Embed Size (px)
Citation preview

Multifunctional Nanoemulsion System for Combination Paclitaxel and Curcumin Delivery in
Human Glioblastoma Cells
Master’s Thesis
By
Ms. Sindhura Ganga
Advisor: Mansoor M. Amiji, PhD
to
The Bouve’ Graduate School of Health Sciences in Partial Fulfillment of the Requirement for the Degree of Master of Science
in Pharmaceutical Science with Specialization in Pharmaceutics and Drug Delivery
Department of Pharmaceutical Sciences Northeastern University
August, 2008
1

PROJECT SUMMARY Glioblastoma multiforme (GBM) is an aggressive form of primary brain tumor that
afflicts as many as 12,000 patients per year in the United States. GBM is a very aggressive
disease and the average patient life-span is reduced to 6 months after diagnosis. Some of the
clinical challenges in GBM include difficulty in initial diagnosis, presence of tumor cell “islets”
in the critical areas of the brain, and the role of blood-brain barrier (BBB) in effective transport
and distribution of drugs in the brain. Due to the presence of tumor cell islets, complete surgical
resection is not always possible. Additionally, poor penetration of many chemotherapeutic agents
across the BBB limits availability and distribution for effective treatment of GBM. In addition to
the presence of a physical barrier in the form of tight endothelial cell junctions, BBB is also
comprised of chemical barriers in the form of expression of the efflux pump, P-glycoprotein (P-
gp), and the presence of drug metabolizing enzymes and the vascular endothelial cell surface.
GBM also has high incidence of resistance development due to the expression of multidrug
resistant protein (MRP).
In this study, our main objective was to evaluate co-administration of paclitaxel (PTX)
and curcumin (CUR), a modulator of MRP and an inhibitor of nuclear factor kappa B (NFκB)
signaling, in oil-in-water nanoemulsion formulations in brain tumor cells. CUR is known to
down-regulate both P-gp and MRP expression and also enhances apoptotic activity by inhibition
of NFκB mediated intracellular signaling. The nanoemulsion formulations in this study are
specifically composed of oils rich in polyunsaturated fatty acids (PUFA) and the surface was
modified to enhance BBB penetration.
As part of the study, PTX and CUR containing nanoemulsions have been formulated with
pine-nut oil, which is rich in linoleic and linolenic acids, an example of omega-6 and omega-3
2

PUFA, respectively. The optimized formulations were characterized for oil droplet size, surface
charge, and observed with transmission electron microscopy (TEM). Human wild-type
glioblastoma cells U87 and the MRP-1 expressing resistant glioblastoma cells, T98G, were
grown in culture. Baseline expression of MRP-1 in T98G cells was confirmed by Western blot
and immunocytochemistry analysis. Intracellular delivery of fluorescently-labeled PTX and CUR
using nanoemulsion formulation was examined by fluorescence microscopy. The cell viability
upon treatment with PTX and CUR, either alone or in combination, was examined in both of
these cell lines using the MTT (conversion of tetrazolium salt to formazan) assay. MRP-1 down-
regulation and the inhibition of NFκB pathway was confirmed by Western blot analysis.
Qualitative apoptotic activity in U87 and T98G cells was evaluated using TUNEL staining.
Pine-nut oil nanoemulsions were successfully formulated with an approximate
hydrodynamic diameter of 120 nm and a -40 mV surface charge. Both PTX and CUR were
efficiently encapsulated in the oil phase of the nanoemulsion. TEM analysis showed that the oil
droplets of the nanoemulsion had a spherical shape and smooth surface morphology. Due to
rapid cellular uptake and internalization of PTX and CUR, as assessed by fluorescence
microscopy, the nanoemulsion formulations were able to significantly enhance cytotoxicity in
both cell lines. Combination of PTX and CUR were found especially to enhance cytotoxicity in
MRP-1 gene expressing T98G cells. CUR was found to down-regulate MRP-1 and was found to
inhibit NFκB pathway. TUNEL staining images confirmed the apoptosis in the treated U87 and
T98G cells. The overall results allowed us to understand the synergistic therapeutic effect upon
administration of PTX and CUR using nanoemulsions made with PUFA-rich oil.
3

NORTHEASTERN UNIVERSITY
Graduate School of Bouvé College of Health Sciences
Thesis Title: Multifunctional Nanoemulsion System for Combination Paclitaxel and Curcumin Delivery in Human Glioblastoma Cells
Author: Sindhura Ganga
Department: Pharmaceutical Sciences
Approved for Thesis Requirements of the Master of Science Degree in Pharmaceutical Science
Dissertation Committee
____________________________________ __________________ Date
____________________________________ __________________
Date ____________________________________ __________________ (Chairman) Date ____________________________________ __________________
Director of Graduate School Date
4

ACKNOWLEDGEMENTS
I have been waiting eagerly for this moment to express my heartfelt gratitude to those
people, who had a major role in the completion of my thesis project and to everyone who wished
good for me.
First and foremost, I would like to express my deepest gratitude, respect, and appreciation
to my advisor and my role model, Dr. Mansoor Amiji. If it weren’t for his ideas, constant
support, guidance, motivation, and patience, none of this would have been possible. Thank you
for making a difference in my career and for having faith in me that I could complete this
project.
I would like to thank my dissertation committee members, Dr. Rebecca Carrier and Dr.
Ralph Loring, for their time, support, and suggestions throughout the course of this work. I
would also like to thank Dr. Robert Campbell, Dr. Vladimir Torchillin and Bill Fowle for being
kind and for allowing me to use various analytical instruments in their laboratory.
I always felt like being with my family when I was working in the lab. I have received all
means of support, guidance, and encouragement from the lab members. I would like to first
thank Harikrishna Devalapally and Padmaja Magadala for helping me in starting with the
project. Lilian van Vlerken, Luis Brito, Lara Jabr-Milane, Mayank Bhavsar, Srinivas Ganta,
Thomas Barchet, and Sandra Chadwick for their valuable suggestions. I would like to specially
thank Sunaina Pai and Shardool Jain for their amazing company. Last, but not the least, I thank
my parents for all their love and sacrifices and for making my dreams come true. To them, I
dedicate this thesis.
5

TABLE OF CONTENTS
PROJECT SUMMARY 2
ACKNOWLEDGEMENTS 5
TABLE OF CONTENTS 6
List of Tables
8
List of Figures
9
1.
INTRODUCTION………………………………………………………………………
11
1.1 Glioblastoma: Incidence and Mortality ……………………………………............ 11
1.2 Role of the Blood-Brain Barrier in GBM Therapy………………………………… 11
1.3 Paclitaxel and Curcumin in Glioblastoma Therapy………………………………… 13
1.4 Nanotechnology for Brain Delivery…………………………..……………………. 18
1.5 Multifunctional Nanoemulsions for PTX/CUR Combination Therapy …………… 20
2.
OBJECTIVES AND SPECIFIC AIMS…………………………………………………
23
2.1 Statement of the Problem……………………………………………………...........
23
2.2 Objectives and Hypotheses………………………………………………………….
23
2.3 Specific Aims………………………………………………………...……………..
24
3.
MATERIAL AND METHODS……………………………………………………….
26
3.1 Preparation of Oil-in-Water Nanoemulsions ………………………………………. 26
3.2 Characterization of the Nanoemulsions ………………………................................. 26
6

3.3 Determination of Baseline MRP Expression in GBM Cells ……………………..
28
3.4 Fluorescence Microscopic Studies of Intracellular Delivery with Nanoemulsions... 30
3.5 Determination of MRP1 Down-Regulation by Western Blot Analysis ………..….. 31
3.6 Determination of NFκB pathway Inhibition by Western Blot Analysis ………..
31
3.7 Cytotoxicity of Single and Combination Treatments in GBM Cells………………. 33
3.8 Qualitative Apoptosis Study by TUNEL Staining ………………………………… 34
4.
RESULTS AND DISCUSSIONS……………………………………………………...
36
4.1 Preparation and Characterization of Multifunctional Nanoemulsions……………...
36
4.2 Determination of Baseline MRP-1 Expression in GBM Cells…………………….. 37
4.3 Cellular Uptake and Distribution Studies………………………………………….. 39
4.4 Down Regulation of MRP-1 and the Inhibition of NFκB Pathway with CUR……. 41
4.5 Cytotoxicity of Single and Combination Treatments in GBM Cells………………. 43
4.6 Qualitative Apoptosis Study by TUNEL Staining…………………………………. 47
5.
CONCLUSIONS………………………………………………………………………..
51
6.
REFERENCES……………………………………………………………….................
53
7

LIST OF TABLES
Table 1. Physical characteristics of blank and drug-containing nanoemulsions
8

LIST OF FIGURES
Figure 1. Schematic depiction of the blood-brain barrier in brain microvasculature
Figure 2. The chemical structures of paclitaxel and curcumin
Figure 3. Proposed multimodal mechanism of MDR modulation with curcumin
Figure 4. Transmission electron micrograph of pine-nut oil nanoemulsion for the co-administration of paclitaxel and curcumin
Figure 5. Immunocytometric analysis of MRP expression in U87 and T98G glioblastoma
cells Figure 6. Western blot analysis of MRP expression in U87 and T98G glioblastoma cells
Figure 7. Differential interference contrast (DIC) and epi-fluorescence microscopy images (20X) of control and PTX and CUR combination nanoemulsion (PTX+CUR-NE) treated U87 cells. The cells were incubated with the nanoemulsions for 8 hours at 37°C
Figure 8. Differential interference contrast (DIC) and epi-fluorescence microscopy images
(20X) of control and PTX and CUR combination nanoemulsion (PTX+CUR-NE) treated T98G cells. The cells were incubated with the nanoemulsions for 8 hours at 37°C
Figure 9: MRP-1 down-regulation and inhibition of NFκB pathway by Western blot
analysis. Figure 10: Cytotoxicity of PTX alone (A, B) and CUR alone (C, D) on U87 (A, C) and T98G
cells (B, D) when administered in the form of solutions and nanoemulsions Figure 11: Cytotoxicity of PTX and CUR (5, 10, 20 μM) combinations on U87 cells, when
administered in the form of solutions (A, B, C) and nanoemulsions (D, E, F) Figure 12: Cytotoxicity of PTX and CUR (5, 10, 20 μM) combinations on T98G cells, when
administered in the form of solutions (A, B, C) and nanoemulsions (D, E, F) Figure 13: Comparison of cytotoxicity of PTX and CUR combination, administered together
in the form of nanoemulsion with PTX alone on MRP-1 expressing T98G cells Figure 14: TUNEL staining images (20X) of untreated U87 and T98G cells
9

Figure 15: TUNEL staining images (40 X) of apoptotic nuclei of U87 cells upon treatment with solutions (PTX-S, CUR-S, PTX+CUR-S) and nanoemulsions (PTX-NE, CUR-NE, PTX+CUR- NE)
Figure 16: TUNEL staining images (40 X) of apoptotic nuclei of T98G cells upon treatment
with solutions (PTX-S, CUR-S, PTX+CUR-S) and nanoemulsions (PTX-NE, CUR-NE, PTX+CUR- NE)
10

1. INTRODUCTION
1.1 Glioblastoma: Incidence and Mortality
Primary brain tumors comprise only 2% of the malignant tumors in human beings (1, 2).
Glioblastoma multiforme (GBM) and anaplastic astrocytoma are the two malignant gliomas that
occur more frequently than any other types of primary brain tumors (3). They have a combined
incidence of 5-8 per 100,000 populations. Although the incidence of GBM is low, it comprises
the most aggressive form of brain tumor with a median survival rate of less than one year. The
standard treatment of malignant gliomas involve surgical excision followed by radiation therapy
with or without adjuvant chemotherapy (4). Even then, GBM and other brain tumors almost
always relapse and become refractory to traditional therapeutic strategies. Difficulty in early
detection, the low tolerance of the brain tissue to radiation, the presence of “islets” of tumor cells
disseminated in critical regions of the brain, and the presence of the blood-brain barrier (BBB)
makes it very challenging to treat malignant gliomas effectively (2).
1.2 Role of the Blood-Brain Barrier in GBM Therapy
The BBB was first observed by Ehrlich and Goldman (5), when they found that the
hydrophilic compound, tryptan blue, injected into the rat did not get distributed into and out of
the brain (Figure 1). The BBB is a dynamic interface separating blood from the extracellular
fluid in the brain parenchyma. It regulates the influx and efflux of biological molecules thus
eliminating the toxic substances from the endothelial compartment and supplying the brain with
nutrients and endogenous compounds (6). On the whole, the BBB serves as an important barrier
to protect the brain from insults and for regulation of its homeostasis. Lack of effective
permeability across the BBB is a major limitation for delivering drugs to treat brain tumors or
11
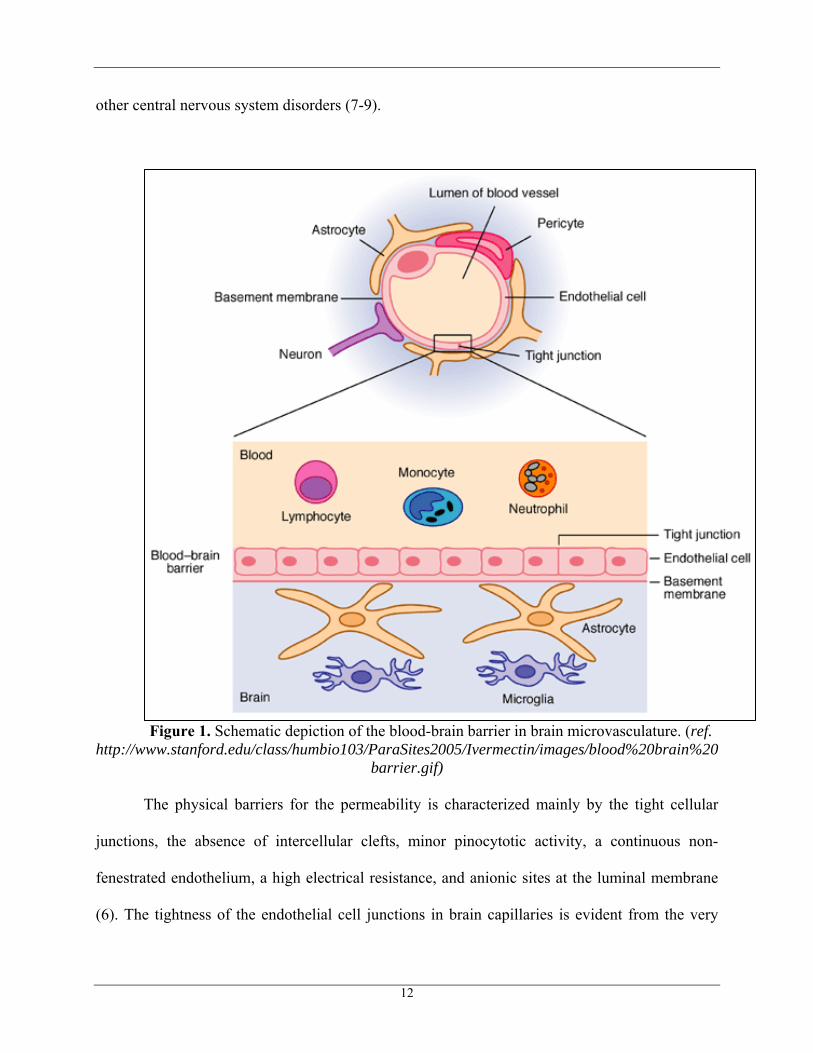
other central nervous system disorders (7-9).
Figure 1. Schematic depiction of the blood-brain barrier in brain microvasculature. (ref.
http://www.stanford.edu/class/humbio103/ParaSites2005/Ivermectin/images/blood%20brain%20barrier.gif)
The physical barriers for the permeability is characterized mainly by the tight cellular
junctions, the absence of intercellular clefts, minor pinocytotic activity, a continuous non-
fenestrated endothelium, a high electrical resistance, and anionic sites at the luminal membrane
(6). The tightness of the endothelial cell junctions in brain capillaries is evident from the very
12

