con la proteına de 40-kD del M. imitans, mas no con su V1hA putativa.La inmunotransferencia puntual de dos dimensiones en cepas de M. gallisepticum demostro que la proteına de 40-kD que reaccionocon el anticuerpo monoclonal 9D4 se expresa mayoritariamente en formas con un punto isoelectrico de aproximadamente 6 y a suvez, en menor numero con formas que difieren en sus puntos isoelectricos. En el M. imitans las formas mayoritarias de la proteına de40-kD reconocida por el anticuerpo monoclonal 9D4 tienen un punto isoelectrico cercano a 6, mientras formas minoritariaspresentaron puntos isoelectricos de 5.5 y 5.8. La secuencia de aminoacidos del extremo N-terminal de la proteına de 40-kD del M.gallisepticum reconocida por el anticuerpo monoclonal 9D4, indica fuertemente que esta proteına se trata de piruvatodeshidrogenasa E1, subunidad a (proteına PdhA, tambien denominada AcoA). La posicion del factor de elongacion (EF-Tu)detectado por el anticuerpo monoclonal de referencia GB8, fue muy similar en los mapas proteomicos de dos dimensiones del M.gallisepticum y el M. imitans (peso molecular de aproximadamente 45-kD; punto isoelectrico de aproximadamente 5.6). En ambos,M. gallisepticum y M. imitans, el anticuerpo monoclonal 7G1 reacciono con proteınas de aproximadamente 36-kD con cargassimilares (formas mayoritarias con un punto isoelectrico de 8). La posicion de esta proteına en el mapa proteomico del M.gallisepticum y la secuencia de aminoacidos de su extremo N-terminal sugieren fuertemente que el anticuerpo monoclonal 7G1reconoce las proteınas lactato (malato) dehidrogenasa (LdH o MdH). La comparacion de los mapas proteomicos de 10 cepas de M.gallisepticum indico que las posiciones de las proteınas Ef-Tu, PdhA y Ldh son constantes y que pueden ser utilizadas como puntosde referencia en futuros analisis de mapas proteomicos para M. gallisepticum.
Key words: Mycoplasma gallisepticum, Mycoplasma imitans, PdhA, Ldh, EF-Tu
Abbreviations: 2D¼ two dimensional; EF-Tu¼ elongation factor Tu; HRP¼ horseradish peroxidase; IEF¼ isoelectric focusing;IIPA ¼ indirect immunoperoxidase assay; Ldh (Mdh) ¼ lactate (malate) dehydrogenase; MAb ¼ monoclonal antibody; MW ¼molecular weight (or molecular mass); PdhA ¼ pyruvate dehydrogenase E1, subunit alpha (termed also AcoA); pI ¼ isoelectricpoint; SDS-PAGE¼ sodium dodecyl sulfate polyacrylamide gel electrophoresis; VlhA¼ variable lipoprotein hemagglutinin (in M.gallisepticum previously termed pMGA)
Mycoplasma gallisepticum is a major respiratory pathogen ofchickens and turkeys (21). Mycoplasma gallisepticum strains varywidely in their pathogenicity, tissue tropism, and in the immuneresponse they induce (17,30). Less pathogenic and immunogenic
M. gallisepticum strains designated as variant or atypical strains lackepitopes on several surface-exposed immunogenic proteins (3,30).Mycoplasma imitans is genetically and antigenically closely related toM. gallisepticum (8). These two species and M. iowae phylogenet-ically belong to the pneumoniae group, whereas other recognizedavian Mycoplasma species belong to the hominis group (16).ACorresponding author.
AVIAN DISEASES 49:507–513, 2005
507

Previous studies have suggested that M. gallisepticum proteins withdifferent molecular masses share epitopes with proteins of otherMycoplasma species (1,2,9,27). Only in a few cases have M.gallisepticum proteins sharing epitopes been identified. Recently, ithas been found that M. gallisepticum dihydrolipoamide acetyltrans-ferase, which is a component of the pyruvate dehydrogenasecomplex, shares epitopes with a similarly sized protein of M. imitans(15). These two species also share epitopes on their hemagglutininsVlhA (in M. gallisepticum termed previously pMGA) and on variableperipheral membrane proteins of 41 kD that share epitopes with thevariably expressed cytadhesin PvpA of M. gallisepticum (22,27).
Monoclonal antibodies (MAbs) such as MAb 9D4 have beenfound to recognize variable epitopes on proteins of about 67–70 kDof typical M. gallisepticum strains, as well as invariable epitopespresent on 40–44-kD proteins of typical and variant M. gallisepticumstrains (3,12). Further study showed that 67–70 kD-proteins wereantigenic variants of the hemagglutinin pMGA (VlhA), whereas aninvariable 40-kD protein sharing epitopes with VlhA was notidentified (23). In all M. gallisepticum strains examined, MAb 7G1detected an invariably expressed protein of about 36 kD, but it wasnot identified (3).
