Embed Size (px)
Citation preview

Mutations Mutations de novode novoIntroduction to genetics
Department of Medical GeneticsMedical University of Warsaw

De novo mutations
- importance
• so far omitted from our course –
to make things simpler
• very rare indeed, BUT ...
• historical necessity
(population size vs allele heterogeneity vs
consequences of homozygosity)
• suprising frequency in diseases with genetic
background (today’s topic)
• causes of mutations?

Mutations vs allele frequency
(mutation-retromutation equilibrium)
p → q
Generation 0: p0=1; q0=0
Generation 1: q1=
Generation n: qn=qn-1+µpn-1; pn=(1-µ)n
Generation 2: q2=
(mutation rate)
with frequency µµµµ
µp0=µ; p1=(1-µ)
q1 p2=(1-µ)2+µp1;
p ← q
qn=qn-1+µpn-1
(retromutation rate)
with frequency νννν
-νqn-1
���� µpn-1 – νqn-1 = 0
since p+q=1 ���� p=(1-q)
equilibrium: qn=qn-1 ���� µpn-1 = νqn-1
q = µ / (µ+ν)
p = ν / (µ+ν)
���� µ(1-q)=vq���� µ-µq=νq ���� µ=νq+µq=q(µ+ν)

For a mutation with a strong phenotypic effect the frequency of a mutated allele
is determined by selectionrather than retromutation
selection coefficient (S) –
denotes a degree of reproductive deficiency
of a given genotype in relation to a genotype
with the highest reproductive rate in a population
S jest dopełnieniem wartości przystosowawczej (W) genotypu do jedności (S = 1 – W).
Selection & allele frequency
(selection in medical genetics)
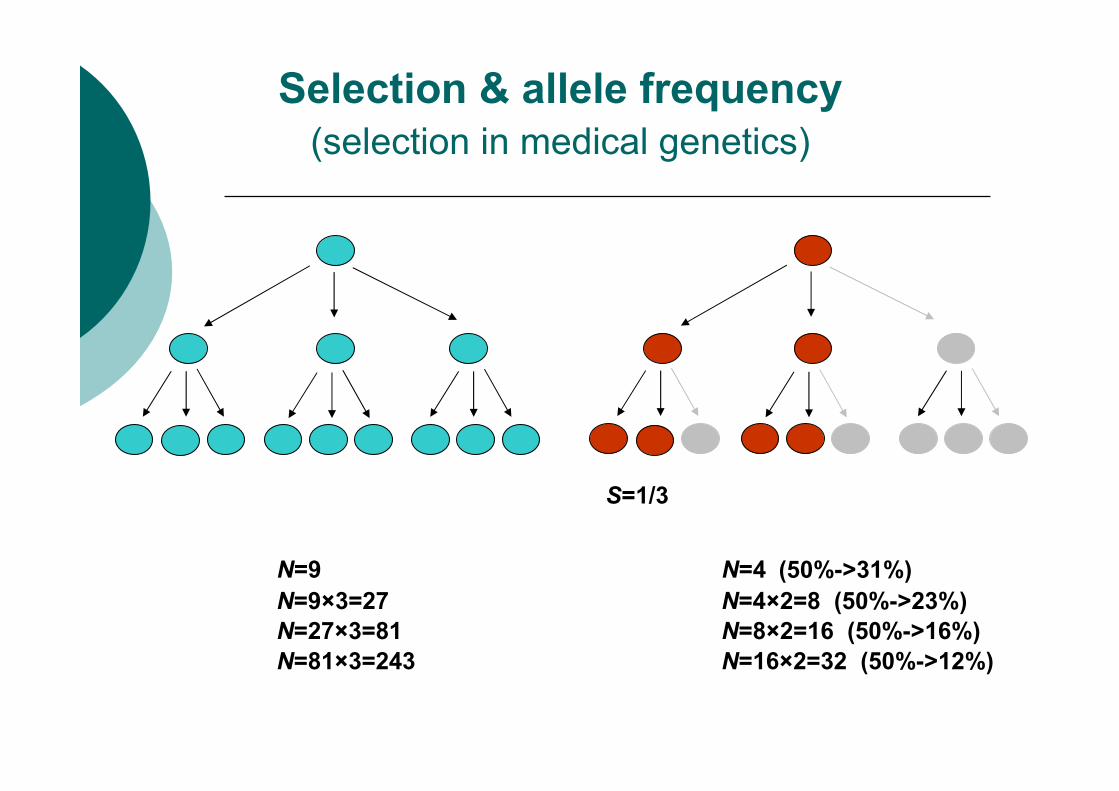
S=1/3
Selection & allele frequency
(selection in medical genetics)
N=9×3=27 N=4×2=8 (50%->23%)
N=27×3=81 N=8×2=16 (50%->16%)
N=81×3=243 N=16×2=32 (50%->12%)
N=9 N=4 (50%->31%)
S=1/3
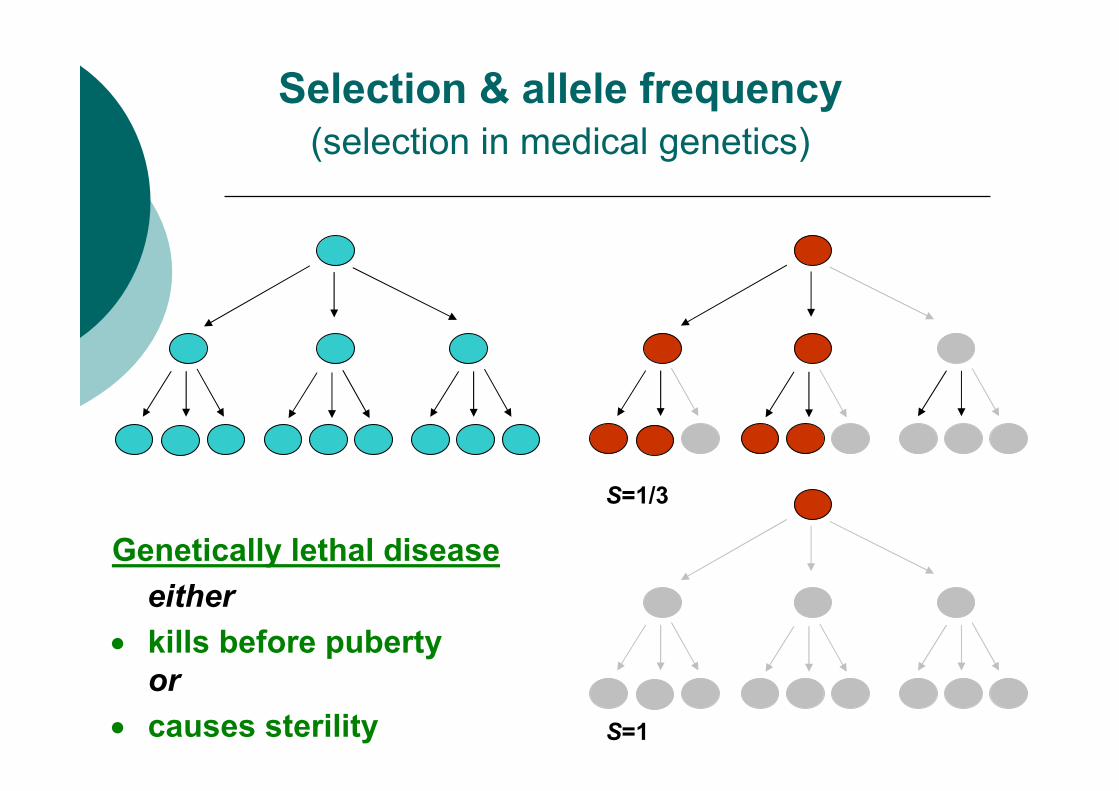
Selection & allele frequency
(selection in medical genetics)
S=1
Genetically lethal disease
either
• kills before puberty
or
• causes sterility

