Embed Size (px)
Citation preview

MURDOCH RESEARCH REPOSITORY
This is the author’s final version of the work, as accepted for publication following peer review but without the publisher’s layout or pagination.
The definitive version is available at http://dx.doi.org/10.1016/j.neuroscience.2014.09.046
Drummond, E.S., Dawson, L.F., Finch, P.M., Li, W., Guo, T-Z, Kingery, W.S. and Drummond, P.D. (2014) Increased bilateral expression of α1-adrenoceptors on peripheral nerves, blood
vessels and keratinocytes does not account for pain or neuroinflammatory changes after distal tibia fracture in rats.
Neuroscience, 281 . pp. 99-109.
http://researchrepository.murdoch.edu.au/24332/
Copyright: © 2014 Elsevier BV.
It is posted here for your personal use. No further distribution is permitted.

1
2
3
4
5
6 Q1
7
8
9
10 Q2
11
12
13
14
Q3
Please cite this article in press as: Drummond ES et al. Increased bilateral expression of a1-adrenoceptors on peripheral nerves, blood vessels and keratinocytes does
not account for pain or neuroinflammatory changes after distal tibia fracture in rats. Neuroscience (2014), http://dx.doi.org/10.1016/j.neuroscience.2014.09.046
NSC 15719 No. of Pages 11
3 October 2014
Neuroscience xxx (2014) xxx–xxx
INCREASED BILATERAL EXPRESSION OF a1-ADRENOCEPTORSON PERIPHERAL NERVES, BLOOD VESSELS AND KERATINOCYTESDOES NOT ACCOUNT FOR PAIN OR NEUROINFLAMMATORYCHANGES AFTER DISTAL TIBIA FRACTURE IN RATS
E. S. DRUMMOND, a L. F. DAWSON, a P. M. FINCH, a
W. LI, b,c T.-Z. GUO, b W. S. KINGERY b ANDP. D. DRUMMOND a*
aCentre for Research on Chronic Pain and Inflammatory
Diseases, Murdoch University, Perth, Western Australia, AustraliabPhysical Medicine and Rehabilitation Services, VAPAHCS,
Palo Alto, CA, USA
cDepartment of Anesthesia, Stanford University, Stanford, CA, USA
15
16
17
18
19
20
21
22
23
24
25
26
27
28
29
30
31
32
33
34
35
36
37
38
39
40
41
Abstract—In certain forms of nerve injury and inflammation,
noradrenaline augments pain via actions on up-regulated
a1-adrenoceptors (a1-ARs). The aim of this study was to
use immunohistochemistry to examine a1-AR expression
on peripheral neurons, cutaneous blood vessels and kerati-
nocytes after distal tibia fracture and cast immobilization, a
model of complex regional pain syndrome type 1. We
hypothesized that there would be increased a1-AR expres-
sion on neurons and keratinocytes in the injured limb in
comparison to the contralateral unaffected limb after distal
tibia fracture, in association with inflammatory changes
and pain. a1-AR expression was increased on plantar kerat-
inocytes, dermal blood vessels and peripheral nerve fibers
at 16 weeks after injury both in the fractured and contralat-
eral uninjured limb. Similar changes were seen in controls
whose limb had been immobilized in a cast for 4 weeks but
not fractured. Neurofilament 200 (NF200), a marker of mye-
linated neurons, and calcitonin gene-related peptide
(CGRP), a neuropeptide involved in neuro-inflammatory sig-
naling, decreased 4 weeks after fracture and casting but
then increased at the 16-week time point. As some of these
changes were also detected in the contralateral hind limb,
they probably were triggered by a systemic response to frac-
ture and casting. Soon after the cast was removed, intrapl-
antar injections of the a1-AR antagonist prazosin released
local vasoconstrictor tone but had no effect on pain behav-
iors. However, systemic injection of prazosin inhibited
behavioral signs of pain, suggesting that fracture and/or
casting triggered an up-regulation of a1-ARs in central noci-
ceptive pathways that augmented pain. Together, these find-
ings indicate that a1-AR expression increases in the hind
http://dx.doi.org/10.1016/j.neuroscience.2014.09.0460306-4522/� 2014 Published by Elsevier Ltd. on behalf of IBRO.
*Corresponding author. Address: School of Psychology and ExerciseScience, Murdoch University, 6150 Western Australia, Australia. Tel:+61-8-93602415; fax: +61-8-93606492.
E-mail address: [email protected] (P. D. Drummond).Abbreviations: a1-AR, a1-adrenoceptor; CGRP, calcitonin gene-relatedpeptide; CRPS, complex regional pain syndrome; IL, interleukin;NF200, neurofilament 200; PBS, phosphate-buffered saline; PGP9.5,protein gene product 9.5; SMA, smooth muscle actin; TNF, tumornecrosis factor-a.
1
limbs after distal tibia fracture and cast immobilisation.
However, these peripheral increases do not contribute
directly to residual pain. � 2014 Published by Elsevier Ltd.
on behalf of IBRO.
Key words: distal tibia fracture, alpha-1 adrenoceptor,
immunohistochemistry, calcitonin gene-related peptide,
neurofilament 200, sympathetically maintained pain.
42
43
44
45
46
47
48
49
50
INTRODUCTION
Distal limb fracture is the most common trigger of complex
regional pain syndrome (CRPS) type 1 (de Mos et al.,
2007), a condition associated with acute neuro-
inflammatory changes and chronic sensory, motor and
autonomic disturbances (Marinus et al., 2011). Investiga-
tion of the neuro-inflammatory changes in an animal model
of distal tibia fracture/cast immobilisation has shown that
pain after fracture is associatedwith signaling of neuropep-
tides such as substance P and calcitonin gene-related
peptide (CGRP), and key inflammatory mediators such
as tumor necrosis factor-a (TNF), interleukin-1b (IL-1b),IL-6 and nerve growth factor (Guo et al., 2004;
Sabsovich et al., 2008a,b; Li et al., 2009, 2010, 2012;
Wei et al., 2009a,b). Many of these neuro-inflammatory
changes after fracture are reduced in transgenic mice defi-
cient in CGRP receptors or substance P, and are inhibited
by neurokinin-1 receptor blockade (Wei et al., 2009b; Guo
et al., 2012). Thus, distal limb fracture appears to trigger a
neuro-inflammatory cascade that might contribute to the
development of CRPS.
We recently reported that noradrenaline, released
from sympathetic vasomotor neurons or produced
locally in the epidermis, may augment inflammation after
distal tibia fracture by activating keratinocytes or
vascular endothelial cells (Li et al., 2013). In particular,
intradermal injection of the b2-adrenoceptor agonist ter-
butaline provoked mechanical allodynia and triggered
the production of IL-6 in the skin. Conversely, chemical
sympathectomy with guanethidine and selective blockade
of b2-adrenoceptors inhibited edema, vascular distur-
bances and behavioral signs of pain. Together, these
findings suggest that sympathetic efferent signaling
contributes to nociceptive sensitization after distal limb
fracture by acting on b2-adrenoceptors.