In this study we examined M. imitans strains for proteinsrecognized by MAbs 9D4, 7G1, and GB8, which react with majorproteins of M. gallisepticum. Positions of those proteins in 2Dproteome map of M. gallisepticum were determined as well as theirN-terminal (amino acid) sequences.
MATERIALS AND METHODS
Mycoplasma species and their strains. Mycoplasma gallisepticumstrains used have already been examined with MAbs in previous studies(3,12,23). Reference standard strains were type strain PG 31 (x95) andstrains R (K781), F (K810), A5969, S6 (culture 208), and S6B (3).Other strains were ‘‘typical’’ strains G11, PET2, IHB1, Alex, and PPT2and ‘‘atypical’’ or variant strains K503 and K703 (3,12). Mycoplasmaimitans strains 4229 (type strain) and B2/85 were kindly donated byJanet M. Bradbury (8). Type strains of Mycoplasma synoviae (WVU1853) and Mycoplasma iowae (I-695) were also used (6,12). In addition,reference strains of Mycoplasma pneumoniae (M129) and Mycoplasmagenitalium (G37) were assayed for epitopes reacting with MAbs 9D4,7G1, and GB8 (5,20).
Monoclonal antibodies. MAbs 9D4 (IgG2a isotype) and 7G1(IgG1) have already been used in studies of M. gallisepticum strains(3,12,23). MAb GB8 (IgG1), recognizing EF-Tu proteins of allMycoplasma species examined (18), was obtained from Birgit Henrich.MAb 1E5 (IgM), recognizing M. gallisepticum variably expressedcytadhesin PvpA and a 41-kD peripheral-membrane protein undergoing
variable expression in M. gallisepticum and M. imitans, was received fromRenate Rosengarten (27). Rabbit anti-pMGA antibodies recognizing anapproximately 67-kD pMGA (i.e., VlhA) of M. gallisepticum and anapproximately 35-kD protein (putative VlhA) of M. imitans (22) werereceived from Philip F. Markham. In addition, rabbit antibodies to M.gallisepticum conjugated with HRP were used in immunoblots to revealmajor immunogenic proteins of M. gallisepticum separated by 2D SDS-PAGE (3,13)
Immunoblotting. Immunoblot procedures used to detect M.gallisepticum and M. imitans proteins by MAbs were described in detailelsewhere (3,23) Usually, the PhastSystem (Pharmacia-LKB, Uppsala,Sweden) was used to determine molecular masses and isoelectric points(pIs) of mycoplasma proteins reacting with MAbs 9D4, 7G1, GB8, and1E5 (3,5,23). However, for improved mapping of M. gallisepticumproteins recognized by MAbs, proteins were separated in larger gels (1103 170 mm) using 2D PAGE (3,13). They were transferred ontoImmobilon-P membrane (Millipore, Billerica, MA) and probed byappropriate antibodies (3,4).
SDS-PAGE. 2D PAGE for protein-mapping of 10 M. gallisepticumstrains and of M. imitans (strain 4229) was performed in 110 3 170 mmgels and their proteins were stained using sensitive silver staining(3,7,13).
Indirect immunoperoxidase assay (IIPA). IIPA was used todetermine whether proteins and their corresponding epitopes recognizedby MAbs 9D4, 7G1, GB8, and 1E5 were exposed on the surface ofMycoplasma colonies (3).
Amino-terminal sequencing of proteins. Mycoplasma galli-septicum cells (strains R[K781] or A5969) were treated with 1% TritonX-100 for 30 min. Following centrifugation (20,000 3 g for 20 min) thesupernatants were analyzed in immunoblots to locate the position ofproteins recognized with appropriate MAbs. Proteins were separated bySDS-PAGE (120 3 100 3 0.5 mm) and transferred onto ImmobilonP membrane (4). The positions of the proteins detected with appro-priate MAbs (9D4, 7G1, or GB8) were determined on the right half ofthe strip bearing M. gallisepticum proteins. The corresponding proteinband was cut out from the second half of the strip and submitted forN-terminal sequencing with automated Edman degradation using anABI (model 492A; Applied Biosystems, Foster City, CA) proteinsequencer.
Identification of proteins. The N-terminal sequences deter-mined for the M. gallisepticum proteins detected by MAbs 9D4, 7G1, orGB8 were compared with protein sequences in databases, particularlywith those derived from the M. gallisepticum genome sequence (24).
The predicted amino acid sequence of an appropriate protein wasused to predict theoretical MW and pI, using Compute pI/MW tool ofthe ExPASy Proteomic tools and ExPASy server (http://www.expasy.org/tools/). Predicted molecular masses and pIs of proteins withcorresponding N-terminal sequences were compared with thoseestimated for proteins reacting with MAbs in 2D immunoblots (seeFigs. 3, 4, 5). We then analyzed whether an appropriate protein, i.e., EF-Tu, PdhA, or Ldh could be found at similar position in 2D map ofdifferent M. gallisepticum strains.