high trans-endothelial electrical resistance of 1,500-2,000 Ωcm2 as compared to 3-33 Ωcm2 in
peripheral tissues (8, 9). Additionally, the astrocytes surround about 85% of the surface of the
capillaries, adding one more lipid barrier to the system. The endothelial cells that line the luminal
surface of the brain capillaries also express ATP-binding cassette (ABC) drug transporters, P-
glycoprotein (P-gp), and members of the multidrug resistance protein (MRP) family at a high
density (10). These efflux pumps remove a wide range of molecules from the intracellular
compartment of the endothelial cell before they can enter the brain parenchyma. Many cytotoxic
chemotherapeutic agents that have shown very high efficacy in brain tumor, such as paclitaxel
(PTX), vincristine, and vinblastine are pumped out by the P-gp efflux pump, leading to
inefficient anti-cancer therapy.
Drug transport across the BBB depends on its systemic pharmacokinetics, which include
absorption, distribution, metabolism and excretion. The physicochemical properties of the drug,
like lipophilicity, hydrophilicity, and hydrogen bonding potential plays an important role in the
passive transport of drugs across the BBB. Hydrophilic paracellular transport and the lipophilic
transcellular transport are the two kinds of passive diffusive transports, drugs undergo to cross
the BBB and they are mainly dependent on the size and charge of the molecule and the hydrogen
bonding potential. Drugs which are highly lipophilic and partially permeable through the tight
junctions of BBB are restricted because of the factors like P-gp efflux and cytochrome P-450
metabolizing enzymes, whereas hydrophilic drugs are completely impermeable by passive
diffusion through the tight junctions the BBB. Therefore, the BBB is an extremely important
physical and chemical hurdle that restricts delivery of drugs for CNS disorders (6).
1.3 Paclitaxel and Curcumin in Glioblastoma Therapy
Paclitaxel (PTX) is a very potent anticancer drug, originally derived from the bark of the
13

pacific Yew tree (Taxus brevifolia). It has got tremendous potential against the breast, ovarian,
cervical and many other tumors. PTX interferes with the dynamic instability of the microtubules
by inducing tubulin polymerization and thereby interferes with the mitotic spindle formation and
arrests the cell cycle at G2/M phase, finally leading to apoptosis (11, 12). Although PTX is
highly effective in treatment of glioblastoma in vitro, the in vivo delivery to the brain is restricted
due to BBB presence and the expression of P-glycoprotein efflux mechanism (13). As such, PTX
efficiency in treating GBM and other brain tumors is hindered by lack of efficacy and potential
chemoresistance development. Most importantly, PTX activates NFκB pathway and hence
promotes cell survival, proliferation and metastatis (14-16). PTX is also found to down-regulate
the cellular proteins that promote cell survival and block apoptosis such as BcL-2 and Bcl-XL
(17) . Another important chemoresistance phenomenon acquired by cytotoxic drugs, like PTX, is
the resistance involving P-gp expression. PTX is a P-gp substrate and this allows less drug to get
accumulated inside the cell and, hence results in reduced cell-kill efficacy (18).
Curcumin (CUR) is a yellow-colored polyphenolic compound extracted from the
turmeric rhizome (Curcumin longa) which is an Indian spice and also a coloring agent used in
variety of Asian foods. CUR widely exhibits antioxidant, anti-inflammatory, and anti-cancer
properties (19, 20). It has been found that free CUR induces apoptosis in many tumor cell lines
derived from breast, colorectal, lung and prostate carcinoma. CUR is also found to inhibit anti-
apoptotic, proliferative, and metastatic proteins in breast cancer cell lines (21). Additionally,
CUR has the potential to modulate multidrug resistance (MDR) in cancer (22, 23). MDR is a
phenomenon in cancer therapy, where the tumor cells develop resistance to a variety of
structurally and functionally dissimilar chemotherapeutic agents (24). MDR phenotype of cancer
cells is caused by various molecular mechanisms. They include over expression of the ATP-
14
dependent drug efflux pump of ABC transporter family like P-glycoprotein (P-gp) and the
members of MRP family. MDR mechanisms also include modifications of the apoptotic
signaling, alterations in DNA repair mechanisms, and modifications in drug metabolism through
over-expression of glutathione-S-transferase (GST) and cytochrome P450 activity (24, 25).
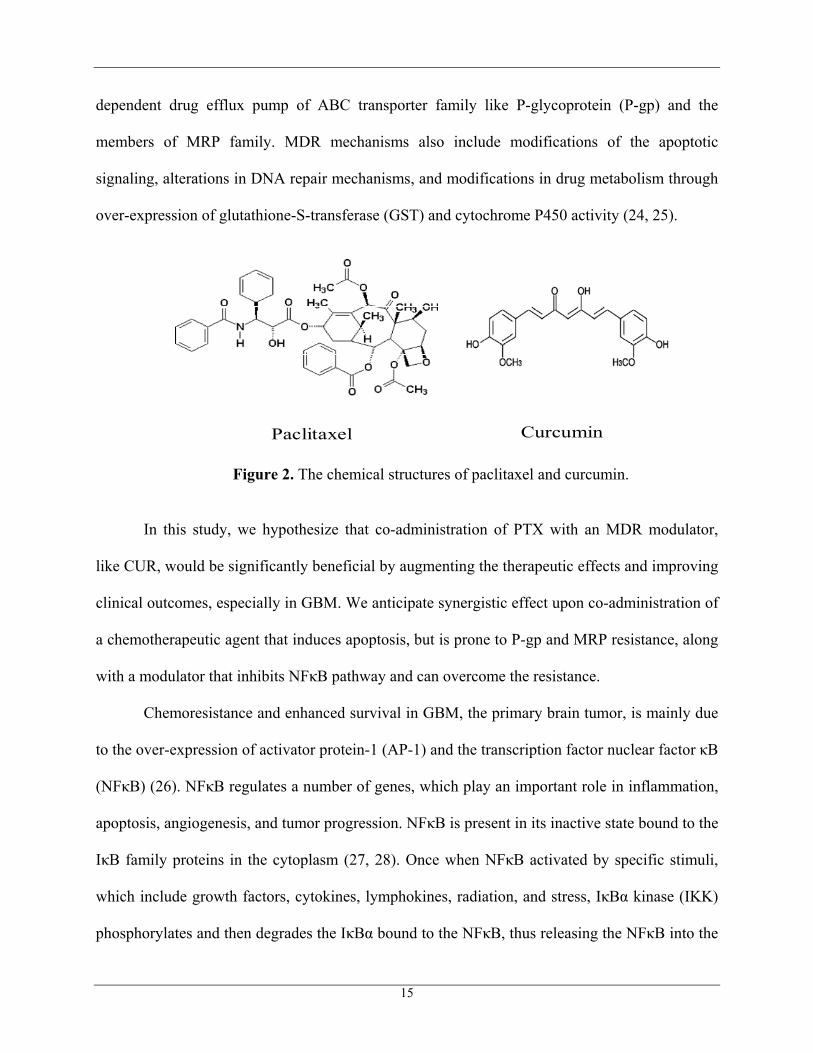
Paclitaxel Curcumin
Figure 2. The chemical structures of paclitaxel and curcumin.
In this study, we hypothesize that co-administration of PTX with an MDR modulator,
like CUR, would be significantly beneficial by augmenting the therapeutic effects and improving
clinical outcomes, especially in GBM. We anticipate synergistic effect upon co-administration of
a chemotherapeutic agent that induces apoptosis, but is prone to P-gp and MRP resistance, along
with a modulator that inhibits NFκB pathway and can overcome the resistance.
Chemoresistance and enhanced survival in GBM, the primary brain tumor, is mainly due
to the over-expression of activator protein-1 (AP-1) and the transcription factor nuclear factor κB
(NFκB) (26). NFκB regulates a number of genes, which play an important role in inflammation,
apoptosis, angiogenesis, and tumor progression. NFκB is present in its inactive state bound to the
IκB family proteins in the cytoplasm (27, 28). Once when NFκB activated by specific stimuli,
which include growth factors, cytokines, lymphokines, radiation, and stress, IκBα kinase (IKK)
phosphorylates and then degrades the IκBα bound to the NFκB, thus releasing the NFκB into the
15

nucleus, which in turn binds to the DNA and activates transcription of various genes, which
helps in cell proliferation and survival. In other way, AP-1 a transcription factor is activated by
JNK pathway (29). C-Jun N-terminal kinases (JNKs) phosphorylate c-Jun, which combines with
c-Fos to form the AP-1. Together, AP-1 and NFκB were found to be the potential targets for
GBM therapy and CUR has been found to decrease the activity of AP-1 and inhibit the NFκB
pathway (26). In this case, the apoptosis induced by CUR was found to be p53 and caspase
independent. CUR inhibits the NFκB by inhibiting the activation of IKK and, thus inhibiting the
subsequent phosphorylation and degradation of IκBα. Additionally, CUR is known to down-
regulates the NFκB-regulated gene products such as Bcl-2, Bcl-XL, cyclin D1, matrix
metalloproteinase-9, cyclooxygenase-2, and interleukin-6, resulting in cell cycle arrest,
suppression of proliferation, and induction of apoptosis (26). Curcumin also inhibited the AP-1
signalling pathway by decreasing the constitutive phosphorylation of the AP-1 transcription
factor, c-Jun.
The other factors making the glioma therapy ineffective is the intrinsic or the acquired
drug resistance phenomenon with the over expression of P-gp and other members of the MRP
family, especially MRP-1, MRP-3 and MRP-5 and glutathione-S-transferase pi (GST-pi) (30). P-
gp is a 170-kDa membrane glycoprotein with 12 membrane spanning domain and is encoded by
the mdr1 gene (31, 32). P-gp serves as a xenobiotic pump in lung, intestine, kidney, and placenta
and is overly expressed on physiological barriers including the BBB. Many cationic and neutral
compounds, in addition to many anti-cancer drugs like taxanes and vinca alkaloids, are substrates
for the drug transporters. Over-expression of P-gp, especially in tumor cells, will not allow drugs
to accumulate in the cells and results in reduced cytotoxicity. Combination of P-gp inhibitors and
conventional chemotherapeutic drugs was found to be efficacious in treating tumors.
16

Curcuminoids were found to inhibit both P-gp function and expression in KB-V1 cells and can
reverse the multidrug resistance phenomenon (33, 34). Commercially available curcumin is a
mixture of curcuminoids, curcumin I, II & III. All three of these curcuminoids inhibit P-gp, but
curcumin I is more potent due to the presence of two methoxy groups and two hydroxyl groups,
which helped in binding more to P-gp. It was also found that P-gp does not confer resistance to
CUR and also modulation of P-gp by CUR was not cell type dependent. On the other hand,
members of the MRP family show the MDR phenomenon, being capable of carrying wide range
of anticancer drugs. MRP-1 is a 190-kDa protein with 17 transmembrane spanning domains
encoded by mrp1 gene (32, 35). Its expression was also found high in BBB. It mainly effluxes
drugs, that are either conjugated or co-transported with glutathione. It prefers transporting
organic anions. Its function is found entirely different from the P-gp of the same ABC transporter
family.
Figure 3. Proposed multimodal mechanisms of MDR modulation with curcumin.
17

