Embed Size (px)
Citation preview

RESEARCH ARTICLE
Mouse ZAR1-Like (XM_359149) ColocalizesWith mRNA Processing Components and ItsDominant-Negative Mutant CausedTwo-Cell-Stage Embryonic ArrestJianjun Hu,1,2 Fengchao Wang,2 Xiaoquan Zhu,2 Ye Yuan,2 Mingxiao Ding,1* and Shaorong Gao2*
Maternal effect genes and encoding proteins are necessary for nuclear reprogramming and zygotic genomeactivation. However, the mechanisms that mediate these functions are largely unknown. Here we identifiedXM_359149, a Zar1-like gene that is predominantly expressed in oocytes and zygotes, which we designatedZar1-like (Zar1l). ZAR1L-EGFP formed multiple cytoplasmic foci in late two-cell-stage embryos. Expressionof the ZAR1L C-terminus induced two-cell-stage embryonic arrest, accompanied with abnormal methylationof histone H3K4me2/3 and H3K9me2/3, and marked down-regulation of a group of chromatin modificationfactors including Dppa2, Dppa4, and Piwil2. When ectopically expressed in somatic cells, ZAR1L colocalizedwith P-body components including EIF2C1(AGO1), EIF2C2(AGO2), DDX6 and LSM14A, and germline-specificchromatoid body components including PIWIL1, PIWIL2, and LIN28. ZAR1L colocalized with ZAR1 andinteracted with human LIN28. Our data suggest that ZAR1L and ZAR1 may comprise a novel family of proc-essing-body/chromatoid-body components that potentially function as RNA regulators in early embryos. De-velopmental Dynamics 239:407–424, 2010. VC 2009 Wiley-Liss, Inc.
Key words: mRNA processing body; two-cell block; Zar1l; Zar1; zygotic genome activation
Accepted 19 October 2009
INTRODUCTION
The maternal factors that accumulate
during oogenesis play pivotal roles in
nuclear reprogramming, zygotic ge-
nome activation, and preimplantation
embryonic development (Schultz, 1993;Aoki et al., 1997; Latham, 1999;Latham and Schultz, 2001; Ma et al.,2001; Hamatani et al., 2004; Minamiet al., 2007; Stitzel and Seydoux,2007). Oogenesis involves a number of
critical events because a growingmouse oocyte is transcriptionally andtranslationally active. A large numberof mRNAs are synthesized and storedto support oocyte maturation and earlypreimplantation embryogenesis and
ABBREVIATIONS AAs amino acids C-body chromatoid-body DAB 3,30-Diaminobenzidine DAPI 40-6-Diamidino-2-phenylindole EGFPenhanced green fluorescent protein FBS fetal bovine serum H3K4me1/2/3 Mono-/Di-/Tri-methylated Histone H3 at Lysine 4 H3K9me2/3Di/Tri- methylated Histone H3 at Lysine 9HRP horseradish peroxidase mRNP mRNA ribonucleoprotein ORF open reading frame P-bodymRNA processing body PFA paraformaldehyde PHD plant homeodomain PHDL cells pancreatic PNA-HSA double-low cells Phospho-Rpb1 phosphorylated Rpb1 at C-terminal Ser2/5 repeats RFP red fluorescent protein RT-PCR reverse transcription polymerase chainreaction.
Additional Supporting Information may be found in the online version of this article.1The Department of Cell Biology and Genetics, College of Life Sciences, Peking University, Beijing, People’s Republic of China2National Institute of Biological Sciences, Beijing, People’s Republic of ChinaGrant sponsor: National High Technology 863; Grant number: 2008AA022311; Grant sponsor: National Natural Science Foundation;Grant number: 30670302.*Correspondence to: Shaorong Gao, National Institute of Biological Sciences, NIBS, Beijing, 102206, P.R. China. E-mail:[email protected] or Mingxiao Ding Department of Cell Biology and Genetics, College of Life Sciences, PekingUniversity, Beijing, 100871, P.R. China E-mail: [email protected]
DOI 10.1002/dvdy.22170Published online 11 December 2009 in Wiley InterScience (www.interscience.wiley.com).
DEVELOPMENTAL DYNAMICS 239:407–424, 2010
VC 2009 Wiley-Liss, Inc.
Dev
elop
men
tal D
ynam
ics

are not used for immediate translation
(Bachvarova, 1985; Wassarman and
Kinloch, 1992). Mature oocytes arrest
in metaphase during their second mei-
otic division (MII stage), which is asso-
ciated with transcriptional shut-down
and reduced translation. Fertilization
triggers the completion of meiosis,
which is followed by the formation of a
one-cell embryo (zygote) containing
haploid paternal and maternal pronu-
clei (Schultz, 1993; Aoki et al., 1997;
Latham, 1999; Latham and Schultz,
2001; Ma et al., 2001; Hamatani et al.,
2004; Minami et al., 2007; Stitzel and
Seydoux, 2007). Each pronucleus
undergoes DNA replication before
entering the first mitosis to produce a
two-cell embryo.Global expression profiling revealed
distinct patterns of maternal RNAdegradation and zygotic genome acti-vation, which includes three transientwaves of de novo transcription: (1) aminor activation before cleavage(minor ZGA), (2) a major activation atthe two-cell-stage (major ZGA), and(3) a major activation preceding thedynamic morphological and func-tional changes that occur during thetransition from morula to blastocyst,which is termed mid-preimplantationgene activation (MGA) (Hamataniet al., 2004). The major ZGA promotesdramatic reprogramming of geneexpression, coupled with the genera-tion of novel transcripts that are notexpressed in oocytes. Thus, thegenetic program governed by mater-nal transcripts/proteins must beswitched to one dominated by tran-scripts/proteins derived from thenewly formed zygotic genome (Schultz,1993; Aoki et al., 1997; Latham, 1999;Latham and Schultz, 2001; Ma et al.,2001; Hamatani et al., 2004; Minamiet al., 2007; Stitzel and Seydoux,2007).
A great amount of maternal effectgenes produce mRNAs or proteinsthat accumulate in the egg duringoogenesis. Nevertheless, a limitednumber of maternal-effect genes havebeen identified in mice: Nlrp5 (Mater,maternal antigen that embryos re-quire) (Tong et al., 2000); Hsf1 (heat-shock factor 1) (Christians et al.,2000); Dnmt1 (DNA methyltransfer-ase 1, oocyte isoform) (Howell et al.,2001); Npm2 (nucleoplasmin 2)
(Burns et al., 2003; De La Fuenteet al., 2004); Dppa3 (Stella) (Payeret al., 2003); Zar1 (zygotic arrest 1)(Wu et al., 2003a); Cdh1 (E-cadherin)(De Vries et al., 2004); Pms2 (Gurtuet al., 2002); Ezh2 (enhancer of zeste2) (Erhardt et al., 2003); Dnmt3a(DNA methyltransferase 3A) (Kanedaet al., 2004); Ube2a (HRA6A) (Roestet al., 2004); and Smarca4 (Brg1)(Bultman et al., 2006). Apart fromSmarca4, Cdh1, Pms2, Ezh2, Dnmt3a,and Ube2a, all of these maternal-effect genes are exclusively expressedin oocytes. In addition, only Nlrp5and Ube2a mutants have similar phe-notypes (two-cell arrest) to that ofSmarca4 maternally depleted embryos.The other mutants primarily arrest atthe one-cell stage (Npm2, Dppa3, Zar1,Hsf1), later stages of preimplantation(Dppa3, Pms2), or during post-implan-tation (Dnmt3a, Dnmt1o) development.Ezh2 mutant exhibits a postnatal phe-notype. Cdh1 mutant appears pheno-typically normal because of rescuing bythe wild-type paternal allele. Meioticmaturation triggers the degradation ofmaternal transcripts. About 90% of thematernal mRNAs have been degradedby the two-cell stage. However, themechanisms that regulate the transla-tion and degradation of maternal tran-scripts are largely unknown.
In the present study, we identifiedXM_359149, a Zar1-like gene that ispredominantly expressed in oocytesand early preimplantation embryos,which we named Zar1-like (Zar1l). Wecharacterized its sub-cellular localiza-tion and its effect on preimplantationdevelopment. Our data showed thatZAR1L formed cytoplasmic foci in latetwo-cell-stage embryos. Its mutantform ZAR1L Cter-Flag-EGFP inducedabnormal epigenetic modifications andgene expression changes in late two-cell-stage embryos, and finally causedtwo-cell-stage arrest. When ectopicallyexpressed in somatic cells, ZAR1L colo-calized with P-body components includ-ing EIF2C1(AGO1), EIF2C2(AGO2),DDX6, and LSM14A, and germline-specific chromatoid body componentsincluding PIWIL1, PIWIL2, andLIN28. ZAR1L colocalized with ZAR1and interacted with human LIN28.Our data suggested that ZAR1L andZAR1 comprise a novel family of P-body/C-body-like structure componentsin late 2-cell embryos.
RESULTS
Zar1l Gene and Protein
Information and Expression
Pattern
The XM_359419 sequence (GeneID:545824; LOCUS: XM_359419) wasidentified in a search for genesthat are preferentially expressed inoocytes and early embryos. Proteinblast analysis showed that theXM_359419 ORF encodes for a ZAR1-like protein, which we have namedZAR1L. By genomic analysis and RT-PCR analysis, we successfully clonedthe full-length ORF of Zar1l (Fig. 1A).Its orthologs have been found in otherorganisms, including humans, dogs,cows, and rats. Mouse ZAR1L exhibitsgreater homology with the predictedhuman ZAR1L protein than with themouse ZAR1 protein (Fig. 1A and B).RT-PCR analysis showed that mouseZar1l has two transcript isoforms andis specifically expressed in adult ovar-ian tissue (Fig. 2A). Moreover, it ispredominantly expressed in oocytesand zygotes. Cloning and sequencingof the full-length ORF showed thatone of the mouse Zar1l isoforms is876 bp (encodes a 291 AA polypeptide)and the other is 982 bp with out-of-frame reading (Fig. 2A and data notshown). RT-PCR analysis showed thatmouse Zar1 gene was also predomi-nantly expressed in oocytes andzygotes. In order to determine theprotein levels of ZAR1L, Western blotwas performed. The results showedthat mouse ZAR1L protein was pre-dominantly expressed in oocytes andzygotes, and was also maintained at acertain level in 2-cell- and 4-cell-stageembryos (Fig. 2B). Only the 876-bptranscript isoform of Zar1l was char-acterized in this study.
Sub-Cellular Localization of
Mouse ZAR1L Protein and Its
Mutants
In order to predict protein domain/structure of ZAR1L, we performedprotein sequence analysis by usingthe PreDom 2006.1 and Super-family 1.73 protein domain predictionprograms, respectively. The PreDomprogram analysis showed that mouseZAR1L protein might have threefunctional domains, one N-terminal
408 HU ET AL.
Dev
elop
men
tal D
ynam
ics

domain (51–103 AAs), one C-terminaldomain (193–241 AAs), and one largerdomain (130–291 AAs, which mightcontain the middle region and the C-terminal region). The Superfamily1.73 program analysis showed thatmouse ZAR1L protein might containone CSE2-like domain (159–190 AAs)
in the middle region, and one atypicalFYVE/PHD zinc finger domain (227–280 AAs) in the C-terminal. Based onthese protein domain predictionresults, we supposed that mouseZAR1L protein should have threefragments that each contained onefunctional domain: a well-conserved
C-terminal region (191–291 AAs, con-taining the FYVE/PHD zinc finger do-main), a conserved N-terminal region(1–111 AAs, containing one functionalunknown domain), and a relativelyconserved middle region (112–190AAs, containing the CSE2-like do-main). Various constructs were de-signed to express wild-type ZAR1Land dominant-negative mutants ofZAR1L (Fig. 2C). To study the sub-cel-lular localization of ZAR1L and itsmutant proteins, as well as their rolesin preimplantation development, invitro transcribed mRNAs were micro-injected into the cytoplasm of GVoocytes, MII oocytes (followed byICSI), and zygotes. The resultsshowed that the ZAR1L-EGFP signalformed cloud-like structures in thecytoplasm of some fully grown GVoocytes (7.8%, 4/51, Fig. 2E) andformed cytoplasmic foci in most(94.9%, 111/117) of the late two-cell-stage embryos (Fig. 2I). ZAR1L-EGFPcytoplasmic foci could not be observedin MI, MII oocytes, and zygotes (Fig.2D, F–H). The C-terminus fragment,however, showed similar cytoplasmicfoci localization (99.2%, 118/119; Fig.2J) with the full-length ZAR1L pro-tein. The N-terminus deleted form,ZAR1L DN-EGFP, was observed toform weak cytoplasmic foci in most ofthe late 2-cell-stage embryos (81.5%,66/81; Fig. 2K). The C249S/C254S-Flag-EGFP point mutant form com-pletely lost its capacity to form cyto-plasmic foci (Fig. 2L). The full-lengthZAR1 predominantly localized to cyto-plasmic foci in late 2-cell-stageembryos (Fig. 2M).