2
Fq 1p
2p
Fq ≈→≅∩=
Autosomal dominant inheritance
Disease frequency F
Mutated allele frequency q
Autosomal recesive inheritance
X-linked recessive inheritance
males: Fm=
Fq =
Hardy & Weinberg
principle
p2+2pq+q2=1
p+q=1
Disease frequency vs
allele frequency
F =
F =
2pq
q2
q q = Fm

All defective alleles
are being lost in each generation
and must be replaced via mutations
All cases of disease are due to new mutations !!! Very low recurrence risk apart from germline mosaicism
de novo mutation
selection
q = µ
p – wild-type allele frequency
q – mutated allele frequency
µ – mutation frequency
Autosomal dominant diseases
(genetically lethal)

Some defective allelesare being lost in each generationand must be replaced via mutations
S×q = µ
p – wild-type allele frequency
q – mutated allele frequency
S – selection coefficient
µ – mutation frequency
Autosomal dominant diseases
(all)
de novo mutation
selection

• population: N
• disease frequency: q2
• No of individuals not passing their alleles: q2×N
(only q are lost)
• No of alleles lost: 2 × q2×N
• No of all alleles: 2 × N
• frequency of alleles being lost: 2 × q2×N / 2 × N = q2
• only q are lost
• lost alleles replaced due to de novo mutations: µ=q2
All homozygotes are sterile,
their alleles are lost in each generation
and must be replaced via mutations
Autosomal recessive diseases
(genetically lethal)
de novo mutation
selection

Practically in autosomal recessive diseases
mutations de novo need not be considered !
Autosomal recessive diseases
(genetically lethal)
example: q=0.01 => µ=0.0001 (µ is very small, ≈0)
q2 = µ
de novo mutation
selection
All homozygotes are sterile,
their alleles are lost in each generation
and must be replaced via mutations
p – wild-type allele frequency
q – mutated allele frequency
µ – mutation frequency
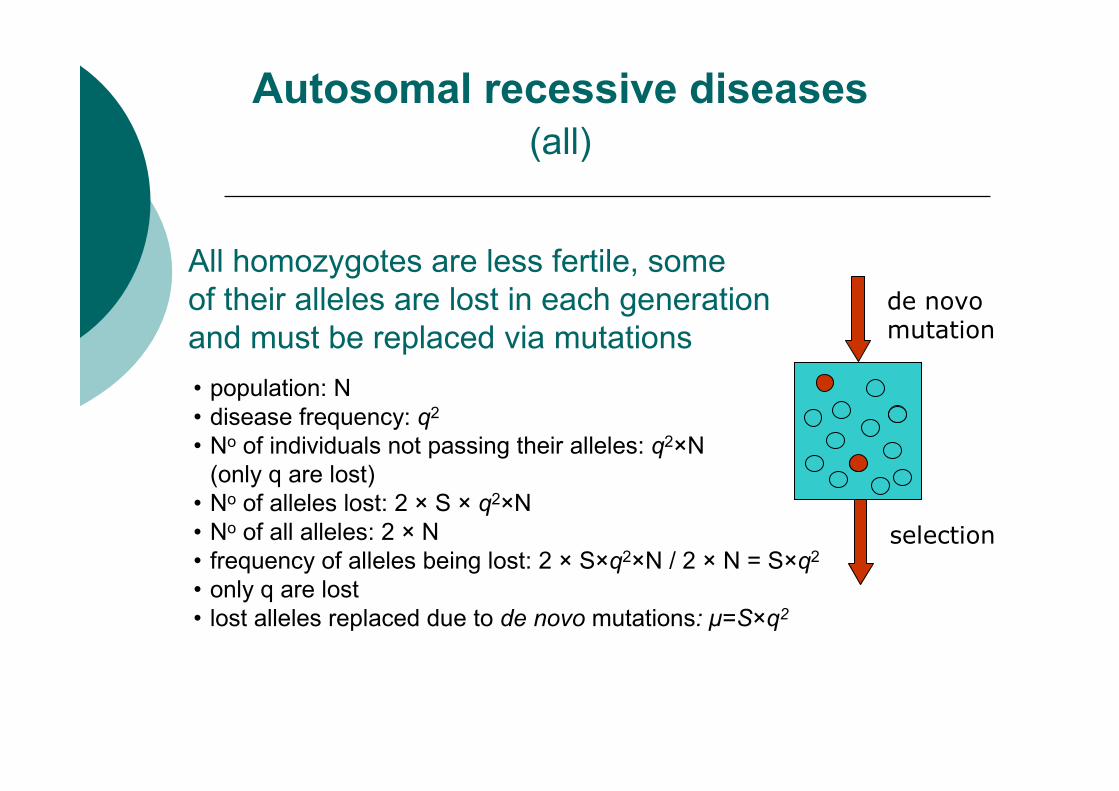
All homozygotes are less fertile, some
of their alleles are lost in each generation
and must be replaced via mutations
Autosomal recessive diseases
(all)
de novo mutation
selection
• population: N
• disease frequency: q2
• No of individuals not passing their alleles: q2×N
(only q are lost)
• No of alleles lost: 2 × S × q2×N
• No of all alleles: 2 × N
• frequency of alleles being lost: 2 × S×q2×N / 2 × N = S×q2
• only q are lost
• lost alleles replaced due to de novo mutations: µ=S×q2