CUR and its glutathione conjugates are found to inhibit MRP-1 and MRP-2 (36). The
mechanism of inhibition is still unknown. Even the MRP modulators can be given along with
anti-cancer agents for effective cancer chemotherapy.
In the effective management of GBM, therefore, PTX can be given in combination with
CUR, which can show the synergistic effect, by inhibiting the NFκB pathway and
downregulating the P-gp and other multidrug resistance proteins, which are over-expressed in the
tumor cells and the blood brain barrier. It was found that in breast cancer, PTX activated NFκB
pathway involving IKK activation, IκBα phosphorylation and degradation and NFκB-regulated
gene expression, whereas, CUR completely suppressed IKK activation and thus suppressing
NFκB activation. CUR also suppressed the various PTX-induced anti-apoptotic gene products
(14). So it is expected that CUR is having the ability to sensitize the GBM cells to PTX- induced
apoptosis.
1.4 Nanotechnology for Brain Delivery
Nanotechnology involves the design of materials, devices and the delivery systems that
have the functional organization at least in one dimension on the nano scale, typically around
100 nm (12, 37, 38). In medicine and biology, nanosystems are engineered in such a way to
interface the cells and tissues at the molecular level. They can carry out many functions at a time
or in a preferred sequence and in a controlled way, an important property for the successful
delivery of drugs across anatomical and physiological barriers, including BBB (39, 40).
Significant research in finding nanotechnology approaches to cross the BBB is done in effective
delivery of anti-neoplastic drugs and other treatments for CNS therapy. This system should cause
less toxic side effects to the healthy tissues, and then cross the BBB, reach the tumor site, and
18

then be effective in killing the tumor cells. Many of the anti-neoplastic drugs have poor
physicochemical properties. PTX, for instance, is poorly soluble in water and the drug has very
low bioavailability. As such, there is a critical need to develop nanotechnology-based strategies
for effective delivery of PTX across the BBB.
Nanoparticles like liposomes, micelles and polymeric systems are extensively developed
in the recent years, which provided organ- and tissue-specific as well as intracellular delivery of
drugs and genes (41). Because of their small size and surface modification, they can cross the
barriers and penetrate through the capillaries into the cells and allow efficient accumulation of
the drug at the target site. In general, nanotechnology-based delivery systems passively or
actively target the tumor site. Passive targeting relies on the size, charge, and surface properties
of the nanocarrier, while active targeting involves attachment of a specific ligand that is
recognized by the cells at the disease site. Using passive targeting, nanocarriers can
preferentially accumulate at the tumor site due to leaky and fenestrated vasculature and poor
lymphatic drainage, which is termed as enhanced permeability and retention (EPR). The leaky
vasculature allows for accumulation of nanoparticles, whereas the poor lymphatic drainage helps
in retention at the required site (42). A number of examples of polymeric-, lipid-, and dendrimer-
based nanocarriers have utilized the EPR effect to concentrate the drug at the tumor mass and
enhance the residence time of the drug (43).
Nanotechnology based formulations also include, encapsulating the drug in the form of a
nanoemulsions (39). Edible plant-seed and fish oils containing high percentage of
polyunsaturated fatty acids (PUFA) have the potential to cross the BBB, thus mimicking the
uptake of natural fatty acids in the brain (44). Nanoemulsions made with oils rich in
polyunsaturated fatty acids (PUFA) were found to solubilize PTX efficiently in the oil droplet
19

and enhanced its oral bioavailability (12). There are two important PUFAs found abundantly in
the brain and are essential for maintaining the neurological health. They are 1) omega -3 PUFA
and the 2) omega-6 PUFA (45).
One of the proposed models for the mechanism of uptake of fatty acids (FA) explains that
the FA diffuses across the BBB and reaches brain cells, without the involvement of transporters.
They cross the luminal and the transluminal leaflets of the brain endothelium and the plasma
membrane by reversible flip-flop. Once the FA reaches the neural cells, acetyl-CoA synthetase
traps the FA by forming acetyl-CoA. This model works when the plasma membrane is
permeable to the FA, but there is also significant of evidence explaining that the transport of FA
across the BBB is derived from FA/albumin complexes (44).
Our preliminary studies have shown that nanoemulsions made with pine-nut oil and
Lipoid® E80 were found to enhance the transport across the BBB (12). Pine-nut oil is rich in
omega-3 and omega-6 PUFAs and specifically pinolenic acid and linoleic acid, respectively. Of
the total fat content in commercially-available extra virgin pine-nut oil, at least 45% of its total
unsaturated fatty acid content as pinolenic acid and 87% as linoleic acid (46). In a study reported
by Edmond (47), it was shown that linoleic acid with 18-carbon backbone and two cis-double
bonds was imported to the brain, while oleic acid containing 18 carbons and one cis-double bond
was not. This suggests exclusive selectivity of essential PUFA across the BBB.
1.5 Multifunctional Nanoemulsions for PTX/CUR Combination Therapy
Nanoemulsions are defined as thermodynamically stable dispersions of oil-in-water or
water-in-oil stabilized by an interfacial film of surfactant and co-surfactant molecules, with the
droplet size in the nanoscale ranging from 100 to 200 nm (37). This drug delivery system mainly
helps in overcoming the difficulty in delivering poorly soluble species and those that have poor
20

bioavailability, by encapsulating them in the inner phase (37, 38). The properties of
nanoemulsions are governed by the factors like the particle size and shape, surface charge,
surfactant resident time at the nanoemulsion surface and particle to particle interactions. These
factors in turn can be varied with the selection of oil phase, water phase and the surfactants. The
versatility of the nanoemulsions is also well described from the fact that the surface of the
droplet can also be modified with target-specific ligands for disease-specific localization.
Nanoemulsions are spontaneously formed when the surface tension is reduced approximately to
zero by applying mechanical energy input. Microfluidizers, ultra-sound and ultra-high shear high
pressure mixing devices are needed to provide significant energy for producing nano-scale
emulsions. Higher the energy applied results in smaller particle size of the oil droplet (37).
In the drug formulation and product optimization, another important reason for the
rejection of many new chemical entities at formulation stage itself is related to poor aqueous
solubility of the species and, thus the difficulty in delivering to systemic targets. PTX, for
instance, is formulated in an ethanolic solution containing Cremophor® EL, which is found to
cause adverse reactions upon systemic administration (48). Apart from the poor solubility, being
a P-gp substrate, PTX also has poor bioavailability and cannot cross BBB. In addition, CUR is
also a poorly soluble drug and has very low oral bioavailability. Its therapeutic effects are hence
limited to the gastrointestinal tract. In a Phase I clinical study, it was reported that patients were
administered with 8.0 grams of CUR per day orally in order to achieve detectable systemic levels
(21). Administration of CUR intravenously will enhance its potential as an anticancer agent.
Nanoemulsions can enhance the solubility of PTX and CUR by specifically dissolving in the oil
phase.
In this study, our objective was to evaluate co-administration of PTX and CUR in
21

nanoemulsion formulations. We anticipate that PTX and CUR will be effectively solubilized in
the oil phase of the emulsions and allow us to achieve intracellular availability. Furthermore, co-
administration of CUR will significantly improve PTX therapeutic effective, especially in MRP-
1 expressing T98G glioblastoma cells. Lastly, the use of PUFA-rich oil for preparation of
nanoemulsion also has an advantage in enhancing BBB transport.
22

2. OBJECTIVES AND SPECIFIC AIMS
2.1 Statement of the Problem
Glioblastoma multiforme (GBM), the most lethal form of brain tumor, affects as many as
12,000 patients in the United States and more than 200,000 world-wide every year (1). The high
mortality rate of GBM is partly due to lack of efficacious therapeutic outcomes upon systemic
chemotherapy administration. The blood-brain barrier (BBB) restricts optimum permeability and
availability of chemotherapeutic agents in the effective treatment of GBM. Nanotechnology-
based delivery systems, such as oil-in-water nanoemulsions, have been shown to enhance drug
availability in the brain upon systemic administration. As such, nanoemulsion formulations can
be very effective in therapeutic management of GBM, especially when combination therapies are
used that can augment the efficacy and improve clinical outcomes.
2.2 Objectives and Hypotheses
The main objective of this Master’s thesis project was to evaluate the potential of oil-in-
water nanoemulsion delivery system for combination therapy in GBM. The nanoemulsions in
this study are specifically prepared with oils rich in omega-3 and omega-6 polyunsaturated fatty
acids (PUFA) to enhance drug transport across the BBB. An additional objective is to evaluate
the potential of combination chemotherapeutic agent (i.e., paclitaxel) and a multidrug resistant
(MDR) modulator (i.e., curcumin). Curcumin is known to have several beneficial effects
including down-regulation of MRP-1 gene and enhancement of apoptosis in tumor cells through
inhibition of the NFκB signaling. Our experimental hypothesis is that the combination of
paclitaxel (PTX) and curcumin (CUR), when administered in nanoemulsions, will significantly
enhance therapeutic efficacy in GBM by down-regulation of drug resistance and augmentation of
cellular apoptotic response.
23

Our experimental hypotheses for this project were: (1) PUFA-rich oil-in-water
nanoemulsions can efficiently solubility PTX and CUR and allow for enhanced intracellular
delivery in GBM cells and (2) co-administration of CUR with PTX will enhance therapeutic
efficacy in GBM cells by simultaneous down-regulation of MRP-1 expression and inhibition of
NFkB signaling.
2.3 Specific Aims
The specific aims of this MS thesis project were:
Aim 1: Preparation, Characterization, and Optimization of Pine Nut Oil-Containing
Oil-in-Water Nanoemulsions:
(a) Blank, PTX- and CUR-containing nanoemulsions, as single agent and in combination,
will be prepared using pine-nut oil as the internal oil phase.
(b) The nanoemulsion formulations will be characterized for particle size, surface charge,
and oil droplet morphology.
(c) Optimization of the nanoemulsion formulations based on droplet size and stability.
Aim 2: Intracellular Delivery in U87 (wild-type) and T98G (MRP positive) GBM
Cells
(a) Establishment of U87 and T98G human glioblastoma cells in culture.
(b) Qualitative evaluations of intracellular PTX and CUR delivery with nanoemulsion
formulations by fluorescence microscopy.
Aim 3: Down-Regulation of MRP-1, Inhibition of NFκB pathway and Enhancement
in Cytotoxicity and Apoptosis in U87 (wild-type) and T98G (MRP-1 positive) GBM Cells
24

(a) Determination of baseline MRP-1 expression in U87 and T98G cells by
immunocytometry and Western blot analysis.
(b) Analysis of MRP-1 expression down-regulation and Inhibition of NFκB pathway with
CUR treatments using Western blot analyses.
(c) Cytotoxicity of single and combination PTX and CUR therapy in solution and in
nanoemulsion formulations using the MTT (formazan) assay.
(d) Qualitative apoptosis measurements by TUNEL staining.
25

3. MATERIALS AND METHODS
3.1 Preparation of Oil-in-Water Nanoemulsions
Oil-in-water nanoemulsion formulations were prepared using an ultrasonication method
that has been optimized in our laboratory (12). Briefly, the aqueous phase was prepared using 4
mL of deionized distilled water and 120 mg of egg phosphotidylcholine (Lipoid® E80, Lipoid
GmbH, Ludwigshafen, Germany), which were mixed and stirred for at least 30 minutes to insure
complete dissolution. Separately, the oil phase, consisting of 1 mL of extra virgin pine-nut oil
(Siberian Tigers Inc., Springfield, VT) was heated to 75-80oC for 2-3 minutes. Gradually, the
aqueous phase was added to the oil phase under stirring conditions. The oil droplet particle size
in the course emulsion formed was further reduced by ultrasonication at 21% amplitude and 50%
duty cycle using Vibra-Cell VC 505® (Sonics Instruments, Newtown, CT) ultrasound instrument
for 10 minutes. The stable nanoemulsions thus formed were stored in a refrigerator at
approximately 4°C for further use.
For the preparation of drug-containing nanoemulsions, stock solutions of both the drugs,
PTX and CUR, were made in dehydrated ethanol. Desired amount of PTX and CUR powders
were weighed and dissolved in ethanol. The stock solution was added to the oil phase (1 mL).
Ethanol in the oil phase was evaporated under high vacuum prior to the mixing of aqueous
phase. The mixture was then ultrasonicated to obtain the 1.0 mg/mL PTX- and CUR-containing
nanoemulsions.
3.2 Characterization of the Nanoemulsions
3.2.1 Particle Size Analysis: Blank and drug-containing nanoemulsions were
characterized for the hydrodynamic particle size and size distribution using dynamic light
scattering method with the Brookhaven Instrument’s ZetaPALS® 90Plus (Holtsville, NY). The
26

nanoemulsion sample was diluted a 1,000 times with deionized distilled water and the oil droplet
particle size was determined at 90° scattering angle and 25°C temperature. Average count rate
was maintained constant between 50-500 kcps to achieve reproducibility in the particle size
measurements. The hydrodynamic diameter of the oil droplet is determined and compared by
considering the mean effective diameter on log-normal size distribution mode.
3.2.2 Surface Charge Measurements: Blank and drug-containing nanoemulsions were
diluted with deionized distilled water and the zeta potential values were measured using
Brookhaven Instrument’s ZetaPALS® 90Plus (Holtsville, NY). The refractive index was kept at
1.33 and the viscosity at 1 cps to mimic the conditions of water. Out of the diluted nanoemulsion
(2.5 mL), small volume of approximately 1.5 mL was connected to an electrode at 4.0 volts and
2.0 Hz field frequency for surface charge determination. Electrophoretic mobility of the nano-
droplets was measured and converted to zeta potential values with a built-in software that uses
the Smoluchowski equation to calculate zeta potential values.
3.2.3. Transmission Electron Microscopy (TEM): To observe the morphology of the oil
droplets in the nanoemulsions, each batch was also characterized by TEM using a negative
staining technique. Approximately, 50 µL of the control and drug-loaded nanoemulsion
formulations were added to the 200 mesh Formwar-coated copper grids (Electron Microscopy
Science, Hatfield, PA). The samples were allowed to dry by draining off the excess liquid with
Whatman filter paper, which was placed on the edge of the copper grid. Then they were
negatively stained with 50 µL of 1.5% uranyl acetate and allowing the staining to proceed for 10
minutes at room temperature. Then the excess uranyl acetate was drained off and the copper grid
containing the nanoemulsion sample as a dry film was placed on a TEM sample holder and
27

observed with a JEOL 100-X (Peabody, MA) transmission electron microscope equipped with a
20 µm aperture and at 60 kV accelerating voltage.
3.3 Determination of Baseline MRP-1 Expression in GBM Cells
3.3.1. Cell Culture Conditions: Wild type human GBM cells (U87-MG) and MRP-1
expressing resistant human GBM cells (T98-G), purchased from American Type Culture
Collections (ATCC, Rockville, MD), were cultured in Minimum Essential Medium (MEM) at
37°C in 5.0% CO2 atmosphere. When the cells in the flask reached confluency, they were
detached with trypsin-EDTA and were then splitted into two flasks. For the immunocyto-
chemistry experiment, when the cells reached confluency, they were detached and plated into 6-
well microplates with a sterile glass coverslip at a density of 5,000 cells per well.
3.3.2 Immunocytometric Evaluation of MRP Expression: Cultured U87 and T98G cells
were grown on sterile glass cover slips overnight at 37oC. On the following day, the cover slips
were gently washed with sterile PBS. The adherent cells were fixed with 10% formalin in PBS
for 10 minutes while keeping the cells wet and the excess formalin was rinsed with subsequent
washes with sterile PBS. To avoid any non-specific binding of the immunoglobulin, the cover
slips were incubated for 30 minutes in normal goat serum blocking solution. After which, the
cover slips were incubated with primary rabbit MRP-1 antibody (Cell Signaling, Danvers, MA)
diluted in 1:100 primary antibody dilution buffer for an hour at room temperature. The cover
slips were then rinsed twice for 2 minutes each in washing buffer. To block the endogenous
peroxidase activity, the cover slips were further incubated in peroxidase blocking solution for 10
minutes and rinsed three times with the washing solution. The cells were then incubated for 30
minute at room temperature with the secondary antibody 1:200 (i.e., goat anti-rabbit antibody)
28

