Embed Size (px)
Citation preview

J. Cell Sci. 10, 563-583 (1972) 563
Printed in Great Britain
MOVEMENT, FINE STRUCTURE, AND FUSION
OF PSEUDOPODS OF AN ENCLOSED AMOEBA,
DIFFLUGIELLA SP.
J. L. GRIFFIN
Department of Anatomy, Harvard Medical School, Boston, Massachusetts 02115, andthe * Armed Forces Institute of Pathology, Washington, D.C. 20305, U.S.A.
SUMMARY
In Difflugiella sp., strain F-20, a small amoeba enclosed in a flexible mantle, pseudopodsextended through a mouth or aperture and seemed to function only for movement and feeding.Pseudopods from different cells fused on contact and cell clumps shared common pseudopodsand moved in a co-ordinated way. During locomotion, pseudopods or pseudopod complexesusually exhibited an activity cycle of 3 phases: anterior extension with the tip firmly adhering;stable hold as other pseudopods advanced; and flaccid posterior retraction. While distaladhesive tips advanced, proximal unattached parts of pseudopods simultaneously shortened asthe cell body advanced.
Microtubules were numerous in pseudopods within the mouth but extended for only1-2 /an into pseudopods up to 20-30 /an long. Microfilaments were present where pseudopodsadhered to the substratum, to the mantle, or to bacteria and were also associated withpinocytotic invaginations. Pseudopod ground plasm was either reticulate or amorphous; noaxial rods or aligned filaments related to pseudopod rigidity were seen.
Simultaneous pseudopod adhesion, extension, and proximal shortening apparently accountfor locomotion or cell body translation of Difflugiella. While some similarities to other amoeboidsystems were noted, the need for detailed studies on different types of organisms or cells isemphasized.
INTRODUCTION
Much recent evidence indicates that actin is present in Acanthamoeba (Pollard,Shelton, Weihing & Korn, 1970), Amoeba (Pollard & Korn, 1971), acellular slimemoulds (Hatano, Totsuka & Oosawa, 1967; Adelman & Taylor, 1969; Nachmias,Huxley & Kessler, 1970), cellular slime moulds (Woolley, 1970), and metazoan cells(Ishikawa, Bischoff& Holtzer, 1969). Myosin is also present in slime moulds (Hatano& Ohnuma, 1970; Adelman & Taylor, 1969; Nachmias & Ingram, 1970) and may bein thick filaments in amoebae (Pollard & Ito, 1970; Holberton & Preston, 1970).
Although amoeboid movement apparently involves energy-linked alterations oforganized substructure, the new molecular information does not necessarily clarifythe relationship between motility and visualized fine structure.
Bundles of actin microfilaments in the slime mould Physarum polycephalum areappropriately located to contract and squeeze out streaming cytoplasm (Wohlfarth-Bottermann, 1964a, b; Rhea, 1966; Porter, Kawakami & Ledbetter, 1965; Nagai &
• Present address.

564 J. L. GriffinKamiya, 1966), but the presence of filament bundles large enough to be seen bypolarizing microscopy is inversely related to streaming and vigorously streamingplasmodia may have none (Nakajima, 1964; Nakajima & Allen, 1965); aggregatesappear during pinocytosis (Bauer, 1967; Griffin, 1967).
Filaments of 2 sizes are present in amoebae fixed in locomotion (Pollard & Ito,1970) but are more numerous in non-motile, pinocytosing amoebae [Chaos andAmoeba) and in amoebae broken during fixation (Nachmias, 1964; Griffin, 1965).Numerous filaments seen after enzyme treatments (Komnick & Wohlfarth-Botterman,1965) are probably related to pinocytosis induced by the enzymes rather than tobetter fixation permitted by membrane changes. Schafer-Danneel (1967) bestvisualized filaments in amoebae fixed while cold and unable to move. In the abovesystems, filament aggregates seem to differentiate under conditions in which move-ment slows and stops. Combining isolated thin (F-actin) and thick filaments fromA. proteus under appropriate conditions does produce dramatic movements (Wolpert,1965; Pollard & Ito, 1970).
Filaments (about 6-nm) have been seen in the enclosed amoebae Difflugia(Wohlman & Allen, 1968) and Hyalosphenia (Joyon & Charret, 1962) and a flabellulidamoeba (probably Vannella) called Hyalodiscus simplex (Wohlfarth-Botterman,1964a, b). Bhowmick (1967) visualized plasma filaments of about 20 nm in Saccamoebasp., strain F-13, Griffin; this amoeba also contains numerous 6-nm filaments (personalobservation). Various slime moulds (McManus & Roth; 1965), the giant amoebaPelomyxa palustris (Griffin, 1965; Daniels & Breyer, 1967), and other amoebae, nakedand shelled (personal observation), also contain microfilaments and microtubules,which could play structural, contractile or other roles.
Microtubules seem to serve a structural or skeletal role in Heliozoa (Tilney &Byers, 1969; Roth & Shigenaka, 1970), Foraminifera (McGee-Russell & Allen, 1971),pigment cells (Bikle, Tilney & Porter, 1966) and blood cells (Fawcett & Witebsky,1964), and microfilaments (F-actin) may have a similar role in microspikes andmicrovilli (Taylor, 1966; Ishikawa et al. 1969) and acanthopodia (Pollard et al. 1970).In cells of the plant Nitella, filament bundles lie parallel to the stream at the shearinterface (Nagai & Rebhun, 1966; Rebhun, 1967).
At present, correlated light- and electron-microscopic studies suggest directinvolvement of filament aggregates in one tension-generating system (Wohlman &Allen, 1968) and an active shearing system (Nagai & Rebhun, 1966). Cytochalasin Bhas recently been used to probe structure-function relationships. Wessells et al.(1971) review the literature and unpublished work and conclude that this compoundreversibly inhibits contractile microfilaments in many systems and that reticulatenetworks, not aligned aggregates, are contractile in individual cells. However,membrane changes in pinocytosis influence microfilaments and cytochalasin can altercell-environment interactions (Carter, 19676), which suggests that secondary as wellas primary changes might be detected after application of this chemical. Studies ofcytochalasin effects at the molecular level should be interesting.
Comparative studies are needed because of the great diversity of parameters ofamoeboid movement (see Allen & Kamiya, 1964). It seems important to relate