ZAR1L C-Terminus
Expression Induced
Embryonic Arrest at the
Two-Cell Stage
In order to study the function ofZAR1L and its mutants in preimplan-tation embryonic development, themRNAs that encoded full-length andmutant ZAR1L were microinjectedinto zygotes. The results showed thatthe zygotes injected with EGFP (Fig.3A–D), ZAR1L-EGFP (Fig. 3E–H),ZAR1L C249S/C254S-Flag-EGFP (Fig.3Q–T), as well as the full-lengthZAR1-EGFP (Fig. 3U–X), could de-velop to blastocyst stage in vitro.
Fig. 1. Sequence alignment of mouse ZAR1 and ZAR1L proteins, human ZAR1L, and mouseZAR1L proteins. A: Sequence alignment of mouse ZAR1 and ZAR1L proteins. The eight well-conserved cysteines (with # above them) form an atypical plant homeodomain (PHD) zinc fingerdomain. The N-terminal sequences exhibit low homology between mouse ZAR1 and ZAR1L pro-tein (analyzed by DNAMAN). B: Sequence alignment of predicted human and mouse ZAR1L pro-teins. Sequence analysis showed that the ZAR1L protein contains three domains/motifs: one N-terminal function unknown domain, one middle CSE2-like domain, and one C-terminal atypicalFVYE/PHD zinc finger domain. The domains/motifs were analyzed by PreDom (2006.1) programand Superfamily (1.73) program. Based on the domain/motif analysis and the BLASTP alignmentresults, we designed ZAR1L mutants with domain/motif containing fragments. The ZAR1L-Cterfragment contains the well-conserved atypical PHD zinc finger domain (191–291 AAs in mouseZAR1L, boxed). The ZAR1L-DN fragment contains the middle domain (112–190 AAs) and C-ter-minal domain (191–291 AAs), but lacks the N-terminal domain (1–111 AAs, underlined).
ZAR1L REGULATES PREIMPLANTATION DEVELOPMENT 409
Dev
elop
men
tal D
ynam
ics
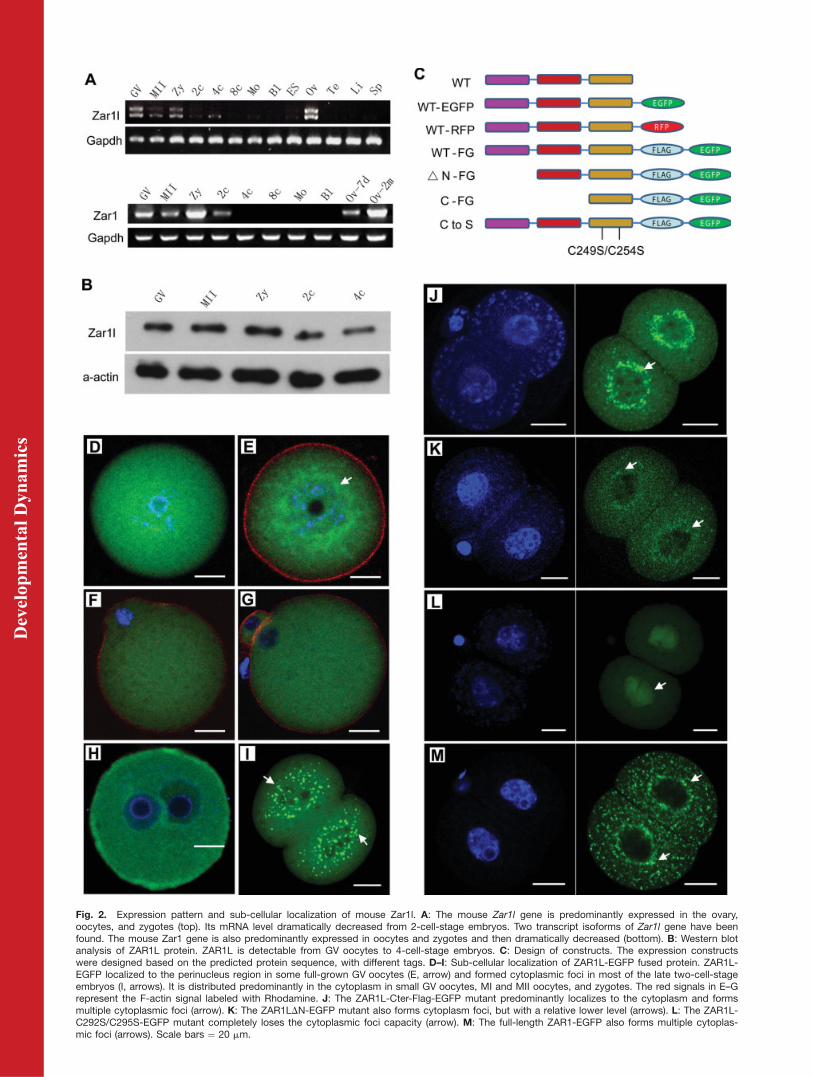
Fig. 2. Expression pattern and sub-cellular localization of mouse Zar1l. A: The mouse Zar1l gene is predominantly expressed in the ovary,oocytes, and zygotes (top). Its mRNA level dramatically decreased from 2-cell-stage embryos. Two transcript isoforms of Zar1l gene have beenfound. The mouse Zar1 gene is also predominantly expressed in oocytes and zygotes and then dramatically decreased (bottom). B: Western blotanalysis of ZAR1L protein. ZAR1L is detectable from GV oocytes to 4-cell-stage embryos. C: Design of constructs. The expression constructswere designed based on the predicted protein sequence, with different tags. D–I: Sub-cellular localization of ZAR1L-EGFP fused protein. ZAR1L-EGFP localized to the perinucleus region in some full-grown GV oocytes (E, arrow) and formed cytoplasmic foci in most of the late two-cell-stageembryos (I, arrows). It is distributed predominantly in the cytoplasm in small GV oocytes, MI and MII oocytes, and zygotes. The red signals in E–Grepresent the F-actin signal labeled with Rhodamine. J: The ZAR1L-Cter-Flag-EGFP mutant predominantly localizes to the cytoplasm and formsmultiple cytoplasmic foci (arrow). K: The ZAR1LDN-EGFP mutant also forms cytoplasm foci, but with a relative lower level (arrows). L: The ZAR1L-C292S/C295S-EGFP mutant completely loses the cytoplasmic foci capacity (arrow). M: The full-length ZAR1-EGFP also forms multiple cytoplas-mic foci (arrows). Scale bars ¼ 20 mm.
Dev
elop
men
tal D
ynam
ics
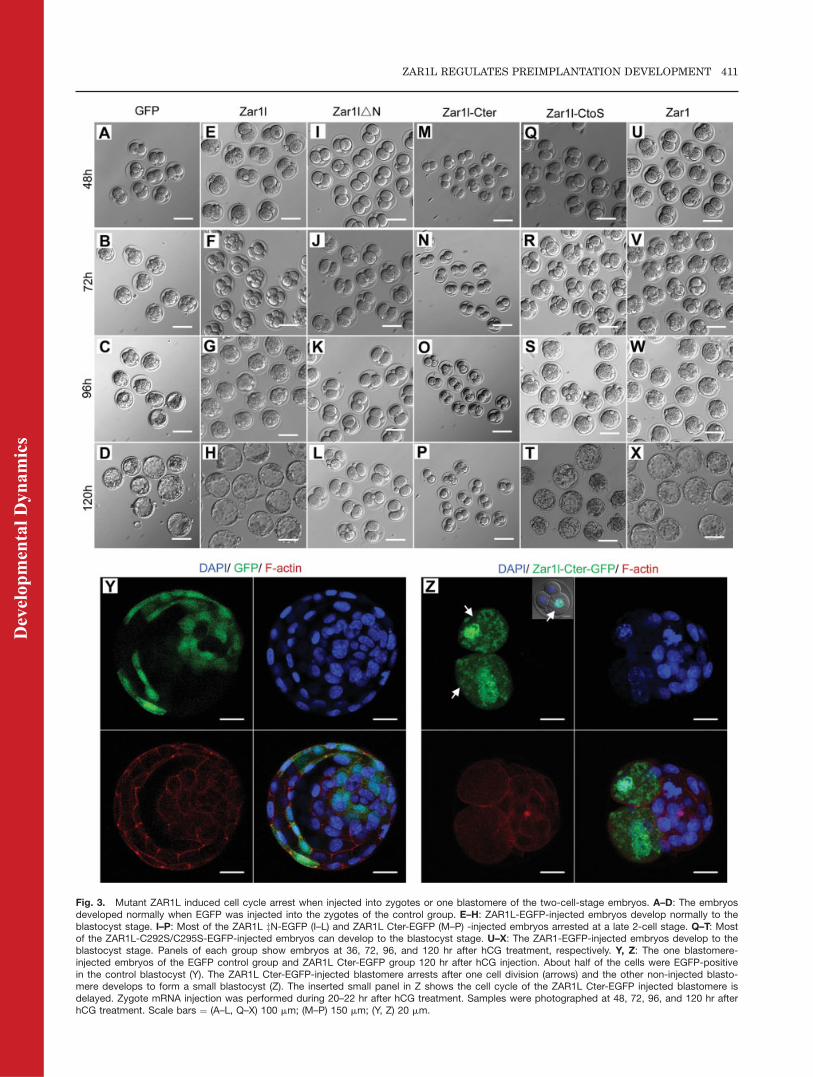
Fig. 3. Mutant ZAR1L induced cell cycle arrest when injected into zygotes or one blastomere of the two-cell-stage embryos. A–D: The embryosdeveloped normally when EGFP was injected into the zygotes of the control group. E–H: ZAR1L-EGFP-injected embryos develop normally to theblastocyst stage. I–P: Most of the ZAR1L zN-EGFP (I–L) and ZAR1L Cter-EGFP (M–P) -injected embryos arrested at a late 2-cell stage. Q–T: Mostof the ZAR1L-C292S/C295S-EGFP-injected embryos can develop to the blastocyst stage. U–X: The ZAR1-EGFP-injected embryos develop to theblastocyst stage. Panels of each group show embryos at 36, 72, 96, and 120 hr after hCG treatment, respectively. Y, Z: The one blastomere-injected embryos of the EGFP control group and ZAR1L Cter-EGFP group 120 hr after hCG injection. About half of the cells were EGFP-positivein the control blastocyst (Y). The ZAR1L Cter-EGFP-injected blastomere arrests after one cell division (arrows) and the other non-injected blasto-mere develops to form a small blastocyst (Z). The inserted small panel in Z shows the cell cycle of the ZAR1L Cter-EGFP injected blastomere isdelayed. Zygote mRNA injection was performed during 20–22 hr after hCG treatment. Samples were photographed at 48, 72, 96, and 120 hr afterhCG treatment. Scale bars ¼ (A–L, Q–X) 100 mm; (M–P) 150 mm; (Y, Z) 20 mm.
ZAR1L REGULATES PREIMPLANTATION DEVELOPMENT 411
Dev
elop
men
tal D
ynam
ics

However, most of the zygotes injectedwith ZAR1L DN-EGFP (80.2%, 89/111; Fig. 3I–L) and ZAR1L Cter-Flag-EGFP (96.9%, 156/161; Fig. 3M–P)arrested at the two-cell stage. WhenEGFP was injected into one blasto-mere of a two-cell-stage embryo, theinjected blastomere was not affectedand developed to blastocyst normally(Fig. 3Y). When ZAR1L Cter-Flag-EGFP was injected into one blasto-mere of a two-cell-stage embryo, theinjected blastomere arrested after onecell division. The other non-injectedblastomere, however, grew to form asmall blastocyst (Fig. 3Z).
