Embed Size (px)
Citation preview

This article was downloaded by: [California Institute of Technology]On: 04 October 2014, At: 04:37Publisher: Taylor & FrancisInforma Ltd Registered in England and Wales Registered Number: 1072954 Registeredoffice: Mortimer House, 37-41 Mortimer Street, London W1T 3JH, UK
New Zealand Journal of Marine andFreshwater ResearchPublication details, including instructions for authors andsubscription information:http://www.tandfonline.com/loi/tnzm20
Morphology and growth characteristicsof Chrysochromulina species(Haptophyceae = Prymnesiophyceae)isolated from New Zealand coastalwatersLesley Rhodes a & Brendon Burke aa Cawthron Institute , Private Bag 2, Nelson, New ZealandPublished online: 30 Mar 2010.
To cite this article: Lesley Rhodes & Brendon Burke (1996) Morphology and growth characteristicsof Chrysochromulina species (Haptophyceae = Prymnesiophyceae) isolated from New Zealandcoastal waters, New Zealand Journal of Marine and Freshwater Research, 30:1, 91-103, DOI:10.1080/00288330.1996.9516699
To link to this article: http://dx.doi.org/10.1080/00288330.1996.9516699
PLEASE SCROLL DOWN FOR ARTICLE
Taylor & Francis makes every effort to ensure the accuracy of all the information (the“Content”) contained in the publications on our platform. However, Taylor & Francis,our agents, and our licensors make no representations or warranties whatsoever as tothe accuracy, completeness, or suitability for any purpose of the Content. Any opinionsand views expressed in this publication are the opinions and views of the authors,and are not the views of or endorsed by Taylor & Francis. The accuracy of the Contentshould not be relied upon and should be independently verified with primary sourcesof information. Taylor and Francis shall not be liable for any losses, actions, claims,proceedings, demands, costs, expenses, damages, and other liabilities whatsoeveror howsoever caused arising directly or indirectly in connection with, in relation to orarising out of the use of the Content.
This article may be used for research, teaching, and private study purposes. Anysubstantial or systematic reproduction, redistribution, reselling, loan, sub-licensing,systematic supply, or distribution in any form to anyone is expressly forbidden. Terms &

Conditions of access and use can be found at http://www.tandfonline.com/page/terms-and-conditions
Dow
nloa
ded
by [
Cal
ifor
nia
Inst
itute
of
Tec
hnol
ogy]
at 0
4:37
04
Oct
ober
201
4

New Zealand Journal of Marine and Freshwater Research, 1996: Vol. 30: 91-1030028-8330/96/3001-0091 $2.50/0 © The Royal Society of New Zealand 1996
91
Morphology and growth characteristics of Chrysochromulinaspecies (Haptophyceae = Prymnesiophyceae) isolated fromNew Zealand coastal waters
LESLEY RHODESBRENDON BURKE
Cawthron InstitutePrivate Bag 2Nelson, New Zealand
Abstract Chrysochromulina acantha, C. hirta,C. pachycylindra, and C. cf. polylepis (Hapto-phyceae = Prymnesiophyceae) are new records,based on transmission electron microscopy, forNew Zealand coastal waters. The morphology andgrowth characteristics of C. acantha and C. hirta,and of C. simplex and C. ericina (also New Zealandisolates), are described. C. ericina of the presentstudy has a shorter haptonema (12-24 urn; non-coiling), larger plate and spine scales, and fewerspine scales (with broader bases), than northernhemisphere isolates. Scales of C. simplex fit theemended type description, whereas the large spinescales of C. hirta are shorter ( 11 µm) than thetype description (20-30 µm). C. ericina and C. hirtafall into a temperate (15-20°C), and C. acanthaand C. simplex into a sub-tropical (20-25°C),temperature group. All species tested grew well inbrackish salinities (c. 23 through to 36); C. hirtagrew optimally at a salinity of 23. All tolerated 25-150 umol m-2 s-1 photon flux and pH of 7.5 to c.8.4. C. acantha had significantly faster growthrates (doublings d-1) with nitrate than with urea orammonium chloride as nitrogen source, and C. hirtawith urea and ammonium chloride rather than withnitrate; none required selenium. None of the speciestested was toxic to Artemia salina. C. ericina andC. acantha were phagotrophic. Calcofluor assistedin the identification of C. quadrikonta from otherspine-bearing species and FITC-conjugated wheatgerm lectin aided differentiation of the Chryso-chromulina species from Prymnesium parvum andP. patelliferum.
Keywords Chrysochromulina; Haptophyceae;Prymnesiophyceae; growth; electron microscopy
M95043Received 3 July 1995; accepted 16 October 1995
INTRODUCTION
The genus Chrysochromulina Lackey (Hapto-phyceae = Prymnesiophyceae) has had littleattention in New Zealand waters since the recordingof 13 species, identified by transmission electronmicroscopy of scales, in 1974 (Moestrup 1979). Anew species, C. novae-zelandiae (Moestrup), wasrecorded at that time. Since then, blooms of bothC. pringsheimii (Parke & Manton) (Stewart Island,November 1990, C.O'Kelly, Bigelow Laboratoryfor Ocean Sciences, ME, USA, pers. comm.) andthe quadriflagellate species C. quadrikonta(Kawachi & Inouye) (Hauraki Gulf, May 1994:6.0 x 104 cells I"1 and June 1995: 3.5 x 105 cellsI"1) have been observed (unpubl. data). C. quad-rikonta has been shown to produce low levels of ahaemolysin in culture (Rhodes 1994).
The present study describes the taxonomy andgrowth characteristics of C. acantha (Leadbeater& Manton), C. ericina (Parke & Manton), C. hirta(Manton), and C. simplex (Estep, Davis, Hargraves& Sieburth), all isolated from New Zealand coastalwaters, and compares them with the previouslydescribed New Zealand isolates, C. quadrikontaand C. camella (Leadbeater & Manton) (Rhodes etal. 1993). It also records the presence of C. pachy-cylindra (Manton, Oates & Course) and C. cf.polylepis (Manton & Parke) in New Zealand waters,as identified by electron microscopy.
