Embed Size (px)
Citation preview

Annals of the Rheumatic Diseases, 1988; 47, 232-240
Morphology of surface synoviocytes in situ at normaland raised joint pressure, studied by scanningelectron microscopyJ N McDONALD AND J R LEVICK
From the Department of Physiology, St George's Hospital Medical School, London
SUMMARY The synovial surface in rabbit knees was examined by scanning electron microscopy(SEM) to define normal surface contour, cell shape, and interstitial exposure. Comparison was
made between specimens excised before immersion fixation (I), fixed in situ by vascularperfusion (V) before excision, or fixed in situ under an effusion pressure of 5-25 cmH20 (E). Thedeeply convoluted appearance of rabbit areolar-muscular synovium fixed after excision (I) was
found to be an artefact; areolar-muscular synovium fixed in situ (V) was much smoother. Thewell documented cobblestone contour of immersion fixed adipose synovium was present afterfixation in situ, but may be exaggerated by the SEM preparative process. At higher magnificationthe synoviocytes showed evidence of considerable surface activity (smooth granules, largercauliflower-like excrescences, thin lamelliform filopodia). Cell shape was variable but many
synoviocytes extended long cytoplasmic processes along the surface, producing fibroblastoid andeven stellate outlines. At an intra-articular pressure of 25 cmH20 (E) the cytoplasmic processeswere elongated and branched, creating a highly dendritic outline. Also, the exposure ofinterstitium increased markedly at the higher pressure. It is concluded that e,xtension of lengthycytoplasmic processes is a feature of normal healthy synoviocytes, and that an increase ininterstitial area with joint pressure contributes to the increased hydraulic permeability ofsynovium at raised pressure.
Key words: synovial surface structure, dendritic morphology.
The surface morphology of the synovial intima is ofinterest in relation to two areas of current research.Firstly, the hydraulic permeability of the intima hasbeen shown to increase at raised intra-articularpressures and fluid volumes,' and intimal structurecould provide clues to the cause. Secondly, asubpopulation of dendritic (stellate) synovial cells inrheumatoid synovium has recently excited consider-able attention,2-7 rendering it a matter of someurgency to define the shape of normal synovial cellsin situ for comparison.The synovial lining consists of a discontinuous
layer of cells (synoviocytes), which mostly lackintercellular junctions and are separated by a fibrousinterstitium.8 9 The shape of normal synovial lining
Accepted for publication 18 September 1987.Correspondence to Dr J R Levick, Department of Physiology,St George's Hospital Medical School, London SW17 ORE.
cells is evidently complex and not easy to visualisefrom histological and transmission electron micro-graphs (TEMs). Cytoplasmic processes -1 ,um wide,apparently separated from cell bodies, are oftencaught in transverse or oblique section at thesynovial surface, e.g., in man,8 ") pig,' and rabbit,12indicating that at least some normal intimal cellshave long, finger-like processes extending over thesurface. Such processes are quite distinct fromintraluminal projections such as the leaf-like filo-podia of 'A' cells. In contrast, however, lining cellsreconstructed from serial TEM sections showed asimpler morphology, without extensive cytoplasmicprocesses. 13
Scanning electron microscopy (SEM) offers amore direct picture of synovial surface morphology.Many early low power studies of human, rabbit,dog, pig, and rat synovium concentrated mainly onsurface contour,1421 though Wysocki and Brinkhous
232
copyright. on D
ecember 3, 2021 by guest. P
rotected byhttp://ard.bm
j.com/
Ann R
heum D
is: first published as 10.1136/ard.47.3.232 on 1 March 1988. D
ownloaded from

Surface synoviocytes in situ at normal and raised joint pressure 233
briefly noted 'dendritic cytoplasmic processes' overcanine fibrous synovium in addition to projectionsinto the joint space.19 The surface of human andcanine areolar synovium appeared folded andfurrowed,16 17 19 21 but as the tissue was not fixedin situ the significance of this is not clear. Adiposesynovium by contrast had a regular cobblestoneappearance. 14 17 19 Date studied the surface atsufficient magnification to reveal detailed cellmorphology,2 and in an excellent but little quotedpaper he described numerous flat interdigitating cellprocesses, cauliflower-like excrescences, andexposed interstitium in human and rabbit synovium.Cell surface protrusions and intercellular spaceswere earlier reported in human synovium21 23 andtenosynovium. All these tissues were excisedbefore fixation.There seem to have been no scanning electron
microscopic studies of synovium fixed in situ. Thepresent SEM study aims to clarify (a) the effect ofexcision on synovial surface contour; (b) the normalmorphology of surface synoviocytes in situ; and (c)the effect of intra-articular distension by fluid onsurface morphology. Morphometric investigationsby light and transmission electron microscopy are inprogress to quantify further the structural con-sequences of distensions.