ZAR1L Cter-Flag-EGFP
Mutant Did Not Affect BrdU
Incorporation But Affected
BrUTP Incorporation
In order to investigate whether DNAreplication was affected by the ZAR1LCter-Flag-EGFP, BrdU incorporationassay was performed after the first cellcleavage. BrdU staining with anti-BrdU antibody showed that the BrdUincorporation in the ZAR1L Cter-Flag-EGFP group was similar to the NLS-EGFP control group (Fig. 4A and B).
Further, to investigate whether RNAsynthesis was affected by the ZAR1LCter-Flag-EGFP, BrUTP incorporationassay was performed at late 2-cellstage (major zygotic genome activa-tion). BrUTP staining with the anti-BrdU antibody from middle 2-cell stageto late 2-cell stage showed that BrUTPincorporation was moderately down-regulated by ZAR1L Cter-Flag-EGFP(Fig. 4I and J) at late 2-cell embryos.Statistics analysis showed that BrUTPincorporation levels in Zar1l Cter-EGFP injected embryos were down-regulated significantly from middle tolate 2-cell stages, as compared with thecontrol group (Supp. Fig. 1, which isavailable online).
ZAR1L Cter-Flag-EGFP
Reduced H3K4me2 and
H3K4me3 Methylation Levels
and Active RNA Polymerase
II (phosphor-Rpb1) Level
Histones H3K4 and H3K9 methylationplay important roles in regulating ge-nome structure and gene transcription.
In order to confirm and explain howthe BrUTP incorporation was affectedby the ZAR1L Cter-Flag-EGFP mu-tant, the RNA transcription-relatedhistone H3K4 methyl-modificationswere determined by immunostaining.The results showed that when ZAR1LCter-Flag-EGFP mutant was injectedinto zygotes, H3K4me2 and H3K4me3were down-regulated significantly(Fig. 5C to F), while H3K4me1 was up-regulated moderately (Fig. 5A and B).It is interesting that in about half ofthe late 2-cell embryos, the H3K4me2and H3K4me3 level in one blastomereusually down-regulated earlier/fasterthan the other one (Fig. 5G and H).Quantification and statistics analysisshowed that H3K4me1 levels were lowin 81.8% of the EGFP-injected late 2-cell embryos. And H3K4me1 levelswere middle or high in 89.9% of theZar1l Cter-EGFP-injected late 2-cellembryos (Fig. 5I). H3K4me2 andH3K4me3 levels were high in 70.7 and72.3% of the EGFP injected late 2-cellembryos (Fig. 5J and K). However,H3K4me2 and H3K4me3 levels werehigh in only 18.2 and 14.1% of theZar1l Cter-EGFP injected late 2-cellembryos (Fig. 5J and K).
Activated RNA polymerase II (phos-phorylated Rpb1 at C-terminal Ser2/5repeats, Phospho-Rpb1) was one ofthe key factors that represented themRNA transcription/synthesis level.In order to confirm the down-regula-tion of RNA synthesis (as indicated byBrUTP incorporation assay), we fur-ther performed Phospho-Rpb1 immu-nostaining. The results showed thatthe Phospho-Rpb1 signal was alsodown-regulated significantly (Fig. 5Gand H). Quantification and statisticsanalysis showed that Phospho-Rpb1levels were high in 82.3% of theEGFP injected late 2-cell embryos.However, Phospho-Rpb1 levels werelow in 67.3% of the Zar1l Cter-EGFP-injected late 2-cell embryos (Fig. 5L).
ZAR1L Cter-Flag-EGFP
Significantly Down-Regulated
H3K9me2 But Up-Regulated
H3K9me3 in Late Two-Cell-
Stage Embryos
To investigate whether Histone H3K9methylation levels changed according
to the phenotypes observed above,we performed immunostaining ofH3K9me2 and H3K9me3 at late 2-cell-stage embryos. The results dem-onstrated that the H3K9me2 signalwas dramatically down-regulated (Fig.6A–C) in most of the late 2-cell embryosthat were injected with ZAR1L Cter-Flag-EGFP. The H3K9me3 signal,however, was dramatically up-regu-lated and formed a perinuclear ringin the ZAR1L Cter-Flag-EGFP group(Fig. 6D–F). The H3K9me3 signalwas dramatically down-regulated fora short period of time in normal latetwo-cell-stage embryos (Fig. 6D). Sta-tistics analysis demonstrated thatabout 77% of the EGFP controlgroup embryos showed a high levelof H3K9me2. In contrast, 74% ofthe ZAR1L Cter-Flag-EGFP groupembryos showed very weak levels ofH3K9me2. Our results showed thatabout 5% (3/57) of the late two-cell-stage embryos in the control groupsexhibited strong staining of anti-H3K9me3. However, more than 80%(83.9 6 6.0%, n ¼ 4; 49/58) of the latetwo-cell-stage embryos in the ZAR1LCter-Flag-EGFP group had stronganti-H3K9me3 staining (Fig. 6E andF). DAPI staining showed thatcrescent-like nuclei were observedin about a half of the ZAR1L Cter-Flag-EGFP-injected two-cell-stageembryos (Figs. 5B, D, 6D). The cres-cent-like nucleus and H3K9me3perinuclear ring had not beenobserved in the control groups (Figs.5A,C, 6A,D).
ZAR1L Colocalized With
P-Body Components and
Germline-Specific
Chromatoid Body
Components in Somatic Cells
In order to explore the functionalproperties and molecular mechanismsof ZAR1L protein, we over-expressedZAR1L-Flag-EGFP in 293T cells andthen performed co-immunoprecipita-tion with the anti-Flag monoclonalantibody. A set of widely expressed P-body components was found in theimmunoprecipitates with ZAR1L-Flag-EGFP, including EIF2C1, EIF2C2,DDX6, and LSM14A. To test colocali-zation between ZAR1L and the P-bodycomponents, we co-expressed ZAR1L
412 HU ET AL.
Dev
elop
men
tal D
ynam
ics
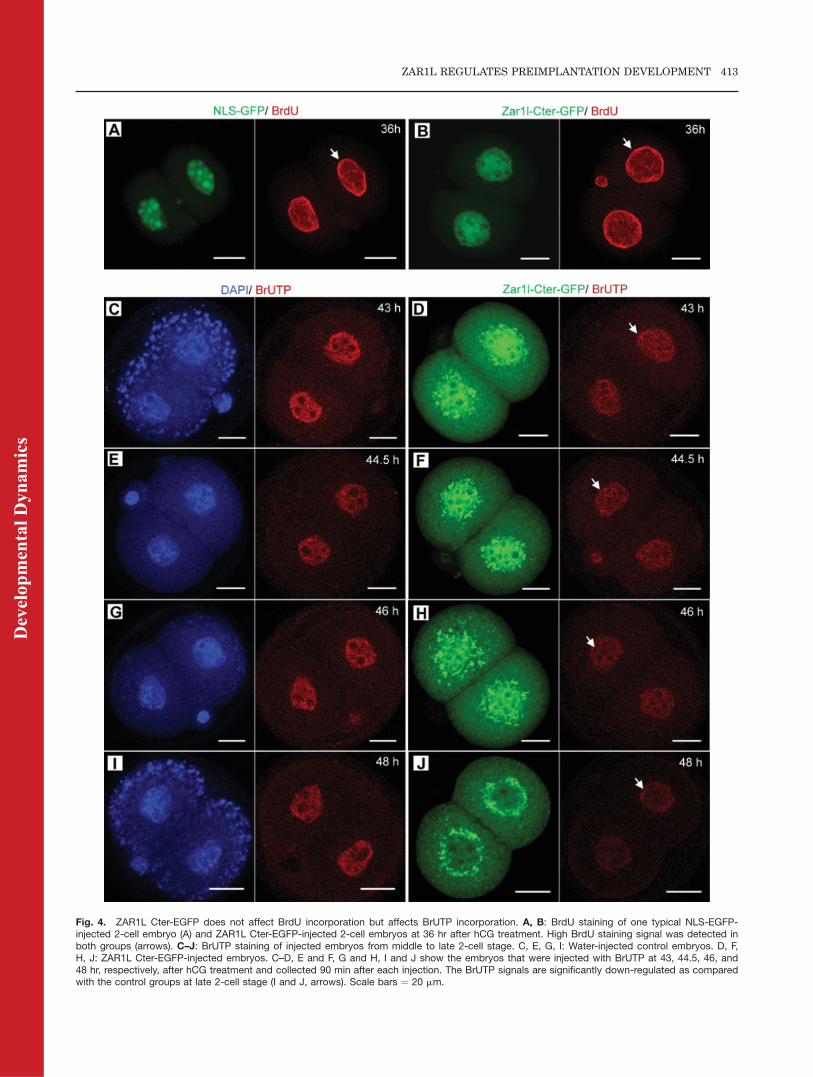
Fig. 4. ZAR1L Cter-EGFP does not affect BrdU incorporation but affects BrUTP incorporation. A, B: BrdU staining of one typical NLS-EGFP-injected 2-cell embryo (A) and ZAR1L Cter-EGFP-injected 2-cell embryos at 36 hr after hCG treatment. High BrdU staining signal was detected inboth groups (arrows). C–J: BrUTP staining of injected embryos from middle to late 2-cell stage. C, E, G, I: Water-injected control embryos. D, F,H, J: ZAR1L Cter-EGFP-injected embryos. C–D, E and F, G and H, I and J show the embryos that were injected with BrUTP at 43, 44.5, 46, and48 hr, respectively, after hCG treatment and collected 90 min after each injection. The BrUTP signals are significantly down-regulated as comparedwith the control groups at late 2-cell stage (I and J, arrows). Scale bars ¼ 20 mm.
ZAR1L REGULATES PREIMPLANTATION DEVELOPMENT 413
Dev
elop
men
tal D
ynam
ics
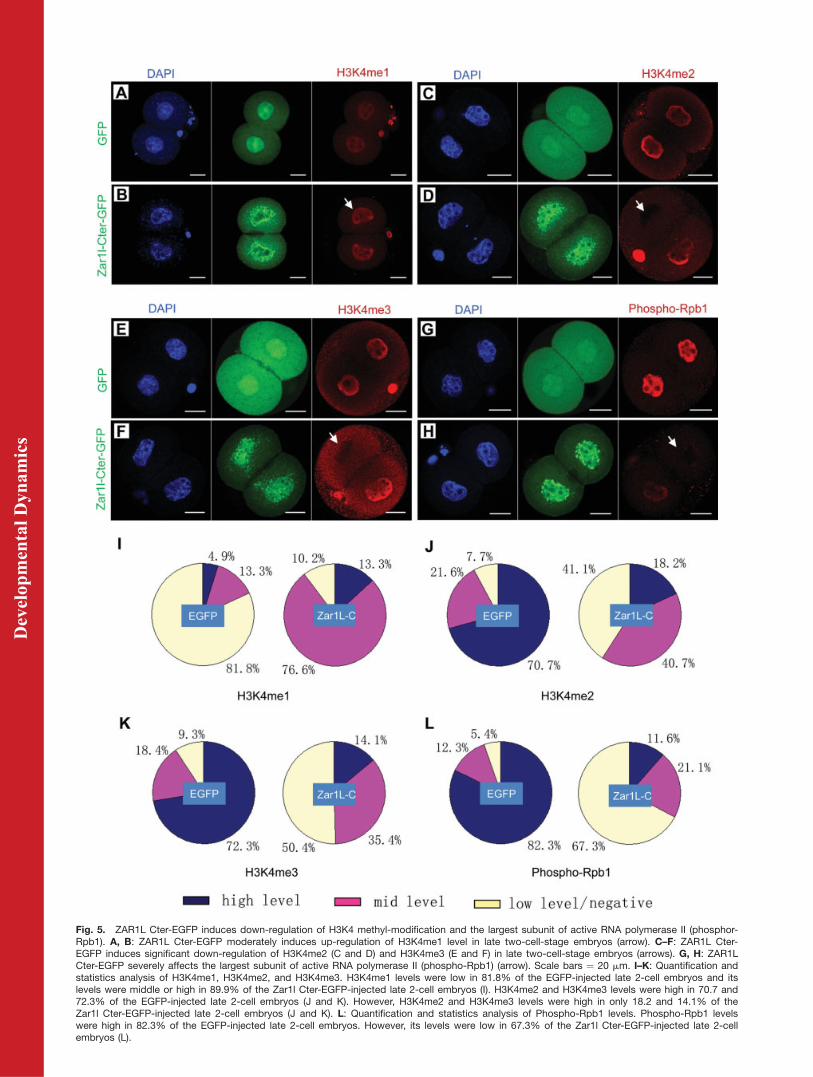
Fig. 5. ZAR1L Cter-EGFP induces down-regulation of H3K4 methyl-modification and the largest subunit of active RNA polymerase II (phosphor-Rpb1). A, B: ZAR1L Cter-EGFP moderately induces up-regulation of H3K4me1 level in late two-cell-stage embryos (arrow). C–F: ZAR1L Cter-EGFP induces significant down-regulation of H3K4me2 (C and D) and H3K4me3 (E and F) in late two-cell-stage embryos (arrows). G, H: ZAR1LCter-EGFP severely affects the largest subunit of active RNA polymerase II (phospho-Rpb1) (arrow). Scale bars ¼ 20 mm. I–K: Quantification andstatistics analysis of H3K4me1, H3K4me2, and H3K4me3. H3K4me1 levels were low in 81.8% of the EGFP-injected late 2-cell embryos and itslevels were middle or high in 89.9% of the Zar1l Cter-EGFP-injected late 2-cell embryos (I). H3K4me2 and H3K4me3 levels were high in 70.7 and72.3% of the EGFP-injected late 2-cell embryos (J and K). However, H3K4me2 and H3K4me3 levels were high in only 18.2 and 14.1% of theZar1l Cter-EGFP-injected late 2-cell embryos (J and K). L: Quantification and statistics analysis of Phospho-Rpb1 levels. Phospho-Rpb1 levelswere high in 82.3% of the EGFP-injected late 2-cell embryos. However, its levels were low in 67.3% of the Zar1l Cter-EGFP-injected late 2-cellembryos (L).