conjugated with horse radish peroxidase (Abcam, Inc., Cambridge, MA) and washed three times
with the washing solution. Subsequently, diaminobenzidine (DAB) solution was added and
allowed to incubate for 5-10 minutes for the peroxidase catalyzed reaction to occur resulting in
the formation of brown precipitates. The samples were observed with a bright field microscope
under 20X magnification.
3.3.3. Western Blot Analysis: In order to determine the MRP levels in U87 and T98G
glioblastoma cells, we have carried out Western blot analysis as follows.
The first step involves extraction of cytosolic proteins from U87 and T98G cells using
freshly prepared lysis buffer (i.e., 0.5 ml of 1.0 M Tris pH 7.4, 0.2 ml of 0.5 M EDTA pH 8.0,
1.5 ml of 5.0 M NaCl, 4.38 ml of 10%, Brij®-97, 0.625 ml of 10% Tween®-20 dissolved in 50 ml
of deionized distilled water) for a period of 5 minutes followed by centrifugation at 3,000 rpm
for an additional 5 minutes. The supernatant was then collected and the protein concentration
was estimated using NanoOrange® (Invitrogen, Carlsbad, CA) protein quantitation assay. A
standard curve for NanoOrange® quantitation was constructed using bovine serum albumin. The
total protein extract was then diluted 1:100 and stored at -80oC. The second step involves loading
and running the gel where 100 micrograms and 25 micrograms of the protein extract was
denatured at 90oC for 10 minutes in a water-bath, followed by 1:1 dilution with Laemmeli’s
buffer and then loaded onto pre-cast 4-15% SDS-polyacrylamide gel electrophoresis (PAGE)
gradient gel (Bio-Rad, Hercules, CA).
The gel was then run using Tris/Glycine/SDS running buffer at 125 volts for 90 minutes.
Subsequently, the protein bands on the gel were transferred onto a nitrocellulose membrane
(BioRad, Hercules, CA) in Tris/Glycine/SDS transfer buffer supplemented with 10% methanol at
25 volts for two on ice-bath. The transferred protein bands on the membrane were then blocked
29

with 3% milk in Tween®-containing Tris buffer saline (T-BST) for 45 minutes. The
nitrocellulose membrane was incubated overnight with agitation at 4oC with 1:1,000 dilution of
the primary mouse MRP-1 specific monoclonal antibody in milk. Following morning, the milk
was decanted and the membrane was washed twice with water and then incubated with 1:2000
dilution of the secondary anti-mouse HRP-conjugated IgG in T-BST for 1 hour at room
temperature. After rinsing the excess secondary antibody with T-BST and water, the reagent was
added that is cleaved by perodixase to give a chemiluminescent product. The chemiluminescent
bands were then visualized with a Kodak imager (Carestream Health, Rochester, NY).
3.4 Fluorescence Microscopic Studies of Intracellular Delivery with Nanoemulsions
3.4.1 Preparation of Fluorescently-Labeled Nanoemulsions: PTX was replaced by
rhodamine-labeled paclitaxel conjugate (rhodamine-PTX) (at 0.1% (w/w)) and was added to the
oil phase. The aqueous phase was added to the oil phase and ultrasonicated, as mentioned above,
to get the fluorescently labeled formulation. CUR is intrinsically fluorescent in the visible green
region (21) and therefore, was directly added to the oil phase and the nanoemulsions were
formulated as previously described with the final CUR concentration to 1 mg/mL.
3.4.2 Fluorescence Microscopy Studies: Nanoemulsion containing rhodamine-PTX and
CUR were diluted with MEM and aliquots were added to the U87 and T98G cells and were
compared with the fluorescently labeled solutions of PTX and CUR respectively. At regular time
intervals, the medium was removed and the cells were washed with fresh medium and then with
sterile PBS. After the final wash, the coverslips were mounted on glass slides with Fluromount-
G® mounting medium. Differential interference contrast (DIC) and epi-fluorescent images were
30

acquired at 2 0X magnifications using fluorescence microscope and the digital images were
processed using the Adobe Photoshop® software.
3.5 Determination of MRP1 Down-Regulation with CUR by Western Blot Analysis
The U87and T98G cells were treated with 20 µM CUR in the form of solutions and
Nanoemulsion formulation and incubated for 24 hours at 37ºC. Following treatments, the total
protein was extracted and the protein content was measured with NanoOrange® assay. Thirty-
micrograms of extracted protein was mixed in a 1:1 ratio with Laemmeli’s buffer and then
loaded onto a pre-cast 4-15% SDS-polyacrylamide gel electrophoresis (PAGE) gradient gel. The
gel was then run for 90 minutes at 125 V and the protein bands on the gel were transferred onto a
nitrocellulose membrane using transfer buffer supplemented with 10% methanol at 25 volts for
two-and-half hours on ice-bath. The transferred protein bands on the membrane were then
blocked with 3% milk in Tween®-containing Tris buffer saline (T-BST) for 45 minutes and then
incubated overnight with agitation at 4oC with 1:1,000 dilution of the primary mouse monoclonal
MRP-1 antibody in milk. The next day, milk was decanted and the membrane was washed twice
with water and then incubated with 1:2,000 dilution of the secondary anti-mouse horse radish
perodixase-conjugated IgG in T-BST for 1 hour at room temperature. After rinsing the excess
secondary antibody with T-BST and water, the ECL reagent was added, which was cleaved by
the horseradish perodixase enzyme to give a chemiluminescent product. The chemiluminescent
bands were visualized using a Kodak imager.
3.6 Determination of NFκB Pathway Inhibition with CUR by Western Blot Analysis
U87and T98G cells were treated with 20µM CUR in the form of solutions and
Nanoemulsion formulation and incubated for 24 hours at 37ºC. Following treatments, the total
31

protein was extracted and the protein content was measured with NanoOrange® assay. Thirty
micrograms of extracted protein was mixed in a 1:1 ratio with Laemmeli’s buffer and then
loaded onto a pre-cast 4-15% sodium dodecylsulfate-polyacrylamide gel electrophoresis (SDS-
PAGE) gradient gel. The gel was then run for 90 minutes at 125 V and the protein bands on the
gel were transferred onto a nitrocellulose membrane using transfer buffer supplemented with
10% methanol at 25 volts for two-and-half hours on ice-bath. The transferred protein bands on
the membrane were then blocked with 3% milk in Tween®-containing Tris buffer saline (T-BST)
for 45 minutes and then incubated overnight with agitation at 4oC with 1:1,000 dilution of the
primary rabbit NFκB p65 (Cell Signaling, Danvers, MA) antibody in milk. The next day, milk
was decanted and the membrane was washed twice with water and then incubated with 1:2,000
dilution of the secondary anti-rabbit horse radish perodixase-conjugated IgG in T-BST for 1 hour
at room temperature. After rinsing the excess secondary antibody with T-BST and water, the
ECL reagent was added, which was cleaved by the horse radish perodixase enzyme to give a
chemiluminescent product. The chemiluminescent bands were visualized using a Kodak imager.
The entire procedure was repeated for Primary antibody incubation with rabbit polyclonal
antibody against IκBα followed by secondary antibody incubation.
In addition, to visualize the control beta-actin bands, the nitrocellulose membrane with
proteins was cut into two halves and the upper half was incubated with primary antibody against
NFκB and the lower second half was then incubated overnight with primary antibody against
beta-actin. Following these treatments, the secondary antibody was added and the bands were
visualized as described above.
32

3.7 Cytotoxicity of Single and Combination Treatments in GBM Cells
The cytotoxicity studies were performed with both solutions and the nanoemulsion
formulations containing different concentrations of PTX, CUR and combination of PTX and
CUR. Solutions were prepared by dissolving the PTX, CUR in DMSO and then adding them to
culture medium to obtain the desired concentrations of PTX (1 nM, 5 nM, 10 nM, 50 nM, 100
nM, and 500 nM) and CUR (5 µM, 10 µM, and 20 µM) respectively and maintaining the DMSO
at 0.1% concentration in the well. Similarly nanoemulsions were also prepared at the above
mentioned concentrations of both PTX and CUR in MEM. The U87 cells and resistant T98G
cells were allowed to adhere on the surface of 96-well microplates at a density of 5,000 cells per
well. After 48 hours, when the cells in the wells were approximately confluent, the culture media
was replaced with the solutions and nanoemulsions of PTX and CUR separately and also in
combinations. After dosing, the U87 cells and T98G cells were incubated at 37°C for 3 days.
Since PTX is a cell cycle specific drug, the 3-day time period was used to insure that all cells in
the microplates can succumb to the drug effect. Cells treated with MEM containing 0.1% DMSO
and also cells treated with MEM containing Blank nanoemulsion formulation alone was used as
negative controls in determining the cytotoxicity of solutions and nanoemulsion formulations,
respectively, and those treated with 250 µg/mL of poly(ethyleneimine) (Mol. Wt. 10 kDa), a
cytotoxic cationic polymer, were used as positive control in both the cases.
For the cytotoxicity study, the MTT reagent was prepared at a concentration of 1 mg/mL
by dissolving the yellow dye (3-(4, 5-dimethylthiazol-2-yl)-2,5-diphenyltetrazolium bromide, a
tetrazole dye) in MEM. Following a 3-day incubation period, the media from each well was
replaced with 50µL of freshly prepared MTT reagent into each well and again incubated for 2
hours in the CO2 chamber at 37°C. Then 150 µL DMSO was added into each well to dissolve
33

the insoluble purple-colored formazan crystals formed in the mitochondria of living cells. The
absorbance in each well was measured using a BioTek-HT (Winooski, VT)
UV/Visible/fluorescence microplate reader at 570 nm. The percentage cell viability was
calculated at each concentration of PTX, CUR and at each combination of PTX and CUR by
dividing the absorbance of the treated cells over the control cells and multiplying it by one
hundred. Then IC50 was found for the solutions and the nanoemulsions on both U87 cells and
T98G cells with the help of bar charts plotted using Microsoft Excel® and Graph-Pad Prism®
softwares. The IC50 was also calculated for the PTX and CUR combinations on both wild-type
and MRP-1 expressing glioblastoma cell lines.
3.8 Qualitative Apoptosis Studies by TUNEL Staining
The TUNEL assay (terminal dUTP nick-end labeling) is an assay used to detect apoptosis
in-situ by measuring nuclear DNA fragmentation, a biochemical indicator of apoptosis at the
single-cell level. This system, end-labels fragmented DNA with biotinylated nucleotide at the
3’OH ends using the terminal deoxynucleotidyl transferase, recombinant (rTdT) enzyme.
Horseradish peroxidase-conjugated streptavidin (streptavidin-HRP) bound to the biotinylated
nucleotide was detected using the peroxidase substrate, hydrogen peroxide and the chromogen
diaminobenzidine, which stains apoptotic nuclei dark brown. A DeadEndTM Colorimetric
TUNEL assay system kit (Promega Corporation, Madison, WI) was used for this assay.
Both U87 and T98G glioma cells grown on Lab-Tek® chamber slides were incubated
with solutions and Nanoemulsions of both 20 µM CUR, 100 nM PTX and combination of 20
µM CUR and 100 nM PTX for 24 hours at 37oC and 5% CO2 to induce apoptosis. The cells were
then washed twice with PBS and were fixed by immersing them in 10% buffered formalin for 25
minutes at room temperature followed by immersing twice in PBS for 5 minutes each at room
34

temperature. The cells were then permeabilized by immersing in 0.2% Triton® X-100 solution in
PBS for 5 minutes at RT. The cells were again immersed twice in PBS for 5 minutes each at RT.
The cells were then equilibrated for 5-10 minutes at RT with 100 µL of equilibrium buffer. One
hundred-μL of rTdT reaction mix per slide was prepared using 98 μl of equilibration buffer + 1
μL of biotinylated nucleotide mix + 1 μL of rTdT enzyme. One hundred-μL of the rTdT reaction
mix was then added to the previously equilibrated sections on the slide without allowing the
sections to dry. Plastic coverslips were used to cover the sections to ensure even distribution. For
the end-labeling reaction to occur the coverslips were then incubated at 37oC, 5% CO2 for 60
minutes. The reaction was then terminated by removing the coverslips and immersing the slides
is 2X sodium chloride-sodium citrate buffer (SSC) buffer for 15 minutes at RT. The slides were
then washed twice by immersing in PBS for 5 minutes at room temperature. The slides were
again immersed in 0.3% hydrogen peroxide solution in PBS at room temperature to block
endogenous peroxidases and were then washed twice by immersing in PBS for 5 minutes at
room temperature. The streptavidin-HRP provided is diluted 1:500 in PBS and 100 μL of it was
added to each slide followed by 30 minute incubation at RT. The slides were then washed twice
by immersing in PBS for 5 minutes at room temperature. DAB solution was prepared just before
use and 100 μL of it was added to the sections until a light brown color develops. The sections
were then rinsed several times in deionized water. The slides were then mounted in permanent
mounting medium and observed under a light microscope.
35
4. RESULTS AND DISCUSSIONS
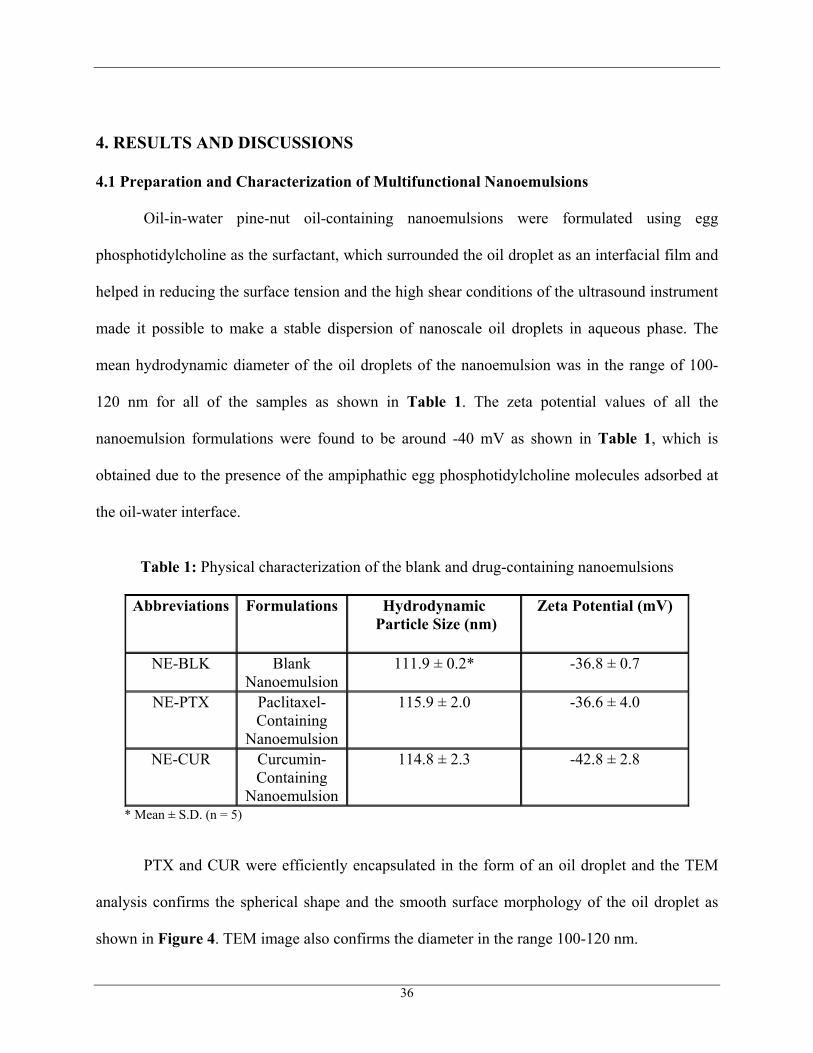
4.1 Preparation and Characterization of Multifunctional Nanoemulsions
Oil-in-water pine-nut oil-containing nanoemulsions were formulated using egg
phosphotidylcholine as the surfactant, which surrounded the oil droplet as an interfacial film and
helped in reducing the surface tension and the high shear conditions of the ultrasound instrument
made it possible to make a stable dispersion of nanoscale oil droplets in aqueous phase. The
mean hydrodynamic diameter of the oil droplets of the nanoemulsion was in the range of 100-
120 nm for all of the samples as shown in Table 1. The zeta potential values of all the
nanoemulsion formulations were found to be around -40 mV as shown in Table 1, which is
obtained due to the presence of the ampiphathic egg phosphotidylcholine molecules adsorbed at
the oil-water interface.
Table 1: Physical characterization of the blank and drug-containing nanoemulsions
Abbreviations Formulations Hydrodynamic Particle Size (nm)
Zeta Potential (mV)
NE-BLK Blank Nanoemulsion
111.9 ± 0.2*
-36.8 ± 0.7
NE-PTX Paclitaxel- Containing
Nanoemulsion
115.9 ± 2.0
-36.6 ± 4.0
NE-CUR Curcumin- Containing
Nanoemulsion
114.8 ± 2.3
-42.8 ± 2.8
* Mean ± S.D. (n = 5)
PTX and CUR were efficiently encapsulated in the form of an oil droplet and the TEM
analysis confirms the spherical shape and the smooth surface morphology of the oil droplet as
shown in Figure 4. TEM image also confirms the diameter in the range 100-120 nm.
36