Materials and methods
New Zealand rabbits (2-3 kg) were anaesthetisedwith intravenous pentobarbitone and urethane.Three kinds of synovium8 25 were sampled in each ofsix knees from three animals, namely areolar-muscular synovium from the medial aspect of thesuprapatellar pouch, adipose synovium from theinfrapatellar fat pad, and mixed synovium from theposteromedial pouch.
TECHNIQUES OF FIXATIONSynovial samples of surface area 2 mmx3 mm andthickness 2 mm were excised by scalpel undervarious conditions.
Immersion fixationIn two joints soft unfixed tissue was excised, thenimmersed at once in fixative at room temperaturefor two hours ('post-fixed' control). No intra-articular injection was made into these joints.
Vascular perfusionIn two joints the synovial lining was first fixedin situ, then excised and reimmersed in fixative atroom temperature for two hours ('pre-fixed'control). No intra-articular infusion was made intothese joints. Fixation in situ was achieved by
vascular perfusion. The abdominal aorta andvena cava were cannulated. After an initial flushingof the knee vasculature with buffer solution (0-05 Msodium cacodylate, 50 g/l sucrose) fixative wasinfused through the aorta from a reservoir 163 cmabove heart level (120 mmHg infusion pressure) andallowed to drain away through the caval cannula.The joint very quickly became stiff. After sevenminutes of vascular perfusion the knee was openedand samples taken of the firm, fixed tissue.
Articular perfusionIn two, volume expanded joints the synovial liningwas fixed in situ for one hour at a raised intra-articular fluid pressure (5 cmH2O and 25 cmH2Orespectively) before sample excision, then immer-sion fixed at room temperature for a further onehour. For these experiments the joint cavity wascannulated by two 21G cannulae. Intra-articularpressure was recorded through one cannula andKrebs' solution ion infused through the other from areservoir whose height governed the intra-articularpressure, as described previously.26 Krebs' solution,27which contains 0*9 g glucose/l as energy source, waspre-equilibrated with 95% 02/5% CO2 at pH 7-4.Intra-articular temperature was 35°C, as is normal inthe rabbit knee.The desired intra-articular pressure was
approached in three or four pressure steps, allowing15 minutes a step to reproduce the conditions of anearlier study of trans-synovial fluid exchange.26 Theinfusate was then switched to fixative and theinfused fluid allowed to drain out continuouslythrough a third cannula connected to a collectionreservoir with an overspill. By appropriate setting ofthe heights of the infusion reservoir and collectionreservoir, and adjustment of the resistance of theoverflow line by a screw clamp, the intra-articularpressure was held constant at 5 or 25 cmH2O duringcontinuous perfusion of the synovial cavity withfixative at 1 ml/min. At this flow the rate constantfor the rise in intra-articular fixative concentrationwas 0X5-1X43 min-1. A longer period (one hour) wasrequired for intra-articular fixation than for per-vascular fixation because of the slower permeationof fixative into the periarticular tissues (e.g.,muscle), which had to be fixed too in order toprevent recoil on opening the joint cavity.The fixative in all cases was half strength
Karnovsky solution-namely, 2% paraformalde-hyde, 2-5% glutaraldehyde in 0-05 M sodium caco-dylate buffer (pH 7.4) with 50 g/l sucrose as osmoticbuffer.28
PROCESSING OF FIXED SAMPLES FOR SEMSamples were washed with three changes of buffer
copyright. on D
ecember 3, 2021 by guest. P
rotected byhttp://ard.bm
j.com/
Ann R
heum D
is: first published as 10.1136/ard.47.3.232 on 1 March 1988. D
ownloaded from

234 McDonald, Levick
(0.05 M sodium cacodylate with 50 g/l sucrose,osmolarity 250 mmol/l), diced finely and post-fixedin 1% osmium tetroxide in buffer for three hours atroom temperature. After double rinsing in bufferthe samples were dehydrated in an ethanol series,substituted with fluorocarbon, dried at the criticalpoint with liquid CO2 (E300, Polaron Ltd), andmounted on stubs with Araldite. Specimens weresplutter coated with gold (ES100 series II, PolaronLtd) and examined in a Coates and Welter fieldemission scanning electron microscope at 16-5 kV.
