Embed Size (px)
Citation preview

BioOne sees sustainable scholarly publishing as an inherently collaborative enterprise connecting authors, nonprofit publishers,academic institutions, research libraries, and research funders in the common goal of maximizing access to critical research.
Morphology and Identification of First Instars of African BlowFlies (Diptera: Calliphoridae) Commonly of Forensic ImportanceAuthor(s): Krzysztof Szpila and Martin H. VilletSource: Journal of Medical Entomology, 48(4):738-752. 2011.Published By: Entomological Society of AmericaDOI: http://dx.doi.org/10.1603/ME10238URL: http://www.bioone.org/doi/full/10.1603/ME10238
BioOne (www.bioone.org) is a nonprofit, online aggregation of core research in thebiological, ecological, and environmental sciences. BioOne provides a sustainable onlineplatform for over 170 journals and books published by nonprofit societies, associations,museums, institutions, and presses.
Your use of this PDF, the BioOne Web site, and all posted and associated content indicatesyour acceptance of BioOne’s Terms of Use, available at www.bioone.org/page/terms_of_use.
Usage of BioOne content is strictly limited to personal, educational, and non-commercialuse. Commercial inquiries or rights and permissions requests should be directed to theindividual publisher as copyright holder.

MORPHOLOGY, SYSTEMATICS, EVOLUTION
Morphology and Identification of First Instars of African Blow Flies(Diptera: Calliphoridae) Commonly of Forensic Importance
KRZYSZTOF SZPILA1 AND MARTIN H. VILLET2
J. Med. Entomol. 48(4): 738Ð752 (2011); DOI: 10.1603/ME10238
ABSTRACT Scanning electron microscopy images of the Þrst instars of Calliphora croceipalpisJaennicke, 1876; Chrysomya chloropyga (Wiedemann, 1818); Chrysomya marginalis (Wiedemann,1830); andChrysomya putoria (Wiedemann, 1830) (Diptera: Calliphoridae) are presented for the Þrsttime, and the following morphological structures are documented: pseudocephalon, antenna, max-illary palpus, facial mask, labial lobe, thoracic and abdominal spinulation, spiracular Þeld, posteriorspiracles, and anal pads. Light microscopy photographs and line illustrations are provided for thecephaloskeleton in lateral and ventral views, and the “ectostomal sclerite” and “chitinized teeth” ofthe cephaloskeleton are recognized as integral parts of the mouthhooks. New diagnostic features ofthe cephaloskeleton and the spinulation of the abdominal segments are described. These results allowreÞnement, clariÞcation, and correction of earlier descriptions, which are reviewed. The relativetaxonomic importance of various morphological characters of the Þrst instars of necrophagous blowßies is discussed, and details of the cephaloskeleton and the spinulation of the abdominal segmentsare highlighted as the characters most useful for species identiÞcation. Finally, a key for identifyingÞrst instars of common African carrion blow ßies is provided.
KEY WORDS Calliphoridae, larvae, morphology, identiÞcation, Africa
The recent rapid development and popularization offorensic entomology (Amendt et al. 2010) has prior-itized detailed studies of forensically important in-sects. Although great progress in molecular methodsof identiÞcation has occurred, the classic morpholog-ical approach to species identiÞcation is still faster andcheaper for most laboratories. In cases where insectshave colonized a body only recently, the larvae maystill be very young, which poses challenges for mor-phological identiÞcation because of the scarcity andfragmentary nature of relevant literature. Larvae inthe Þrst instar are as relevant for forensic purposes asany later stage, and they offer a better degree ofprecision in estimating a postmortem interval thanolder larvae because they have had less time to accu-mulate random variation in the variables use to makethe estimate (Villet et al. 2010). Of course, they shouldbe used to estimate a postmortem interval only if theyare the oldest immature specimens of their speciespresent in a case.
Contemporary knowledge of the morphology ofÞrst instars of necrophagous blow ßies is reasonablegood in comparison to other groups of Diptera.Several studies on this topic have been published inthe past 5 yr (Thyssen and Linhares 2007, Szpila etal. 2008, Sukontason et al. 2008, Ekanem and
Umoetuk 2009, Florez and Wolff 2009, Mendonca etal. 2010). Good descriptions are accumulating buteven some new descriptions treat early instars su-perÞcially and provide insufÞcient useful informa-tion for taxonomic purposes, let alone phylogeneticand other subsidiary research. Drawings and pho-tographs of morphological details can be small andunclear (Thyssen and Linhares 2007, Florez andWolff 2009, Ekanem and Umoetuk 2009) or thematerial may be badly preserved (Sukontason et al.2008). In almost all of these studies, the relevantdescriptions are vague and lack mention of taxo-nomically important characters, apparently mod-eled on an outdated tradition of brief descriptionsin older studies (Schumann 1954, Prins 1982,Erzinclioglu 1985, Greenberg and Szyska 1984). Thereasons for this shallow treatment probably lie in thesmall size and brief duration of the Þrst instar, withthe molt into second instar often taking place onlyhours after hatching from the egg (Szpila et al.2008), and in the relatively poor resolution of oldermicroscopy equipment.
ThenecrophagousblowßyfaunaofAfrica is relativelyrich (Zumpt 1956, 1965; Pont 1980; Braack 1986; Rognesand Paterson 2005; Kurahashi and Kirk-Spriggs 2006;VervesandKhrokalo2009),butdetailsof thebiologyandmorphology of the larval stages of many of them remainunknown. However, the assemblage of species regularlydeveloping on large vertebrate carrion (including hu-man corpses) seems to be quite consistent and is largelyrestricted to the following species: Calliphora croceipal-
1 Corresponding author: Department of Animal Ecology, Instituteof Ecology and Environmental Protection, Nicolaus Copernicus Uni-versity, Gagarina 9, 87-100 Torun, Poland (e-mail: [email protected]).
2 Department of Zoology and Entomology, Rhodes University,Grahamstown, 6140 South Africa.
0022-2585/11/0738Ð0752$04.00/0 � 2011 Entomological Society of America

pis Jaennicke, 1876; Chrysomya albiceps (Wiedemann,1819); Chrysomya chloropyga (Wiedemann, 1818);Chrysomya marginalis (Wiedemann, 1830); Chrysomyamegacephala (F., 1794); Chrysomya putoria (Wiede-mann, 1830);Hemipyrellia fernandica (Macquart, 1855);Lucilia cuprina (Wiedemann, 1830); and Lucilia sericata(Meigen, 1826) (Prins 1982, Braack 1987, Louw and vander Linde 1993, Tantawi et al. 1996, Kurahashi and Kirk-Spriggs 2006, Ekanem and Dike 2010). Calliphora vomi-toria (L., 1758) occurs in North Africa (Erzinclioglu1987a), and Calliphora vicina Robineau-Desvoidy, 1830is now found in South Africa (Williams and Villet 2006),and both may therefore become signiÞcant in forensiccontexts in Africa. The immature stages of most of thesespecies are already described, sometimes repeatedly.The larva ofC. croceipalpiswas described and illustratedby Prins (1982), but he provided only a brief descriptionof the Þrst instar and a superÞcial Þgure of its pseudo-cephalon, and Erzinclioglu (1987a) later provided a de-tailed description of the cephaloskeleton and spinula-tion. The morphology of the Þrst instar of C. vicina wasrecently revised (Szpila et al. 2008). Good descriptionsand illustrations of the Þrst instars ofCh. albiceps andCh.megacephala have been published by Carvalho Queirozet al. (1997), Fan et al. (1997), and Sukontason et al.(2003). Ekanem and Umoetuk (2009) provided verysuperÞcial Þgures of the cephaloskeleton of the Þrstinstar ofH. fernandica.The morphologies of the larvae ofL. cuprina and L. sericata are also well known from theworks of Schumann (1954), Erzinclioglu (1989), andOÕFlynn and Moorhouse (1980). The recent reestablish-ment of Ch. chloropyga and Ch. putoria as separate spe-cies(RognesandPaterson2005,Harveyetal. 2008)givesstimulus for further investigation of the characters oftheir larvae to clarify which species were used in thedescriptions published when the two were treated assynonyms (Greenberg and Szyska 1984, Erzinclioglu1989, Ekanem and Usua 2000). Although Prins (1982)described the Þnal instar of Ch. marginalis under thenameCh. regalisRobineau-Desvoidy, 1830 its Þrst instarhave yet to be described.
The aim of the present article is to provide a usefulkey to the Þrst instars of African blow ßies that arecommonly of forensic, medical and veterinary impor-tance. To this end, descriptions of the morphologies ofthe Þrst instar ofC. croceipalpis,Ch. chloropyga andCh.putoria are thoroughly revised or provided for the Þrsttime for Ch. marginalis. We also show that even asimple technique such as light microscopy, when usedproÞciently, can reveal taxonomically useful charac-ters in the morphology of the early preimaginal stages.An identiÞcation key to the majority of the commoncarrion-breeding blow ßies is provided that is based onfeatures that should be easily observable even inpoorly equipped laboratories. The key should be usedwith caution in tropical African countries where ad-ditional species of necrophagous blow ßies [e.g.,Hemipyrellia pulchra (Wiedemann, 1830), Lucilia in-fernalis (Villeneuve 1914), or Chrysomya inclinataWalker, 1861) may be expected.
Materials and Methods
First instars of C. croceipalpis, Ch. chloropyga, Ch.marginalis, andCh. putoriawere reared from eggs laidon chicken liver by females freshly collected in Gra-hamstown (33� 17� S, 26� 29� E), South Africa. Larvaewere killed by soaking in hot (�95�C) distilled waterto extend the body and avoid subsequent deformationwhen stored in 80% ethanol (Adams and Hall 2003).Ten larvae of each species were slide-mounted inHoyerÕs medium for light microscopy. A Nikon 8400digital camera mounted on an Eclipse E200 micro-scope (Nikon, Tokyo, Japan) was used for photomi-crography. Another 10 larvae of each species wereprepared for scanning electron microscopy (SEM) bydehydration through 80, 90, and 99.5% ethanol; criticalpoint drying in CO2; and sputter-coating with plati-num. SEM pictures were taken using a JEOL JSM-6335F SEM (JEOL, Tokyo, Japan). All larvae of eachexamined species were the offspring of a single female.