Dev
elop
men
tal D
ynam
ics
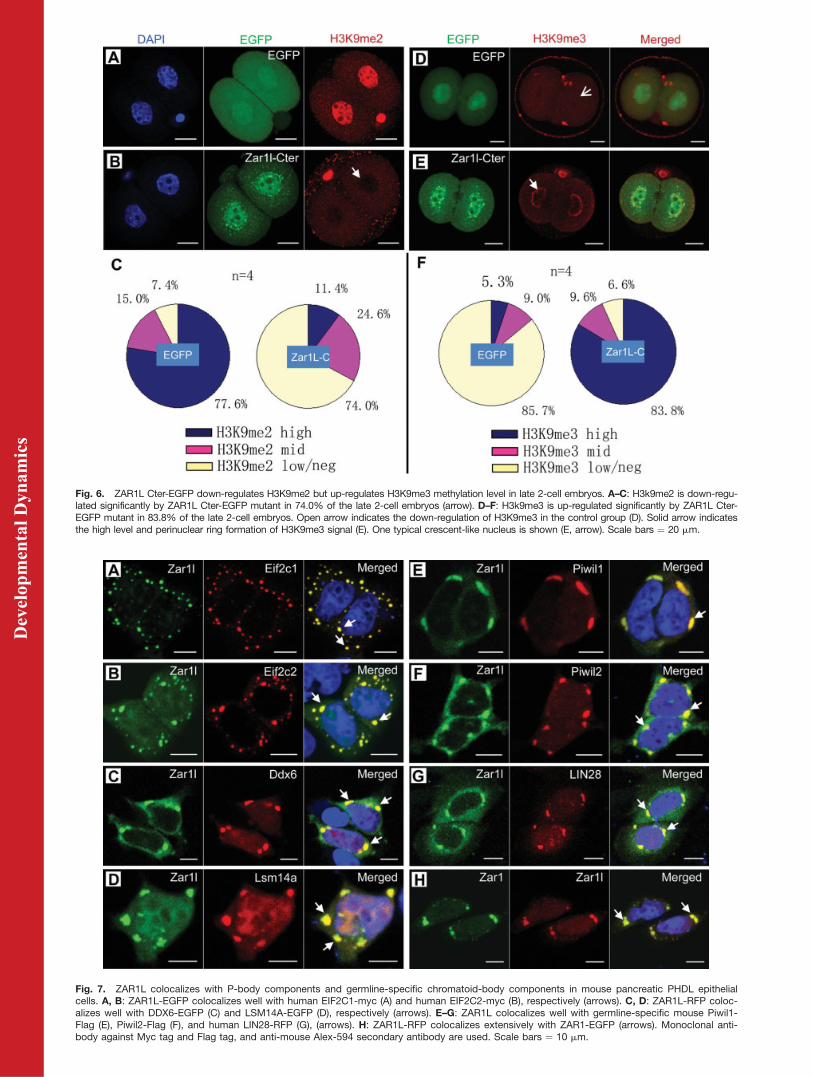
Fig. 6. ZAR1L Cter-EGFP down-regulates H3K9me2 but up-regulates H3K9me3 methylation level in late 2-cell embryos. A–C: H3k9me2 is down-regu-lated significantly by ZAR1L Cter-EGFP mutant in 74.0% of the late 2-cell embryos (arrow). D–F: H3k9me3 is up-regulated significantly by ZAR1L Cter-EGFP mutant in 83.8% of the late 2-cell embryos. Open arrow indicates the down-regulation of H3K9me3 in the control group (D). Solid arrow indicatesthe high level and perinuclear ring formation of H3K9me3 signal (E). One typical crescent-like nucleus is shown (E, arrow). Scale bars ¼ 20 mm.
Fig. 7. ZAR1L colocalizes with P-body components and germline-specific chromatoid-body components in mouse pancreatic PHDL epithelialcells. A, B: ZAR1L-EGFP colocalizes well with human EIF2C1-myc (A) and human EIF2C2-myc (B), respectively (arrows). C, D: ZAR1L-RFP coloc-alizes well with DDX6-EGFP (C) and LSM14A-EGFP (D), respectively (arrows). E–G: ZAR1L colocalizes well with germline-specific mouse Piwil1-Flag (E), Piwil2-Flag (F), and human LIN28-RFP (G), (arrows). H: ZAR1L-RFP colocalizes extensively with ZAR1-EGFP (arrows). Monoclonal anti-body against Myc tag and Flag tag, and anti-mouse Alex-594 secondary antibody are used. Scale bars ¼ 10 mm.
Dev
elop
men
tal D
ynam
ics

and the P-body components transi-ently in mouse pancreatic PNA-HSAdouble-low (PHDL) cells and human293T cells. The results showed thatZAR1L colocalized well with EIF2C1,EIF2C2, DDX6, and LSM14A inmouse PHDL cells (Fig. 7A–D) andhuman 293T cells (data not shown).
The chromatoid body has a structuresimilar to that of the P-body in themale germline. It was observed in cyto-plasm as cytoplasmic foci. BecauseZAR1L was a female germline-specificprotein, we assessed whether ZAR1Lcolocalized with germline-specific chro-matoid body components. The resultsshowed that ZAR1L colocalized exten-sively with the germline-specificPIWIL1, PIWIL2, and LIN28 whenthey were ectopically expressed inPHDL cells (Fig. 7E–G) and 293T cells(data not shown). In addition, ZAR1Lalso colocalized extensively with thegermline-specific ZAR1.
Colocalization of ZAR1L,
ZAR1L C-ter-Flag-EGFP,
LIN28, LSM14A, and ZAR1 in
2-Cell Embryos
The protein sequence alignment analy-sis suggested that mouse ZAR1L pro-tein might have three functionaldomains. In order to characterizethe function properties of differentdomains, series mutants have beendesigned to analyze their sub-cellularlocalization and colocalization withgermline P-body/C-body components.Our results showed that both ZAR1L-EGFP and ZAR1L Cter-Flag-EGFPcolocalized extensively with LIN28-RFP (Fig. 8A and B) and LSM14A-RFP(Fig. 8C and D) in late 2-cell-stageembryos. In order to characterize thecolocalization between ZAR1L C-termi-nus and ZAR1L or ZAR1, we co-expressed them in zygotes and ana-lyzed them in late 2-cell embryos. Wefound that both ZAR1L-RFP andZAR1-EGFP colocalized extensivelywith the C-terminus of ZAR1L innearly all the 2-cell embryos in thecytoplasmic foci region (Fig. 8E–H).However, we also found that ZAR1LCter-Flag-EGFP and ZAR1L Cter-RFPlocalized to the nucleus in about onefourth of the late 2-cell embryos(23.5%, 12/51 and 27.5%, 19/69, respec-tively, Fig. 8F and H). Further experi-
ment showed that ZAR1L-RFP colocal-ized extensively with ZAR1-EGFP (Fig.8I) and ZAR1L Cter-RFP colocalizedwell with ZAR1 Cter-EGFP (Fig. 8J).Serial observation showed that theZAR1L C-terminus moved from the nu-cleus to the cytoplasmic foci in middleto late 2-cell-stage embryos (Fig. 4D–J,left panels). The colocalization study ofthe Zar1l, Zar1, and Zar1l C-terminusdemonstrated that the full-length Zar1and Zar1l, as well as Zar1l, C-termi-nus, could shuttle from the nucleus tothe cytoplasm. Simultaneously, wefound that co-expression of ZAR1L andC-terminus of ZAR1L could partially(48.2%, 41/85) rescue the 2-cell-blockphenotype that was induced by the C-terminus of ZAR1L at 72 hr after hCGinjection (Fig. 8K and L). However,these embryos still failed to develop tothe blastocyst stage at 120 hr afterhCG injection (Fig. 8M and N).
Mouse ZAR1L Protein
Interacts With Human LIN28
In order to identify some molecularmechanisms of ZAR1L function, weperformed immunoprecipitation andsilver staining experiments to try tofind out some proteins that mightinteract with mouse ZAR1L. Massspectrometry analysis revealed thathuman LIN28 protein was one of theZAR1L immunoprecipitated compo-nents when mouse ZAR1L andhuman LIN28 were co-expressed in293T cells. Moreover, ZAR1L proteinwas rich in the 60-kD band that wasin the lysates generated from LIN28-Flag-EGFP immunoprecipitation (withanti-Flag monoclonal antibody) (Fig.9A). Western blotting analysis furthersupported the interaction betweenmouse ZAR1L and human LIN28 (Fig.9B). Further analysis showed thatboth ZAR1L-RFP and ZAR1L Cter-RFP colocalized extensively withLIN28-EGFP in late two-cell-stageembryos (Fig. 8A and B).
ZAR1L Cter-Flag-EGFP
Down-Regulated a Set of
Chromatin Modification-
Associated Genes in mRNA
Level
It was well known that P-body compo-nents played important roles in regu-lating post-transcriptional mRNA sta-
bility and translation. In order tocharacterize whether the ZAR1L Cter-Flag-EGFP mutant resulted in down-regulation of some zygote and 2-cellembryo highly expressed genes, we per-formed RT-PCR analysis of a group ofselected genes. Our data showed that aset of chromatin-modification genes,including Dppa2, Dppa4, and Piwil2(Mili), were dramatically down-regu-lated by ZAR1L Cter-Flag-EGFP at anmRNA level in late two-cell-stageembryos (Fig. 10A). However, Cbx1,Dppa3, Oct4, Gata6, and Tbpl1 geneswere not affected significantly (Fig.10A). In order to confirm the effect ofZAR1L Cter-Flag-EGFP expression onDPPA2 in the protein level, we per-formed immunostaining with DPPA2-specific antibody. Our results showedthat DPPA2 was dramatically down-regulated at protein levels in about90% (90.8 6 2.2%, n ¼ 4; total 42/54) ofthe late two-cell-stage embryos thatwere injected with ZAR1L Cter-Flag-EGFP (Fig. 10B–D). Only about 4% (2/55) of the late two-cell-stage embryoshad a high staining of DPPA2 (Fig.10D). In the EGFP control group, about95% (94.7 6 3.8%, n ¼ 4; 50/53) of theinjected embryos had strong staining ofDPPA2 (Fig. 10D). To preliminarily testwhether DPPA2 had important rolesin preimplantation development, themRNA corresponding to the N-termi-nus deleted DPPA2-DN-EGFP mutant(Fig. 10E) was injected into zygotes.The DPPA2-DN-EGFPmutant predom-inantly localized in the nucleus andinduced two-cell-stage arrest (Fig. 10Fand G). To further investigate whetherDPPA2 was mainly responsible for theZAR1L Cter-Flag-EGFP-induced 2-cellarrest phenotype, we performed aDPPA2 rescue experiment. We foundthat over-expression of DPPA2 couldnot rescue the 2-cell arrest phenotypethat was induced by ZAR1L Cter-Flag-EGFP (Fig. 10H and I). Simultane-ously, we found that, if co-injected withZAR1L Cter-Flag-EGFP, ectopicallyexpressed DPPA2-RFP protein wasalso down-regulated significantly atlate 2-cell stage (Fig. 10J and K).