Movement and fine structure of an enclosed amoeba 565
visualized fine structure to patterns of movement in living cells. Difflugiella sp. alsooffers some advantages beyond that of a comparative approach. Organelles andinclusions are enclosed within a capsule, from which pseudopods extend, so pseudopodultrastructure should reflect the functions of movement and feeding. Fixation of thinpseudopods should be rapid, thereby improving the chances of preserving ultra-structure in a lifelike state.
MATERIALS AND METHODS
Difflugiella sp., strain F-20, Griffin, was isolated from a mixed culture of Amoeba proteus andgrown on i % non-nutrient agar with Aerobacter, strain A-154, added as food. An unidentifiedspore-farming bacterium was present in these cultures. Assignment of F-20 to the genusDifflugiella is based primarily on a discussion by Page (1966). A flexible test or mantle ischaracteristic of this genus. Since F-20 is small and the pseudopods seem unique, furtherstudies may well justify a generic reassignment. Euglypha sp. used for comparisons is strainF-54, Griffin.
For light microscopy, amoebae were usually observed in the inorganic medium of Prescott& Carrier (1964) with Zeiss (Jena) phase and Nomarski optics. Movement was recorded bycinematography or by sequential photographs, usually at 5-s intervals. An electronic flash(Fawcett & Ito, 1958) was used for some still photography.
Amoebae were fixed at room temperature by rapidly pouring fixative over amoebae growingand migrating on agar. The micrographs presented here were from amoebae fixed either with2 % unbuffered osmium tetroxide plus 0002 M CaCl, for 5 min or with 125 % glutaraldehyde,collidine 01 M, pH 77, with 0002 M CaCla for 2 h, followed by unbuffered OsO4 for 5 min.The fixative is indicated in the caption for each figure. Some material was perhaps extracted bythe procedure used, but the calcium seemed to stabili2e components of interest (see Wood &Luft, 1965, and Griffin, 1963). For in-block staining with uranyl acetate, cells were fixed for1 h in 4% glutaraldehyde in 0-05 M PO4) pH 69, 20 min in OsO4 in 005 M PO4, rinsed twicein 01 N sodium acetate and stained 20 min in 3 % aqueous uranyl acetate (Terzakis, 1968).After fixation, agar wedges with adhering amoebae were cut out, dehydrated in ethanol, passedthrough propylene oxide into Epon 812, and embedded open face. Sections were usuallystained with a saturated solution of uranyl acetate in water followed by lead citrate (Venablc &Coggeshall, 1965).
RESULTS AND OBSERVATIONS
As in other enclosed amoebae, Difflugiella has a relatively stable organelle distri-bution. At the oral end, pseudopods extend from the specialized mouth (Figs. 1, 5-9)near which contractile vacuoles are located (Figs. 1, 5). Granules and food or digestionvacuoles form a band across the middle of the cell (Fig. 1). The nucleus, with a singlecomplex nucleolus, is located at the adoral end (Fig. 5). Clumps of fused cells (as inFigs. 4 and 5) are sometimes made up of a dozen or more cells which share commonpseudopods and move in a co-ordinated way. No indications were seen of nuclear ororganelle exchanges between cell bodies.
Light-microscopic observations on movement
A detailed presentation of specific movement patterns is made in Figs. 1-4 andtheir captions.
Commonly, several pseudopods or pseudopod complexes extended from beneathan erect body, mostly extending within a half circle in the direction of locomotion,remaining until newer, more anterior complexes had formed, then withdrawing from

