Embed Size (px)
Citation preview

at SciVerse ScienceDirect
Neuropharmacology 66 (2013) 355e364
Contents lists available
Neuropharmacology
journal homepage: www.elsevier .com/locate/neuropharm
MPEP, an mGlu5 receptor antagonist, reduces the development of L-DOPA-induced motor complications in de novo parkinsonian monkeys: Biochemicalcorrelates
Nicolas Morin a,b, Laurent Grégoire a, Marc Morissette a, Sandrine Desrayaud c, Baltazar Gomez-Mancilla c,Fabrizio Gasparini c, Thérèse Di Paolo a,b,*
aMolecular Endocrinology and Genomic Research Center, Laval University Medical Center (CHUQ), Quebec, QC, Canadab Faculty of Pharmacy, Laval University, Quebec, QC, CanadacNovartis Institute for BioMedical Research, Basel, Switzerland
a r t i c l e i n f o
Article history:Received 5 March 2012Received in revised form6 July 2012Accepted 9 July 2012
Keywords:mGlu5 receptorDyskinesiaL-DOPAMPTP-lesioned monkeyMPEP
* Corresponding author. Molecular Endocrinology aLaval University Medical Center (CHUQ), 2705 LaurieG1V 4G2. Tel.: þ1 418 654 2296; fax: þ1 418 654 276
E-mail address: [email protected] (
0028-3908/$ e see front matter � 2012 Elsevier Ltd.http://dx.doi.org/10.1016/j.neuropharm.2012.07.036
a b s t r a c t
L-3,4-Dihydroxyphenylalanine (L-DOPA), the gold standard therapy for Parkinson disease (PD), is asso-ciated with motor fluctuations and dyskinesias. This study sought to prevent the development of L-DOPA-induced dyskinesias (LID) with the metabotropic glutamate receptor type 5 (mGlu5 receptor) antagonist2-methyl-6-(phenylethynyl)pyridine (MPEP) in the de novo treatment of monkeys lesioned with 1-methyl-4-phenyl-1,2,3,6-tetrahydropyridine (MPTP) as a PD model. MPTP-lesioned monkeys weretreated once daily for one month with either L-DOPA or L-DOPA þ MPEP (10 mg/kg). MPEP (administered15 min before L-DOPA) plasma concentrations were elevated during all the L-DOPA motor activation anddid not accumulate during a month. The antiparkinsonian effect was maintained throughout the treat-ment period in MPTP-lesioned monkeys treated with L-DOPA þ MPEP, while the duration of this effectdecreased over time in MPTP-lesioned monkeys treated with L-DOPA alone, suggesting wearing-off. Overthe month-long treatment, the mean dyskinesia score increased in L-DOPA-treated monkeys; interest-ingly, this increase was reduced by overall 72% in the L-DOPA þ MPEP group. Mean dyskinesia scores ofmonkeys correlated inversely with plasma MPEP concentrations. Normal control and saline-treatedMPTP-lesioned monkeys were also included for biochemical analyses. All MPTP-lesioned monkeyswere extensively and similarly denervated. [3H]ABP688 specific binding to mGlu5 receptors increased inthe putamen of L-DOPA-treated monkeys compared to control, saline or L-DOPA þ MPEP-treatedmonkeys. Mean dyskinesia scores of MPTP-lesioned monkeys correlated positively with [3H]ABP688specific binding in the putamen. This study showed a beneficial chronic antidyskinetic effect of MPEP inde novo L-DOPA-treated MPTP-lesioned monkeys, supporting the therapeutic use of mGlu5 receptorantagonists in PD to prevent LID.
This article is part of a Special Issue entitled ‘Metabotropic Glutamate Receptors’.� 2012 Elsevier Ltd. All rights reserved.
1. Introduction
Parkinson’s disease (PD) is the most common neurodegenera-tive movement disorder and is likely to increase due to the aging ofpopulations (de Lau and Breteler, 2006; Siderowf and Stern, 2003).Replacing dopamine (DA) deficiency with its precursor L-3,4-dihydroxyphenylalanine (L-DOPA) is the most effective and
nd Genomic Research Center,r Blvd., Quebec, QC, Canada1.T. Di Paolo).
All rights reserved.
commonly used treatment for PD. In the long-term, w80% of L-DOPA-treated patients will develop abnormal involuntary move-ments, including L-DOPA-induced dyskinesias (LID) which can bevery difficult to manage (Fabbrini et al., 2007; Gottwald andAminoff, 2011; Meissner et al., 2011). The mechanisms underlyingLID are still unknown. Altered dopaminergic and nondopaminergicneurotransmission and peptide neuromodulators levels in the basalganglia are observed in LID (Blandini and Armentero, 2012). Morespecifically related to the present study, glutamate neurotrans-mission plays a crucial role in the modulation of corticostriatalinputs and striatal output to downstream nuclei of the basal gangliacircuit (Blandini and Armentero, 2012).

Table 1Description of the MPTP-lesioned monkeys the day before the onset of experiment(day 1).
Monkey (number) Age(years)
Weight(kg)
Total doseof MPTP(mg)
Time afterlast MPTPdose (month)a
Parkinsonianscoreb
L-DOPA group1 7.7 2.7 23.0 6.5 11.32 7.3 4.2 16.0 9.0 9.83 7.0 3.8 31.0 9.3 7.44 6.5 3.9 28.0 4.0 10.4Mean 7.1 3.7 24.5 7.2 9.7SEM 0.3 0.3 3.3 1.2 0.8MPEP þ L-DOPA group
5 6.7 3.8 16.0 9.0 10.16 7.3 3.4 31.5 8.3 9.87 5.8 3.5 30.2 6.5 7.68 5.1 3.9 28.0 4.0 12.69 5.3 3.6 14.0 3.1 8.9Mean 6.0 3.6 23.9 6.2 9.8SEM 0.4 0.1 3.7 1.2 0.8p-value 0.08 0.97 0.91 0.56 0.95
a Time between the last MPTP administration and the onset of the L-DOPAtreatment.
b Basal parkinsonian score after administration of vehicle to monkeys the daybefore the onset of L-DOPA treatment.
N. Morin et al. / Neuropharmacology 66 (2013) 355e364356
Glutamate is the most abundant excitatory neurotransmitter inthe brain, mediating as much as 70% of synaptic transmission in thecentral nervous system (Meldrum, 2000; Platt, 2007). Gluta-matergic transmission is increased in the basal ganglia in PD(Klockgether and Turski, 1993) and is also believed to be involved inLID (Calon et al., 2003b; Chase and Oh, 2000). Anti-glutamatergicdrugs, such as antagonists of the ionotropic glutamate receptors,have been proposed to relieve PD symptoms and decrease LID in PD(Olanow et al., 2009) and in animal models such as the 1-methyl-4-phenyl-1,2,3,6-tetrahydropyridine (MPTP)-lesioned monkey (HadjTahar et al., 2004). Amantadine, which has N-methyl D-aspartate(NMDA) anti-glutamatergic properties, is currently the only drugused in the clinic shown to have modest benefit in LID, and to alsoimprove akinesia, rigidity and tremor (Stocchi et al., 2008;Verhagen Metman et al., 1998). However, these drugs have signif-icant adverse effects in many patients such as cognitive impair-ment, which significantly limits their use (Stocchi et al., 2008).
In addition to ionotropic receptors, the action of glutamate ismediated via metabotropic glutamate receptors (mGlu receptors),a family of G-protein coupled receptors comprising eight subtypes.mGlu receptors display a higher affinity for glutamate than iono-tropic glutamate receptors (Conn and Pin, 1997). There is evidencefor an extrasynaptic diffusion of glutamate in the central nervoussystem, indicating that the activation of mGlu receptors is likelymore complex than mere synaptic receptor activation (Smith et al.,2001). The mGlu5 receptor subtype, which belongs to group 1 ofmGlu receptors, is highly expressed in striatal medium spinyneurons (Conn et al., 2005; Paquet and Smith, 2003; Testa et al.,1994) and is positively linked to the activation of phospholipase Cand phosphoinositide hydrolysis via Gq. The mGlu5 receptor playsa key role in modulating the responses mediated by NMDAreceptors and L-type calcium channels (Gubellini et al., 2004).Moreover, mGlu5 receptor activation could reduce striatal DAuptake by phosphorylation of the transporter via the activation ofcalciumecalmodulin-dependent protein kinase II and proteinkinase C (Page et al., 2001). This regulatory interaction providesevidence that glutamatergic and dopaminergic systems interactclosely in the striatum and that glutamate may potentially regulatedopaminergic neurotransmission (Samadi et al., 2007). Antago-nistic interaction between the D2R DA receptor and mGlu5 recep-tors (Fuxe et al., 2008), mGlu5 receptor activation of NMDAreceptors (Pisani et al., 2001) as well as NMDA and mGlu5 receptorcolocalization have been described (Perroy et al., 2008), singlingout this receptor as an interesting pharmacological target.