Fig. 1. Immunoblot analysis of M. gallisepticum and M. imitansproteins recognized with MAb 9D4. Mycoplasma gallisepticum strains arein the following lanes: 1, K503; 2, PG 31; 5, S6; 7, ULB931; and 8, R.Mycoplasma imitans strains are in these lanes: 3, B2/85; and 4, 4229.Lane 6 is the molecular weight marker. Positions of VlhA (pMGA) of 67kD and 40-kD protein (P40) are indicated at the right side. Positionscorresponding to 67 kD (bovine serum albumin) and 43 kD (chickenovalbumin) are indicated at the left side and in lane 6.
Fig. 2. Immunoblot analysis of M. gallisepticum and M. imitansproteins recognized with MAb 7G1. Mycoplasma gallisepticum strainsare in the following lanes: 1, K503; 2, PG 31; 5, S6; 7 and 8, twodifferent cultures of strain R. Mycoplasma imitans strains are in theselanes: 3, B2/85; and 4, 4229. Lane 6 is the molecular weight marker.Positions corresponding to 43 and 30 kD are indicated at the left sideand in lane 6.
508 M. Lavric et al.

RESULTS
Mycoplasma gallisepticum and M. imitans share epitopesfor MAb 9D4. As expected, in the IIPA, MAb 9D4 revealedvariable staining of colonies of typical M. gallisepticum strains, whichwas positively correlated with the expression of VlhA, i.e., pMGA(3,23). This MAb did not react with colonies of variant strains K503and 703 or with colonies of M. imitans strains 4229 and B2/85. Inimmunoblots MAb 9D4 reacted with proteins of about 40-kD fromM. imitans (strains 4229 and B2/85) and with similarly sizedproteins of all M. gallisepticum strains examined (Fig. 1 and data inreferences 3 and 12). However, in typical M. gallisepticum strains,MAb 9D4 reacted also with the approximately 67-kD VlhA, whichwas not detected in variant strain K503 or M. imitans strains (Fig.1). MAb 9D4 strongly reacted with immunoaffinity purified VlhAof 67 kD (i.e., pMGA) obtained from strain PG31 using MAb K1and from strain S6 (culture 208) using MAb 71 (23). On the otherhand, MAb 9D4 did not react with a putative VlhA of M. imitans(an approximately 35-kD protein in strain 4229) recognized byrabbit antibodies to M. gallisepticum VlhA (22 and data not shown).
In immunoblotting of M. imitans proteins separated by isoelectricfocusing (IEF, PhastGel 3-9), MAb 9D4 reacted with a major
protein band with a pI of about 6, and with less-abundant proteinshaving pIs between 5.5 and 5.8. Similar pI values have beendocumented for isoelectric forms of the invariable 40-kD proteindetected by this MAb in M. gallisepticum strain IHB1 (23). MAb9D4 did not react with M. pneumoniae, M. genitalium, M. synoviae,or M. iowae (3,12).
MAb GB8, which reacted with the EF-Tu protein of Mycoplasmaspecies, including M. gallisepticum (18), detected in this studyproteins of about 45 kD in M. imitans strains (4229 and B2/85) andin all M. gallisepticum strains examined. In both M. gallisepticum andM. imitans, major forms of putative EF-Tu detected by MAb GB8have pIs between 5.5 and 6. However, isoelectric isoforms (two orthree minor bands) were detected by MAb GB8 in several M.gallisepticum strains and in M. imitans.
M. gallisepticum and M. imitans share epitopes for MAb7G1. MAb 7G1, which does not react in the IIPA with colonies ofM. gallisepticum (3), did not react with M. imitans strains colonies.On the other hand, in immunoblots MAb 7G1 recognized abundantproteins of about 36 kD in M. imitans and in all strains of M.gallisepticum (Fig. 2). Immunoblotting following IEF of M. imitansproteins showed that MAb 7G1 recognized a major protein with a pIof about 8. In M. gallisepticum strains, major forms of 36-kD
Fig. 3. 2D proteome of M. gallisepticum, PG 31(X-95) and positions of EF-Tu, PdhA, and Ldh. Panel A, proteins were separated by IEF (pH 4–10) followed by SDS-PAGE and silver staining. Proteins, EF-Tu (label E, ;45 kD, pI of a major spot ;5.6), PdhA (label P, ;40 kD, pI of a majorspot ;6), and Ldh (label L, ;36 kD, pI of a major spot ;7.7) were identified by immunoblotting using MAbs GB8, 9D4, and 7G1. Panel B,positions of proteins recognized by MAbs 9D4 or 7G1. The membrane was first immunostained with rabbit antibodies (conjugated with HRP) toM. gallisepticum to identify positions of certain immunogenic proteins, e.g., of VlhA (indicated by arrow, ;67 kD, pI ;5). Consecutive reaction ofan appropriate part of the membrane with MAb 9D4 revealed the position of PdhA (spots under label P) forms with similar MW (;40 kD), butwith different pI values. Another section from the same membrane was probed with MAb 7G1 which showed the position of Ldh (spots under labelL) having similar MW (;36 kD) but different pI values. Panel C, an appropriate part from another membrane bearing 2D separated proteins ofstrain PG 31 was probed with MAb 7G1 to reveal more clearly the pattern of protein spots recognized by this MAb.