If an autosomal recessive disease is not genetically lethal,
de novo mutations play even lesser role !
p – wild-type allele frequency
q – mutated allele frequency
S – selection coefficient
µ – mutation frequency
Autosomal recessive diseases
(all)
example: q=0.01 & S=0.25 => µ=0.000025=2.5×10-5
µ = S×q2
All homozygotes are less fertile, some
of their alleles are lost in each generation
and must be replaced via mutations
de novo mutation
selection
XY XX
X-linked recessive diseases
(genetically lethal)
If the disease is genetically lethal,
all mutated alleles
carried by hemizygotic males
are lost in each generation
achieving equilibrium is
more complex
de novo mutation
selection

H/2
A H
H/2+µ
A
µ
H=½H+2µ
H-½H=2µ
½H=2µ //×2
H=4µ
H/2+
µ+µH=4µH/2+µA=3µ
A=½H+µA=½(4µ)+µA=2µ+µA=3µ Częstość:
A – chorych mężczyzn
H – kobiet nosicielek
µ – mutacji
X-linked recessive diseases Frequency of mutations (µ), sicked males (A) and female carriers (H)

2µ
3µ 4µ
2µ+µ
3µ
µ
2µ+
µ+µ4µ2µ+µ3µ
Częstość:
A – chorych mężczyzn
H – kobiet nosicielek
µ – mutacji
X-linked recessive diseases Frequency of mutations (µ), sicked males (A) and female carriers (H)

3/1000
3/1000
1/1000
2µ+
µ+µ2µ+µ3/1000
X-linked recessive diseases Frequency of mutations (µ), sicked males (A) and female carriers (H)
N = 1000M+1000K
µ = 1/10004/1000
4/2000(2/1000)
2/1000
+
1/1000
4/1000

X-linked recessive diseases Frequency of mutations (µ), sicked males (A) and female carriers (H)
A=½H+µ
H=½H+µ+µH=4µ
A=3µ
The frequency of female carriers
The frequency of sick males
µ - mutation rate

Sporadic
case 1

Probability of
I2 is
a carrier
I2 isn’t
a carrier
initial 4µ 1
conditional
(sick son)1/2 µ
odds 2 µ µ
final risk 2/3 1/3
Sex-linked recessive disease –mutation de novo (µf=µm) or inheritance? [Bayes theorem]
L1: I2 jest nosicielką
L2: mutacja de novo