Negative allosteric modulators inhibit receptor function at a sitedistal to the actual orthosteric ligand binding domain of thereceptor. These ligands can normalize excessive glutamate trans-mission without blocking the physiological actions of glutamate inthe brain (Rouse et al., 2000; Sheffler et al., 2011). Over the past fewyears, pharmacological antagonism (or more precisely negativeallosteric modulation) of mGlu5 receptor with the selectiveantagonists 2-methyl-6-(phenylethynyl)pyridine (MPEP) and 3-[(2-methyl-1,3-thiazol-4-yl)ethynyl]pyridine (MTEP) has beenshown to inhibit LIDmanifestations in the 6-hydroxydopamine- (6-OHDA) lesioned PD rat model (Gravius et al., 2008; Levandis et al.,2008). Several studies have investigated the acute antiparkinsonianand antidyskinetic effects of MPEP and MTEP in 6-OHDA- orhaloperidol-treated rats tomodel PD (Breysse et al., 2002; Dekundyet al., 2006; Ossowska et al., 2005). mGlu5 receptor specific bindingis also increased in the basal ganglia of parkinsonian monkeys thathave developed LID following chronic L-DOPA treatment and inparkinsonian patients with motor complications (Ouattara et al.,2010, 2011; Samadi et al., 2008). Moreover a study in 6-OHDArats showed the antidyskinetic effect of MTEP (5 mg/kg) witha chronic treatment of 3 weeks (Rylander et al., 2009). The mGlu5
receptor antagonists MPEP, MTEP and AFQ056 have been recentlyshown to reduce LID in MPTP-lesioned monkeys (Gregoire et al.,2011; Johnston et al., 2010; Morin et al., 2010). Moreover, AFQ056reduces LID in parkinsonian patients (Berg et al., 2011). In fourMPTP-lesioned rhesus male monkeys, Rylander et al. (2010)observed that LID were less pronounced upon a 17-day adjuncttreatment with the mGlu5 receptor antagonist fenobam, comparedto three vehicle þ L-DOPA-treated MPTP-lesioned monkeys.
The present study aimed at preventing LID development witha one-month de novo treatment with the prototypical mGlu5receptor antagonist MPEP in combination with L-DOPA in fiveovariectomized MPTP-lesioned Macaca fascicularis monkeys tomodel aging women, compared to four MPTP-lesioned monkeysreceiving L-DOPA treatment. Brain mGlu receptor levels have notbeen investigated following treatment with mGlu5 receptor-targeted drugs in any species. mGlu5 receptor antagonists arevery promising drugs for the management of various brain disor-ders, and one of their potential applications is LID therapy. Sincethese drugswould be used for long-term treatment, it is essential toknow the long-term behavioral effects of this class of compoundsand ensuing possible biochemical adaptations of the brain. In thepresent study, we measured mGlu5 receptor specific binding in theputamen to evaluate the effects of a chronic treatment with MPEPon mGlu5 receptors as well as their correlations with LID.
2. Materials and methods
2.1. Animals and treatments
Nine female ovariectomized cynomolgusmonkeys (M. fascicularis) were used forthe behavioral experiments (Table 1). In addition, the experiment comprised 9 otherovariectomized cynomolgus monkeys (M. fascicularis), including 4 monkeys used asnormal controls (age: 5.13 � 0.16 y; weight: 3.7 � 0.2 kg) and 5 MPTP-lesionedmonkeys administered saline (age: 5.35 � 0.15 y; weight: 3.6 � 0.1 kg; meanparkinsonian score: 9.9 � 0.4), used for the biochemical analyses. These animalswere drug-naïve at the start of the experiment. Handling of the primates was per-formed in accordance with the National Institute of Health Guide for the Care andUse of Laboratory Animals. All procedures, including means tominimize discomfort,were reviewed and approved by the Institutional Animal Care Committee of LavalUniversity. The animals were rendered parkinsonian by continuous infusion of MPTP(SigmaeAldrich, Canada, Oakville, ON, Canada) using subcutaneous Alzet osmoticminipumps (0.5 mg/24 h) until they developed a stable parkinsonian syndrome(Fig. 1). After an average period of 6.6 months of stabilization, animals were treatedonce daily for one month with L-DOPA/benserazide 100/25 mg capsule p.o. alone

Fig. 1. Time lines for animal treatments, blood sampling and behavioral observations in the present study (BS2: MPEP blood sampling at day 2; BS16: MPEP blood sampling at day16; BS31: MPEP blood sampling at day 31; PK28: complete MPEP pharmacokinetic at day 28; CSF31: MPEP CSF sampling at day 31).
N. Morin et al. / Neuropharmacology 66 (2013) 355e364 357
(Prolopa, Hoffmann-La Roche; a mixture of 100 mg of L-DOPA and 25 mg benser-azide; n ¼ 4 monkeys) or with the combination of L-DOPA/benserazide (100/25 mg)and MPEP (10 mg/kg, administered 15 min prior to L-DOPA; n ¼ 5 monkeys). MPTP-lesioned monkeys of both groups received the same dose of 100 mg L-DOPA/25 mgbenserazide once daily for one month, a dose that was comparable on a body weightbasis since the mean weight of each group was similar (Table 1).
2.2. Drugs
MPEP (provided by Novartis Pharma AG, Basel, Switzerland) solutions werefreshly prepared on every experimental day. MPEP was first solubilized in sterilewater and between 12 and 20ml of MPEP solutionwere administered by nasogastricgavage.
2.3. Pharmacokinetic study
Blood and cerebrospinal fluid (CSF) samplings were performed following theadministration of MPEP (10 mg/kg) and L-DOPA. MPEP blood samples were with-drawn 15 min before L-DOPA administration on days 2, 16 and 31, just before theirdaily MPEP administration. MPEP CSF samplings were taken on day 31 just beforeeuthanasia, i.e. 24 h after the last MPEP administration. Moreover, a completepharmacokinetic study was performed on day 28. Blood samples (1 ml) werecollected at 0, 0.5, 1.0, 1.5, 2.5, 4.0 and 6.0 h after MPEP administration in lithiumheparin tubes, and centrifuged (10 min, 1000 � g) at 4 �C. Plasma samples werestored at�80 �C. MPEP plasma concentrations were obtained by plasma partitioningwith an aqueous solution of saturated NaCl and acetonitrile added to each samplebefore analysis of the supernatant by liquid chromatography tandem mass spec-trometry (LC/MS/MS) using atmospheric pressure chemical ionization. The lowerlimit of quantification of the assay was 10 ng/ml.
2.4. Behavioral MPEP chronic study
The animals were first evaluated following vehicle (sterile water) administrationalone. Behavioral measures after L-DOPA/benserazide alone or the combination of L-DOPA/benserazide and MPEP were performed every other day for 30 d. The animalswere observed through a one-way screen and were scored immediately every15 min for antiparkinsonian and dyskinetic responses for the whole duration of theL-DOPA response. The end of the L-DOPA response period corresponds to the fullrestoration of the initial basal parkinsonian score for each monkey. Behavioralmeasures were rated by independent evaluators blind to treatment assignment.
2.4.1. Parkinsonian scoreA disability scale developed in our laboratory was used to evaluate the parkin-
sonian syndrome in MPTP-lesioned monkeys (Hadj Tahar et al., 2004; Samadi et al.,2003). Briefly, behaviors were scored every 15 min (maximal score: 16) for: (a)posture: normal ¼ 0, flexed intermittent ¼ 1, flexed constant ¼ 2, crouched ¼ 3; (b)mobility: normal ¼ 0, mild reduction ¼ 1, moderate reduction ¼ 2, severereduction ¼ 3; (c) climbing: present ¼ 0, absent ¼ 1; (d) gait: normal ¼ 0, slow ¼ 1,very slow¼ 2, very slowwith freezing¼ 3; (e) grooming: present¼ 0, absent¼ 1; (f)vocalization: present¼ 0, absent ¼ 1; (g) social interaction: present¼ 0, absent ¼ 1;(h) tremor: absent ¼ 0, mild action tremor ¼ 1, moderate action tremor ¼ 2, restingtremor ¼ 3.