The contention that haptophyte species whichcan not be differentiated by light microscopicobservation under standard conditions can beidentified with the aid of fluorescent probes istested. Calcofluor White M2R is a fluorescent dyethat binds to cellulose and other P-linked glucans(Hughs & McCully 1975). The scales of Chryso-
Dow
nloa
ded
by [
Cal
ifor
nia
Inst
itute
of
Tec
hnol
ogy]
at 0
4:37
04
Oct
ober
201
4
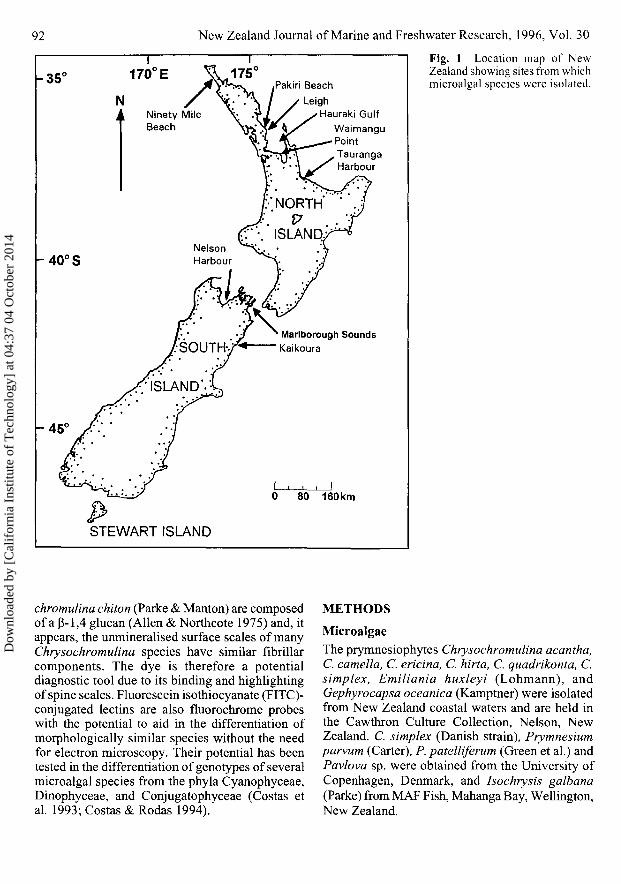
92 New Zealand Journal of Marine and Freshwater Research, 1996, Vol. 30
-35°
- A(\°
170°E
NPakiri Beach
LeighHauraki Gulf
WaitnanguPointTaurangaHarbour
40° S
Marlborough SoundsKaikoura
80 160km
STEWART ISLAND
Fig. 1 Location map of NewZealand showing sites from whichmicroalgal species were isolated.
chromulina chiton (Parke & Manton) are composedof a (3-1,4 glucan (Allen & Northcote 1975) and, itappears, the unmineralised surface scales of manyChrysochromulina species have similar fibrillarcomponents. The dye is therefore a potentialdiagnostic tool due to its binding and highlightingof spine scales. Fluorescein isothiocyanate (FITC)-conjugated lectins are also fluorochrome probeswith the potential to aid in the differentiation ofmorphologically similar species without the needfor electron microscopy. Their potential has beentested in the differentiation of genotypes of severalmicroalgal species from the phyla Cyanophyceae,Dinophyceae, and Conjugatophyceae (Costas etal. 1993; Costas & Rodas 1994).
METHODS
MicroalgaeThe prymnesiophytes Chrysochromulina acantha,C. camella, C. ericina, C. hirta, C. quadrikonta, C.simplex, Emiliania huxleyi (Lohmann), andGephyrocapsa oceanica (Kamptner) were isolatedfrom New Zealand coastal waters and are held inthe Cawthron Culture Collection, Nelson, NewZealand. C. simplex (Danish strain), Prymnesiumparvum (Carter), P. patelliferum (Green et al.) andPavlova sp. were obtained from the University ofCopenhagen, Denmark, and Isochrysis galbana(Parke) from MAF Fish, Mahanga Bay, Wellington,New Zealand.
Dow
nloa
ded
by [
Cal
ifor
nia
Inst
itute
of
Tec
hnol
ogy]
at 0
4:37
04
Oct
ober
201
4

Rhodes & Burke—Chrysochromulina spp. in New Zealand 93
Isolation and culture of microalgaeHaptophyte isolates were obtained from sea watersamples collected from various sites around NewZealand's coastline (Fig. 1) either by vertical haulwith a 10 urn mesh net or with a water-bottlesampler at various discrete depths (0—25 m). Cellswere isolated and cultured as described previously(Rhodes et al. 1993) in modified CHRY (Andersenet al. 1991) or GP media (Loeblich & Smith 1968)in tissue culture wells (Becton Dickinson).
Changes in chlorophyll a over time, determinedby fluorometry (Rhodes et al. 1993), were used tomonitor growth during the exponential phase.Means of cell and cell organelle measurementswere calculated from a minimum of 50 cells (insettling chambers, observed live with an invertedmicroscope); growth rates (number of doublingsper day) were derived from the means of fourreplicate cultures. ANOVAs were used to determinethe statistical significance of the results, with theTukey-Kramer method applied for comparison ofmeans.
Fluorescent probesCalcofluor White M2R: 1% Calcofluor solution(Sigma Chemical Co.) was added to microalgae onglass slides (final concentration 0.05%).Lectins: Fresh solutions of FITC-conjugated lectins(100 ugmH; Sigma Chemical Co.) were preparedusing filtered (0.4 um Nuclepore), synthetic seawater (Gross 1967), salinity 28, pH 8.0. Microalgalcell cultures were centrifuged(1 14 g; 18min, 15°C)and the lectin (Table 1) added to the microalgalpellet at 1 ml per 1.0 x 105 cells and incubated(15 min, 20°C). Unbound lectin was removed bywashing in synthetic sea water. Fluorescence oftreated cells, mounted on siliconised glass slides,was determined qualitatively by epifluorescence
microscopy and recorded as —, no fluorescence; +,bright fluorescence.
MicroscopyLight and electron microscopy were carried out asdescribed previously (Rhodes et al. 1993).
Microalgae, treated with Calcofluor or FITC-conjugated lectin, were examined with an AHBTNew Vanox Olympus photomicrographic epi-fluorescence microscope using UV (excitation 375nm; emission > 420 nm) and blue light (excitation490nm; emission 520 nm), respectively.
PhagotrophyMicroalgae were cultured in tissue culture plates,fluorescently-labelled glass beads (0.8 um diameter,soaked overnight in bovine serum albumin) added,and uptake observed under UV light.
Toxicity bioassayChrysochromulina species were tested for toxicityusing a modified Artemia salina bioassay (Persoone& Wells 1987; Rhodes et al. 1993).