Results
GENERAL SURFACE CONTOURThe cobblestone profile of adipose synovium
surface'4 19 was observed in tissue fixed in situ aswell as in post-fixed samples (Fig. 1), i.e., it wasindependent of the fixation method. Examination ofthe cut edge of the specimen showed that eachsurface 'cobble' (diameter 50 ,um) was due to a fatcell some distance below the intima. The surfaceconformation of non-adipose synovium depended,however, on the fixation method. Tissue excisedbefore fixation had a wrinkled, convoluted surfacewith roughly parallel furrows 0-01 to 0-15 mm apart(Fig. 1A), very like immersion fixed human andcanine areolar synovium. 16 17 19 20 In contrast, thesurface of samples fixed in situ was much smoother,though a few, wider spaced furrows were still seen inpre-fixed control and 5 cmH2O samples. In samplesfixed at 25 cmH20, surface furrows were elimin-
Fig. 1 Lowerpower scanning electron micrographs ofrabbit synovial surface to illustrate effect offixation method andsubstructure on surface contour. Scale bar=50 tum. Red cells liberated during excision ofsample have setted on the surfacein places. (A) Areolarsynovium from medial suprapatellarpouch excised beforefixation then immersion fixed (post-fixedcontrol). Surface is deeply furrowed. (B) Areolarsynovium from medial suprapatellarpouchfixed in situ by vascularperfusion before excision (pre-fixed control). Surface is much smoother. (C) Adipose synovium over infrapatellar fatpad,fixed in situ by vascular perfusion. The cobblestone surface persists and is due to underlyingfat cells. (D) Areolar synoviumfrom suprapatellar pouch fixed in situ in a joint distended to 25 cmH20 intra-articular pressure. The long branchedhummocks are probably due to blood vessels below the synovial surface.
copyright. on D
ecember 3, 2021 by guest. P
rotected byhttp://ard.bm
j.com/
Ann R
heum D
is: first published as 10.1136/ard.47.3.232 on 1 March 1988. D
ownloaded from
Surface synoviocytes in situi at normtal and raised joint pressure 235
ated, but long branching cylindrical ridges wereconspicuous (Fig. ID).
CEI.UILAR MORPHOLOGY AT NORMAL JOINTPRESSUREThe following description applies to synovium fixedin situ by vascular perfusion. At magnifications of>l100x the remarkably complex nature of thesynovial surface became apparent. The surfaceconsisted of cell bodies, cell processes, and, inbetween. interstitium exposed directly to the jointcavity. The general appearance varied considerably,some regions resembling a patchwork quilt (corres-ponding to relatively regular, non-dissected celloutlines) and others resembling a mass of worms(corresponding to irregular cells with extensive
t'5' , ' z = f.
Fig. 2 Tracings ofintimal cell shapefrom the surface ofsynovium fixed in situ at normal joint pressure (cells a-f,pre-fixed controls) and at 25 cmH2O joint pressure(cells g-i). Scale bars=5 tm. Dashed lines indicateobscuration by adjacent overlying cells. Cells a-f wereselected to illustrate the diversity and gradation ofnormalcell morphology from (a) ovoid plate through (c, d)fibroblastoid and (e) stag headfern to (f) stellate. Intra-articular distension caused a great increase in dendritic(stellate) forms (e.g. g-i).
processes), with intermediate gradations. Highermagnifications showed the cell outline was veryvariable, and a progressive, continuous gr.adation incomplexity could be discerned (Fig. 2).
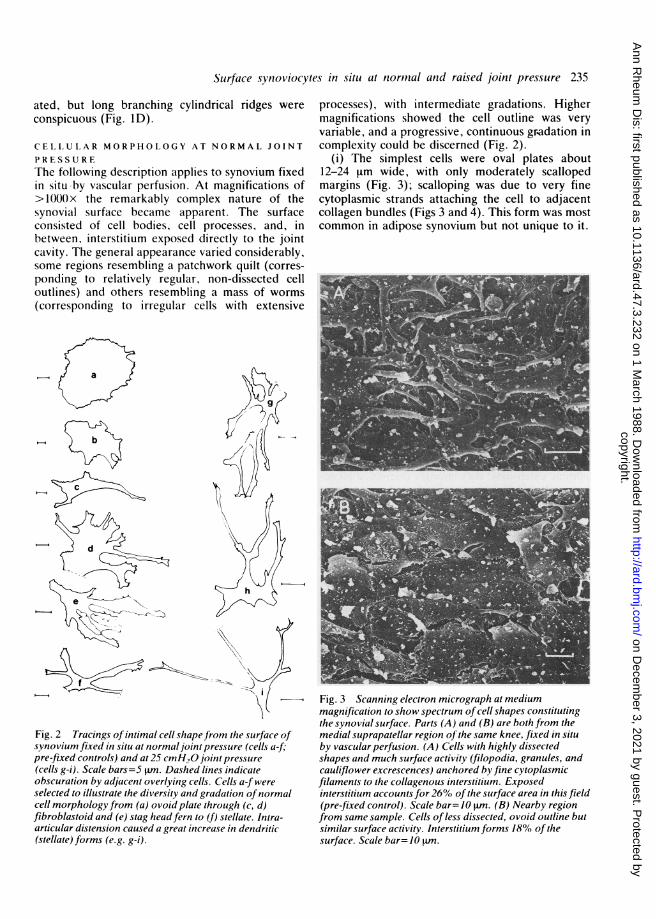
(i) The simplest cells were oval plates about12-24 Lrm wide, with only moderately scallopedmargins (Fig. 3); scalloping was due to very finecytoplasmic strands attaching the cell to adjacentcollagen bundles (Figs 3 and 4). This form was mostcommon in adipose synovium but not unique to it.