2.4.2. Dyskinetic responseLID were scored every 15 min for the whole duration of the L-DOPA effect (Hadj
Tahar et al., 2004; Samadi et al., 2003). LID were rated for the face, neck, trunk, arms
and legs as follows: none ¼ 0; mild (occasional) ¼ 1; moderate (intermittent) ¼ 2;severe (continuous)¼ 3. The dyskinetic score obtained was the sum of the scores forall body segments (maximal score: 21).
2.4.3. Locomotor responseLocomotor activity was monitored continuously with an electronic motility
monitoring system fixed on each cage (Datascience, St. Paul, MN). Computerizedmobility counts were obtained every 5 min.
2.5. Tissue preparation
At the end of experiments, all 9 drug-treatedmonkeys, along with the 4 controlsand 5 saline-treated MPTP-lesioned monkeys, were killed by an overdose of sodiumpentobarbital. The time interval between the last drug treatment and euthanasiawas 24 h. Brains were removed, immersed in isopentane for <30 s (�40 �C), andkept frozen at �80 �C until use. Brains were cut into 12-mm coronal sections ona cryostat (�18 �C). Samples of the posterior putamen from levels A15eA18 asdefined in the atlas of Szabo and Cowan (Szabo and Cowan, 1984) were investigated.Slices were thaw-mounted onto SuperFrost Plus slides (Brain Research Laboratories,Newton, MA) and stored at �80 �C until use for assays.
2.5.1. DA concentrationsSmall frozen tissue pieces of coronal brain sections were taken, the putamen
dissected and homogenized in 250 ml of 0.1 M HClO4 at 4 �C and centrifuged at10,000 � g for 20 min to precipitate proteins. The supernatants were kept at �80 �Cin small polyethylene tubes until time of assay, while the pellets were solubilized in100 ml of 0.1 M NaOH for determination of protein content. DA concentrations in theputamen were measured by high-performance liquid chromatography with elec-trochemical detection, according to previously published procedures (Goulet et al.,1999; Morissette et al., 2006).
2.5.2. Dopamine transporter (DAT) autoradiographyThe DAT was labeled with 3b-[4-125I-iodophenyl]tropane-2b-carboxylic acid
isopropyl ester ([125I]RTI-121; 2200 Ci/mmol, Mandel, Boston, MA) bindingaccording to our previously published procedure (Calon et al., 2002; Morissetteet al., 2006). Specific binding to the DAT was measured using 25 pM [125I]RTI-121.Non-specific binding was determined in the presence of 100 nM mazindol (SandozPharmaceuticals, Dorval, Quebec). All experimental groups were simultaneouslyanalyzed in the same assay.
2.6. [3H]ABP688 autoradiography
mGlu5 receptor specific binding was evaluated with [3H]ABP688 binding(81.6 Ci/mmol, Novartis, Switzerland), a high-affinity and selective mGlu5 receptorantagonist (Hintermann et al., 2007), according to our published conditions(Ouattara et al., 2010, 2011) using 5 nM [3H]ABP688 and 10 mM MPEP to estimatenon-specific binding. Slide-mounted tissue sections were exposed to BioMax MRfilms (Kodak; along with Amersham standard [3H]-microscales) for 20 d at roomtemperature.
2.7. Autoradiography data analysis
The intensity of [125I]RTI-121 and [3H]ABP688 autoradiographic labeling wasquantified on X-ray films with a Power Macintosh G4 connected to a video camera(XC-77; Sony) and a constant-illumination light table using computerized densi-tometry (NIH image v.1.63). Optical gray densities were transformed into nCi per mg
N. Morin et al. / Neuropharmacology 66 (2013) 355e364358
of tissue using a standard radioactivity curve. Results were finally converted intofmol/mg of tissue using the radioligand specific activity.
2.8. Statistical analysis
An unpaired Student’s t-test was used to compare age, weight, dose of MPTP andtime of stabilization after MPTP administration between the L-DOPA and L-DOPA þ MPEP groups. The ManneWhitney test was used to compare the initialbasal parkinsonian scores between the L-DOPA and L-DOPAþMPEP groups. For eachtreatment day and for each monkey, a mean parkinsonian score and a meandyskinetic score (total period) were obtained by averaging all 15-min scores
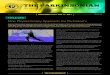
Fig. 2. Motorbehaviorof femaleMacaca fascicularisMPTP-lesionedmonkeys treatedoncedaily for onemonthwith L-DOPA alone (100mg; 4monkeys) orwith L-DOPA (100mg) andMPEP (10 mg/kg, administered 15 min prior to L-DOPA; 5 monkeys). (A) Duration of theantiparkinsonian response; (B) meanweekly parkinsonian score; (C) locomotion.
obtained for the duration of the response. Mean daily parkinsonian and dyskineticscores were then pooled for each week of treatment. For intra-group comparisons,the mean scores/week/animal were analyzed using the Friedman non-parametrictest followed by a BonferronieDunn’s multiple comparison test. TheManneWhitney test was used for between-group comparisons. The median andrange of parkinsonian and dyskinesia scores are represented with a horizontal lineand a box, respectively, in the Figures. Values for locomotor activity and duration ofL-DOPA response (in min) were analyzed with an ANOVA for repeated measuresfollowed by a Dunnett’s multiple comparison test. The results of locomotor activityand duration of L-DOPA response are presented as the mean � SEM. Statisticalcomparisons for autoradiographic labeling intensities and catecholamine concen-trations were performed by an ANOVA, followed by post-hoc pairwise comparisonswith the Fisher’s least significant difference test. Data are expressed as themean � SEM in the putamen as a percentage of controls. A value of p < 0.05 wasconsidered significant.
3. Results
3.1. Characteristics of the L-DOPA- and L-DOPA þ MPEP-treatedmonkeys
Table 1 shows the age, weight, MPTP dose, time of stabilizationafter MPTP administration and basal parkinsonian scores of the L-
Fig. 3. Dyskinetic behavior of female Macaca fascicularis MPTP-lesioned monkeystreated once daily for one month with L-DOPA alone (100 mg; 4 monkeys) or with L-DOPA (100 mg) and MPEP (10 mg/kg, given 15 min prior to L-DOPA; 5 monkeys). (A)Mean daily dyskinesia score; (B) mean weekly dyskinesia score.
N. Morin et al. / Neuropharmacology 66 (2013) 355e364 359
DOPA and the L-DOPA þ MPEP groups of MPTP-lesioned monkeysthe day before the beginning of the behavioral experiment (day 1);no statistically significant difference was observed for theseparameters between the two experimental groups.
3.2. MPEP effect on dyskinesia and parkinsonism
The duration of the L-DOPA antiparkinsonian effect significantlydecreased in the L-DOPA-treated group on weeks 3 and 4 (areduction of 49 and 56 min, respectively) compared to the firstweek (F3,9 ¼ 4.86, p < 0.05), while the decrease was not significantin the presence of MPEP (F3,12 ¼ 2.13, p ¼ 0.15; Fig. 2A). No statis-tically significant difference was observed in the duration of the L-DOPA antiparkinsonian effect between the L-DOPA and L-DOPA þ MPEP groups for each respective week.