566 J. L. Griffinthe region behind the cell body. Amoebae in locomotion showed characteristicpseudopodial patterns (Figs. 1-4). On glass, pseudopods were frequently straight withsmooth outlines (Figs. 2-4), while more irregular outlines seemed to result from acloser adhesion to agar. Thin webs frequently formed and retracted between pseudo-pods, particularly larger fused pseudopods (Fig. 4). In motion pictures, sequentialphotographs and direct observations, only pseudopods extending in contact with thesubstratum were seen to influence locomotion. F-20 can form free, unattachedpseudopods, but, as in flabellulid amoebae (Bovee, 1964), these are not involved inlocomotion. An extending pseudopod (moderate adhesion) was preceded by anadhesive tip that was flattened, somewhat irregular, and relatively narrow (Figs. 1-4).The cell body moved in a direction determined by the advance of multiple pseudopodsand the distance between the cell body and stationary branches of individual pseudo-pods shortened up to 50% while the tip was still extending (Figs. 1, 2). This earlyproximal shortening was distinct in time and position from posterior withdrawal. Asa pseudopod became fully formed and reached a temporarily stable stage, it seemed tothicken and straighten (Fig. 2, top). Withdrawing pseudopods were usually detachedand flaccid (Fig. 2B, c). Zipper-like fusion of adjacent pseudopods frequently precededretraction (Fig. 2C, D). Retraction was normally faster than extension (Fig. 4A, B);pseudopods up to 30 fim in length sometimes withdrew within 1-2 s. The ingestion ofa bacterium seemed to involve a simple movement of adhering membrane over theparticle, without formation of a food cup (Fig. 4).
When pseudopods touched, fusion was immediate, whether they originated fromthe same cell mass or from difFerent cells (Fig. 1). Although fusion occurred whenamoebae met, the possibility exists that aggregates may have derived in part fromdivisions in which the cell body, but not the pseudopods, separated. The separation offused pseudopods seemed similar to normal retraction. Usually, as amoebae movedapart, the fused pseudopod detached from the substratum. Such fused pseudopodssometimes seemed to be under tension, but then parted slowly near the middlewithout elastic rebound. Some contacts between pseudopods seemed to initiate animmediate response that caused separation within 10—15 s after fusion (Fig. 1). Non-compressed amoebae did not differ in fusion and separation patterns from the gentlycompressed cells in Fig. 1.
Ultrastructure of pseudopods
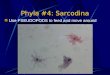
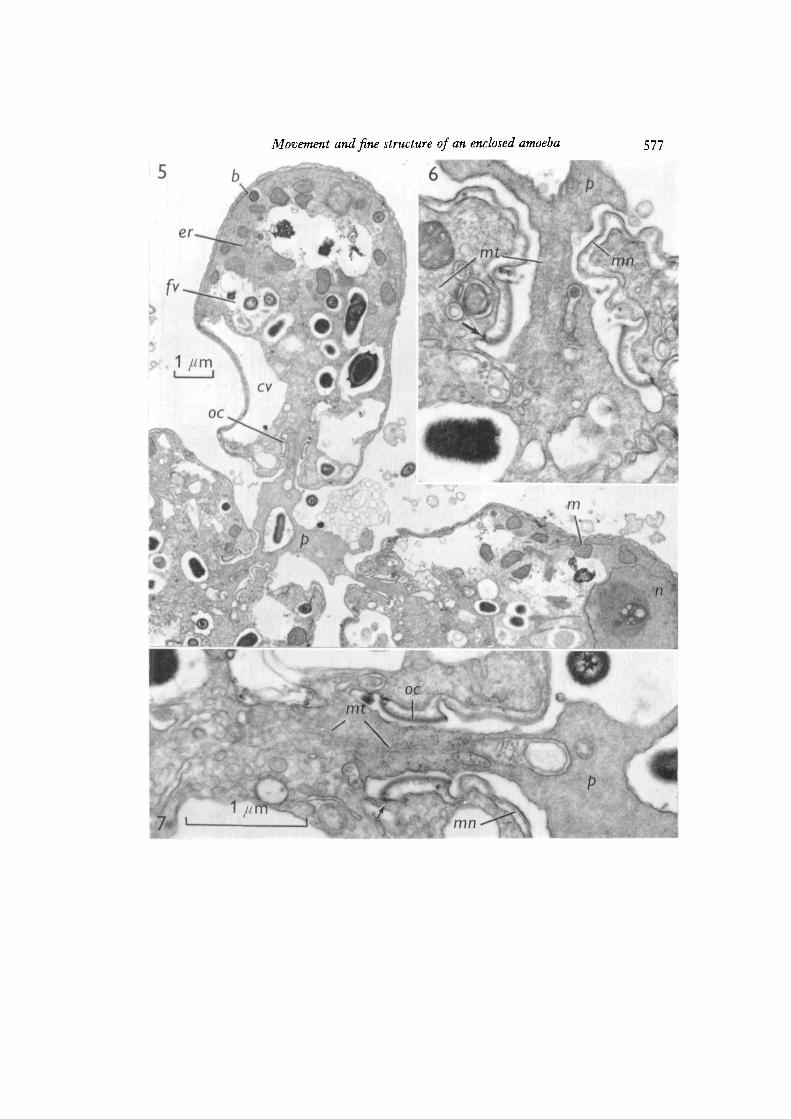
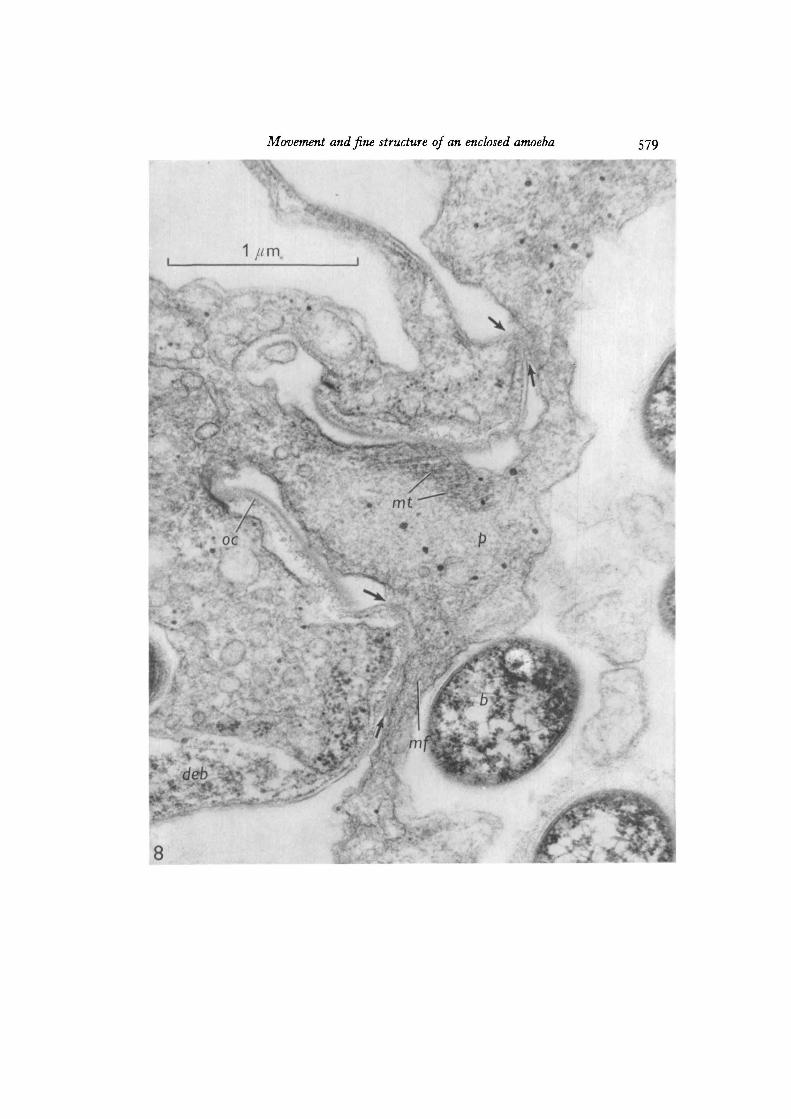
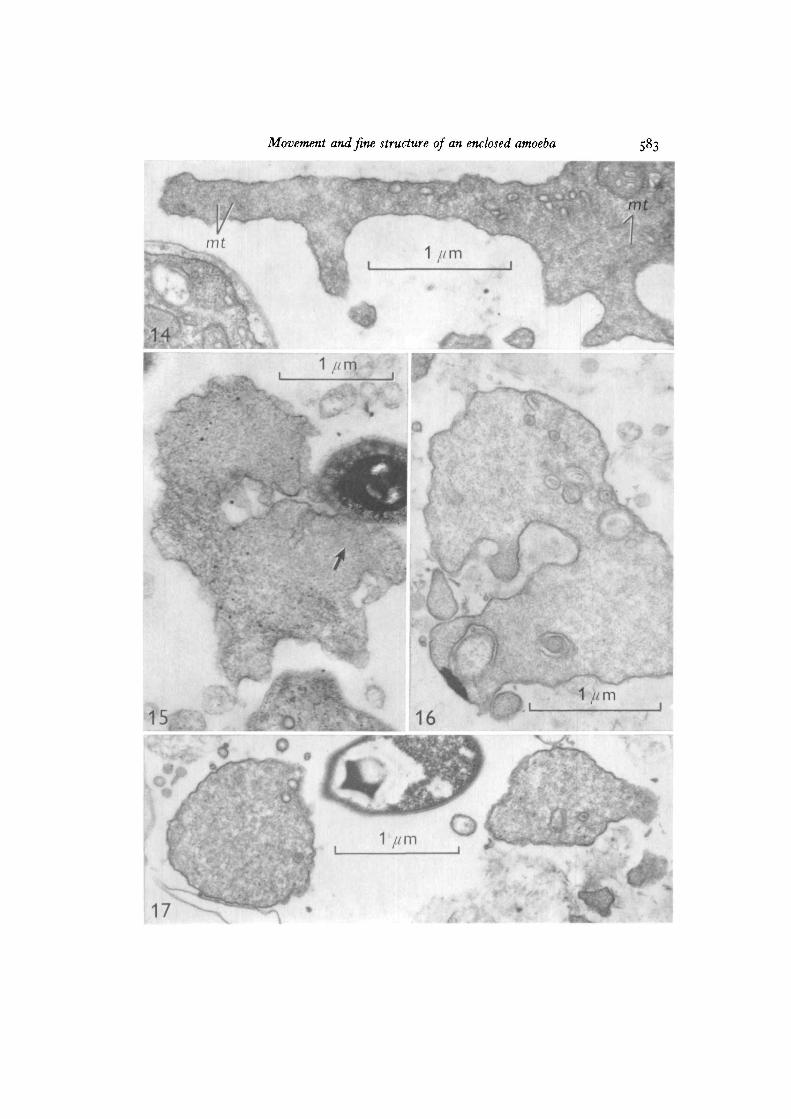
Electron micrographs showed that full cytoplasmic continuity was establishedbetween cells sharing pseudopods (Fig. 5). Pseudopods contained few organelles orgranules, although mitochondria were seen near the mouth (Figs. 9, 14). Microtubules,microfilaments, areas with a reticulate texture, and gradients in electron density wereseen within the pseudopods after fixation with osmium or glutaraldehyde (Figs. 6-9,14-17). Although numerous (over 40 in one cross-section), microtubules were presentin pseudopods emerging from the mouth (Figs. 6—8), at a distance of 1 /*ra they wereindistinct (Figs. 6, 7, 14), and none were observed more than 2 /tm away from themouth (Figs. 6-9, 14-17). Some microtubules seemed to anchor the junction of themembrane and the oral collar (Fig. 9).