Fig. 3 Scanning electron micrograph at mediummagnification to show spectrum of cell shapes constitutingthe synovial surface. Parts (A) and (B) are both from themedial suprapatellar region of the same knee, fixed in situby vascular perfusion. (A) Cells with highly dissectedshapes and much surface activity (filopodia, granules, andcauliflower excrescences) anchored by fine cytoplasmicfilaments to the collagenous interstitium. Exposedinterstitium accounts for 26% ofthe surface area in this field(pre-fixed control). Scale bar=-10 tm. (B) Nearby regionfrom same sample. Cells of less dissected, ovoid outline butsimilar surface activity. Interstitium forms 18% of thesurface. Scale bar= 10 .m.
copyright. on D
ecember 3, 2021 by guest. P
rotected byhttp://ard.bm
j.com/
Ann R
heum D
is: first published as 10.1136/ard.47.3.232 on 1 March 1988. D
ownloaded from

236 McDonald, Levick
Fig. 4 HighI power scamminiiieleC tron mnicro)grap1hlt()osIhowCexposed Syniov'ial iiitervilitumobetween two cells. Ani irregularcollagen mesh work ypnhe/citntercellulalr gaps. I'Iie cells arcattached to tlie collageni netiwork 1bvfine cytoplasmnic strnl(Is (arrows)ws'hich p)rodlce the scallopedimargin. The cell on i/ic lef skhowslvo surjace features-a cu.sp-likeleaf o)f cvtoplasni (ltianellifOlI-tifilopodium, If) andI a smoothsulrface grauilile 0-5>tu in (lialdialter(g). Po.steromnedial s silovil'li fixedini sitli at 2'S cmH.0. Scale bar= 'rtun. Thle insert s/iow's (1 tlir hlkilithof 'sutfacefce leature, i/ic largecauliflowscr-like excrescen(c.
(ii) Cells of 'fibroblastoid' outline were moreirregular, with two to six short, flat, broad cyto-plasmic extensions: they strongly resembled mono-layer cultured fibroblasts as illustrated in a standardSEM atlas.29
(iii) Cells with greater elongation of the flattened,cytoplasmic processes were termed stag head ferncells (Fig. 2) owing to their strong resemblance tothat plant. Such processes interdigitated with othersand sometimes ran over or under adjacent cells.
(iv) The most dissected cell outlines (stellate ordendritic) comprised ;two to six cytoplasmicprocesses 1-2 jtm wide and typically 6-12 jim long(max 25 Rm). The processes entwined and over-lapped, sometimes bifurcated, and often ended asflattened expansions with fine attachment filamentsproducing scalloped margins.
Because cell shape comprised a continuousspectrum, the relative prevalence was difficult tomeasure, but forms (ii) and (iii) seemed mostcommon. The whole cell outline was not alwaysvisible-some bodies emerged from beneath aninterstitial layer or had a thin skein of fibrils partiallycovering them. The shape of the cell below thesurface plane was not of course revealed by SEM;nor were nuclear regions detected as these areusually 5 jtm below the cell surface (TEM). Distinc-tive forms which might correspond to classical 'A'and 'B' cells of TEM¶ were not recognised by SEM,as noted by others.23
CELI SURFACE STRUCTURFSCell surface structures abounded and were of threekinds (Figs 3 and 4).
(a) Fine fingers and flaps of cytoplasm (thickness0-O1-2 jtm) protruded into the joint cavity (Fig. 4).corresponding to the lamelliform filopodiai seeni inTEMs. "
(b) Most cells had one or more larger (1 0-4.5 jimwide), knobbly excrescences protuding into the jointcavity ('cauliflower-like protrusion' as described byDate22). At high magnification these resolved intocomplicated folds of cytoplasm, fingers and clustersof small smooth granules of 02-0-3 jm diameter.
(c) Small smooth round granules were alsocommon in isolation on cell and cell process surfacesand, being 0-2-0-5 jtm wide, were probably the samestructures as those occurring in clusters on thecauliflower excrescences.