The antiparkinsonian response in MPTP-lesioned monkeystreated with L-DOPA þ MPEP was maintained during the month asmeasured with locomotion and antiparkinsonian scores ascompared to the L-DOPA-treated group. L-DOPA administered aloneto MPTP-lesioned monkeys induced a decrease of parkinsonianscores (Friedman statistics (Q) ¼ 9.40, p < 0.001); this was main-tained upon the addition of MPEP for the whole duration of thestudy (Q ¼ 13.60, p < 0.01; Fig. 2B). L-DOPA administered alone toMPTP-lesioned monkeys significantly increased their locomotoractivity on weeks 3 and 4 (F4,19 ¼ 3.18, p < 0.05) and this was also
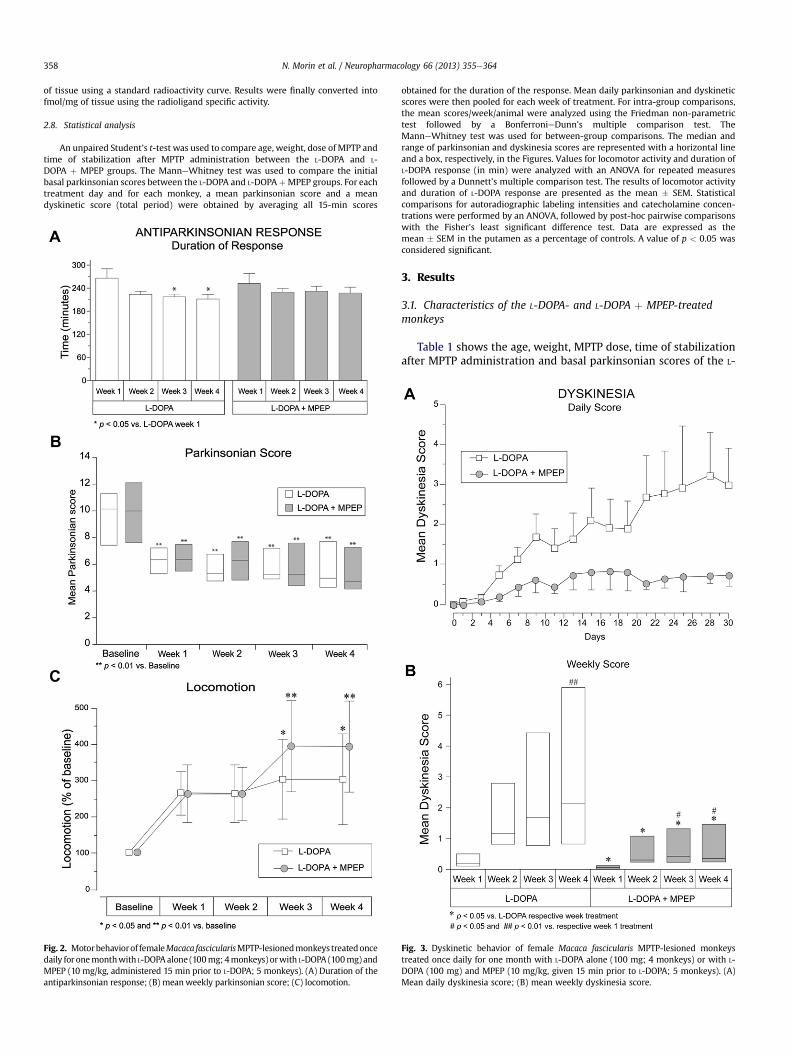
Fig. 4. Time course of the development of LID after L-DOPA treatment in MPTP-lesioned moalone (A and C; 4 monkeys) or with the combination of L-DOPA þ MPEP (B and D; 5 monk
observed upon the further addition of MPEP (F4,19 ¼ 6.36, p < 0.01;Fig. 2C). No statistically significant difference was observed forlocomotion and parkinsonian scores between the L-DOPA and L-DOPA þ MPEP groups.
L-DOPA induced choreic and dystonic LID in the MPTP-lesionedmonkeys, which were prevented by the addition of MPEP,compared to L-DOPA alone. Our dyskinesia score did not discrimi-nate choreic from dystonic LID, which prevented a clear assessmentof an association between the choreic and/or dystonic nature of LIDand the effect of MPEP. The mean dyskinesia score during the fullinterval of L-DOPA antiparkinsonian motor effect was lower withthe addition of MPEP for thewhole duration of the study, comparedto the L-DOPA-treated group (Fig. 3A). The mean dyskinesia scoresignificantly increased over a month in the L-DOPA-treated group(Q ¼ 10.80, p < 0.01). By contrast, that score was lower by 81%, 65%,68% and 74% in L-DOPA þ MPEP-treated monkeys, respectively, foreach consecutiveweek of the one-month treatment, with amean of72% for the four weeks combined (Q ¼ 11.45, p < 0.01; Fig. 3B).
Fig. 4 displays the time course of LID development in L-DOPA-and L-DOPAþMPEP-treatedmonkeys during the time of the L-DOPAantiparkinsonian effect on days 7, 14, 21 and 28, respectively, ascompared to the parkinsonian score for the sameperiod. The furtheraddition of MPEP to L-DOPA reduced the intensity and duration ofLID during the effect of L-DOPA (approximately 2.3 h). No abnormalbehavior such as dizziness, hyperactivity, hallucination, orthostatic
nkeys. Each curve represents the average parkinsonian and dyskinesia score of L-DOPAeys) on days 7, 14, 21 and 28, respectively.
N. Morin et al. / Neuropharmacology 66 (2013) 355e364360
hypotension, falling and stereotypywas observed at any timeduringL-DOPA or L-DOPA þ MPEP chronic treatment.
3.3. Pharmacokinetics of MPEP
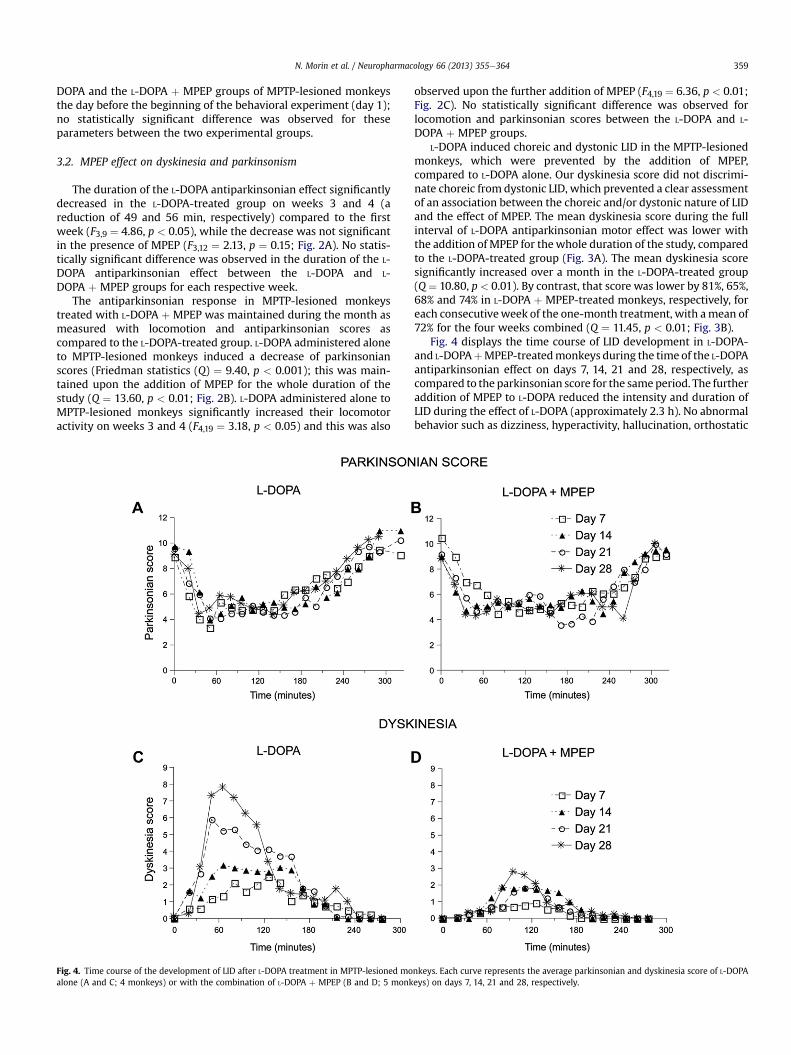
In the pharmacokinetic study, plasma MPEP levels weremeasured in the five monkeys as a function of time following theadministration of MPEP on day 28. Fig. 5 displays the time course ofthe mean LID and of the mean plasma MPEP concentrations for thefive individual L-DOPA þ MPEP-treated monkeys as well as thecorresponding means for the group on day 28. It shows thatadministration of MPEP 15min before L-DOPA led to an elevation ofplasmaMPEP concentration that closely paralleledmotor activationby L-DOPA in the MPTP-lesioned monkeys (Fig. 5).
Fig. 5. Time course of LID in individual L-DOPA þ MPEP-treated monkeys compared to theirof observation in the 5 monkeys are also shown.
Plasma MPEP concentrations had detectably risen as early as30 min after MPEP administration p.o. (608 � 311 nM), and peakedat 90e120 min (Cmax ¼ 1233� 288 nM). During the month of MPEPtreatment, plasma MPEP levels had returned to basal values by8e12 h after administration and remained low for as long as 24 hafter administration (19 � 4, 21 � 6 and 39 � 21 nM, respectively,on days 2, 16 and 31). MPEP was very low (4 nM for one monkey) orundetectable (for the othermonkeys) in the CSF taken 24 h after thelast MPEP administration on day 31 just before euthanasia (data notshown).