Movement and fine structure of an enclosed amoeba 567
Microfilaments in pseudopods were usually near regions, presumably adhesive,adjacent to the agar substratum (Figs. 9, 16), to the mantle of the amoeba (Figs. 8, 9)or to bacteria (Fig. 15). Microfilaments also oriented towards membrane imaginations(Fig. 9), but otherwise seemed to be present only at or near sites of adhesion.
The mantle surrounding the cell body is composed of 3 layers: (1) an externalelectron-dense layer about 9-10 nm thick, (2) a directly underlying amorphous layerof about 20 nm, and (3) just within the amorphous layer, punctate and circular pro-files, possibly representing filaments and tubules in cross-section (Figs. 8, 9). At themouth, the mantle is continuous with the slightly more complex oral collar (Figs. 5—9).The membrane at oral collar-membrane junctions stained intensely (Figs. 5-9) andserved as an attachment for 20-nm microtubules (Fig. 9).
In micrographs of most species of amoebae processed and photographed as inFigs. 6-9 and 14-17, internal membranes usually appear as single lines, while thickerexternal membranes are relatively easy to resolve as trilaminar 'unit membranes', asshown in micrographs of Euglypha (Figs. 11, 12). In micrographs of F-20 a trilaminarstructure of external membranes is hard to detect (Fig. 13) except after uranium inblock (Fig. 10).
In other samples there is slight variation in dimensions as reported for Figs. 10—13,but differences in both thickness and contrast are reproducible. The external membraneon pseudopods and inside the mantle thus differs from that of most other smallamoebae in both structure and behaviour (fusion).
DISCUSSION
Patterns of movement
While it is clear that specimens of Difflugiella exhibit a distinct type or pattern ofamoeboid movement and characteristic fine structure, direct evidence related tomolecular interactions or mechanisms of movement is lacking. No internal or externalparticles that might reveal relative movements of pseudopod protoplasm or membranewere seen in Difflugiella, except during retraction. The dissociation of cellular systems,useful in studies of other amoebae (Pollard et al. 1970; Pollard & Ito, 1970; Pollard &Korn, 1971; Wolpert, 1965; Allen, Cooledge & Hall, i960; Griffin, 1964), was notattempted.
The locomotion of Difflugia, a much larger enclosed amoeba, involves sequentialextension of unattached pseudopods, adhesion at the tip, and shortening to exerttraction (Wohlman & Allen, 1968; Mast, 1931). Because Difflugia drags a heavy shellmade of sand grains, its locomotory organelles or mechanisms are presumed to bespecialized for heavy work loads.
In Difflugiella, adhesion, pseudopod extension, and shortening (to exert traction ?)occur simultaneously rather than sequentially. While the adherent tips of pseudopodsare still extending, proximal shortening decreases the distance between the cell bodyand the stationary regions of attachment. Posterior withdrawal of flaccid pseudopodsoccurs later and does not contribute to cell body advance. Proximal shortening duringpseudopod extension presumably accounts for cell-body translation in the directionof locomotion.

568 J. L. GriffinBecause similarities are lacking, it seems unlikely that Difflugiella could utilize,
without modification, mechanisms proposed to account for movement of loboseamoebae (Allen, 1961; Allen, Francis & Nakajima, 1965; Alien & Kamiya, 1964;Jahn & Bovee, 1969; Noland, 1957) or Foraminifera (McGee-Russell & Allen, 1971;Allen, 1964; Jahn & Rinaldi, 1959).
Difflugiella can form relatively rigid free or unattached hyaline pseudopods. Thesemay be analogous to the hyaline pseudopods of Vannella and other mayorellids (Bovee,1964), although their movements are not as complex. The free pseudopods ofDifflugiella do not contribute to locomotion. In locomotion, all extending pseudopodsadhere and normally exhibit a finely irregular or fuzzy appearance at the advancingtip-
A significant feature of movement of Difflugiella is the close relationship betweenpseudopod extension and adhesion, a correlation also seen in Vannella and otheramoebae normally led by a flattened anterior fan (Bovee, 1964). In these amoebae,the membrane moves forward over the cell and remains stationary under the advancingbody like a tank tread (Griffin & Allen, i960; Griffin, 1970). In an abstract (Griffin,1970), a modified frontal contraction (see Allen, 1961; Allen et al. 1965; Allen &Kamiya, 1964) was suggested as compatible with the advance of the adhering hyalinefan of Vanella. This conceptual model also seems compatible with the advance of theadhering tip of hyaline Difflugiella pseudopods, but it is quite possible that thecompatibility is apparent only because little direct evidence is available. Of course,internal consistency and compatibility with available evidence are not enough toprove a theory correct. Metazoan cells also show a close correlation between adhesionand extension of hyaline peripheral fans (Taylor, 1966; Buckley & Porter, 1967;Carter, 1967a; Wessells et al. 1971).
Possible functional roles of visualized fine structure
It seems reasonable to assume that the fine structure visualized was present in theliving state. Light microscopy revealed no gross distortion and the amoebae weremoving normally just prior to rapid fixation at room temperature. The thin pseudopodsand simple membranes of Difflugiella should present no special problems in fixation.It is encouraging that fixation with either glutaraldehyde or osmium preservedessentially similar patterns in pseudopods of Difflugiella. A second assumption is thatthe activity of the cell at the instant of fixation was correctly inferred from themorphology of the embedded cell, and from light- and electron-microscopic sections.The cells sectioned were not directly visualized during fixation and processing(Griffin, 1963).
Ground cytoplasm of pseudopods. Regions of pseudopods interpreted as groundcytoplasm (not adjacent to adhesive regions) appeared either amorphous or reticulate,with reticulate filamentous elements less than 5 nm in diameter. The difference be-tween reticulate and amorphous areas may reflect either different physical states of theprotoplasm or merely differences in preservation and visualization. For example,complete preservation and staining of soluble proteins could make it impossible tovisualize a diffuse reticulate ground structure.