RESULTS
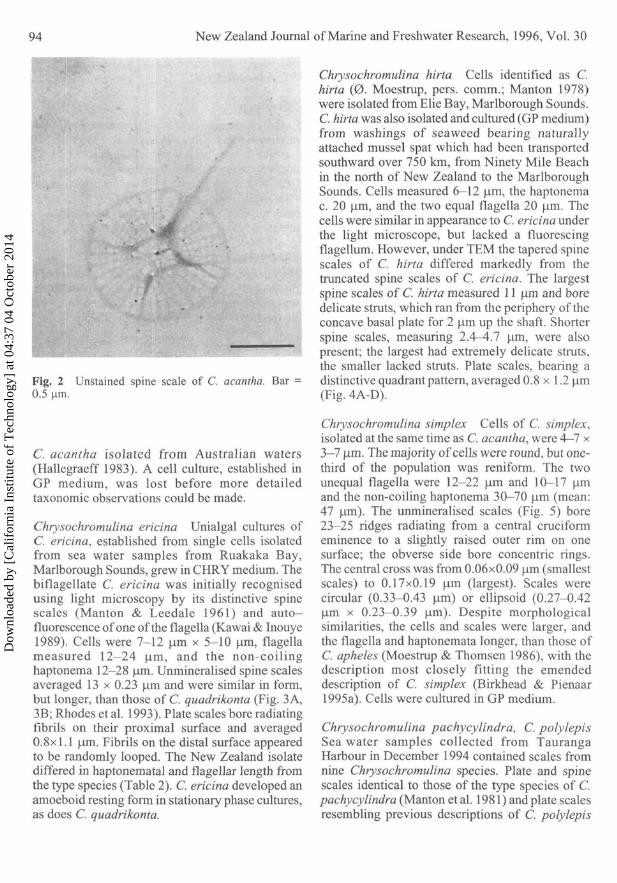
Isolation and identificationChrysochromulina acantha Several Chryso-chromulina species were observed in sea watersamples taken from Pakiri Beach in the HaurakiGulf, February 1994, including C. acantha. Cellsizes were 7—10 um x 5—7 nm. The two equalflagella measured 15 um and the haptonema 20-34 urn. The species was identified, using trans-mission electron microscopy (TEM), by itsdistinctive plate and spine scales (Fig. 2; Leadbeater& Manton 1971); scales fitted the description of
Table 1 FITC-conjugated lectins used as probes.
Name Source Specificity
ConAECAHPAPEAPHAPNAPWMSBAUEAWGA
Canavalia ensiformisErythrina cristagalliHelix pomatiaPisum sativumPhaseolus limensisArachis hypogaeaPhytolacca americanaGlycine maxUlex europaeusTriticum vulgaris
Methyl a-D-mannopyranoside; D-mannose; D-Glucosea-Lactose; N-Acetyl-D-galactosamine; D-GalactoseN-Acetyl-D-glucosamine; N-Acetyl-D-galactosamine; D-GalactoseMethyl a-D-mannopyranoside; D-Mannose; D-GlucoseN-Acetyl-D-galactosaminea-Lactose; D-GalactoseN-Acetyl-D-glucosamineN-Acetyl-D-galactosamine; D-Galactose; Methyl a-D-galactopyranosideL-FucoseN-Triacetylchitotriose; N-Diacetylchitobiose; Sialic acid
Dow
nloa
ded
by [
Cal
ifor
nia
Inst
itute
of
Tec
hnol
ogy]
at 0
4:37
04
Oct
ober
201
4
94 New Zealand Journal of Marine and Freshwater Research, 1996, Vol. 30
Fig. 2 Unstained spine scale of C. acantha. Bar =0.5 urn.
C. acantha isolated from Australian waters(Hallegraeff 1983). A cell culture, established inGP medium, was lost before more detailedtaxonomic observations could be made.
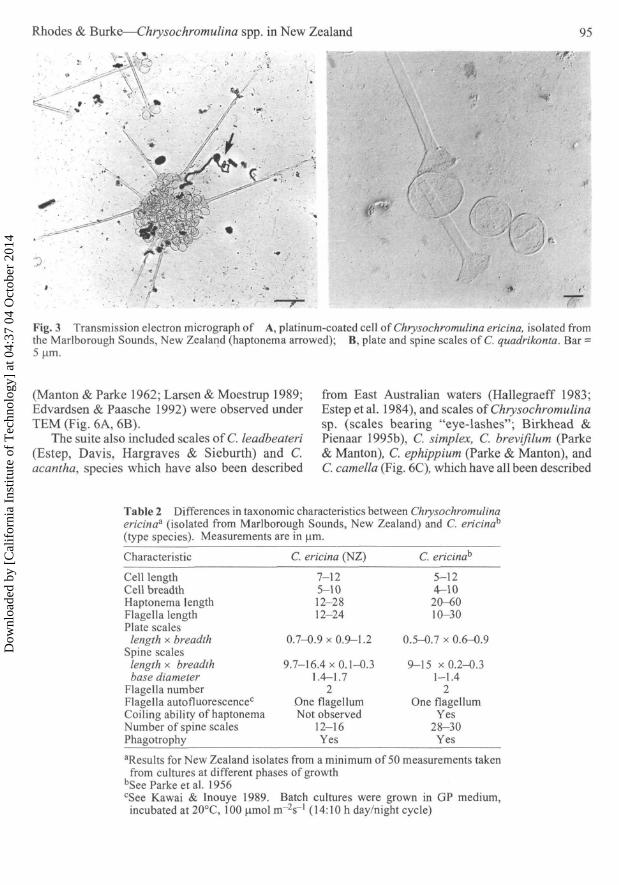
Chrysochromulina ericina Unialgal cultures ofC. ericina, established from single cells isolatedfrom sea water samples from Ruakaka Bay,Marlborough Sounds, grew in CHRY medium. Thebiflagellate C. ericina was initially recognisedusing light microscopy by its distinctive spinescales (Manton & Leedale 1961) and auto-fluorescence of one of the flagella (Kawai & Inouye1989). Cells were 7-12 um x 5-10 urn, flagellameasured 12—24 um, and the non-coilinghaptonema 12-28 urn. Unmineralised spine scalesaveraged 13 x 0.23 um and were similar in form,but longer, than those of C. quadrikonta (Fig. 3A,3B; Rhodes et al. 1993). Plate scales bore radiatingfibrils on their proximal surface and averaged0.8x1.1 um. Fibrils on the distal surface appearedto be randomly looped. The New Zealand isolatediffered in haptonematal and flagellar length fromthe type species (Table 2). C. ericina developed anamoeboid resting form in stationary phase cultures,as does C. quadrikonta.