51
52
5354
55
56
57
58
59
60
61
62
63
64
65
66
67
68
69
70
71
72
73
74
75
76
7778
79
80
81
82
8384
85
86
87
88
89
90
91
92
93
94
95
96
97
98
99
100
101
102
103
104
105
106
107
108
109
110
111Q4
112
113
114
115
116
117
118
119
120
121
122
123
124
125
126
127
128
129
130
131
132
133
134
135
136
137
138
139
140
141
142
2 E. S. Drummond et al. / Neuroscience xxx (2014) xxx–xxx
NSC 15719 No. of Pages 11
3 October 2014
In certain forms of nerve injury and inflammation,
noradrenaline may also augment pain via actions on
a1-adrenoceptors (a1-AR) (Hong and Abbott, 1996; Ali
et al., 1999; Nam et al., 2000; Hord et al., 2001; Xie
et al., 2001; Kim et al., 2005; Dogrul et al., 2006; Maruo
et al., 2006; Kopp et al., 2007; Meisner et al., 2007;
Trevisani et al., 2007; Gibbs et al., 2008; Dawson et al.,
2011; Donello et al., 2011). Notably, we recently reported
that a1-AR expression increased in dermal nerve bundles
4–28 days after partial sciatic nerve lesion (Drummond
et al., 2014b), and on nociceptive nerve fibers and kerati-
nocytes 7 days after chronic constriction injury of the sci-
atic nerve (Drummond et al., 2014a). This is consistent
with an aberrant involvement of these receptors in CRPS
(Drummond et al., 1996, 2014b; Finch et al., 2014), neu-
roma pain (Chabal et al., 1992; Lin et al., 2006), post-her-
petic neuralgia (Choi and Rowbotham, 1997; Schmidt
et al., 2003) and other neuropathic pain conditions (Lee
et al., 2000). Thus, the first aim of this study was to look
for signs of nerve injury and to examine a1-AR expression
on peripheral neurons, cutaneous blood vessels and kerat-
inocytes after distal tibia fracture and cast immobilization.
We hypothesized that a1-ARs would be expressed more
strongly on neurons and keratinocytes in the injured than
contralateral limb, in association with signs of inflammation
and pain (Li et al., 2013). Likewise, we hypothesized that
a1-AR up-regulation would be greater after distal tibia
fracture and cast immobilization than in a cast-only group.
Our second aim was to determine whether
sympathetic signaling in the skin might contribute to
pain after distal tibia fracture by activating a1-ARs on
neurons or keratinocytes. As local administration of
a1-AR agonists intensifies pain behaviors associated
with acute inflammation (Hong and Abbott, 1996), we
hypothesized that injection of the a1-AR antagonist prazo-
sin into the hind paw skin of the injured limb would allevi-
ate pain associated with distal tibia fracture.
143144
145
146
147
148
149
150
151
152
153
154
155
156
157
158
159
160
EXPERIMENTAL PROCEDURES
Animals
These experiments were approved by the Veterans
Affairs Palo Alto Health Care System Institutional Animal
Care and Use Committee (Palo Alto, CA, USA) and the
Murdoch University Animal Research Ethics Committee,
and followed the guidelines of the International
Association for the Study of Pain. Adult (9-month-old)
male Sprague–Dawley rats (Simonsen Laboratories,
Gilroy, CA, USA) were housed individually in isolator
cages with solid floors covered with 3 cm of soft
bedding and were given food and water ad libitum.
During the experimental period the animals were fed
Lab Diet 5012 (PMI Nutrition Institute, Richmond, IN,
USA), which contains 1.0% calcium, 0.5% phosphorus,
and 3.3 IU/g vitamin D3, and were kept under standard
conditions with a 12-h light–dark cycle.
161
162
163
164
165
Surgery
Tibia fracture was performed under 2–4% isoflurane to
maintain surgical anesthesia (Guo et al., 2004). The right
Please cite this article in press as: Drummond ES et al. Increased bilateral expression o
not account for pain or neuroinflammatory changes after distal tibia fracture in rats. Ne
hind limb was wrapped in a stockinet (2.5 cm wide) and
the distal tibia was fractured using pliers with an adjust-
able stop (Visegrip, Petersen Manufacturing, Dewitt, NE,
USA) that had been modified with a three-point jaw. The
hind limb was wrapped in casting tape ([delta]-Lite,
Johnson & Johnson, New Brunswick, NJ, USA) so that
the hip, knee, and ankle were flexed. The cast extended
from the metatarsals of the hind paw up to a spica formed
around the abdomen. The cast over the paw was
applied only to the plantar surface; a window was left
open over the dorsum of the paw and ankle to minimize
constriction when post-fracture edema developed. To pre-
vent the animals from chewing at their casts, the cast
material was wrapped in a galvanized wire mesh. The rats
were given subcutaneous saline and buprenorphine
immediately after the procedure (0.03 mg/kg) and on
the first day after fracture for postoperative hydration
and analgesia. At 4 weeks the rats were anesthetized with
isoflurane and the cast was removed with a vibrating cast
saw. All rats used in this study had union at the fracture
site after 4 weeks of cast immobilization.
As only one limb was fractured, values recorded from
the contralateral limb were used as an internal control. A
cast-only group served as an additional external control
as the limb was immobilized but not fractured.
Behavioral and physiological measures
Behavioral measures were obtained the day after the cast
was removed at 4 weeks, and again at 16 weeks in a
subgroup of animals. To measure mechanical allodynia,
an up-down von Frey testing paradigm was used as
previously described. In addition, an incapacitance
device (IITC Inc. Life Science, Woodland Hills, CA, USA)
was used to measure hind paw unweighting. The rats
were manually held in a vertical position over the
apparatus with the hind paws resting on separate metal
scale plates, and the entire weight of the rat was
supported on the hind paws. The duration of each
measurement was 6 s, and 10 consecutive
measurements were taken at 60-s intervals. Eight
readings (excluding the highest and lowest) were
averaged to calculate the hind paw weight-bearing
values (Kingery et al., 2003; Guo et al., 2004, 2006).
Room temperature was maintained at 23 �C and
humidity ranged between 25% and 45%. The
temperature of the hind paw was measured using a fine
wire thermocouple (Omega, Stamford, CT, USA) applied
to the paw skin, as previously described (Kingery et al.,
2003; Guo et al., 2004, 2006). In addition, a laser sensor
technique was used to determine the dorsal-ventral thick-
ness of the hind paw, as previously described (Kingery
et al., 2003; Guo et al., 2004, 2006).
Administration of a1-AR agonists and antagonists4–6 weeks after distal tibia fracture
All drugs were dissolved in normal saline. Baseline
behavioral testing was performed prior to drug injection
and 1 h after intraperitoneal injection of the a1-AR
agonists phenylephrine (3 mg/kg) and cirazoline (1 mg/
kg), or the a1-AR antagonist prazosin (2 mg/kg). This
f a1-adrenoceptors on peripheral nerves, blood vessels and keratinocytes does
uroscience (2014), http://dx.doi.org/10.1016/j.neuroscience.2014.09.046

166
167
168
169
170
171
172
173
174
175
176
177
178
179
180
181
182
183
184
185
186
187
188
189
190
191
192
193
194
195 Q5
196
197
198
199
200
201
202
203
204
205
206
207
208
209210
211
212
213
214
215
216
217
218
219
220
221
222
223
224
225
226227
228
229
230
231
232
233
234
235
236
237
238
239
240
241
242
243
244
245
246
247
248
249
250
251
E. S. Drummond et al. / Neuroscience xxx (2014) xxx–xxx 3
NSC 15719 No. of Pages 11
3 October 2014
time point was chosen based on our previous experience
with intraperitoneal injection of these drugs (Guo et al.,
1991; Kingery et al., 2002). In addition, von Frey allodynia
and skin temperature were measured bilaterally before
and after unilateral injection of prazosin (20 lg in 25 llsaline) into the plantar hind paw. Prazosin was injected
into the right paw (ipsilateral to the cast and fracture) on
day one and into the left paw 3 days later. Von Frey allo-
dynia was investigated in both hind paws at 0.5, 1, 2 and
3 h after each injection, and skin temperature was mea-
sured at 0.5, 1, 2, 3, 4, 6 and 24 h. These time points were
selected based on preliminary tests.
Tissue processing and immunohistochemistry
Animals were euthanized and immediately perfused with
Zamboni’s fixative (FD NeuroTechnologies, Inc., Ellicott,
MD, USA), via the ascending aorta; the hind paw dorsal
and plantar skin including sub-dermal layers, and
approximately 10 mm of both sciatic nerves and tibia
nerves were collected from intact controls (n= 5), week
4 (n= 5) and week 16 (n= 5) cast only rats, and week
4 (n= 8) and week 16 (n= 7) post-fracture rats.