Mycoplasma gallisepticum PdhA, Ldh, and EF-Tu 509

proteins recognized by MAb 7G1 had similar pIs, whereas minorforms had less basic pIs (see Figs. 3, 4). This MAb did not react withM. pneumoniae, M. genitalium, M. synoviae, or M. iowae (3).
MAb 1E5 revealed variable expression of a putative cytadhesinPvpA in M. gallisepticum strains as has been reported (27). However,in M. gallisepticum strains, MAb 1E5 did not react with proteins ofabout 40 kD, which have been described as peripheral-membraneproteins undergoing variation in expression among clonal isolates ofM. gallisepticum and M. imitans (27). In addition, M. imitanscultures, which abundantly synthesized the 40-kD protein definedby MAb 9D4, did not react with MAb 1E5 (data not shown).
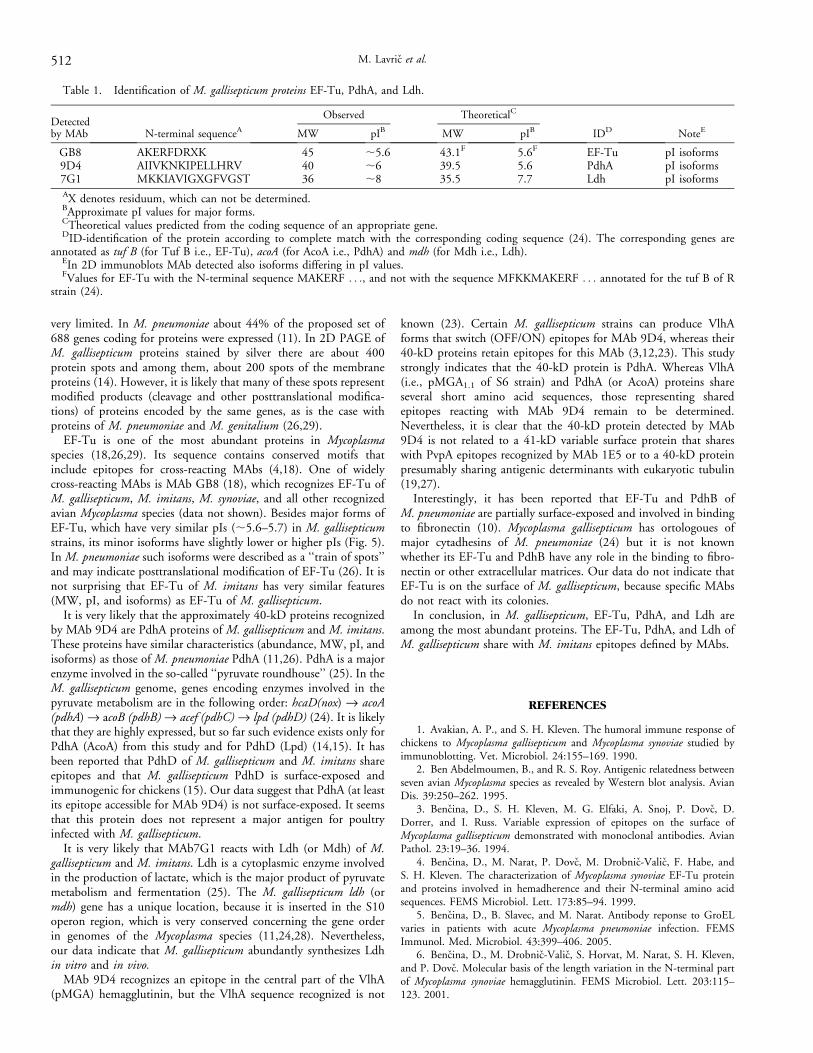
N-terminal sequences of M. gallisepticum EF-Tu, PdhA,and Ldh. N-terminal amino acid sequences were determined forM. gallisepticum (strains R[K781] and A5969) proteins reacting withMAbs GB8, 9D4 (40-kD proteins), and 7G1 (Table 1). Thesequence of the 45-kD protein reacting with MAb GB8 was identicalto the sequence of the EF-Tu protein. The approximately 40-kDprotein reacting with MAb 9D4 had the sequence AIIVKNKI-PELLHRV, which is identical to the sequence predicted for M.gallisepticum PdhA (or AcoA, amino acids 2–16). The approximately36-kD protein recognized by MAb 7G1 revealed the followingsequence: MKKIAVIGXGFKGST (the ninth residuum could not bedetermined but was probably C). This sequence is identical in all
determined residues to the sequence predicted from the ldh gene ofthe A5969 strain (GenBank accession number AF 036708) and tothe mdh gene of strain R (GenBank accession number NP 825855).