There was a significant negative correlation between meandyskinesia scores for the L-DOPA and L-DOPA þ MPEP-treatedmonkeys (n ¼ 9) and MPEP plasma concentrations on day 28(r ¼ �0.732, p ¼ 0.025; Fig. 6).
plasma MPEP concentrations on day 28 (n ¼ 5). The mean values for the combined sets
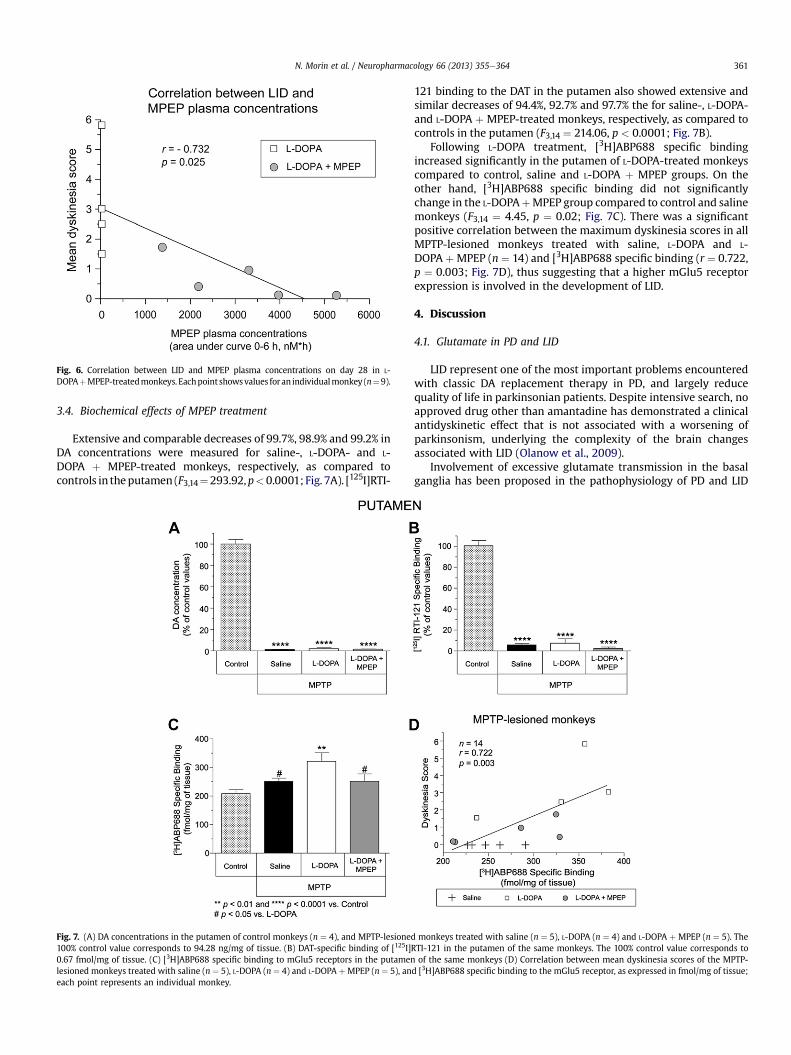
Fig. 6. Correlation between LID and MPEP plasma concentrations on day 28 in L-DOPAþMPEP-treatedmonkeys. Eachpoint showsvalues for an individualmonkey (n¼ 9).
N. Morin et al. / Neuropharmacology 66 (2013) 355e364 361
3.4. Biochemical effects of MPEP treatment
Extensive and comparable decreases of 99.7%, 98.9% and 99.2% inDA concentrations were measured for saline-, L-DOPA- and L-DOPA þ MPEP-treated monkeys, respectively, as compared tocontrols in theputamen (F3,14¼293.92,p<0.0001; Fig. 7A). [125I]RTI-
Fig. 7. (A) DA concentrations in the putamen of control monkeys (n ¼ 4), and MPTP-lesioned100% control value corresponds to 94.28 ng/mg of tissue. (B) DAT-specific binding of [125I]R0.67 fmol/mg of tissue. (C) [3H]ABP688 specific binding to mGlu5 receptors in the putamenlesioned monkeys treated with saline (n ¼ 5), L-DOPA (n ¼ 4) and L-DOPA þMPEP (n ¼ 5), aneach point represents an individual monkey.
121 binding to the DAT in the putamen also showed extensive andsimilar decreases of 94.4%, 92.7% and 97.7% the for saline-, L-DOPA-and L-DOPA þ MPEP-treated monkeys, respectively, as compared tocontrols in the putamen (F3,14 ¼ 214.06, p < 0.0001; Fig. 7B).
Following L-DOPA treatment, [3H]ABP688 specific bindingincreased significantly in the putamen of L-DOPA-treated monkeyscompared to control, saline and L-DOPA þ MPEP groups. On theother hand, [3H]ABP688 specific binding did not significantlychange in the L-DOPAþMPEP group compared to control and salinemonkeys (F3,14 ¼ 4.45, p ¼ 0.02; Fig. 7C). There was a significantpositive correlation between the maximum dyskinesia scores in allMPTP-lesioned monkeys treated with saline, L-DOPA and L-DOPA þMPEP (n ¼ 14) and [3H]ABP688 specific binding (r ¼ 0.722,p ¼ 0.003; Fig. 7D), thus suggesting that a higher mGlu5 receptorexpression is involved in the development of LID.
4. Discussion
4.1. Glutamate in PD and LID
LID represent one of the most important problems encounteredwith classic DA replacement therapy in PD, and largely reducequality of life in parkinsonian patients. Despite intensive search, noapproved drug other than amantadine has demonstrated a clinicalantidyskinetic effect that is not associated with a worsening ofparkinsonism, underlying the complexity of the brain changesassociated with LID (Olanow et al., 2009).
Involvement of excessive glutamate transmission in the basalganglia has been proposed in the pathophysiology of PD and LID
monkeys treated with saline (n ¼ 5), L-DOPA (n ¼ 4) and L-DOPA þ MPEP (n ¼ 5). TheTI-121 in the putamen of the same monkeys. The 100% control value corresponds toof the same monkeys (D) Correlation between mean dyskinesia scores of the MPTP-
d [3H]ABP688 specific binding to the mGlu5 receptor, as expressed in fmol/mg of tissue;

N. Morin et al. / Neuropharmacology 66 (2013) 355e364362
(Bezard et al., 2001; Blandini and Armentero, 2012). Althoughpsychiatric and cognitive impairments in humans limit theirusefulness (Krystal et al., 1994; Meldrum, 1998), the preclinical andclinical use of ionotropic glutamate receptor antagonists hasconfirmed the role of excessive glutamatergic transmission in thepathology of parkinsonian symptoms and LID (Blanchet et al., 2003).Hence, drugs acting at mGlu receptors are alternative therapeutictools for LID treatment (Berg et al., 2011; Meissner et al., 2011).
4.2. mGlu5 receptor and LID
Based on our previous biochemical results showing high mGlu5receptor specific binding in the putamen of parkinsonian patientsand MPTP-lesioned monkeys with LID (Ouattara et al., 2011;Samadi et al., 2008) and neurochemical correlates of mGlu5receptor antagonism in a rat model of LID (Mela et al., 2007;Rylander et al., 2009), we addressed the mGlu5 receptor subtypemore specifically in our assessment of the potential of drugs tar-geting mGlu receptors to improve LID.
Moreover, we recently showed the acute antidyskinetic effectsof the mGlu5 receptor antagonists MPEP, MTEP and AFQ056 inMPTP-lesioned monkeys (Gregoire et al., 2011; Morin et al., 2010).Finally, a recent randomized, double-blind, placebo-controlled,parallel-group and in-patient study of parkinsonian patients withmoderate to severe LID and severe LID on stable dopaminergictherapy has been performed, with patients receiving 25e150 mg ofAFQ056 or placebo twice daily for 16 d (Berg et al., 2011). AFQ056showed a clinically relevant and significant antidyskinetic effectwithout changing the antiparkinsonian effects of dopaminergictherapy. AFQ056 was well tolerated, and adverse events such asdizziness and worsening of LID associated with loss of adherence totreatment or discontinuation were the most common adverseevents reported in the study (Berg et al., 2011).