Figure 4: Transmission electron micrograph of pine-nut oil nanoemulsion for the co-administration of paclitaxel and curcumin
4.2 Determination of Baseline MRP-1 Expression in GBM Cells
4.2.1 Immunocytometric Evaluation of MRP-1 expression: Qualitative baseline
expression of MRP-1 in U87 and T98G human glioblastoma cells was examined by
Immunocytometric analsyis. U87 and T98G cells which are not treated with anti-body against
MRP-1, but treated with DAB stain are used as control. U87 cells and T98G cells were treated
with anti-MRP-1 primary antibody and HRP-linked secondary antibody and were then treated
with DAB stain. The peroxide catalyzed reaction that occurs following the addition of DAB
results in the formation of a brown precipitate on the cell surface and within the cell, indicating
the localization of MRP-1 in U87 and T98G cells as shown in Figure 5. Dark brown precipitate
observed on T98G cells as shown in Figure 5 confirms the over-expression of MRP-1 in
37

accordance with the Western blot results as shown in Figure 6. Controls cells did not show any
brown precipitates indicating the absence of non-specific binding of primary antibody.
A B
C D
A B
C D
Figure 5. Immunocytometric analysis of MRP-1 expression in U87 and T98G glioblastoma cells. The figures represent T98G cells (A&B) and U87 cells (C&D). A&C are images of unstained
control cells, while B&D represent cells that were treated with MRP-1 antibody and stained with diaminobenzidine
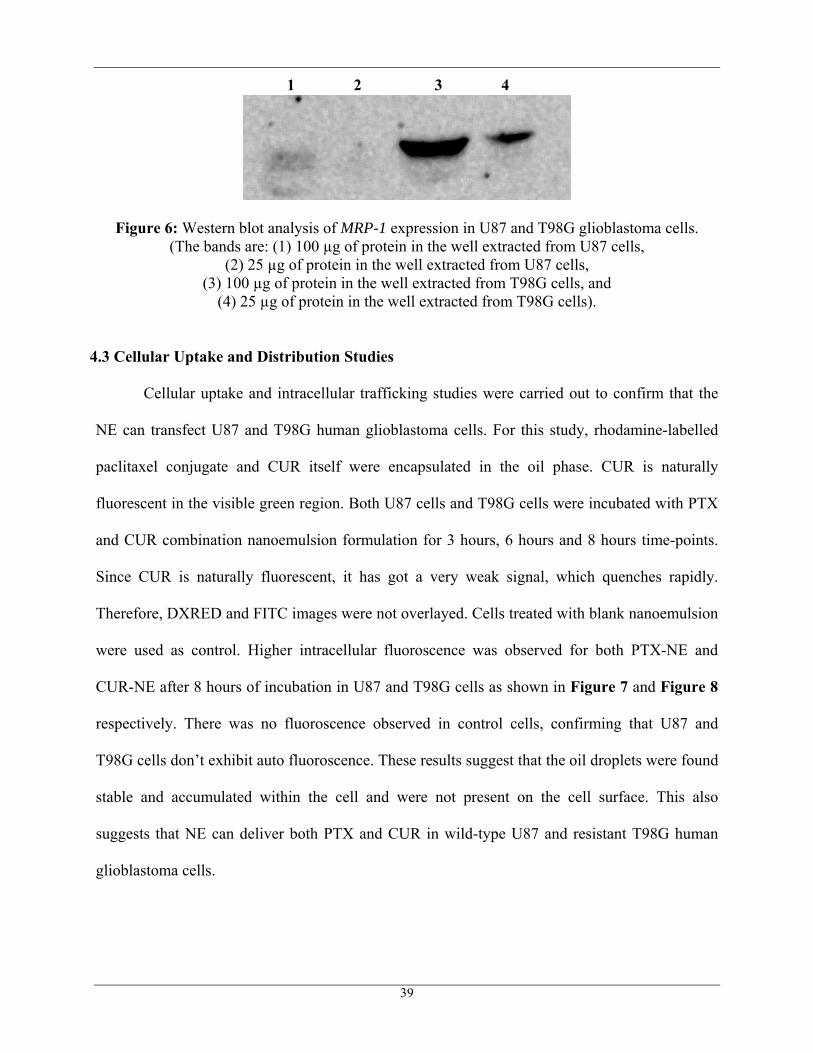
4.2.2 Western Blot Analysis: The main purpose of this study is to further confirm the
over-expression of MRP-1 in resistant T98G cells when compared to that of wild-type U87
human glioblastoma cells. As shown in Figure 6, MRP-1 expression was observed in both U87
(lane 1) and T98G (lane 3), but there was far higher level of expression in T98G cells as
compared to U87 cells. Hence the results of Western blot analysis, confirms that T98G cell line
would serve as a best model to study the down regulation of MRP-1 by CUR.
38
39
Figure 6: Western blot analysis of MRP-1 expression in U87 and T98G glioblastoma cells. (The bands are: (1) 100 µg of protein in the well extracted from U87 cells,
(2) 25 µg of protein in the well extracted from U87 cells, (3) 100 µg of protein in the well extracted from T98G cells, and
(4) 25 µg of protein in the well extracted from T98G cells).
4.3 Cellular Uptake and Distribution Studies
Cellular uptake and intracellular trafficking studies were carried out to confirm that the
NE can transfect U87 and T98G human glioblastoma cells. For this study, rhodamine-labelled
paclitaxel conjugate and CUR itself were encapsulated in the oil phase. CUR is naturally
fluorescent in the visible green region. Both U87 cells and T98G cells were incubated with PTX
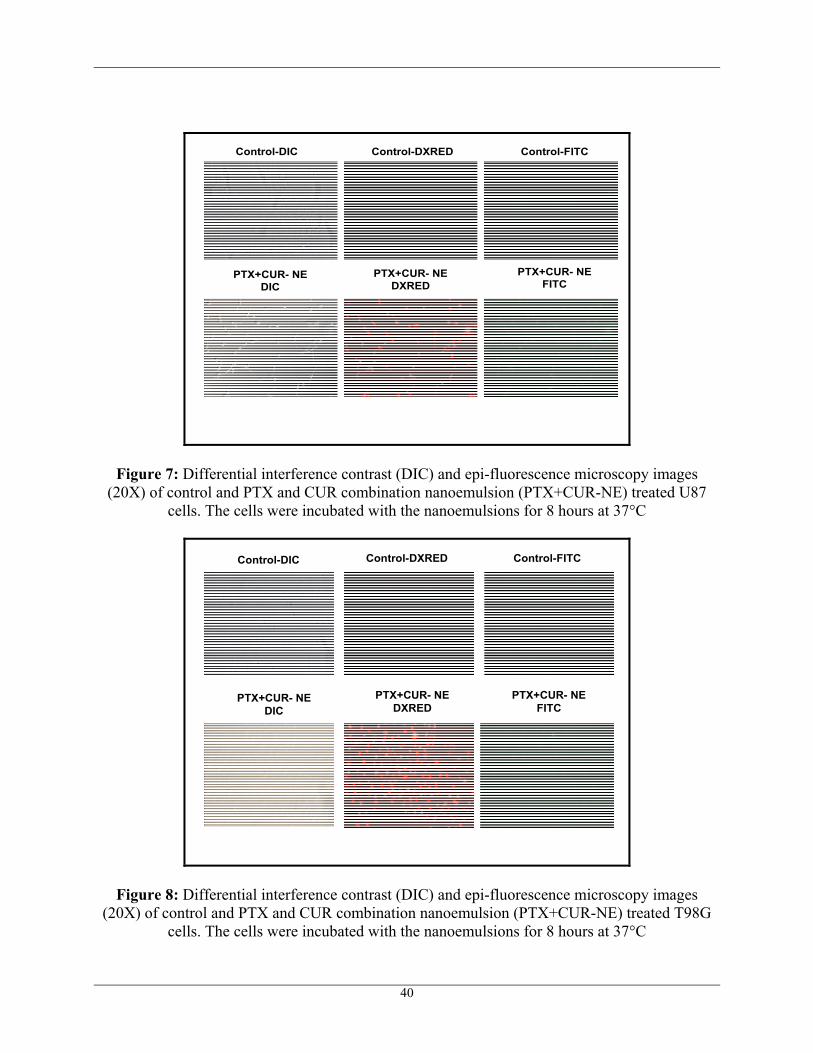
and CUR combination nanoemulsion formulation for 3 hours, 6 hours and 8 hours time-points.
Since CUR is naturally fluorescent, it has got a very weak signal, which quenches rapidly.
Therefore, DXRED and FITC images were not overlayed. Cells treated with blank nanoemulsion
were used as control. Higher intracellular fluoroscence was observed for both PTX-NE and
CUR-NE after 8 hours of incubation in U87 and T98G cells as shown in Figure 7 and Figure 8
respectively. There was no fluoroscence observed in control cells, confirming that U87 and
T98G cells don’t exhibit auto fluoroscence. These results suggest that the oil droplets were found
stable and accumulated within the cell and were not present on the cell surface. This also
suggests that NE can deliver both PTX and CUR in wild-type U87 and resistant T98G human
glioblastoma cells.
1 2 3 4
Control-DIC Control-DXRED Control-FITC
PTX+CUR- NEDIC
PTX+CUR- NEDXRED
PTX+CUR- NEFITC
Figure 7: Differential interference contrast (DIC) and epi-fluorescence microscopy images (20X) of control and PTX and CUR combination nanoemulsion (PTX+CUR-NE) treated U87
cells. The cells were incubated with the nanoemulsions for 8 hours at 37°C
Control-DIC Control-DXRED Control-FITC
PTX+CUR- NEDIC
PTX+CUR- NEDXRED
PTX+CUR- NEFITC
Figure 8: Differential interference contrast (DIC) and epi-fluorescence microscopy images (20X) of control and PTX and CUR combination nanoemulsion (PTX+CUR-NE) treated T98G
cells. The cells were incubated with the nanoemulsions for 8 hours at 37°C
40