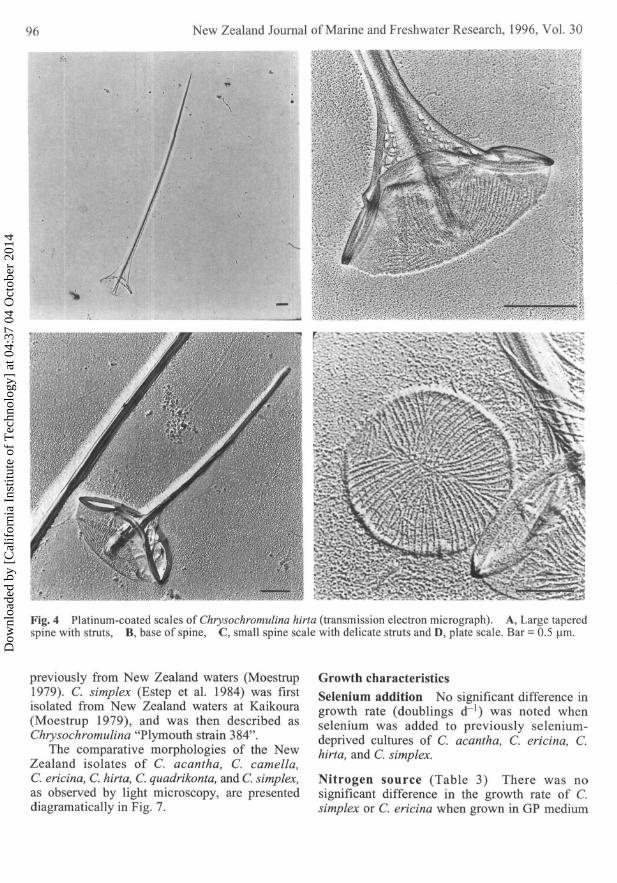
Chrysochromulina hirta Cells identified as C.hirta (0. Moestrup, pers. comm.; Manton 1978)were isolated from Elie Bay, Marlborough Sounds.C. hirta was also isolated and cultured (GP medium)from washings of seaweed bearing naturallyattached mussel spat which had been transportedsouthward over 750 km, from Ninety Mile Beachin the north of New Zealand to the MarlboroughSounds. Cells measured 6—12 um, the haptonemac. 20 um, and the two equal flagella 20 um. Thecells were similar in appearance to C. ericina underthe light microscope, but lacked a fluorescingflagellum. However, under TEM the tapered spinescales of C. hirta differed markedly from thetruncated spine scales of C. ericina. The largestspine scales of C. hirta measured 11 um and boredelicate struts, which ran from the periphery of theconcave basal plate for 2 um up the shaft. Shorterspine scales, measuring 2.4—4.7 um, were alsopresent; the largest had extremely delicate struts,the smaller lacked struts. Plate scales, bearing adistinctive quadrant pattern, averaged 0.8 x 1.2 um(Fig. 4A-D).
Chrysochromulina simplex Cells of C. simplex,isolated at the same time as C. acantha, were 4—7 x3—7 um. The majority of cells were round, but one-third of the population was reniform. The twounequal flagella were 12-22 um and 10-17 umand the non-coiling haptonema 30—70 um (mean:47 um). The unmineralised scales (Fig. 5) bore23—25 ridges radiating from a central cruciformeminence to a slightly raised outer rim on onesurface; the obverse side bore concentric rings.The central cross was from 0.06x0.09 um (smallestscales) to 0.17x0.19 urn (largest). Scales werecircular (0.33-0.43 um) or ellipsoid (0.27-0.42um x 0.23—0.39 um). Despite morphologicalsimilarities, the cells and scales were larger, andthe flagella and haptonemata longer, than those ofC. apheles (Moestrup & Thomsen 1986), with thedescription most closely fitting the emendeddescription of C. simplex (Birkhead & Pienaar1995a). Cells were cultured in GP medium.
Chrysochromulina pachycylindra, C. polylepisSea water samples collected from TaurangaHarbour in December 1994 contained scales fromnine Chrysochromulina species. Plate and spinescales identical to those of the type species of C.pachycylindra (Manton et al. 1981) and plate scalesresembling previous descriptions of C. polylepis
Dow
nloa
ded
by [
Cal
ifor
nia
Inst
itute
of
Tec
hnol
ogy]
at 0
4:37
04
Oct
ober
201
4
Rhodes & Burke—Chrysochromulina spp. in New Zealand
!
•»..
Fig. 3 Transmission electron micrograph of A, platinum-coated cell of Chrysochromulina ericina, isolated fromthe Marlborough Sounds, New Zealand (haptonema arrowed); B, plate and spine scales of C. quadrikonta. Bar =5 \im.
(Manton & Parke 1962; Larsen & Moestrup 1989;Edvardsen & Paasche 1992) were observed underTEM (Fig. 6A, 6B).
The suite also included scales of C. leadbeateri(Estep, Davis, Hargraves & Sieburth) and C.acantha, species which have also been described
from East Australian waters (Hallegraeff 1983;Estep et al. 1984), and scales of Chrysochromulinasp. (scales bearing "eye-lashes"; Birkhead &Pienaar 1995b), C. simplex, C. brevifilum (Parke& Manton), C. ephippium (Parke & Manton), andC. camella (Fig. 6C), which have all been described
Table 2 Differences in taxonomic characteristics between Chrysochromulinaericina1* (isolated from Marlborough Sounds, New Zealand) and C. ericinab
(type species). Measurements are in ^im.
Characteristic
Cell lengthCell breadthHaptonema lengthFlagella lengthPlate scales
length x breadthSpine scales
length x breadthbase diameter
Flagella numberFlagella autofluorescence0
Coiling ability of haptonemaNumber of spine scalesPhagotrophy
C. ericina (NZ)
7-125-1012-2812-24
0.7-0.9x0.9-1.2
9.7-16.4x0.1-0.31.4-1.7
2One flagellumNot observed
12-16Yes
C. ericina^
5-124-1020-6010-30
0.5-0.7 x 0.6-0.9
9-15 x 0.2-0.31-1.4
2One flagellum
Yes28-30Yes
aResults for New Zealand isolates from a minimum of 50 measurements takenfrom cultures at different phases of growth
bSee Parke et al. 1956cSee Kawai & Inouye 1989. Batch cultures were grown in GP medium,incubated at 20°C, 100 nmol r r r V 1 (14:10 h day/night cycle)
95
Dow
nloa
ded
by [
Cal
ifor
nia
Inst
itute
of
Tec
hnol
ogy]
at 0
4:37
04
Oct
ober
201
4
96 New Zealand Journal of Marine and Freshwater Research, 1996, Vol. 30
' ^ ' • • - ' - • ^ ' ' " ' •
gmmmwmm
Fig. 4 Platinum-coated scales of Chrysochromulina hirta (transmission electron micrograph). A, Large taperedspine with struts, B, base of spine, C, small spine scale with delicate struts and D, plate scale. Bar = 0.5 (im.
previously from New Zealand waters (Moestrup1979). C. simplex (Estep et al. 1984) was firstisolated from New Zealand waters at Kaikoura(Moestrup 1979), and was then described asChrysochromulina "Plymouth strain 384".
The comparative morphologies of the NewZealand isolates of C. acantha, C. camella,C. ericina, C. hirta, C. quadrikonta, and C. simplex,as observed by light microscopy, are presenteddiagramatically in Fig. 7.
Growth characteristicsSelenium addition No significant difference ingrowth rate (doublings d~ ') was noted whenselenium was added to previously selenium-deprived cultures of C. acantha, C. ericina, C.hirta, and C. simplex.