Details of the morphology of Þrst instars of otherspecies that occur in Africa were gathered from thefollowing sources: C. vicina, Szpila et al. (2008); C.vomitoria, Erzinclioglu (1985, 1987a); Ch. albiceps,Lopes (1983), Carvalho Queiroz et al. (1997), Florezand Wolff (2009), and Mendonca et al. (2010); Ch.megacephala, Fan et al. (1997) and Florez and Wolff(2009); L. sericata, Schumann (1954), Hall (1948),Kano and Sato (1952), Erzinclioglu (1989), Liu andGreenberg (1989), and Fan et al. (1997); and L. cu-prina,Kano and Sato (1952), OÕFlynn and Moorhouse(1980), Greenberg and Szyska (1984), Erzinclioglu(1989), and Fan et al. (1997). These data were veriÞedby observation of new material made available foranalysis through the courtesy of Prof. Tarek Tantawi(Alexandria University, Alexandria, Egypt), Profs.Kabkaew L. Sukontason and Kom Sukontason(Chiang Mai University, Chiang Mai, Thailand), andDr. Soccoro Gomez Dorante (COMEXA, Chiapa deCorzo, Chiapas, Mexico). Unfortunately, morpholog-ical information provided by Ekanem and Umoetuk(2009) for the Þrst instar of H. fernandica is insufÞ-ciently detailed to include in the present key.
Morphological terminology follows Courtney et al.(2000) and Szpila et al. (2008). The term “ectostomalsclerite” used in the past is treated as a basal part of themouthhook (see discussion).
Results
Descriptions of First Instar of C. croceipalpis,Ch. chloropyga, Ch. marginalis, and Ch. putoria
The Þrst instars of C. croceipalpis, Ch. chloropyga,Ch. marginalis, and Ch. putoria possess the generalpattern characteristic for the Calyptratae in beingdivided into a bilobed pseudocephalon (termed pc),three thoracic segments (t1Ðt3), seven abdominal seg-ments (a1Ða7), and an anal division (ad) carrying theposterior spiracles (Figs. 1A; 2A; 4A; 6A; and 8AÐD).Each abdominal segment a1Ða7 has a transverse crev-ice on the ventral surface (Fig. 5A) and lateral creep-
July 2011 SZPILA AND VILLET: MORPHOLOGY OF FIRST INSTARS OF AFRICAN BLOW FLIES 739
ing welts (often termed intersegmental insertions)(Fig. 8A).
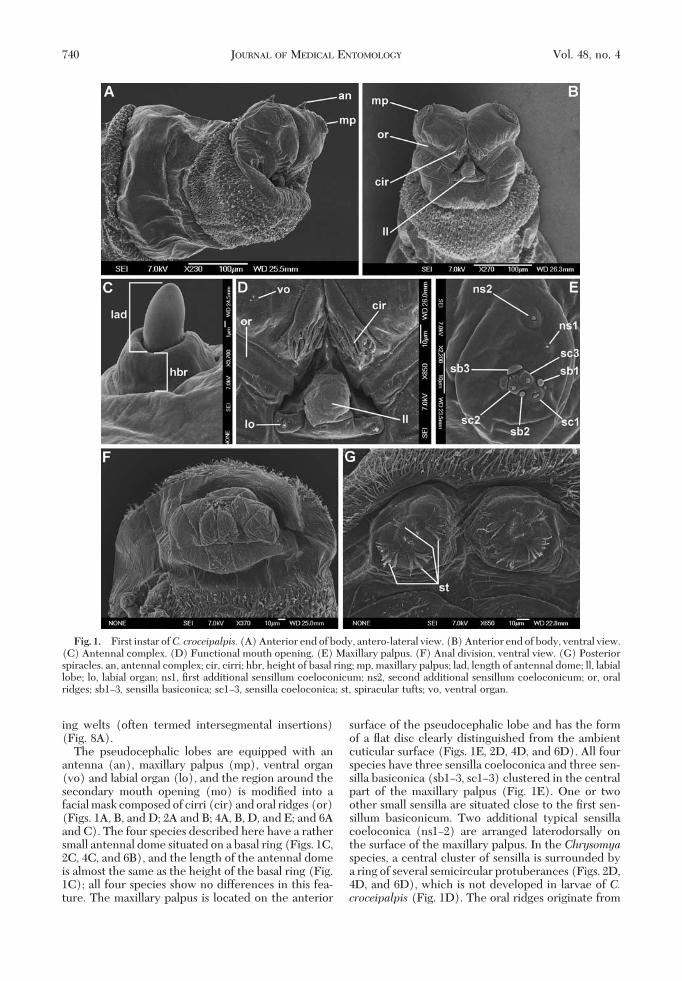
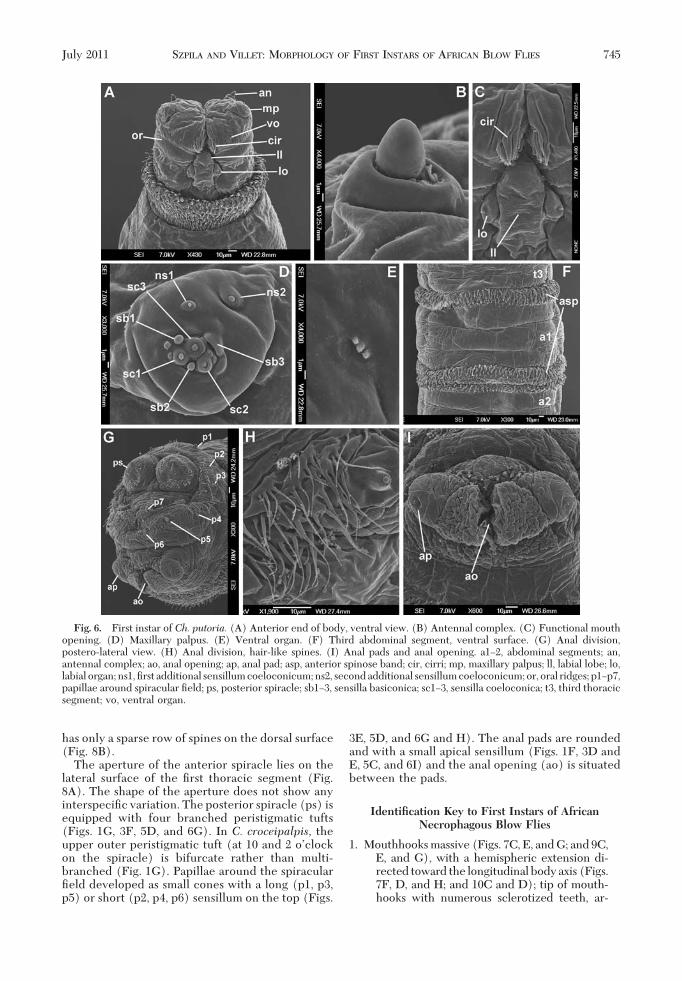
The pseudocephalic lobes are equipped with anantenna (an), maxillary palpus (mp), ventral organ(vo) and labial organ (lo), and the region around thesecondary mouth opening (mo) is modiÞed into afacial mask composed of cirri (cir) and oral ridges (or)(Figs. 1A, B, and D; 2A and B; 4A, B, D, and E; and 6Aand C). The four species described here have a rathersmall antennal dome situated on a basal ring (Figs. 1C,2C, 4C, and 6B), and the length of the antennal domeis almost the same as the height of the basal ring (Fig.1C); all four species show no differences in this fea-ture. The maxillary palpus is located on the anterior
surface of the pseudocephalic lobe and has the formof a ßat disc clearly distinguished from the ambientcuticular surface (Figs. 1E, 2D, 4D, and 6D). All fourspecies have three sensilla coeloconica and three sen-silla basiconica (sb1Ð3, sc1Ð3) clustered in the centralpart of the maxillary palpus (Fig. 1E). One or twoother small sensilla are situated close to the Þrst sen-sillum basiconicum. Two additional typical sensillacoeloconica (ns1Ð2) are arranged laterodorsally onthe surface of the maxillary palpus. In the Chrysomyaspecies, a central cluster of sensilla is surrounded bya ring of several semicircular protuberances (Figs. 2D,4D, and 6D), which is not developed in larvae of C.croceipalpis (Fig. 1D). The oral ridges originate from
Fig. 1. First instar ofC. croceipalpis. (A) Anterior end of body, antero-lateral view. (B) Anterior end of body, ventral view.(C) Antennal complex. (D) Functional mouth opening. (E) Maxillary palpus. (F) Anal division, ventral view. (G) Posteriorspiracles. an, antennal complex; cir, cirri; hbr, height of basal ring; mp, maxillary palpus; lad, length of antennal dome; ll, labiallobe; lo, labial organ; ns1, Þrst additional sensillum coeloconicum; ns2, second additional sensillum coeloconicum; or, oralridges; sb1Ð3, sensilla basiconica; sc1Ð3, sensilla coeloconica; st, spiracular tufts; vo, ventral organ.