Positions of EF-Tu and putative PdhA and Ldh in theproteome of M. gallisepticum. We examined whether the 40-kDprotein defined by MAb 9D4 and the 36-kD protein recognized byMAb 7G1 had in the M. gallisepticum proteome positionscorresponding to theoretical (predicted) molecular masses and pIsof PdhA and Ldh (see Table 1). In the 2D proteome of the typestrain PG 31 (x95), MAb 9D4 detected a major protein of 40 kDwith a pI of about 6, and also an isoform with a pI of about 5.8 (Fig.3A,B). For comparison, the EF-Tu protein (about 45 kD) revealedmajor form with a pI of 5.6, which is consistent with its predicted pI(see Fig. 3A and Table 1).
In M. gallisepticum strain G11, MAb 9D4 detected a majorprotein of 40 kD that had a pI similar to that of strain PG 31, as wellas isoforms (Fig. 4). Major form of the 40-kD protein was slightlyless acidic in comparison with the major form of the EF-Tu protein(45 kD, pI ; 5.6). In other M. gallisepticum strains (A 5969, R,S6[208], S6B) the position of the 40-kD protein reacting with MAb9D4 was very similar to that described for strains PG 31 and G11(see Fig. 5). It is very likely that this MAb recognized an epitope onPdhA of M. gallisepticum and probably of M. imitans.
Fig. 4. 2D proteome of M. gallisepticum strain G11 and positions of EF-Tu, PdhA, and Ldh. Panel A, proteins were separated by isoelec-tric focusing (pH 4–10) followed by SDS-PAGE and stained with silver. Proteins, EF-Tu (label E, ;45 kD, pI of major spot 5.6). PdhA (label P,;40 kD, pI of major spot ;6), and Ldh (label L, ;36 kD, pI of major spot , 8) were identified by immunoblotting using MAbs GB8, 9D4, and7G1. Panel B, positions of proteins recognized by MAbs 9D4 or 7G1. The membrane was first immunostained with antibodies to M. galli-septicum to identify positions of certain immunogenic proteins e.g., of an approximately 50-kD protein appearing as a ‘‘train of spots’’ with differentpI values (under label P50). In panel A, more acidic forms of P50 (pI ;5.6) are positioned above EF-Tu. An appropriate part of the mem-brane was then probed with MAb 9D4 to show the position of PdhA (;40 kD, pI of major spot ;6). Panel C, in the next immunostainingusing MAb7G1, the position of Ldh was revealed. Most forms had similar MW (;36 kD), but differed in pI values.
510 M. Lavric et al.

In a 2D immunoblot of the PG 31 strain, MAb 7G1 reacted withthe 36-kD protein that appeared in isoforms differing in pI (Fig.3B,C). Major forms had pIs between 7.5 and 8, whereas minorisoforms had lower pIs. In the 2D proteome of the G11 strain, thepattern of proteins (isoforms) recognized by this MAb was similar tothat of the type strain PG 31. These data strongly indicate that MAb7G1 reacted with M. gallisepticum Ldh (or Mdh), which seems toshare epitopes with related proteins of M. imitans (similar size and pI).
Comparison of the proteomes of M. gallisepticum andM. imitans. Ten M. gallisepticum strains and M. imitans (4229)were analyzed by 2D SDS-PAGE and their proteins stained bysensitive silver staining. Whereas there were notable differencesconcerning the expression of many proteins, e.g., VlhA forms, in thispaper the focus is on the proteome region that includes EF-Tu,PdhA, and Ldh (MW 30–55 kD, pI 5–8.5). Fig. 5 shows relevantinserts of five M. gallisepticum strains and of M. imitans strain 4229.Notably, the positions of EF-Tu and of major forms of PdhA andLdh seem to be rather similar in all M. gallisepticum strains. This isobvious for EF-Tu (major form, 45 kD, pI about 5.6) which is
below a major protein (about 50 kD) that revealed three to fivemajor isoforms. Notably, in the 2D proteome map of the humanpathogen M. pneumoniae the positions of EF-Tu and PdhA are verysimilar to the positions of EF-Tu and putative PdhA in M.gallisepticum strains (Fig. 5 and Figs. 1, 2 in reference 26). Moreover,in the M. pneumoniae proteome Ldh has similar position as putativeLdh of M. gallisepticum. On the other hand, in M. pneumoniaeabundantly expressed PdhB is some 5 kD under the position ofPdhA (26), whereas there is no major protein in this region in theproteome of M. gallisepticum (Fig. 5).