Tissue samples were post-fixed in Zamboni’s fixative for
4 h at 4 �C, then rinsed with 0.1 M phosphate buffer (pH
7.4) and 50% ethanol followed by embedding in
TissurePrep2 paraffin (Fisher Scientific, Fair Lawn, NJ,
USA). Some results were not available in the hind paw
skin for the 4-week cast-only group because of technical
difficulties with the fixation of these samples.
Ten-micrometer sections were collected onto silane-
coated slides (Hurst Scientific). Sections were
deparaffinized through xylene and a descending series
of ethanol washes to 0.1 M phosphate- buffered saline
(PBS). Antigen retrieval was performed by incubation
with 1 mg/ml porcine trypsin (Sigma–Aldrich) for 30 min
at 37 �C. Sections were then treated with 0.5% Triton
X-100 for 5 min and incubated with blocking solution for
2 h (10% normal donkey serum in 0.1 M PBS). The
following combinations of primary antibodies were then
applied: a1-AR/protein gene product 9.5 (PGP9.5) to
examine a1-AR expression on all nerve fibers; a1-AR/
CGRP to examine a1-AR expression on peptidergic
nerve fibers; a1-AR/ neurofilament 200 (NF200) to
examine a1-AR expression on myelinated fibers; and
Table 1. Primary and secondary antibodies used in rat tissues: dilutions and s
Antigen and host species Dilution – skin D
Primary antibodies
Anti a1-AR, rabbit polyclonal 1:200 1
Anti CGRP, goat polyclonal 1:400 1
Anti NF200, chicken polyclonal 1:3000 1
Anti SMA, mouse monoclonal 1:1000 1
Anti PGP9.5, mouse monoclonal 1:500 1
Secondary antibodies
Donkey anti-rabbit Dylight 488 1:600 1
Donkey anti-goat Cy3 1:750 n
Donkey anti-goat Dylight 549 n/a 1
Donkey anti-chicken Dylight 647 1:1000 1
Donkey anti-mouse Dylight 647 1:1000 1
Abbreviations: a1-AR, a1-adrenoceptor; CGRP, calcitonin gene related peptide; SMA, sm
Please cite this article in press as: Drummond ES et al. Increased bilateral expression o
not account for pain or neuroinflammatory changes after distal tibia fracture in rats. Ne
a1-AR/smooth muscle actin (SMA) to examine a1-AR
expression on blood vessels (details and dilutions are
shown in Table 1). Antibodies were co-incubated for
48 h at 4 �C, diluted in blocking solution. Sections were
then washed with PBS (3 � 15 min) and incubated with
the appropriate species- specific secondary antibodies
(Table 1) diluted in 5% donkey serum (Sigma) in PBS
for 4 h at room temperature. Sections were washed with
PBS (3 � 15 min) and coverslipped with Prolong Gold
anti-fade mounting media (Invitrogen). The pattern of
staining produced by the a1-AR antibody on blood
vessels, nerves and epidermal cells in rat skin tissue
resembles the staining pattern produced by BODIPY
FL-prazosin, a fluorescent a1-AR antagonist (Dawson
et al., 2011). In addition, staining was eliminated following
pre-adsorption of the anti-sera with an a1-AR-specific
peptide, indicating that the antibody binds specifically to
a1-ARs (Dawson et al., 2011). In this study, no staining
was observed on negative control sections that had all
primary antibodies omitted.
Confocal imaging and quantification ofimmunohistochemistry
Images of immunostained sections were collected using a
Nikon Eclipse Ti multiphoton confocal microscope. All
images were collected at 200� magnification and
consisted of a stack of optical sections (Z-series)
acquired at 2-lm intervals throughout the depth of the
tissue section. Each label was collected sequentially at
the appropriate excitation and emission spectra to
prevent bleed-through between channels, and imaging
settings were consistent for all sections in each staining
run. For sciatic and tibial nerve sections, two images
were collected per sample for each staining combination.
For skin sections, two images containing the epidermis
and underlying papillary dermis and two images of the
deeper dermis containing large nerve bundles were
collected per sample for each staining combination.
Quantification of immunohistochemistry was performed
using ImageJ software (ImageJ, version 1.47t, National
Institute of Health, USA) on the maximum projection
image of each z-stack. The investigator who performed
the quantification was blinded to the experimental group
and to the laterality of each slide.
ource
ilution – sciatic/tibial nerve Product code and source
:200 A270, Sigma–Aldrich
:400 1720-9007, AbD Serotec
:3000 Jackson ImmunoResearch
:1000 A2547, Sigma–Aldrich
:500 MCA2084, AbD Serotec
:600 Jackson ImmunoResearch
/a Jackson ImmunoResearch
:1200 Jackson ImmunoResearch
:1000 Jackson ImmunoResearch
:1000 Jackson ImmunoResearch
ooth muscle actin; NF200, neurofilament 200; PGP9.5, protein gene product 9.5.
f a1-adrenoceptors on peripheral nerves, blood vessels and keratinocytes does
uroscience (2014), http://dx.doi.org/10.1016/j.neuroscience.2014.09.046

252
253
254
255
256
257
258
259
260
261
262
263
264
265
266
267
268
269
270
271
272
273
274
275
276
277
278
279
280
281
282
283
284
285
286
287
288
289
290
291
292
293
294
295
296
297
298
299
300
301
302
303
304
305
306
307
308
4 E. S. Drummond et al. / Neuroscience xxx (2014) xxx–xxx
NSC 15719 No. of Pages 11
3 October 2014
To examine cutaneous CGRP+ nerve fiber
populations, a standardized threshold level that
identified CGRP+ pixels was applied to all images of
dermal nerve bundles. The number of CGRP+ pixels in
each nerve bundle above this threshold was expressed
as a percentage of the total nerve bundle area to allow
for comparison between nerve bundles of different
sizes. This same method was also used to determine
the expression of NF200+ fibers in nerve bundles. The
expression of CGRP and NF200 in sciatic and tibial
nerve fibers was examined using a similar method in a
standard rectangular area that was applied to all images.
a1-AR expression was quantified in the sciatic nerve,
tibial nerve and epidermis, and in dermal nerve fibers
and blood vessels. The average a1-AR staining intensity
in sciatic and tibial nerves was quantified in a standard
rectangular area that was consistent across all images.
This area was representative of the whole nerve with
regard to a1-AR staining intensity and only contained
nerve fibers. In skin sections, the epidermis was
identified by morphology and the average a1-AR
staining intensity in this region was quantified. To
identify a1-AR expression on nerve fibers, PGP9.5
staining was used to create a mask of all nerve fibers,
allowing a1-AR staining intensity to be measured only
on these nerve fibers. Similarly, SMA staining was used
to identify dermal blood vessels and a1-AR staining
intensity was measured on these identified regions.
The average a1-AR staining intensity was also
quantified in CGRP+ and NF200+ fiber populations.
Fig. 1. Behavioral responses in week 4 (n= 5) and week 16 (n= 5) cast onl
the behavioral measures differed between the ipsilateral and contralateral hin
In A–D, #represents a significant difference between the ipsilateral and contra
the ipsilateral hind paw denote significant differences between groups tha
differences were detected among the four groups for the contralateral hind p
Please cite this article in press as: Drummond ES et al. Increased bilateral expression o
not account for pain or neuroinflammatory changes after distal tibia fracture in rats. Ne
CGRP or NF200 staining was used to create a mask of
nerve fiber location, which was then applied to the
corresponding a1-AR image and the average a1-AR
staining intensity in this defined region was measured.
The average a1-AR staining intensity in CGRP+ and
NF200+ fibers was also determined in the sciatic and
tibial nerves using the same method.
Statistical approach
Differences between the fracture and cast-only groups in
hind paw allodynia, unweighting, temperature and
swelling were investigated for the ipsilateral versus the
contralateral hind paw in repeated measures analyses
of variance. A similar approach was used to investigate
differences in PGP, CGRP, NF200 and a1-AR
expression across the experimental and control groups.