740 JOURNAL OF MEDICAL ENTOMOLOGY Vol. 48, no. 4
the lateral parts of the functional mouth opening andrun perpendicular to the longitudinal axis of the bodytoward the lateral parts of the pseudocephalon (Figs.1A, 2A, 4A and B, and 6A). Laterally on the upper (oranterior)partof the functionalmouthopeningare twoclusters or tufts of ßeshy, cuticular outgrowths calledcirri, which hide the apical sclerotized teeth of themouthhooks situated more posteriorly in the func-tional mouth opening (Figs. 1D, 2B, 4E, and 6C).Because of the opaqueness of tissue to the SEM beam,differences in the degree of sclerotization of the cirriand the teeth of the mouthhooks are impossible todiscern on SEM images, but under the light micro-scope the more strongly sclerotized teeth of themouthhooks are clearly differentiated from the trans-parent cirri (Figs. 9AÐH and 10AÐD). The functional
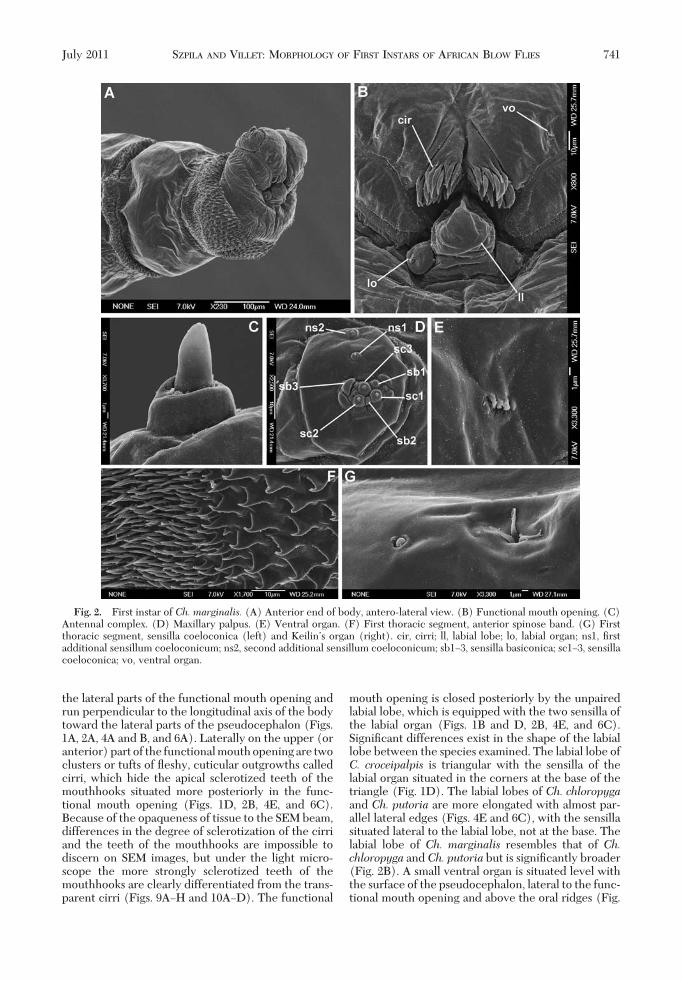
mouth opening is closed posteriorly by the unpairedlabial lobe, which is equipped with the two sensilla ofthe labial organ (Figs. 1B and D, 2B, 4E, and 6C).SigniÞcant differences exist in the shape of the labiallobe between the species examined. The labial lobe ofC. croceipalpis is triangular with the sensilla of thelabial organ situated in the corners at the base of thetriangle (Fig. 1D). The labial lobes of Ch. chloropygaand Ch. putoria are more elongated with almost par-allel lateral edges (Figs. 4E and 6C), with the sensillasituated lateral to the labial lobe, not at the base. Thelabial lobe of Ch. marginalis resembles that of Ch.chloropyga and Ch. putoria but is signiÞcantly broader(Fig. 2B). A small ventral organ is situated level withthe surface of the pseudocephalon, lateral to the func-tional mouth opening and above the oral ridges (Fig.
Fig. 2. First instar of Ch. marginalis. (A) Anterior end of body, antero-lateral view. (B) Functional mouth opening. (C)Antennal complex. (D) Maxillary palpus. (E) Ventral organ. (F) First thoracic segment, anterior spinose band. (G) Firstthoracic segment, sensilla coeloconica (left) and KeilinÕs organ (right). cir, cirri; ll, labial lobe; lo, labial organ; ns1, Þrstadditional sensillum coeloconicum; ns2, second additional sensillum coeloconicum; sb1Ð3, sensilla basiconica; sc1Ð3, sensillacoeloconica; vo, ventral organ.
July 2011 SZPILA AND VILLET: MORPHOLOGY OF FIRST INSTARS OF AFRICAN BLOW FLIES 741
1D and 2B). This structure does not show any inter-speciÞc variation.
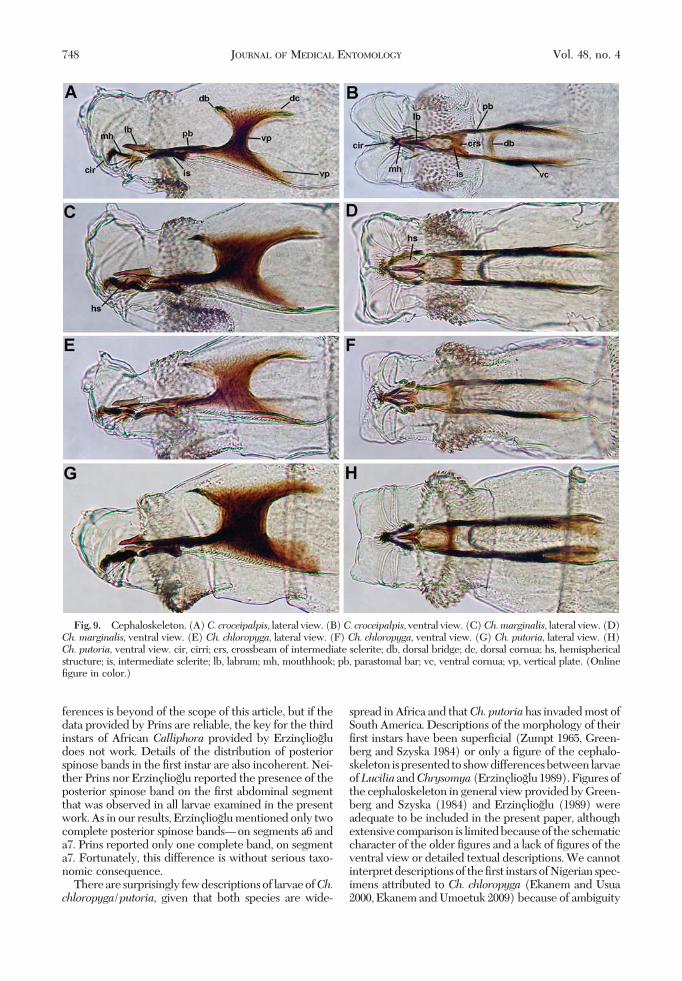
The cephaloskeleton of the Þrst instars of the fourspecies follows the general pattern found in othernecrophagous blow ßies. The cephaloskeleton con-sists of an unpaired labrum (lb), paired mouthhooks(mh), an unpaired intermediate sclerite (is) and abasal sclerite with parastomal bars (pb), vertical plates(vp) and ventral and dorsal cornua (vc, dc) (Figs.7AÐH and 9AÐH). The mouthhooks are the most an-
terior structures of the cephaloskeleton. Each mouth-hook is shaped likea straightbar,with the tipequippedwith various numbers of sharp teeth directed ven-trally. The weakly sclerotized basal part of the mouth-hooks of C. croceipalpis is equipped with a lateral arm(“ectostomal sclerite,” “chitinized teeth”). The ante-rior part is slender and has four or Þve sclerotizedteeth at the tip, arranged in a single row, sometimeswith one additional tooth in front of this row. Themouthhooks of the Chrysomya species are more ro-
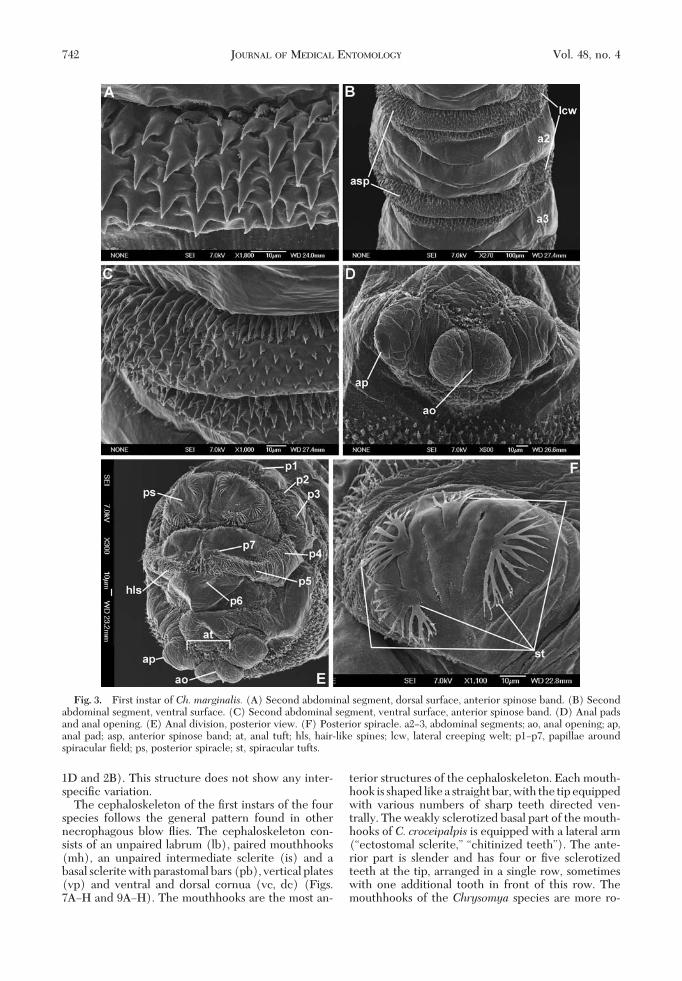
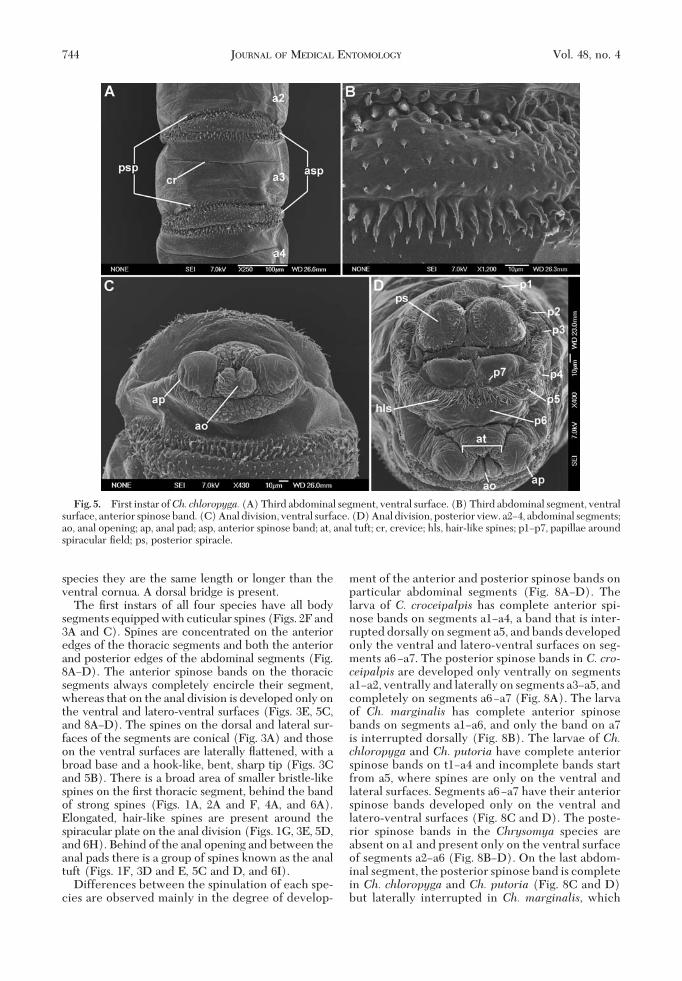
Fig. 3. First instar of Ch. marginalis. (A) Second abdominal segment, dorsal surface, anterior spinose band. (B) Secondabdominal segment, ventral surface. (C) Second abdominal segment, ventral surface, anterior spinose band. (D) Anal padsand anal opening. (E) Anal division, posterior view. (F) Posterior spiracle. a2Ð3, abdominal segments; ao, anal opening; ap,anal pad; asp, anterior spinose band; at, anal tuft; hls, hair-like spines; lcw, lateral creeping welt; p1Ðp7, papillae aroundspiracular Þeld; ps, posterior spiracle; st, spiracular tufts.