SYNOVIAI INTERSTITIUMCells and processes were not in contact butseparated by gaps filled with interstitium, visible as arandom meshwork of collagen (Fig. 3-low pressure;Fig. 4-high pressure). A thin lacework of fibrilsextended partly over some cell surfaces. The areabetween cells, estimated by a standard morpho-metric point counting method,3' averaged 26(relative standard error (RSE) 3)% over 7x 103 jim2areolar synovium. The range was from 18% to 29%of the surface for prefixed control tissue, the lower
copyright. on D
ecember 3, 2021 by guest. P
rotected byhttp://ard.bm
j.com/
Ann R
heum D
is: first published as 10.1136/ard.47.3.232 on 1 March 1988. D
ownloaded from

Surface synoviocytes in situ at normal and raised joint pressure 237
Fig. 5 Scanning electronimicrograph ofposteromedialsynovial surface fixed in situt at ajoint distended to 25 cmH,O intra-articular pressure. Cellmorphology is more dendritic andmany processes have one or morebifurcations. Interstitium, seetn as acollagen mesh work, constitutes agreater proportion of the surfacearea (50 3%). Many surfaceexcrescences are presenit, appearingas white structures protrudinig initothe joint cavity. Scale bar= I0 m.
value corresponding to a region of plate-like cells(cell type (a) of Fig. 2) and the higher value to a
region of more dissected cells (types (d), (e), and (f)of Fig. 2). Adipose and posteromedial synovia hadinterstitial areas of 24 (3)% and 29 (3)% respect-ively.
EFFECT OF INTRA-ARTICULAR DISTENSION
The general appearance of the surface did not differmarkedly between tissue fixed in situ at normal jointpressure and tissue fixed in situ at 5 cmH2O. Thelatter is well below the 'yield point' at whichhydraulic permeability begins to increase.' 26Distension by 25 cmH2U intra-articular pressure
(above yield point), however, produced dramaticchanges in surface shape. Although some cells stillretained scalloped oval outlines (especially, postero-medial samples), most became much more dendritic(Fig. 5). The cytoplasmic processes extending alongthe synovial surface (typically three to four) were
much elongated (20-25 ,um) and thinned(0.4-0.8 ism wide), resembling short unmyelinatedaxons. Many were branched and possessed smallsmooth granules or cauliflower-like excrescences, or
both, on their surfaces.Between the cell processes the area of exposed
interstitium was very obviously increased at25 cmH2O. The point counting method applied over
3-8x103 [Lm2 areolar synovium gave an interstitialarea of 53 (RSE 2)%. Preliminary results from a
morphometric study of TEMs also gave a high value(45%). For adipose and posteromedial synoviumthe interstitial areas were 43 (3)% and 35 (3)%respectively.
Structures just beneath the surface created longcylindrical hummocks along the surface (Fig. 1), andinspection of the cut edge of the sample indicatedthat these ridges were raised by microvessels veryclose to the surface.
Discussion
SURFACE CONTOURS
Comparison of tissue excised before fixation withtissue fixed in situ showed considerable deformationof areolar-muscular synovium in post-fixed samples,due presumably to elastic retraction of the soft tissueas it is excised. Inspection of the unfixed, exposedareolar synovial surface in vitro through a dissectingmicroscope in the course of previous studies27indicated that the normal hydrated areolar surfacein situ resembled Fig. 1B rather than Fig. 1A. Weconclude that the highly convoluted surface of rabbitareolar-muscular synovium excised before fixation isto a large extent an artefact. It is important to note,however, that synovium fixed in situ is not com-pletely flat-some furrows persist, and may be func-tionally important in permitting synovium to stretchwith minimal stress during joint movement.
For adipose synovium the fixation method did notinfluence surface contour (cobblestone pattern),presumbly owing to the incompressible nature ofpacked fat globules. The cobblestone appearancemay have been greatly exaggerated, however, by theSEM preparative technique. Rabbit infrapatellaradipose synovium in conventional paraffinembedded histological sections (excised tissue or
copyright. on D
ecember 3, 2021 by guest. P
rotected byhttp://ard.bm
j.com/
Ann R
heum D
is: first published as 10.1136/ard.47.3.232 on 1 March 1988. D
ownloaded from

238 McDonald, Levick
whole knee) has a predominantly smooth convexsurface with only an occasional cobblestone corres-ponding to a fat cell (unpublished observation).Similarly, resin embedded adipose synovium has amainly smooth surface not conforming to underlyingfat cells, both on light microscopy of semithinsections (see Fig. 1 of reference 25) and on TEM(unpublished observation, Fig. 1 of reference 32,and Fig. 2 of reference 19), even when the fat cellsare very superficial.19 The exaggerated cobblestonepattern of SEM may be due to collapse of synoviuminto the dehydrated angles between adjacent fatcells.