Significant main effects and interactions were explored
across groups in a one-way analyses of variance and
post hoc Newman–Keuls tests, and within each group
with paired t-tests. Results are reported as the
mean ± standard error, and the criterion of statistical
significance was p< 0.05.
RESULTS
Behavioral and physiological changes after distaltibia fracture
At 4 weeks, allodynia and unweighting were greater in the
ipsilateral than contralateral hind paw in the fracture and
cast-only groups (p< 0.001) (Figs. 1A, B). These
y, and week 4 (n= 8) and week 16 (n= 7) post-fracture rats. Each of
d paws across the four groups (Group � Side interactions, p< 0.01).
lateral hind paw (p< 0.05). The letters a, b and c above the bars for
t do not share the same letter (p< 0.05, Newman–Keuls test). No
aw (Newman–Keuls test). Error bars represent standard errors.
f a1-adrenoceptors on peripheral nerves, blood vessels and keratinocytes does
uroscience (2014), http://dx.doi.org/10.1016/j.neuroscience.2014.09.046

309
310
311
312
313
314
315
316
317
318
319
320
321
322
323
324
325
326
327
328
329
330
331
332
333
334
335
336
337
338
339
340
341
342
343
344
345
346
347
348Q6
349
350
351
352
353
354
355
356
357
358
359
360
361
E. S. Drummond et al. / Neuroscience xxx (2014) xxx–xxx 5
NSC 15719 No. of Pages 11
3 October 2014
behavioral signs of pain persisted at 16 weeks in the
fracture group but had resolved in the cast-only group.
The temperature of the ipsilateral hind paw was
elevated in both groups at 4 weeks (p< 0.001)
(Fig. 1C), but this response had disappeared by
16 weeks. In addition, the dorsal-ventral thickness was
greater in the ipsilateral than contralateral hind paw of
the fracture group at 4 weeks (p< 0.001), but the
swelling had gone by 16 weeks (Fig. 1D).
Four weeks after fracture, intraperitoneal injection of
prazosin inhibited allodynia and unweighting in the
ipsilateral hind paw (Fig. 2). In addition, temperature
increased in both hind paws after the prazosin injection;
this increase was greater in the contralateral than
ipsilateral hind paw possibly because temperature was
already elevated in the ipsilateral hind paw at the
baseline (Fig. 2). Allodynia did not change in either hind
paw after intraplantar injections of prazosin; however,
increases in temperature were greater when prazosin
was injected into the ipsilateral than contralateral
hind paw (Fig. 3). Allodynia did not increase after
intraperitoneal injection of phenylephrine or cirazoline,
perhaps due to ceiling effects as allodynia was already
pronounced before the injections (data not shown).
362
363
364
365
366
367
368
369
370
371
Changes in PGP, CGRP and NF200 expression afterdistal tibia fracture
The percentage of PGP+ pixels was not affected either by
fracture or casting, either in the ipsilateral or contralateral
sciatic nerve. However, the percentage of CGRP+ pixels
in sciatic nerves was greater in the 16-week fracture
group than in the other four groups (p< 0.05) (Fig. 4).
CGRP staining also differed among groups in the
Fig. 2. Effect of intraperitoneal injection of prazosin (2 mg/kg) on behavior
allodynia and unweighting were greater in the ipsilateral than contralateral h
swollen than the contralateral hind paw (⁄p< 0.05). Allodynia and unweigh
p< 0.05). In addition, increases in skin temperature were greater for the cont
thereby offsetting differences at baseline. #represents a significant difference
the prazosin injection (p< 0.05). Error bars represent standard errors.
Please cite this article in press as: Drummond ES et al. Increased bilateral expression o
not account for pain or neuroinflammatory changes after distal tibia fracture in rats. Ne
plantar dermal nerves, primarily in the contralateral hind
paw. Increases were most pronounced in the 16-week
cast group; in addition, the percentage of CGRP+ pixels
was lower on the ipsilateral than contralateral side in the
4- and 16-week fracture groups (p< 0.05) (Fig. 4).
However, CGRP expression did not after fracture or
cast-immobilisation in the tibial nerve or in dorsal dermal
nerve bundles (data not shown).
The percentage of NF200+ pixels in sciatic nerves
also differed among groups (Fig. 5). Specifically,
NF200+ expression decreased in the 4-week fracture
group, particularly on the contralateral side (p< 0.001
compared with the ipsilateral side), but then increased in
the 16-week fracture group such that levels were
greater on the ipsilateral than contralateral side
(p< 0.05). Similar trends in the tibial and dermal nerves
did not achieve statistical significance.
Changes in a1-AR expression after distal tibiafracture
a1-AR expression increased significantly in the sciatic
nerve in all groups (Fig. 6). In the 16-week cast and
fracture groups, a1-AR expression also increased
significantly in plantar dermal blood vessels and
keratinocytes in both hind paws (Figs. 7 and 8).
a1-AR expression was also examined in individual
nerve populations (Fig. 9). a1-AR staining intensity on
CGRP+ and NF200+ axons trended upward in the hind
limbs of the 16-week cast and fracture groups. For
CGRP+ fibers, a1-AR expression differed among
groups for the tibial (p< 0.05) and plantar dermal
nerves (p< 0.05). For NF200+ fibers, a1-AR
al responses 4 weeks after distal tibia fracture (n= 7). At baseline,
ind paw; in addition, the ipsilateral hind paw was warmer and more
ting decreased after the prazosin injection (Time � Side interactions,
ralateral than ipsilateral hind paw (Time � Side interaction, p< 0.05),
between levels at baseline and levels in the same hind paw 1 h after
f a1-adrenoceptors on peripheral nerves, blood vessels and keratinocytes does
uroscience (2014), http://dx.doi.org/10.1016/j.neuroscience.2014.09.046

Fig. 3. Effect of intraplantar injection of prazosin (20 lg in 25 ll saline) on allodynia (n= 7) and hind paw temperature (n= 4) 6 weeks after distal
tibia fracture. Allodynia did not change after injections into the ipsilateral or contralateral hind paw. However, shortly after prazosin administration,
increases in skin temperature on the injured side were greater after ipsilateral than contralateral injections (Side Injected � Time � Side Injured
interaction, p< 0.01; ⁄p< 0.05 for differences between the ipsilateral and contralateral sides). Error bars represent standard errors.
Fig. 4. CGRP expression (in terms of % total area) in peripheral nerve fibers. The images illustrate CGRP expression in nerve bundles (outlined in
white) in plantar skin (Scale = 20 lm). The main effect for Group was statistically significant for the sciatic nerve (p< 0.001), and main effects for
Group (p< 0.001) and Side (p< 0.05) were statistically significant for dermal nerves in plantar skin. For the sciatic nerve, the ⁄denotes a significant
difference in CGRP expression, averaged across the ipsilateral and contralateral sides, between the 16-week fracture group and each other group;
for plantar dermal nerves, the ⁄denotes a significant difference in CGRP expression, averaged across sides, between the 16-week cast-only group
and each other group (p< 0.05, Newman–Keuls test). In addition, #represents a significant difference between the ipsilateral and contralateral hind
paw in plantar dermal nerves (p< 0.05). Error bars represent standard errors.
6 E. S. Drummond et al. / Neuroscience xxx (2014) xxx–xxx
NSC 15719 No. of Pages 11
3 October 2014
Please cite this article in press as: Drummond ES et al. Increased bilateral expression of a1-adrenoceptors on peripheral nerves, blood vessels and keratinocytes does
not account for pain or neuroinflammatory changes after distal tibia fracture in rats. Neuroscience (2014), http://dx.doi.org/10.1016/j.neuroscience.2014.09.046

372
373
374
375
376
377
378
379
380
381
382
383
384
385
386
387
388
389
390
391
392
393
394
395
396
397
398
399
400
401
402
403
404
Fig. 5. NF200 expression in sciatic nerves. The main effect for Group
(p< 0.001) and the Group � Side interaction (p< 0.01) were
statistically significant, due primarily to a decrease in NF200 expres-
sion 4 weeks after fracture followed by an increase above baseline on
the injured side at 16 weeks (p< 0.05, Newman–Keuls test). Error
bars represent standard errors, and #represents a significant differ-
ence between sides.