742 JOURNAL OF MEDICAL ENTOMOLOGY Vol. 48, no. 4
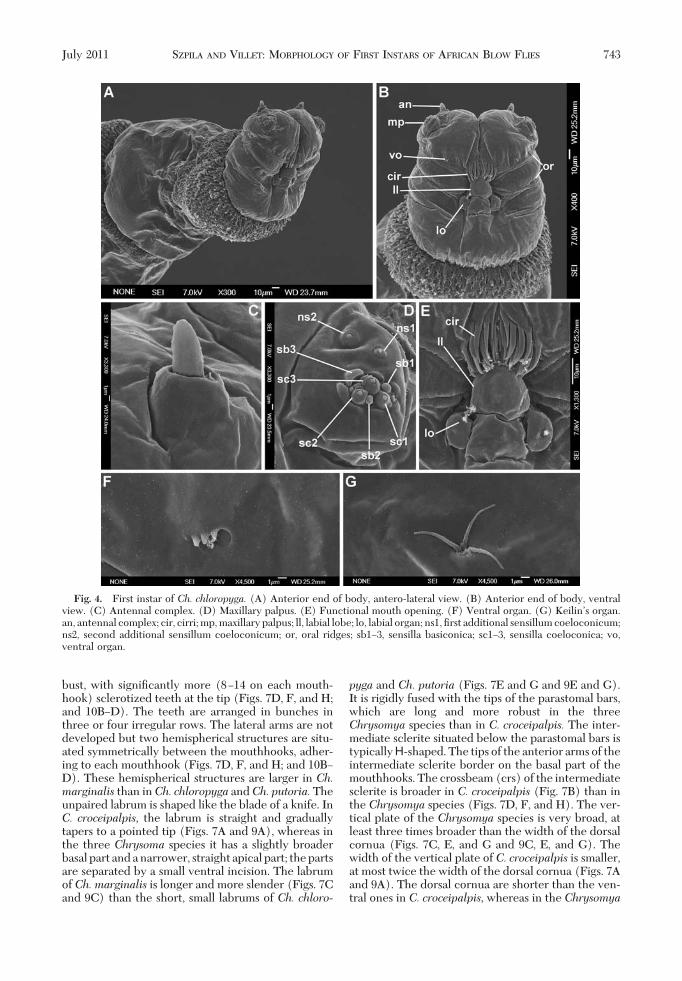
bust, with signiÞcantly more (8Ð14 on each mouth-hook) sclerotized teeth at the tip (Figs. 7D, F, and H;and 10BÐD). The teeth are arranged in bunches inthree or four irregular rows. The lateral arms are notdeveloped but two hemispherical structures are situ-ated symmetrically between the mouthhooks, adher-ing to each mouthhook (Figs. 7D, F, and H; and 10BÐD). These hemispherical structures are larger in Ch.marginalis than inCh. chloropyga andCh. putoria.Theunpaired labrum is shaped like the blade of a knife. InC. croceipalpis, the labrum is straight and graduallytapers to a pointed tip (Figs. 7A and 9A), whereas inthe three Chrysoma species it has a slightly broaderbasal part and a narrower, straight apical part; the partsare separated by a small ventral incision. The labrumof Ch. marginalis is longer and more slender (Figs. 7Cand 9C) than the short, small labrums of Ch. chloro-
pyga and Ch. putoria (Figs. 7E and G and 9E and G).It is rigidly fused with the tips of the parastomal bars,which are long and more robust in the threeChrysomya species than in C. croceipalpis. The inter-mediate sclerite situated below the parastomal bars istypically H-shaped. The tips of the anterior arms of theintermediate sclerite border on the basal part of themouthhooks. The crossbeam (crs) of the intermediatesclerite is broader in C. croceipalpis (Fig. 7B) than inthe Chrysomya species (Figs. 7D, F, and H). The ver-tical plate of the Chrysomya species is very broad, atleast three times broader than the width of the dorsalcornua (Figs. 7C, E, and G and 9C, E, and G). Thewidth of the vertical plate of C. croceipalpis is smaller,at most twice the width of the dorsal cornua (Figs. 7Aand 9A). The dorsal cornua are shorter than the ven-tral ones in C. croceipalpis, whereas in the Chrysomya
Fig. 4. First instar of Ch. chloropyga. (A) Anterior end of body, antero-lateral view. (B) Anterior end of body, ventralview. (C) Antennal complex. (D) Maxillary palpus. (E) Functional mouth opening. (F) Ventral organ. (G) KeilinÕs organ.an, antennal complex; cir, cirri; mp, maxillary palpus; ll, labial lobe; lo, labial organ; ns1, Þrst additional sensillum coeloconicum;ns2, second additional sensillum coeloconicum; or, oral ridges; sb1Ð3, sensilla basiconica; sc1Ð3, sensilla coeloconica; vo,ventral organ.
July 2011 SZPILA AND VILLET: MORPHOLOGY OF FIRST INSTARS OF AFRICAN BLOW FLIES 743
species they are the same length or longer than theventral cornua. A dorsal bridge is present.
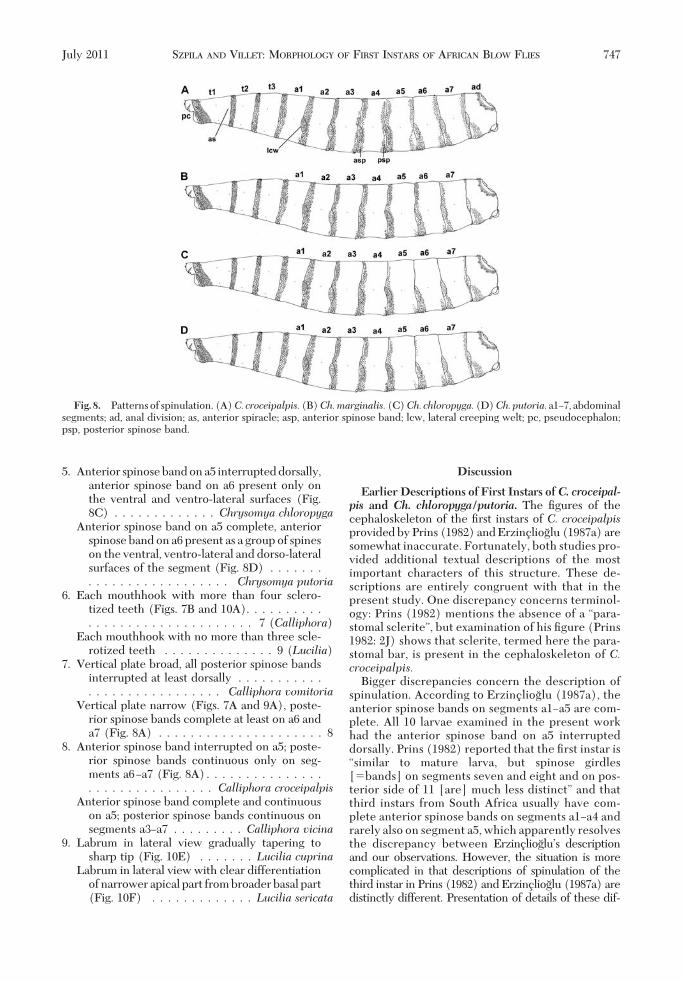
The Þrst instars of all four species have all bodysegments equipped with cuticular spines (Figs. 2F and3A and C). Spines are concentrated on the anterioredges of the thoracic segments and both the anteriorand posterior edges of the abdominal segments (Fig.8AÐD). The anterior spinose bands on the thoracicsegments always completely encircle their segment,whereas that on the anal division is developed only onthe ventral and latero-ventral surfaces (Figs. 3E, 5C,and 8AÐD). The spines on the dorsal and lateral sur-faces of the segments are conical (Fig. 3A) and thoseon the ventral surfaces are laterally ßattened, with abroad base and a hook-like, bent, sharp tip (Figs. 3Cand 5B). There is a broad area of smaller bristle-likespines on the Þrst thoracic segment, behind the bandof strong spines (Figs. 1A, 2A and F, 4A, and 6A).Elongated, hair-like spines are present around thespiracular plate on the anal division (Figs. 1G, 3E, 5D,and 6H). Behind of the anal opening and between theanal pads there is a group of spines known as the analtuft (Figs. 1F, 3D and E, 5C and D, and 6I).