DISCUSSION
Wu et al. (2003a) have reported thatZar1 (�/�) mice are viable and grosslynormal, but the Zar1(�/�) females are
416 HU ET AL.
Dev
elop
men
tal D
ynam
ics
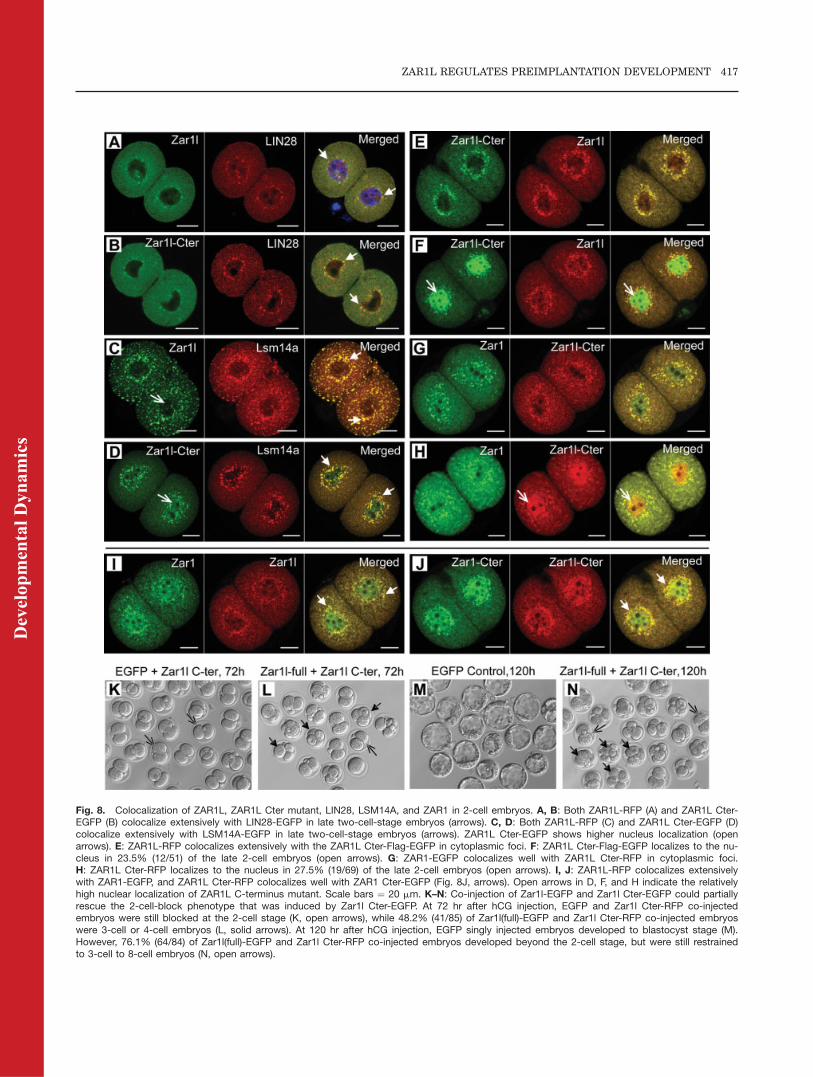
Fig. 8. Colocalization of ZAR1L, ZAR1L Cter mutant, LIN28, LSM14A, and ZAR1 in 2-cell embryos. A, B: Both ZAR1L-RFP (A) and ZAR1L Cter-EGFP (B) colocalize extensively with LIN28-EGFP in late two-cell-stage embryos (arrows). C, D: Both ZAR1L-RFP (C) and ZAR1L Cter-EGFP (D)colocalize extensively with LSM14A-EGFP in late two-cell-stage embryos (arrows). ZAR1L Cter-EGFP shows higher nucleus localization (openarrows). E: ZAR1L-RFP colocalizes extensively with the ZAR1L Cter-Flag-EGFP in cytoplasmic foci. F: ZAR1L Cter-Flag-EGFP localizes to the nu-cleus in 23.5% (12/51) of the late 2-cell embryos (open arrows). G: ZAR1-EGFP colocalizes well with ZAR1L Cter-RFP in cytoplasmic foci.H: ZAR1L Cter-RFP localizes to the nucleus in 27.5% (19/69) of the late 2-cell embryos (open arrows). I, J: ZAR1L-RFP colocalizes extensivelywith ZAR1-EGFP, and ZAR1L Cter-RFP colocalizes well with ZAR1 Cter-EGFP (Fig. 8J, arrows). Open arrows in D, F, and H indicate the relativelyhigh nuclear localization of ZAR1L C-terminus mutant. Scale bars ¼ 20 mm. K–N: Co-injection of Zar1l-EGFP and Zar1l Cter-EGFP could partiallyrescue the 2-cell-block phenotype that was induced by Zar1l Cter-EGFP. At 72 hr after hCG injection, EGFP and Zar1l Cter-RFP co-injectedembryos were still blocked at the 2-cell stage (K, open arrows), while 48.2% (41/85) of Zar1l(full)-EGFP and Zar1l Cter-RFP co-injected embryoswere 3-cell or 4-cell embryos (L, solid arrows). At 120 hr after hCG injection, EGFP singly injected embryos developed to blastocyst stage (M).However, 76.1% (64/84) of Zar1l(full)-EGFP and Zar1l Cter-RFP co-injected embryos developed beyond the 2-cell stage, but were still restrainedto 3-cell to 8-cell embryos (N, open arrows).
ZAR1L REGULATES PREIMPLANTATION DEVELOPMENT 417
Dev
elop
men
tal D
ynam
ics

infertile. They found that ovarian de-velopment, oogenesis, and fertiliza-tion are unimpaired in Zar1 (�/�)mice. Most of the embryos generatedfrom Zar1(�/�) females, however,arrest at the one-cell and two-cellstages (Wu et al., 2003a). Theyobserved that pronuclei formationand DNA replication occur, but thematernal and paternal genomesremain separate entities in arrestedzygotes. The mechanism by whichnull Zar1 expression caused zygoteand two-cell-stage arrestis unknown. In this study, we charac-terized a Zar1-like gene,XM_359149, which encoded a ZAR1-like protein and we have designatedit as Zar1l. Zar1l ORF sequenceshave a poor similarity to the Zar1ORF. However, they exhibit a highsimilarity in protein sequence intheir C-terminus. When our Zar1lproject was ongoing, Sangiorgioet al. (2008) reported the expressionand preliminary sequence analysis ofBovine ZAR1-like gene and its ortho-logs in vertebrates. Unlike bovineZAR1-like gene, we found thatmouse Zar1-like gene was predomi-nantly expressed in ovaries, oocytes,and early preimplantation embryos.The differences in expression pat-terns suggest that ZAR1L may havespecies-variant functions. The ZAR1-like orthologous proteins are con-
served in their predicted N-termi-nus, middle-region, and C-terminusdomains. The C-terminus zinc-fin-ger-containing domains are well con-served from zebrafish to human(data not shown). These results sug-gested that the mouse Zar1l genemay play some important roles infemale germline and/or embryonicdevelopment.
Mutant ZAR1L Induced
Cell Cycle Arrest in
Preimplantation Embryos
Protein sequence analysis suggestedthat mouse ZAR1L might containthree functional domains: an N-termi-nus unknown domain, a CSE2-likemiddle domain, and a C-terminusatypical PHD zinc finger domain (Wuet al., 2003b). Series mutations weredesigned for functional analysis ofZAR1L protein in preimplantationembryos. Our data showed that boththe C-terminus ZAR1L (ZAR1L Cter-Flag-EGFP) and the N-terminusdeleted ZAR1L (ZAR1L DN-EGFP)caused two-cell-stage arrest. TheZAR1L Cter-Flag-EGFP induced amore severe phenotype. When ZAR1LCter-Flag-EGFP was injected into oneblastomere of the two-cell embryos, itresulted in arrest of the injected blas-tomere after one time cell division,whereas the other blastomere grew to
form a small blastocyst. These datasuggested that mouse Zar1l mightplay important roles in preimplanta-tion development in mice.
Mutant ZAR1L Did Not Affect
DNA Replication But Affected
RNA Synthesis
To find out whether DNA replicationand/or RNA synthesis was affected bythe ZAR1L Cter-Flag-EGFP mutant,we performed BrdU incorporationassay (Vitale et al., 1989) and BrUTPincorporation assay (Aoki et al.,1997). Our results demonstrated thatBrdU incorporation was not affected,but the BrUTP incorporation was sig-nificantly down-regulated in late 2-cell embryos. These data indicatedthat RNA synthesis during late 2-cellembryos was affected by ZAR1L Cter-Flag-EGFP mutant. RNA synthesis isone of the key events during major zy-gotic gene activation. To further con-firm the influence of ZAR1L Cter-Flag-EGFP mutant on zygotic geneactivation, we detected the largestactive RNA polymerase II subunit,Phospho-Rpb1(Ser2/5) (Svejstrup,2002; Shilatifard et al., 2003). Dra-matic down-regulation of phosphor-Rpb1 indicated the RNA synthesis ac-tivity was affected by ZAR1L Cter-Flag-EGFP.
ZAR1L Cter-Flag-EGFP
Induced Abnormalities in
H3K4me1/2/3 and H3K9me2/3
Modifications in Two-Cell-
Stage Embryos
It was reported that Histone H3K4methylation levels correlated withthe genome activation (Lepikhovand Walter, 2004; Lepikhov et al.,2008; VerMilyea et al., 2009), whileH3K9me3 methylation (Lachner andJenuwein, 2002; Horn and Peterson,2006; Grewal and Jia, 2007) corre-lated with genome inactivation. So wefurther determined the H3K4me1/2/3levels and H3K9me2/3 levels. Ourresults showed that H3K4me2/3 wassignificantly down-regulated, whileH3K4me1 was moderately up-regu-lated. These data consisted with thetranscription activity as indicatedby a BrUTP incorporation test andphospho-Rpb1 staining. H3K9me2/3
Fig. 9. Biochemical analysis of interaction between mouse ZAR1L and human LIN28 in 293Tcells. A: Silver staining of samples obtained by immunoprecipitation. Left: ZAR1L Cter-EGFPimmunoprecipitation and silver staining. Middle: ZAR1L-Flag-EGFP was co-transfected withhLIN28-RFP into 293T cells and immunoprecipitation was performed with anti-Flag monoclonalantibody. Right: ZAR1L-RFP was co-transfected with hLIN28-Flag-EGFP into 293T cells andimmunoprecipitation was performed with anti-Flag monoclonal antibody. Arrows indicate thebands of ZAR1L (top) and human LIN28 (bottom). B: Western blotting analysis of samplesobtained by immunoprecipitation. ZAR1L-RFP and LIN28-Flag-EGFP are co-transfected into293T cells. ZAR1L-RFP and LIN28-Flag-EGFP are used as bait proteins. Before elution, samplesare treated with or without RNaseA (20 mg/ml) for 30 min, respectively. Anti-ZAR1L polyclonalantibody and anti-Flag monoclonal antibody have been used.