Fig. 6. a1-AR expression on PGP+ neurons in the sciatic nerves
(main effect for Group, p< 0.01). The ⁄above the bars (averaged
across the ipsilateral and contralateral sides) denote significant
differences between each experimental group and the control (naı̈ve)
group (p< 0.05, Newman–Keuls test). Error bars represent standard
errors.
Fig. 7. a1-AR expression in the epidermis. The images illustrate a1-AR exp
Group was statistically significant in plantar skin (p< 0.001). The ⁄above the
greater expression than bars without the ⁄(p< 0.05, Newman–Keuls tes
references to color in this figure legend, the reader is referred to the web ve
E. S. Drummond et al. / Neuroscience xxx (2014) xxx–xxx 7
NSC 15719 No. of Pages 11
3 October 2014
Please cite this article in press as: Drummond ES et al. Increased bilateral expression o
not account for pain or neuroinflammatory changes after distal tibia fracture in rats. Ne
expression differed among groups for the sciatic
(p< 0.05) and tibial nerves (p< 0.01).
DISCUSSION
Distal tibia fracture and casting produced a range of
behavioral, physiological and anatomical changes,
including an increase in a1-AR expression on nerve
fiber populations at 4 and 16 weeks, and on plantar
keratinocytes and dermal blood vessels at the 16-week
time point (Table 2). The increase in a1-AR expression
involved not only the fractured limb but also the
contralateral uninjured limb; furthermore, a1-AR
expression was elevated in the cast-only group
12 weeks after the cast had been removed, when pain
had resolved. Four to six weeks after the fracture,
systemic injection of the a1-AR antagonist prazosin
inhibited behavioral signs of pain; however, intraplantar
injections of prazosin did not. Thus, it seems unlikely
that an increase in a1-AR expression on peripheral
nociceptive neurons or other dermal tissues was the
primary source of pain after distal tibia fracture.
Behavioral and physiological changes
As in previous studies (Guo et al., 2004), both distal tibia
fracture and cast immobilisation generated behavioral
signs of pain which resolved more quickly after cast
immobilisation than distal tibia fracture. In addition, the
hind paw was swollen in the fracture group and hind
paw temperature was elevated in both groups at 4 weeks.
Pain in the distal tibia fracture model can be attenuated by
LY303870 (a substance P antagonist) (Guo et al., 2004),
soluble TNF receptor type 1 (an antagonist for TNF)
(Sabsovich et al., 2008a), anti-nerve growth factor anti-
bodies (Sabsovich et al., 2008b), the cytokine inhibitor
pentoxifylline (Wei et al., 2009b), the IL-6 antagonist
ression (green) in plantar skin (Scale = 50 lm). The main effect for
bars (averaged across the ipsilateral and contralateral sides) indicates
t). Error bars represent standard errors. (For interpretation of the
rsion of this article.)
f a1-adrenoceptors on peripheral nerves, blood vessels and keratinocytes does
uroscience (2014), http://dx.doi.org/10.1016/j.neuroscience.2014.09.046
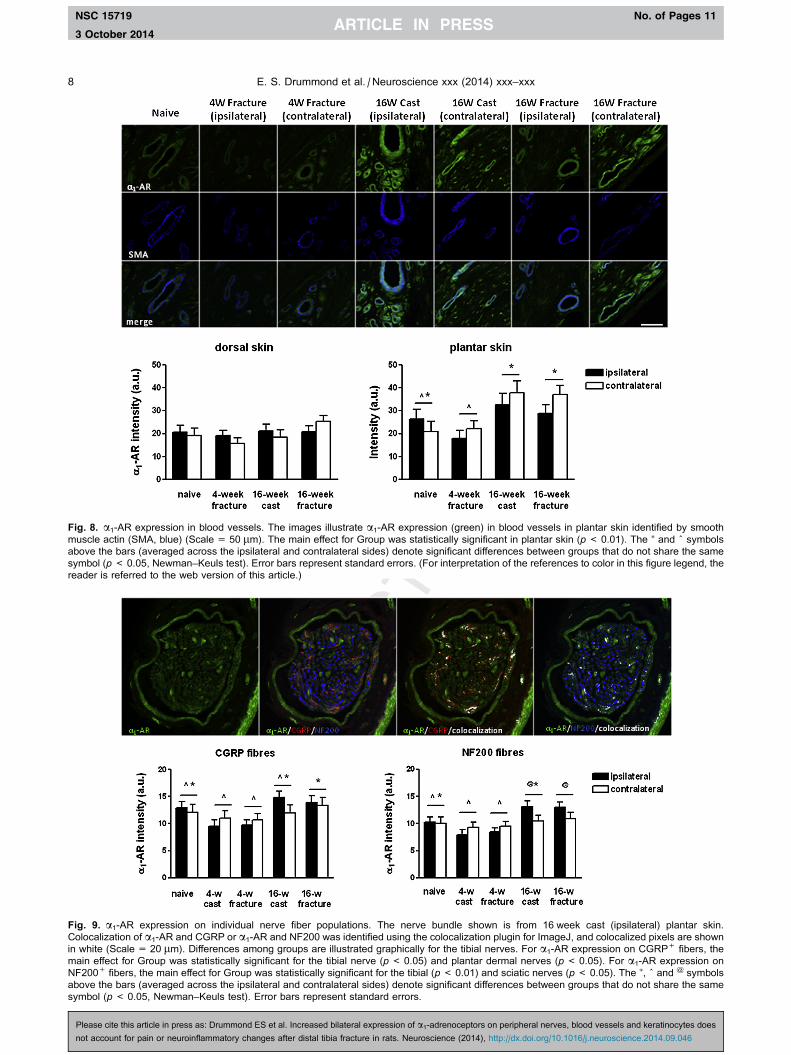
Fig. 8. a1-AR expression in blood vessels. The images illustrate a1-AR expression (green) in blood vessels in plantar skin identified by smooth
muscle actin (SMA, blue) (Scale = 50 lm). The main effect for Group was statistically significant in plantar skin (p< 0.01). The ⁄ and ^ symbols
above the bars (averaged across the ipsilateral and contralateral sides) denote significant differences between groups that do not share the same
symbol (p< 0.05, Newman–Keuls test). Error bars represent standard errors. (For interpretation of the references to color in this figure legend, the
reader is referred to the web version of this article.)
Fig. 9. a1-AR expression on individual nerve fiber populations. The nerve bundle shown is from 16 week cast (ipsilateral) plantar skin.
Colocalization of a1-AR and CGRP or a1-AR and NF200 was identified using the colocalization plugin for ImageJ, and colocalized pixels are shown
in white (Scale = 20 lm). Differences among groups are illustrated graphically for the tibial nerves. For a1-AR expression on CGRP+ fibers, the
main effect for Group was statistically significant for the tibial nerve (p< 0.05) and plantar dermal nerves (p< 0.05). For a1-AR expression on
NF200+ fibers, the main effect for Group was statistically significant for the tibial (p< 0.01) and sciatic nerves (p< 0.05). The ⁄, ^ and @ symbols
above the bars (averaged across the ipsilateral and contralateral sides) denote significant differences between groups that do not share the same
symbol (p< 0.05, Newman–Keuls test). Error bars represent standard errors.