Differences between the spinulation of each spe-cies are observed mainly in the degree of develop-
ment of the anterior and posterior spinose bands onparticular abdominal segments (Fig. 8AÐD). Thelarva of C. croceipalpis has complete anterior spi-nose bands on segments a1Ða4, a band that is inter-rupted dorsally on segment a5, and bands developedonly the ventral and latero-ventral surfaces on seg-ments a6Ða7. The posterior spinose bands in C. cro-ceipalpis are developed only ventrally on segmentsa1Ða2, ventrally and laterally on segments a3Ða5, andcompletely on segments a6Ða7 (Fig. 8A). The larvaof Ch. marginalis has complete anterior spinosebands on segments a1Ða6, and only the band on a7is interrupted dorsally (Fig. 8B). The larvae of Ch.chloropyga and Ch. putoria have complete anteriorspinose bands on t1Ða4 and incomplete bands startfrom a5, where spines are only on the ventral andlateral surfaces. Segments a6Ða7 have their anteriorspinose bands developed only on the ventral andlatero-ventral surfaces (Fig. 8C and D). The poste-rior spinose bands in the Chrysomya species areabsent on a1 and present only on the ventral surfaceof segments a2Ða6 (Fig. 8BÐD). On the last abdom-inal segment, the posterior spinose band is completein Ch. chloropyga and Ch. putoria (Fig. 8C and D)but laterally interrupted in Ch. marginalis, which
Fig. 5. First instar ofCh. chloropyga. (A) Third abdominal segment, ventral surface. (B) Third abdominal segment, ventralsurface, anterior spinose band. (C) Anal division, ventral surface. (D) Anal division, posterior view. a2Ð4, abdominal segments;ao, anal opening; ap, anal pad; asp, anterior spinose band; at, anal tuft; cr, crevice; hls, hair-like spines; p1Ðp7, papillae aroundspiracular Þeld; ps, posterior spiracle.
744 JOURNAL OF MEDICAL ENTOMOLOGY Vol. 48, no. 4
has only a sparse row of spines on the dorsal surface(Fig. 8B).
The aperture of the anterior spiracle lies on thelateral surface of the Þrst thoracic segment (Fig.8A). The shape of the aperture does not show anyinterspeciÞc variation. The posterior spiracle (ps) isequipped with four branched peristigmatic tufts(Figs. 1G, 3F, 5D, and 6G). In C. croceipalpis, theupper outer peristigmatic tuft (at 10 and 2 oÕclockon the spiracle) is bifurcate rather than multi-branched (Fig. 1G). Papillae around the spiracularÞeld developed as small cones with a long (p1, p3,p5) or short (p2, p4, p6) sensillum on the top (Figs.
3E, 5D, and 6G and H). The anal pads are roundedand with a small apical sensillum (Figs. 1F, 3D andE, 5C, and 6I) and the anal opening (ao) is situatedbetween the pads.
Identification Key to First Instars of AfricanNecrophagous Blow Flies
1. Mouthhooks massive (Figs. 7C, E, and G; and 9C,E, and G), with a hemispheric extension di-rected toward the longitudinal body axis (Figs.7F, D, and H; and 10C and D); tip of mouth-hooks with numerous sclerotized teeth, ar-
Fig. 6. First instar of Ch. putoria. (A) Anterior end of body, ventral view. (B) Antennal complex. (C) Functional mouthopening. (D) Maxillary palpus. (E) Ventral organ. (F) Third abdominal segment, ventral surface. (G) Anal division,postero-lateral view. (H) Anal division, hair-like spines. (I) Anal pads and anal opening. a1Ð2, abdominal segments; an,antennal complex; ao, anal opening; ap, anal pad; asp, anterior spinose band; cir, cirri; mp, maxillary palpus; ll, labial lobe; lo,labial organ;ns1,Þrst additional sensillumcoeloconicum;ns2, secondadditional sensillumcoeloconicum;or, oral ridges; p1Ðp7,papillae around spiracular Þeld; ps, posterior spiracle; sb1Ð3, sensilla basiconica; sc1Ð3, sensilla coeloconica; t3, third thoracicsegment; vo, ventral organ.
July 2011 SZPILA AND VILLET: MORPHOLOGY OF FIRST INSTARS OF AFRICAN BLOW FLIES 745

ranged in bunches consisting of two to fourirregular rows; vertical plate of basal scleritevery broad, at least three times broader thanwidth of dorsal cornua. Posterior spinose bandon a4Ða6 developed only on ventral surface ofsegments (Fig. 8B, C, and D) . . . . . . . . . .. . . . . . . . . . . . . . . . . . . . 2 (Chrysomya)
Mouthhooks slender, with a weakly sclerotizedbasal part and clearly visible lateral arm; tip ofmouthhooks with a few sclerotized teeth, ar-ranged in one row, sometimes with one scle-rotized tooth situated in front of the row; ver-tical plate narrow, at least twice as broad aswidth of dorsal cornua (Figs. 7A and B, 9A andB, and 10A). Posterior spinose band on a6developed at least on ventral and dorso-lateralsurfaces (Fig. 8A) . . . . . . . . . . . . . . . . . 6
2. Basal part of mouthhook much broader than tip.Anterior spinose bands on a3Ða4 interrupteddorsally; posterior spinose band on a7 presentonly as a single row of spines on the ventralsurfaces of the segment . . . . . . . . . . . . . .. . . . . . . . . . . . . . . . . . Chrysomya albiceps
Mouthhook almost the same width along itswhole length (Figs. 7C, E, and G; and 9C, E,and G). Anterior spinose bands on a3-a4 com-plete, continuous (Fig. 8BÐD); posterior spi-nose band on a7 complete or with spines onlyon the dorsal and ventral surfaces . . . . . . . 3
3. Labrum of cephaloskeleton long and slender(Figs. 7C and 9C). Anterior spinose band on a6complete, continuous; posterior spinose bandon a7 incomplete on the lateral surfaces (Fig.8B) . . . . . . . . . . . . . Chrysomya marginalis
Labrum of cephaloskeleton short and small(Figs. 7E and G and 9E and G). Anterior spi-nose band on a6 incomplete, interrupted atleast dorsally; posterior spinose band on a7complete (Fig. 8C and D) . . . . . . . . . . . . 4
4. Apical part of labrum narrow, clearly differenti-ated from broad basal part as on Fig. 10F. . .. . . . . . . . . . . . . . . Chrysomya megacephala
Apical part of labrum broad, poorly differenti-ated from broad basal part (Figs. 7E and G and9E and G) . . . . . . . . . . . . . . . . . . . . . . 5
Fig. 7. Cephaloskeleton. (A) C. croceipalpis, lateral view. (B) C. croceipalpis, anterior part, ventral view. (C) Ch. marginalis,lateralview.(D)Ch.marginalis,anteriorpart,ventralview.(E)Ch.chloropyga, lateralview.(F)Ch.chloropyga,anteriorpart,ventralview. (G) Ch. putoria, lateral view. (H) Ch. putoria, anterior part, ventral view. am, anterior part of mouthhook; bm, basal part ofmouthhook; cir, cirri; crs, crossbeam of intermediate sclerite; db, dorsal bridge; dc, dorsal cornua; hs, hemispherical structure; is,intermediate sclerite; la, lateral arm; lb, labrum; mh, mouthhook; pb, parastomal bar; th, teeth; vc, ventral cornua; vp, vertical plate.