Nitrogen source (Table 3) There was nosignificant difference in the growth rate of C.simplex or C. ericina when grown in GP medium
Dow
nloa
ded
by [
Cal
ifor
nia
Inst
itute
of
Tec
hnol
ogy]
at 0
4:37
04
Oct
ober
201
4
Rhodes & Burke—Chrysochwmulina spp. in New Zealand
ffl&k
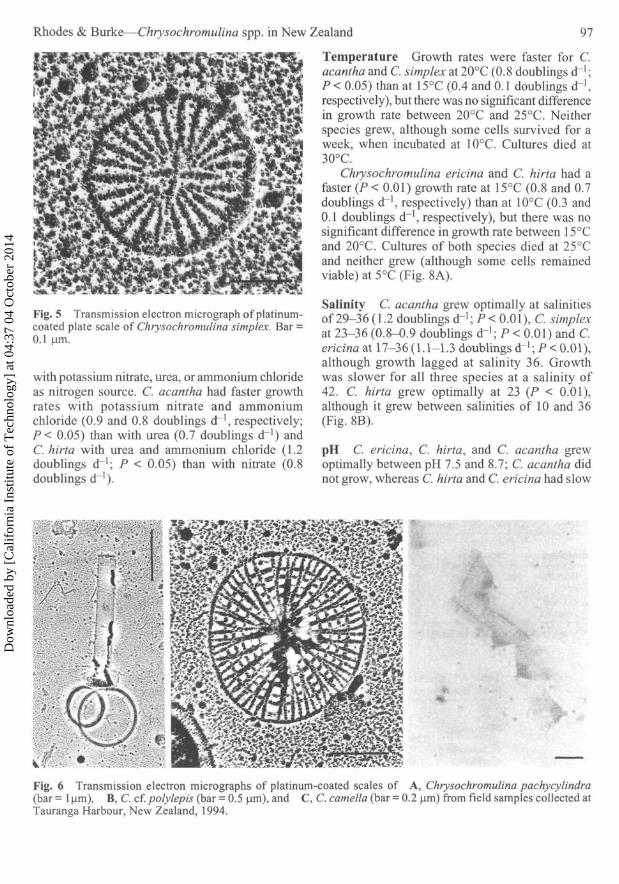
Fig. 5 Transmission electron micrograph of platinum-coated plate scale of Chrysochromulina simplex. Bar =0.1 urn.
with potassium nitrate, urea, or ammonium chlorideas nitrogen source. C. acantha had faster growthrates with potassium nitrate and ammoniumchloride (0.9 and 0.8 doublings d~', respectively;P< 0.05) than with urea (0.7 doublings d"1) andC. hirta with urea and ammonium chloride (1.2doublings d"1; P < 0.05) than with nitrate (0.8doublings d~').
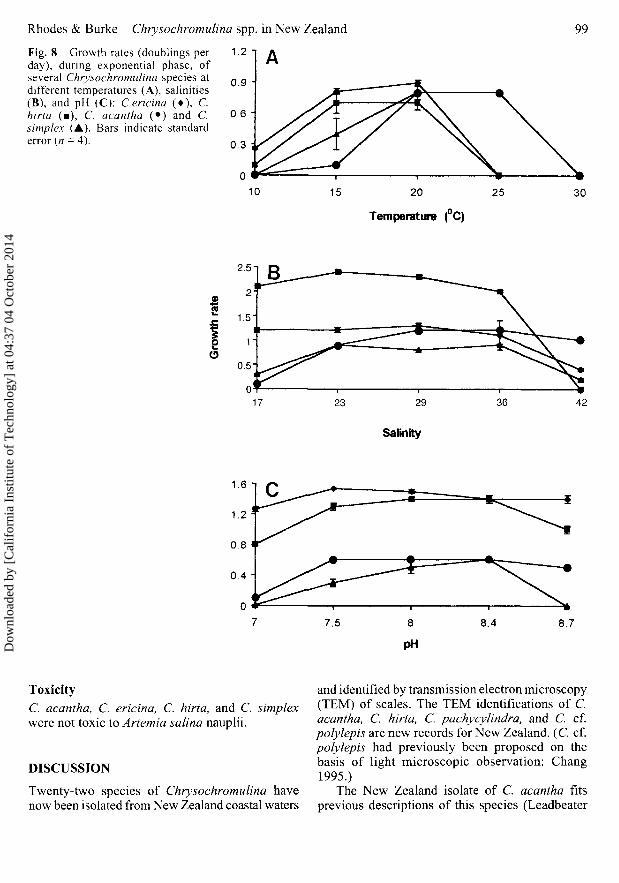
Temperature Growth rates were faster for C.acantha and C. simplex at 20°C (0.8 doublings dr];P< 0.05) than at 15°C (0.4 and 0.1 doublings dr\respectively), but there was no significant differencein growth rate between 20°C and 25°C. Neitherspecies grew, although some cells survived for aweek, when incubated at 10°C. Cultures died at30°C.
Chrysochromulina ericina and C. hirta had afaster (P < 0.01) growth rate at 15°C (0.8 and 0.7doublings d"1, respectively) than at 10°C (0.3 and0.1 doublings cH, respectively), but there was nosignificant difference in growth rate between 15°Cand 20°C. Cultures of both species died at 25°Cand neither grew (although some cells remainedviable) at 5°C (Fig. 8A).
Salinity C. acantha grew optimally at salinitiesof 29-36 (1.2 doublings d"1; P < 0.01), C. simplexat 23-36 (0.8-0.9 doublings d"1; P < 0.01) and C.ericina at 17-36(1.1-1.3 doublings d ~ ] ;P< 0.01),although growth lagged at salinity 36. Growthwas slower for all three species at a salinity of42. C. hirta grew optimally at 23 (P < 0.01),although it grew between salinities of 10 and 36(Fig. 8B).
pH C. ericina, C. hirta, and C. acantha grewoptimally between pH 7.5 and 8.7; C. acantha didnot grow, whereas C. hirta and C. ericina had slow
Fig. 6 Transmission electron micrographs of platinum-coated scales of A, Chrysochromulina pachycylindra(bar = 1 (im), B, C. cf. polylepis (bar = 0.5 |xm), and C, C. camella (bar = 0.2 um) from field samples collected atTauranga Harbour, New Zealand, 1994.
97
Dow
nloa
ded
by [
Cal
ifor
nia
Inst
itute
of
Tec
hnol
ogy]
at 0
4:37
04
Oct
ober
201
4

New Zealand Journal of Marine and Freshwater Research, 1996, Vol. 30
Fig. 7 Diagram illustratingdifferences in cell size andorganelle length, observable underthe light microscope: A,Chrysochromulinaquadrikonta; B, C. ericina; C,C. hirta; D, C. camella; E, C.acantha; and F, C. simplex. Bar= 10 urn.
B
growth rates, at pH 7.0 (P < 0.01). C. simplex grewoptimally at pH 7.5-8.4 (P < 0.01; Fig. 8C).
Light (Table 4) No significant difference ingrowth rate was recorded for C. ericinabetween 25 and 150 umol mr2 s~'. C. acanthaand C. hirta had a slower growth rate at 25 than at50 umol m~2 s"1 and C. acantha at 50 than75 umol m"2 s~x. C, simplex grew faster at> 100 umol m 2s~' .
Table 3 Growth rates (doublings d ' ) of Chryso-chromulina species cultured with different nitrogensources. Results are means of quadruplicates.