4.4 Down Regulation of MRP-1 and Inhibition of NFκB Pathway with CUR
There was literature evidence suggesting that Curcumin and its glutathione conjugates
have the potential to inhibit MRP-1 (36). From the above results, it was also shown that U87 and
T98G cell line can serve as a best model to study MRP-1 down-regulation. Both U87 and T98G
cells were treated with CUR solution and CUR nanoemulsions and the cytosolic proteins were
extracted from both treated and untreated cells (control). The chemiluminiscent bands observed
for the MRP-1 were shown in Figure 9. Lanes 1, 2 and 3 indicates protein extracted from U87
cells. Lanes 4, 5 and 6 indicates protein extracted from T98G cells. Lane 1 and 4 indicate protein
extracted from untreated cells. Lanes 2 and 5 indicate protein extracted from CUR solution
(CUR-S) treated cells. Lanes 3 and 6 indicate protein extracted from CUR nanoemulsion (CUR-
NE) treated cells. MRP-1 bands were observed at 190 kDa. Bands in lane 2 and 3 were less
intense when compared to that of lane 1 and band in lane 6 was less intense when compared to
that of lane 4 and 5 suggesting the MRP-1 down-regulation in CUR treated U87 and T98G cells.
Bands in lane 3 & 6 were less intense when compared to that of 2 and 5 indicating that,
nanoemulsion formulation was much more efficient in MRP-1 down-regulation.
For studying the inhibition of NFκB pathway, two different antibodies were choosen.
One antibody was against NFκB p65 domain. Most abundantly occurring NFκB dimers in
cytoplasm contain p65 domain. For certain stimuli, IKK gets activated in a tumor cell, which
further phosphorylates and degrades IκBα and thus NFκB translocates into the nucleus and
initiates the transcription of several genes responsible for tumor cell survival. CUR inhibits
NFκB pathway by inhibiting IKK. Therefore high intense NFκB bands at 64 kDa were observed
in CUR treated U87 cells when compared to that of untreated U87 cells as shown in Figure 9.
There was not such inhibtion observed in T98G cells treated with CUR. This result confirms the
41
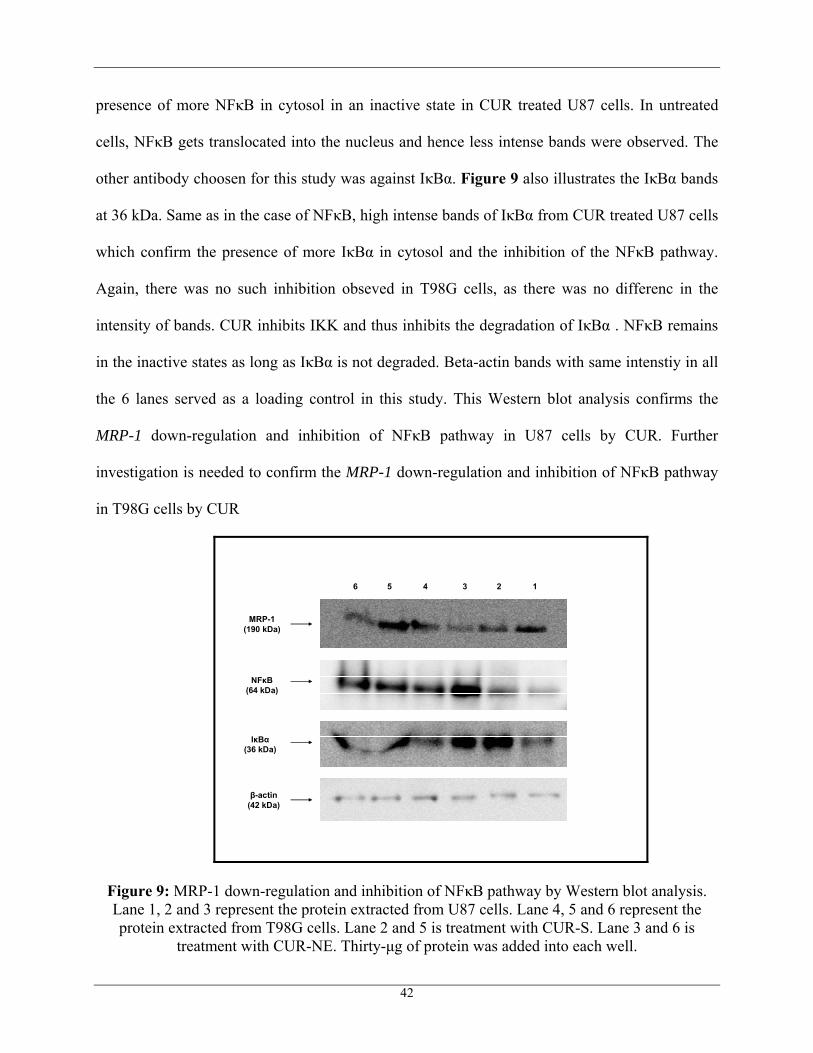
presence of more NFκB in cytosol in an inactive state in CUR treated U87 cells. In untreated
cells, NFκB gets translocated into the nucleus and hence less intense bands were observed. The
other antibody choosen for this study was against IκBα. Figure 9 also illustrates the IκBα bands
at 36 kDa. Same as in the case of NFκB, high intense bands of IκBα from CUR treated U87 cells
which confirm the presence of more IκBα in cytosol and the inhibition of the NFκB pathway.
Again, there was no such inhibition obseved in T98G cells, as there was no differenc in the
intensity of bands. CUR inhibits IKK and thus inhibits the degradation of IκBα . NFκB remains
in the inactive states as long as IκBα is not degraded. Beta-actin bands with same intenstiy in all
the 6 lanes served as a loading control in this study. This Western blot analysis confirms the
MRP-1 down-regulation and inhibition of NFκB pathway in U87 cells by CUR. Further
investigation is needed to confirm the MRP-1 down-regulation and inhibition of NFκB pathway
in T98G cells by CUR
NFκB(64 kDa)
IκBα(36 kDa)
β-actin(42 kDa)
6 5 4 3 2 1
MRP-1(190 kDa)
Figure 9: MRP-1 down-regulation and inhibition of NFκB pathway by Western blot analysis. Lane 1, 2 and 3 represent the protein extracted from U87 cells. Lane 4, 5 and 6 represent the protein extracted from T98G cells. Lane 2 and 5 is treatment with CUR-S. Lane 3 and 6 is
treatment with CUR-NE. Thirty-μg of protein was added into each well.
42