746 JOURNAL OF MEDICAL ENTOMOLOGY Vol. 48, no. 4
5. Anterior spinose band on a5 interrupted dorsally,anterior spinose band on a6 present only onthe ventral and ventro-lateral surfaces (Fig.8C) . . . . . . . . . . . . . Chrysomya chloropyga
Anterior spinose band on a5 complete, anteriorspinose band on a6 present as a group of spineson the ventral, ventro-lateral and dorso-lateralsurfaces of the segment (Fig. 8D) . . . . . . .. . . . . . . . . . . . . . . . . . Chrysomya putoria
6. Each mouthhook with more than four sclero-tized teeth (Figs. 7B and 10A). . . . . . . . . .. . . . . . . . . . . . . . . . . . . . . 7 (Calliphora)
Each mouthhook with no more than three scle-rotized teeth . . . . . . . . . . . . . . 9 (Lucilia)
7. Vertical plate broad, all posterior spinose bandsinterrupted at least dorsally . . . . . . . . . . .. . . . . . . . . . . . . . . . . Calliphora vomitoria
Vertical plate narrow (Figs. 7A and 9A), poste-rior spinose bands complete at least on a6 anda7 (Fig. 8A) . . . . . . . . . . . . . . . . . . . . . 8
8. Anterior spinose band interrupted on a5; poste-rior spinose bands continuous only on seg-ments a6Ða7 (Fig. 8A). . . . . . . . . . . . . . .. . . . . . . . . . . . . . . . Calliphora croceipalpis
Anterior spinose band complete and continuouson a5; posterior spinose bands continuous onsegments a3Ða7 . . . . . . . . . Calliphora vicina
9. Labrum in lateral view gradually tapering tosharp tip (Fig. 10E) . . . . . . . Lucilia cuprina
Labrum in lateral view with clear differentiationof narrower apical part from broader basal part(Fig. 10F) . . . . . . . . . . . . . Lucilia sericata
Discussion
Earlier Descriptions of First Instars of C. croceipal-pis and Ch. chloropyga/putoria. The Þgures of thecephaloskeleton of the Þrst instars of C. croceipalpisprovided by Prins (1982) and Erzinclioglu (1987a) aresomewhat inaccurate. Fortunately, both studies pro-vided additional textual descriptions of the mostimportant characters of this structure. These de-scriptions are entirely congruent with that in thepresent study. One discrepancy concerns terminol-ogy: Prins (1982) mentions the absence of a “para-stomal sclerite”, but examination of his Þgure (Prins1982: 2J) shows that sclerite, termed here the para-stomal bar, is present in the cephaloskeleton of C.croceipalpis.
Bigger discrepancies concern the description ofspinulation. According to Erzinclioglu (1987a), theanterior spinose bands on segments a1Ða5 are com-plete. All 10 larvae examined in the present workhad the anterior spinose band on a5 interrupteddorsally. Prins (1982) reported that the Þrst instar is“similar to mature larva, but spinose girdles[�bands] on segments seven and eight and on pos-terior side of 11 [are] much less distinct” and thatthird instars from South Africa usually have com-plete anterior spinose bands on segments a1Ða4 andrarely also on segment a5, which apparently resolvesthe discrepancy between ErzincliogluÕs descriptionand our observations. However, the situation is morecomplicated in that descriptions of spinulation of thethird instar in Prins (1982) and Erzinclioglu (1987a) aredistinctly different. Presentation of details of these dif-
Fig. 8. Patternsof spinulation. (A)C. croceipalpis.(B)Ch.marginalis.(C)Ch. chloropyga.(D)Ch.putoria.a1Ð7, abdominalsegments; ad, anal division; as, anterior spiracle; asp, anterior spinose band; lcw, lateral creeping welt; pc, pseudocephalon;psp, posterior spinose band.
July 2011 SZPILA AND VILLET: MORPHOLOGY OF FIRST INSTARS OF AFRICAN BLOW FLIES 747
ferences is beyond of the scope of this article, but if thedata provided by Prins are reliable, the key for the thirdinstars of African Calliphora provided by Erzincliogludoes not work. Details of the distribution of posteriorspinose bands in the Þrst instar are also incoherent. Nei-ther Prins nor Erzinclioglu reported the presence of theposterior spinose band on the Þrst abdominal segmentthat was observed in all larvae examined in the presentwork. As in our results, Erzinclioglu mentioned only twocomplete posterior spinose bandsÑon segments a6 anda7. Prins reported only one complete band, on segmenta7. Fortunately, this difference is without serious taxo-nomic consequence.
Therearesurprisingly fewdescriptionsof larvaeofCh.chloropyga/putoria, given that both species are wide-
spread in Africa and thatCh. putoriahas invaded most ofSouth America. Descriptions of the morphology of theirÞrst instars have been superÞcial (Zumpt 1965, Green-berg and Szyska 1984) or only a Þgure of the cephalo-skeletonispresentedtoshowdifferencesbetweenlarvaeofLucilia andChrysomya(Erzinclioglu 1989). Figures ofthe cephaloskeleton in general view provided by Green-berg and Szyska (1984) and Erzinclioglu (1989) wereadequate to be included in the present paper, althoughextensivecomparison is limitedbecauseof theschematiccharacter of the older Þgures and a lack of Þgures of theventral view or detailed textual descriptions. We cannotinterpretdescriptionsof theÞrst instarsofNigerianspec-imens attributed to Ch. chloropyga (Ekanem and Usua2000, Ekanem and Umoetuk 2009) because of ambiguity
Fig. 9. Cephaloskeleton. (A)C. croceipalpis, lateral view. (B)C. croceipalpis,ventral view. (C)Ch.marginalis, lateral view. (D)Ch. marginalis, ventral view. (E) Ch. chloropyga, lateral view. (F) Ch. chloropyga, ventral view. (G) Ch. putoria, lateral view. (H)Ch. putoria, ventral view. cir, cirri; crs, crossbeam of intermediate sclerite; db, dorsal bridge; dc, dorsal cornua; hs, hemisphericalstructure; is, intermediate sclerite; lb, labrum; mh, mouthhook; pb, parastomal bar; vc, ventral cornua; vp, vertical plate. (OnlineÞgure in color.)
748 JOURNAL OF MEDICAL ENTOMOLOGY Vol. 48, no. 4
in the descriptions, Þgures, and identiÞcation. The ma-terial was identiÞed in 1992 (M. S. Ekanem, personalcommunication) during a period when Ch. chloropygaand Ch. putoriawere treated as synonyms (Rognes andPaterson 2005), and the voucher material was severelydamaged in the mail and not preserved (N. Wyatt, per-sonalcommunication).Regarding theÞrst instarof(pre-sumably)Ch. chloropyga,Zumpt (1965) mentioned onlyanteriorspinosebands(“elevenbandsofminutespines”)and wrote that the bands on a4Ða7 and the anal divisionwere well developed only on the ventral surface withonly a few spines on the dorsal surface. Unfortu-nately, he did not state which bands are complete orinterrupted. For Ch. putoria, Greenberg and Szyska
(1984) only mentioned that the “spine pattern [is]similar to third instar.” They reported complete an-terior spinose bands on segments a1Ða3 and some-times also on a4 of the third instar, which does notexactly correspond with our observations. Again, theposterior spinose bands are treated together with theanterior spinose bands as the “intersegmental spinessurrounding posterior segments.” Unsurprisingly(given the prevailing contemporary taxonomic styleand confusion about the distinctness of the taxa), theinformation provided by Zumpt (1965) and Green-berg and Szyska (1984) is generally insufÞciently de-tailed for differentiation of the Þrst instars of Ch.chloropyga and Ch. putoria.
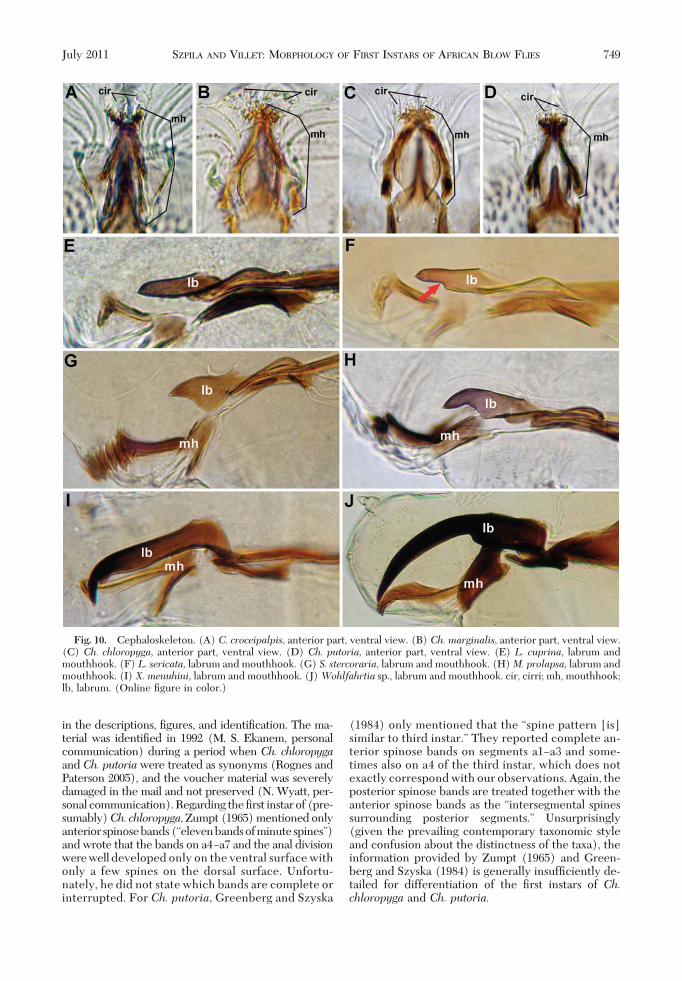
Fig. 10. Cephaloskeleton. (A) C. croceipalpis, anterior part, ventral view. (B) Ch. marginalis, anterior part, ventral view.(C) Ch. chloropyga, anterior part, ventral view. (D) Ch. putoria, anterior part, ventral view. (E) L. cuprina, labrum andmouthhook. (F) L. sericata, labrum and mouthhook. (G) S. stercoraria, labrum and mouthhook. (H)M. prolapsa, labrum andmouthhook. (I) X. menuhini, labrum and mouthhook. (J)Wohlfahrtia sp., labrum and mouthhook. cir, cirri; mh, mouthhook;lb, labrum. (Online Þgure in color.)