418 HU ET AL.
Dev
elop
men
tal D
ynam
ics
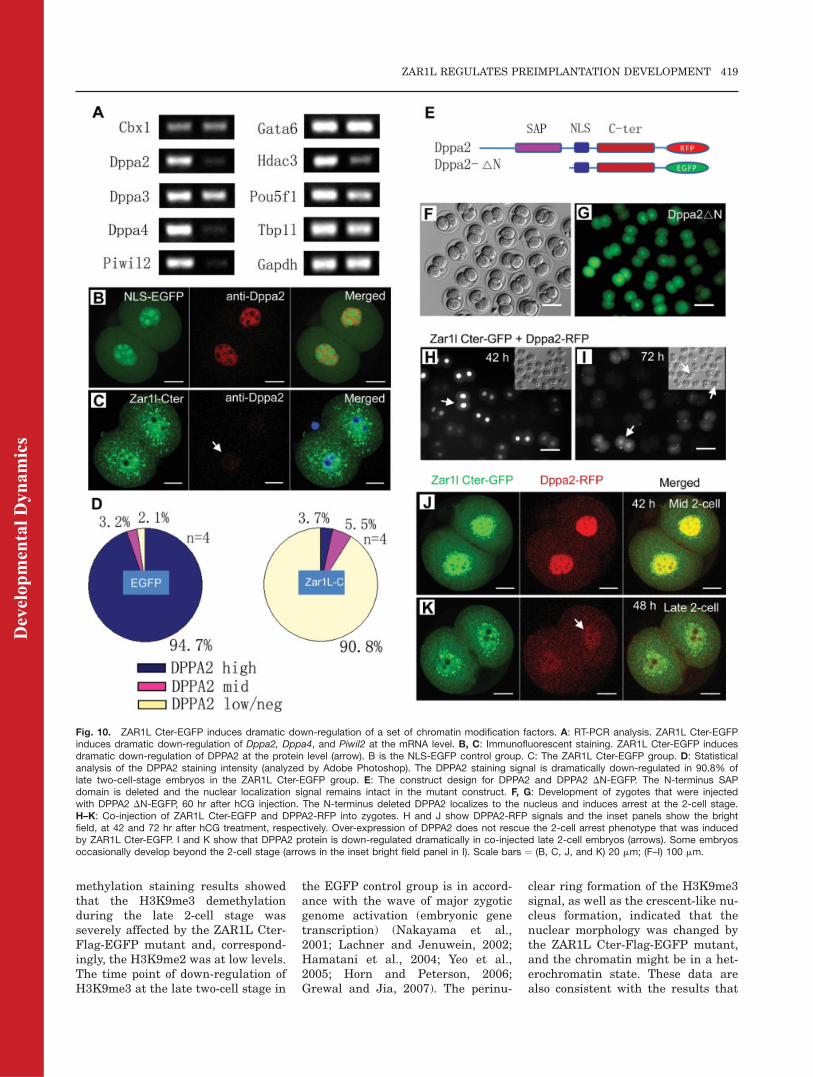
methylation staining results showedthat the H3K9me3 demethylationduring the late 2-cell stage wasseverely affected by the ZAR1L Cter-Flag-EGFP mutant and, correspond-ingly, the H3K9me2 was at low levels.The time point of down-regulation ofH3K9me3 at the late two-cell stage in
the EGFP control group is in accord-ance with the wave of major zygoticgenome activation (embryonic genetranscription) (Nakayama et al.,2001; Lachner and Jenuwein, 2002;Hamatani et al., 2004; Yeo et al.,2005; Horn and Peterson, 2006;Grewal and Jia, 2007). The perinu-
clear ring formation of the H3K9me3signal, as well as the crescent-like nu-cleus formation, indicated that thenuclear morphology was changed bythe ZAR1L Cter-Flag-EGFP mutant,and the chromatin might be in a het-erochromatin state. These data arealso consistent with the results that
Fig. 10. ZAR1L Cter-EGFP induces dramatic down-regulation of a set of chromatin modification factors. A: RT-PCR analysis. ZAR1L Cter-EGFPinduces dramatic down-regulation of Dppa2, Dppa4, and Piwil2 at the mRNA level. B, C: Immunofluorescent staining. ZAR1L Cter-EGFP inducesdramatic down-regulation of DPPA2 at the protein level (arrow). B is the NLS-EGFP control group. C: The ZAR1L Cter-EGFP group. D: Statisticalanalysis of the DPPA2 staining intensity (analyzed by Adobe Photoshop). The DPPA2 staining signal is dramatically down-regulated in 90.8% oflate two-cell-stage embryos in the ZAR1L Cter-EGFP group. E: The construct design for DPPA2 and DPPA2 DN-EGFP. The N-terminus SAPdomain is deleted and the nuclear localization signal remains intact in the mutant construct. F, G: Development of zygotes that were injectedwith DPPA2 DN-EGFP, 60 hr after hCG injection. The N-terminus deleted DPPA2 localizes to the nucleus and induces arrest at the 2-cell stage.H–K: Co-injection of ZAR1L Cter-EGFP and DPPA2-RFP into zygotes. H and J show DPPA2-RFP signals and the inset panels show the brightfield, at 42 and 72 hr after hCG treatment, respectively. Over-expression of DPPA2 does not rescue the 2-cell arrest phenotype that was inducedby ZAR1L Cter-EGFP. I and K show that DPPA2 protein is down-regulated dramatically in co-injected late 2-cell embryos (arrows). Some embryosoccasionally develop beyond the 2-cell stage (arrows in the inset bright field panel in I). Scale bars ¼ (B, C, J, and K) 20 mm; (F–I) 100 mm.
ZAR1L REGULATES PREIMPLANTATION DEVELOPMENT 419
Dev
elop
men
tal D
ynam
ics

significant down-regulation of BrUTPincorporation and phosphor-Rpb1 lev-els in the nucleus have been inducedby Zar1l Cter-EGFP.
ZAR1L Colocalized With the
P-body and C-body
Components in Both Somatic
Cells and Late 2-Cell-Stage
Embryos
Many maternal mRNAs accumulatein growing oocytes and are stored inMII oocytes. They are translationallyrepressed until fertilization. Most ofthe maternal mRNAs have beendegraded by the end of the two-cellstage. Correspondingly, major zygoticgenome activation occurs at the latetwo-cell stage. How the maternalmRNAs are tightly controlled interms of their stability, translationalrepression, and/or initiation, and deg-radation is largely unknown. Recentstudies have revealed the existence ofspecific mRNA processing bodies (P-bodies) as multiple cytoplasmic foci insomatic cells (Hannon, 2002; van Dijket al., 2002; Sheth and Parker, 2003;Cougot et al., 2004; Brengues et al.,2005; Liu et al., 2005). P-bodies con-tain untranslated mRNAs and canserve as sites of mRNA translationalrepression and degradation. P-bodiesare highly dynamic structures, andthe components are altered dependingon the cell state. Many proteins havebeen reported to be localized to P-body structures (Hannon, 2002; vanDijk et al., 2002; Sheth and Parker,2003; Cougot et al., 2004; Brengueset al., 2005; Fillman and Lykke-Andersen, 2005; Liu et al., 2005; Yanget al., 2006; Eulalio et al., 2007;Parker and Sheth, 2007; Pressmanet al., 2007). In the male germline,chromatoid-body (C-body) structureshave been found to be similar to theP-body structures (Matsumoto et al.,2005; Kotaja et al., 2006; Kotaja andSassone-Corsi, 2007).
The male germline-specific cytoplas-mic foci (chromatoid body) share com-ponents found in somatic cell P-bodies,such as the Agonaute proteins, some ofthe RNA enzymes, and ribosomal pro-teins. We found that the maternaleffect gene Zar1l encoded a femalegermline-specific protein ZAR1L,which localized to the cytoplasmic
foci structures in late two-cell-stageembryos. Based on the knowledge thatmRNA processing mechanisms areevolutionarily conserved from germcells to somatic cells, we speculatedthat ZAR1L might be involved inP-body- or C-body-like structures.Indeed, we found that mouse ZAR1Lwas colocalized extensively with widelyexpressed P-body components, includ-ing EIF2C1, EIF2C2, DDX6, andLSM14A and with germline-specificchromatoid-body components includingPIWIL1, PIWIL2, and LIN28 in so-matic cells. We also confirmed thatmouse ZAR1L colocalized extensivelywith LIN28 and LSM14A in late 2-cellembryos. Our data further indicatedco-expression of ZAR1L could partiallyrescue the 2-cell-block phenotype thatwas caused by ZAR1L C-terminus. Thecolocalization study of the Zar1l, Zar1,and Zar1l C-terminus demonstratedthat the full-length Zar1 and Zar1l, aswell as Zar1l C-terminus, could shuttlefrom the nucleus to the cytoplasm.These results indicate that the C-ter-minus of ZAR1L may induce 2-cellblocks through a dominant-negativeeffect. Because of the lack of commer-cialized ZAR1L antibody, we custom-ized peptide antibody against mouseZAR1L. It works well for Western blot-ting but not for immunostaining. Weconfirmed the interaction betweenmouse ZAR1L and human LIN28 in so-matic cells using immunoprecipitationand Western blotting. Our results alsoshowed that ZAR1L extensively colo-calized with ZAR1. These data demon-strated that ZAR1L, as well as ZAR1,may play some roles in P-body and/orC-body structures and might havefunctions in regulating oocyte-to-embryo transition.
The ZAR1L Cter-EGFP Down-
Regulated a Set of Chromatin
Modification Factors,
Including Dppa2, Dppa4, and
Piwil2 at the mRNA Level
Nuclear reprogramming is a criticalevent that occurs during zygotic ge-nome activation (Schultz, 1993; Aokiet al., 1997; Latham, 1999; Lathamand Schultz, 2001; Ma et al., 2001;Hamatani et al., 2004; Minami et al.,2007; Stitzel and Seydoux, 2007),through which the transcriptionally
inactive genome changes into an activegenome. Several maternal factors areassociated with nuclear reprogram-ming. Depletion of Smarca4 (Brg1), achromatin-remodeling factor, causedarrest at the two-cell stage, which wasaccompanied by down-regulation of amultitude of mRNAs (Bultman et al.,2006). Dppa3(Stella) is required forprotection of the maternal genomefrom DNA-demethylation during earlyembryonic development (Nakamuraet al., 2007). Similar to DPPA3, DPPA2and DPPA4 have one DNA-bindingSAP domain and one uncharacterizedC-terminal domain, and are associatedwith chromatin (Aravind and Koonin,2000; Maldonado-Saldivia et al., 2007;Masaki et al., 2007). Recent studieshave indicated that mouse DPPA4 pro-tein associated with transcriptionallyactive chromatin in ES cells (Masakiet al., 2007). Comprehensive ChIP-on-chip analysis demonstrated thatPOU5F1 (OCT-3/4), SOX-2, andNANOG each bind to the Dppa4 pro-moter region in human ES cells (Boyeret al., 2005). However, the roles ofDPPA2 and DPPA4 in early embryonicdevelopment remain largely unknown.Our results showed that Dppa2 andDppa4 were dramatically down-regu-lated by the mutant ZAR1L Cter-EGFP in late two-cell-stage embryos.In order to test whether DPPA2 pro-tein plays important roles in preim-plantation development, we designed adominant-negative mutant of mouseDPPA2 and injected it to the zygotes.Our data showed that deletion of theN-terminal SAP domain of mouseDPPA2 caused arrest at the two-cellstage in vitro. Our data suggested thatDPPA2 may play an important role inembryonic development. We tried torescue the 2-cell arrest phenotypethrough over-expression of DPPA2 butfailed. These data indicated thatDPPA2 is one of the important but notthe dominant factors affected byZAR1L Cter-Flag-EGFP.Piwi family members regulate chro-
matin structure, transposon control,mRNA transcription and translation,and mRNA degradation throughinteractions with piRNAs and associ-ated complexes (Kuramochi-Miya-gawa et al., 2004; Parker et al., 2004;Kavi et al., 2006; Lau et al., 2006;Aravin et al., 2007; Brower-Tolandet al., 2007; Carmell et al., 2007;
420 HU ET AL.
Dev
elop
men
tal D
ynam
ics

Hartig et al., 2007; Houwing et al.,2007; Lin, 2007; Klattenhoff and The-urkauf, 2008). Recently, piRNAs havebeen isolated from murine matureoocytes (Brennecke et al., 2008; Tamet al., 2008; Watanabe et al., 2008).Piwil2 (Mili), but not Piwil1 (Miwi),or Piwil4 (Miwi2) is specificallyexpressed in mature oocytes (Wata-nabe et al., 2008). Piwi family pro-teins and piRNAs play importantroles in chromatin modification andgenome stability (Aravin et al., 2007,2008; O’Donnell and Boeke, 2007).Our data showed that ZAR1L Cter-EGFP dramatically induced down-regulation of Piwil2 mRNAs in two-cell embryos. Down-regulation ofDppa2, Dppa4, and Piwil2 mRNA byZAR1L Cter-Flag-EGFP in vitro inlate two-cell-stage embryos indicatedthat ZAR1L may correlate with a setof mRNAs’ stability or degradation.