DISCUSSION
Recently, the complete genome sequence of M. gallisepticum (Rstrain) has been determined (24). The M. gallisepticum genomeencodes about 50 proteins more than that of M. pneumoniae (11,24).However, in comparison with the well-characterized M. pneumoniaeproteome, the current knowledge of the M. gallisepticum proteome is
Fig. 5. Comparisons of the proteome regions containing EF-Tu, PdhA, and Ldh of five M. gallisepticum strains and M. imitans. 2D SDS-PAGE–separated proteins of M. gallisepticum strains F(K810), R(K781), S6 (culture 208), S6B, PET2, and M. imitans (4229) were stained by silver. Regionsincluding EF-Tu, PdhA, and Ldh (proteins with MW between ;35–55 kD and pI between ;4.5–9) are presented. EF-Tu (;45 kD, pI ;5.6)forms are under label E. PdhA (under label P) of about 40 kD, revealed isoforms differing in pI values. Ldh (under label L) of about 36 kD alsorevealed isoforms with different pI values, but its major forms had pI between 7 and 8 in all strains examined and also in M. imitans.
Mycoplasma gallisepticum PdhA, Ldh, and EF-Tu 511
very limited. In M. pneumoniae about 44% of the proposed set of688 genes coding for proteins were expressed (11). In 2D PAGE ofM. gallisepticum proteins stained by silver there are about 400protein spots and among them, about 200 spots of the membraneproteins (14). However, it is likely that many of these spots representmodified products (cleavage and other posttranslational modifica-tions) of proteins encoded by the same genes, as is the case withproteins of M. pneumoniae and M. genitalium (26,29).
EF-Tu is one of the most abundant proteins in Mycoplasmaspecies (18,26,29). Its sequence contains conserved motifs thatinclude epitopes for cross-reacting MAbs (4,18). One of widelycross-reacting MAbs is MAb GB8 (18), which recognizes EF-Tu ofM. gallisepticum, M. imitans, M. synoviae, and all other recognizedavian Mycoplasma species (data not shown). Besides major forms ofEF-Tu, which have very similar pIs (;5.6–5.7) in M. gallisepticumstrains, its minor isoforms have slightly lower or higher pIs (Fig. 5).In M. pneumoniae such isoforms were described as a ‘‘train of spots’’and may indicate posttranslational modification of EF-Tu (26). It isnot surprising that EF-Tu of M. imitans has very similar features(MW, pI, and isoforms) as EF-Tu of M. gallisepticum.
It is very likely that the approximately 40-kD proteins recognizedby MAb 9D4 are PdhA proteins of M. gallisepticum and M. imitans.These proteins have similar characteristics (abundance, MW, pI, andisoforms) as those of M. pneumoniae PdhA (11,26). PdhA is a majorenzyme involved in the so-called ‘‘pyruvate roundhouse’’ (25). In theM. gallisepticum genome, genes encoding enzymes involved in thepyruvate metabolism are in the following order: hcaD(nox) fi acoA(pdhA) fi acoB (pdhB) fi acef (pdhC) fi lpd (pdhD) (24). It is likelythat they are highly expressed, but so far such evidence exists only forPdhA (AcoA) from this study and for PdhD (Lpd) (14,15). It hasbeen reported that PdhD of M. gallisepticum and M. imitans shareepitopes and that M. gallisepticum PdhD is surface-exposed andimmunogenic for chickens (15). Our data suggest that PdhA (at leastits epitope accessible for MAb 9D4) is not surface-exposed. It seemsthat this protein does not represent a major antigen for poultryinfected with M. gallisepticum.
It is very likely that MAb7G1 reacts with Ldh (or Mdh) of M.gallisepticum and M. imitans. Ldh is a cytoplasmic enzyme involvedin the production of lactate, which is the major product of pyruvatemetabolism and fermentation (25). The M. gallisepticum ldh (ormdh) gene has a unique location, because it is inserted in the S10operon region, which is very conserved concerning the gene orderin genomes of the Mycoplasma species (11,24,28). Nevertheless,our data indicate that M. gallisepticum abundantly synthesizes Ldhin vitro and in vivo.
MAb 9D4 recognizes an epitope in the central part of the VlhA(pMGA) hemagglutinin, but the VlhA sequence recognized is not
known (23). Certain M. gallisepticum strains can produce VlhAforms that switch (OFF/ON) epitopes for MAb 9D4, whereas their40-kD proteins retain epitopes for this MAb (3,12,23). This studystrongly indicates that the 40-kD protein is PdhA. Whereas VlhA(i.e., pMGA1.1 of S6 strain) and PdhA (or AcoA) proteins shareseveral short amino acid sequences, those representing sharedepitopes reacting with MAb 9D4 remain to be determined.Nevertheless, it is clear that the 40-kD protein detected by MAb9D4 is not related to a 41-kD variable surface protein that shareswith PvpA epitopes recognized by MAb 1E5 or to a 40-kD proteinpresumably sharing antigenic determinants with eukaryotic tubulin(19,27).
Interestingly, it has been reported that EF-Tu and PdhB ofM. pneumoniae are partially surface-exposed and involved in bindingto fibronectin (10). Mycoplasma gallisepticum has ortologoues ofmajor cytadhesins of M. pneumoniae (24) but it is not knownwhether its EF-Tu and PdhB have any role in the binding to fibro-nectin or other extracellular matrices. Our data do not indicate thatEF-Tu is on the surface of M. gallisepticum, because specific MAbsdo not react with its colonies.