4.5 Cytotoxicity of Single and Combination Treatments in GBM Cells
The cytotoxicity experiments were carried out with both solutions and nanoemulsion
formulations. IC50 values were determined and the efficiency of nanoemulsions over solutions
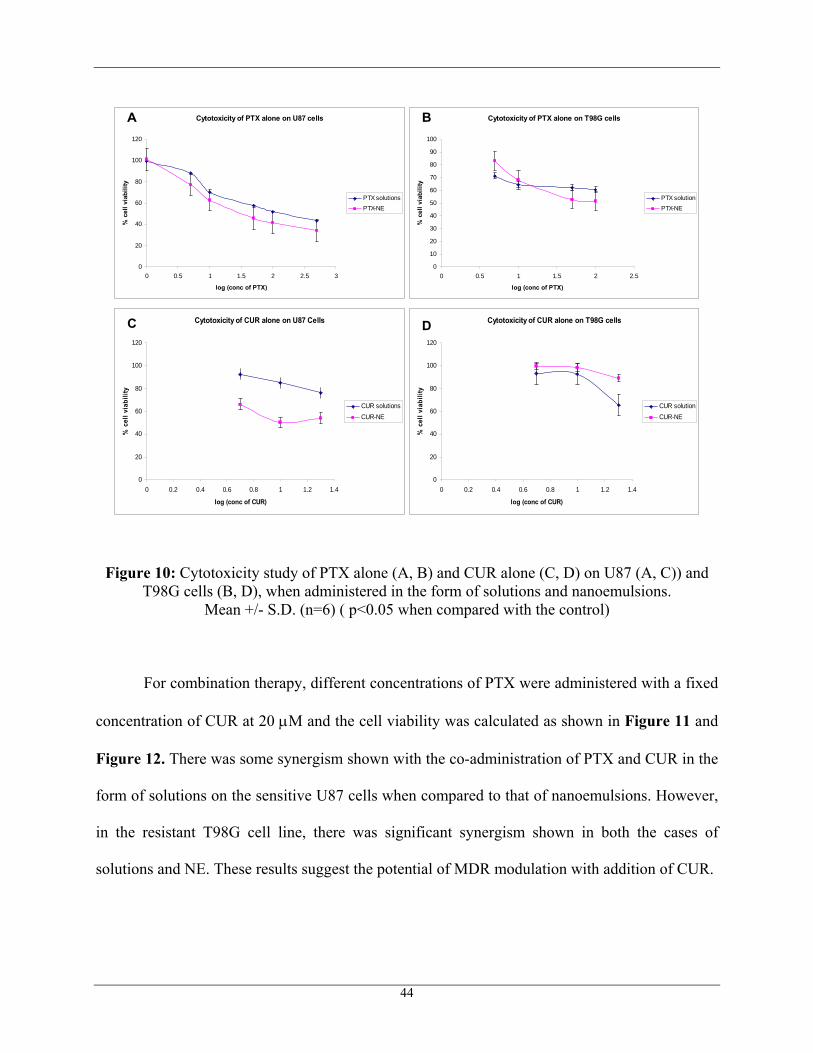
and the single therapy over co-therapy was evaluated. From semi-logarthmic plots as shown in
Figure 10, it was clearly observed that the IC50 of PTX solution alone when administered on
U87 cells was found approximately between 100 nM and 500 nM, whereas the PTX-NE (PTX
nanoemulsion) was found approximately between 10 nM and 50 nM showing a 10-fold increase
in the cytotoxicity with NE when compared to that of solutions. Figure 10 also showed that there
was an increase in cell cytotoxicity with NE (IC50 between 50 nM and 100 nM) on T98G cells
when compared to that of the solution (IC50 above 100 nM). No significant change in cell
viability at three different concentrations 10 nM, 50 nM and 100 nM (around 60% cell viability)
was probably due to the resistance shown by MRP-1 to PTX. At low concentrations of CUR,
there was no significant cell death as shown in Figure 10. CUR was found to have some
cytotoxicity in the sensitive U87 cell line, when administered in the form of NE.
43
Cytotoxicity of PTX alone on U87 cells
0
20
40
60
80
100
120
0 0.5 1 1.5 2 2.5 3
log (conc of PTX)
% c
ell v
iabi
lity
PTX solutionsPTX-NE
Cytotoxicity of PTX alone on T98G cells
0
10
20
30
40
50
60
70
80
90
100
0 0.5 1 1.5 2 2.5
log (conc of PTX)
% c
ell v
iabi
lity
PTX solutionPTX-NE
Cytotoxicity of CUR alone on U87 Cells
0
20
40
60
80
100
120
0 0.2 0.4 0.6 0.8 1 1.2 1.4
log (conc of CUR)
% c
ell v
iabi
lity
CUR solutionsCUR-NE
Cytotoxicity of CUR alone on T98G cells
0
20
40
60
80
100
120
0 0.2 0.4 0.6 0.8 1 1.2 1.4
log (conc of CUR)
% c
ell v
iabi
lity
CUR solutionCUR-NE
A B
C D
Figure 10: Cytotoxicity study of PTX alone (A, B) and CUR alone (C, D) on U87 (A, C)) and T98G cells (B, D), when administered in the form of solutions and nanoemulsions.
Mean +/- S.D. (n=6) ( p<0.05 when compared with the control)
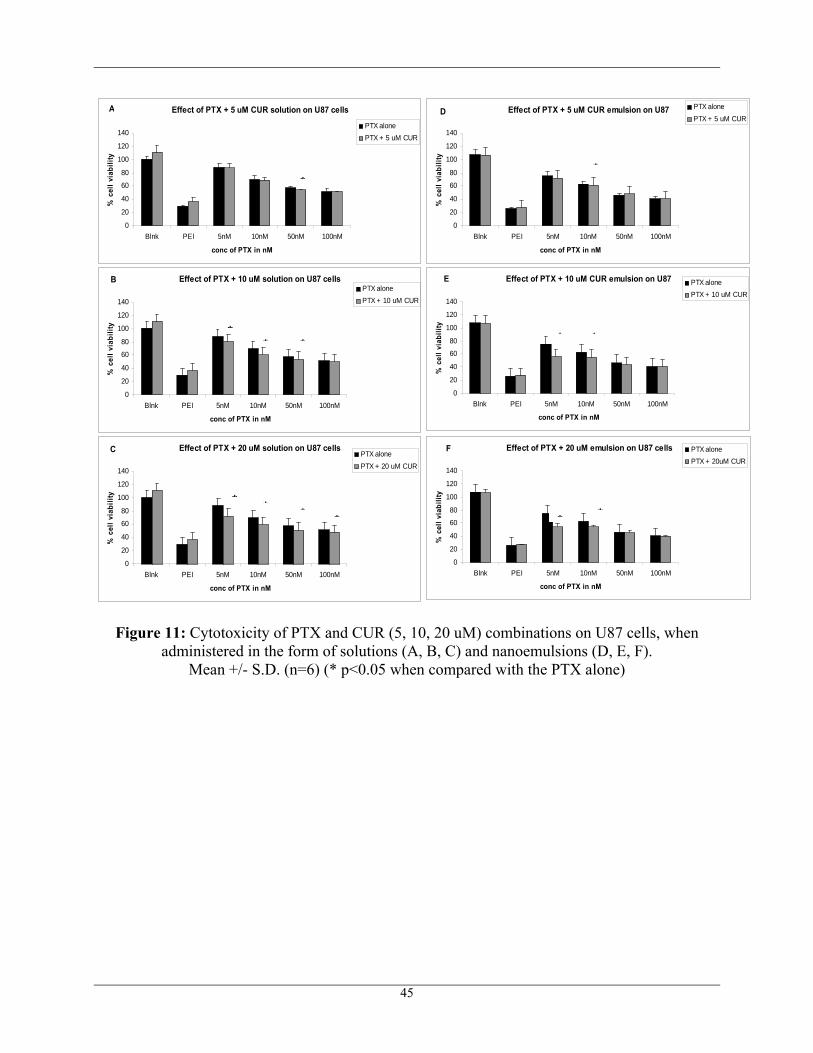
For combination therapy, different concentrations of PTX were administered with a fixed
concentration of CUR at 20 μM and the cell viability was calculated as shown in Figure 11 and
Figure 12. There was some synergism shown with the co-administration of PTX and CUR in the
form of solutions on the sensitive U87 cells when compared to that of nanoemulsions. However,
in the resistant T98G cell line, there was significant synergism shown in both the cases of
solutions and NE. These results suggest the potential of MDR modulation with addition of CUR.
44
45
Figure 11: Cytotoxicity of PTX and CUR (5, 10, 20 uM) combinations on U87 cells, when administered in the form of solutions (A, B, C) and nanoemulsions (D, E, F).
Mean +/- S.D. (n=6) (* p<0.05 when compared with the PTX alone)
Effect of PTX + 5 uM CUR solution on U87 cells
0
20
40
60
80
100
120
140
Blnk PEI 5nM 10nM 50nM 100nM
conc of PTX in nM
% c
ell v
iabi
lity
PTX alonePTX + 5 uM CUR
A
Effect of PTX + 10 uM solution on U87 cells
0
20
40
60
80
100
120
140
Blnk PEI 5nM 10nM 50nM 100nM
conc of PTX in nM
% c
ell v
iabi
lity
PTX alonePTX + 10 uM CUR
B
Effect of PTX + 20 uM solution on U87 cells
0
20
40
60
80
100
120
140
Blnk PEI 5nM 10nM 50nM 100nM
conc of PTX in nM
% c
ell v
iabi
lity
PTX alonePTX + 20 uM CUR
C
Effect of PTX + 5 uM CUR emulsion on U87
0
20
40
60
80
100
120
140
Blnk PEI 5nM 10nM 50nM 100nM
conc of PTX in nM
% c
ell v
iabi
lity
PTX alonePTX + 5 uM CUR
D
*
** *
* * * *
*
Effect of PTX + 10 uM CUR emulsion on U87
0
20
40
60
80
100
120
140
Blnk PEI 5nM 10nM 50nM 100nM
conc of PTX in nM
% c
ell v
iabi
lity
PTX alonePTX + 10 uM CUR
E
* *
Effect of PTX + 20 uM emulsion on U87 cells
0
20
40
60
80
100
120
140
Blnk PEI 5nM 10nM 50nM 100nM
conc of PTX in nM
% c
ell v
iabi
lity
PTX alonePTX + 20uM CUR
F
* *
46
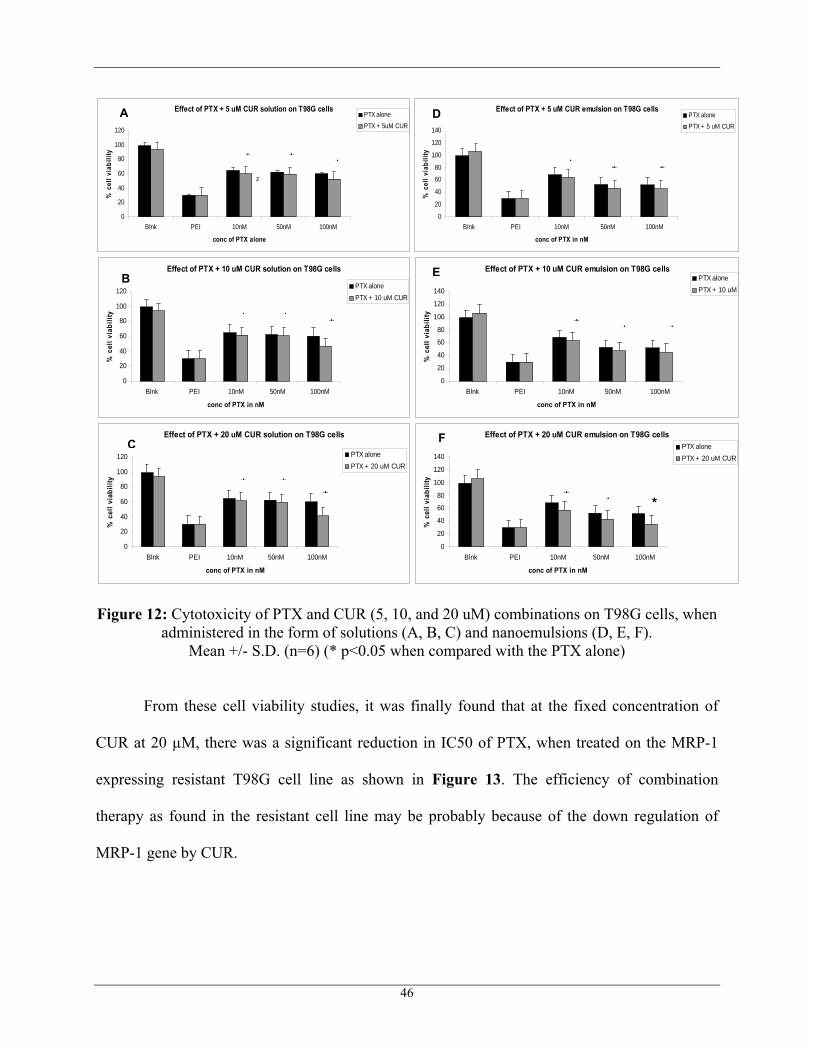
Figure 12: Cytotoxicity of PTX and CUR (5, 10, and 20 uM) combinations on T98G cells, when administered in the form of solutions (A, B, C) and nanoemulsions (D, E, F).
Mean +/- S.D. (n=6) (* p<0.05 when compared with the PTX alone)
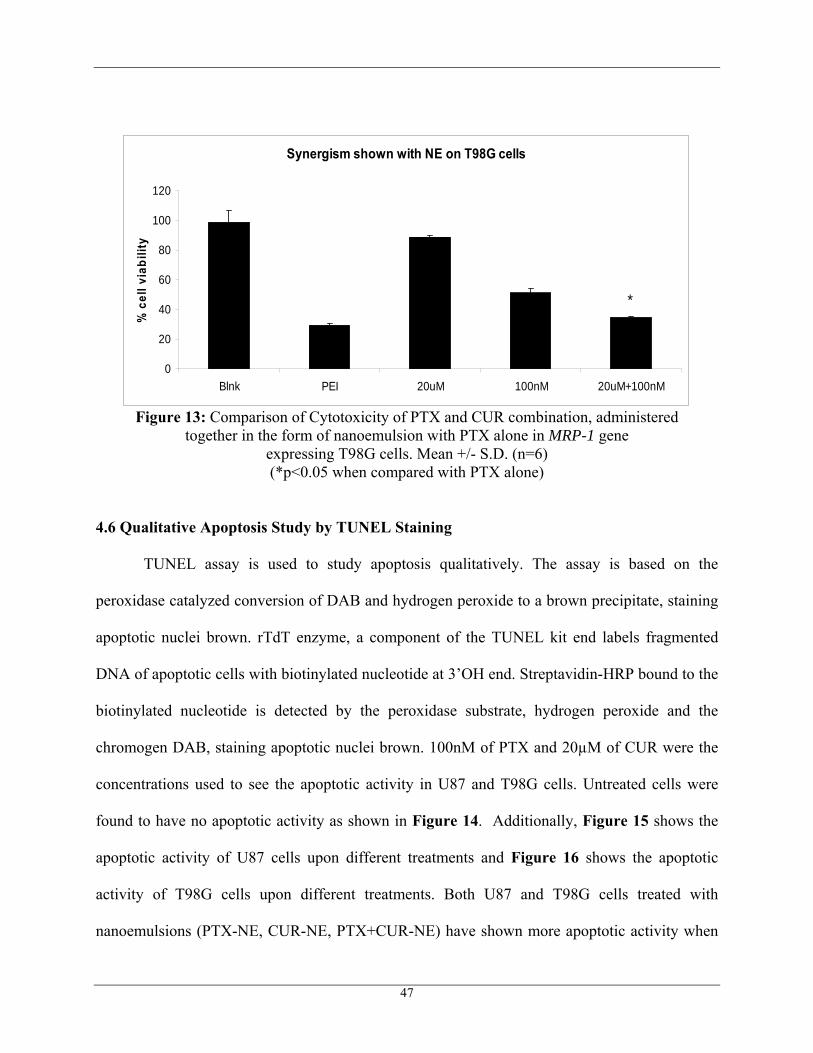
From these cell viability studies, it was finally found that at the fixed concentration of
CUR at 20 µM, there was a significant reduction in IC50 of PTX, when treated on the MRP-1
expressing resistant T98G cell line as shown in Figure 13. The efficiency of combination
therapy as found in the resistant cell line may be probably because of the down regulation of
MRP-1 gene by CUR.
Effect of PTX + 5 uM CUR solution on T98G cells
0
20
40
60
80
100
120
Blnk PEI 10nM 50nM 100nM
conc of PTX alone
% c
ell v
iabi
lity
PTX alonePTX + 5uM CUR
z
Effect of PTX + 10 uM CUR solution on T98G cells
0
20
40
60
80
100
12
Blnk PEI 10nM 50nM 100nM
conc of PTX in nM
% c
ell v
iabi
lity
0PTX alonePTX + 10 uM CUR
Effect of PTX + 20 uM CUR solution on T98G cells
0
20
40
60
80
100
120
Blnk PEI 10nM 50nM 100nM
conc of PTX in nM
% c
ell v
iabi
lity
PTX alonePTX + 20 uM CUR
Effect of PTX + 5 uM CUR emulsion on T98G cells
0
20
40
60
80
100
120
140
Blnk PEI 10nM 50nM 100nM
conc of PTX in nM
% c
ell v
iabi
lity
PTX alonePTX + 5 uM CUR
Effect of PTX + 10 uM CUR emulsion on T98G cells
0
20
40
60
80
100
120
140
Blnk PEI 10nM 50nM 100nM
conc of PTX in nM
% c
ell v
iabi
lity
PTX alonePTX + 10 uM
Effect of PTX + 20 uM CUR emulsion on T98G cells
0
20
40
60
80
100
120
140
Blnk PEI 10nM 50nM 100nM
conc of PTX in nM
% c
ell v
iabi
lity
PTX alonePTX + 20 uM CUR
A
* * *
B
* * *
C
* **
D
E
F
* * *
* * *
* * *
Synergism shown with NE on T98G cells
0
20
40
60
80
100
120
Blnk PEI 20uM 100nM 20uM+100nM
% c
ell v
iabi
lity
*
Figure 13: Comparison of Cytotoxicity of PTX and CUR combination, administered together in the form of nanoemulsion with PTX alone in MRP-1 gene
expressing T98G cells. Mean +/- S.D. (n=6) (*p<0.05 when compared with PTX alone)
4.6 Qualitative Apoptosis Study by TUNEL Staining
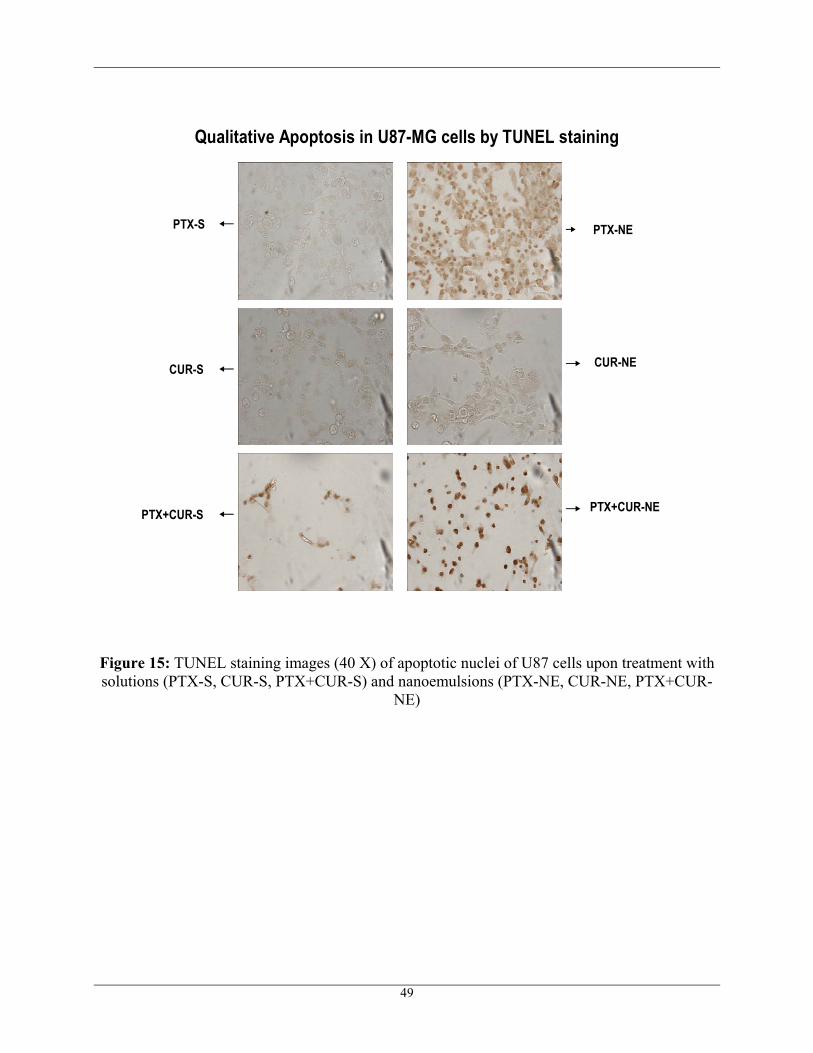
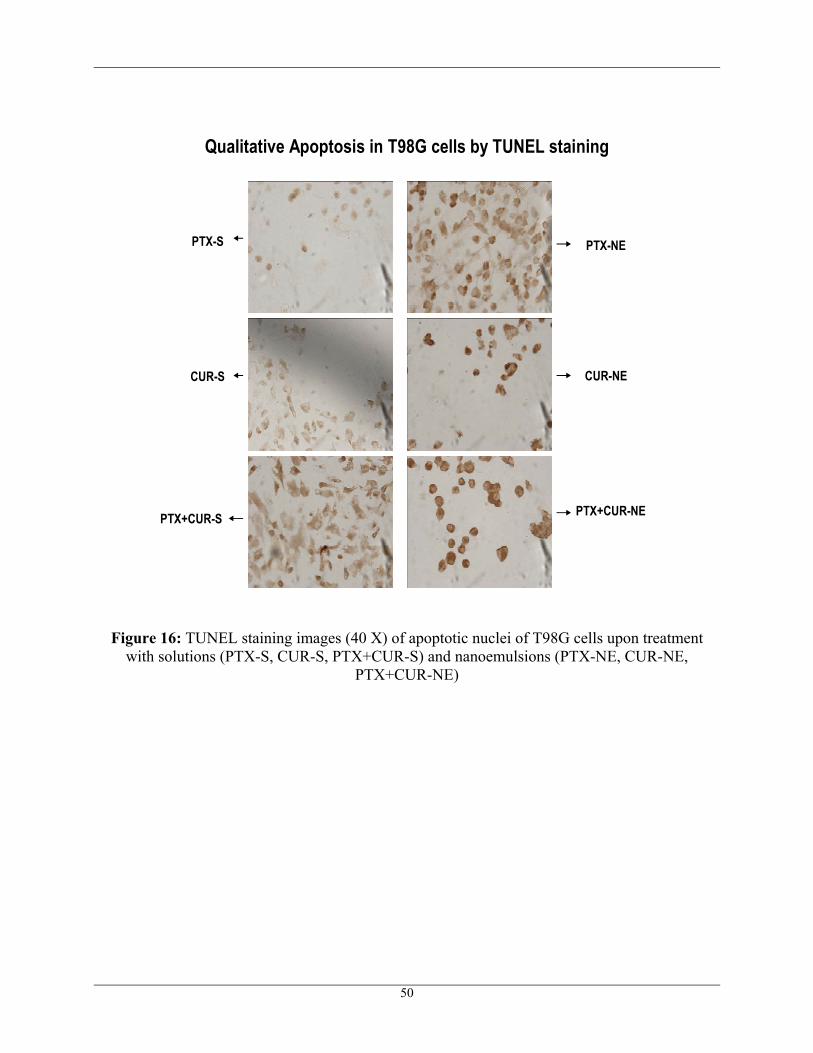
TUNEL assay is used to study apoptosis qualitatively. The assay is based on the
peroxidase catalyzed conversion of DAB and hydrogen peroxide to a brown precipitate, staining
apoptotic nuclei brown. rTdT enzyme, a component of the TUNEL kit end labels fragmented
DNA of apoptotic cells with biotinylated nucleotide at 3’OH end. Streptavidin-HRP bound to the
biotinylated nucleotide is detected by the peroxidase substrate, hydrogen peroxide and the
chromogen DAB, staining apoptotic nuclei brown. 100nM of PTX and 20µM of CUR were the
concentrations used to see the apoptotic activity in U87 and T98G cells. Untreated cells were
found to have no apoptotic activity as shown in Figure 14. Additionally, Figure 15 shows the
apoptotic activity of U87 cells upon different treatments and Figure 16 shows the apoptotic
activity of T98G cells upon different treatments. Both U87 and T98G cells treated with
nanoemulsions (PTX-NE, CUR-NE, PTX+CUR-NE) have shown more apoptotic activity when
47

compared to that of cells treated with aqueous solutions (PTX-S, CUR-S, PTX+CUR-S). The
most apoptotic activity was observed when 100nM of PTX was co-administered with 20µM of
CUR in the form of nanoemulsion in both sensitive U87 and resistant T98G human glioblastoma
cells. Figure 15 and Figure 16 also show that there were fewer cells when treated with
combination, which confirms that most of the cells were dead and got washed away. The
TUNEL results were found to be in correlation with Cytotoxicity results. TUNEL results suggest
that the apoptotic activity was enhanced with combination therapy and hence improved the
efficiency of glioblastoma chemotherapy.
U87 untreated cells T98G untreaetd cells
20X 20X
Figure 14: TUNEL staining images (20X) of untreated U87 and T98G cells
48
Qualitative Apoptosis in U87-MG cells by TUNEL staining
PTX-S
CUR-S
PTX+CUR-S
PTX-NE
CUR-NE
PTX+CUR-NE
Figure 15: TUNEL staining images (40 X) of apoptotic nuclei of U87 cells upon treatment with solutions (PTX-S, CUR-S, PTX+CUR-S) and nanoemulsions (PTX-NE, CUR-NE, PTX+CUR-
NE)
49
Qualitative Apoptosis in T98G cells by TUNEL staining
PTX-S
CUR-S
PTX+CUR-S
PTX-NE
CUR-NE
PTX+CUR-NE
Figure 16: TUNEL staining images (40 X) of apoptotic nuclei of T98G cells upon treatment with solutions (PTX-S, CUR-S, PTX+CUR-S) and nanoemulsions (PTX-NE, CUR-NE,
PTX+CUR-NE)
50