LR=L1/L2=2 (2:1):
Every third sporadic case is due to de novo mutation!!!
Sex-linked recessive disease –mutation de novo (µf=µm) or inheritance? [LR]
~4µ
1/2
L1 = 4µ×0,5 = 2µ
L2 = 1×µ = µ
µ
~1
L1: I2 is a carrier
L2: mutation de novo

Results of dystrophin gene mutation in DMD:
a sample of a muscle of the healthy person (upper)
and the affected one (lower) stained with HE (left) or
immunostained with antibody against dystrophin (right)
Calves pseudohypertrophy
muscles are replaced with
fat and connective tissue
in 8-years old boy with
Duchenne muscular dystrophy
Duchenne muscular dystrophy myopathy due to DMD mutations (1/3500 M)
Gowers maneuver: boy with DMD rises from the ground

Duchenne muscular dystrophy
myopathy due to DMD mutations
• A third of mothers who have
a single son affected will not themselves be
carriers of a mutation in the DMD gene !!!
• Thus they have very low risk (µ)
of subsequent problems

α - male/female ratio of mutations)
Is evolution male-driven?
De novo mutation rates of male & female
origin in the diseases studied so far

Nat Rev Gen 2001
Meiosis in females
and in males

H/2
A H
H/2
+µF
A
µM
H=½H+µM+µF
H-½H=µM+µF
½H=µM+µF /×2
H=2µM+2µF
H/2+
µF+µM
H=
2µF+
2µM
H/2+µF
A=
2µF+µM
A=½H+µF
A=½(µM+µF)+µF
A=2µF+µM
Częstość:
A – chorych mężczyzn
H – kobiet nosicielek
µ – mutacji
X-linked recessive diseases Frequency of mutations (µ), sicked males (A) and female carriers (H)
when µF≠µM

2µf +µmµf +µm
µm
2µf +µm
µf +µm
+µf +µm
2µf +µm2(µf +µm)
µf +µm+µf
=2µf +µm
X linked recessive inheritance: relationship between µµµµ,
H, and A (cntd.), µµµµ differes between sexes (µf =µm) cntd.
gametes
chromosomes
people
people
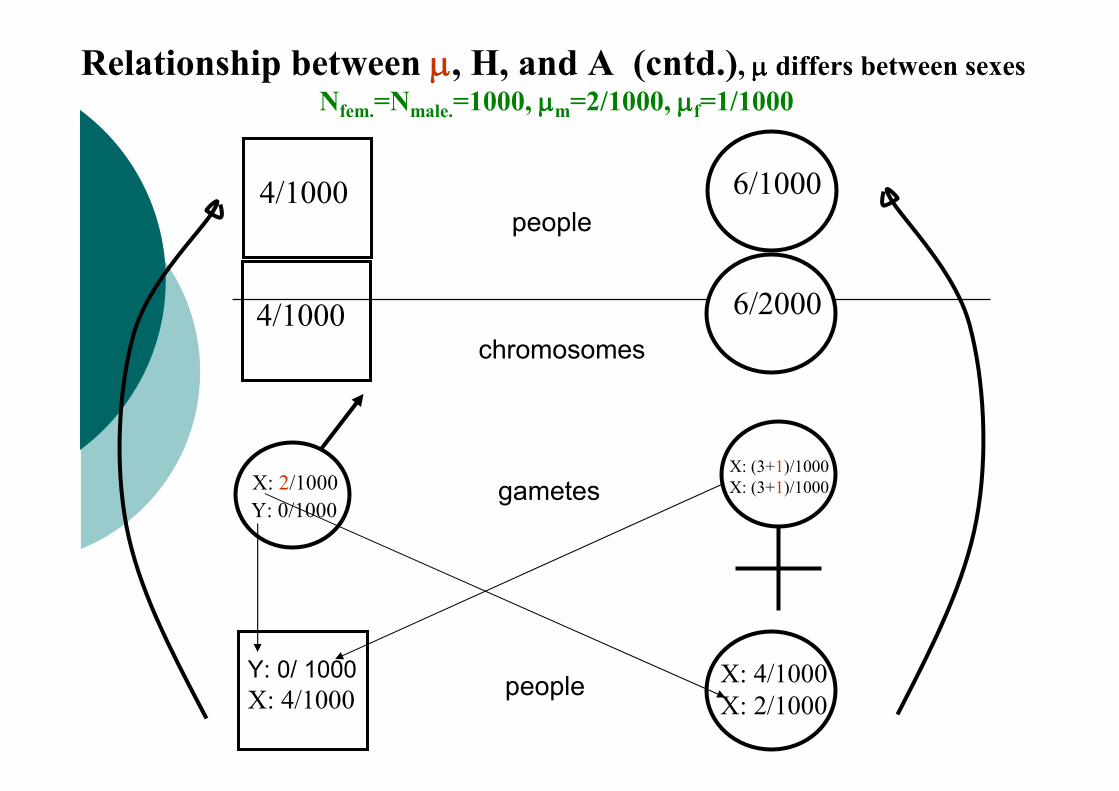
4/1000 6/2000
X: 2/1000
Y: 0/ 1000
X: 4/1000
4/1000 6/1000
Relationship between µµµµ, H, and A (cntd.), µµµµ differs between sexes
Nfem.=Nmale.=1000, µµµµm=2/1000, µµµµf=1/1000
Y: 0/1000
X: 4/1000
X: 2/1000
X: (3+1)/1000
X: (3+1)/1000gametes
chromosomes
people
people