In conclusion, in M. gallisepticum, EF-Tu, PdhA, and Ldh areamong the most abundant proteins. The EF-Tu, PdhA, and Ldh ofM. gallisepticum share with M. imitans epitopes defined by MAbs.
REFERENCES
1. Avakian, A. P., and S. H. Kleven. The humoral immune response ofchickens to Mycoplasma gallisepticum and Mycoplasma synoviae studied byimmunoblotting. Vet. Microbiol. 24:155–169. 1990.
2. Ben Abdelmoumen, B., and R. S. Roy. Antigenic relatedness betweenseven avian Mycoplasma species as revealed by Western blot analysis. AvianDis. 39:250–262. 1995.
3. Bencina, D., S. H. Kleven, M. G. Elfaki, A. Snoj, P. Dovc, D.Dorrer, and I. Russ. Variable expression of epitopes on the surface ofMycoplasma gallisepticum demonstrated with monoclonal antibodies. AvianPathol. 23:19–36. 1994.
4. Bencina, D., M. Narat, P. Dovc, M. Drobnic-Valic, F. Habe, andS. H. Kleven. The characterization of Mycoplasma synoviae EF-Tu proteinand proteins involved in hemadherence and their N-terminal amino acidsequences. FEMS Microbiol. Lett. 173:85–94. 1999.
5. Bencina, D., B. Slavec, and M. Narat. Antibody reponse to GroELvaries in patients with acute Mycoplasma pneumoniae infection. FEMSImmunol. Med. Microbiol. 43:399–406. 2005.
6. Bencina, D., M. Drobnic-Valic, S. Horvat, M. Narat, S. H. Kleven,and P. Dovc. Molecular basis of the length variation in the N-terminal partof Mycoplasma synoviae hemagglutinin. FEMS Microbiol. Lett. 203:115–123. 2001.
Table 1. Identification of M. gallisepticum proteins EF-Tu, PdhA, and Ldh.
Detectedby MAb N-terminal sequenceA
Observed TheoreticalC
IDD NoteEMW pIB MW pIB
GB8 AKERFDRXK 45 ;5.6 43.1F 5.6F EF-Tu pI isoforms9D4 AIIVKNKIPELLHRV 40 ;6 39.5 5.6 PdhA pI isoforms7G1 MKKIAVIGXGFVGST 36 ;8 35.5 7.7 Ldh pI isoformsAX denotes residuum, which can not be determined.BApproximate pI values for major forms.CTheoretical values predicted from the coding sequence of an appropriate gene.DID-identification of the protein according to complete match with the corresponding coding sequence (24). The corresponding genes are
annotated as tuf B (for Tuf B i.e., EF-Tu), acoA (for AcoA i.e., PdhA) and mdh (for Mdh i.e., Ldh).EIn 2D immunoblots MAb detected also isoforms differing in pI values.FValues for EF-Tu with the N-terminal sequence MAKERF . . ., and not with the sequence MFKKMAKERF . . . annotated for the tuf B of R
strain (24).
512 M. Lavric et al.

7. Blum, H., H. Beie, and H. J. Gross. Improved silver stainingof plant proteins, RNA and DNA in polyacrylamide gels. Electrophoresis8:93–99. 1987.
8. Bradbury, J. M., O. M. S. Abdul-Wahab, C. A. Yavari, J. P.Dupiellet, and J. M. Bove. Mycoplasma imitans sp. nov is related toMycoplasma gallisepticum and found in birds. Int. J. Sytem. Bacteriol. 43:721–728. 1993.
9. Bradley, L. D., D. B. Snyder, and R. A. Van Deusen. Identification ofspecies-specific and interspecies-specific polypeptides of Mycoplasma galli-septicum and Mycoplasma snoviae. Am. J. Vet. Res. 49:511–515. 1988.
10. Dallo, S. H., T. R. Kannan, M. W. Blaylock, and J. B. Baseman.Elongation factor Tu and 1b subunit of pyruvate dehydrogenase complex actas fibronectin binding proteins in Mycoplasma pneumoniae. Mol. Microbiol.46:1041–1051. 2002.
11. Dandekar, T., B. Snel, S. Schmidt, W. Lathe, M. Suyama, M.Huynen, and P. Bork. (2002).Comparative genome analysis of Mollicutes.In: Molecular biology and pathogenicity of mycoplasmas. S. Razin andR. Herrmann, eds. Kluwer Academic/Plenum Publishers, New York.pp. 255–278. 2002.
12. Garcia, M., M. G. Elfaki, and S. H. Kleven. Analysis of the variabilityin expression of Mycoplasma gallisepticum surface antigens. Vet. Microbiol.42:147–158. 1994.
13. Gorg, A., W. Postel, and S. Gunther. The current state of two-dimensional electrophoresis with immobilized pH gradients. Electrophoresis9:531–546. 1988.
14. Jan, G., C. Fontenelle, M. Le Henaff, and H. Wroblewski. Acylationand immunological properties of Mycoplasma gallisepticum membraneproteins. Res. Microbiol. 146:1739–745. 1995.