MicroalgaPotassium
nitrateAmmonium
chloride Urea
C. acanthaC. hirtaC. ericinaC. simplex
0.9 "0.8"1.10.4
0.8 a
1.21.10.4
0.71.20.90.3
aValues are significantly different (P < 0.05)
Phagotrophy Cells of C. ericina could beobserved, under UV excitation, taking up, and afterc. 20 seconds ejecting, fluorescently-labelled beads.Fluorescent beads were observed within cells of C.acantha, although rarely. Phagotrophy was notobserved in C. simplex nor C. hirta.
Fluorescent probesCalcofluor White: C. ericina, C. hirta, C.quadrikonta, and C. simplex (Danish strain)fluoresced on the addition of Calcofluor; C. simplex(NZ strain) and C. camella did not. The spinescales of C. ericina, C. quadrikonta, and C. hirtawere highlighted and could be clearly observedwith epifluorescence microscopy.
FITC-conjugated lectins: C. simplex (bothstrains), C. ericina, C. hirta, and C. quadrikontabound WGA, but none of the other lectins tested.The fluorescence was clearly observed around thecell body in each case, but no fluorescence wasnoted on the spine scales (Table 5). None of theother haptophytes investigated fluoresced with anyof the lectins tested.
Table 4 Growth rates (doublings d ') of Chrysochromulina species culturedat different photon flux. NT, not tested. Results are means of quadruplicates.
Microalga
C. acanthaC. ericinaC. hirtaC. simplex
25
0.7 a
0.60.70.8"
50
1.0"0.90.71.2
Photon
75
1.21.10.91.3
flux (|imol
100
1.11.0"1.01.4
m-V1)125
1.21.51.01.2
150
1.31.40.91.2
175
1.21.2NTNT
aValues are significantly different (P < 0.05)
98
Dow
nloa
ded
by [
Cal
ifor
nia
Inst
itute
of
Tec
hnol
ogy]
at 0
4:37
04
Oct
ober
201
4
Rhodes & Burke—Chrysochromulina spp. in New Zealand
Fig. 8 Growth rates (doublings perday), during exponential phase, ofseveral Chrysochromulina species atdifferent temperatures (A), salinities(B), and pH (C): C.ericina (•) , C.hirta (•), C. acantha (•) and C.simplex (A). Bars indicate standarderror (n = 4).
99
15 20
Temperature (C)
25 30
17
Salinity
Toxicity
C. acantha, C. ericina, C. hirta, and C. simplexwere not toxic to Anemia salina nauplii.
DISCUSSION
Twenty-two species of Chrysochromulina havenow been isolated from New Zealand coastal waters
and identified by transmission electron microscopy(TEM) of scales. The TEM identifications of C.acantha, C. hirta, C. pachycylindra, and C. cf.polylepis are new records for New Zealand. (C cf.polylepis had previously been proposed on thebasis of light microscopic observation: Chang1995.)
The New Zealand isolate of C. acantha fitsprevious descriptions of this species (Leadbeater
Dow
nloa
ded
by [
Cal
ifor
nia
Inst
itute
of
Tec
hnol
ogy]
at 0
4:37
04
Oct
ober
201
4

100 New Zealand Journal of Marine and Freshwater Research, 1996, Vol. 30
& Manton 1971; Hallegraeff 1983). The scales ofC. simplex have close similarities to those describedfor Chrysochromulina "Plymouth 384", identifiedby TEM of scales from the coastal waters ofKaikoura, New Zealand (Moestrup 1979), and alsoobserved in eastern Australian waters (Hallegraeff1983). The isolate most closely fits the typedescription of C. simplex (Estep et al. 1984) asemended by Birkhead & Pienaar (1995a), ratherthan that of C. apheles, scales of which have alsobeen described from New Zealand waters(Moestrup & Thomsen 1986).
Plate scales of C. ericina (New Zealand isolate)are comparable with previous descriptions fromNew Zealand and Norwegian waters (Moestrup1979; Leadbeater 1972a), but are larger than thetype description (Parke et al. 1956). Spine scalesare longer and fewer in the New Zealand strain,although spine scale number could be related tocell age (Manton & Leedale 1961); variability intrumpet-scale dimensions has been noted previously(Leadbeater 1972b). The haptonema is shorter thanin northern hemisphere isolates and it is also non-coiling. The New Zealand isolate of C. hirta hasmarkedly shorter spine scales than its type species(Manton 1978).
During the present study members of the genusChrysochromulina were most commonly present(although usually in low numbers) in coastal watersduring the early summer and late autumn,presumably when water temperatures and day
Table 5 Specificity of FITC-conjugated wheat germlectin (WGA) and Calcofluor used as fluorescent probes.N, not tested; +, bright fluorescence; —, no fluorescence.
WGA Calcofluor
PrymnesialesC. camella N —C. ericina + +C. hirta + +C. quadrikonta + +C. simplex (NZ isolate) + -C. simplex (Danish isolate) + +Prymnesium parvum — +P. patelliferum — +PavlovalesPavlova sp — +
IsochrysidalesIsochrysis sp - +CoccolithophoralesGephyrocapsa oceanica — —Emiliania huxleyi — —
length were most favourable to growth. C.quadrikonta bloomed in the Firth of Thames,Hauraki Gulf in May 1994, when sea surfacetemperatures (SSTs) were 16.5°C (MAF RegulatoryAuthority operational research phytoplanktonprogramme unpubl. report). In June 1995, SSTswere 13.7°C. Winter SSTs dropped to 12°C andsummer temperatures reached 22°C in 1993; the insitu temperatures recorded at the time C.quadrikonta bloomed closely match the temperaturerange within which C. quadrikonta survives andgrows in vitro.
Despite the different maxima and minima forthe two temperature groups, all would have beenable to survive throughout the year in the north-eastern waters of the North Island and the northernwaters of the South Island. The range of averagemonthly temperatures in the Marlborough Sounds,from which C. ericina, C. camella, and C. hirtahave been isolated, was from 18°C in summer to10°C in winter for 1987-93 (Regal Salmon Limited,unpubl. data). It appears that at least some of NewZealand's coastal haptophytes are existing at thelower end of their temperature range and this mightexplain why they are usually recorded in extremelylow numbers. Most species in this study wereisolated from sea water samples obtained betweenMarch and May, before temperatures had droppedbelow 16°C. The consistently lower temperaturerange of 10-16°C recorded in Big Glory Bay,Stewart Island (MacKenzie 1991), where C.pringsheimii has been recorded in high numbers(C. O'Kelly, Bigelow Laboratory for OceanSciences, ME, USA, pers. comm.), suggests thatthe warmer temperature group of Chryso-chromulina species will be excluded from southernNew Zealand waters.
Chrysochromulina simplex and C. ericina ofthis study grew equally well on a variety of nitrogensources, as did C. quadrikonta and C. camella(Rhodes etal. 1993), whereas C. acantha showed apreference for nitrate and C. hirta for ammoniumand urea. Unlike C. quadrikonta, none of the speciestested in this study had a selenium requirement.