In summary, our data demon-strated that ZAR1L plays importantroles in regulating oocyte-to-embryotransition and preimplantation devel-opment. The ZAR1L Cter-Flag-EGFPmutant induced epigenetic abnormal-ities and down-regulation of a groupof chromatin modification factors inlate two-cell-stage embryos andfinally caused arrest at the two-cellstage. ZAR1L colocalized with multi-ple mRNA chromatoid-body/process-ing-body components in somatic cellsand late two-cell-stage embryos, andit interacted with LIN28. ZAR1Lcould be the first tissue- and stage-specific chromatoid-body/processing-body component that has been identi-fied in 2-cell-stage mouse embryos.Zar1l knockout mice will be gener-ated to analyze the functional role ofZAR1L in the female germline andduring embryonic development. Thebiochemical nature of the domains ofZAR1L, as well as ZAR1, remains tobe further characterized.
EXPERIMENTAL
PROCEDURES
Animals, Collection of
Oocytes and Embryos
B6D2F1 (C57BL/6JxDBA2) femalemice (8–10 weeks old) were used forthe collection of fully grown germinalvesicles (GV) and MII oocytes. GVoocytes were collected according to a
previous study (Wang et al., 2008).Zygotes were collected from success-fully mated B6D2F1 females. Allstudies adhered to procedures consist-ent with the National Institute of Bio-logical Sciences Guide for the careand use of laboratory animals.
In Vitro Transcription,
Microinjection, and
Preimplantation Embryo
Incubation
The predicted Zar1l ORF was clonedinto pBS-RN3 (Lemaire et al., 1995),a modified in vitro transcription vec-tor, in which the wild type and mu-tant ZAR1L were expressed with Flagand/or EGFP as fused proteins. Inbrief, zygotes were collected fromadult male B6D2F1 mice at 18 hr af-ter hCG treatment. mRNA injectionwas performed during 20–22 hr afterhCG treatment. The surviving zygoteswere cultured in KSOM medium.Samples were photographed at 48, 72,96, and 120 hr after hCG treatmentfor developmental recording. Late 2-cell embryos were collected during 48–50 hr after hCG treatment. CappedRNAs were transcribed under the con-trol of a T3 promoter with mMessagemMachine (Ambion, Austin, TX), asthe protocol dictated. In vitro tran-scribed mRNAs were injected into GV,MII oocytes, and zygotes using aPIEZO micro-injector.
Cell Culture
293T cells were cultured in DMEM-based medium, which contained 10%FBS (Hyclone, Logan, UT), 2 mM glu-tamine, 1� nucleosides (Gibco, Gai-thersburg, MD), 1� nonessentialamino acids (Gibco), 1� beta-mercap-toethanol (Gibco), 2 mM glutamine(Gibco), 100 IU/ml penicilLin, and 100mg/ml streptomycin (Gibco). Pancre-atic PNA-HSA double-low cells wereincubated under similar conditions asdescribed above, with 3% FBS added.
Construction of Transient
Expression Vectors and
Transfection
The mouse Zar1l ORF was obtainedby performing RT-PCR from adultovarian tissue. The sequences wereconfirmed by sequencing. The DNA
sequences of Zar1l ORF reported inthis study have been deposited in theGenbank database (www.ncbi.nlm.nih.gov/Genbank, accession no. FJ858201).The ORF and mutants were cloned intothe pEGFP-N1 vector for transientexpression. Eukaryotic expression vec-tors inserted with human EIF2C1,human EIF2C2, mouse Ddx6, mouseLsm14a, mouse Piwil1 (Miwi), mousePiwil2 (Mili), and human LIN28 wereco-transfected with Zar1l into PHDLcells or 293T cells to analyze their coloc-alization with mouse ZAR1L. Vigofectreagent was used according to the man-ufacturer’s protocol (Vigorous). Cellswere collected 24 or 36 hr after trans-fection. Please see Supp. Table S1 toview the primer sequences used.
RT-PCR
Total RNA samples were preparedfrom adult ovary, testis, liver,spleen, oocytes, and preimplantationembryos. The RNA was extractedusing conventional methods for adulttissues. The PicoPure RNA isolationkit (Arcturus) was used to extractRNA from collected oocytes andpreimplantation embryos. Reversetranscription and PCR were per-formed by conventional methodsusing MMLV reverse transcriptase(Promega, Madison, WI). GenomicDNA was extracted by conventionalmethods. The RNA was reverse-tran-scribed by MMLV reverse transcrip-tase (Promega) and amplified by PCRfor 25 or 30 cycles. Primers wereselected that encompassed the in-tronic sequences. PCR cycling wasperformed at 98�C for 2 min followedby 98�C for 15 sec, 54–60�C for 15 sec,72�C for 40 sec, and finally 72�C for 8min, using the PrimeSTAR HS DNApolymerase (Takara). Please seeSupp. Table S2 to view the primersequences.
Immunofluorescent Staining
and Confocal Microscopy
Conventional immunostaining wereperformed for H3K4me1, H3K4me2,H3K4me3, H3K9me2, H3K9me3,Phospho-Rpb1, and Dppa2 antibodies.In brief, samples were fixed by 4%paraformeldehyde for 20 min. Then,the samples were permeabilized with0.5% triton X-100 and blocked with
ZAR1L REGULATES PREIMPLANTATION DEVELOPMENT 421
Dev
elop
men
tal D
ynam
ics

5% normal horse serum for 2 hr.The primary antibodies were incu-bated overnight at 4�C. The antibod-ies against H3K4me1, H3K4me2,H3K4me3, H3K9me2, H3K9me3 (allfrom Upstate, Billerica, MA), andDppa2 (gifted by Dr. Western at theARC Centre, Australia) were dilutedat 1:300. Phospho-Rpb1 antibody(Cell Signaling Technology, Danvers,MA) was diluted at 1:100. For BrdUstaining, BrdU was added to the me-dium at 28 hr after hCG treatment.The BrdU-labeled samples were col-lected 36 hr after hCG treatment.Samples were treated with 2 M HClfor 10 min after permeabilization andwashing. Then, samples were washedonce with pH 8.0 Tris-HCl and thenwith pH 7.4 PBS four times after HCltreatment. Conventional methodswere used for the other staining steps.For BrUTP staining, a similaramount of 100 mM BrUTP (Sigma,St. Louis, MO) was injected intoZAR1L Cter-EGFP or EGFP pre-injected (at zygote stage) 2-cellembryos at 43, 44.5, 46, and 48 hr af-ter hCG treatment. Samples were col-lected 90 min after each injection ofBrUTP (Sigma). Both the incorpo-rated BrdU and BrUTP were stainedwith anti-BrdU monoclonal antibody.The Alexa-594-conjugated secondaryantibodies were incubated for 1 hr atroom temperature. Samples were fur-ther counterstained with 100 ng/ml ofDAPI. Images were obtained with anOlympus IX 71 microscope equippedwith a CCD camera (DVC, Austin,TX), or LSM510 Meta confocal micro-scope (Zeiss, Oberkochen, Germany).The staining intensity was analyzedby Adobe Photoshop.
Immunoprecipitation and
Western Blotting
293T cells were transfected to transi-ently express ZAR1L Cter-Flag-EGFP,ZAR1L-Flag-EGFP, ZAR1L-RFP, hu-man LIN28-Flag-EGFP, and LIN28-RFP, either independently or in combi-nation. Samples were collected 36 hrafter transfection. Immunoprecipita-tions were performed according to themanufacturer’s protocol (FLAGIPT-1;Sigma) or with anti-ZAR1L polyclonalantibody prepared using a specificpolypeptide (134-RRPQDGE DEES-QEE-147). The final concentration of
the ZAR1L antibody was 1 mg/ml. Pro-teins were separated in 10% SDS-poly-acrylamide gels and transferred toPVDF membrane. Immunoblottinganalysis was performed using anti-Flag monoclonal and anti-ZAR1L poly-clonal antibodies. Blots were detectedusing ECL (Amersham Biosciences,Pittsburgh, PA) according to the manu-facturer’s protocol. Blots were detectedusing ECL (GE Healthcare, London,UK).
Silver Staining and Mass
Spectrometry Analysis
Proteins were separated in 10% SDS-polyacrylamide gels. Silver stainingwas performed according to the man-ufacturer’s protocol (PROTSIL2-1KT,Sigma). The bands of interest wereanalyzed by a LTQ linear ion trapmass spectrometer (Thermo Electron,Waltham, MA).
ACKNOWLEDGMENTSWe greatly appreciate Dr. Gurdon atthe Gurdon Institute for providing thepBS-RN3 vector. We thank Dr. West-ern at the ARC Centre (Australia) forproviding the DPPA2 antibody. Wethank Dr. Satomi at Osaka Universityfor providing the Piwil1 and Piwil2cDNA. We thank the lab members andthe two anonymous reviewers for help-ful comments on themanuscript.
REFERENCES
Aoki F, Worrad DM, Schultz RM. 1997.Regulation of transcriptional activityduring the first and second cell cyclesin the preimplantation mouse embryo.Dev Biol 181:296–307.
Aravin AA, Hannon GJ, Brennecke J.2007. The Piwi-piRNA pathway pro-vides an adaptive defense in the trans-poson arms race. Science 318:761–764.
Aravin AA, Sachidanandam R, Bourc’hisD, Schaefer C, Pezic D, Toth KF, BestorT, Hannon GJ. 2008. A piRNA pathwayprimed by individual transposons islinked to de novo DNA methylation inmice. Mol Cell 31:785–799.
Aravind L, Koonin EV. 2000. SAP: a puta-tive DNA-binding motif involved inchromosomal organization. Trends Bio-chem Sci 25:112–114.
Bachvarova R. 1985. Gene expressionduring oogenesis and oocyte develop-ment in mammals. Dev Biol 1:453–524.
Boyer LA, Lee TI, Cole MF, JohnstoneSE, Levine SS, Zucker JP, GuentherMG, Kumar RM, Murray HL, JennerRG, Gifford DK, Melton DA, Jaenisch
R, Young RA. 2005. Core transcriptionalregulatory circuitry in human embry-onic stem cells. Cell 122:947–956.
Brengues M, Teixeira D, Parker R. 2005.Movement of eukaryotic mRNAsbetween polysomes and cytoplasmicprocessing bodies. Science 310:486–489.
Brennecke J, Malone CD, Aravin AA,Sachidanandam R, Stark A, HannonGJ. 2008. An epigenetic role for mater-nally inherited piRNAs in transposonsilencing. Science 322:1387–1392.
Brower-Toland B, Findley SD, Jiang L,Liu L, Yin H, Dus M, Zhou P, Elgin SC,Lin H. 2007. Drosophila PIWI associateswith chromatin and interacts directlywith HP1a. Genes Dev 21:2300–2311.
Bultman SJ, Gebuhr TC, Pan H, SvobodaP, Schultz RM, Magnuson T. 2006.Maternal BRG1 regulates zygotic ge-nome activation in the mouse. GenesDev 20:1744–1754.
Burns KH, Viveiros MM, Ren Y, Wang P,DeMayo FJ, Frail DE, Eppig JJ, MatzukMM. 2003. Roles of NPM2 in chromatinand nucleolar organization in oocytesand embryos. Science 300:633–636.
Carmell MA, Girard A, van de Kant HJ,Bourc’his D, Bestor TH, de Rooij DG,Hannon GJ. 2007. MIWI2 is essentialfor spermatogenesis and repression oftransposons in the mouse male germ-line. Dev Cell 12:503–514.
Christians E, Davis AA, Thomas SD, Ben-jamin IJ. 2000. Maternal effect of Hsf1on reproductive success. Nature 407:693–694.
Cougot N, Babajko S, Seraphin B. 2004.Cytoplasmic foci are sites of mRNAdecay in human cells. J Cell Biol 165:31–40.
De La Fuente R, Viveiros MM, Burns KH,Adashi EY, Matzuk MM, Eppig JJ.2004. Major chromatin remodeling inthe germinal vesicle (GV) of mamma-lian oocytes is dispensable for globaltranscriptional silencing but requiredfor centromeric heterochromatin func-tion. Dev Biol 275:447–458.