8 E. S. Drummond et al. / Neuroscience xxx (2014) xxx–xxx
NSC 15719 No. of Pages 11
3 October 2014
Please cite this article in press as: Drummond ES et al. Increased bilateral expression of a1-adrenoceptors on peripheral nerves, blood vessels and keratinocytes does
not account for pain or neuroinflammatory changes after distal tibia fracture in rats. Neuroscience (2014), http://dx.doi.org/10.1016/j.neuroscience.2014.09.046

405
406
407
408
409
410
411
412
413
414
415
416417
418
419
420421
422
423
424
425
426
427
428
429
430
431
432
433
434
435
436
437
438
439
440
441
442
443
444
445
446
447
448
449
450
451
452
453
454
455
456
457
458
459
460
461
462
463
464
465
466
467
468
469
470
471
472
Table 2. Summary of findings
4-Week cast 4-Week fracture 16-Week cast 16-Week fracture
Allodynia and
unweighting
" Ipsilaterally " Ipsilaterally No abnormality detected " Ipsilaterally
Hind paw temperature " Ipsilaterally " Ipsilaterally No abnormality detected No abnormality detected
Edema No abnormality
detected
" Ipsilaterally No abnormality detected No abnormality detected
CGRP+ pixels No abnormality
detected
; Ipsilaterally in plantar
dermal nerves
" Most prominent in
contralateral plantar
dermal nerves
" In sciatic nerve and
greater in contralateral
than ipsilateral plantar
dermal nerves
NF200+ pixels No abnormality
detected
; Bilateral trend in the
sciatic and tibial nerves
No abnormality detected " Ipsilaterally in sciatic
nerve
a1-AR staining intensity
in the sciatic nerves
" Bilaterally " Bilaterally " Bilaterally " Bilaterally
a1-AR staining intensity
in the epidermis and
dermal blood vessels
Not measured No abnormality detected " Bilaterally in plantar
skin
" Bilaterally in plantar
skin
E. S. Drummond et al. / Neuroscience xxx (2014) xxx–xxx 9
NSC 15719 No. of Pages 11
3 October 2014
TB-2-081, chemical sympathectomy with 6-OHDA or gua-
nethidine, and butoxamine (a b2-adrenoceptor antagonist)
(Li et al., 2013). Together, these findings suggest that
neuropeptides released from nociceptive afferents and
noradrenaline secreted by sympathetic nerve fibers trig-
ger the production of inflammatory mediators (e.g., from
keratinocytes, vascular endothelial cells, mast cells or
immune cells); in turn, these inflammatory mediators sen-
sitize nociceptors and cause pain to persist (Li et al.,
2013).
In the present study, systemic administration of the
a1-AR antagonist prazosin inhibited behavioral signs of
pain 4 weeks after fracture whereas intraplantar
injections two weeks later did not. One possibility is that
fracture and/or casting triggered an up-regulation of
a1-ARs in central nociceptive pathways that augmented
pain (Holden et al., 1999; Brightwell and Taylor, 2009;
Jeong and Holden, 2009). Alternatively, the intraplantar
dose might not have blocked local a1-AR signaling com-
pletely. However, this seems unlikely as hind paw temper-
ature increased significantly after the intraplantar
injections, implying release of noradrenergic vasocon-
strictor tone. This increase was more prominent in the
ipsilateral than contralateral hind paw, possibly because
release of noradrenergic vasoconstrictor tone unmasked
underlying inflammatory vasodilator influences on the
plantar microvasculature in the injured limb.
473
474
475
476
477
478
479
480
481
482483
484
485
486
Neural markers
To determine whether distal tibia fracture-cast
immobilisation evoked signs of nerve injury, we
investigated changes in expression of the pan-neuronal
marker PGP9.5 and the selective markers NF200 and
CGRP. PGP9.5 staining did not differ significantly from
control levels after fracture or casting indicating that, on
the whole, nerve fiber populations remained stable.
NF200 is a marker for neurofilaments, which provide a
cytoskeleton for myelinated neurons and regulate
axonal diameter. Not only do they influence conduction
velocity, but they are also involved in neuronal
differentiation, axon outgrowth and regeneration (Perrot
et al., 2008; Shea et al., 2009). Axonally-transported neu-
Please cite this article in press as: Drummond ES et al. Increased bilateral expression o
not account for pain or neuroinflammatory changes after distal tibia fracture in rats. Ne
rofilament proteins decrease in injured neurons whereas
the expression of neurofilament subunits is strongly up-
regulated during regeneration (Perrot et al., 2008). Thus,
the bilateral decrease in NF200 in the sciatic nerves
4 weeks after distal tibia fracture, succeeded at 16 weeks
by heightened NF200 expression, is consistent with nerve
injury followed by repair. However, this perturbation was
relatively minor because it did not involve dermal nerves
in the hind paw. Possibly a systemic response to fracture
temporarily suppressed the production or transport of
neurofilaments in myelinated sciatic neurons.
Four weeks after distal tibia fracture, expression of the
neuropeptide CGRP had decreased in plantar dermal
nerves in the injured hind paw but 12 weeks later this
expression had increased in the contralateral hind paw
both in the cast-only and fracture groups. Although this
might indicate a cycle of denervation and reinnervation,
these changes could also reflect a temporary reduction
in CGRP due to a lag in replenishing depleted neural
stores. In particular, the increase in CGRP in
contralateral plantar dermal neurons both after fracture
and cast immobilisation suggests that a systemic
response to immobilisation triggered an increase in
CGRP production. This would be consistent with
previous findings that distal tibia fracture increased
CGRP gene expression in the dorsal root ganglia
supplying the ipsilateral hind limb without a significant
change in epidermal neurite counts (Wei et al., 2009a).
Neuropeptide signaling is actively involved in the neuro-
inflammatory response triggered by distal fracture and
immobilisation, as blocking this signaling inhibits post-
fracture allodynia, vascular changes and production of
inflammatory mediators such as IL-1b, TNF and nerve
growth factor (Wei et al., 2009b; Guo et al., 2012).
a1-AR expression
After peripheral nerve injury, heightened expression of
a1-ARs on nociceptive afferents contributes to an
increase in neural excitability and pain (Ali et al., 1999;
Xie et al., 2001; Maruo et al., 2006). Similar effects on
keratinocytes could stimulate the production and/or
release of inflammatory mediators that sensitize
f a1-adrenoceptors on peripheral nerves, blood vessels and keratinocytes does
uroscience (2014), http://dx.doi.org/10.1016/j.neuroscience.2014.09.046

487
488
489
490
491
492
493
494
495
496
497
498
499
500
501
502
503
504
505
506
507
508509
510
511
512
513
514
515
516
517
518
519
520
521
522
523
524
525
526
527
528
529
530
531
532
533
534
535
536
537
538
539
540
541
542
543 Q7
544 Q8
545
546
547
548
549
550
551
552
553
554
555
556
557
558
559
560
561
562
563
564
565
566
567
568
569
570
571
572
573
574
575
576
577
578
10 E. S. Drummond et al. / Neuroscience xxx (2014) xxx–xxx
NSC 15719 No. of Pages 11
3 October 2014
nociceptive afferents, whereas up-regulation of a1-ARs on
cutaneous blood vessels might cause pain by altering
microvascular blood flow (Coderre and Bennett, 2010).
Thus, it is tempting to speculate that an increase in expres-
sion of a1-ARs on neurons, blood vessels and keratino-
cytes contributed to pain in the fracture group. However,
it seems unlikely that this was the primary source of pain
as a1-AR expression increased bilaterally in plantar hind
paw skin both in the 16-week fracture group (associated
with signs of residual pain in the affected limb) and in the
16-week cast-only group where pain had resolved. Fur-
thermore, a1-AR expression did not increase in dorsal
hind paw skin either on the fractured/immobilized or con-
tralateral side. It might be relevant that constriction due
to post-fracture edema was minimized by providing a win-
dow in the cast over the dorsum of the paw and ankle.