Movement and fine structure of an enclosed amoeba 569
The resistance to deformation of pseudopods seems to be based on a reticulatesubstructure, rather than on components such as microfilaments or microtubules. Thesubstructure of Difflugiella pseudopods seems not to differ in any significant way fromthe protoplasmic substructure of hyaline regions of many different cell types.
Microfilaments. Relatively straight microfilaments of about 6 nm were seen nearwhere membranes adhered to the substratum, the mantle, or bacteria and in regionsof pinocytosis. They seemed not to be involved in maintaining configuration ofelongate pseudopods or other structures. In many micrographs a gradual transitionof cytoplasmic texture was seen, ranging from a diffuse reticulate background to moredistinct microfilaments adjacent to apparent areas of adhesion. Microfilaments maybe an alternative configuration of reticulate material.
The microfilaments seem analagous to those formed in larger amoebae in responseto pinocytosis inducers, such as alcian blue (Nachmias, 1964) and enzymes (Komnick& Wohlfarth-Botterman, 1965), and may reflect a cytoplasm-membrane bondingrequired either for pinocytosis or to reinforce a site of adhesion to the substratum.Taylor (1966) saw both microfilaments and microtubules adjacent to sites of adhesionof tissue culture cells. Light-microscopic views of pseudopod complexes of Difflugiella(as in Fig. 4) with webs between the pseudopods look like webs between stress fibresin tissue culture cells (Buckley & Porter, 1967), but fine-structural similarities werenot seen. Thick microfilaments (Pollard & Ito, 1970) were not seen.
Microtubules. Microtubules in the mouth of Difflugiella could act as skeletal elementsor support translational machinery for moving vacuoles through the mouth (compareRebhun, 1967). Microtubules anchoring the collar-membrane junction probably helpto maintain the configuration of the cell body. Microtubules were not seen in pseudo-pods so apparently do not contribute to pseudopod rigidity, as in heliozoans (Tilney &Byers, 1969; Roth & Shigenaka, 1970) and foraminiferans (McGee-Russell & Allen,1971). The microtubules are clear and distinct only in the region within 1 fim of themouth, suggesting that something in that region may act as an organizer for micro-tubule differentiation. Their absence in pseudopods does not seem to be a fault inpreservation, since both types of fixative give the same distribution.
Membrane. The membrane of Difflugiella differs from external membranes of mostamoebae both in behaviour and in thickness and contrast. Immediate fusion was seenwhenever pseudopod contact was observed. In most species of amoebae, the externalmembrane shows clear differentiation into a trilaminar' unit membrane' configuration.A trilaminar configuration was seen in the external membranes of Difflugiella onlyafter uranyl acetate in block or when osmium-fixed material was sectioned verycarefully.
Cellular fusion is important in many examples of morphogenesis, but reversiblefusion of pseudopods only has apparently not been reported. The larger pseudopodsof clumps may permit more effective food gathering or locomotion under certaincircumstances.

570 J. L. Griffin
CONCLUSIONS
That pseudopod adhesion, extension, and shortening account for locomotion is notsurprising, since it is difficult to imagine any amoeboid progression without similarprocesses. Although micronlaments were adjacent to sites of membrane adhesion,neither micronlaments nor microtubules seem involved in other pseudopodialfunction. Reticulate pseudopod substructure may represent some aspect of themachinery for movement, if such machinery was preserved and visualized. Thisconclusion would be compatible with concepts advanced by Wessells et al. (1971).
Note. Some of the material herein was presented to the American Society of Cell Biology(Griffin, 1968). In a recent review, Jahn & Bovee (1969) wrote 'This stereoplasmic track inDifflugiella is a bundle of micronlaments that develop in only the attached portions of thefilopods (194).' Although reference 194 is to Griffin (1968), there is no stereoplasmic track inDifflugiella and no known reason for their statement.
This study was supported in part by NIH Research Grant AIO 3410, an NIH SpecialFellowship, an NIH Departmental Training grant, and small grants from the Milton Fund(all at Harvard Medical School); and a Research Contract, Project No. 3A062110A822, fromthe Medical Research and Development Command, U.S. Army, Washington, D.C. Theopinions or assertions contained herein are the private views of the author and are not to beconstrued as official or as reflecting the views of the Department of the Army or the Departmentof Defense.
REFERENCESADELMAN, M. R. & TAYLOR, E. W. (1969). Further purification and characterization of slime
mold myosin and slime mold actin. Biochemistry 8, 4976-4988.ALLEN, R. D. (1961). Ameboid movement. In The Cell, vol. 2 (ed. J. Brachet & A. E. Mirsky),
PP- !3S~2i6. New York and London: Academic Press.AXLEN, R. D. (1964). Cytoplasmic streaming and locomotion in marine Foraminifera. In
Primitive Motile Systems in Cell Biology (ed. R. D. Allen & N. Kamiya), pp. 407-432. NewYork: Academic Press.
ALLEN, R. D., COOLEDGE, J. W. & HALL, P. J. (i960). Streaming in cytoplasm dissociated fromthe giant amoeba Chaos chaos. Nature, Lond. 187, 896-899.
ALLEN, R. D., FRANCIS, D. W. & NAKAJIMA, H. (1965). Cyclic birefringence changes inpseudopods of Chaos carolinensis revealing the localization of the motive force in pseudopodextension. Proc. natn. Acad. Sci. U.S.A. 54, 1153-1161.
ALLEN, R. D. & KAMIYA, N., eds. (1964). Primitive Motile Systems in Cell Biology. New York:Academic Press.
BAUER, L. G. (1967). On the similar orientation of fibrillar structures and rows of esterolyticinvaginations in plasmodia of the slime mold, Physarum confertum Macbr. J. exp. Zool. 164,60-80.
BHOWMICK, D. K. (1967). Electron microscopy of Trichamoeba villosa and amoeboid move-ment. Expl Cell Res. 45, 570-589.
BIKLE, D., TILNEY, L. G. & PORTER, K. R. (1966). Microtubules and pigment migration in themelanophores of Fundulus heteroclitus L. Protoplasma 61, 322-345.
BOVEE, E. C. (1964). Morphological differences among pseudopodia of various small amoebaeand their functional significance. In Primitive Motile Systems in Cell Biology (ed. R. D. Allen& N. Kamiya), pp. 189-219. New York: Academic Press.
BUCKLEY, I. K. & Porter, K. R. (1967). Cytoplasmic fibrils in living cultured cells. A light andelectron microscopic study. Protoplasma 64, 349-380.