Sporadic
case 2

LR= (µf +µm) / µf.
X-linked recessive disease –
mutation de novo (µf≠µm) or inherited?
~2×(µf+µm)
1/2
L1= µf+µm
L2=1×µf=µf
µf
~1
L1: I2 is a carrier
L2: de novo mutation
if µf=0.2µm, then LR=1.2/0.2=6, tzn.:
Only 1/7 of sporadic cases are due to de novo mutations

P m
oth
er
as a
carr
ier
µµµµm
/µµµµf
0,67
0,72
0,77
0,82
0,87
0,92
2 3 4 5 6 7 8 9 101
X-linked recessive disease –
mutation de novo (µf≠µm) or inherited?

� Hemophilia A and hemophilia B are X-linked disorders of coagulation caused by mutations in the F8 and F9 genes, respectively. Mutations of F8 cause deficiency or dysfunction of clotting factor VIII; mutations of F9 cause deficiency or dysfunction of clotting factor IX.
� Hemophilia A has an incidence of 1 in 5000 to 10,000 newborn males.
� Hemophilia B is far rarer, with an incidence of 1 in 100,000.
Subcutaneous hematoma of the forehead
in a young heophiliac boy, days after a minor contusion.
The appearance of the forehead returned to normal in 6 months
Hemophilia B Factor IX α ~4 Sommer 2001.
Hemophilia A Factor VIII α ~3 Rosendaal 1990.
Hemophilia

� De novo mutations arise with comparable frequency during oogenesis and spermatogenesis
� The dystrophin gene is a very large (>2mB) and contains many repetitive regions. Furthermore, a greater than 30-fold(!) reduction in dystrophin activity must occur before the DMD phenotype is observed Sommer 2001.
� High frequency of large deletions (60% to 65%), large duplications (5% to 10%), and small deletions, insertions, or nucleotide changes (25% to 30%).
� Most de novo large deletions arise during oogenesis, whereas most de novo nucleotide changes arise during spermatogenesis
Duchenne muscular dystrophy
an exception to the rule?
![Identification of De Novo JAK2 and MAPK7 Mutations Related ... · (Ronemus et al. 2014) and TADA [Transmission And De novo Association] (Li et al. 2016), which prioritize the list](https://img.pdfslide.us/doc/110x75/5fcc9e0fbd3c59693e1845bb/identification-of-de-novo-jak2-and-mapk7-mutations-related-ronemus-et-al-2014.jpg)




