INTERCELLULAR SPACESThe substantial area of synovial surface composedsimply of interstitium at normal joint pressureagrees with previous SEM studies.21-23 The possi-bility of artefact must be considered as some tissueshrinkage occurs during preparation for SEM,though the critical point drying method is designedto minimise postdehydration changes. The linearshrinkage of synovial samples after ethanol dehydra-tion, measured with a Vernier travelling micro-scope, averaged 7%. We consider it unlikely,however, that the interstitial exposure was primarilyan artefact of differential tissue shrinkage, forseveral reasons. Firstly, the observations agree withthe findings by light microscopy that the intimal cellsform a discontinuous layer embedded in a fibrousinterstitium.8l' 30 The interstitial exposure inFig. 3B bears a close resemblance to that shown bysilver nitrate staining of hydrated synovium(e.g., Fig. 12 of reference 8, Fig. 7 of reference 9).Secondly, the observation is supported by TEM;17-21% of the surface of unstretched rabbit kneesynovium was reported to consist of interstitium in amorphometric TEM study.12 Thirdly, the obser-vations are compatible with physiological proper-ties-namely, permeability of the intima to quitelarge extracellular tracers like colloidal carbon.33Fourthly, cell bodies were not infrequently partiallyoverlapped by interstitium, a situation unlikely toarise from cell retraction.Specimens prepared by the alternative method of
freezing and freeze drying do not seem to havediffered markedly from those prepared by criticalpoint drying as here.19 2( A low temperature SEMstudy of unfixed hydrated synovium might,however, be of interest.
CELL SHAPEWe must consider whether the observed cell shapestruly represent those in vivo. Opportunities for cellshape to change might arise (a) during exposure tointra-articular fluid for one hour, (b) in the course of
fixation, if slow, and (c) owing to shrinkage duringprocessing.
(a) The intra-articular fluid was of physiologicalionic composition (Krebs' solution), of normal pH,temperature, and osmolarity, with glucose as energysource, and pre-equilibrated with an oxygen/CO2mixture. Moreover, intimal gas tensions and nutri-tion in vivo were maintained by the synovialcapillary network, which exceeds that of skeletalmuscle in density.25 Quantitative physiologicalstudies show that the permeability of the intima isunaltered by several hours of intra-articular Krebsinfusion.1 In addition, TEM studies (unpublished)show a similar synovial structure after per-vascularfixation with no intra-articular infusion and afterone hour of intra-articular Krebs infusion belowyield pressure followed by intra-articular fixation.For these various reasons it seems unlikely that cellshape was changed by the 'stress' of intra-articularinfusion of Krebs' solution under low pressure.
(b) It seems unlikely that the novel synovialfixation methods changed cell shape. Fixation byvascular perfusion was very rapid because thesynovial capillaries deliver fixative in seconds towithin 6-11 jm of the synovial surface,25 acrosswhich distances diffusion times are very short.Although the half life for fixative concentrationgrowth was slower with the intra-articular fixationmethod (30 seconds at 5 cmH20), the SEM resultsat 5 cmH20 were similar to those after per-vascularfixation. Synoviocytes in culture are capable ofreversibly transforming between fibroblastoid andmore stellate morphologies, but such changes takemore than three to four hours (typically 16-18 hours),4 6 a time scale far longer than allowedhere.
(c) The demonstration of extensive cell processesfits with deductions from histology.8-1 As long agoas 1930 Sigurdson, using the silver nitrate method onunfixed hydrated tissue, noted that some synovialcell surface profiles were relatively regular whileothers were 'large and irregular with numerousprocesses'.The observed cell shapes, with intermeshing
cytoplasmic process of variable length and width,agree with studies of immersed, fixed tissue.8 22 34Many cells strongly resembled fibroblasts in mono-layer culture,35 and some were more dendritic. Itseems clear that even normal synovial intimal cellsextend many cytoplasmic processes, and that theseare made longer and thinner by intra-articulardistension. Moreover, living surface synoviocytesfreshly detached from normal pig synovium alsohave a dendritic or stellate outline.3b The demon-stration of a multiplicity of cytoplasmic processes innormal intimal cells in situ raises an interesting
copyright. on D
ecember 3, 2021 by guest. P
rotected byhttp://ard.bm
j.com/
Ann R
heum D
is: first published as 10.1136/ard.47.3.232 on 1 March 1988. D
ownloaded from

Surface synoviocytes in situ at normal and raised joint pressure 239
question-Does the dendritic subpopulation of cellscultured from rheumatoid synovium2-7 represent aspecial class of cells involved specifically in thedisease process or are such cells relatively normal,deformable intimal cells expressing their inherentphenotypic tendency to extend cytoplasmic pro-cesses? Reversible transformation of culturedrheumatoid dendritic cells into fibroblastoidmorphology,"6 collagenase and prostaglandin E2synthetic ability,4 6 and surface antigenicity7 haveled to suggestions that such cells are closely relatedto fibroblasts-as are normal intimal 'B' cells-rather than to dendritic antigen presenting macro-phages. Our observations of the synoviocyte in situsupport that view. It appears that diverse cell typescan adopt a stellate morphology given the appro-priate conditions (e.g., tissue macrophages, bloodmonocytes in culture, synovial intimal cells, astro-cytes).