5. CONCLUSIONS
The result of this study has shown that PTX and CUR combination therapy in the form of
a nanoemulsion has significant potential for the treatment of human glioblastoma. Pine-nut oil-
containing nanoemulsions, which is rich in omega-3 PUFA, formulated using the ultrasound
method has proved to be a suitable carrier for both PTX and CUR. The results of
immunocytometric and Western blot analyses have shown the over-expression of MRP-1 in
resistant T98G cells when compared to that of sensitive U87 cells. The cytotoxicity results have
demonstrated that the nanoemulsions were more efficient in killing cells when compared to the
free drug in aqueous solution. Synergistic effect was observed especially in the resistant T98G
cells when 20 μM CUR was co-administered with 100 nM of PTX in the form of nanoemulsion,
making the system efficient for the treatment of glioblastoma. The results of the cellular uptake
and distribution study have shown that the designed nanoemulsion system can effectively deliver
PTX and CUR within the U87 and T98G cells. Western blot analysis has also shown the MRP-1
down regulation in both U87 and T98G cells and has also shown
the inhibition of NFκB pathway by CUR in U87 cells, but not to a great extent in T98G
cells which confirms that the synergism observed in the co-therapy is due to the MRP-1 down-
regulation by CUR which helps by increasing the accumulation of PTX within the tumor cell and
also overcoming the MDR phenomenon by inhibiting the NFκB pathway. Apoptosis induced by
PTX and CUR in U87 and T98G cells was well understood using the Qualtitative apoptosis
TUNEL assay.
In conclusion, it has been shown in this study that the nanotechnology based oil-in-water
pine-nut oil nanoemulsions improved the therapeutic efficacy by successfully delivering
combination therapeutic agents in GBM cells. It was found that CUR has the ability to sensitize
51

the glioblastoma cells to PTX-induced apoptosis. This dual therapeutic strategy has significant
potential in clinical management of GBM.
52

6. REFERENCES
1. S. A. Grossman and J. F. Batara. Current management of glioblastoma multiforme. Semin
Oncol 31: 635-44 (2004).
2. R. Stupp, P. Y. Dietrich, S. Ostermann Kraljevic, A. Pica, I. Maillard, P. Maeder, R.
Meuli, R. Janzer, G. Pizzolato, R. Miralbell, F. Porchet, L. Regli, N. de Tribolet, R. O.
Mirimanoff, and S. Leyvraz. Promising survival for patients with newly diagnosed
glioblastoma multiforme treated with concomitant radiation plus temozolomide followed
by adjuvant temozolomide. J Clin Oncol 20: 1375-82 (2002).
3. P. C. Burger, F. S. Vogel, S. B. Green, and T. A. Strike. Glioblastoma multiforme and
anaplastic astrocytoma. Pathologic criteria and prognostic implications. Cancer 56: 1106-
11 (1985).
4. N. G. Avgeropoulos and T. T. Batchelor. New treatment strategies for malignant gliomas.
Oncologist 4: 209-24 (1999).
5. D. Ribatti, B. Nico, E. Crivellato, and M. Artico. Development of the blood-brain barrier:
a historical point of view. Anat Rec B New Anat 289: 3-8 (2006).
6. A. G. de Boer, I. C. van der Sandt, and P. J. Gaillard. The role of drug transporters at the
blood-brain barrier. Annu Rev Pharmacol Toxicol 43: 629-56 (2003).
7. M. M. Amiji, P. K. Lai, D. B. Shenoy, and M. Rao. Intratumoral administration of
paclitaxel in an in situ gelling poloxamer 407 formulation. Pharm Dev Technol 7: 195-
202 (2002).
8. W. M. Pardridge. The blood-brain barrier: bottleneck in brain drug development.
NeuroRx 2: 3-14 (2005).
53

9. W. M. Pardridge. Molecular biology of the blood-brain barrier. Mol Biotechnol 30: 57-70
(2005).
10. R. Bendayan, G. Lee, and M. Bendayan. Functional expression and localization of P-
glycoprotein at the blood brain barrier. Microsc Res Tech 57: 365-80 (2002).
11. M. Pennati, A. J. Campbell, M. Curto, M. Binda, Y. Cheng, L. Z. Wang, N. Curtin, B. T.
Golding, R. J. Griffin, I. R. Hardcastle, A. Henderson, N. Zaffaroni, and D. R. Newell.
Potentiation of paclitaxel-induced apoptosis by the novel cyclin-dependent kinase
inhibitor NU6140: a possible role for survivin down-regulation. Mol Cancer Ther 4:
1328-37 (2005).
12. A. Desai, T. Vyas, and M. Amiji. Cytotoxicity and apoptosis enhancement in brain tumor
cells upon coadministration of paclitaxel and ceramide in nanoemulsion formulations. J
Pharm Sci (2007).
13. M. J. Glantz, M. C. Chamberlain, S. M. Chang, M. D. Prados, and B. F. Cole. The role of
paclitaxel in the treatment of primary and metastatic brain tumors. Semin Radiat Oncol 9:
27-33 (1999).
14. B. B. Aggarwal, S. Shishodia, Y. Takada, S. Banerjee, R. A. Newman, C. E. Bueso-
Ramos, and J. E. Price. Curcumin suppresses the paclitaxel-induced nuclear factor-
kappaB pathway in breast cancer cells and inhibits lung metastasis of human breast
cancer in nude mice. Clin Cancer Res 11: 7490-8 (2005).
15. S. V. Bava, V. T. Puliappadamba, A. Deepti, A. Nair, D. Karunagaran, and R. J. Anto.
Sensitization of taxol-induced apoptosis by curcumin involves down-regulation of
nuclear factor-kappaB and the serine/threonine kinase Akt and is independent of tubulin
polymerization. J Biol Chem 280: 6301-8 (2005).
54

16. H. L. Pahl. Activators and target genes of Rel/NF-kappaB transcription factors.
Oncogene 18: 6853-66 (1999).
17. E. Chun and K. Y. Lee. Bcl-2 and Bcl-xL are important for the induction of paclitaxel
resistance in human hepatocellular carcinoma cells. Biochem Biophys Res Commun 315:
771-9 (2004).
18. E. M. Kemper, A. E. van Zandbergen, C. Cleypool, H. A. Mos, W. Boogerd, J. H.
Beijnen, and O. van Tellingen. Increased penetration of paclitaxel into the brain by
inhibition of P-Glycoprotein. Clin Cancer Res 9: 2849-55 (2003).
19. A. J. Ruby, G. Kuttan, K. D. Babu, K. N. Rajasekharan, and R. Kuttan. Anti-tumour and
antioxidant activity of natural curcuminoids. Cancer Lett 94: 79-83 (1995).
20. M. T. Huang, T. Lysz, T. Ferraro, T. F. Abidi, J. D. Laskin, and A. H. Conney. Inhibitory
effects of curcumin on in vitro lipoxygenase and cyclooxygenase activities in mouse
epidermis. Cancer Res 51: 813-9 (1991).
21. S. Bisht, G. Feldmann, S. Soni, R. Ravi, C. Karikar, A. Maitra, and A. Maitra. Polymeric
nanoparticle-encapsulated curcumin ("nanocurcumin"): a novel strategy for human
cancer therapy. J Nanobiotechnology 5: 3 (2007).
22. B. H. Choi, C. G. Kim, Y. Lim, S. Y. Shin, and Y. H. Lee. Curcumin down-regulates the
multidrug-resistance mdr1b gene by inhibiting the PI3K/Akt/NFkappaB pathway. Cancer
Lett 259: 111-8 (2008).
23. P. Limtrakul. Curcumin as chemosensitizer. Adv Exp Med Biol 595: 269-300 (2007).
24. G. Bradley, P. F. Juranka, and V. Ling. Mechanism of multidrug resistance. Biochim
Biophys Acta 948: 87-128 (1988).
55

25. A. L. Harris and D. Hochhauser. Mechanisms of multidrug resistance in cancer treatment.
Acta Oncol 31: 205-13 (1992).
26. K. M. Dhandapani, V. B. Mahesh, and D. W. Brann. Curcumin suppresses growth and
chemoresistance of human glioblastoma cells via AP-1 and NFkappaB transcription
factors. J Neurochem 102: 522-38 (2007).
27. F. Kuhnel, L. Zender, Y. Paul, M. K. Tietze, C. Trautwein, M. Manns, and S. Kubicka.
NFkappaB mediates apoptosis through transcriptional activation of Fas (CD95) in
adenoviral hepatitis. J Biol Chem 275: 6421-7 (2000).
28. V. Maldonado, J. Melendez-Zajgla, and A. Ortega. Modulation of NF-kappa B, and Bcl-2
in apoptosis induced by cisplatin in HeLa cells. Mutat Res 381: 67-75 (1997).
29. E. Shaulian and M. Karin. AP-1 in cell proliferation and survival. Oncogene 20: 2390-
400 (2001).
30. C. Calatozzolo, M. Gelati, E. Ciusani, F. L. Sciacca, B. Pollo, L. Cajola, C. Marras, A.
Silvani, L. Vitellaro-Zuccarello, D. Croci, A. Boiardi, and A. Salmaggi. Expression of
drug resistance proteins Pgp, MRP1, MRP3, MRP5 and GST-pi in human glioma. J
Neurooncol 74: 113-21 (2005).
31. K. Ueda, M. M. Cornwell, M. M. Gottesman, I. Pastan, I. B. Roninson, V. Ling, and J. R.
Riordan. The mdr1 gene, responsible for multidrug-resistance, codes for P-glycoprotein.
Biochem Biophys Res Commun 141: 956-62 (1986).
32. T. A. Brooks, H. Minderman, K. L. O'Loughlin, P. Pera, I. Ojima, M. R. Baer, and R. J.
Bernacki. Taxane-based reversal agents modulate drug resistance mediated by P-
glycoprotein, multidrug resistance protein, and breast cancer resistance protein. Mol
Cancer Ther 2: 1195-205 (2003).
56

33. W. Chearwae, S. Anuchapreeda, K. Nandigama, S. V. Ambudkar, and P. Limtrakul.
Biochemical mechanism of modulation of human P-glycoprotein (ABCB1) by curcumin
I, II, and III purified from Turmeric powder. Biochem Pharmacol 68: 2043-52 (2004).
34. P. Limtrakul, S. Anuchapreeda, and D. Buddhasukh. Modulation of human multidrug-
resistance MDR-1 gene by natural curcuminoids. BMC Cancer 4: 13 (2004).
35. P. Borst, R. Evers, M. Kool, and J. Wijnholds. A family of drug transporters: the
multidrug resistance-associated proteins. J Natl Cancer Inst 92: 1295-302 (2000).
36. H. M. Wortelboer, M. Usta, A. E. van der Velde, M. G. Boersma, B. Spenkelink, J. J. van
Zanden, I. M. Rietjens, P. J. van Bladeren, and N. H. Cnubben. Interplay between MRP
inhibition and metabolism of MRP inhibitors: the case of curcumin. Chem Res Toxicol
16: 1642-51 (2003).
37. D. K. Sarker. Engineering of nanoemulsions for drug delivery. Curr Drug Deliv 2: 297-
310 (2005).
38. S. Shafiq, F. Shakeel, S. Talegaonkar, F. J. Ahmad, R. K. Khar, and M. Ali. Development
and bioavailability assessment of ramipril nanoemulsion formulation. Eur J Pharm
Biopharm 66: 227-43 (2007).
39. G. A. Silva. Nanotechnology approaches for drug and small molecule delivery across the
blood brain barrier. Surg Neurol 67: 113-6 (2007).
40. U. Schroeder, P. Sommerfeld, S. Ulrich, and B. A. Sabel. Nanoparticle technology for
delivery of drugs across the blood-brain barrier. J Pharm Sci 87: 1305-7 (1998).
41. S. B. Tiwari and M. M. Amiji. A review of nanocarrier-based CNS delivery systems.
Curr Drug Deliv 3: 219-32 (2006).
57

58
42. H. Maeda, J. Wu, T. Sawa, Y. Matsumura, and K. Hori. Tumor vascular permeability and
the EPR effect in macromolecular therapeutics: a review. J Control Release 65: 271-84
(2000).
43. L. E. van Vlerken, T. K. Vyas, and M. M. Amiji. Poly(ethylene glycol)-modified
nanocarriers for tumor-targeted and intracellular delivery. Pharm Res 24: 1405-14
(2007).
44. J. A. Hamilton and K. Brunaldi. A model for fatty acid transport into the brain. J Mol
Neurosci 33: 12-7 (2007).
45. J. A. Hamilton, C. J. Hillard, A. A. Spector, and P. A. Watkins. Brain uptake and
utilization of fatty acids, lipids and lipoproteins: application to neurological disorders. J
Mol Neurosci 33: 2-11 (2007).
46. E. Ryan, K. Galvin, T. P. O'Connor, A. R. Maguire, and N. M. O'Brien. Fatty acid
profile, tocopherol, squalene and phytosterol content of brazil, pecan, pine, pistachio and
cashew nuts. Int J Food Sci Nutr 57: 219-28 (2006).
47. J. Edmond. Essential polyunsaturated fatty acids and the barrier to the brain: the
components of a model for transport. J Mol Neurosci 16: 181-93; discussion 215-21
(2001).
48. S. Khandavilli and R. Panchagnula. Nanoemulsions as versatile formulations for
paclitaxel delivery: peroral and dermal delivery studies in rats. J Invest Dermatol 127:
154-62 (2007).




