De Vries WN, Evsikov AV, Haac BE,Fancher KS, Holbrook AE, Kemler R, Sol-ter D, Knowles BB. 2004. Maternal beta-catenin and E-cadherin in mouse develop-ment. Development 131:4435–4445.
Erhardt S, Su IH, Schneider R, Barton S,Bannister AJ, Perez-Burgos L, Jenu-wein T, Kouzarides T, Tarakhovsky A,Surani MA. 2003. Consequences of thedepletion of zygotic and embryonicenhancer of zeste 2 during preimplanta-tion mouse development. Development130:4235–4248.
Eulalio A, Behm-Ansmant I, IzaurraldeE. 2007. P bodies: at the crossroads ofpost-transcriptional pathways. Nat RevMol Cell Biol 8:9–22.
Fillman C, Lykke-Andersen J. 2005. RNAdecapping inside and outside of process-ing bodies. Curr Opin Cell Biol 17:326–331.
Grewal SI, Jia S. 2007. Heterochromatinrevisited. Nat Rev Genet 8:35–46.
Gurtu VE, Verma S, Grossmann AH, Lis-kay RM, Skarnes WC, Baker SM. 2002.
422 HU ET AL.
Dev
elop
men
tal D
ynam
ics

Maternal effect for DNA mismatchrepair in the mouse. Genetics 160:271–277.
Hamatani T, Carter MG, Sharov AA, KoMS. 2004. Dynamics of global geneexpression changes during mouse pre-implantation development. Dev Cell 6:117–131.
Hannon GJ. 2002. RNA interference. Na-ture 418:244–251.
Hartig JV, Tomari Y, Forstemann K.2007. piRNAs--the ancient hunters ofgenome invaders. Genes Dev 21:1707–1713.
Horn PJ, Peterson CL. 2006. Heterochro-matin assembly: a new twist on an oldmodel. Chromosome Res 14:83–94.
Houwing S, Kamminga LM, Berezikov E,Cronembold D, Girard A, van den ElstH, Filippov DV, Blaser H, Raz E, MoensCB, Plasterk RH, Hannon GJ, DraperBW, Ketting RF. 2007. A role for Piwiand piRNAs in germ cell maintenanceand transposon silencing in Zebrafish.Cell 129:69–82.
Howell CY, Bestor TH, Ding F, LathamKE, Mertineit C, Trasler JM, ChailletJR. 2001. Genomic imprinting dis-rupted by a maternal effect mutation inthe Dnmt1 gene. Cell 104:829–838.
Kaneda M, Okano M, Hata K, Sado T,Tsujimoto N, Li E, Sasaki H. 2004.Essential role for de novo DNA methyl-transferase Dnmt3a in paternal andmaternal imprinting. Nature 429:900–903.
Kavi HH, Fernandez HR, Xie W, BirchlerJA. 2006. Polycomb, pairing and PIWI–RNA silencing and nuclear interactions.Trends Biochem Sci 31:485–487.
Klattenhoff C, Theurkauf W. 2008. Bio-genesis and germline functions of piR-NAs. Development 135:3–9.
Kotaja N, Sassone-Corsi P. 2007. Thechromatoid body: a germ-cell-specificRNA-processing centre. Nat Rev MolCell Biol 8:85–90.
Kotaja N, Bhattacharyya SN, JaskiewiczL, Kimmins S, Parvinen M, FilipowiczW, Sassone-Corsi P. 2006. The chroma-toid body of male germ cells: similaritywith processing bodies and presence ofDicer and microRNA pathway compo-nents. Proc Natl Acad Sci USA 103:2647–2652.
Kuramochi-Miyagawa S, Kimura T, IjiriTW, Isobe T, Asada N, Fujita Y, IkawaM, Iwai N, Okabe M, Deng W, Lin H,Matsuda Y, Nakano T. 2004. Mili, amammalian member of piwi familygene, is essential for spermatogenesis.Development 131:839–849.
Lachner M, Jenuwein T. 2002. The manyfaces of histone lysine methylation.Curr Opin Cell Biol 14:286–298.
Latham KE. 1999. Mechanisms and con-trol of embryonic genome activation inmammalian embryos. Int Rev Cytol193:71–124.
Latham KE, Schultz RM. 2001. Embry-onic genome activation. Front Biosci 6:D748–759.
Lau NC, Seto AG, Kim J, Kuramochi-Miyagawa S, Nakano T, Bartel DP,
Kingston RE. 2006. Characterization ofthe piRNA complex from rat testes. Sci-ence 313:363–367.
Lemaire P, Garrett N, Gurdon JB. 1995.Expression cloning of Siamois, a Xeno-pus homeobox gene expressed in dorsal-vegetal cells of blastulae and able toinduce a complete secondary axis. Cell81:85–94.
Lepikhov K, Walter J. 2004. Differentialdynamics of histone H3 methylation atpositions K4 and K9 in the mouse zy-gote. BMC Dev Biol 4:12.
Lepikhov K, Zakhartchenko V, Hao R,Yang F, Wrenzycki C, Niemann H, WolfE, Walter J. 2008. Evidence for con-served DNA and histone H3 methylationreprogramming in mouse, bovine andrabbit zygotes. Epigenet Chromatin 1:8.
Lin H. 2007. piRNAs in the germ line.Science 316:397.
Liu J, Valencia-Sanchez MA, Hannon GJ,Parker R. 2005. MicroRNA-dependentlocalization of targeted mRNAs to mam-malian P-bodies. Nat Cell Biol 7:719–723.
Ma J, Svoboda P, Schultz RM, Stein P.2001. Regulation of zygotic gene act-ivation in the preimplantation mouseembryo: global activation and repres-sion of gene expression. Biol Reprod 64:1713–1721.
Maldonado-Saldivia J, van den Bergen J,Krouskos M, Gilchrist M, Lee C, Li R,Sinclair AH, Surani MA, Western PS.2007. Dppa2 and Dppa4 are closelylinked SAP motif genes restricted to plu-ripotent cells and the germ line. StemCells 25:19–28.
Masaki H, Nishida T, Kitajima S, AsahinaK, Teraoka H. 2007. Developmental plu-ripotency-associated 4 (DPPA4) local-ized in active chromatin inhibits mouseembryonic stem cell differentiation intoa primitive ectoderm lineage. J BiolChem 282:33034–33042.
Matsumoto K, Kwon OY, Kim H, Akao Y.2005. Expression of rck/p54, a DEAD-box RNA helicase, in gametogenesisand early embryogenesis of mice. DevDyn 233:1149–1156.
Minami N, Suzuki T, Tsukamoto S. 2007.Zygotic gene activation and maternalfactors in mammals. J Reprod Dev 53:707–715.
Nakamura T, Arai Y, Umehara H, Masu-hara M, Kimura T, Taniguchi H, Seki-moto T, Ikawa M, Yoneda Y, Okabe M,Tanaka S, Shiota K, Nakano T. 2007.PGC7/Stella protects against DNAdemethylation in early embryogenesis.Nat Cell Biol 9:64–71.
Nakayama J, Rice JC, Strahl BD, AllisCD, Grewal SI. 2001. Role of histoneH3 lysine 9 methylation in epigeneticcontrol of heterochromatin assembly.Science 292:110–113.
O’Donnell KA, Boeke JD. 2007. MightyPiwis defend the germline against ge-nome intruders. Cell 129:37–44.
Parker R, Sheth U. 2007. P bodies andthe control of mRNA translation anddegradation. Mol Cell 25:635–646.
Parker JS, Roe SM, Barford D. 2004.Crystal structure of a PIWI protein sug-
gests mechanisms for siRNA recogni-tion and slicer activity. Embo J 23:4727–4737.
Payer B, Saitou M, Barton SC, ThresherR, Dixon JP, Zahn D, Colledge WH,Carlton MB, Nakano T, Surani MA.2003. Stella is a maternal effect generequired for normal early developmentin mice. Curr Biol 13:2110–2117.
Pressman S, Bei Y, Carthew R. 2007.Posttranscriptional gene silencing. Cell130:570.
Roest HP, Baarends WM, de Wit J, vanKlaveren JW, Wassenaar E, Hooger-brugge JW, van Cappellen WA, Hoeij-makers JH, Grootegoed JA. 2004. Theubiquitin-conjugating DNA repair en-zyme HR6A is a maternal factor essen-tial for early embryonic development inmice. Mol Cell Biol 24:5485–5495.
Sangiorgio L, Strumbo B, Brevini TA,Ronchi S, Simonic T. 2008. A putativeprotein structurally related to zygotearrest 1 (Zar1), Zar1-like, is encoded bya novel gene conserved in the verte-brate lineage. Comp Biochem Physiol BBiochem Mol Biol 150:233–239.
Schultz RM. 1993. Regulation of zygoticgene activation in the mouse. Bioessays15:531–538.
Sheth U, Parker R. 2003. Decapping anddecay of messenger RNA occur in cyto-plasmic processing bodies. Science 300:805–808.
Shilatifard A, Conaway RC, Conaway JW.2003. The RNA polymerase II elonga-tion complex. Annu Rev Biochem 72:693–715.
Stitzel ML, Seydoux G. 2007. Regulationof the oocyte-to-zygote transition. Sci-ence 316:407–408.
Svejstrup JQ. 2002. Mechanisms of tran-scription-coupled DNA repair. Nat RevMol Cell Biol 3:21–29.
Tam OH, Aravin AA, Stein P, Girard A,Murchison EP, Cheloufi S, Hodges E,Anger M, Sachidanandam R, SchultzRM, Hannon GJ. 2008. Pseudogene-derived small interfering RNAs regu-late gene expression in mouse oocytes.Nature 453:534–538.
Tong ZB, Gold L, Pfeifer KE, DorwardH, Lee E, Bondy CA, Dean J, NelsonLM. 2000. Mater, a maternal effectgene required for early embryonic de-velopment in mice. Nat Genet 26:267–268.
van Dijk E, Cougot N, Meyer S, BabajkoS, Wahle E, Seraphin B. 2002. HumanDcp2: a catalytically active mRNAdecapping enzyme located in specificcytoplasmic structures. Embo J 21:6915–6924.
VerMilyea MD, O’Neill LP, Turner BM.2009. Transcription-independent herit-ability of induced histone modificationsin the mouse preimplantation embryo.PLoS One 4:e6086.
Vitale M, Neri LM, Manzoli L, Galanzi A,Rana R, Antonucci A, Papa S. 1989.Improved bromodeoxyuridine/DNA anal-ysis by anti-BudR monoclonal antibodyversus right angle light scatter. Histo-chemistry 93:9–11.
ZAR1L REGULATES PREIMPLANTATION DEVELOPMENT 423
Dev
elop
men
tal D
ynam
ics

Wang S, Hu J, Guo X, Liu JX, Gao S.2008. ADP-Ribosylation factor 1 regu-lates asymmetric cell division in femalemeiosis in the mouse. Biol Reprod 80:555–562
Wassarman PM, Kinloch RA. 1992. Geneexpression during oogenesis in mice.Mutat Res 296:3–15.
Watanabe T, Totoki Y, Toyoda A,Kaneda M, Kuramochi-Miyagawa S,Obata Y, Chiba H, Kohara Y, Kono T,Nakano T, Surani MA, Sakaki Y,
Sasaki H. 2008. Endogenous siRNAsfrom naturally formed dsRNAs regu-late transcripts in mouse oocytes.Nature 453:539–543.
Wu X, Viveiros MM, Eppig JJ, Bai Y, Fitz-patrick SL, Matzuk MM. 2003a. Zygotearrest 1 (Zar1) is a novel maternal-effect gene critical for the oocyte-to-embryo transition. Nat Genet 33:187–191.
Wu X, Wang P, Brown CA, Zilinski CA,Matzuk MM. 2003b. Zygote arrest 1
(Zar1) is an evolutionarily conservedgene expressed in vertebrate ovaries.Biol Reprod 69:861–867.
Yang WH, Yu JH, Gulick T, Bloch KD,Bloch DB. 2006. RNA-associated pro-tein 55 (RAP55) localizes to mRNAprocessing bodies and stress granules.Rna 12:547–554.
Yeo S, Lee KK, Han YM, Kang YK. 2005.Methylation changes of lysine 9 of his-tone H3 during preimplantation mousedevelopment. Mol Cells 20:423–428.
424 HU ET AL.
Dev
elop
men
tal D
ynam
ics





