CELL SURFACE FEATURESThe marked surface activity of a high proportion ofthe cells was suggestive of endocytosis or exocytosis,or both. The structure of filopodia, such as that inFig. 4, seems compatible with endocytosis. Thesmooth granules of diameter 02-0-5 [tm have beenreported also in tenosynovium23 and rheumatoidsynovium.20' 23 They may be secretory vesicles. Thelarger complex cauliflower protruberances, de-scribed by Date22 and confirmed here, could beeither phagocytic or secretory in function. As theycontain clusters of smaller granules a secretoryfunction is perhaps more likely.
EFFECT OF INTRA-ARTICULAR VOLUMEEXPANSION ON STRUCTUREThe structural changes at 25 cmH2O fluid pressurehelp account for the increase in hydraulicpermeability of synovium above yield pressure(9.5 cmH2O). Several possible explanations havebeen advanced for the latter, including increase inintercellular area available for fluid exchange,' butdirect evidence has until now been lacking. Here amarked increase in interstitial area was visualiseddirectly. The increased area for fluid exchange mustcontribute to the reported increases in intimalhydraulic permeability at raised pressure; but inter-stitial area changes alone (-2x) do not seemquantitatively great enough to account fully for thefar greater permeability changes.' 26 Studies ofstructural changes by TEM are in progress to assesswhether path length as well as area is modified byintra-articular distension. Wide intercellular spacesare also prominent in scanning electron micrographsof the rheumatoid synovial surface.23 37
This research was supported by the Arthritis and RheumatismCouncil.
References1 Knight A D, Levick J R. Effects of fluid pressure on thehydraulic conductance of interstitium and fenestrated endo-thelium in the rabbit knee. J Physiol (Lond) 1985; 360: 311-32.
2 Woolley D E, Brinckerhoff C E, Mainardi C L, Vater C A,Evanson J M, Harris E D. Collagenase production by rheuma-toid synovial cells: morphological and immunohistochemicalstudies of the dendritic cell. Ann Rheu,n Dis 1979; 38: 262-70.
3 Winchester R J, Burmester G R. Demonstration of la antigenson certain dendritic cells and a novel elongate cell found inhuman synovial tissue. Scand J Immunol 1981; 14: 439-44.
4 Baker D G, Dayer J M, Roelke M, Schumacher H R,Krane S M. Rheumatoid synovial cell morphologic changesinduced by a mononuclear cell factor in culture. ArthritisRheum 1983; 26: 8-14.
5 Hendler P L, Lavoie P E, Werb Z, Chan J, Seaman W E.Human synovial dendritic cells. Direct observation of transitionto fibroblasts. J Rheumatol 1985; 12: 660-4.
6 Gadher S J, Woolley D E. Comparative studies of adherentrheumatoid synovial cells in primary culture: characterisation ofthe dendritic (stellate) cell. Rheumatol Int 1987; 7: 13-22.
7 Heino J, Viander M, Peltonen J, Kouri T. Adherent cells fromrheumatoid synovia; identity of HLA-DR positive stellate cells.Ann Rheumn Dis 1987; 46: 114-20.
8 Key J A. The synovial membrane of joints and bursae. In:Cowdry E V, ed. Special cytology. Vol 2. New York:P B Hoeber, 1928: 1054-74.
9 Sigurdson L A. The structure and function of articular synovialmembrane. Journal of Bone and Joint Surgery 1930; 12: 603-39.
10 Hamerman D, Stephens M, Barland P. Comparative histologyand metabolism of synovial tissue in normal and arthritic joints.In: Mills L C, ed. Inflammationi and disease of connective tissue.Philadelphia: Saunders, 1961: 158-68.
11 Fell H B, Glauert A M, Barratt M E J, Green R. The pigsynovium. I. The intact synovium in vivo and in organ culture.J Anat 1976; 122: 663-80.
12 Knight A D, Levick J R. Morphometry of the ultrastructure ofthe blood-joint barrier in the rabbit knee. Q J Exp Physiol 1984;69: 271-88.
13 Graabaek P M. Ultrastructural evidence for two distinct typesof synoviocytes in rat synovial membranes. J Ultrastruct Res1982; 78: 321-39.
14 Woodward D H, Gryfe A, Gardner D L. Comparative study byscanning electron microscopy of synovial surfaces of fourmammalian species. Experientia 1969; 25: 1301-3.
15 Gryffe A, Gardner D L, Woodward D H. Scanning electronmicroscopy of normal and inflamed synovial tissue from arheumatoid patient. Lancet 1969; ii: 156-7.