Movement and fine structure of an enclosed amoeba 571
CARTER, S. B. (1967a). Haptotaxis and the mechanism of cell motility. Nature, Lond. 213,256—260.
CARTER, S. B. (1967ft). Effects of cytochalasins on mammalian cells. Nature, Lond. 213, 261-264.DANIELS, E. W. & BREYER, E. P. (1967). Ultrastructure of the giant amoeba Pelomyxa palustris.
J. Protozool. 14, 167-179.FAWCETT, D. W. & ITO, S. (1958). Observations on the cytoplasmic membranes of testicular
cells, examined by phase contrast and electron microscopy. J. biophys. biochem. Cytol. 4,I3S-H2.
FAWCETT, D. W. & WITEBSKY, F. (1964). Observations on the ultrastructure of nucleatederythrocytes and thrombocytes, with particular reference to the structural basis of theirdiscoidal shape. Z. Zellforsch. mikrosk. Anat. 62, 785-806.
GRIFFIN, J. L. (1963). Motion picture analysis of fixation for electron microscopy: Amoebaproteus.jf. Cell Biol. 19, 77 A.
GRIFFIN, J. L. (1964). The comparative physiology of movement in the giant multinucleateamebae. In Primitive Motile Systems in Cell Biology (ed. R. D. Allen & N. Kamiya), pp. 303-321. New York: Academic Press.
GRIFFIN, J. L. (1965). Fixation and visualization of microfilaments and microtubules and theirsignificance in the movement of four types of ameboid cells. J. Cell Biol. 27, 39 A.
GRIFFIN, J. L. (1967). The functional role of microfilament aggregates in the slime moldPhysarum polycephalum. J. Cell Biol. 35, 50—51 A.
GRIFFIN, J. L. (1968). An example of ameboid movement compatible with a mechanism basedon differential adhesion.,?. Cell Biol. 39, 56 A.
GRIFFIN, J. L. (1970). Ameboid movement in anteriorly-flattened amebae. J. Protozool.17 (Suppl.), 15.
GRIFFIN, J. L. & ALLEN, R. D. (i960). The movement of particles attached to the surface ofamebae in relation to current theories of ameboid movement. Expl Cell Res. 20, 619-622.
HATANO, S. & OHNUMA, J. (1970). Purification and characterization of myosin A from themyxomycete plasmodium. Bioc/iim. biophys. Ada 205, 110-120.
HATANO, S., TOTSUKA, T. & OOSAWA, F. (1967). Polymerization of plasmodium actin. Biochim.biophys. Ada 140, 109-122.
HOLBERTON, D. V. & PRESTON, T. M. (1970). Arrays of thick filaments in ATP-activatedAmoeba model cells. Expl Cell Res. 62, 473-477.
ISHIKAWA, H., BISCHOFF, R. & HOLTZER, H. (1969). Formation of arrowhead complexes withheavy meromyosin in a variety of cell types. J. Cell Biol. 43, 312-328.
JAHN, T. L. & BOVEE, E. C. (1969). Protoplasmic movements within cells. Physiol. Rev. 49,793-862.
JAHN, T. L. & RINALDI, R. A. (1959). Protoplasmic movement in the foraminiferan, AUogromialaticollaris; and a theory of its mechanism. Biol. Bull. mar. biol. Lab., Woods Hole 117,100-118.
JOYON, L. & CHARRET, R. (1962). Sur I'ultra8tructure du Thecamoebien Hyalosphenia papilio(Leidy). C. r. hebd. Sianc. Acad. Set., Paris 255, 2661-2663.
KOMNICK, H. & WOHLFARTH-BOTTERMANN, K. E. (1965). Das Grundplasma und diePlasmafilamente de AmSbe Chaos chaos nach enzymatischer Behandlung der Zellmembran.Z. Zellforsch. mikrosk. Anat. 66, 434-456.
MCGEE-RUSSELL, S. M. & ALLEN, R. D. (1971). Reversible stabilization of labile microtubulesin the reticulopodial network of AUogromia. Advances in Cell and Molecular Biology 1,I53-I84-
MCMANUS, Sister M. A. & ROTH, L. E. (1965). Fibrillar differentiation in myxomyceteplasmodia. J. Cell Biol. 25, 305-318.
MAST, S. O. (1931). Movement and response in Difflugia with special reference to the natureof cytoplasmic contraction. Biol. Bull. mar. biol. Lab., Woods Hole 61, 223-241.
NACHMIAS, V. T. (1964). Fibrillar structures in the cytoplasm of Chaos chaos. J. Cell Biol. 23,183-188.
NACHMIAS, V. T., HUXLEY, H. E. & KESSLER, D. (1970). Electron microscope observations onactomyosin and actin preparations from Physarum polycephalum, and on their interactionwith heavy meromyosin subfragment I from muscle myosin. J. molec. Biol. 50, 83-90.
37 CE L 10