All the New Zealand Chrysochromulina isolatesinvestigated grow over a wide range of salinities,flourishing in the slightly brackish coastal waters.On both occasions when C. quadrikonta bloomedin the Hauraki Gulf (May 1994 and June 1995),salinities were between 30.5 and 30.7. Chryso-chromulina species also tolerate a wide range oflight intensities (Rhodes 1994). This flexibility,combined with the ability of C. ericina and
Dow
nloa
ded
by [
Cal
ifor
nia
Inst
itute
of
Tec
hnol
ogy]
at 0
4:37
04
Oct
ober
201
4

Rhodes & Burke—Chrysochromulina spp. in New Zealand 101
C. acantha to phagocytose, might give these speciesa competitive advantage over other phytoplanktongenera.
The Chrysochromulina species are difficult todifferentiate by light microscopy, although acombination of flagella number and length, spinescale length, cell size and shape, and haptonematallength can aid identification (Fig. 7). Fluorescentprobes are also an aid in the identification of somemorphologically similar species, and can confirmgenus differentiation.
The fluorescence of spine scales of C. ericinaand C. hirta with Calcofluor White M2R aidsdiscrimination of these species using lightmicroscopy. However, they might still requireelectron microscopy for final species confirmationas their spine scales are of a similar length andtheir differences in form are not always clearlydefined at the lower magnifications of lightmicroscopy, despite fluorescence.
FITC-conjugated lectins enable the differ-entiation of those Chrysochromulina species testedfrom the other haptophyte genera. The genusChrysochromulina was originally separated fromthe genus Prymnesium (Massart ex Conrad) on thebasis of longer haptonematal length and haptone-matal coiling in Chrysochromulina (Manton &Leadbeater 1974) and on the existence of only onescale type in Prymnesium. However it is nowrecognised that long haptonemata do not alwaysexhibit coiling, for example in C. parkeae (Green& Leadbeater) (Green & Leadbeater 1972) and C.spinifera ((Fournier) Pienaar & Norris (cf. Parke& Dixon)) (Pienaar & Norris 1979), and that thereare species with short non-coiling haptonematathat have dimorphic scales (Pienaar & Kleizen1976). The discrimination of the genera on thesebases is therefore untenable and the possibility ofcombining them has been raised (Pienaar & Norris1979). Differentiation of Prymnesium andChrysochromulina at the ultrastructural level isalso open to question. For example, mostChrysochromulina species possess broad flagellarroots with relatively few micro tubules, and manymembers of the genus Prymnesium have multi-membered flagellar roots (Moestrup & Thomsen1986). However, there are exceptions, as in thelarge, compound roots of Chrysochromulina sp.,as described by Birkhead & Pienaar (1995b).
The binding of wheat germ lectin (WGA) to allof the Chrysochromulina species tested, but toneither of the Prymnesium species, supports anydecision to retain the separation of the genera.
However, more species, and strains from differentgeographic locations, should be tested to confirmthis finding. WGA appears to bind to the cellmembrane rather than to the unmineralised scales,as no spine scales have been observed fluorescing,although it is possible that lectin receptor sites arepresent on the scales, but inaccessible.
The marine haptophytes of New Zealand'swaters are, as elsewhere in the world, a notablecomponent of the phytoplankton. Already 22species of the 55 described in the genusChrysochromulina have been observed in NewZealand coastal waters (Table 6). Several hapto-phyte species not yet recorded in New Zealandhave been observed in the east Australian current(Hallegraeff 1983) and it is likely that they will beobserved here over the next few years. The abilityof some haptophytes to develop resting forms could
Table 6 Chrysochromulina species identified in NewZealand coastal waters by transmission electronmicroscopy of their unmineralised scales.
Species Site
C. acanthaC. alifera a
C. apheles h
C. brevifilum a
C. camella a
C. chiton a
C. ephippium a
C. ericina a
C. aff. fragilisaC. hirtaC. leadbeateriC. mactra 3
C. novae-zelandiae3
C. parkeae a
C. pachycylindraC. cf. polylepisC. pringsheimii d
C. quadrikonta c
C. simplex a
C. sp. ("eyelash scales")''C. spinifera a
C. strobilus e
"Pakiri BeachKaikouraKaikoura#Kaikoura"Leigh; Marlborough
SoundsKaikoura"Kaikoura#Kaikoura; Marlborough
Sounds; Stewart IslandLeigh"Marlborough Sounds"Tauranga HarbourLeighKaikouraLeigh, Kaikoura#Tauranga Harbour"Tauranga HarbourStewart IslandNelson Harbour,
Waimangu Point"Pakiri Beach, Tauranga
Harbour; Kaikoura* Tauranga HarbourKaikouraMarlborough Sounds
# Described from New Zealand waters in this studyaMoestrup 1979bMoestrup & Thomsen 1986cRhodesetal. 1993dC.O' Kelly, pers. comm.eRhodes 1994
Dow
nloa
ded
by [
Cal
ifor
nia
Inst
itute
of
Tec
hnol
ogy]
at 0
4:37
04
Oct
ober
201
4

102 New Zealand Journal of Marine and Freshwater Research, 1996, Vol. 30
assist in their dispersal via ship's ballast water(Hallegraeff & Bolch 1992); transport over longdistances has already been demonstrated onseaweed bearing mussel spat.
ACKNOWLEDGMENTS
The technical support of Maggie Atkinson, CawthronInstitute, Nelson, New Zealand, has been appreciated,as has the support of Charley O'Kelly, BigelowLaboratory for Ocean Sciences, Maine, USA; Julie Hall,NIWA, Hamilton, New Zealand and David Fountain,Department of Plant Biology and Biotechnology, MasseyUniversity, Palmerston North, New Zealand. Thanks toDoug Hopcroft and Raymond Bennett, Horticulture andFood Research Institute, NZ, Ltd, Palmerston North, forvalued assistance with electron microscopy. This studywas supported by a contract with the Foundation forResearch, Science and Technology and a Lottery ScienceResearch grant. L. Rhodes was personally supported bya New Zealand Universities Postgraduate Scholarshipand a Cawthron Scholarship.
REFERENCES
Allen, D. M.; Northcote, D. H. 1975: The scales ofChrysochromulina chiton. Protoplasma 83:389-412.
Andersen, R. A.; Jacobson, D. M.; Sexton, J. P. 1991:Catalog of strains. Provasoli-Guillard Centre forculture of marine phytoplankton, West BoothbayHarbor, Maine, USA. 98 p.
Birkhead, M.; Pienaar, R. N. 1995a: The taxonomy andultrastructure of Chrysochromulina simplex(Prymnesiophyceae). Phycologia 34: 145-156.
Birkhead, M.; Pienaar, R. N. 1995b: The flagellarapparatus of Chrysochromulina sp. (Prym-nesiophyceae). Journal of phycology 31: 96-108.