16 Redler I, Zimmy M L. Scanning electron microscopy of normaland abnormal articular cartilage and synovium. J Bonie JointSurg fAmi 1970; 52A: 1395-404.
17 Highton T C, Donaldson L A. Scanning electron microscopy ofsynovial joints with preservation of mucopolysaccharides. Pro-ceedings of the University of Otago Medical School 197t); 48:64-5.
18 Highton T C. A scanning electron microscopic study of synovialjoints. J R Coll Physicians Lond 1971; 6: 25-32.
19 Wysocki G P, Brinkhous K M. Scanning electron microscopy ofsynovial membranes. Archives of Pathology 1972; 93: 172-7.
20 Cameron H U, MacNab I. Scanning electron microscopicstudies of the hip joint capsule and synovial membrane. Cant JSurg 1973; 16: 388-92.
21 Gaucher A, Faure G, Netter P, Pouvel J, Duheille J. Apport dela microscopie electronique a balayage a l'tude de la synovialehumaine normale et pathologique. Rev Rhum Mal Osteoartic1976; 43: 51-60.
copyright. on D
ecember 3, 2021 by guest. P
rotected byhttp://ard.bm
j.com/
Ann R
heum D
is: first published as 10.1136/ard.47.3.232 on 1 March 1988. D
ownloaded from

240 McDonald, Levick
22 Date K. Scanning electron microscope studies on the synovialmembrane. Arch Histol Jpni 1979; 42: 517-31.
23 Fujita T, Inoue H, Kodama T. Scanning electron microscopy ofthe normal and rheumatoid synovial membranes. Arch His-tolJpat 1968; 29: 511 -22.
24 Inoue H, Takasugi H. Akahori 0. Surface study of teno-synovium in hens and humans by electron microscopy. thieHand 1976; 8: 222-7.
25 Knight A D, Levick J R. The density and distribution ofcapillaries around a synovial cavity. Q J Exp Physiol 1983; 68:629-44.
26 Levick J R. The influence of hydrostatic prcssure on trans-synovial fluid movement and on capsular cxpansion in therabbit knee. J Physiol (Lotnd) 1979; 289: 69-X2.
27 Knight A D, Levick J R, McDonald J N. Relation betwecntrans-synovial flow and plasma osmotic pressure. with anestimation of the albumin reflection coefficient in the rabbitknee. Q J Fxp Phs'sisol (in press).
28 Karnovsky M J. A formaldchydc-glutaraldehydc fixative ofhigh osmolality for use in electron microscopy. J Cell Biol 1965;27: 137A.
29 Kessel R G, Kardon R H. Tiosoies atid organs: a text-atlas o)fscanning electroni inicroscopY. San Francisco: Frceman & Co,1979: 12.
30 Ghadially F N. Fine structure of Joints. In: Sokoloff L. cd. 7he
joint.S antd ss'noiaflfid. Vol 1. Ncw York: Ac,idcmic l'i-css,1978: 105-76.
31 Aherne W A. Dunnill M S. Morphooietr-v. Londoii: Ariiold,1982.
32 Ghadially F N. Roy S. Ultrastructurc of rabbit sv ros aimembrane. Annii Rheion Dio 1966: 25: 318-25.
33 Levick J R. Blood flow and mass transport in svin-ovial joilnts.In: Rcnkin E M, Michel C C. cds. Hanidbook of plisvsioloyv.Vol IV. 7he inicrocirculation. Bethesda M D: AmicricianPhysiological Socicty. 1984: 917-47.
34 Elliott S. Cookc r D V. Scanning clectroni microscops ofantigen induccd artht-itic joints. 1. Inflarrimmatory cell intcactionat synovial-meniscal surf.accs durinig the 1Arthus e-esponsc.J Rhetiomiatol 1986: 13: 401-7.
35 Pratt R M. Yaimada K M. Oldenl K. Ohani'an S H. HlasclIl V C.Tunamycin-induced alteration in svnthcsis of sulphattcd pi-o-teoglycans and ccll sur-face miorphology in the chick emrnbyofibroblast. Exp Cell Res 1978; 118: 245-52.
36 Barratt M E J, Fcll I1 B. Coombs R R A. Glaucrt A M. Thc pigsynovium. II. Some propertics of isol;itcd intlimal cells. J Anat1977; 123: 47-66.
37 Hayashi T. Three-dimensionail observations of rhcumaitoidsynovial membranc.so-yachli 1976: 16: 3>5-6. (Englishabstract. )
copyright. on D
ecember 3, 2021 by guest. P
rotected byhttp://ard.bm
j.com/
Ann R
heum D
is: first published as 10.1136/ard.47.3.232 on 1 March 1988. D
ownloaded from




