15. Jan, G., M. Le Henaff, C. Fontenelle, and H. Wroblevski.Biochemical and antigenic characterization of Mycoplasma gallisepticummembrane proteins P52 and P67 (pMGA). Arch. Microbiol. 177:81–90.2001.
16. Johansson, K. E. and B. Pettersson. Taxonomy of Mollicutes. In:Molecular biology and pathogenicity of mycoplasmas. S. Razin and R.Herrmann, eds. Kluwer Academic/Plenum Publishers, New York. pp. 1–29.2002.
17. Jordan, F. T. W. Avian mycoplasmosis. In: Poultry diseases. F.T.W.Jordan and M. Pattison, eds. W.B. Saunders, London. pp. 81–93. 1996.
18. Kamla, V., B. Henrich, and U. Hadding. Species differentiation ofmycoplasmas by EF-Tu specific monoclonal antibodies. J. Immunol. Meth.147:73–81. 1992.
19. Korolev, E. V., A. V. Nikonov, M. S. Brudnaya, E. S. Snigirevskaya,G. V. Sabinin, Y. Y. Komissarchik, P. I. Ivanov, and S. N. Borchenius.Tubular structures of Mycoplasma gallisepticum and their possible participa-tion in cell motility. Microbiology. 140:671–681. 1994.
20. Krause, D., and M. F. Balish. Structure, function and assembly ofthe terminal organelle of Mycoplasma pneumoniae. FEMS Microbiol. Lett.198:1–7. 2001.
21. Ley, D. H., and H. W. Yoder. Mycoplasma gallisepticum infection. In:
Diseases of poultry, 9th ed. B. W. Calnek, H. J. Barnes, C. W. Beard, L. R.
McDougald, and Y. M. Saif, eds. Iowa State University Press, Ames, IA.
pp. 194–207. 1997.22. Markham, P. F., M. F. Duffy, M. D. Glew, and G. F. Browning.
A gene family in Mycoplasma imitans closely related to the pMGA family
of Mycoplasma gallisepticum. Microbiology. 145:2095–2103. 1999.23. Milosevic-Berlic, T., D. Bencina, and P. Dovc. Sequence poly-
morphisms within the pMGA genes and pMGA antigenic variants in
Mycoplasma gallisepticum. FEMS Microbiol. Lett. 184:133–139. 2000.24. Papazisi, L., T. Gorton, G. Kutish, P. F. Markham, G. Browning,
D. K. Nguyen, S. Swartzell, A. Madan, G. Mahairas, and S. J. Geary. The
complete genome sequence of avian pathogen Mycoplasma gallisepticum
strain Rlow. Microbiology. 149:2307–2316. 2003.25. Pollack, J. D. Central carbohydrate pathways: metabolic flexibility
and the extra role of some ‘‘housekeeping’’ enzymes. In: Molecular biology
and pathogenicity of mycoplasmas. S. Razin and R. Herrmann, eds. Kluwer
Academic/Plenum Publishers, New York. pp. 163–199. 2002.26. Regula, J. T., B. Ueberle, G. Boguth, A. Gorg, M. Schnolzer, R.
Herrmann, and R. Frank. Towards a proteome map of Mycoplasma
pneumoniae. Electrophoresis. 21:3765–3780. 2000.27. Rosengarten, R., S. Levisohn, and D. Yogev. A 41-kD variable
surface protein of Mycoplasma gallisepticum has a counterpart in
Mycoplasma imitans and Mycoplasma iowae. FEMS Microbiol. Lett. 132:
115–123. 1995.28. Skamrov, A., E. Feoktistova, M. Goldman, and R. Beabealashvilli.
Mycoplasma gallisepticum rpoA gene cluster. FEMS Microbiol. Lett. 208:
281–285. 2002.29. Wasinger, V. C., J. D. Pollack, and J. Humphery-Smith. The
proteome of Mycoplasma genitalium. Chaps soluble component. Eur. J.
Biochem. 267:1571–1582. 2000.30. Yoder, H. W., Jr. A historical account of the diagnosis and
characterization of strains of Mycoplasma gallisepticum of low virulence.
Avian Dis. 30:510–518. 1986.
ACKNOWLEDGMENTS
This study was supported by grant J4-6115-0481-04 from theMinistry of Education and Science of the Republic of Slovenia. Wethank Drs. J. M. Bradbury, D. C. Krause, B. Henrich, R. Rosengarten,P. F. Markham, and S. H. Kleven for MAbs and cultures of Mycoplasmaspecies. We thank Dr. Igor Krizaj for the amino acid analysis of theM. gallisepticum proteins recognized by MAbs. Technical assistance ofI. Beretic, I. Russ, and B. Slavec is also acknowledged.
Mycoplasma gallisepticum PdhA, Ldh, and EF-Tu 513





