Chang, F. H. 1995: The first records of Gymnodiniumsp. nov. (cf. breve) (Dinophyceae) and otherharmful phytoplankton species in the early 1993blooms in New Zealand. In: Lassus, P.; Arzul,G.; Erard, E.; Gentien, P. Marcaillou, C. ed.Harmful marine algal blooms. Lavoisier,Intercept Ltd. pp. 27- 32.
Costas, E.; Gonzalez-Chavarri, E.; Aguilera, A.;Gonzalez-Gil, S.; Lopez-Rodas, V. 1993: Useof lectins to recognise and differentiateunicellular algae. Botanica marina 36: 1-4.
Costas, E.; Rodas, V. L. 1994: Identification of marinedinoflagellates using fluorescent lectins. Journalof phycology 30: 987-990.
Edvardsen, B.; Paasche, E. 1992: Two motile stages ofChrysochromulina polylepis (Prymnesio-phyceae): morphology, growth, and toxicity.Journal of phycology 28: 104-114.
Estep, K. W.; Davis, P. G.; Hargreaves, P. E.; Sieburth,J. McN. 1984: Chloroplast containing micro-flagellates in natural populations of NorthAtlantic nanoplankton, their identification anddistribution; including a description of five newspecies of Chrysochromulina (Prymnesio-phyceae). Protistologica 20: 613-634.
Green, J. C.; Leadbeater, B. S. C. 1972: Chryso-chromulina parkeae sp. nov. (Haptophyceae) anew species recorded from S.W. England andNorway. Journal of the Marine BiologicalAssociation of the United Kingdom 52: 469-474.
Gross, M. G. 1967: Oceanography. Merrill Books Inc.,Ohio.
Hallegraeff, G. M. 1983. Scale-bearing loricatenanoplankton from the east Australian current.Botanica marina 26: 493-515.
Hallegraeff, G. M.; Bolch, C. J. 1992: Transport ofdiatom and dinoflagellate resting spores in ship'sballast water: implications for planktonbiogeography and aquaculture. Journal ofplankton research 14: 1067-1084.
Hughs, J.; McCully, M. E. 1975: The use of an opticalbrightener in the study of plant structure. Staintechnology 50: 319-329.
Kawai, H.; Inouye, I. 1989: Flagellar autofluorescencein forty-four chlorophyll c-containing algae.Phycologia 28: 222-227.
Larsen, J.; Moestrup, Ø. 1989: Guide to toxic andpotentially toxic marine algae. Ministry ofFisheries, Denmark, pp. 46-49.
Leadbeater, B. S. C. 1972a: Fine structural observationson six new species of Chrysochromulina(Haptophyceae) from Norway. Sarsia 49: 65-80.
Leadbeater, B. S. C. 1972b: Identification, by means ofelectron microscopy, of flagellate nanoplanktonfrom the coast of Norway. Sarsia 49: 107-124.
Leadbeater, B. S. C.; Manton, I. 1971: Fine structureand light microscopy of a new species ofChrysochromulina (C.acantha). Archiv fürMikrobiologie 78: 58-69.
Loeblich, A. R.; Smith, V. E. 1968: Chloroplast pigmentsof the marine dinoflagellate Gyrodiniumresplendens. Lipids 3: 5-13
MacKenzie, L. 1991: Toxic and noxious phytoplanktonin Big Glory Bay, Stewart Island, New Zealand.Journal of applied phycology 3: 19-34.
Manton, I. 1978: Chrysochromulina hirta sp. nov., awidely distributed species with unusual spines.British phycological journal 13: 3-14.
Manton, I.; Leadbeater, B. S. C. 1974: Fine-structuralobservations on six species of Chrysochromulinafrom wild Danish marine nanoplankton,including a description of C. campanulifera sp.nov. and a preliminary summary of thenanoplankton as a whole. Det Kongelige Danskevidenskabernes selskab biologiske skrifter 20:1-26.
Dow
nloa
ded
by [
Cal
ifor
nia
Inst
itute
of
Tec
hnol
ogy]
at 0
4:37
04
Oct
ober
201
4

Rhodes & Burke—Chrysochromulina spp. in New Zealand 103
Manton, I.; Leedale, G. F. 1961: Further observationson the fine structure of Chrysochromulina ericinaParke & Manton. Journal of the MarineBiological Association of the United Kingdom41: 145-155.
Manton, I; Oates, K.; Course, P. A. 1981: Cylinder-scales in marine flagellates from the genusChrysochromulina (Haptophyceae=Prym-nesiophyceae) with a description of C. pachy-cylindra sp. nov. Journal of the MarineBiological Association of the United Kingdom61: 17-26.
Manton, I.; Parke, M. 1962: Preliminary observationson scales and their mode of origin in Chryso-chromulina polylepis sp. nov. Journal of theMarine Biological Association of the UnitedKingdom 42: 565-578.
Moestrup, Ø. 1979: Identification by electron microscopyof marine nanoplankton from New Zealand,including the description of four new species.New Zealand journal of botany 17: 61-95.
Moestrup, Ø.; Thomsen, H. A. 1986: Ultrastructure andreconstruction of the flagellar apparatus inChrysochromulina apheles sp. nov. (Prym-nesiophyceae = Haptophyceae). Canadianjournal of botany 64: 593-610.
Parke, M.; Manton, I.; Clark, B. 1956: Studies on marineflagellates. 3. Three further species of Chryso-chromulina. Journal of the Marine BiologicalAssociation of the United Kingdom 35: 387-414.
Persoone, G.; Wells, P. G. 1987: Anemia in aquatictoxicology: a review. In: Sorgeloos, P.; Bengston,D. A.; Decleir, W.; Jaspers, E. ed. Artemiaresearch and its applications. Vol. 1. UniversaPress, Belgium, pp. 259- 275.
Pienaar, R. N.; Kleizen, H. G. 1976: A comparativestudy of two species of the toxic alga Prym-nesium. Proceedings of the Electron MicroscopySociety of Southern Africa 6: 55-56.
Pienaar, R. N.; Norris, R. E. 1979: The ultrastructure ofthe flagellate Chrysochromulina spinifera(Fournier) comb. nov. (Prymnesiophyceae) withspecial reference to scale production. Phycologia18: 99-108.
Rhodes, L. L.; O'Kelly, C. J.; Hall, J. A. 1993:Comparison of growth characteristics of NewZealand isolates of Chrysochromulina quad-rikonla and C. camella with those of theichthyotoxic species C. polylepis. Journal ofplankton research 16: 69-82.
Rhodes, L. L. 1994: Prymnesiophytes of New Zealandcoastal waters. Unpublished PhD thesis, MasseyUniversity, Palmerston North, New Zealand.
Dow
nloa
ded
by [
Cal
ifor
nia
Inst
itute
of
Tec
hnol
ogy]
at 0
4:37
04
Oct
ober
201
4





