572 J. L. Griffin
NACHMIAS, V. T. & INGRAM, W. C. (1970). Actomyosin from Physarum polyceplialum: electronmicroscopy of myosin-enriched preparations. Science, N.Y. 170, 743-745.
NAGAI, R. & KAMIYA, N. (1966). Movement of the myxomycete plasmodium. II . Electronmicroscopic studies on fibrillar structures in the plasmodium. Proc. Japan Acad. 42,934-939-
NAGAI, R. & REBHUN, L. I. (1966). Cytoplasmic microfilaments in streaming Nitella cells. J.Ultrastruct. Res. 14, 571-589.
NAKAJIMA, H. (1964). The mechanochemical system behind streaming in Physarum. InPrimitive Motile Systems in Cell Biology (ed. R. D. Allen & N. Kamiya), pp. m - 1 2 0 . NewYork: Academic Press.
NAKAJIMA, H. & ALLEN, R. D. (1965). The changing pattern of birefringence in plasmodia ofthe slime mold, Physarum polyceplialum. J. Cell Biol. 25, 361-374.
NOLAND, L. E. (1957). Protoplasmic streaming: a perennial puzzle.,?. Protozool. 4, 1-7.PAGE, F. C. (1966). Cryptodifflugia operctilatan.sp. (Rhizopodea: Arcellinida, Cryptodifflugiidae)
and the status of the genus Cryptodifflugia. Trans. Am. microsc. Soc. 85, 506-515.POLLARD, T. D. & ITO, S. (1970). Cytoplasmic filaments of Amoeba proteus. I. The role of
filaments in consistency changes and movement. J. Cell Biol. 46, 267-289.POLLARD, T. D. & KORN, E. D. (1971). Filaments of Amoeba proteus. II. Binding of heavy
meromyosin by thin filaments in motile cytoplasmic extracts. J. Cell Biol. 48, 216—219.
POLLARD, T. D., SHELTON, E., WEIHING, R. R. & KORN, E. D. (1970). Ultrastructural
characterization of F-actin isolated from Acanthamoeba castellanii and identification ofcytoplasmic filaments as F-actin by reaction with rabbit heavy meromyosin. J. molec. Biol.50, 91-97.
PORTER, K. R., KAWAKAMI, N. & LEDBETTER, M. C. (1965). Structural basis of streaming inPhysarum polycephalum. J. Cell Biol. 27, 78 A.
PRESCOTT, D. M. & CARRIER, R. F. (1964). Experimental procedures and cultural methods forEuplotes eurystomus and Amoeba proteus. In Methods in Cell Physiology, vol 1 (ed. D. M.Prescott), pp. 85-95. N e w York: Academic Press.
REBHUN, L. I. (1967). Structural aspects of saltatory particle movement. J. gen. Physiol. 50,223-239-
RHEA, P. (1966). Electron microscopic observations on the slime mold Physarum polyceplialumwith specific reference to fibrillar structures. J. Ultrastruct. Res. 15, 349-379.
ROTH, L. E. & SHIGENAKA, Y. (1970). Microtubules in the heliozoan axopodium. II. Rapiddegradation by cupric and nickelous iona.jf. Ultrastruct. Res. 31, 356—374.
SCHAFER-DANNEEL, S. (1967). Strukturelle und functionelle Voraussetzungen, fur dieBewegung von Amoeba proteus. Z. Zellforsch. mikrosk. Anat. 78, 441-462.
TAYLOR, A. C. (1966). Microtubules in the microspikes and cortical cytoplasm of isolated cells.J. Cell Biol. 28, 155-168.
TERZAKIS, J. A. (1968). Uranyl acetate, a stain and a fixative. J. Ultrastruct. Res. 22, 168-184.
TILNEY, L. G. & BYERS, B. (1969). Studies on the microtubules in Heliozoa. V. Factorscontrolling the organization of microtubules in the axonemal pattern in Ecliinospliaerium(Actinosphaerium) nucleofilum. J. Cell Biol. 43, 148-165.
VENABLE, J. H. & CoGGESHALL, R. (1965). A simplified lead citrate stain for use in electronmicroscopy. J. Cell Biol. 25 (No. 1), 407-408.
WESSELLS, N. K., SPOONER, B. S., ASH, J. F., BRADLEY, M. O., LUDUENA, M. A., TAYLOR, E. L.,
WRENN, J. T . & YAMADA, K. M. (1971). Microfilaments in cellular and developmentalprocesses. Science, N.Y. 171, 135-143.
WOHLFARTH-BOTTERMANN, K. E. (1964a). Cell structures and their significance for ameboidmovement. Int. Rev. Cytol. 16, 61-131.
WOHLFARTH-BOTTERMANN, K. E. (19646). Differentiations of the ground cytoplasm and theirsignificance for the generation of the motive force of ameboid movement. In Primitive MotileSystems in Cell Biology (ed. R. D. Allen & N. Kamiya), pp. 79-108. New York: AcademicPress.
WOHLMAN, A. & ALLEN, R. D. (1968). Structural organization associated with pseudopodextension and contraction during cell locomotion in Difflugia.J. Cell Sci. 3, 105-114.

Movement and fine structure of an enclosed amoeba 573
WOLPERT, L. (1965). Cytoplasmic streaming and amoeboid movement. Symp. Soc. gen.Microbiol. 15, 270-293.
WOOD, L. & LUFT, J. H. (1965). The influence of buffer systems on fixation with osmiumtetroxide. J. Ultrastrtict. Res. 12, 22-45.
WOOLLEY, D. E. (1970). Extraction of an actomyosin-like protein from amoebae of Dictyo-stelium discoideum.J. cell. Pliysiol. 76, 185-190.
[Received 21 June 1971)
37-2

574 J- L- Griffin
Fig. i A, B. Two sequential phase-contrast photographs of living Diffktgiella sp. gentlycompressed against soft agar so that locomotion continued. In each cell, pseudopodsemerged from the mouth and extended in close contact with the agar. Contractilevacuoles lie adjacent to the mouth (top left of cell a). Nuclei are not in clear focus.
Two different responses to pseudopodial fusion were exhibited by cell a. Contactand pseudopod fusion with cell b (Fig. i A, left arrow) was immediately followed by awithdrawal response. Within 3 s (Fig. 1 B), both parts of the shared pseudopod haddetached from the agar and formed a straight connecting pseudopod. Some irregularbeading followed, and the pseudopods parted without rebound about 9 s after the firstcontact. In contrast, pseudopodial fusion between cells a and c (Fig. 1 A, right arrow)was not followed by rapid withdrawal and remained for nearly 60 s, apparently partingonly as part of the normal extension-withdrawal cycle. Cells a and d did not fuse,since only the cell mantles touched. Cell d exhibits 2 pseudopodial complexes, oneextending (left), one withdrawing (right); the left pseudopod exhibits slight proximalshortening with movement of the cell body about 05 fim to the left between A and B.(Illustrations: AFIP Negs. 69-7644-1-8) x 1350.
Figs. 2-4. Phase-contrast pictures, focused on the pseudopods, of cells moving on aglass slide. Alphabetic sequences were photographed at 5-s intervals.
Fig. 2A-D. In A and B the upper pseudopod is extending and is preceded by aflattened, somewhat irregular, adhesive region. In c and D this pseudopod is fullyextended, straight, somewhat thicker near the tip, and lacks any obvious adhesiveregion. In the bottom pseudopod, minute adhesive fans are present at the tip in A andB. In c the distal part is detached and folded upward and continues shortening in D,while a secondary adhesion at the bend is maintained. During the sequence A-D anotherpseudopodial complex advanced toward the right, just above the centre of the cell.New extensions were relatively narrow and preceded by formation of small adhesivefans. No pseudopods extended into the medium above the slide. Proximal shorteningcan be measured in sequence B-D in the pseudopod complex to the right. The base ofthe branch pseudopod is stationary with reference to bacteria adhering to the slide.The cell body is about 6 fim from this branch in B and about 3 /tm from it in D,10 s later, x 750.
Fig. 3 A—D. The cell and field of view are the same as in Fig. 2. Fig. 3 A was 30 slater than Fig. 2D. During the interval, the upper pseudopod retracted by about halfits length and re-extended to the left of its position in Fig. 2. In Fig. 3 A the upperpseudopod is in the hold position. Five seconds later (B) small lateral adhesionsformed on this pseudopod to become, within 5 s (c), a branch pseudopod; 5 s later (D)the 2 had fused and retraction had started. The lower right pseudopodial complex inFig. 3 was derived from the centre right complex in Fig. 2. In 3A-C the lower branchis extending, led by a minute adhesive flattening. In c 2 pseudopodial branches of thiscomplex were fused, and in D all 3 had fused in a zipper-like action, prior to retraction.X7S°-
Fig. 4A-C. Bacteria adhering to the substratum form reference points for 3 fusedcells moving by means of shared pseudopods. Features noted in Figs. 2 and 3 can alsobe seen in shared pseudopods in this sequence. The larger amount of cytoplasmavailable to multiple cell clusters permits more extensive protoplasmic sheets, whichform and retract between pseudopods (top right). The 4 bacteria (2 at arrows) withinthe pseudopods were initially attached to the glass and were covered and detachedas pseudopods extended. A bacterium, just contacted in Fig. 4A (arrow), was coveredand detached within 5 s (Fig. 4B). The bacteria were carried under the cell bodies aspseudopods retracted. Two single pseudopods pointing toward the upper left andbottom right can be seen in Fig. 4A; within 5 s (B) both these pseudopods hadwithdrawn, x 750.