July 2011 SZPILA AND VILLET: MORPHOLOGY OF FIRST INSTARS OF AFRICAN BLOW FLIES 749

Taxonomically Important Features in Identifica-tion of the First Instars of Necrophagous Blow Flies.IdentiÞcation of the early instars of necrophagousblow ßies is difÞcult because larvae are very homo-geneous in morphology even at the level of subfamily.This creates the additional problem that relatively fewmorphological characters of larvae are available tosystematics and phylogenetics. Rognes (1997) deÞned43 phylogenetically informative characters in the mor-phology of adult blow ßies, whereas Szpila and Pape(2008) found only 21 phylogenetically signiÞcantcharacters of Þrst instars in the Miltogramminae,which are recognized as having very diverse larvalmorphology (Szpila 2010). The number of usefullyvariable characters in the Þrst instars of necrophagousCalliphoridae is reasonably low. The combination ofavailable features is restricted mainly to small varia-tions in the morphology of the cephaloskeleton andthe distribution of spines (Schumann 1954, OÕFlynnand Moorhouse 1980, Liu and Greenberg 1989,Erzinclioglu 1987a, 1989). Detailed analysis of accu-mulated data from previous studies (Schumann 1954;OÕFlynn and Moorhouse 1980; Liu and Greenberg1989; Erzinclioglu 1987a, 1989) and ongoing work ofthe Þrst author on a broad spectrum of larval materialhas allowed the deÞnition of an expanded set of usefulmorphological characters of Þrst instars for systemat-ics and phylogenetics. There are as follows.Mouthhooks of Cephaloskeleton. Thorough observa-
tion of the shape of the mouthhooks is difÞcult be-cause of the weak sclerotization of some of their el-ements. In the opinion of K.S., the true shape of themouthhook should encompass not only its apical teeth[discussed in Szpila et al. (2008))] but also the“ectostomal sclerite” of Schumann (1954) and the“chitinized teeth” ofErzinclioglu(1985).Comparativeanalyses of the additional taxa suggest that these smallarticles are probably the last sclerotized remains of thebasal part of the mouthhooks. Whole mouthhooks arestill entirely sclerotized in some species of Scatophagi-dae [e.g., Scathophaga stercoraria (L., 1758)], Musci-dae [e.g., Muscina prolapsa (Harris, 1780)] and Sar-cophagidae (e.g., Xiphidiella menuhini Lehrer, 1993;Wohlfahrtia spp.) and their complete shape is easilyobservable (Fig. 10GÐJ). The “chitinized teeth” of theÞrst instars of necrophagous Calliphoridae are homol-ogous to the lateral arms of the mouthhook describedin Miltogramminae (Szpila 2010), and they are anintegral part of the mouthhook. Differences in thelevel of sclerotization of the mouthhooks between Þrstinstars of particular species have taxonomic value. TheÞrst instar of Holarctic species Phormia regina (Mei-gen, 1826) is easy recognizable by the transparentanterior part of its mouthhooks (Hall 1948, Szpila et al.2008). Species of Chrysomya (Chrysomyinae) havethe basal part of their mouthhooks strongly sclerotized(but with reduced sclerotization of the lateral arm), incontrast with larvae of Calliphora and Lucilia (Calli-phorinae, Luciliinae). The number of sclerotized api-cal teeth on the mouthhook also has great taxonomicimportance. This character was almost entirely ne-glected in earlier studies (e.g., Hall 1948, Schumann
1954, OÕFlynn and Moorhouse 1980, Erzinclioglu 1985,Liu and Greenberg 1989), but the ventral view of theapical part of the mouthhooks reveals large differ-ences between species and genera in the number andarrangement of their teeth (Szpila et al. 2008; also seethe key provided here). The hemispherical structuressituated between the mouthhooks of Chrysomya alsoseem to be a good character for separating the Þrstinstars of this genus from those of other genera.Labrum of Cephaloskeleton. In most species ofChrysomya the labrum is small in relation to the sizeof the basal sclerite, with simultaneous developmentof strong mouthhooks. IntraspeciÞc variation is pres-ent in the shape of the labrum. In some species, thelabrum gradually tapers to a sharp tip in lateral view,whereas in others differentiation of a narrower apicalpart and a broader basal part is clear. Species with bothof these forms of labrum may be present in the samegenus (e.g., Lucilia and Chrysomya).Intermediate Sclerite of Cephaloskeleton. Like the
mouthhook, the shape of the intermediate sclerite isdifÞcult to observe because of its weak sclerotization.However, at least the width of the crossbeam in ven-tral view may be useful in separating larvae of speciesof Chrysomya from those of Lucilia and Calliphora.The thickness of the intermediate sclerite in lateralview (Erzinclioglu 1985, 1987a) is not very reliablebecause the parallax effect of even slight tilting of aslide-mounted specimen may signiÞcantly change theapparent dimensions of this sclerite.Basal Sclerite of Cephaloskeleton. The large width of
the vertical plate in relation to other elements of thecephaloskeleton was used to differentiate Þrst instars ofChrysomyinae from those of other blow ßies (Liu andGreenberg 1989). The width of this structure also wassuggested by Erzinclioglu (1987a) to separate C. vomi-toria from other species of Calliphora in Africa, but thedifference in width is not as obvious as it is betweenChrysomyinae and species representing other subfami-lies. Szpila et al. (2008) reported the presence of thesclerotized transverse bridge between the posterior tipsof the ventral cornua in Þrst instars of C. vicina and P.regina. This structure is more widespread in larvae ofCalliphora species, but not in other Chrysomyinae ex-cept Protophormia terraenovae (Robineau-Desvoidy,1830).Thesuggestion that thedegreeof sclerotizationofthe dorsal bridge could differentiate the Þrst instars ofL.cuprina and L. sericata (Erzinclioglu 1989) is contra-dicted by the results presented here.Spinose Bands on Abdominal Segments. The distri-
bution of the spinose bands, and their fragmentationand reduction on particular segments, are very usefultaxonomic characters that have been used in almost allprevious keys (Schumann 1954, OÕFlynn and Moor-house 1980, Prins 1982, Erzinclioglu 1987a, Liu andGreenberg 1989). Use of these features is complicatedby discrepancies between various authors in their de-scriptions of spinulation (Szpila et al. 2008, also seediscussion above). Some interspeciÞc variation doesoccur, but the question is whether it represents con-sistent differences or only artifacts of differing meth-ods of preparation and precision of observation.
750 JOURNAL OF MEDICAL ENTOMOLOGY Vol. 48, no. 4

Spinulation is deÞnitely a good diagnostic featurebut demands the proper methods of preparation ofmaterial. Killing in hot water (Adams and Hall2003), storage in 70Ð80% ethanol and using mediacontaining chloral hydrate (like HoyerÕs medium orBerlese ßuid) for preparation of microscope slidesseems to be best.Anal Division. In contrast to that of third instars
(Schumann 1954, Prins 1982; Erzinclioglu 1985, Wall-man 2001, Florez and Wolff 2009, Szpila 2010), the analdivision of Þrst instars very rarely bears taxonomicallyuseful characters. Even using SEM techniques prob-ably cannot change this situation. Currently, the shapeof the anal pads successfully separates only Þrst instarsof Ch. albiceps and Ch. rufifacies (in which they pro-trude strongly) from other necrophagous blow ßies.Erzinclioglu (1987b) suggested using the shape anddensity of hair-like spines around the spiracular Þeldto differentiate Þrst instars of Calliphora and Luciliabut recently published SEM pictures (Szpila et al.2008) do not show any signiÞcant difference in thischaracter between C. vicina and L. illustris.However,the density of hair-like spines seems to be reliable foridentiÞcation at least of some species of Calliphora(Liu and Greenberg 1989). The posterior spiracles ofthe Þrst instars of necrophagous blow ßies are uselessfor taxonomic purposes because they are very homo-geneous. Only the shape of the spiracular tufts (ob-servable only using SEM) gives some hope for Þndingrelevant differences between species. However, thesimpliÞed form of one spiracular tuft mentioned ascharacteristic of the genus Lucilia is problematic be-cause a similar reduction of this structure occurs inlarvae of C. croceipalpis (Fig. 1G).
In conclusion, a variety of adult ßies are found onthe carrion of large vertebrates in Africa (e.g., Braack1986, 1987), but the maggot community is dominatedby blow ßies, which are therefore most commonly offorensic signiÞcance. The descriptions and identiÞca-tion key provided here represent a consolidation andextension of taxonomic information about these mag-gots, and an important contribution to the develop-ment of forensic entomology in Africa. Because someof the species have already been introduced intoSouth America, and because blow ßies are being trans-ported synanthropically around the world, this workhas potentially much broader geographical applica-bility.
The Þrst instars of necrophagous blow ßies havelong been regarded as taxonomically challenging toidentify, but this review of the African fauna showsthat the impediment is not a lack of distinguishingcharacters, but the small size of the specimens. Ourkey has been designed to be easy for staff of forensicand quarantine laboratories to use successfully. It iscertain that further research on the carrion blow ßiesof Africa will lengthen the list of forensically relevantspecies present on this continent. As details of thelarvae of these species are discovered, they will beincorporated into future keys.
Acknowledgments
We are very grateful to Thomas Pape (Natural HistoryMuseum of Denmark, Zoological Museum) for providingaccess to excellent SEM equipment and to Mfon Ekanem(Ahmadu Bello University, Nigeria) and Nigel Wyatt (Nat-ural History Museum, London, United Kingdom) for clari-fying the taxonomic status of some specimens. This work wasÞnancially supported by the Polish Ministry of Science andHigher Education (grant N N303 470838) and Rhodes Uni-versity.
References Cited
Adams, Z.J.O., and M.J.R. Hall. 2003. Methods used for thekilling and preservation of blowßy larvae, and their effecton post-mortem larval length. Forensic Sci. Int. 138:50Ð61.
Amendt, J., R. Zehner, D. G. Johnson, and J. Wells. 2010.Future trends in forensic entomology, pp. 353Ð368. InJ. Amendt, M. L. Goff, C. P. Campobasso, and M. Grass-berger (eds.), Current concepts in forensic entomology.Springer, Dordrecht, The Netherlands.