Together, these findings suggest that a non-specific
effect of the procedure (e.g., psychological stress or
changes in activity or weight distribution on the hind
paws after fracture/immobilisation) induced circulatory or
systemic neural or inflammatory changes that altered
a1-AR expression. In particular, inflammatory mediators,
growth factors or hormones released after fracture and
cast immobilisation (Li et al., 2013) might initiate a1-AR
up-regulation. For example, the inflammatory mediators
TNF and IL-1b trigger the expression of the a1A-AR sub-
type in the THP-1 human monocyte cell line (Heijnen
et al., 2002). In addition, glucocorticoids and the b2-AR
agonist terbutaline increase the expression of messenger
RNA (mRNA) for a1B-AR and a1D-AR on THP-1 cells
(Rouppe van der Voort et al., 1999), and nerve growth
factor increases levels of a1B-AR mRNA and protein in
cultured dorsal root ganglion cells (Zhang and Tan, 2011).
579
580
581
582
583
584
585
586
587
588
589
590
591
592
593Q9
594
595
596
597
598
599
600
601
602
603
604
605
606
607
608
609
610
611
Limitations and conclusions
Several limitations apply to this study. In particular,
additional quantitative methods will be required to more
precisely delineate the triggers and sequence of change
in a1-AR expression after fracture and cast
immobilisation. Furthermore, effects of a1-AR blockade
were examined at only one time point, and only a single
concentration of antagonist was used.
Despite these limitations, our findings clearly show
that distal tibia fracture and cast immobilisation
produced transient reductions followed by increases in
the expression of the neural markers NF200 and CGRP,
and non-specific increases in a1-AR expression in
peripheral neurons, blood vessels and keratinocytes.
These changes are unlikely to account for persistent
pain after limb fracture. Nevertheless, further
examination of the systemic responses that generated
these changes seems warranted, to determine whether
they aggravate symptoms (e.g., by compromising neural
structure or function) or delay recovery (e.g., by
augmenting the neuroinflammatory responses that
mediate symptoms).
Acknowledgment—This work was supported by grants from the
National Health and Medical Research Council of Australia (grant
numbers APP1030379, 437205), the Australian and New Zea-
land College of Anaesthetists (grant number 10/021), and the
Please cite this article in press as: Drummond ES et al. Increased bilateral expression o
not account for pain or neuroinflammatory changes after distal tibia fracture in rats. Ne
Department of Veterans Affairs Rehabilitation and Research
Development grant F7137R.
REFERENCES
Ali Z, Ringkamp M, Hartke TV, Chien HF, Flavahan NA, Campbell JN,
Meyer RA (1999) Uninjured C-fiber nociceptors develop
spontaneous activity and alpha-adrenergic sensitivity following
L6 spinal nerve ligation in monkey. J Neurophysiol 81:455–466.
Brightwell JJ, Taylor BK (2009) Noradrenergic neurons in the locus
coeruleus contribute to neuropathic pain. Neuroscience
160:174–185.
Chabal C, Jacobson L, Russell LC, Burchiel KJ (1992) Pain response
to perineuromal injection of normal saline, epinephrine, and
lidocaine in humans. Pain 49:9–12.
Choi B, Rowbotham MC (1997) Effect of adrenergic receptor
activation on post-herpetic neuralgia pain and sensory
disturbances. Pain 69:55–63.
Coderre TJ, Bennett GJ (2010) A hypothesis for the cause of complex
regional pain syndrome-type I (reflex sympathetic dystrophy):
pain due to deep-tissue microvascular pathology. Pain Med
11:1224–1238.
Dawson LF, Phillips JK, Finch PM, Inglis JJ, Drummond PD (2011)
Expression of alpha1-adrenoceptors on peripheral nociceptive
neurons. Neuroscience 175:300–314.
de Mos M, de Bruijn AG, Huygen FJ, Dieleman JP, Stricker BH,
Sturkenboom MC (2007) The incidence of complex regional pain
syndrome: a population-based study. Pain 129:12–20.
Dogrul A, Coskun I, Uzbay T (2006) The contribution of alpha-1 and
alpha-2 adrenoceptors in peripheral imidazoline and adrenoceptor
agonist-induced nociception. Anesth Analg 103:471–477. table of
contents.
Donello JE, Guan Y, Tian M, Cheevers CV, Alcantara M, Cabrera S,
Raja SN, Gil DW (2011) A peripheral adrenoceptor-mediated
sympathetic mechanism can transform stress-induced analgesia
into hyperalgesia. Anesthesiology 114:1403–1416.
Drummond PD, Skipworth S, Finch PM (1996) Alpha 1-
adrenoceptors in normal and hyperalgesic human skin. Clin Sci
91:73–77.
Drummond ES, Dawson LF, Finch PM, Bennett GJ, Drummond PD
(2014a) Increased expression of cutaneous a1-adrenoceptors
after chronic constriction injury in rats. J Pain 15:188–196.
Drummond PD, Drummond ES, Dawson LF, Mitchell V, Finch PM,
Vaughan CW, Phillips JK (2014b) Up-regulation of a1-
adrenoceptors on cutaneous nerve fibres after partial sciatic
nerve ligation and in complex regional pain syndrome type II. Pain
155:606–616.
Finch PM, Drummond ES, Dawson LF, Phillips JK, Drummond PD
(2014) Up-regulation of cutaneous alpha1-adrenoceptors in
complex regional pain syndrome type I. Pain Med [in press].
Gibbs GF, Drummond PD, Finch PM, Phillips JK (2008) Unravelling
the pathophysiology of complex regional pain syndrome: focus on
sympathetically maintained pain. Clin Exp Pharmacol Physiol
35:717–724.
Guo TZ, Tinklenberg J, Oliker R, Maze M (1991) Central alpha 1-
adrenoceptor stimulation functionally antagonizes the hypnotic
response to dexmedetomidine, an alpha 2-adrenoceptor agonist.
Anesthesiology 75:252–256.
Guo TZ, Offley SC, Boyd EA, Jacobs CR, Kingery WS (2004)
Substance P signaling contributes to the vascular and nociceptive
abnormalities observed in a tibial fracture rat model of complex
regional pain syndrome type I. Pain 108:95–107.
Guo TZ, Wei T, Kingery WS (2006) Glucocorticoid inhibition of
vascular abnormalities in a tibia fracture rat model of complex
regional pain syndrome type I. Pain 121:158–167.
Guo TZ, Wei T, Shi X, Li WW, Hou S, Wang L, Tsujikawa K, Rice KC,
Cheng K, Clark DJ, Kingery WS (2012) Neuropeptide deficient
mice have attenuated nociceptive, vascular, and inflammatory
f a1-adrenoceptors on peripheral nerves, blood vessels and keratinocytes does
uroscience (2014), http://dx.doi.org/10.1016/j.neuroscience.2014.09.046

612
613
614
615
616
617
618
619
620
621
622
623
624
625
626
627
628
629
630
631
632
633
634
635
636
637
638
639
640
641
642
643
644
645
646
647
648
649
650
651
652
653
654
655
656
657
658
659
660
661
662
663
664
665
666
667
668
669
670
671
672
673
674
675
676
677
678
679
680
681
682
683
684
685
686
687
688
689
690
691
692
693
694
695
696
697
698
699
700
701
702
703
704
705
706
707
708
709
710
711
712
713
714
715
716
717
718
719
720
721
722
723
724
725
726
727
728
729
730
E. S. Drummond et al. / Neuroscience xxx (2014) xxx–xxx 11
NSC 15719 No. of Pages 11
3 October 2014
changes in a tibia fracture model of complex regional pain
syndrome. Mol Pain 8:85.
Heijnen CJ, Rouppe van der Voort C, van de Pol M, Kavelaars A
(2002) Cytokines regulate alpha(1)-adrenergic receptor mRNA
expression in human monocytic cells and endothelial cells. J
Neuroimmunol 125:66–72.