Movement and fine structure of an enclosed amoeba 575

576 J. L. Griffin
Fig. 5. Electron micrograph of parts of 3 cells that share a pseudopod (p). Each cell iscovered by a mantle that joins an oral collar (oc) at the mouth. Within the cell bodiesare seen mitochondria (m), endoplasmic reticulum (er), food vacuoles(/t>), cytoplasmicbacteria (6), contractile vacuoles (cv), and a nucleus («) with a complex nucleolus.Glutaraldehyde. x 10000.Figs. 6, 7. Enlargements of the mouths of the left and upper cells in Fig. 5. One sideof the oral collar (oc) is directly connected to the mantle (mri), while the other isanchored to the plasma membrane at a region of increased density (arrow). In thepseudopod (p) within the mouth are 16—18 nm microtubules (mt). No microtubuleswere seen in the shared pseudopod outside the mouth. Vesicular material within theemerging pseudopods is seen in tangential section to be in close contact with micro-tubules. Glutaraldehyde. x 3Z000.
Movement and fine structure of an enclosed amoeba 577

578 J. L. Griffin
Fig. 8. In this longitudinal section a pseudopod emerges from the mouth and divergesup and down. A parallel array of 20—22 run microtubules (mt) lies within thepseudopod (p). Above and below the mouth are regions of apparent pseudopod-mantleadhesion (between arrows) associated with microfilamentous densities (ntf) in thepseudopod. The wall of the bacterium (b) can be resolved into 5 layers. Apparentdebris (deb) lies between the mantle and the plasma membrane, oc, oral collar.Osmium tetroxide. x 50000.
Movement and fine structure of an enclosed amoeba
« \
S79
\
oc
mt•
;*•./
deb
8
S

580 J. L. Griffin
Fig. 9. In this longitudinal section through the mouth, 18-22 nm microtubules (mi)extend into the cytoplasm from the junction of the oral collar and the cell membrane.Microfilaments (mf) are concentrated along the right side of the pseudopod, where thepseudopod is adjacent to and apparently adheres to the agar. Increased membranedensity and associated microfilaments are present in an apparent pinocytoticinvagination (pi). Dense microfilaments (arrow) in the upper part of the pseudopodseem to be adjacent to a site of pseudopod-mantle adhesion. A mitochondrion (in)lies just outside the mouth within the pseudopod. Osmium tetroxide. x 38000.
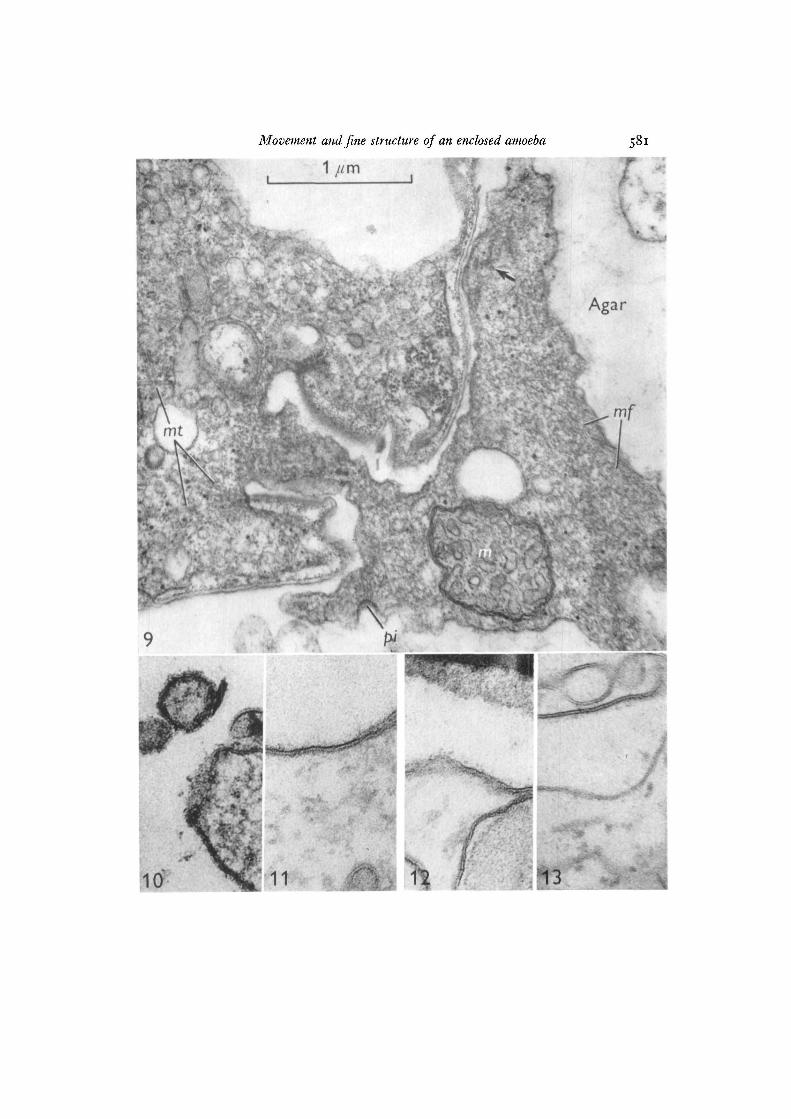
Figs. 10-13. Micrographs printed at the same magnification to show trilaminar unitmembranes of Difflugiella and Eiiglyplia.
Fig. 10 shows pseudopod membrane (9-10 nm) of Difflugiella fixed with glutaralde-hyde and stained in block with uranyl acetate. This special stain reveals the trilaminarstructure but the membrane is not as thick or as easily seen as the pseudopodmembrane (12-13 nm), osmium fixed, of Euglyplia in Fig. 11. Fig. 12 shows themembrane (9-10 nm) of Euglypha inside the shell (shell visible at top), fixed withosmium, compared with the membrane of Difflugiella inside the mantle, Fig. 13, alsofixed with osmium. In Fig. 13, note the pale appearance of the Difflugiella membrane(centre, 8-10 nm) as compared with the myelin figure membrane (top, 9-11 nm),apparently produced by digestion of bacteria and excreted beneath the mantle,x 114000.
Movement and fine structure of an enclosed amoeba
1 /ivn
581
UAgar
mf
10
Movement and fine structure of an enclosed amoeba S83
15
17
\16
* o
1 //m