Braack,L. 1986. Arthropods associated with carcasses in thenorthern Kruger National Park. S. Afr. J. Wildl. Res. 16:91Ð98.
Braack, L. 1987. Community dynamics of carrion-attendantarthropods in tropical African woodland. Oecologia 72:402Ð409.
CarvalhoQueiroz,M.M., R. Pinto deMello, andM.M.Lima.1997. Morphological aspects of the larval instars ofChrysomya albiceps (Diptera, Calliphoridae) reared inthe laboratory. Mem. Inst. Oswaldo Cruz 92: 187Ð196.
Courtney, G. W., B. J. Sinclair, and R. Meier. 2000. Mor-phology and terminology of Diptera larvae, pp. 85Ð161. InL. Papp and B. Darvas (eds.), Contributions to a manualof Palaearctic Diptera (with special reference to ßies ofeconomic importance). Science Herald Press, Budapest,Hungary.
Ekanem,M. S., andM. C.Dike. 2010. Arthropod successionon pig carcasses in southeastern Nigeria. Pap. AvulsosZool. 50: 561Ð570.
Ekanem,M. S., and S. Umoetuk. 2009. The immature stagesof three carrion breeding blowßies (Diptera: Calliphori-dae) in South Eastern Nigeria. Zoologist 7: 152Ð161.
Ekanem, M. S., and E. J. Usua. 2000. Immature stages andbiology of to blowßy species (Diptera: Calliphoridae) inAkwa Ibom state, Nigeria. Nig. J. Entomol. 17: 1Ð11.
Erzinclioglu, Y. Z. 1985. Immature stages of British Calli-phora and Cynomya,with re-evaluation of the taxonomiccharacters of larval Calliphoridae (Diptera). J. Nat. Hist.19: 69Ð96.
Erzinclioglu, Y. Z. 1987a. The larval instars of the Africanblowßy, Calliphora croceipalpis Jaennicke, with a key tothe genera of the third instars of African carrion-breedingCalliphoridae (Diptera). Bull. Entomol. Res. 77: 575Ð580.
Erzinclioglu, Y. Z. 1987b. Recognition of the early instarlarvae of the genera Calliphora and Lucilia (Dipt., Cal-liphoridae). Entomol. Mon. Mag. 123: 97Ð98.
Erzinclioglu, Y. Z. 1989. The early larval instars of Luciliasericata andLucilia cuprina (Diptera: Calliphoridae): my-iasis blowßies of Africa and Australia. J. Nat. Hist. 23:1133Ð1136.
Fan, Z., C. Zhizi, F. Jianming, Z. Shensheng, and T. Zhen-liang. 1997. Diptera: Calliphoridae. Fauna Sinica, In-secta, 6: x � 1Ð707.
Florez, E., and M. Wolff. 2009. Descripcion y clave de losestadios inmaduros de las principales especies de Calli-
July 2011 SZPILA AND VILLET: MORPHOLOGY OF FIRST INSTARS OF AFRICAN BLOW FLIES 751

phoridae de importancia forense en Colombia. Neotrop.Entomol. 38: 418Ð429.
Greenberg, B., and M. Szyska. 1984. Immature stages andbiology of Þfteen species of Peruvian Calliphoridae (Dip-tera). Ann. Entomol. Soc. Am. 77: 488Ð517.
Hall, D. G. 1948. The blowßies of North America. TheThomas Say Foundation, Baltimore, MD.
Harvey, M. L., S. Gaudieri, M. H. Villet, and I. R. Dadour.2008. A global study of forensically signiÞcant calli-phorids: implications for identiÞcation. Forensic Sci. Int.177: 66Ð76.
Kano, R., and K. Sato. 1952. Notes on the ßies of medicalimportance in Japan. (Part VI) Larvae of Luciliini inJapan. Jpn. J. Exp. Med. 22: 33Ð42.
Kurahashi, H., andA. Kirk-Spriggs. 2006. The Calliphoridaeof Namibia (Diptera: Oestroidea). Zootaxa 1322: 1Ð131.
Liu, D., and B. Greenberg. 1989. Immature stages of someßies of forensic importance. Ann. Entomol. Soc. Am. 82:80Ð93.
Lopes, H. S. 1983. The importance of the mandible andclypeal arch of the Þrst instar larvae in the classiÞcationof the Sarcophagidae (Diptera). Rev. Bras. Entomol. 26:293Ð326.
Louw, S. M., and T. C. van der Linde. 1993. Insects fre-quenting decomposing corpses in central South Africa.Afr. Entomol. 1: 265Ð269.
Mendonca,P.M., J.R.DosSantos-Mallet, andM.M.CarvalhoQueiroz. 2010. Ultramorphological characteristics ofimmature stages of Chrysomya albiceps (Wiedemann1819) (Diptera: Calliphoridae), a ßy species of forensicimportance. Mircosc. Res. Tech. 73: 779Ð784.
O’Flynn, M. A., and D. E. Moorhouse. 1980. IdentiÞcationof early immature stages of some common Queenslandcarrion ßies. J. Aust. Entomol. Soc. 19: 53Ð61.
Prins, A. J. 1982. Morphological and biological notes on sixSouth African blow-ßies (Diptera, Calliphoridae) andtheir immature stages. Ann. S. Afr. Mus. 90: 201Ð217.
Pont, A. 1980. 90. Family Calliphoridae, pp. 779Ð800. InR. W. Crosskey (ed.), Catalogue of the Diptera of theAfrotropical Region. British Museum (Natural History),London, United Kingdom.
Rognes, K. 1997. The Calliphoridae (blowßies) (Diptera:Oestroidea) are not a monophyletic group. Cladistics 13:27Ð66.
Rognes, K., and H.E.H. Paterson. 2005. Chrysomya chloro-pyga (Wiedemann, 1818) and C. putoria (Wiedemann,1830) are two different species. Afr. Entomol. 13: 49Ð70.
Schumann, H. 1954. Morphologish-systematische Studienan Larven von hygienisch wichtigen mitteleuropaischenDipteren der Familien CalliphoridaeÐMuscidae. Wiss. Z.Univ. Greifswald Jahrgang III, 1953/54 Mathematisch-naturwissenschaftliche Reihe Nr. 4/5: 245Ð274.
Sukontason, K. L., K. Sukontason, S. Piangjai, N. Boonchu, T.Chaiwong,R.C.Vogtsberger, B.Kuntalue,N.Thijuk, and
J. K. Olson. 2003. Larval morphology of Chrysomyamegacephala (Fabricius) (Diptera: Calliphoridae) usingscanning electron microscopy. J. Vector Ecol. 28: 47Ð52.
Sukontason, K. L., P. Sribanditmongkol, T. Chaiwong, R. C.Vogtsberger, S. Piangjai, and K. Sukontason. 2008. Mor-phology of immature stages of Hemipyrellia ligurriens(Wiedemann) (Diptera: Calliphoridae) for use in foren-sic entomology applications. Parasitol. Res. 103: 877Ð887.
Szpila, K., andT. Pape. 2008. Morphological diversity of Þrstinstar larvae in Miltogramma subgenus Pediasiomyia(Diptera: Sarcophagidae, Miltogramminae). Zool. Anz.247: 259Ð273.
Szpila, K., T. Pape, andA. Rusinek. 2008. Morphology of theÞrst instar larva of Calliphora vicina, Phormia regina, andLucilia illustris (Diptera, Calliphoridae). Med. Vet. En-tomol. 22: 16Ð25.
Szpila, K. 2010. The Þrst instar of European Miltogrammi-nae (Diptera: Sarcophagidae). Wydawnictwo NaukoweUMK, Torun, Poland.
Tantawi, T., E. M. El-Kady, B. Greenberg, and H. A. El-Ghaffar. 1996. Arthropod succession on exposed rabbitcarrion in Alexandria, Egypt. J. Med. Entomol. 33: 566Ð580.
Thyssen, P. J., and A. X. Linhares. 2007. First description ofthe immature stages ofHemilucilia segmentaria (Diptera:Calliphoridae). Biol. Res. 40: 271Ð280.
Verves, Yu., G., and L. A. Khrokalo. 2009. Superfamily Oes-troidea. family Calliphoridae, pp. 251Ð270. In J. Gerlach(ed.), The Diptera of the Seychelles islands. Pensoft,SoÞa, Bulgaria.
Villet, M. H., C. S. Richards, and J. M. Midgley. 2010. Con-temporary precision, bias and accuracy of minimum post-mortem intervals estimated using development of carri-on-feeding insects, pp. 109Ð137. In J. Amendt, M. L. Goff,C. P. Campobasso, and M. Grassberger (eds.), Currentconcepts in forensic entomology. Springer, Dordrecht,The Netherlands.
Wallman, J. F. 2001. Third instar larvae of common carrion-breeding blowßies of the genusCalliphora (Diptera: Cal-liphoridae) in South Australia. Invertebr. Taxon. 15:37Ð51.
Williams, K. A., and M. H. Villet. 2006. A new and earlierrecord of Chrysomya megacephala in South Africa, withnotes on another exotic species, Calliphora vicina (Dip-tera: Calliphoridae). Afr. Invertebr. 47: 347Ð350.
Zumpt,F. 1956. Calliphoridae (Diptera Cyclorrhapha) PartI: Calliphorini and Chrysomyiini. Exploration du ParcNational Abert Mission G. F. De Witte (1933Ð1935), Fas-cicule. 87: 1Ð201.
Zumpt, F. 1965. Myiasis in Man and animals in the OldWorld. A textbook for physicians, veterinarians and zo-ologists. ButterworthÕs, London, United Kingdom.
Received 29 October 2010; accepted 17 February 2011.
752 JOURNAL OF MEDICAL ENTOMOLOGY Vol. 48, no. 4





