Holden JE, Schwartz EJ, Proudfit HK (1999) Microinjection of
morphine in the A7 catecholamine cell group produces opposing
effects on nociception that are mediated by alpha1- and alpha2-
adrenoceptors. Neuroscience 91:979–990.
Hong Y, Abbott FV (1996) Contribution of peripheral alpha 1A-
adrenoceptors to pain induced by formalin or by alpha-methyl-5-
hydroxytryptamine plus noradrenaline. Eur J Pharmacol
301:41–48.
Hord AH, Denson DD, Stowe B, Haygood RM (2001) Alpha-1 and
alpha-2 adrenergic antagonists relieve thermal hyperalgesia in
experimental mononeuropathy from chronic constriction injury.
Anesth Analg 92:1558–1562.
Jeong Y, Holden JE (2009) Lateral hypothalamic-induced alpha-
adrenoceptor modulation occurs in a model of inflammatory pain
in rats. Biol Res Nurs 10:331–339.
Kim SK, Min BI, Kim JH, Hwang BG, Yoo GY, Park DS, Na HS (2005)
Effects of alpha1- and alpha2-adrenoreceptor antagonists on cold
allodynia in a rat tail model of neuropathic pain. Brain Res
1039:207–210.
Kingery WS, Agashe GS, Guo TZ, Sawamura S, Davies MF, Clark
JD, Kobilka BK, Maze M (2002) Isoflurane and nociception: spinal
alpha2A adrenoceptors mediate antinociception while supraspinal
alpha1 adrenoceptors mediate pronociception. Anesthesiology
96:367–374.
Kingery WS, Davies MF, Clark JD (2003) A substance P receptor
(NK1) antagonist can reverse vascular and nociceptive
abnormalities in a rat model of complex regional pain syndrome
type II. Pain 104:75–84.
Kopp UC, Cicha MZ, Smith LA, Mulder J, Hokfelt T (2007) Renal
sympathetic nerve activity modulates afferent renal nerve activity
by PGE2-dependent activation of alpha1- and alpha2-
adrenoceptors on renal sensory nerve fibers. Am J Physiol
Regul Integr Comp Physiol 293:R1561–R1572.
Lee YH, Ryu TG, Park SJ, Yang EJ, Jeon BH, Hur GM, Kim KJ
(2000) Alpha1-adrenoceptors involvement in painful diabetic
neuropathy: a role in allodynia. Neuroreport 11:1417–1420.
Li WW, Sabsovich I, Guo TZ, Zhao R, Kingery WS, Clark JD (2009)
The role of enhanced cutaneous IL-1beta signaling in a rat tibia
fracture model of complex regional pain syndrome. Pain
144:303–313.
Li WW, Guo TZ, Li XQ, Kingery WS, Clark JD (2010) Fracture
induces keratinocyte activation, proliferation, and expression of
pro-nociceptive inflammatory mediators. Pain 151:843–852.
Li WW, Guo TZ, Liang DY, Sun Y, Kingery WS, Clark JD (2012)
Substance P signaling controls mast cell activation,
degranulation, and nociceptive sensitization in a rat fracture
model of complex regional pain syndrome. Anesthesiology
116:882–895.
Li W, Shi X, Wang L, Guo T, Wei T, Cheng K, Rice KC, Kingery WS,
Clark JD (2013) Epidermal adrenergic signaling contributes to
inflammation and pain sensitization in a rat model of complex
regional pain syndrome. Pain 154:1224–1236.
Please cite this article in press as: Drummond ES et al. Increased bilateral expression o
not account for pain or neuroinflammatory changes after distal tibia fracture in rats. Ne
Lin EE, Horasek S, Agarwal S, Wu CL, Raja SN (2006) Local
administration of norepinephrine in the stump evokes dose-
dependent pain in amputees. Clin J Pain 22:482–486.
Marinus J, Moseley GL, Birklein F, Baron R, Maihofner C, Kingery
WS, van Hilten JJ (2011) Clinical features and pathophysiology of
complex regional pain syndrome. Lancet Neurol 10:637–648.
Maruo K, Yamamoto H, Yamamoto S, Nagata T, Fujikawa H, Kanno
T, Yaguchi T, Maruo S, Yoshiya S, Nishizaki T (2006) Modulation
of P2X receptors via adrenergic pathways in rat dorsal root
ganglion neurons after sciatic nerve injury. Pain 120:106–112.
Meisner JG, Waldron JB, Sawynok J (2007) Alpha1-adrenergic
receptors augment P2X3 receptor-mediated nociceptive
responses in the uninjured state. J Pain 8:556–562.
Nam TS, Yeon DS, Leem JW, Paik KS (2000) Adrenergic sensitivity
of uninjured C-fiber nociceptors in neuropathic rats. Yonsei Med J
41:252–257.
Perrot R, Berges R, Bocquet A, Eyer J (2008) Review of the multiple
aspects of neurofilament functions, and their possible contribution
to neurodegeneration. Mol Neurobiol 38:27–65.
Rouppe van der Voort C, Kavelaars A, van de Pol M, Heijnen CJ
(1999) Neuroendocrine mediators up-regulate alpha1b- and
alpha1d-adrenergic receptor subtypes in human monocytes. J
Neuroimmunol 95:165–173.
Sabsovich I, Guo TZ, Wei T, Zhao R, Li X, Clark DJ, Geis C, Sommer
C, Kingery WS (2008a) TNF signaling contributes to the
development of nociceptive sensitization in a tibia fracture model
of complex regional pain syndrome type I. Pain 137:507–519.
Sabsovich I, Wei T, Guo TZ, Zhao R, Shi X, Li X, Yeomans DC,
Klyukinov M, Kingery WS, Clark JD (2008b) Effect of anti-NGF
antibodies in a rat tibia fracture model of complex regional pain
syndrome type I. Pain 138:47–60.
Schmidt M, Kress M, Heinemann S, Fickenscher H (2003) Varicella-
zoster virus isolates, but not the vaccine strain OKA, induce
sensitivity to alpha-1 and beta-1 adrenergic stimulation of sensory
neurones in culture. J Med Virol 70(suppl. 1):S82–S89.
Shea TB, Chan WK, Kushkuley J, Lee S (2009) Organizational
dynamics, functions, and pathobiological dysfunctions of
neurofilaments. Results Prob Cell Differ 48:29–45.
Trevisani M, Campi B, Gatti R, Andre E, Materazzi S, Nicoletti P,
Gazzieri D, Geppetti P (2007) The influence of alpha1-
adrenoreceptors on neuropeptide release from primary sensory
neurons of the lower urinary tract. Eur Urol 52:901–908.
Wei T, Li WW, Guo TZ, Zhao R, Wang L, Clark DJ, Oaklander AL,
Schmelz M, Kingery WS (2009a) Post-junctional facilitation of
substance P signaling in a tibia fracture rat model of complex
regional pain syndrome type I. Pain 144:278–286.
Wei T, Sabsovich I, Guo TZ, Shi X, Zhao R, Li W, Geis C, Sommer C,
Kingery WS, Clark DJ (2009b) Pentoxifylline attenuates
nociceptive sensitization and cytokine expression in a tibia
fracture rat model of complex regional pain syndrome. Eur J
Pain 13:253–262.
Xie J, Ho Lee Y, Wang C, Mo Chung J, Chung K (2001) Differential
expression of alpha1-adrenoceptor subtype mRNAs in the dorsal
root ganglion after spinal nerve ligation. Brain Res Mol Brain Res
93:164–172.
Zhang Q, Tan Y (2011) Nerve growth factor augments neuronal
responsiveness to noradrenaline in cultured dorsal root ganglion
neurons of rats. Neuroscience 193:72–79.
(Accepted 19 September 2014)(Available online xxxx)
f a1-adrenoceptors on peripheral nerves, blood vessels and keratinocytes does
uroscience (2014), http://dx.doi.org/10.1016/j.neuroscience.2014.09.046





