4.3. MPEP effect on dyskinesia and parkinsonism
In the present chronic study, L-DOPA administered to MPTP-lesioned monkeys relieved their parkinsonian symptoms. Theantiparkinsonian activity of L-DOPA and MPEP combination wasgenerally maintained in MPTP-lesioned monkeys during the wholemonth of treatment, as shown by similar locomotion and anti-parkinsonian scores, compared to L-DOPA alone, suggesting thatmonkeys were exposed to similar levels of L-DOPA in the brain.Mean dyskinesia scores significantly increased over the one-monthtreatment in the L-DOPA-treated group compared to L-DOPA þMPEP-treated monkeys for each of the 4 respective weeks.The duration of the L-DOPA antiparkinsonian effect was alsomeasured as it is well known that this duration shortens inparkinsonian patients after prolonged use, a motor complicationcalled wearing-off (Jankovic and Stacy, 2007; Pahwa and Lyons,2009). In the present study, the duration of the L-DOPA anti-parkinsonian effect decreased in the L-DOPA-treated group,consistent with wearing-off, while no significant change was notedupon the further addition of MPEP. Except for LID, no abnormalbehavior was observed at any time during L-DOPA or L-DOPA þ MPEP chronic treatments.
The present study represents the first report of the pharmaco-logical antagonism of mGlu5 receptors with the selective antago-nist MPEP in female primates lesioned with MPTP using de novochronic (one-month) treatment that shows inhibition of LIDexpression without shortening the duration of the L-DOPA anti-parkinsonian effect (i.e. wearing-off). There is only a single othersimilar report in male rhesus MPTP-lesioned monkeys showingthat de novo administration of the mGlu5 receptor antagonistfenobam for 17 d to four drug-naïve animals attenuated the
development of peak-dose LID without compromising L-DOPAantiparkinsonian effect (Rylander et al., 2010). In the latter de novostudy, development of LID in MPTP-lesioned primates treated withL-DOPA plus fenobam was reduced by w50% compared to L-DOPA,but the duration of the L-DOPA antiparkinsonian effect was notinvestigated. We also observed an inhibition of LID developmentwith MPEP with an averaged 72% less LID for the four weeks in fivede novo MPTP-lesioned monkeys during our one-month experi-ment. Hence, in MPTP-lesioned monkeys of both sexes, an mGlu5receptor antagonist is able to reduce the development of LID in L-DOPA-treated animals.
4.4. Pharmacokinetics of MPEP
The plasma MPEP concentrations measured during and at theend of theMPEP treatment showed noMPEP accumulation over themonth of treatment and indicated that the most elevated MPEPblood concentrations coincided with the occurrence of the LIDpeak. The latter observation shows that MPEP was administered atan appropriate time, i.e. immediately before L-DOPA administra-tion, and that its plasma titer did not require to be maintainedbeyond the duration of the L-DOPA antiparkinsonian effect toprevent development of LID. Moreover, this is the first studyshowing a negative correlation between LID and the blood levels ofan mGlu5 receptor antagonist (MPEP). Our observation stronglysuggests that the antidyskinetic effect observed was related toMPEP activity per se and not to interindividual differencesbetween-groups and/or animals in response to L-DOPA treatment.
4.5. Biochemical effects of MPEP treatment
In human postmortem brains, we previously reported that thespecific binding to ionotropic glutamate receptors NMDA andAMPA (Calon et al., 2003b) as well as to the mGlu5 receptor(Ouattara et al., 2011) increases in parkinsonian patients thatdeveloped LID and in those that displayed wearing-off, suggestingsome commonmechanism involving glutamate in these twomotorcomplications. By contrast, other receptors such as adenosine A2A(Calon et al., 2004) and GABAA (Calon et al., 2003a) or peptidemRNA such as preproenkephalin (Calon et al., 2002) were reportedto be increased in parkinsonian patients with LID, but not in thoseshowing wearing-off. In the present study, MPEP prevented thedevelopment of LID. Moreover, MPEP maintained the duration ofthe L-DOPA antiparkinsonian effect in MPTP-lesioned monkeyswhile it was decreased in the group of monkeys treated with L-DOPA alone, thus exhibiting a wearing-off effect. This is the firstobservation of a dual effect of an mGlu5 receptor antagonist on LIDdevelopment and on the duration of L-DOPA antiparkinsonianeffect in primates, which is in accordance with human postmortemfindings in the brain of parkinsonian patients with LID andwearing-off. This dual effect of MPEP is better than that observedwith the positive comparator amantadine, the glutamatergic drugused to inhibit LID in parkinsonian patients (Goetz et al., 2005;Sawada et al., 2010; Thomas et al., 2004). Indeed, amantadine hasbeen reported to reduce LID in MPTP-lesioned monkeys whileworsening their antiparkinsonian response (Blanchet et al., 1998;Rylander et al., 2010).
mGlu5 receptor specific binding increased in the putamen in theL-DOPA-treated dyskinetic monkeys compared to control and salinemonkeys, as previously reported by our group using two differentradioligands (Ouattara et al., 2011; Samadi et al., 2008). By contrast,[3H]ABP688 specific binding did not increase in monkeys treatedwith L-DOPA þ MPEP compared to control monkeys and untreatedMPTP-lesioned monkeys, suggesting that MPEP prevented the LID-related increase in mGlu5 receptor levels and associated LID.

N. Morin et al. / Neuropharmacology 66 (2013) 355e364 363
Moreover, there was a significant negative correlation betweenmean dyskinesia scores and [3H]ABP688 specific binding in theputamen of MPTP-lesioned monkeys. In rodent models of PD,changes of striatal mGlu5 receptors in LID have not been investi-gated, but striatal molecular changes relevant to LID have beenreported to be reversed by MPEP or MTEP, including delta FosBprotein (Jimenez et al., 2009), prodynorphin mRNA (Mela et al.,2007), glutamic acid decarboxylase (GAD65 and GAD67) mRNA(Yamamoto and Soghomonian, 2009) and phosphorylated extra-cellular signal-regulated kinases 1 and 2 (ERK1/2) protein levels(Rylander et al., 2009). By counteracting abnormal molecularresponses in striatal neurons, mGlu5 receptor antagonists mightnormalize a hyperactive GABAergic transmission in the basalganglia (Mela et al., 2007), thus opposing LID expression (Cenci andLindgren, 2007). In the present study with MPTP-lesionedmonkeys, a chronic treatment with L-DOPA þ MPEP was able tostabilize mGlu5 receptor levels in the putamen at control values,thus consequently could maintain a normal glutamate stimulatoryneurotransmission and its downstream molecular changes associ-ated with LID.
4.6. Conclusion
The present study shows a beneficial chronic antidyskineticeffect of mGlu5 receptor antagonism in L-DOPA-treated MPTP-lesioned monkeys. This supports the therapeutic use of an mGlu5receptor antagonist to normalize brain glutamate neurotransmis-sion in PD and prevent LID. Chronic treatment with an mGlu5receptor antagonist could prevent the development of LID andmight constitute the first therapy available to control LID in PD.Taken together, results in rodent models and results presentedherein with MPTP-lesioned monkeys argue in favor of the contri-bution of mGlu5 receptors in the pathophysiology of LID and clearlyindicate that blockade of this receptor might constitute a noveltherapeutic treatment for LID in PD. To this date, no therapy has yetbeen approved for LID treatment.
Acknowledgments
We would like to thank Dr. Richard Poulin for providinglanguage help, writing assistance and proof reading the article.
References
Berg, D., Godau, J., Trenkwalder, C., Eggert, K., Csoti, I., Storch, A., Huber, H., Morelli-Canelo, M., Stamelou, M., Ries, V., Wolz, M., Schneider, C., Di Paolo, T.,Gasparini, F., Hariry, S., Vandemeulebroecke, M., Abi-Saab, W., Cooke, K.,Johns, D., Gomez-Mancilla, B., 2011. AFQ056 treatment of levodopa-induceddyskinesias: results of 2 randomized controlled trials. Mov. Disord. 26,1243e1250.
Bezard, E., Brotchie, J.M., Gross, C.E., 2001. Pathophysiology of levodopa-induceddyskinesia: potential for new therapies. Nat. Rev. Neurosci. 2, 577e588.
Blanchet, P.J., Konitsiotis, S., Chase, T.N., 1998. Amantadine reduces levodopa-induced dyskinesias in parkinsonian monkeys. Mov. Disord. 13, 798e802.
Blanchet, P.J., Metman, L.V., Chase, T.N., 2003. Renaissance of amantadine in thetreatment of Parkinson’s disease. Adv. Neurol. 91, 251e257.
Blandini, F., Armentero, M.T., 2012. New pharmacological avenues for the treatmentof L-DOPA-induced dyskinesias in Parkinson’s disease: targeting glutamate andadenosine receptors. Expert Opin. Investig. Drugs 21, 153e168.
Breysse, N., Baunez, C., Spooren, W., Gasparini, F., Amalric, M., 2002. Chronic but notacute treatment with a metabotropic glutamate 5 receptor antagonist reversesthe akinetic deficits in a rat model of parkinsonism. J. Neurosci. 22, 5669e5678.
Calon, F., Birdi, S., Rajput, A.H., Hornykiewicz, O., Bedard, P.J., Di Paolo, T., 2002.Increase of preproenkephalin mRNA levels in the putamen of Parkinson diseasepatients with levodopa-induced dyskinesias. J. Neuropathol. Exp. Neurol. 61,186e196.
Calon, F., Dridi, M., Hornykiewicz, O., Bedard, P.J., Rajput, A.H., Di Paolo, T., 2004.Increased adenosine A2A receptors in the brain of Parkinson’s disease patientswith dyskinesias. Brain 127, 1075e1084.
Calon, F., Morissette, M., Rajput, A.H., Hornykiewicz, O., Bedard, P.J., Di Paolo, T.,2003a. Changes of GABA receptors and dopamine turnover in the postmortem
brains of parkinsonians with levodopa-induced motor complications. Mov.Disord. 18, 241e253.
Calon, F., Rajput, A.H., Hornykiewicz, O., Bedard, P.J., Di Paolo, T., 2003b. Levodopa-induced motor complications are associated with alterations of glutamatereceptors in Parkinson’s disease. Neurobiol. Dis. 14, 404e416.
Cenci, M.A., Lindgren, H.S., 2007. Advances in understanding L-DOPA-induceddyskinesia. Curr. Opin. Neurobiol. 17, 665e671.
Chase, T.N., Oh, J.D., 2000. Striatal mechanisms and pathogenesis of parkinsoniansigns and motor complications. Ann. Neurol. 47, S122eS129. discussionS129eS30.
Conn, P.J., Battaglia, G., Marino, M.J., Nicoletti, F., 2005. Metabotropic glutamatereceptors in the basal ganglia motor circuit. Nat. Rev. Neurosci. 6, 787e798.
Conn, P.J., Pin, J.P., 1997. Pharmacology and functions of metabotropic glutamatereceptors. Annu. Rev. Pharmacol. Toxicol. 37, 205e237.
de Lau, L.M., Breteler, M.M., 2006. Epidemiology of Parkinson’s disease. LancetNeurol. 5, 525e535.
Dekundy, A., Pietraszek, M., Schaefer, D., Cenci, M.A., Danysz, W., 2006. Effects ofgroup I metabotropic glutamate receptors blockade in experimental models ofParkinson’s disease. Brain Res. Bull. 69, 318e326.
Fabbrini, G., Brotchie, J.M., Grandas, F., Nomoto, M., Goetz, C.G., 2007. Levodopa-induced dyskinesias. Mov. Disord. 22, 1379e1389. quiz 1523.
Fuxe, K., Marcellino, D., Rivera, A., Diaz-Cabiale, Z., Filip, M., Gago, B.,Roberts, D.C., Langel, U., Genedani, S., Ferraro, L., de la Calle, A., Narvaez, J.,Tanganelli, S., Woods, A., Agnati, L.F., 2008. Receptorereceptor interactionswithin receptor mosaics. Impact on neuropsychopharmacology. Brain Res.Rev. 58, 415e452.
Goetz, C.G., Poewe, W., Rascol, O., Sampaio, C., 2005. Evidence-based medicalreview update: pharmacological and surgical treatments of Parkinson’s disease:2001 to 2004. Mov. Disord. 20, 523e539.
Gottwald, M.D., Aminoff, M.J., 2011. Therapies for dopaminergic-induced dyskine-sias in Parkinson disease. Ann. Neurol. 69, 919e927.
Goulet, M., Morissette, M., Grondin, R., Falardeau, P., Bedard, P.J., Rostene, W., DiPaolo, T., 1999. Neurotensin receptors and dopamine transporters: effects ofMPTP lesioning and chronic dopaminergic treatments in monkeys. Synapse 32,153e164.
Gravius, A., Dekundy, A., Nagel, J., More, L., Pietraszek, M., Danysz, W., 2008.Investigation on tolerance development to subchronic blockade of mGluR5 inmodels of learning, anxiety, and levodopa-induced dyskinesia in rats. J. NeuralTransm. 115, 1609e1619.
Gregoire, L., Morin, N., Ouattara, B., Gasparini, F., Bilbe, G., Johns, D., Vranesic, I.,Sahasranaman, S., Gomez-Mancilla, B., Di Paolo, T., 2011. The acute anti-parkinsonian and antidyskinetic effect of AFQ056, a novel metabotropicglutamate receptor type 5 antagonist, in L-Dopa-treated parkinsonian monkeys.Parkinsonism Relat. Disord. 17, 270e276.
Gubellini, P., Pisani, A., Centonze, D., Bernardi, G., Calabresi, P., 2004. Metabotropicglutamate receptors and striatal synaptic plasticity: implications for neuro-logical diseases. Prog. Neurobiol. 74, 271e300.
Hadj Tahar, A., Gregoire, L., Darre, A., Belanger, N., Meltzer, L., Bedard, P.J., 2004.Effect of a selective glutamate antagonist on L-dopa-induced dyskinesias indrug-naive parkinsonian monkeys. Neurobiol. Dis. 15, 171e176.
Hintermann, S., Vranesic, I., Allgeier, H., Brulisauer, A., Hoyer, D., Lemaire, M.,Moenius, T., Urwyler, S., Whitebread, S., Gasparini, F., Auberson, Y.P., 2007.ABP688, a novel selective and high affinity ligand for the labeling of mGlu5receptors: identification, in vitro pharmacology, pharmacokinetic and bio-distribution studies. Bioorg. Med. Chem. 15, 903e914.
Jankovic, J., Stacy, M., 2007. Medical management of levodopa-associated motorcomplications in patients with Parkinson’s disease. CNS Drugs 21, 677e692.
Jimenez, A., Bonastre, M., Aguilar, E., Marin, C., 2009. Effect of the metabotropicglutamate antagonist MPEP on striatal expression of the Homer family proteinsin levodopa-treated hemiparkinsonian rats. Psychopharmacology (Berl.) 206,233e242.
Johnston, T.H., Fox, S.H., McIldowie, M.J., Piggott, M.J., Brotchie, J.M., 2010. Reductionof L-DOPA-induced dyskinesia by the selective metabotropic glutamate receptor5 antagonist 3-[(2-methyl-1,3-thiazol-4-yl)ethynyl]pyridine in the 1-methyl-4-phenyl-1,2,3,6-tetrahydropyridine-lesioned macaque model of Parkinson’sdisease. J. Pharmacol. Exp. Ther. 333, 865e873.
Klockgether, T., Turski, L., 1993. Toward an understanding of the role of glutamate inexperimental parkinsonism: agonist-sensitive sites in the basal ganglia. Ann.Neurol. 34, 585e593.
Krystal, J.H., Karper, L.P., Seibyl, J.P., Freeman, G.K., Delaney, R., Bremner, J.D.,Heninger, G.R., Bowers Jr., M.B., Charney, D.S., 1994. Subanesthetic effects of thenoncompetitive NMDA antagonist, ketamine, in humans. Psychotomimetic,perceptual, cognitive, and neuroendocrine responses. Arch. Gen. Psychiatry 51,199e214.
Levandis, G., Bazzini, E., Armentero, M.T., Nappi, G., Blandini, F., 2008. Systemicadministration of an mGluR5 antagonist, but not unilateral subthalamic lesion,counteracts L-DOPA-induced dyskinesias in a rodent model of Parkinson’sdisease. Neurobiol. Dis. 29, 161e168.
Meissner, W.G., Frasier, M., Gasser, T., Goetz, C.G., Lozano, A., Piccini, P., Obeso, J.A.,Rascol, O., Schapira, A., Voon, V., Weiner, D.M., Tison, F., Bezard, E., 2011.Priorities in Parkinson’s disease research. Nat. Rev. Drug Discov. 10, 377e393.
Mela, F., Marti, M., Dekundy, A., Danysz, W., Morari, M., Cenci, M.A., 2007. Antag-onism of metabotropic glutamate receptor type 5 attenuates L-DOPA-induceddyskinesia and its molecular and neurochemical correlates in a rat model ofParkinson’s disease. J. Neurochem. 101, 483e497.

N. Morin et al. / Neuropharmacology 66 (2013) 355e364364
Meldrum, B.S., 1998. The glutamate synapse as a therapeutical target: perspectivesfor the future. Prog. Brain Res. 116, 441e458.
Meldrum, B.S., 2000. Glutamate as a neurotransmitter in the brain: review ofphysiology and pathology. J. Nutr. 130, 1007Se1015S.
Morin, N., Gregoire, L., Gomez-Mancilla, B., Gasparini, F., Di Paolo, T., 2010. Effect ofthe metabotropic glutamate receptor type 5 antagonists MPEP and MTEP inparkinsonian monkeys. Neuropharmacology 58, 981e986.
Morissette, M., Dridi, M., Calon, F., Hadj Tahar, A., Meltzer, L.T., Bedard, P.J., DiPaolo, T., 2006. Prevention of levodopa-induced dyskinesias by a selectiveNR1A/2B N-methyl-D-aspartate receptor antagonist in parkinsonian monkeys:implication of preproenkephalin. Mov. Disord. 21, 9e17.
Olanow, C.W., Stern, M.B., Sethi, K., 2009. The scientific and clinical basis for thetreatment of Parkinson disease. Neurology 72, S1e136.
Ossowska, K., Konieczny, J., Wolfarth, S., Pilc, A., 2005. MTEP, a new selectiveantagonist of the metabotropic glutamate receptor subtype 5 (mGluR5),produces antiparkinsonian-like effects in rats. Neuropharmacology 49, 447e455.
Ouattara, B., Gasparini, F., Morissette, M., Gregoire, L., Samadi, P., Gomez-Mancilla, B., Di Paolo, T., 2010. Effect of L-Dopa on metabotropic glutamatereceptor 5 in the brain of parkinsonian monkeys. J. Neurochem. 113, 715e724.
Ouattara, B., Gregoire, L., Morissette, M., Gasparini, F., Vranesic, I., Bilbe, G.,Johns, D.R., Rajput, A., Hornykiewicz, O., Rajput, A.H., Gomez-Mancilla, B., DiPaolo, T., 2011. Metabotropic glutamate receptor type 5 in levodopa-inducedmotor complications. Neurobiol. Aging 32, 1286e1295.
Page, G., Peeters, M., Najimi, M., Maloteaux, J.M., Hermans, E., 2001. Modulation ofthe neuronal dopamine transporter activity by the metabotropic glutamatereceptor mGluR5 in rat striatal synaptosomes through phosphorylation medi-ated processes. J. Neurochem. 76, 1282e1290.
Pahwa, R., Lyons, K.E., 2009. Levodopa-related wearing-off in Parkinson’s disease:identification and management. Curr. Med. Res. Opin. 25, 841e849.
Paquet, M., Smith, Y., 2003. Group I metabotropic glutamate receptors in themonkey striatum: subsynaptic association with glutamatergic and dopami-nergic afferents. J. Neurosci. 23, 7659e7669.
Perroy, J., Raynaud, F., Homburger, V., Rousset, M.C., Telley, L., Bockaert, J., Fagni, L.,2008. Direct interaction enables cross-talk between ionotropic and group Imetabotropic glutamate receptors. J. Biol. Chem. 283, 6799e6805.
Pisani, A., Gubellini, P., Bonsi, P., Conquet, F., Picconi, B., Centonze, D., Bernardi, G.,Calabresi, P., 2001. Metabotropic glutamate receptor 5 mediates the potentia-tion of N-methyl-D-aspartate responses in medium spiny striatal neurons.Neuroscience 106, 579e587.
Platt, S.R., 2007. The role of glutamate in central nervous system health and diseasee a review. Vet. J. 173, 278e286.
Rouse, S.T., Marino, M.J., Bradley, S.R., Awad, H., Wittmann, M., Conn, P.J., 2000.Distribution and roles of metabotropic glutamate receptors in the basal gangliamotor circuit: implications for treatment of Parkinson’s disease and relateddisorders. Pharmacol. Ther. 88, 427e435.
Rylander, D., Iderberg, H., Li, Q., Dekundy, A., Zhang, J., Li, H., Baishen, R., Danysz, W.,Bezard, E., Cenci, M.A., 2010. A mGluR5 antagonist under clinical developmentimproves L-DOPA-induced dyskinesia in parkinsonian rats and monkeys. Neu-robiol. Dis. 39, 352e361.
Rylander, D., Recchia, A., Mela, F., Dekundy, A., Danysz, W., Cenci, M.A., 2009.Pharmacological modulation of glutamate transmission in a rat model of L-DOPA-induced dyskinesia: effects on motor behavior and striatal nuclearsignaling. J. Pharmacol. Exp. Ther. 330, 227e235.
Samadi, P., Gregoire, L., Bedard, P.J., 2003. Opioid antagonists increase the dyskineticresponse to dopaminergic agents in parkinsonian monkeys: interactionbetween dopamine and opioid systems. Neuropharmacology 45, 954e963.
Samadi, P., Gregoire, L., Morissette, M., Calon, F., Hadj Tahar, A., Dridi, M.,Belanger, N., Meltzer, L.T., Bedard, P.J., Di Paolo, T., 2008. mGluR5 metabotropicglutamate receptors and dyskinesias in MPTP monkeys. Neurobiol. Aging 29,1040e1051.
Samadi, P., Rouillard, C., Bedard, P.J., Di Paolo, T., 2007. Functional neurochemistry ofthe basal ganglia. Handb. Clin. Neurol. 83, 19e66.
Sawada, H., Oeda, T., Kuno, S., Nomoto, M., Yamamoto, K., Yamamoto, M.,Hisanaga, K., Kawamura, T., 2010. Amantadine for dyskinesias in Parkinson’sdisease: a randomized controlled trial. PLoS One 5, e15298.
Sheffler, D.J., Gregory, K.J., Rook, J.M., Conn, P.J., 2011. Allosteric modulation ofmetabotropic glutamate receptors. Adv. Pharmacol. 62, 37e77.
Siderowf, A., Stern, M., 2003. Update on Parkinson disease. Ann. Intern. Med. 138,651e658.
Smith, Y., Charara, A., Paquet, M., Kieval, J.Z., Pare, J.F., Hanson, J.E., Hubert, G.W.,Kuwajima, M., Levey, A.I., 2001. Ionotropic and metabotropic GABA and gluta-mate receptors in primate basal ganglia. J. Chem. Neuroanat. 22, 13e42.
Stocchi, F., Tagliati, M., Olanow, C.W., 2008. Treatment of levodopa-induced motorcomplications. Mov. Disord. 23 (Suppl. 3), S599eS612.
Szabo, J., Cowan, W.M., 1984. A stereotaxic atlas of the brain of the cynomolgusmonkey (Macaca fascicularis). J. Comp. Neurol. 222, 265e300.
Testa, C.M., Standaert, D.G., Young, A.B., Penney Jr., J.B., 1994. Metabotropic gluta-mate receptor mRNA expression in the basal ganglia of the rat. J. Neurosci. 14,3005e3018.
Thomas, A., Iacono, D., Luciano, A.L., Armellino, K., Di Iorio, A., Onofrj, M., 2004.Duration of amantadine benefit on dyskinesia of severe Parkinson’s disease.J. Neurol. Neurosurg. Psychiatry 75, 141e143.
Verhagen Metman, L., Del Dotto, P., van den Munckhof, P., Fang, J., Mouradian, M.M.,Chase, T.N., 1998. Amantadine as treatment for dyskinesias and motor fluctu-ations in Parkinson’s disease. Neurology 50, 1323e1326.
Yamamoto, N., Soghomonian, J.J., 2009. Metabotropic glutamate mGluR5receptor blockade opposes abnormal involuntary movements and theincreases in glutamic acid decarboxylase mRNA levels induced by L-DOPA instriatal neurons of 6-hydroxydopamine-lesioned rats. Neuroscience 163,1171e1180.